
Morphogenic effects of abiotic stress: reorientation of growth in seedlings
Upload
nottinghamCategory
view
0download
0
lable at ScienceDirect
Biomaterials 31 (2010) 1242–1250
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
The effect of the delivery of vascular endothelial growth factor and bonemorphogenic protein-2 to osteoprogenitor cell populations on bone formation
Janos M. Kanczler a, Patrick J. Ginty b, Lisa White b, Nicholas M.P. Clarke c, Steven M. Howdle d,Kevin M. Shakesheff b, Richard O.C. Oreffo a,*
a Bone & Joint Research Group, Centre for Human Development, Stem Cells and Regeneration, Developmental Origins of Health and Disease, Institute of Developmental Sciences,University of Southampton, General Hospital, Southampton, SO16 6YD, UKb School of Pharmaceutical Sciences, University of Nottingham, Nottingham, UKc University Orthopaedics, Southampton General Hospital, Southampton SO16 6YD, UKd School of Chemistry, University of Nottingham, University Park Nottingham, UK
a r t i c l e i n f o
Article history:Received 10 August 2009Accepted 26 October 2009Available online 18 November 2009
Keywords:Bone tissue regenerationHuman bone marrow stromal cellsSupercritical CO2
Critical sized femur defectVEGFBMP-2
* Corresponding author. Tel.: þ44 2380 798502; faE-mail address: [email protected] (R.O.C. Oreffo).URL: http://www.mesenchymalstemcells.org
0142-9612/$ – see front matter � 2009 Elsevier Ltd.doi:10.1016/j.biomaterials.2009.10.059
a b s t r a c t
Regenerating bone tissue involves complex, temporal and coordinated signal cascades of which bonemorphogenic protein-2 (BMP-2) and vascular endothelial growth factor (VEGF165) play a prominent role.The aim of this study was to determine if the delivery of human bone marrow stromal cells (HBMSC)seeded onto VEGF165/BMP-2 releasing composite scaffolds could enhance the bone regenerative capa-bility in a critical sized femur defect. Alginate-VEGF165/PDLLA-BMP-2 scaffolds were fabricated usinga supercritical CO2 mixing technique and an alginate entrapment protocol. Increased release of VEGF165
(750.4� 596.8 rg/ml) compared to BMP-2 (136.9� 123.4 rg/ml) was observed after 7-days in culture.Thereafter, up till 28 days, an increased rate of release of BMP-2 compared to VEGF165 was observed. Thealginate-VEGF165/PDLLA-BMP-2þHBMSC group showed a significant increase in the quantity of regen-erated bone compared to the alginate-VEGF165/PDLLA-BMP-2 and alginate/PDLLA groups respectively ina critical sized femur defect study as indices measured by mCT. Histological examination confirmedsignificant new endochondral bone matrix in the HBMSC seeded alginate-VEGF165/PDLLA-BMP-2 defectgroup in comparison to the other groups. These studies demonstrate the ability to deliver a combinationof HBMSC with angiogenic and osteogenic factors released from biodegradable scaffold compositesenhances the repair and regeneration of critical sized bone defects.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Segmental bone defects and non-unions resulting from trauma,resection or pathology represent significant clinical challengesworld-wide, for the orthopaedic, reconstructive and maxillofacialsurgeons [1,2]. To date, bone grafting techniques that use materialssuch as autografts, allografts or metallic implants face considerablelimitations for bone repair due to the lack of available bone tissue,disease transmission, cell-mediated immune responses, andsignificant costs. These difficulties have resulted in the search foralternative bone graft substitutes to be used in repairing non-unionfractures, spinal fusions and bone tumour resections.
The development and regeneration of natural bone tissue isa complex, coordinated temporal process involving a myriad of
x: þ44 2380 5525.
All rights reserved.
molecular, cellular, biochemical and mechanical cues [3]. Advancesin the field of regenerative medicine have led to the possibility ofsuccessfully repairing and restoring the function in damaged ordiseased tissues [4,5]. The release of growth factors and osteopro-genitor cell populations by biocompatible scaffolds have been shownto enhance the regenerative capacity of bone [6–8] as a consequence,in part, of replicating the natural bone environment by provision ofan appropriate synthetic extracellular matrix scaffolds [9], presen-tation of growth factors [10] and osteoprogenitor cells. The presen-tation of these cells and cues, at appropriate time intervals offers, theopportunity to mimic the repair sequences in bone.
However, critical for regenerative tissue is the generation andexistence of a functional microvascular network within the engi-neered constructs to provide oxygen and nutrients to facilitategrowth, differentiation, and tissue functionality [3,11]. Thus, thelack of an adequate microvascular network will result in thehypoxic cell death of the implanted engineered tissue. Factors thatare involved in angiogenesis are also fundamental to the osteogenic

J.M. Kanczler et al. / Biomaterials 31 (2010) 1242–1250 1243
response in fracture healing. VEGF165, a potent growth factorinvolved in angiogenesis has been extensively investigated andshown to be involved in osteogenesis and bone repair [12]. Duringendochondral bone formation VEGF165 modulates angiogenesis,chondrocyte apoptosis, cartilage remodelling, osteoblast migrationand endochondral growth plate ossification [13–16]. Moreover,BMP-2 has been shown to upregulate VEGF165 [17]. Conversely,inhibition of angiogenesis prevents fracture healing [18] and inhi-bition of VEGF165 decreases angiogenesis, callus mineralisation andbone healing [19].
A number of studies have shown that the presentation of indi-vidual growth factors encapsulated in scaffolds lead to successfulengineering of bone like constructs. Indeed the release of activeBMP-2 & VEGF165 respectively from polymer scaffolds such as PDLLAhas been shown to induce HBMSC osteogenesis [6,20]. However,mimicking the signalling cascade, release and presentation of thesegrowth factors in a fracture repair by developing composite scaf-folds that can release a number of growth factors at varying kineticrates in situ will, ultimately, enhance the regenerative capacity ofbone. Recently, scaffolds have been developed to release dual setsof growth factors for regeneration of tissue [10]. Simmons et al. [21]showed that TGF-b3 and BMP-2 released from a scaffold andtransplanted bone marrow stromal cells enhanced bone formationin vivo. Additionally, Huang et al. [7] fabricated Poly(lactic-co-gly-colic acid) scaffolds which were able to release VEGF and a plasmidDNA encoding BMP-4 resulting in the generation of ectopic bone byHBMSC within these scaffolds when implanted into the subcuta-neous tissue of SCID mice.
The presentation of multiple growth factors released at differentkinetic rates from 3D biodegradable scaffolds within close prox-imity of osteoprogenitor cells offers an attractive strategy for theaugmentation and repair of bone defects. Using a supercritical CO2
and alginate entrapment technique, composite scaffolds consistingof alginate and poly(D,L-lactic acid) (PLA) were generated to allowdifferent biodegradation rates and release of selected factors atdifferent rates [22]. For this study VEGF165 was encapsulated in thealginate fibres and BMP-2 was encapsulated into PDLLA inducing anaccelerated rate of release of VEGF165 and slower rate of release ofBMP-2. In this current study we have used a critical sized femurdefect model (load–bearing) to investigate the potential boneregeneration capacity of such a combination therapy of humanbone marrow stromal cells (HBMSC) transplanted onto thecomposite alginate-VEGF165/PDLLA-BMP-2 scaffolds.
2. Methods and materials
2.1. Materials
Recombinant human BMP-2 was kindly provided by Professor Walter Sebald,University of Wurzburg, Germany. Recombinant Human VEGF165 was purchasedfrom Tebu-bio, UK. Vibrant� carboxy fluorescein diacetate, succinimidyl ester (CFDASE) cell tracer kit and Ethidium Homodimer-1 were purchased from MolecularProbes, UK. BMP-2 and VEGF165 ELISA kits were purchased from R&D Systems, UK.Phosphate Buffered Saline (PBS) was purchased from Lonza Biologics, UK; endo-thelial cell culture growth supplement was purchased from Promocell, Germany.Fetal calf serum was purchased from Invitrogen, UK. Poly D,L-lactic acid (Mw
w65 kDa was supplied by Lakeshore Biomaterials (Birmingham, Alabama, US). Allother materials were obtained form Sigma–Aldrich, UK unless stated.
