
Menstrual blood transplantation for ischemic stroke: Therapeutic mechanisms and practical issues

bFGF induces an earlier expression of nephrogenic proteins after ischemicacute renal failure
Sandra Villanueva, Carlos Cespedes, Alexis Gonzalez, and Carlos P. VioDepartamento de Fisiologia, Pontificia Universidad Catolica de Chile, Santiago, Chile
Submitted 11 January 2006; accepted in final form 20 July 2006
Villanueva, Sandra, Carlos Cespedes, Alexis Gonzalez, andCarlos P. Vio. bFGF induces an earlier expression of nephrogenicproteins after ischemic acute renal failure. Am J Physiol Regul IntegrComp Physiol 291: R1677–R1687, 2006. First published July 27,2006; doi:10.1152/ajpregu.00023.2006.—Recovery from acute renalfailure (ARF) requires the replacement of injured cells with new cellsthat restore tubule epithelial integrity. We described recently theexpression of a wide range of nephrogenic proteins in tubular cellsafter ARF induced by ischemia-reperfusion (I/R) (Villanueva S,Cespedes C, and Vio CP. Am J Physiol Regul Integr Comp Physiol290: R861–R870, 2006). These markers, namely, Vimentin, neuralcell adhesion molecules (Ncam), basic fibroblast growth factor(bFGF), paired homeobox-2 (Pax-2), bone morphogene protein-7(BMP-7), Noggin, Lim-1, Engrailed, Smad, phospho-Smad, hypoxia-induced factor-1� (HIF-1�), VEGF, and Tie-2, are expressed in a timeframe similar to that observed in normal kidney development. bFGFparticipates in early kidney development as a morphogen involved inmesenchyme/epithelial transition, and it is reexpressed in the recoveryphase of ARF. To test the hypothesis that bFGF can accelerate theregeneration after renal damage, we used recombinant bFGF andstudied the expression pattern of the above described morphogens inARF. Male Sprague-Dawley rats were subjected to 30 min of renalischemic injury and were injected with bFGF 30 �g/kg followed byreperfusion. Rats were killed and the expression of nephrogenicproteins were analyzed by immunohistochemistry and Western blotanalysis. In the animals subjected to I/R treated with bFGF, weobserved a 12- to 24-h earlier and more abundant reexpression of theproteins Ncam, bFGF, Pax-2, BMP-7, Noggin, Lim-1, Engrailed,VEGF, and Tie-2 than the I/R untreated rats. In addition, we observeda reduction in renal damage markers ED-1 and �-smooth muscleactin. These results indicate that bFGF can participate in the regen-eration process and suggest that the treatment with bFGF can inducean earlier regeneration process after ischemic acute renal failure.
kidney; morphogen; regeneration
ACUTE RENAL FAILURE IS A CLINICAL syndrome characterized by arapid decline in glomerular filtration rate and is associated withhigh morbidity and mortality; however, it is potentially revers-ible if patients survive the initial insult (47). Acute tubularnecrosis (ATN) due to poor perfusion is the most commoncauses of ARF, accounting for two-thirds of the intrinsiccauses (15). The principal cause of ATN is hypoxia induced byI/R, which can be caused by clinical conditions, such ashemorrhagic shock or sepsis (35). ATN is characterized by aregeneration phase (29), which involves recovery of kidneyfunction, with a sequence of events, including epithelial celldedifferentiation and proliferation, followed by differentiationand restoration of the nephron functional integrity (1). Al-though the morphological characteristics of this process have
been described (55), the molecular basis of the events leadingto regeneration after ATN is not completely understood (48).
Many genes are modulated in response to kidney damage (53).The expression of transcription factors like c-myc (10), c-jun (4)and EGR-1 (21); growth factors human growth factor (HGF) andIGF (46), Adam, HO1, UCP-2, thymosine b4 (64); and proapop-totic factors FADD, DAXX, BAX, BAD, and p53 (53) areupregulated after kidney damage, whereas EGF, cytochromeP-450, Iid6, cyp 2d9, and ADH B2 expression are downregulated(64). Yet, the expression control of these genes is a common eventto several pathological processes and cannot explain the regener-ation processes that occur during ATN.
One possibility is that renal regeneration may recapitulatepart of the kidney genetic program during organogenesis,including apoptosis (2). Several reports have shown that neph-rogenic proteins present in the regeneration process after in-duced ATN, such as mesenchymal proteins Ncam (1), WT-1(62), and Vimentin (63); epithelial proteins paired homeobox-2(Pax-2) (45) and zone occludens-1 (ZO-1); and tubular proteinsLim-1 and Engrailed, which play a crucial role during earlymetanephric development and remain detectable only in col-lecting tubular cells in adulthood (10), are reexpressed inregenerating tubular epithelium after renal damage (20, 57).
Other examples are the reports on bone morphogen protein7 (BMP-7) (12), its antagonist Noggin (9, 56), and the tran-scription factors Smad 1 and 5 (28), which are involved inphosphorylation of the BMP transcription pathway. Thesemorphogenic proteins are expressed by mesenchymal cells inthe nephrogenic zone and are downregulated once these cellsbegin to form epithelium (13, 61); however, we have recentlydescribed the reexpression of these proteins in the regenerationphase, after ATN induced by I/R in the same sequential patternthan the observed during kidney development (57).
On the other hand, there are other proteins expressed duringkidney development, such as vascular endothelial growth fac-tor (VEGF), expressed in epithelial and endothelial cells of therenal corpuscle (7) and the angiopoietin receptor, Tie-2, thatare reexpressed in later maturation stages of mouse metane-phros, when interstitial and glomerular capillaries begin toform (27). Both proteins stimulate glomerulus development(17, 25) and maturation of blood vessels during embryonicdevelopment (22, 27). Under physiological conditions, VEGFand Tie-2 are induced by hypoxia-induced factor-1 � (HIF-1�;18, 44). We have described recently the reexpression of thesethree proteins: HIF-1�, VEGF, and Tie-2 in the regenerationphase of ATN induced by I/R (57).
Another important protein secreted early by the ureteric budis the basic fibroblast growth factor (bFGF). bFGF is necessary
Address for reprint requests and other correspondence: S. Villanueva, Dept.de Fisiologia, Pontificia Universidad Catolica de Chile, Casilla 114-D, San-tiago, Chile (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Regul Integr Comp Physiol 291: R1677–R1687, 2006.First published July 27, 2006; doi:10.1152/ajpregu.00023.2006.
0363-6119/06 $8.00 Copyright © 2006 the American Physiological Societyhttp://www.ajpregu.org R1677
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

for the induction of mesenchymal cells aggregation (23); how-ever, it is not capable of turning these aggregates into epithelialcells (28). The main effect of bFGF is the inhibition ofapoptosis, the promotion of the condensation of mesenchymalcells, the maintenance of synthesis of WT-1, a transcriptionfactor that induces the transformation of mesenchymal cellsinto metanephrogenic tissue (42), and the early tubulogenesisin kidney embryonic development (41, 49). The expression ofthis protein is not observed in adult kidney, but a stronginduction is observed in the regeneration phase of ATN asdescribed recently by us (57). In addition, bFGF promotes theconversion of tubular epithelium to a cell with mesenchymalcharacteristics and can participate in the epithelial mesenchy-mal transition (52).
In this regard, we have recently described the reexpressionof important morphogenes in the regeneration phase of ARFinduced by I/R, in a temporal pattern similar to that observedin kidney development (57). This theory opens the possibilitythat these morphogenes can be used to accelerate the repairprocess induced by ARF. BMP-7 is the only morphogenobserved in our study that has previously been used in a renalfibrosis model, and it was capable of blunting the progressionof the fibrotic disease and can accelerate the return to normalrenal function (36, 65).