2.2. Cell culture
Adult human bone marrow samples were obtained from hematological normalpatients undergoing routine elective hip replacement surgery for osteoarthritis.Only tissue that would have been discarded was used with the approval of theSouthampton & South West Hampshire Local Research Ethics Committee (Ref: 194/99/w). Primary cultures of bone marrow cells were established as previouslydescribed [23]. Cultures were maintained in basal medium (a-MEM containing 10%FCS, penicillin-100 U/ml and streptomycin-100 U/ml) at 37 �C in humidified air with5% CO2. C2C12 cells (ECACC 91031101) were cultured in basal medium (DMEM
supplemented with 10% FCS, penicillin-100 U/ml and streptomycin-100 U/ml) andcultured at 37 �C in humidified air with 5% CO2. Human Umbilical Vein EndothelialCells (HUVEC) were isolated from umbilical cords, collected from the Princess AnneHospital, Southampton, UK, with the approval of the Southampton and South WestHampshire Local Research Ethics Committee (Ref: 05/Q1702/102). HUVEC wereisolated and cultured as described by Jaffe et al, [24] with minor modifications.Briefly, fresh umbilical cords, after informed consent, were collected in 1� PBS. Thecords were cleaned and then cannulated at each end and washed gently with 1�PBS. Collagenase Type 2 (1 mg/ml) in 1� PBS was injected into the umbilical vein,and left at 37 �C for 40 min. The cells were collected by flushing the vein with 1�PBS, continued with centrifuging the suspension at 400 g for 10 min. The pellet wasresuspended in endothelial cell basal medium (MEM 199 supplemented with 10%FCS, penicillin-100 U/ml and streptomycin-100 U/ml and endothelial cell growthsupplement) and cultured at 37 �C in humidified air with 5% CO2.
2.3. Fabrication of dual growth factor releasing alginate-VEGF165/PDLLA-BMP-2synthetic scaffolds
In models of bone repair it has been shown that angiogenesis occurs beforeosteogenesis where strong elevated expression of VEGF is observed at 5–14 days[25,26]. Hence, to mimic growth factor presentation in bone regeneration usinga controlled release scaffold we required a new scaffold that could release VEGF165
relatively quickly (peak release in 1 week) and BMP-2 over a longer time frame(more than one month). Therefore, in a single porous scaffold we required twocompartments, one compartment releasing VEGF165 rapidly and the secondcompartment retarding the early release of BMP-2.
To achieve these independent release kinetics we used alginate fibres asa hydrogel carrier for VEGF165 and PDLLA as a biodegradable carrier for BMP-2. ThePDLLA component forms the continuous phase and the scaffold possessesthe compressive properties of the PDLLA with the alginate fibre embedded in thecontinuous phase. The alginate/VEGF165 phase was manufactured using pre-formedalginate fibres (Advanced Medical Solutions, UK) of approx. 1 mm in thickness [23].The fibres were swollen in a sodium-rich NaCl/CaCl2 solution (NaCl/CaCl2 ratio 20:1,NaCl concentration 10% w/v) for 5 min before exposure to a 20 mg/ml solution ofVEGF165 for 30 min. The fibres were then immersed in a barium chloride solution (5%w/v) for 15 min, followed by washing with distilled water (3 times for 5 min) anddrying in air at room temperature. The VEGF165 loaded fibres were then cut intofibrils of 1–2 mm in length
The PDLLA/BMP-2 phase was formed using a supercritical carbon dioxide (scCO2)mixing technique [27,28]. Briefly, 20 mg of BMP-2 solution was added to PDLLApolymer powder and the mixture freeze-dried overnight to remove the water. Atthis stage the primary PDLLA/BMP-2 mixture was physically mixed with the VEGF165
loaded fibres and processed using scCO2 (200 bar/35 �C) for 1 h in a clamp-sealed60 ml pressure vessel. After a 1 h decompression cycle the scaffolds were taken outof the pressure vessel and the non-porous skin removed before assay. The compositescaffolds were scanned at 20 kV with a Philips 505 scanning electron microscope todetermine the levels of porosity. An average pore size of 250 mm and an averageporosity of 70% was determined by micro-CT.
2.4. Release kinetics of VEGF165 and BMP-2 from alginate/PDLLA scaffoldsrespectively
The alginate-VEGF165/PDLLA-BMP-2 scaffolds were cut into quarters (z40 mg)and were incubated with 1� antimycotic/antibiotic solution (Sigma, UK), washed 3times in 1� PBS before being placed in 24 well plates containing 0.5 ml 1� PBS (pH7.4) in an incubator (37 �C, 5% CO2 balanced air). At each time point (4, 7, 14, 21, & 28days) the supernatants were collected and stored at �80 �C. The scaffolds wereresuspended in fresh 1� PBS and incubated until the next following time point. Therelease of VEGF165 and BMP-2 were quantified with antibody-specific enzyme-linkedimmunosorbent assay (ELISA) kits according to the manufacturer’s instructions.
2.5. Activity of BMP-2 released from alginate-VEGF165/PDLLA-BMP-2 scaffolds
C2C12 cells (a mouse C3H muscle myoblast cell line) express alkaline phos-phatase activity when stimulated by BMP-2 and consequently are a good marker todetermine the activity of BMP-2 released from the composite scaffolds in situ overa period of time. Therefore, to test the activity of BMP-2 released from the scaffoldsC2C12 cells (1�105) were seeded either onto sterilised alginate/PDLLA or alginate-VEGF165/PDLLA-BMP-2 scaffolds and placed in 6 well culture plates and incubatedwith DMEM supplemented with 10% FCS, penicillin-100 U/ml; streptomycin-100 mg/mL and incubated at 37 �C in humidified air/5% CO2 where culture mediumwas replenished every 3–4 days. At day 28, scaffold degradation results in therelease of BMP-2 which seeded with C2C12 cells come in contact with. The cells onthe scaffolds were tested for alkaline phosphatase enzyme activity. Briefly, sampleswere rinsed in 1� PBS and fixed in 95% ethanol for 15 min prior to histochemicalstaining. Naphthol AS-MX Phosphate and Fast Violet-B Salts were combined inaccordance with a modified Sigma–Aldrich protocol 104 and applied to the culturedscaffold samples seeded with C2C12 cells [29].
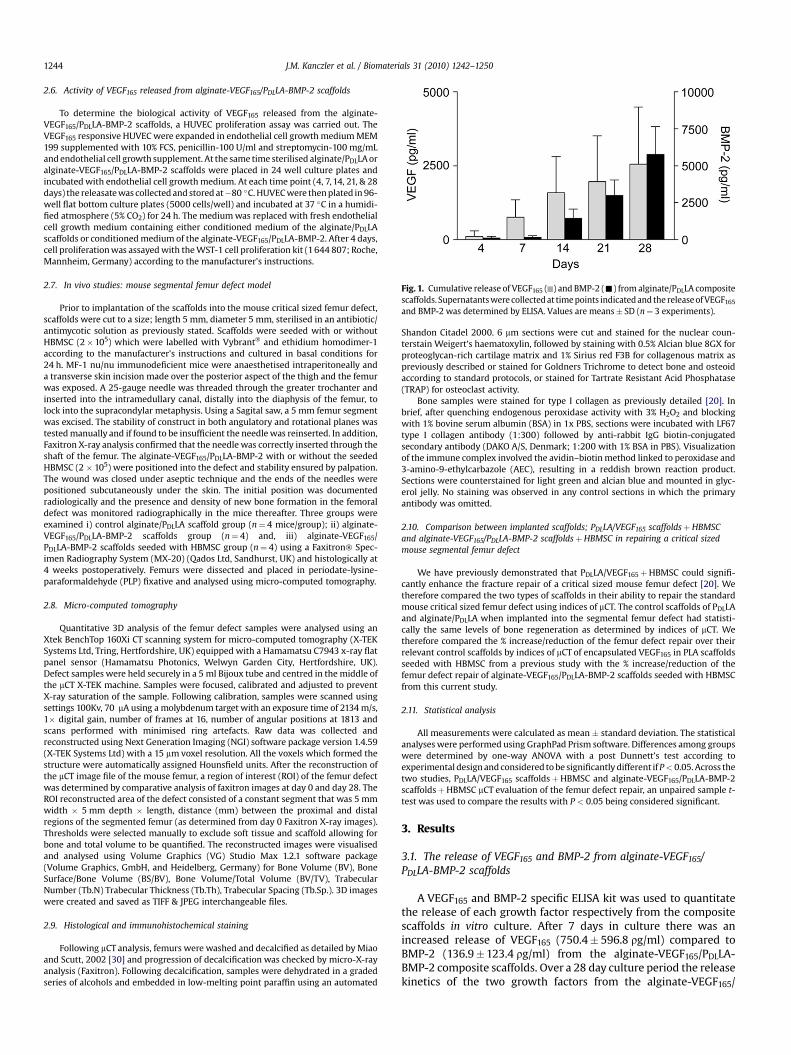
Fig. 1. Cumulative release of VEGF165 ( ) and BMP-2 (-) from alginate/PDLLA compositescaffolds. Supernatants were collected at time points indicated and the release of VEGF165
and BMP-2 was determined by ELISA. Values are means� SD (n¼ 3 experiments).