In this study, we evaluated the effect of morphogen bFGF inthe repair of kidney damage; this is a protein expressed early inkidney development (23, 41, 52) and is reexpressed in theregeneration phase after ARF (57). We studied the expressionpattern of several morphogenes and the transcription factorsmentioned previously in ARF in kidneys pretreated with arecombinant bFGF. We observed that the reexpression of theanalyzed morphogens was 12 to 24 h earlier in time and higherin intensity, than the one observed in rats with I/R but withoutbFGF administration. These results indicate that bFGF partic-ipates in the regeneration process and suggests that the treat-ment with bFGF can accelerate the regeneration process of ratkidneys after ischemic renal failure.
MATERIALS AND METHODS
Animals. Adult male Sprague-Dawley rats (220–250 g), werehoused in 12:12-h light-dark cycle and maintained at the Universityanimal care facilities; food and water were supplied ad libitum. Allexperimental procedures were in accordance with institutional andinternational standards for the human care and use of laboratoryanimals (Animal Welfare Assurance Publication A5427–01, Officefor Protection from Research Risks, Division of Animal Welfare,National Institutes of Health), as previously described (57). In thisstudy, the protocol of use of animals was reviewed and approved bythe institutional and independent Ethical Committee of the PontificiaUniversidad Catolica de Chile.
Renal I/R injury and bFGF treatment. An established model ofrenal I/R injury was performed recently by us (57); this resemblesstructural and functional consequences of renal ischemia, includingapoptotic tubular epithelial cells (6). Animals (n � 5 for each I/Rgroup) were anesthetized with ketamine:xylazine (25:2.5 mg/kg ip).Body temperature was maintained at 37°C. Both kidneys were ex-posed by a flank incision, and both renal arteries were occluded withnontraumatic vascular clamps for 30 min. After 30 min of clamping,the left kidney was injected intrarenally with bFGF (30 �g/kg) (30,31, 54) in a total volume of 200 �l and was considered to be treated;the right kidney was injected with the same amount of saline andserved as a control. After the injection, both clamps were removed,
renal blood flow was reestablished, and the incisions were sutured. Athird group of sham animals was included; these animals weresubjected to the same surgical procedure and conditions, withoutclamping the renal arteries. Rats were allowed to recover in a warmroom with water and food ad libitum. Rats were killed under anes-thesia (ketamine:xylazine) 24, 48, 72, and 96 h after reperfusion; bothkidneys were removed and processed for immunohistochemistry andWestern blot analysis.
Tissue processing and immunohistochemical analysis. Immunohis-tochemical studies in Paraplast-embedded sections, were performed aspreviously described (57, 58). For cryosections, the kidney sections (3to 4 mm thick) were processed as recently described (57).
Immunolocalization studies were performed using an indirect im-munoperoxidase technique as recently described (57). Briefly, thetissue sections were incubated with the primary antibody, followed byincubation with the corresponding secondary antibody and with theperoxidase-antiperoxidase (PAP) complex and revealed using 3,3�-diaminobenzidine (DAB). For some specific antibodies, immunoreac-tivity was revealed using a secondary antibody conjugated to alkaline-phosphatase, in the presence of nitroblue tetrazolium chloride: 5-bro-mo-4chloro-3indolyl phosphate p-toludine salt (4.5:3.5 �l/ml) inbuffer Tris 100 mM pH 9.5. Controls for the immunostaining proce-dure were prepared by omission of the first antibody by its replace-ment with normal or preimmune serum of the same species (59).
Antibodies and chemicals. The primary antibodies used correspondto the same antibodies used recently by us (57): the monoclonalantibodies against Lim 1�2 (clone 4F2), Engrailed (clone 4G11),Vimentin (clone 40E-C), neural cell adhesion molecules (Ncam; clone5B8), ZO-1 (clone R26.4C) were obtained from the DevelopmentalStudies Hybridoma Bank developed under the auspices of the NICHDand maintained by the University of Iowa, Department of BiologicalSciences (Iowa City, IA). The goat polyclonal antibodies against Pax2, BMP-7, Smads 1–5-8, phospho-Smad (p-Smad) 2–3; the rabbitpolyclonal antibodies against Tie-2, bFGF, and the monoclonal anti-body against VEGF (clone C-1) were obtained from Santa CruzBiotechnology (Santa Cruz, CA). The monoclonal antibodies againstmacrophages (clone ED-1) were obtained from Biosource (Camarillo,CA), �-smooth muscle actin (�-SMA; clone 1A4) was obtained fromSigma-Aldrich (St. Louis, MO), HIF-1� (clone H1�67) was obtainedfrom Novus Biological (Littleton, CO), and Noggin was a gift fromDr. R. Harland.
Secondary antibodies and the corresponding PAP complexes werepurchased from ICN Pharmaceuticals-Cappel (Aurora, OH). TritonX-100, DAB, carrageenan, Tris �HCl, hydrogen peroxide, phosphatesalts, and other chemicals were purchased from Sigma-Aldrich.
Immunoblotting. Whole kidney sections (�1 mm thick) werehomogenized, and the protein concentration was determined as pre-viously described (57). Western blot analysis was performed asdescribed by Harlow and Lane (19). For SDS-PAGE, proteins weremixed with sample buffer (100 mM Tris �HCl, pH 6.8, 200 mMdithiothreitol, 4% SDS, 0.2% bromophenol blue, 20% glycerol),transferred to nitrocellulose membranes and blocking, as previouslydescribed (57). After blocking, the membranes were probed with thecorresponding antibody, washed with Tris-buffered saline-Tween, andincubated with horseradish peroxidase-conjugated secondary antibodyfor 1 h at room temperature. Immunoreactivity was detected usingenhanced chemiluminescence technique obtained from Perkin-Elmer,Life Sciences (Boston, MA). The positive control was an embryo at17 days of development, and the negative control was obtained fromsham-operated rat kidneys. The Western blots were run with the totalsamples (n � 5) in each time-period, and the selected blot correspondsto a one representative from the group.
The blots were scanned, and densitometric analysis was performedusing the public domain National Institutes of Health (NIH) Imageprogram v1.61 (U.S. NIH, http://rsb.info.nih.gov/nih-image). The ex-pression of tubulin was used to correct for variation in sample loading.
R1678 EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from
Detection and quantification of renal cell apoptosis by in situ endlabeling of fragmented DNA and caspase-3 detection. Apoptotic cellsin kidney tissue slices were visualized using the apop tag fluoresceinin situ apoptosis detection kit by the indirect terminal deoxynucleo-tidyl transferase-mediated dUTP nick-end labeling (TUNEL) methodfrom Chemicon (Temecula, CA) following the manufacturer’s proto-col. Caspase-3 was evaluated with the appropriate antibody obtainedfrom Promega (Madison, WI), which was previously described (57).The fluorescence was viewed by microscopy using appropriate exci-tation and emission filters.
Determination of functional and tissue damage. As previouslydescribed (57), functional damage was assessed through serum creat-inine levels (51), and tissue damage was evaluated using periodicacid-Schiff (PAS) staining and immunolocalization of interstitialmacrophages (ED-1) and myofibroblasts (�-SMA).
The �-SMA immunoreactive area in each image was determinedby image analysis using Simple PCI software (Compix). The valuescorresponding to total immunostained (brown) cells were averagedand expressed as the mean absolute values and the mean percentage ofstained cells area per field (0.064 mm2) with a modification of apreviously described method (60).
Statistical analysis. The differences were assessed with the nonpara-metric Mann-Whitney U-test for pairwise comparisons when overallsignificance was detected. The significance level was P � 0.05.
RESULTS
Determination of functional and tissue damage. Renal dam-age by I/R verified by serum creatinine levels showed thatcreatinine levels in rats subjected to the I/R protocol injectedwith saline was 2.3 � 1.8 mg/dl and was lower (1.5 � 0.5mg/dl) in animals injected with bFGF during the early phase ofATN (24 to 48 h after I/R).
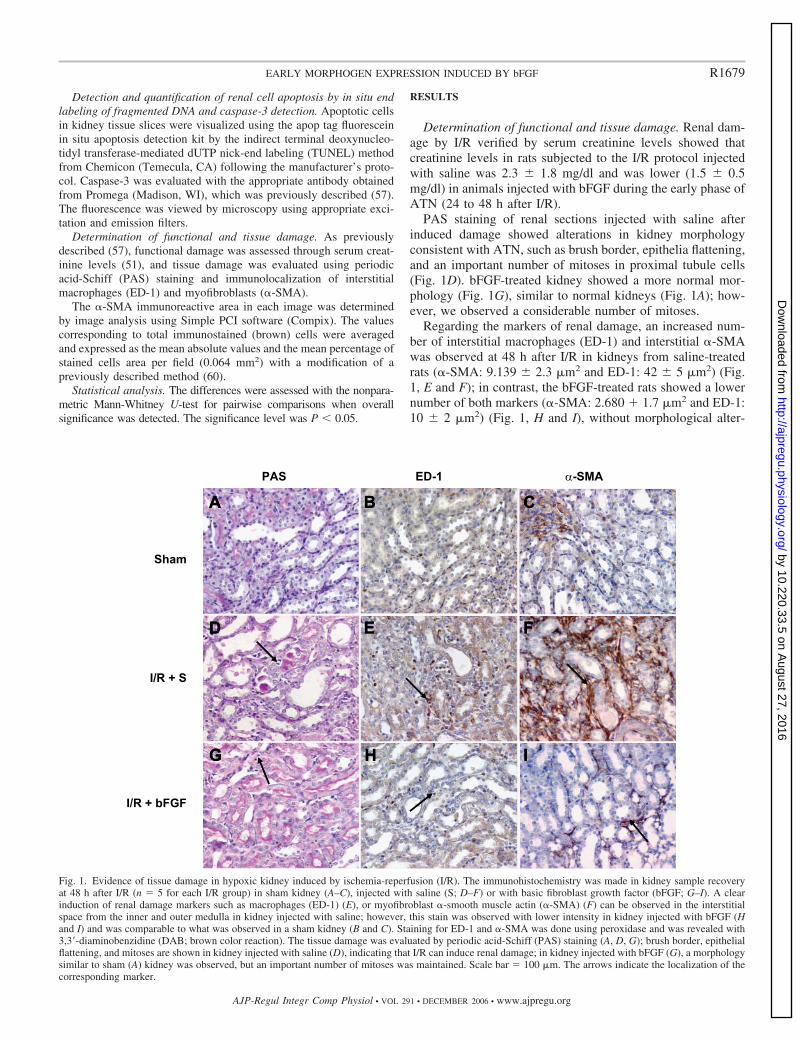
PAS staining of renal sections injected with saline afterinduced damage showed alterations in kidney morphologyconsistent with ATN, such as brush border, epithelia flattening,and an important number of mitoses in proximal tubule cells(Fig. 1D). bFGF-treated kidney showed a more normal mor-phology (Fig. 1G), similar to normal kidneys (Fig. 1A); how-ever, we observed a considerable number of mitoses.
Regarding the markers of renal damage, an increased num-ber of interstitial macrophages (ED-1) and interstitial �-SMAwas observed at 48 h after I/R in kidneys from saline-treatedrats (�-SMA: 9.139 � 2.3 �m2 and ED-1: 42 � 5 �m2) (Fig.1, E and F); in contrast, the bFGF-treated rats showed a lowernumber of both markers (�-SMA: 2.680 � 1.7 �m2 and ED-1:10 � 2 �m2) (Fig. 1, H and I), without morphological alter-
Fig. 1. Evidence of tissue damage in hypoxic kidney induced by ischemia-reperfusion (I/R). The immunohistochemistry was made in kidney sample recoveryat 48 h after I/R (n � 5 for each I/R group) in sham kidney (A–C), injected with saline (S; D–F) or with basic fibroblast growth factor (bFGF; G–I). A clearinduction of renal damage markers such as macrophages (ED-1) (E), or myofibroblast �-smooth muscle actin (�-SMA) (F) can be observed in the interstitialspace from the inner and outer medulla in kidney injected with saline; however, this stain was observed with lower intensity in kidney injected with bFGF (Hand I) and was comparable to what was observed in a sham kidney (B and C). Staining for ED-1 and �-SMA was done using peroxidase and was revealed with3,3�-diaminobenzidine (DAB; brown color reaction). The tissue damage was evaluated by periodic acid-Schiff (PAS) staining (A, D, G); brush border, epithelialflattening, and mitoses are shown in kidney injected with saline (D), indicating that I/R can induce renal damage; in kidney injected with bFGF (G), a morphologysimilar to sham (A) kidney was observed, but an important number of mitoses was maintained. Scale bar � 100 �m. The arrows indicate the localization of thecorresponding marker.
R1679EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

ations (Fig. 1, B and C). These differences were significant(P � 0.05).
In situ cell death detection and caspase-3 immunohisto-chemistry. TUNEL was used to detect DNA double-strandbreaks. In control kidneys injected with saline, TUNEL stain-ing was predominantly localized to proximal tubule cells and tosome collecting duct cells from the inner and outer medulla(Fig. 2A). In addition, bFGF-treated kidneys did not show anyreaction by the TUNEL assays (9.82/hpf) (Fig. 2B). In animalsinjected with saline, 24 h after I/R, a markedly increasedsignal, staining the whole nucleus, was observed (�22.46/highpower factor) (Fig. 2A), this signal decreased gradually there-after. The majority of TUNEL-positive cells exhibited mor-phological features of apoptotic death (shrinkage of cytoplasmand condensation of nucleus).
To verify whether this TUNEL-positive signal was due toapoptosis, we examined caspase-3 cleavage (activation) inischemic rat kidney with or without bFGF. Caspase-3 immu-nostaining dramatically increased in the inner and outer me-dulla area of the kidney injected with saline, at 24 h (24.85/high power factor) after I/R (Fig. 2C), and decreased after 48 hafter I/R. At 24 h, the majority of caspase-3 positive cells hadapoptotic morphology, and the staining was predominantlypresent in the proximal tubular cells and to a smaller degree incollecting duct cells. The correlation between caspase-3 andTUNEL was evident at 24 h postischemia. We determined thepositive correlation (r � 0.68) between caspase-3 protein andTUNEL (P � 0.05). bFGF-treated kidney did not showcaspase-3 immunostaining (Fig. 2D).
Detection of hypoxic tissue by markers induced by hypoxia:HIF-1�, VEGF, and Tie-2. As reported previously, followingrenal artery clamping and injection with saline, a strong nu-clear accumulation of HIF-1� occurred within 30 min. HIF-1�staining increased and reached its maximum at 48 h after I/R inthe kidney injected with saline (Figs. 3.1A and 4A); from 48 honward, the number of HIF-1�-positive cell nuclei declined,disappearing at 96 h (Fig. 3.1A). In the kidney treated withbFGF, HIF-1� immunostaining was observed at 24 h after I/R;48 h after I/R, the area of staining was larger and with a higher
intensity (Fig. 4D). The expression of HIF-1� was observed inpapillary collecting ducts, thick ascending limb, and proximaltubular cells, mainly located in the inner and outer medulla ofkidneys with I/R.
VEGF and Tie-2 are markers induced by hypoxia andpromote angiogenesis; thus, we observed a marked expressionof VEGF at 24 h after I/R in the kidney injected with saline(Figs. 3.1B and 4B), which was significantly decreased by72–96 h. In the kidney treated with bFGF, we observed anincreased expression of VEGF at 24 h after I/R; this level wasmaintained up to 48 h and then decreased by 72–96 h (Figs.3.1B and 4E). Tie-2 immunoreactivity in the kidney injectedwith saline had the highest-level expression at 48 h after I/R(Figs. 3.1C and 4C), decreased at 72 h, and was completelyabsent at 96 h. In the kidney treated with bFGF, we observedan increased immunostaining for Tie-2 at 24 h after I/R, whichpeaked at 48 h and decreased by 72–96 h, but with superiorlevels to what was observed with saline (Figs. 3.1C and 4F).The expression of all these proteins was observed in proximaltubule cells, mainly localized in the inner and outer medulla(Fig. 4, A–F).