J.M. Kanczler et al. / Biomaterials 31 (2010) 1242–12501244
2.6. Activity of VEGF165 released from alginate-VEGF165/PDLLA-BMP-2 scaffolds
To determine the biological activity of VEGF165 released from the alginate-VEGF165/PDLLA-BMP-2 scaffolds, a HUVEC proliferation assay was carried out. TheVEGF165 responsive HUVEC were expanded in endothelial cell growth medium MEM199 supplemented with 10% FCS, penicillin-100 U/ml and streptomycin-100 mg/mLand endothelial cell growth supplement. At the same time sterilised alginate/PDLLA oralginate-VEGF165/PDLLA-BMP-2 scaffolds were placed in 24 well culture plates andincubated with endothelial cell growth medium. At each time point (4, 7, 14, 21, & 28days) the releasate was collected and stored at�80 �C. HUVEC were then plated in 96-well flat bottom culture plates (5000 cells/well) and incubated at 37 �C in a humidi-fied atmosphere (5% CO2) for 24 h. The medium was replaced with fresh endothelialcell growth medium containing either conditioned medium of the alginate/PDLLAscaffolds or conditioned medium of the alginate-VEGF165/PDLLA-BMP-2. After 4 days,cell proliferation was assayed with the WST-1 cell proliferation kit (1 644 807; Roche,Mannheim, Germany) according to the manufacturer’s instructions.
2.7. In vivo studies: mouse segmental femur defect model
Prior to implantation of the scaffolds into the mouse critical sized femur defect,scaffolds were cut to a size; length 5 mm, diameter 5 mm, sterilised in an antibiotic/antimycotic solution as previously stated. Scaffolds were seeded with or withoutHBMSC (2�105) which were labelled with Vybrant� and ethidium homodimer-1according to the manufacturer’s instructions and cultured in basal conditions for24 h. MF-1 nu/nu immunodeficient mice were anaesthetised intraperitoneally anda transverse skin incision made over the posterior aspect of the thigh and the femurwas exposed. A 25-gauge needle was threaded through the greater trochanter andinserted into the intramedullary canal, distally into the diaphysis of the femur, tolock into the supracondylar metaphysis. Using a Sagital saw, a 5 mm femur segmentwas excised. The stability of construct in both angulatory and rotational planes wastested manually and if found to be insufficient the needle was reinserted. In addition,Faxitron X-ray analysis confirmed that the needle was correctly inserted through theshaft of the femur. The alginate-VEGF165/PDLLA-BMP-2 with or without the seededHBMSC (2 � 105) were positioned into the defect and stability ensured by palpation.The wound was closed under aseptic technique and the ends of the needles werepositioned subcutaneously under the skin. The initial position was documentedradiologically and the presence and density of new bone formation in the femoraldefect was monitored radiographically in the mice thereafter. Three groups wereexamined i) control alginate/PDLLA scaffold group (n¼ 4 mice/group); ii) alginate-VEGF165/PDLLA-BMP-2 scaffolds group (n¼ 4) and, iii) alginate-VEGF165/PDLLA-BMP-2 scaffolds seeded with HBMSC group (n¼ 4) using a Faxitron� Spec-imen Radiography System (MX-20) (Qados Ltd, Sandhurst, UK) and histologically at4 weeks postoperatively. Femurs were dissected and placed in periodate-lysine-paraformaldehyde (PLP) fixative and analysed using micro-computed tomography.
2.8. Micro-computed tomography
Quantitative 3D analysis of the femur defect samples were analysed using anXtek BenchTop 160Xi CT scanning system for micro-computed tomography (X-TEKSystems Ltd, Tring, Hertfordshire, UK) equipped with a Hamamatsu C7943 x-ray flatpanel sensor (Hamamatsu Photonics, Welwyn Garden City, Hertfordshire, UK).Defect samples were held securely in a 5 ml Bijoux tube and centred in the middle ofthe mCT X-TEK machine. Samples were focused, calibrated and adjusted to preventX-ray saturation of the sample. Following calibration, samples were scanned usingsettings 100Kv, 70 mA using a molybdenum target with an exposure time of 2134 m/s,1� digital gain, number of frames at 16, number of angular positions at 1813 andscans performed with minimised ring artefacts. Raw data was collected andreconstructed using Next Generation Imaging (NGI) software package version 1.4.59(X-TEK Systems Ltd) with a 15 mm voxel resolution. All the voxels which formed thestructure were automatically assigned Hounsfield units. After the reconstruction ofthe mCT image file of the mouse femur, a region of interest (ROI) of the femur defectwas determined by comparative analysis of faxitron images at day 0 and day 28. TheROI reconstructed area of the defect consisted of a constant segment that was 5 mmwidth � 5 mm depth � length, distance (mm) between the proximal and distalregions of the segmented femur (as determined from day 0 Faxitron X-ray images).Thresholds were selected manually to exclude soft tissue and scaffold allowing forbone and total volume to be quantified. The reconstructed images were visualisedand analysed using Volume Graphics (VG) Studio Max 1.2.1 software package(Volume Graphics, GmbH, and Heidelberg, Germany) for Bone Volume (BV), BoneSurface/Bone Volume (BS/BV), Bone Volume/Total Volume (BV/TV), TrabecularNumber (Tb.N) Trabecular Thickness (Tb.Th), Trabecular Spacing (Tb.Sp.). 3D imageswere created and saved as TIFF & JPEG interchangeable files.
2.9. Histological and immunohistochemical staining
Following mCT analysis, femurs were washed and decalcified as detailed by Miaoand Scutt, 2002 [30] and progression of decalcification was checked by micro-X-rayanalysis (Faxitron). Following decalcification, samples were dehydrated in a gradedseries of alcohols and embedded in low-melting point paraffin using an automated
Shandon Citadel 2000. 6 mm sections were cut and stained for the nuclear coun-terstain Weigert’s haematoxylin, followed by staining with 0.5% Alcian blue 8GX forproteoglycan-rich cartilage matrix and 1% Sirius red F3B for collagenous matrix aspreviously described or stained for Goldners Trichrome to detect bone and osteoidaccording to standard protocols, or stained for Tartrate Resistant Acid Phosphatase(TRAP) for osteoclast activity.
Bone samples were stained for type I collagen as previously detailed [20]. Inbrief, after quenching endogenous peroxidase activity with 3% H2O2 and blockingwith 1% bovine serum albumin (BSA) in 1x PBS, sections were incubated with LF67type I collagen antibody (1:300) followed by anti-rabbit IgG biotin-conjugatedsecondary antibody (DAKO A/S, Denmark; 1:200 with 1% BSA in PBS). Visualizationof the immune complex involved the avidin–biotin method linked to peroxidase and3-amino-9-ethylcarbazole (AEC), resulting in a reddish brown reaction product.Sections were counterstained for light green and alcian blue and mounted in glyc-erol jelly. No staining was observed in any control sections in which the primaryantibody was omitted.
2.10. Comparison between implanted scaffolds; PDLLA/VEGF165 scaffoldsþHBMSCand alginate-VEGF165/PDLLA-BMP-2 scaffoldsþHBMSC in repairing a critical sizedmouse segmental femur defect
We have previously demonstrated that PDLLA/VEGF165þHBMSC could signifi-cantly enhance the fracture repair of a critical sized mouse femur defect [20]. Wetherefore compared the two types of scaffolds in their ability to repair the standardmouse critical sized femur defect using indices of mCT. The control scaffolds of PDLLAand alginate/PDLLA when implanted into the segmental femur defect had statisti-cally the same levels of bone regeneration as determined by indices of mCT. Wetherefore compared the % increase/reduction of the femur defect repair over theirrelevant control scaffolds by indices of mCT of encapsulated VEGF165 in PLA scaffoldsseeded with HBMSC from a previous study with the % increase/reduction of thefemur defect repair of alginate-VEGF165/PDLLA-BMP-2 scaffolds seeded with HBMSCfrom this current study.
2.11. Statistical analysis
All measurements were calculated as mean � standard deviation. The statisticalanalyses were performed using GraphPad Prism software. Differences among groupswere determined by one-way ANOVA with a post Dunnett’s test according toexperimental design and considered to be significantly different if P< 0.05. Across thetwo studies, PDLLA/VEGF165 scaffoldsþHBMSC and alginate-VEGF165/PDLLA-BMP-2scaffoldsþHBMSC mCT evaluation of the femur defect repair, an unpaired sample t-test was used to compare the results with P < 0.05 being considered significant.