Expression of mesenchymal, epithelial, and tubular markersin kidney after I/R. Representative expression patterns areshown for mesenchymal, epithelial, and tubular markers inkidneys injected with saline or bFGF after I/R in mesenchymal,epithelial, and tubular markers. ATN induced by I/R in ratsinjected with saline or bFGF, leads to an altered distributionpattern of mesenchymal markers (bFGF, Ncam, Pax-2, Vimen-tin, and Noggin), epithelial markers (BMP-7 and Engrailed),and tubular markers (Lim-1 and ZO-1), similar to what hasbeen reported previously (57) (Figs. 3.2, 3.3, 5, A–H, 6, A–H,7B). In the healthy adult kidneys, these proteins could bedetected in collecting duct cell nuclei, with the highest expres-sion in the papilla. Cells from proximal tubules, glomeruli, orperitubular cells localized in the inner/outer medulla and med-ullary rays were devoid of labeling (57).
In kidneys injected with saline, the highest expression forbFGF, Ncam, and Noggin was observed at 24 h; and forBMP-7, Engrailed, Lim-1, and ZO-1 at 48 h after I/R, the
Fig. 2. Distribution of terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) and caspase-3-positive cells in hypoxia-induced by I/R.A and B: TUNEL techniques and immunohistochemistry for caspase-3 (C and D) were performed in kidney samples obtained 48 h after I/R injected with saline(A and C) or bFGF (B and D) (n � 5 for each I/R group). A clear staining localized to proximal tubular cells from the inner and outer medulla area was observedat the first stage of damage induced by acute tubular necrosis (ATN) in kidney injected with saline (A and C), which was diminished in kidney injected withbFGF (B and D). Staining for TUNEL and caspase-3 was done using fluorescence (Texas red and green color reaction, respectively). Scale bar � 100 �m. Thearrows indicate the localization of the corresponding marker.
R1680 EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

expression of these proteins decreased at 72–96 h after I/R(Fig. 3.2, 3.3). These proteins were mainly localized in innerand outer medulla. bFGF and BMP-7 were localized in prox-imal tubule cells. Ncam, Engrailed, Lim-1, ZO-1, and Nogginwere localized in the peritubular area, similar to what wasreported previously (57) (Figs. 5, A–H, 6, A–H).
bFGF-treated kidneys, the highest expression for mesen-chymal and early epithelial markers bFGF, Ncam, Pax-2,Vimentin, and Noggin was observed at 24 h after I/R,maintained up to 72 h, and decreased at 96 h (Fig. 3.2 and3.4L). This expression pattern in the regenerative phase wasconsistent with cell dedifferentiation and coincident withcell proliferation (20, 63) (Fig. 5, E–H). For later epithelialand tubular markers, BMP-7, Engrailed, Lim-1, and ZO-1,the highest expression was observed at 24 h, peaked at 48 h,and was maintained up to 72 h after I/R (Figs. 3.3, 6, E–H).Positive cells could be identified as belonging to regenerat-ing proximal tubules in the inner and outer medulla, accord-ing to the high flattening epithelium and remnants of thedamaged brush border detected in the lumen of the tubules.The expression of all proteins in proximal tubule cell and
peritubular area was decreased at 96 h after I/R (Fig. 3.2 and3.3). At this point, all proteins were barely detectable in theproximal tubules, and the expression was again restricted tocollecting duct cells (data not shown). In summary, weobserved that all morphogenic protein levels were increasedand expressed in the time series in kidneys treated withbFGF (Fig. 3.2, 3.3) compared with kidneys subjected to I/Rand injected with saline.
BMP-7, a survival factor for undifferentiated mesenchyme(13), is antagonized by the Noggin protein (9, 56) and phos-phorylates the transcription factors Smad 1 and 5 (28). Thetranscription factor Smad (total and phosphorylated) was stud-ied in regenerating cells from kidneys injected with bFGF orsaline after I/R (Fig. 7, C, D, G, and H and Fig. 3.4, M and N).Reexpression of both states was observed in proximal tubulecells in inner and outer medulla; the maximum expression ofSmad and p-Smad was observed at 24–48 h after I/R (Fig. 7,C, D, G, H), decreasing at 72–96 h after I/R in kidneys injectedwith saline or bFGF, without apparent differences (Fig. 3.4, Mand N). These results were similar to the ones reported previ-ously (57).
Fig. 3. Immunoblot of nephrogenic proteins in acute renal failure (ARF) induced by I/R. 3.1: Expression of endothelial cell markers in kidney regeneration wasstudied by immunoblot for proteins: hypoxia-induced factor (HIF-1�; A), vascular endothelial growth factor (VEGF; B), Tie-2 (C) at 24, 48, 72, and 96 h after30-min ischemia in kidney injected with saline or bFGF. In kidneys injected with saline, an increase in HIF-1�, VEGF, and Tie-2 levels was observed, with amaximum at 48 h, followed by a decrease that was similar to control levels at 96 h. In kidneys injected with bFGF, an increase in this marker at 24 h was observed,maintained at 48 h after I/R, followed by a decrease at 72–96 h after I/R, but with a superior level that was observed in kidney injected with saline. 3.2: Expressionof metanephric mesenchymal and early epithelial markers in kidney regeneration was studied by immunoblot for morphogenic proteins bFGF (D), neural celladhesion molecule (Ncam; E), Pax-2 (F), and Vimentin (VIM; G) at 24, 48, 72, and 96 h after 30 min of ischemia. In kidneys injected with saline, an increaseof metanephric mesenchymal (MM) markers Vimentin and Ncam with a maximum at 48 h after ischemia was observed, the levels of both markers decreasedat 72 h and returned to a basal level at 96 h. The early epithelial proteins Pax-2 and bFGF are expressed from 24 to 72 h and disappeared at 96 h after ischemia.In kidneys injected with bFGF, an increase in this marker at 24 h was observed, maintained at 48 h after I/R, followed by a slight decrease at 72–96 h after I/R.The expression observed in kidney injected with bFGF is superior to that observed in kidney injected with saline. 3.3: Expression of epithelial and tubular markersin kidney regeneration was studied by immunoblot for morphogenic proteins bone morphogen protein-7 (BMP-7; H), Engrailed (ENG; I) and Lim 1�2 (J) at24, 48, 72, and 96 h after 30 min ischemia. An increase in epithelial and tubular proteins can be observed, with a maximum at 48 h and a decrease at 72 h afterischemia in kidneys injected with saline. In kidneys injected with bFGF, an increase in these markers at 24 h was observed, maintained at 48 h after I/R, followedby a decrease at 72–96 h after I/R. The expression observed in kidneys injected with bFGF was superior to that observed in kidneys injected with saline. 3.4:Expression of transcription pathway for BMP-7 was studied by immunoblot for morphogenic proteins BMP-7 (K), Noggin (L), and the nonphosphorylated (M)and phosphorylated (N) form of the transcription factor Smad at 24, 48, 72, and 96 h after 30 min of ischemia. A maximum increase in BMP-7 and Noggin proteinlevels was observed at 48 h followed by a decrease at 72–96 h after ischemia in kidneys injected with saline. In kidneys injected with bFGF, an increase in bothmarkers at 24 h was observed, which peaked at 48 h after I/R and was followed by a slight decrease at 72–96 h after I/R. The expression observed in kidneysinjected with bFGF is superior to that observed in kidneys injected with saline. The Smads proteins show an increase with a maximum at 48–72 h and decreasedat 96 h after ischemia in kidneys injected with saline or bFGF (n � 5 for each I/R).
R1681EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

Levels of markers for differentiation on ATN kidneys. Tostudy the effect of ATN induced by I/R on kidneys injectedwith bFGF or saline, freshly prepared extracts from inner andouter medulla kidney sections were analyzed with the corre-sponding antibodies. The Western blots were run with the totalsamples (n � 5) in each time period, and the selected blotcorresponds to a representative one from the group. Comparedwith kidney homogenates from I/R injected with saline, kid-neys from I/R rats treated with bFGF showed an increase ofthese proteins and an early expression in time, similar to thatobserved by immunohistochemistry in proximal tubular cells(Fig. 3.1–3.4) (P � 0.05). Twenty-four hours after I/R, in-creased levels of HIF-1�, VEGF, bFGF, Ncam, Pax-2, Vimen-tin, Noggin, Tie-2, BMP-7, Engrailed, and Lim-1 levels wereobserved. This staining peaked at 48 h after I/R and wasmaintained for 72–96 h after I/R (Fig. 3.1–3.4). In addition,Smad and p-Smad levels peaked 48 h after I/R (Fig. 3.4, M andN), 72 h after I/R. The intensity of the bands declined, and theywere not detectable at 96 h (Fig. 3.4, M and N).
DISCUSSION
Embryonic kidney development is characterized by prolif-eration of undedifferentiated cells and later differentiation ofdaughter cells into specific cell phenotype. A similar sequenceof events can be observed during the regeneration process,
opening the possibility that renal regeneration may recapitulatepart of the kidney genetic program, during organogenesis,including apoptosis (2, 57). If this hypothesis is correct, wecould use morphogenes to induce the repairing process orprevent damage in I/R kidney. One morphogen is bFGF, andprevious reports have shown that it participates in the regen-eration process in the retina (66), angiogenesis (3), myocardialinfarction (38), and ischemic heart (33), where a rapid revas-cularization was observed after ischemia. In this study, weevaluated the effect of the morphogen bFGF on the repair ofkidney damage.
Apoptosis is a programmed mode of cell death that plays animportant role in the pathogenesis of the renal ischemia (39).Caspases play a key role in the mammalian apoptotic mecha-nism, caspase-3 being a prototypical component of this mech-anism. Together with this analysis, the presence of importantmarkers induced by damage, such as macrophages (cloneED-1) and interstitial �-SMA is an excellent method to eval-uate the cellular morphology induced by ischemia. We haveshown here that caspase-3 was activated in kidneys with I/R, asjudged by a increase in its immunoreactivity and a higherdegree of colocalization with TUNEL-positive cells in a time-dependent manner. The expression of TUNEL and caspasespeaked at 24 h after I/R. Additionally, PAS staining indicatedthat ischemic tubular cells showed apoptotic features, as ob-
Fig. 4. Immunolocalization of endothelial cell markers in kidney regeneration. Maximum staining intensity for HIF-1�, (A and D), VEGF (B and E), and Tie-2(C and F) was observed at 48 h after 30 min of ischemia in kidneys injected with bFGF (D–F). The staining for each marker was also performed at similar timeperiods in kidneys injected with saline (A–C) (n � 5 for each group). The expression of these markers was observed in papillary collecting ducts, thick ascendinglimb, and proximal tubular cells localized mainly in the inner and outer medulla of kidney. Staining HIF-1� was done using peroxidase and was revealed withDAB (brown color reaction); the staining for VEGF and Tie-2 was done using alkaline phosphatase and was revealed with nitro blue tetrazolium chloride:5-bromo-4chloro-3indolyl phosphate (NBT/BCIP; p-toluidine salt blue color reaction). Scale bar � 100 �m. The arrows indicate the localization of thecorresponding marker.
R1682 EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

Fig. 5. Immunolocalization of MM and early epithelial markers in kidney regeneration. Kidney slices were immunostained for bFGF (A and E), Ncam (B andF), Pax 2–5-8 (C and G), and Vimentin (D and H) 48 h after 30 min of ischemia in kidneys injected with bFGF (E–H). Kidney slices from rats injected withsaline (A–D) were also stained for each marker (n � 5 for each group). The MM markers Vimentin and Ncam were localized in the peritubular area in the innerand outer medulla, and the early epithelial proteins Pax-2 and bFGF were localized in the proximal tubular cells in the inner and outer medulla. Staining for allfour markers was increased after ischemia in kidneys injected with bFGF (E–H) vs. kidney injected with saline (A–D), with a maximum at 24 h. Staining forbFGF, Ncam, Pax 2–5-8, and Vimentin was done using peroxidase and was revealed with DAB (brown color reaction). Scale bar � 100 �m. The arrows indicatethe localization of the corresponding marker.
Fig. 6. Kidneys injected with saline were stained for BMP-7, Engrailed, Lim 1�2, and zone occludens-1 (ZO-1; A–D) (n � 5 for each group).Immunolocalization of epithelial and tubular markers in kidney regeneration. Maximum staining intensity for BMP-7 (E), Engrailed (F), Lim 1�2 (G), and ZO-1(H) was observed 48 h after 30 min of ischemia in kidneys injected with bFGF. Engrailed and Lim1�2 were localized in the peritubular area, and BMP-7 andZO-1 were localized in the proximal tubular cells from the inner and outer medulla. Staining for epithelial and tubular proteins increased, with a maximum at48 h after ischemia in kidney injected with bFGF (E–H) vs. kidney injected with saline (A–D). Staining for BMP-7, Engrailed, Lim 1�2, and ZO-1 was doneusing peroxidase and was revealed with DAB (brown color reaction). Scale bar � 100 �m. The arrows indicate the localization of the corresponding marker.
R1683EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

served with PAS staining, and the strong presence of ED-1 and�-SMA was strongly present at 24–48 h after I/R, a timeperiod that is delayed when compared with the upregulation ofTUNEL and caspase-3. These data are in agreement withprevious reports on ischemic kidneys and neurons, in whichmorphological changes were shown several hours aftercaspase-3 upregulation (39, 42, 57). However, for bFGF-treated kidney with I/R, we observed a marked reduction in thenumber of tubular cell TUNEL and caspase-3-positive cellsafter 24 h. Immunoreactivity was not observed, and the PASstaining showed normal morphological characteristics in tubu-lar cells at 48 h after I/R, coincidently with the disappearanceof damage markers ED-1 and �-SMA, which presented animmunohistochemistry similar to what is observed in a normalkidney. It is noteworthy that an important number of mitoseswas observed and maintained at 72 h after I/R, indicating thatan important proliferating process is maintained after the ap-parent recovery of renal morphology. These results show thatbFGF may prevent tubular apoptosis triggered by ARF, similarto previous reports for other morphogenes, such as BMP-7(65). Coincidentally, in both cases, where the kidneys weretreated with bFGF or saline, the TUNEL and caspase-3-positive staining decrease when the reparation process began,expressing morphogenic and epitheliogenic proteins (57).
Important proteins in the regeneration process are thoseinduced by hypoxia. Hypoxia has been described (2) as beingcapable of inducing the expression of specific proteins in-volved in the repair process. Our results clearly show thepresence of hypoxia at the initial time of I/R in the same areasthat later express morphogenic and epitheliogenic proteins;
however, this expression is different in kidneys treated withbFGF or saline. The induction of HIF-1� is an early event inthe sequence of cellular changes after the interruption of bloodflow, and it is likely to play an important role for initiatingsubsequent reactions. In kidneys injected with saline, HIF-1�induction was visible after 30 min of I/R and was mainlyconfined to a period of 48–72 h, similar to what was reportedpreviously (44, 57), whereas in kidneys treated with bFGF,HIF-1� was observed with a higher intensity at 24–48 h afterI/R and declined at 72–96 h. Because the area of HIF-1�expression was overlapping with areas in which we foundincreased nephrogenic and epitheliogenic proteins (57), it ispossible, that HIF-1� plays an important role in cell deathand/or survival decision, inducing the cell to express morpho-genic proteins, and it is possible that pretreating kidney cellswith bFGF can harness the effect induced by HIF-1�.