3. Results
3.1. The release of VEGF165 and BMP-2 from alginate-VEGF165/PDLLA-BMP-2 scaffolds
A VEGF165 and BMP-2 specific ELISA kit was used to quantitatethe release of each growth factor respectively from the compositescaffolds in vitro culture. After 7 days in culture there was anincreased release of VEGF165 (750.4� 596.8 rg/ml) compared toBMP-2 (136.9�123.4 rg/ml) from the alginate-VEGF165/PDLLA-BMP-2 composite scaffolds. Over a 28 day culture period the releasekinetics of the two growth factors from the alginate-VEGF165/
Alginate/PLA Alginate-VEGF/PLA–BMP-2
0.00
0.25
0.50
0.75 **
A
B
Abso
rban
ce (4
50nm
)
Alginate/PLA+ C2C12 cells
Alginate-VEGF/PLA–BMP-2+ C2C12 cells
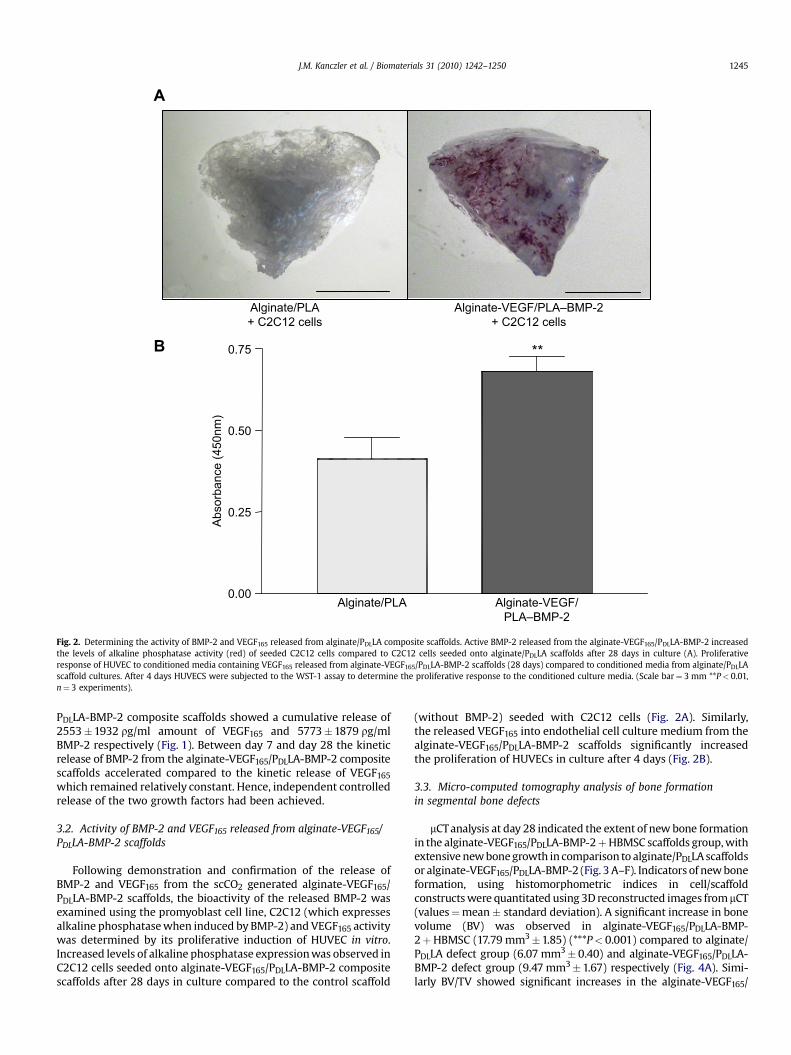
Fig. 2. Determining the activity of BMP-2 and VEGF165 released from alginate/PDLLA composite scaffolds. Active BMP-2 released from the alginate-VEGF165/PDLLA-BMP-2 increasedthe levels of alkaline phosphatase activity (red) of seeded C2C12 cells compared to C2C12 cells seeded onto alginate/PDLLA scaffolds after 28 days in culture (A). Proliferativeresponse of HUVEC to conditioned media containing VEGF165 released from alginate-VEGF165/PDLLA-BMP-2 scaffolds (28 days) compared to conditioned media from alginate/PDLLAscaffold cultures. After 4 days HUVECS were subjected to the WST-1 assay to determine the proliferative response to the conditioned culture media. (Scale bar¼ 3 mm **P< 0.01,n¼ 3 experiments).
J.M. Kanczler et al. / Biomaterials 31 (2010) 1242–1250 1245
PDLLA-BMP-2 composite scaffolds showed a cumulative release of2553�1932 rg/ml amount of VEGF165 and 5773�1879 rg/mlBMP-2 respectively (Fig. 1). Between day 7 and day 28 the kineticrelease of BMP-2 from the alginate-VEGF165/PDLLA-BMP-2 compositescaffolds accelerated compared to the kinetic release of VEGF165
which remained relatively constant. Hence, independent controlledrelease of the two growth factors had been achieved.
3.2. Activity of BMP-2 and VEGF165 released from alginate-VEGF165/PDLLA-BMP-2 scaffolds
Following demonstration and confirmation of the release ofBMP-2 and VEGF165 from the scCO2 generated alginate-VEGF165/PDLLA-BMP-2 scaffolds, the bioactivity of the released BMP-2 wasexamined using the promyoblast cell line, C2C12 (which expressesalkaline phosphatase when induced by BMP-2) and VEGF165 activitywas determined by its proliferative induction of HUVEC in vitro.Increased levels of alkaline phosphatase expression was observed inC2C12 cells seeded onto alginate-VEGF165/PDLLA-BMP-2 compositescaffolds after 28 days in culture compared to the control scaffold
(without BMP-2) seeded with C2C12 cells (Fig. 2A). Similarly,the released VEGF165 into endothelial cell culture medium from thealginate-VEGF165/PDLLA-BMP-2 scaffolds significantly increasedthe proliferation of HUVECs in culture after 4 days (Fig. 2B).
3.3. Micro-computed tomography analysis of bone formationin segmental bone defects
mCT analysis at day 28 indicated the extent of new bone formationin the alginate-VEGF165/PDLLA-BMP-2þHBMSC scaffolds group, withextensive new bone growth in comparison to alginate/PDLLA scaffoldsor alginate-VEGF165/PDLLA-BMP-2 (Fig. 3 A–F). Indicators of new boneformation, using histomorphometric indices in cell/scaffoldconstructs were quantitated using 3D reconstructed images from mCT(values¼mean� standard deviation). A significant increase in bonevolume (BV) was observed in alginate-VEGF165/PDLLA-BMP-2þHBMSC (17.79 mm3�1.85) (***P< 0.001) compared to alginate/PDLLA defect group (6.07 mm3� 0.40) and alginate-VEGF165/PDLLA-BMP-2 defect group (9.47 mm3�1.67) respectively (Fig. 4A). Simi-larly BV/TV showed significant increases in the alginate-VEGF165/
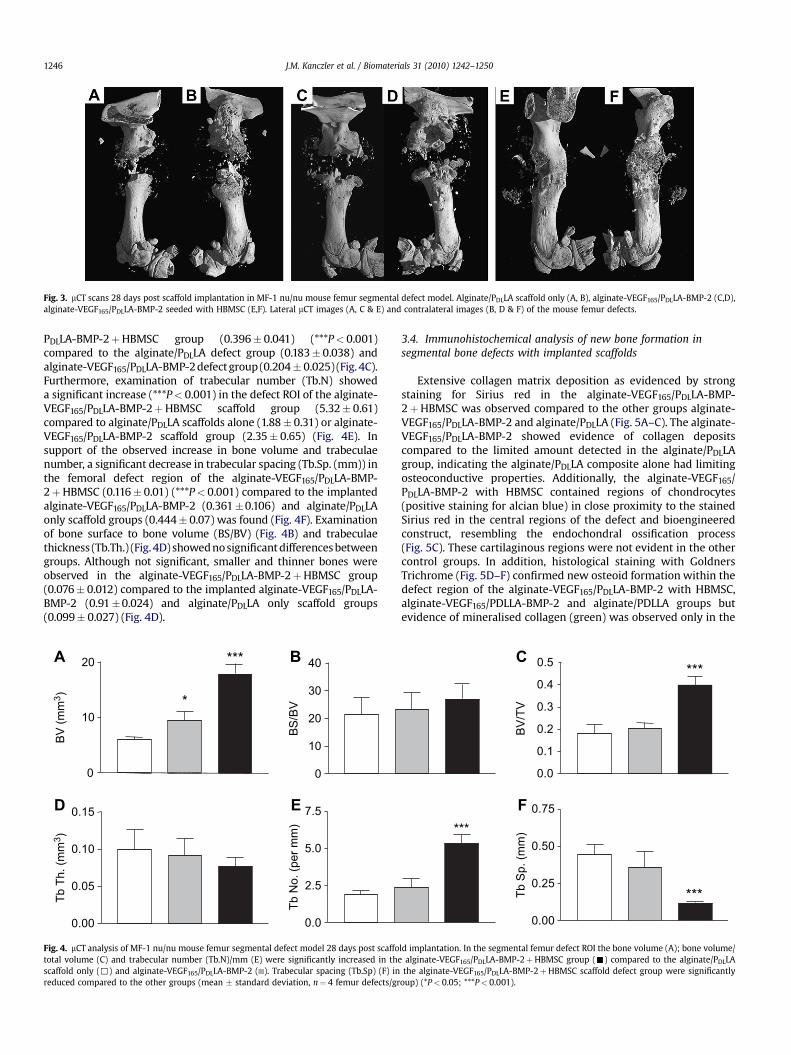
Fig. 3. mCT scans 28 days post scaffold implantation in MF-1 nu/nu mouse femur segmental defect model. Alginate/PDLLA scaffold only (A, B), alginate-VEGF165/PDLLA-BMP-2 (C,D),alginate-VEGF165/PDLLA-BMP-2 seeded with HBMSC (E,F). Lateral mCT images (A, C & E) and contralateral images (B, D & F) of the mouse femur defects.
J.M. Kanczler et al. / Biomaterials 31 (2010) 1242–12501246
PDLLA-BMP-2þHBMSC group (0.396� 0.041) (***P< 0.001)compared to the alginate/PDLLA defect group (0.183� 0.038) andalginate-VEGF165/PDLLA-BMP-2 defect group (0.204� 0.025) (Fig. 4C).Furthermore, examination of trabecular number (Tb.N) showeda significant increase (***P< 0.001) in the defect ROI of the alginate-VEGF165/PDLLA-BMP-2þHBMSC scaffold group (5.32� 0.61)compared to alginate/PDLLA scaffolds alone (1.88� 0.31) or alginate-VEGF165/PDLLA-BMP-2 scaffold group (2.35� 0.65) (Fig. 4E). Insupport of the observed increase in bone volume and trabeculaenumber, a significant decrease in trabecular spacing (Tb.Sp. (mm)) inthe femoral defect region of the alginate-VEGF165/PDLLA-BMP-2þHBMSC (0.116� 0.01) (***P< 0.001) compared to the implantedalginate-VEGF165/PDLLA-BMP-2 (0.361�0.106) and alginate/PDLLAonly scaffold groups (0.444� 0.07) was found (Fig. 4F). Examinationof bone surface to bone volume (BS/BV) (Fig. 4B) and trabeculaethickness (Tb.Th.) (Fig. 4D) showed no significant differences betweengroups. Although not significant, smaller and thinner bones wereobserved in the alginate-VEGF165/PDLLA-BMP-2þHBMSC group(0.076� 0.012) compared to the implanted alginate-VEGF165/PDLLA-BMP-2 (0.91�0.024) and alginate/PDLLA only scaffold groups(0.099� 0.027) (Fig. 4D).
0
10
20 ***
*
BV (m
m3 )
0
10
20
30
40
BS/B
V
0.00
0.05
0.10
0.15
Tb T
h. (m
m3 )
0.0
2.5
5.0
7.5
Tb N
o. (p
er m
m)
A B
D E
Fig. 4. mCT analysis of MF-1 nu/nu mouse femur segmental defect model 28 days post scaffototal volume (C) and trabecular number (Tb.N)/mm (E) were significantly increased in thescaffold only (,) and alginate-VEGF165/PDLLA-BMP-2 ( ). Trabecular spacing (Tb.Sp) (F) inreduced compared to the other groups (mean � standard deviation, n¼ 4 femur defects/gr
3.4. Immunohistochemical analysis of new bone formation insegmental bone defects with implanted scaffolds
Extensive collagen matrix deposition as evidenced by strongstaining for Sirius red in the alginate-VEGF165/PDLLA-BMP-2þHBMSC was observed compared to the other groups alginate-VEGF165/PDLLA-BMP-2 and alginate/PDLLA (Fig. 5A–C). The alginate-VEGF165/PDLLA-BMP-2 showed evidence of collagen depositscompared to the limited amount detected in the alginate/PDLLAgroup, indicating the alginate/PDLLA composite alone had limitingosteoconductive properties. Additionally, the alginate-VEGF165/PDLLA-BMP-2 with HBMSC contained regions of chondrocytes(positive staining for alcian blue) in close proximity to the stainedSirius red in the central regions of the defect and bioengineeredconstruct, resembling the endochondral ossification process(Fig. 5C). These cartilaginous regions were not evident in the othercontrol groups. In addition, histological staining with GoldnersTrichrome (Fig. 5D–F) confirmed new osteoid formation within thedefect region of the alginate-VEGF165/PDLLA-BMP-2 with HBMSC,alginate-VEGF165/PDLLA-BMP-2 and alginate/PDLLA groups butevidence of mineralised collagen (green) was observed only in the
0.0
0.1
0.2
0.3
0.4
0.5 ***
BV/T
V
***
0.00
0.25
0.50
0.75
Tb S
p. (m
m)
***
C
F
ld implantation. In the segmental femur defect ROI the bone volume (A); bone volume/alginate-VEGF165/PDLLA-BMP-2þHBMSC group (-) compared to the alginate/PDLLA
the alginate-VEGF165/PDLLA-BMP-2þHBMSC scaffold defect group were significantlyoup) (*P< 0.05; ***P< 0.001).
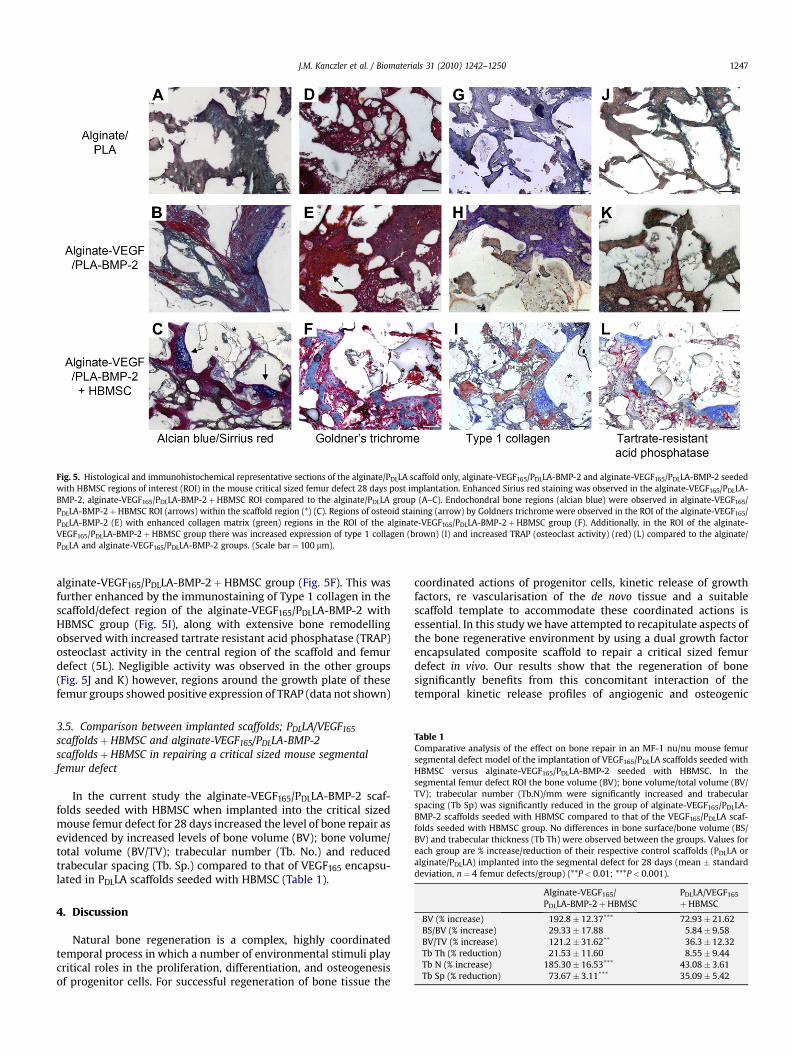
Fig. 5. Histological and immunohistochemical representative sections of the alginate/PDLLA scaffold only, alginate-VEGF165/PDLLA-BMP-2 and alginate-VEGF165/PDLLA-BMP-2 seededwith HBMSC regions of interest (ROI) in the mouse critical sized femur defect 28 days post implantation. Enhanced Sirius red staining was observed in the alginate-VEGF165/PDLLA-BMP-2, alginate-VEGF165/PDLLA-BMP-2þHBMSC ROI compared to the alginate/PDLLA group (A–C). Endochondral bone regions (alcian blue) were observed in alginate-VEGF165/PDLLA-BMP-2þHBMSC ROI (arrows) within the scaffold region (*) (C). Regions of osteoid staining (arrow) by Goldners trichrome were observed in the ROI of the alginate-VEGF165/PDLLA-BMP-2 (E) with enhanced collagen matrix (green) regions in the ROI of the alginate-VEGF165/PDLLA-BMP-2þHBMSC group (F). Additionally, in the ROI of the alginate-VEGF165/PDLLA-BMP-2þHBMSC group there was increased expression of type 1 collagen (brown) (I) and increased TRAP (osteoclast activity) (red) (L) compared to the alginate/PDLLA and alginate-VEGF165/PDLLA-BMP-2 groups. (Scale bar¼ 100 mm).