It is very important to consider that even though ARF hasbeen closely linked to tubular epithelial cell injury, an impor-tant vascular element is also involved, as observed in thedamage of peritubular capillaries in rats subjected to renalischemia (5). An example is the distorted integrity of endothe-lial layers by desquamating or retracting cells (8) or an earlyswelling with narrowing of the vascular lumen (40). For thisreason, ARF can affect the function of the renal endothelium,resulting in prolonged renal hypoperfusion (40). Consideringthese data, we analyzed the expression of VEGF and Tie-2,known to be regulated by HIF-1� in an oxygen-dependentfashion. VEGF is essential for endothelial cell differentiation(vasculogenesis) and for the sprouting of new capillaries frompreexisting vessels (angiogenesis); the evidence has shown that
Fig. 7. Immunolocalization of transcription way BMP-7. Kidney slices from kidneys injected with saline were also stained for BMP-7, Noggin, Smad, andp-Smad (A–D) (n � 5 for each group). Maximum staining intensity for BMP-7 (E), Noggin (F), Smad (G), and p-Smad (H) was observed at 48 h after 30 minof ischemia in kidneys injected with bFGF. Staining for BMP-7 and Noggin proteins increased at 48 h after ischemia and were localized to the proximal tubularcells from the inner and outer medulla. Staining for Smad proteins increased with a maximum at 24–48 h and was localized to the proximal tubular cells fromthe inner and outer medulla. Staining for BMP-7 and Noggin was done using peroxidase and was developed with DAB (brown color reaction); the staining forSmad and p-Smad was done using alkaline phosphatase and was developed with NBT/BCIP (blue color reaction). Scale bar � 100 �m. The arrows indicate thelocalization of the corresponding marker for each figure.
R1684 EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

VEGF is a survival factor that allows various cell types tosurvive and proliferate under conditions of extreme stress suchas hypoxia (24). For this reason, hypoxia induced by I/R is akey regulator of VEGF gene expression and has an importantrole in the vascular response to kidney ischemia (17). WhereasVEGF has a beneficial role in the pathogenesis of somediseases, it produces harmful effects in others (24). In thisstudy in kidneys with I/R injected with saline, VEGF wasreexpressed in the regenerating proximal tubules and peaked at48 h after I/R, whereas in kidneys treated with bFGF, aconsiderable increase in expression was observed at 24–48 hand was maintained at 72–96 h. Because of the considerableimpact of VEGF on angiogenesis, it may be an importantcontributor to the reestablishment of epithelial and endothelialcell integrity after kidney damage, and it is possible that it canaccelerate the regeneration process because of its early in-crease, and so recover before the endothelial function. Theangiopoietin receptor Tie-2 plays an important role in nephro-genesis, angiogenesis, and stabilization of vascular integrity. Inour study in kidneys injected with saline, we observed areexpression of Tie-2 at 48 h after injury induced by I/R,whereas the treatment with bFGF induced Tie-2, 24 h after I/Rand increased its amount. Recent studies have shown thepresence of endothelial progenitor cells Tie-2 positives, 7 daysafter of I/R (40). In this study, we evaluated the presence intubular cells; because Tie-2 has a role in vascular growth in theearly stages of mammalian nephrogenesis, we postulated thatin the kidney regeneration process, Tie-2 could reestablish theintegrity of interstitial and glomerular vessels. The differentialexpression of VEGF and Tie-2 reported previously is notobserved in kidneys treated with bFGF, and the expression ofboth proteins is observed at the same time, 24 h after I/R. It canbe explained, in part, by the high requirements for VEGF andTie-2 to accelerate the regeneration process induced by bFGFtreatment; see Rabie and Lu (43) for more details on theupregulation of VEGF by bFGF.
The results obtained indicate that renal adult cells have thecapacity of reexpressing specific proteins of kidney develop-ment during recovery from a transient episode of ischemia, andthis process can be accelerated by the treatment with bFGF asa specific morphogen, suggesting that these cells participate inrenal repair. bFGF, Ncam, BMP-7, Lim-1, Engrailed, andZO-1 are known to play a crucial role during early metanephrickidney development (20). After kidney injury induced by I/R,these proteins were locally restricted and reexpressed in theregenerating proximal tubules. In kidneys injected with saline,this expression was limited to a time interval of 24 to 72 h,peaking 24–48 h after I/R for bFGF and Ncam and 48 h afterI/R for BMP-7, Lim-1, Engrailed, and ZO-1. These results aresimilar to others reported previously in kidney development(20, 57). In kidneys treated with bFGF, these morphogeneswere all strongly induced at 24 h after I/R and peaked at 48 h;however, the amount induced at 24 h is comparable to theamount induced at 48 h in kidneys treated with saline, indicat-ing that induction of these morphogens is increased and pre-viously expressed in time. This confirms previous reports thathave shown an upregulation of BMP-7, Lim, and Engrailed byFGF (34, 37). In addition, the expression in proximal tubularcells declined after reconstitution of the tubule. This transientreexpression is similar to the one reported for Pax-2 (20).
Vimentin is a marker of mesenchymal cells and therefore isa marker of fully dedifferentiated renal epithelia. It is notpresent in healthy adult renal tubules but its reexpressionoccurs during tubular regeneration and proliferation (63). Theearly presence and high increase of this protein in our results inkidneys treated with bFGF shows that these cells acquire asimilar state of differentiation to the one in early development,mimicking this process in the regeneration phase after I/R inATN. Recent reports have shown that the same intrarenal cellsare the major source for regeneration in postischemic kidneys(32); in this regard, we analyzed the presence of hematopoieticstem cells CD34 positive in kidneys after I/R and could notobserve the presence of this cellular type in the time framestudied, similar to what was reported previously (14), andconsidering the number of mitoses in tubular cells, we believethat the same tubular cells are the ones that are dedifferentiat-ing to repair kidney damage. These examples are consistentwith the hypothesis that during tissue regeneration a cascade ofdevelopmental gene pathways may be reactivated.
We analyzed the BMP pathway because of the importantinteraction between BMP and FGF described in early devel-opment. In this regard, we observed an important reexpressionof BMP-7 and its antagonist Noggin 48 and 24 h, respectively,after I/R in kidneys injected with saline; however, in kidneystreated with bFGF, both were strongly expressed at 24 h afterI/R and maintained in time up to 96 h after I/R. The relevanceof these data is related to the regulatory function of Noggin andBMP. Noggin could be generating specific signals to determineone particular cell type, or it could make cell groups sensitiveto a specific morphogen, such as BMP. Furthermore, thisindicates that Noggin would be involved in kidney develop-ment and in the regeneration process after kidney damage. Inaddition, previous reports have shown the upregulation ofNoggin, BMP, and their transcription pathway Smads by FGF(11, 26, 16, 37).
Recent reports have shown that BMP signaling, mediated bySmad proteins, is important during kidney development (61).The spatial and temporal expression patterns of the Smads havebeen described in mesenchymal cells of the nephrogenic zoneduring kidney development (61), in which Smads are down-regulated once these cells begin to epithelialize. We observedSmad and p-Smad (the phosphorylated form of Smad) arereexpressed after damage in kidneys treated with bFGF orsaline, reaching a maximum at 24–48 h after I/R, at whichpoint, we observed the presence of nephrogenic proteins andthe highest levels of Noggin and BMP. Its expression waslower at 72–96 h after I/R, coincident with the high levels ofepitheliogenic proteins, similar to what was reported in embryo(57). On the basis of the observed patterns of expression, wespeculate that one individual or a combination of Smads couldplay specific roles in the early regeneration phase of ATNduring kidney damage and be involved in nephrogenic process,in a similar way to the one described in kidney development.
In summary, our results suggest that during the regenerationprocesses, bFGF can be reexpressed to restore mature kidneyfunction, in a process similar to the one described in nephro-genesis during embryonic development and suggest that theregeneration process can be induced earlier by the treatmentwith this growth factor.
R1685EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

ACKNOWLEDGMENTS
The authors thank Maria Alcoholado for technical assistance in tissueprocessing, Dr. Victoria Velarde, and Dr. Ricardo Moreno for critical readingof the manuscript.
GRANTS
This work was supported by Fondecyt Grant 3050075 (to S. Villanueva)and 1050977 (to C. P. Vio).