Table 1Comparative analysis of the effect on bone repair in an MF-1 nu/nu mouse femursegmental defect model of the implantation of VEGF165/PDLLA scaffolds seeded withHBMSC versus alginate-VEGF165/PDLLA-BMP-2 seeded with HBMSC. In thesegmental femur defect ROI the bone volume (BV); bone volume/total volume (BV/TV); trabecular number (Tb.N)/mm were significantly increased and trabecularspacing (Tb Sp) was significantly reduced in the group of alginate-VEGF165/PDLLA-BMP-2 scaffolds seeded with HBMSC compared to that of the VEGF165/PDLLA scaf-folds seeded with HBMSC group. No differences in bone surface/bone volume (BS/BV) and trabecular thickness (Tb Th) were observed between the groups. Values foreach group are % increase/reduction of their respective control scaffolds (PDLLA oralginate/PDLLA) implanted into the segmental defect for 28 days (mean � standarddeviation, n¼ 4 femur defects/group) (**P< 0.01; ***P< 0.001).
Alginate-VEGF165/PDLLA-BMP-2þHBMSC
PDLLA/VEGF165
þHBMSC
BV (% increase) 192.8� 12.37*** 72.93� 21.62BS/BV (% increase) 29.33� 17.88 5.84� 9.58BV/TV (% increase) 121.2� 31.62** 36.3� 12.32Tb Th (% reduction) 21.53� 11.60 8.55� 9.44Tb N (% increase) 185.30� 16.53*** 43.08� 3.61Tb Sp (% reduction) 73.67� 3.11*** 35.09� 5.42
J.M. Kanczler et al. / Biomaterials 31 (2010) 1242–1250 1247
alginate-VEGF165/PDLLA-BMP-2þHBMSC group (Fig. 5F). This wasfurther enhanced by the immunostaining of Type 1 collagen in thescaffold/defect region of the alginate-VEGF165/PDLLA-BMP-2 withHBMSC group (Fig. 5I), along with extensive bone remodellingobserved with increased tartrate resistant acid phosphatase (TRAP)osteoclast activity in the central region of the scaffold and femurdefect (5L). Negligible activity was observed in the other groups(Fig. 5J and K) however, regions around the growth plate of thesefemur groups showed positive expression of TRAP (data not shown)
3.5. Comparison between implanted scaffolds; PDLLA/VEGF165
scaffoldsþHBMSC and alginate-VEGF165/PDLLA-BMP-2scaffoldsþHBMSC in repairing a critical sized mouse segmentalfemur defect
In the current study the alginate-VEGF165/PDLLA-BMP-2 scaf-folds seeded with HBMSC when implanted into the critical sizedmouse femur defect for 28 days increased the level of bone repair asevidenced by increased levels of bone volume (BV); bone volume/total volume (BV/TV); trabecular number (Tb. No.) and reducedtrabecular spacing (Tb. Sp.) compared to that of VEGF165 encapsu-lated in PDLLA scaffolds seeded with HBMSC (Table 1).
4. Discussion
Natural bone regeneration is a complex, highly coordinatedtemporal process in which a number of environmental stimuli playcritical roles in the proliferation, differentiation, and osteogenesisof progenitor cells. For successful regeneration of bone tissue the
coordinated actions of progenitor cells, kinetic release of growthfactors, re vascularisation of the de novo tissue and a suitablescaffold template to accommodate these coordinated actions isessential. In this study we have attempted to recapitulate aspects ofthe bone regenerative environment by using a dual growth factorencapsulated composite scaffold to repair a critical sized femurdefect in vivo. Our results show that the regeneration of bonesignificantly benefits from this concomitant interaction of thetemporal kinetic release profiles of angiogenic and osteogenic

J.M. Kanczler et al. / Biomaterials 31 (2010) 1242–12501248
growth factors from a biodegradable composite scaffold in close, insitu presentation of osteoprogenitor cells.
Alginate/PDLLA composite scaffolds were loaded with a solutionof BMP-2 and VEGF165 by combining both a scCO2 mixing techniqueand an alginate entrapment technique. Previously, the adaptationand combination of these two encapsulating techniques alloweddifferent proteins (ribonuclease and horseradish peroxidase) to beencapsulated into a monolith, alginate and PDLLA compositescaffold [22]. Due to the variant release profiles of the polymersalginate and PDLLA, the encapsulated proteins were observed to bereleased at differing kinetic rates. Using this processing techniquewe were able to encapsulate VEGF165 into the alginate portion ofthe composite and BMP-2 into the PDLLA polymer, hence,fabricating a composite alginate-VEGF165/PDLLA-BMP-2 scaffold.VEGF165 and BMP-2 incorporated into these scaffolds allowed forsustained and localised release of these bioactive factors, albeit atdifferent kinetic rates. The alginate/PDLLA composite encapsulatedwith VEGF165 & BMP-2 respectively was designed to enable rapidrelease of VEGF165 compared to the slower, deliberate release rateof the BMP-2 from the composite scaffold; therefore, mimicking,albeit modestly, the events these growth factors are expressed inthe time events of bone fracture repair [25,32].
Our in vitro studies confirmed the different release kinetics ofVEGF165 from the alginate portion of the composite scaffold incomparison to the BMP-2 release from the PDLLA polymer over 7days. However, between 7 and 28 days the kinetic release of BMP-2from the PDLLA was greater than the release of VEGF165 from thealginate portion of the composite scaffold. Additional studiesconfirmed that both growth factors released from the scaffoldsretained their bioactivity over a 28 day period in culture andsubsequently were able to stimulate an osteogenic and anangiogenic response in C2C12 cells and HUVECs respectively. In ourmodel of a critical sized femur defect the combination of local andsustained VEGF165 and BMP-2 release from scaffolds in combina-tion with an osteoprogenitor population induced the strongestbone regenerative response. Moreover, the dual growth factorscaffolds without seeded HBMSC provided no significant advantagein repairing the segmental defect compared to alginate/PDLLAscaffolds alone. Using the segmental defect model we have previ-ously shown that HBMSC cells alone do not enhance the repairmechanism of bone, but, only a combined effect of HBMSC andVEGF165/PDLLA scaffolds proved efficacious [20]. Indeed, when wecompared the increase in bone repair of the alginate-VEGF165/PDLLA-BMP-2 scaffold þ HBMSC groups with VEGF165/PDLLA scaf-foldsþHBMSC groups; the dual growth factor scaffold compositesignificantly enhanced the regenerative capacity of the femurdefect.
Given these are large critical sized defects, cells seeded ontoscaffolds without growth factors are without sufficient local envi-ronmental stimuli to survive and repair the fracture. Similarly,scaffolds containing only encapsulated growth factors provideda minimal effect on the bone regeneration of the segmental defect.This is likely to be a consequence of localised growth factors and thelack of effect on host cells which will be a significant distance fromthe growth factors released.
Angiogenesis is a critical process in bone formation and boneregeneration [12,33]. Induction of a vascular supply in fracturerepair aids in the recruitment and differentiation of osteoprogeni-tor cell populations due to the provision of nutrients and oxygenand the signalling interactions between endothelial cells andosteoprogenitor cells [34]. Critically, the stabilisation of hypoxiainducible factor �1 and the subsequent upregulation of VEGFexpression enables the acceleration of bone healing in a distractionosteogenesis model [35]. Moreover, endothelial cells in closecontact with human osteoprogenitors have the ability to modulate
their proliferation and differentiation [36]. Furthermore, VEGF165
a potent angiogenic factor plays an important role in protectingcells [37]. The close proximity release of VEGF165 to the HBMSCseeded on to the VEGF165/PDLLA-BMP-2 scaffolds may not only actto keep the HBMSC viable sufficiently to enable differentiation andproliferation but, in addition, may also allow for endothelial cellrecruitment. In support of such a role, VEGF165, has been demon-strated to play a critical role in endochondral bone formation [13].Our histological observations of the segmental defect femursshowed that the alginate-VEGF165/PDLLA-BMP-2 with HBMSCdeveloped regions of endochondral bone formation in the centralregion of the scaffold/defect with elevated osteoclastic activitycompared to other scaffold groups. In these studies we observeda trend in the mCT analysis of the trabecular bone to be reduced inthickness in the VEGF165/PDLLA-BMP-2 with HBMSC groupcompared to other scaffold/defect groups concluding that thecombination of VEGF165 and BMP-2 released in conjunction withHBMSC leads to increased remodelling of the new bone which isbeing formed.
The current studies have a number of differences to previouswork in this area with the demonstration of differently kineticrelease profiles of active dual growth factors with in situ closeproximity of seeded osteoprogenitor cells augmenting boneformation in a load bearing critical sized femur defect. Previousstudies have demonstrated that the release of a single growth factorfrom polymeric carriers can enhance the regenerative capacity ofbone [38,39] although at protein concentrations much greater thanat physiological levels with inherent cost implications. Initially,scaffold design has allowed for the encapsulation of growth factorsinto biodegradable (approved by the FDA in synthetic bone graftproducts) scaffolds and released to enhance osteogenesis [6,40];correspondingly, angiogenic factors have also been incorporated toinduce angiogenesis in situ when released from its associatedscaffold [31]. Furthermore, Richardson et al. [10] demonstrated thepotential for the dual release of VEGF165 and platelet-derivedgrowth factor (PDGF)-BB factor from a scaffold at varying kineticrates to induce vascularisation. The combination of angiogenic andosteogenic factors can thus stimulate bone healing and regenera-tion [8,41,42]. Indeed, BMP-2 and TGF-b3, combined with RGD-alginate hydrogel co-delivered to femoral defects within polymerscaffolds showed an increase in bone formation in a critical sizeddefect (8 mm) [43]. Recently, Patel et al,. [45] demonstrated theimportance of the dual delivery of VEGF165 and BMP-2 in enhancingthe bone repair of a rat cranial critical sized defect using gelatinmicroparticles incorporated into poly(propylene fumarate)scaffolds. Additionally, Kempen et al. [45] demonstrated thata composite scaffold allowing the sequential release of VEGF andBMP-2 could enhance angiogenesis and osteogenesis ectopicallyimplanted into rats. Interestingly, when these scaffolds wereimplanted into a critical sized femur defect the VEGF/BMP-2composite showed only a slight increase compared to the BMP-2scaffold alone in repairing the defect.
In our studies we did not see any significant increase in boneformation unless the combination of cells and dual growth factorswere implanted into the femur defect. The differences are possiblydue to the in vivo bone defect models being used to assess the repairof a critical sized bone defect; the differing release mechanisms ofcomposite scaffolds used and the size of the critical defects createdfor repair. Our femur defect model showed an endochondral likerepair mechanism with the implanted dual scaffolds and HBMSCsimilar to the results Kempen et al. [45] showed with their bonedefect studies, whereas Patel et al. [44], cranial defect modeldemonstrated an intramembraneous ossification process. From thenumber of studies in dual release growth factors for bone repairand tissue engineering of a critical sized defect it appears the

J.M. Kanczler et al. / Biomaterials 31 (2010) 1242–1250 1249
enhanced effect of these scaffolds is critically dependent on releaseof growth factors and the location they are acting on and criticallythe addition of osteoprogenitors or mesenchymal stem cells inclose proximity or the rapid recruitment of local mesenchymalstem cells.
The importance of combining essential growth factors,progenitor cells and specific release cues with the aid of intelligentbiodegradable scaffolds offers an efficient strategy to enhance there-engineering of tissue, particularly bone. Hence, polymeric scaf-folds capable of distinct kinetic delivery profiles of multiple growthfactors could be employed to enhance and aid better replication ofthe temporal sequence of growth factor expression that occursduring natural bone repair. The current studies demonstrate theability to deliver, temporally, a combination of VEGF165 and BMP-2released from porous biocompatible scaffolds seeded with HBMSC,to sites of bone defects, enhances the repair and formation of newbone tissue.
The development of three-dimensional engineered skeletaltissue will depend upon the ability to organise appropriate signal(s)into a coordinated, temporal regenerative vehicle and, critically, anappropriate vascular network. The alginate entrapment and scCO2
encapsulation techniques prove a significant step for the prepara-tion of novel scaffolds for tissue engineering and critical in the dualrelease of VEGF165 and BMP-2 to enhance bone regeneration. Thepotential to incorporate progenitor and different cell types withinpolymeric scaffolds to coincide with release of growth factors thatoccur in the repair of a fracture defect is an exciting challenge.
Additionally, improvements to this process can be made byusing high osteocondictive polymers instead of PLA, or the additionof particles such as hydroxyapatite to PLA to improve the osteo-conductivity and the strength of the polymer. Optimisation of theporosity of these composite constructs without compromising itsstructural integrity is vital for the development of a functionalvasculature and the survivability of the engineered tissue. In oursystem we used static seeding of HBMSC onto the scaffold; dynamicseeding in the future may improve the number of cells penetratingthe scaffold and thus enhance the bone regenerative process.Dynamic seeding together of a number of cell types such asendothelial and osteoprogenitor cells with dual releasing growthfactor scaffolds would have an improved outcome in the boneregenerative process. Indeed, co-culture systems of endothelialcells and mesenchymal cells have already been shown to increasethe osteogenic response [46] and multi seeding of endothelialprogenitor and bone marrow stromal cells onto biodegradablescaffolds has improved bone regeneration in a segmental femoraldefect [47].
Finally, the mouse model provides us with valuable informationon the dynamic interactions of scaffold, growth factor release andHBMSC in a critical sized bone defect repair however, successfultranslation of this technology to larger animal bone defect studieswill only then potentially lead to this technology being used forbone regeneration strategies in the clinical orthopaedic arena.
5. Conclusion
In summary, our current studies demonstrate the ability todeliver growth factors locally from biodegradable composite scaf-folds at varying kinetic rates in close proximity to seeded HBMSCcan enhance the reparative mechanism of critical sized bonedefects; thus, mimicking the in vivo bone repair environment.Strategies to enable the coordinated, multiple releases of growthfactors such as VEGF165 and BMP-2 from composite scaffolds offeran opportunity to mimic, in part, the conditions present in therepair sequences of a bone fracture. Such cell and tissue engi-neering strategies offer therapeutic opportunities in orthopaedics
and serve as a model to study the interaction between growthfactors, progenitor cells and scaffolds in hard tissue regenerationand engineering of other complex tissues.
Acknowledgements
The authors would like to thank members of the Bone and JointResearch Group for useful discussions and, in particular, Ms KateMurawski for her technical assistance. We would like to thank DrLarry Fisher for provision of Type I collagen antibody and ProfessorWalter Sebald for the provision of BMP-2 and to the midwives atthe Princess Anne hospital for the collection of umbilical cords. Thiswork was supported by the Engineering and Physical SciencesResearch Council (grant number GR/S27276/01).
Appendix
Figure with essential color discrimination. Fig. 2 of this articlemay be difficult to interpret in black and white. The full colorimages can be found in the online version, at doi:10.1016/j.biomaterials.2009.10.059.
References
[1] Langer R, Vacanti JP. Tissue engineering. Science 1993;260:920–6.[2] Bolland BJ, Tilley S, New AM, Dunlop DG, Oreffo RO. Adult mesenchymal stem
cells and impaction grafting: a new clinical paradigm shift. Expet Rev Med Dev2007;4:393–404.
[3] Kneser U, Schaefer DJ, Polykandriotis E, Horch RE. Tissue engineering of bone:the reconstructive surgeon’s point of view. J Cell Mol Med 2006;10:7–19.
[4] Murphy WL, Simmons CA, Kaigler D, Mooney DJ. Bone regeneration viaa mineral substrate and induced angiogenesis. J Dent Res 2004;83:204–10.
[5] Levenberg S, Rouwkema J, Macdonald M, Garfein ES, Kohane DS, Darland DC, et al.Engineering vascularized skeletal muscle tissue. Nat Biotechnol 2005;23:879–84.
[6] Yang XB, Whitaker MJ, Sebald W, Clarke N, Howdle SM, Shakesheff KM, et al.Human osteoprogenitor bone formation using encapsulated bone morpho-genetic protein 2 in porous polymer scaffolds. Tissue Eng 2004;10:1037–45.
[7] Huang YC, Kaigler D, Rice KG, Krebsbach PH, Mooney DJ. Combined angiogenicand osteogenic factor delivery enhances bone marrow stromal cell-drivenbone regeneration. J Bone Miner Res 2005;20:848–57.
[8] Kaigler D, Wang Z, Horger K, Mooney DJ, Krebsbach PH. VEGF scaffoldsenhance angiogenesis and bone regeneration in irradiated osseous defects.J Bone Miner Res 2006;21:735–44.
[9] Putnam AJ, Mooney DJ. Tissue engineering using synthetic extracellularmatrices. Nat Med 1996;2:824–6.
[10] Richardson TP, Peters MC, Ennett AB, Mooney DJ. Polymeric system for dualgrowth factor delivery. Nat Biotechnol 2001;19:1029–34.
[11] Brey EM, King TW, Johnston C, McIntire LV, Reece GP, Patrick Jr CW. A tech-nique for quantitative three-dimensional analysis of microvascular structure.Microvasc Res 2002;63:279–94.
[12] Carano RA, Filvaroff EH. Angiogenesis and bone repair. Drug Discov Today2003;8:980–9889.