REFERENCES
1. Abbate M, Brown D, and Bonventre J. Expression of NCAM recapit-ulates tubulogenic development in kidneys recovering from acute ische-mia. Am J Physiol Renal Physiol 277: F454–F463, 1999.
2. Al-awqati Q and Oliver JA. Stem cells in the kidney. Kidney Int 61:387–395, 2002.
3. Aviles R, Annex B, and Lederman R. Testing clinical therapeuticangiogenesis using basic fibroblast growth factor (FGF-2). Br J Pharma-col 140: 637–646, 2003.
4. Bardella L and Comolli R. Differential expression of c-jun, c-fos and hsp70 mRNAs after folic acid and ischemia-reperfusion injury: Effect ofantioxidant treatment. Exp Nephrol 2: 158–165, 1994.
5. Basile DP, Donohoe D, Roethe K, and Osborn JL. Renal ischemic injuryresults in permanent damage to peritubular capillaries and influences long-term function. Am J Physiol Renal Physiol 281: F887–F899, 2001.
6. Basile DP, Fredrich K, Alausa M, Vio CP, Liang M, Rieder MR,Greene AS, and Cowley Jr AW. Identification of persistently alteredgene expression in kidney following functional recovery from ischemicacute renal failure. Am J Physiol Renal Physiol 288: F253–F263, 2005.
7. Breier G, Albrecht U, Sterrer S, and Risau W. Expression of vascularendothelial growth factor during embryonic angiogenesis and endothelialcell differentiation. Development 114: 521–532, 1992.
8. Brodsky SV, Yamamoto T, Tada T, Kim B, Chen J, Kajiya F, andGoligorsky MS. Endothelial dysfunction in ischemic acute renal failure:rescue by transplanted endothelial cells. Am J Physiol Renal Physiol 282:F1140–F1149, 2002.
9. Brunet LJ, McMahon JA, McMahon AP, and Harland RM. Noggin,cartilage morphogenesis, and joint formation in the mammalian skeleton.Science 280: 1455–1457, 1998.
10. Cai Q, Dmitrieva N, Ferraris J, Brooks H, van Balkom B, and BurgM. Pax2 expression occurs in renal medullary epithelial cells in vivo andin cell culture, is osmoregulated, and promotes osmotic tolerance. ProcNatl Acad Sci USA 102: 503–508, 2005.
11. Chiba S, Kurokawa MS, Yoshikawa H, Ikeda R, Takeno M, TadokoroM, Sekino H, Hashimoto T, and Suzuki N. Noggin and basic FGF wereimplicated in forebrain fate and caudal fate, respectively, of the neuraltube-like structures emerging in mouse ES cell culture. Exp Brain Res 163:86–99, 2005.
12. Dudley AT, Lyons KM, and Robertson EJ. A requirement for bonemorphogenetic protein-7 during development of the mammalian kidneyand eye. Genes Dev 9: 2795–2807, 1995.
13. Dudley AT, Godin RE, and Robertson EJ. Interaction between FGF andBMP signaling pathways regulates development of metanephric mesen-chyme. Genes Dev 13: 1601–1613, 1999.
14. Duffield JS, Park KM, Hsiao LL, Kelley VR, Scadden DT, IchimuraT, and Bonventre JV. Restoration of tubular epithelial cells during repairof the postischemic kidney occurs independently of bone marrow-derivedstem cells. J Clin Invest 115: 1743–1755, 2005.
15. Esson ML and Schrier RW. Diagnosis and treatment of acute tubularnecrosis. Ann Intern Med 137: 744–752, 2002.
16. Fakhry A, Ratisoontorn C, Vedhachalam C, Salhab I, Koyama E,Leboy P, Pacifici M, Kirschner RE, and Nah HD. Effects of FGF-2/-9in calvarial bone cell cultures: differentiation stage-dependent mitogeniceffect, inverse regulation of BMP-2 and noggin, and enhancement ofosteogenic potential. Bone 36: 254–266, 2005.
17. Freeburg PB and Abrahamson DR. Hypoxia-inducible factors andkidney vascular development. J Am Soc Nephrol 14: 2723–2730, 2003.
18. Goda N, Dozier SJ, and Johnson RS. HIF-1 in cell cycle regulation,apoptosis, and tumor progression. Antioxid Redox Signal 5: 467–473,2003.
19. Harlow E and Lane D. Antibodies: A Laboratory Manual. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory, 1998.
20. Imgrund M, Grone E, Grone HJ, Kretzler M, Holzman L, Schlon-dorff D, and Rothenpieler UW. Re-expression of the developmental
gene Pax-2 during experimental acute tubular necrosis in mice. Kidney Int56: 1423–1431, 1999.
21. Joannidis M, Cantley LG, Spokes K, Stuart-Tilley AK, Alper SL, andEpstein FH. Modulation of c-fos and egr-1 expression in the isolatedperfused kidney by agents that alter tubular work. Kidney Int 52: 130–139,1997.
22. Jones N, Voskas D, Master Z, Sarao R, Jones J, and Dumont DJ.Rescue of the early vascular defects in Tek/Tie2 null mice reveals anessential survival function. EMBO J 2: 438–445, 2001.
23. Karanova I, Dove L, Resau J, and Perantoni A. Conditioned mediumfrom a rat ureteric bud cell line in combination with bFGF inducescomplete differentiation of isolated metanephric mesenchyme. Develop-ment 122: 4159–4167, 1996.
24. Kim BS and Goligorsky MS. Role of VEGF in kidney development,microvascular maintenance and pathophysiology of renal disease. KoreanJ Intern Med 18: 65–75, 2003.
25. Kitamoto Y, Tokunaga H, and Tomita K. Vascular endothelial growthfactor is an essential molecule for mouse kidney development: glomeru-logenesis and nephrogenesis. J Clin Invest 99: 2351–2357, 1997.
26. Khot S and Ghaskadbi S. FGF signaling is essential for the early eventsin the development of the chick nervous system and mesoderm. Int J DevBiol 45: 877–885, 2001.
27. Kolatsi-Joannou M, Li XZ, Suda T, Yuan HT, and Woolf AS. Expres-sion and potential role of angiopoietins and Tie-2 in early development ofthe mouse metanephros. Dev Dyn 222: 120–126, 2001.
28. Kopp JB. BMP receptors in kidney. Kidney Int 58: 2237–2238, 2000.29. Lameire NH and Vanholder R. Pathophysiology of ischaemic acute
renal failure. Best Pract Res Clin Anaesthesiol 18: 21–36, 2004.30. Lazarous DF, Unger EF, Epstein SE, Stine A, Arevalo JL, Chew EY,
and Quyyumi AA. Basic fibroblast growth factor in patients with inter-mittent claudication: results of a phase I trial. J Am Coll Cardiol 36:1239–1244, 2000.
31. Lederman RJ, Mendelsohn FO, Anderson RD, Saucedo JF, TenagliaAN, Hermiller JB, Hillegass WB, Rocha-Singh K, Moon TE, White-house MJ, and Annex BH. Therapeutic angiogenesis with recombinantfibroblast growth factor-2 for intermittent claudication (the TRAFFICstudy): a randomised trial. Lancet 359: 2053–2058, 2002.
32. Lin F, Moran A, and Igarashi P. Intrarenal cells, not bone marrow-derived cells, are the major source for regeneration in postischemickidney. J Clin Invest 115: 1756–1764, 2005.
33. Lutter G, Attmann T, Heilmann C, von Samson P, von Specht B, andBeyersdorf F. The combined use of transmyocardial laser revasculariza-tion (TMLR) and fibroblastic growth factor (FGF-2) enhances perfusionand regional contractility in chronically ischemic porcine hearts. EurJ Cardiothorac Surg 22: 753–761, 2002.
34. Mayordomo R and Alvarez IS. Vertical regulation of En-2 expressionand eye development by FGFs and BMPs. J Craniofac Genet Dev Biol 20:64–75, 2000.