[13] Gerber HP, Vu TH, Ryan AM, Kowalski J, Werb Z, Ferrara N. VEGF coupleshypertrophic cartilage remodeling, ossification and angiogenesis duringendochondral bone formation. Nat Med 1999;5:623–8.
[14] Mayr-Wohlfart U, Waltenberger J, Hausser H, Kessler S, Gunther KP, Dehio C,et al. Vascular endothelial growth factor stimulates chemotactic migration ofprimary human osteoblasts. Bone 2002;30:472–7.
[15] Wang Y, Wan C, Deng L, Liu X, Cao X, Gilbert SR, et al. The hypoxia-induciblefactor alpha pathway couples angiogenesis to osteogenesis during skeletaldevelopment. J Clin Invest 2007;117:1616–26.
[16] Bouletreau PJ, Warren SM, Spector JA, Peled ZM, Gerrets RP, Greenwald JA,et al. Hypoxia and VEGF up-regulate BMP-2 mRNA and protein expression inmicrovascular endothelial cells: implications for fracture healing. PlastReconstr Surg 2002;109:2384–97.
[17] Deckers MM, van Bezooijen RL, van der Horst G, Hoogendam J, van Der Bent C,Papapoulos SE, et al. Bone morphogenetic proteins stimulate angiogenesisthrough osteoblast-derived vascular endothelial growth factor A. Endocri-nology 2002;143:1545–53.
[18] Hausman MR, Schaffler MB, Majeska RJ. Prevention of fracture healing in ratsby an inhibitor of angiogenesis. Bone 2001;29:560–4.
[19] Street J, Bao M, deGuzman L, Bunting S, Peale Jr FV, Ferrara N, et al. Vascularendothelial growth factor stimulates bone repair by promoting angiogenesisand bone turnover. Proc Natl Acad Sci U S A 2002;99:9656–61.
[20] Kanczler JM, Ginty PJ, Barry JJ, Clarke NM, Howdle SM, Shakesheff KM, et al.The effect of mesenchymal populations and vascular endothelial growth factor

J.M. Kanczler et al. / Biomaterials 31 (2010) 1242–12501250
delivered from biodegradable polymer scaffolds on bone formation. Bioma-terials 2008;29:1892–900.
[21] Simmons CA, Alsberg E, Hsiong S, Kim WJ, Mooney DJ. Dual growth factordelivery and controlled scaffold degradation enhance in vivo bone formationby transplanted bone marrow stromal cells. Bone 2004;35:562–9.
[22] Ginty PJ, Barry JJ, White LJ, Howdle SM, Shakesheff KM. Controlling proteinrelease from scaffolds using polymer blends and composites. Eur J PharmBiopharm 2008;68:82–9.
[23] Oreffo RO, Virdi AS, Triffitt JT. Modulation of osteogenesis and adipogenesis byhuman serum in human bone marrow cultures. Eur J Cell Biol 1997;74:251–61.
[24] Jaffe EA, Nachman RL, Becker CG, Minick CR. Culture of human endothelialcells derived from umbilical veins. Identification by morphologic and immu-nologic criteria. J Clin Invest 1973;52:2745–56.
[25] Sojo K, Sawaki Y, Hattori H, Mizutani H, Ueda M. Immunohistochemical studyof vascular endothelial growth factor (VEGF) and bone morphogeneticprotein-2, -4 (BMP-2, -4) on lengthened rat femurs. J Craniomaxillofac Surg2005;33:238–45.
[26] Weiss S, Zimmermann G, Pufe T, Varoga D, Henle P. The systemic angiogenicresponse during bone healing. Arch Orthop Trauma Surg 2009;129:989–97.
[27] Howdle SM, Watson MS, Whitaker MJ, Popov VK, Davies MC, Mandel FS, et al.Supercritical fluid mixing: preparation of thermally sensitive polymercomposites containing bioactive materials. Chem Commun 2001;01:109–10.
[28] Hou LT, Liu CM, Liu BY, Chang PC, Chen MH, Ho MH, et al. Tissue engineeringbone formation in novel recombinant human bone morphogenic protein2-atelocollagen composite scaffolds. J Periodontol 2007;78:335–43.
[29] Bancroft JD. Enzyme histochemistry and its diagnostic applications. In:Bancroft JD, Gamble M, editors. Theory and practise of histological techniques.London: Churchill Livingstone Press; 2002. p. 596–8.
[30] Miao D, Scutt A. Histochemical localization of alkaline phosphatase activity indecalcified bone and cartilage. J Histochem Cytochem 2002;50:333–40.
[31] Kanczler JM, Barry J, Ginty P, Howdle SM, Shakesheff KM, Oreffo RO. Super-critical carbon dioxide generated vascular endothelial growth factor encap-sulated poly(DL-lactic acid) scaffolds induce angiogenesis in vitro. BiochemBiophys Res Commun 2007;352:135–41.
[32] Uchida S, Sakai A, Kudo H, Otomo H, Watanuki M, Tanaka M, et al. Vascular endo-thelial growth factor is expressed along with its receptors during the healing processof bone and bone marrow after drill-hole injury in rats. Bone 2003;32:491–501.
[33] Kanczler JM, Oreffo RO. Osteogenesis and angiogenesis: the potential forengineering bone. Eur Cell Mater 2008;15:100–14.
[34] Kaigler D, Krebsbach PH, Polverini PJ, Mooney DJ. Role of vascular endothelialgrowth factor in bone marrow stromal cell modulation of endothelial cells.Tissue Eng 2003;9:95–103.
[35] Wan C, Gilbert SR, Wang Y, Cao X, Shen X, Ramaswamy G, et al. Activation ofthe hypoxia-inducible factor-1alpha pathway accelerates bone regeneration.Proc Natl Acad Sci U S A 2008;105:686–91.
[36] Guillotin B, Bareille R, Bourget C, Bordenave L, Amedee J. Interaction betweenhuman umbilical vein endothelial cells and human osteoprogenitors triggerspleiotropic effect that may support osteoblastic function. Bone 2008;42:1080–91.
[37] Ferrari G, Pintucci G, Seghezzi G, Hyman K, Galloway AC, Mignatti P. VEGF.a prosurvival factor, acts in concert with TGF-beta1 to induce endothelial cellapoptosis. Proc Natl Acad Sci 2006;103:17260–5.
[38] Saito N, Okada T, Horiuchi H, Ota H, Takahashi J, Murakami N, et al. Local boneformation by injection of recombinant human bone morphogenetic protein-2contained in polymer carriers. Bone 2003;32:381–6.
[39] Bessho K, Carnes DL, Cavin R, Ong JL. Experimental studies on bone inductionusing low-molecular-weight poly (DL-lactide-co-glycolide) as a carrier forrecombinant human bone morphogenetic protein- 2. J Biomed Mater Res2002;61:61–5.
[40] Wei G, Jin Q, Giannobile WV, Ma PX. The enhancement of osteogenesis bynano-fibrous scaffolds incorporating rhBMP-7 nanospheres. Biomaterials2007;28:2087–96.
[41] Geiger F, Bertram H, Berger I, Lorenz H, Wall O, Eckhardt C, et al. Vascularendothelial growth factor gene-activated matrix (VEGF165-GAM) enhancesosteogenesis and angiogenesis in large segmental bone defects. J Bone MinerRes 2005;20:2028–35.
[42] Peng H, Usas A, Olshanski A, Ho AM, Gearhart B, Cooper GM, et al. VEGFimproves, whereas sFlt1 inhibits, BMP2-induced bone formation and bonehealing through modulation of angiogenesis. J Bone Miner Res 2005;20:2017–27.
[43] Oest ME, Dupont KM, Kong HJ, Mooney DJ, Guldberg RE. Quantitativeassessment of scaffold and growth factor-mediated repair of critically sizedbone defects. J Orthop Res 2007;25:941–50.
[44] Patel ZS, Young S, Tabat Y, Jansen JA, Wong ME, Mikos AG. Dual delivery of anangiogenic and an osteogenic growth factor for bone regeneration in a criticalsized defect model. Bone 2008;43:931–40.
[45] Kempen DH, Lu L, Heijink A, Hefferan TE, Creemers LB, Maran A, et al. Effect oflocal sequential VEGF and BMP-2 delivery on ectopic and orthotopic boneregeneration. Biomaterials 2009;30:2816–25.
[46] Kaigler D, Krebsbach PH, West ER, Horger K, Huang YC, Mooney DJ. Endothelcell modulation bone marrow stromal cell osteogenic potential. FASEB J2005;19:665–7.
[47] Yu H, Vandevord PJ, Gong W, Wu B, Song Z, Matthew HW, et al. Promotion ofosteogenesis in tissue-engineered bone by pre-seeding endothelial progenitorcells-derived endothelial cells. J Orthop Res 2008;26:1147–52.
Copyright © 2022 FDOKUMEN




