35. Mehta R. Outcomes research in acute renal failure. Semin Nephrol 23:283–294, 2003.
36. Morrisey M, Hruska K, Guo G, Wang S, Chen Q, and Klahr S. Bonemorphogenetic protein-7 improves renal fibrosis and accelerates the returnof renal function. J Am Soc Nephrol 13: S14–S21, 2002.
37. Nakamura Y, Tensho K, Nakaya H, Nawata M, Okabe T, andWakitani S. Low-dose fibroblast growth factor-2 (FGF-2) enhances bonemorphogenetic protein-2 (BMP-2)-induced ectopic bone formation inmice. Bone 36: 399–407, 2005.
38. Nishida S, Nagamine H, Tanaka Y, and Watanabe G. Protective effectof basic fibroblast growth factor against myocyte death and arrhythmias inacute myocardial infarction in rats. Circ J 67: 334–339, 2003.
39. Oberbauer R, Schwarz C, Regele HM, Hansmann C, Meyer TW, andMayer G. Regulation of renal tubular cell apoptosis and proliferation afterischemic injury to a solitary kidney. Lab Clin Med 138: 343–351, 2001.
40. Patschan D, Krupincza K, Patschan S, Zhang Z, Hamby C, andGoligorsky MS. Dynamics of mobilization and homing of endothelialprogenitor cells after acute renal ischemia: modulation by ischemic pre-conditioning. Am J Physiol Renal Physiol 291: F176–F185, 2006.
41. Perantoni AO, Dove LF, and Karavanova I. Basic fibroblast growthfactor can mediate the early inductive events in renal development. ProcNatl Acad Sci USA 92: 4696–4700, 1995.
42. Qi JP, Wu AP, Wang DS, Wang LF, Li SX, and Xu FL. Correlationbetween neuronal injury and caspase-3 after focal ischemia in humanhippocampus. Chin Med J (Engl) 117: 1507–1512, 2004.
R1686 EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from

43. Rabie AB and Lu M. Basic fibroblast growth factor up-regulates theexpression of vascular endothelial growth factor during healing of allo-geneic bone graft. Arch Oral Biol 49: 1025–1033, 2004.
44. Rosenberger C, Mandriota S, Jurgensen JS, Wiesener MS, HorstrupJH, Frei U, Ratcliffe PJ, Maxwell PH, Bachmann S, and Eckardt KU.Expression of hypoxia-inducible factor-1alpha and -2alpha in hypoxic andischemic rat kidneys. J Am Soc Nephrol 13: 1721–1732, 2002.
45. Rothenpieler UW and Dressler GR. Pax-2 is required for mesenchyme-to-epithelium conversion during kidney development. Development 119:711–720, 1993.
46. Safirstein R. Gene expression in nephrotoxic and ischemic acute renalfailure. J Am Soc Nephrol 4: 1387–1395, 1994.
47. Schrier RW, Wang W, Poole B, and Mitra A. Acute renal failure:definitions, diagnosis, pathogenesis, and therapy. J Clin Invest 114: 5–14,2004.
48. Singri N, Ahya SN, and Levin ML. Acute renal failure. JAMA 289:747–751, 2003.
49. Sakurai H, Barros EJ, Tsukamoto T, Barasch J, and Nigam SK. An invitro tubulogenesis system using cell lines derived from the embryonickidney shows dependence on multiple soluble growth factors. Proc NatlAcad Sci USA 94: 6279–6284, 1997.
50. Stanley E, Biben C, Kotecha S, Fabri L, Tajbakhsh S, Wang CC,Hatzistavrou T, Roberts B, Drinkwater C, Lah M, Buckingham M,Hilton D, Nash A, Mohun T, and Harvey RP. DAN is a secretedglycoprotein related to Xenopus cerberus. Mech Dev 77: 173–184, 1998.
51. Stanton B and Koeppen B. The kidney. In: Physiology (5th ed.), editedby Berne R, Levi M, Koeppen B, and Stanton B. St. Louis, MO: Mosby,2004.
52. Strutz F, Zeisberg M, Ziyadeh FN, Yang CQ, Kalluri R, Muller GA,and Neilson EG. Role of basic fibroblast growth factor-2 in epithelialmesenchymal transformation. Kidney Int 61: 1714–1728, 2002.
53. Supavekin S, Zhang W, Kucherlapati R, Kaskel FJ, Moore LC, andDevarajan P. Differential gene expression following early renal ische-mia/reperfusion. Kidney Int 63: 1714–1724, 2003.
54. Unger EF, Goncalves L, Epstein SE, Chew EY, Trapnell CB, Cannon3rd RO, and Quyyumi AA. Effects of a single intracoronary injection ofbasic fibroblast growth factor in stable angina pectoris. Am J Cardiol85:1414–1419, 2000.
55. Venkatachalam MA, Rennke HG, and Sandstrom DJ. The vascularbasis for acute renal failure in the rat. Preglomerular and postglomerularvasoconstriction. Circ Res 38: 267–279, 1976.
56. Villanueva S, Glavic A, Ruiz P, and Mayor R. Posteriorization by FGF,Wnt and retinoic acid is required for neural crest induction. Dev Biol 241:289–301, 2002.
57. Villanueva S, Cespedes C, and Vio CP. Ischemic acute renal failureinduces the expression of a wide range of nephrogenic proteins. Am JPhysiol Regul Integr Comp Physiol 290: R861–R870, 2006.
58. Vio CP, An SJ, Cespedes C, McGiff JC, and Ferreri NR. Induction ofcyclooxygenase-2 in thick ascending limb cells by adrenalectomy. J AmSoc Nephrol 12: 649–658, 2001.
59. Vio CP, Cespedes C, Gallardo P, and Masferrer JL. Renal identifica-tion of cyclooxygenase-2 in a subset of thick ascending limb cells.Hypertension 30: 687–692, 1997.
60. Vio CP and Figueroa CD. Evidence for a stimulatory effect of highpotassium diet on renal kallikrein. Kidney Int 31: 1327–1334, 1987.
61. Vrljicak P, Myburgh D, Ryan AK, Van Rooijen MA, Mummery CL,and Gupta IR. Smad expression during kidney development. Am JPhysiol Renal Physiol 286: F625–F633, 2004.
62. Wagner KD, Wagner N, Wellmann S, Schley G, Bondke A, Theres H,and Scholz H. Oxygen-regulated expression of the Wilms tumor suppres-sor Wt1 involves hypoxia-inducible factor-1 (HIF-1). FASEB J 17: 1364–1366, 2003.
63. Witzgall R, Brown D, Schwarz C, and Bonventre JV. Localization ofproliferating cell nuclear antigen, vimentin, c-Fos, and clusterin in thepostischemic kidney. Evidence for a heterogeneous genetic responseamong nephron segments, and a large pool of mitotically active anddedifferentiated cells. J Clin Invest 93: 2175–2188, 1994.
64. Yoshida T, Kurella M, Beato F, Min H, Ingelfinger JR, Stears RL,Swinford RD, Gullans SR, and Tang SS. Monitoring changes in geneexpression in renal ischemia-reperfusion in the rat. Kidney Int 61: 1646–1654, 2002.
65. Zeisberg M, Shah AA, and Kalluri R. Bone morphogenic protein-7induces mesenchymal to epithelial transition in adult renal fibrosis andfacilitates regeneration of injured kidneys. J Biol Chem 280: 8094–8100,2005.
66. Zhang Q, Wang G, and Sun J. Pharmacokinetics of recombinant humanbasic fibroblast growth factor in rabbits and mice serum and rabbitsaqueous humor. Acta Pharmacol Sin 25: 991–995, 2004.
R1687EARLY MORPHOGEN EXPRESSION INDUCED BY bFGF
AJP-Regul Integr Comp Physiol • VOL 291 • DECEMBER 2006 • www.ajpregu.org
by 10.220.33.5 on August 27, 2016
http://ajpregu.physiology.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















