
From Main to High: Consumers, Class, and the Spatial Reorientation of an Industrial City
Upload
uni-freiburgCategory
view
0download
0
Environmental and Experimental Botany 53 (2005) 299–314
Morphogenic effects of abiotic stress: reorientation of growthin Arabidopsis thaliana seedlings
Taras Pasternaka,b, Volodymir Rudasa, Geert Pottersb,∗, Marcel A.K. Jansenb,c
a Institute of Cell Biology and Genetic Engineering Natl. Acad. Sci. of Ukraine, Zabolotnogo Str. 148, U-03650 Kiev, Ukraineb Laboratory for Plant Physiology, Department of Biology, University of Antwerp, Groenenborgerlaan 171, B 2020 Antwerp, Belgium
c Department of Plant Sciences (ZEPS), University College Cork, Butler Building, Distillery Field, North Mall, Cork, Ireland
Accepted 9 April 2004
Abstract
Abiotic stress responses include changes in physiological and biochemical processes as well as morphological and devel-opmental patterns. It has remained an enigma which mechanisms are responsible for stress-induced morphogenesis. In thispaper we demonstrate that stress induced phenotypes comprise a re-orientation rather than a cessation of growth. Moreover,strong similarities between the phenotypes induced by excess copper, paraquat, salicylic acid and a hydrogen peroxide analogue,indicate that a common molecular-physiological response system mediates these morphogenic stress responses. It is proposedthat reactive oxygen species play a key role in controlling the architectural changes in stressedArabidopsis thaliana seedlings.
We found that phenotypes of plants exposed to stress resemble, in terms of the redistribution of growth, plants altered inphytohormone metabolism. We also found that plants in which polar auxin transport is blocked with TIBA, strongly resemble,but are not identical to, plants exposed to abiotic stress. Based on the stress induced formation of lateral roots, we surmise thatstress induces local auxin accumulation near the root pericycle.© 2004 Elsevier B.V. All rights reserved.
Keywords: Abiotic stress; Plant morphogenesis; Auxin;Arabidopsis thaliana; Reactive oxygen species
1. Introduction
Plants are exposed to a wide and diverse range ofabiotic and biotic stress conditions. However, mecha-nisms have evolved that diminish the impact of thesestresses on plant development and growth. Someof these so-called protective responses are highlystress-specific. Thus, photolyases very specifically
∗ Corresponding author. Tel.:+32 3 265 34 21;fax: +32 3 265 34 17.
E-mail address: [email protected] (G. Potters).
restore DNA damage resulting from the exposure toUV-radiation (Britt and Fiscus, 2003), phytochelatinsspecifically remove heavy metals from the cytoplasm(Sauge-Merle et al., 2003), solutes accumulate tocounter drought (Patakas et al., 2002), and xantho-phylls protect photosystem II from excessive radiationand heat (Davison et al., 2002). In contrast, enzymaticscavengers of free reactive oxygen species (ROS)like superoxide dismutase, catalase, and ascorbateperoxidase, or low molecular weight antioxidantslike ascorbate (ASC) and glutathione (GSH), preventoxidative damage in general. Thus, these scavengers
0098-8472/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.envexpbot.2004.04.009

300 T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314
confer tolerance to a range of stresses (Allen, 1995;Mittler, 2002). Conyza bonarienses plants that wereselected under field conditions as being tolerant toparaquat, are co-tolerant against a wide range of otherabiotic stresses, including SO2, ozone, photoinhibi-tion and others (Jansen et al., 1990; Ye and Gressel,1994). Conversely, plants with reduced antioxidativecapacity are relatively sensitive to a range of differentstresses (Mittler et al., 1999; Veljovic-Jovanovic et al.,2001). The up-regulation of free radical scavengingsystems is a common component of the response ofplants to many distinct stresses. Indeed, free radicaldamage is associated with a particularly broad rangeof different biotic and abiotic stresses including heatstress (Dat et al., 1998; Larkindale and Knight, 2002),UV-radiation stress (Brosche and Strid, 2003), pho-toinhibition (Muller-Moule et al., 2003), heavy metalstress (Pinto et al., 2003) and anoxia (Blokhina et al.,2001).
A second response common to many distinct abi-otic stresses, is the induction of architectural changes.Mechanistically distinct abiotic stresses can all giverise to alterations in plant morphology (Eerens et al.,1998; Malinowski and Belesky, 2000; Schikora andSchmidt, 2001; Bahaji et al., 2002; Lee, 2002; Jouiliand El Ferjani, 2003). The molecular mechanisms un-derlying these responses are often poorly understood.In this paper we detail how excess Cu impacts on thephenotype ofArabidopsis thaliana. Our data showthat the stress-induced alterations in plant architec-ture are not simply a consequence of a stress-inducedcessation of growth, but rather involve a specific re-orientation of growth. Changes in ROS metabolismcommonly follow exposure of plants to a wide varietyof biotic or abiotic stresses. In this paper, we spec-ulate that changes in ROS metabolism, possibly viaeffects on auxin homeostasis, control stress-inducedmorphogenic growth responses.
2. Materials and methods
2.1. Plant material
Seeds ofA. thaliana var. Columbia were used inthe experiments. Seeds were surface-sterilized for10 min in 20% commercial bleach and rinsed threetimes in sterile water. Then they were placed on 1/2
MS (Murashige and Skoog, 1962) medium containing1.5% sucrose and 0.8% agar and incubated at 16 hlight (40 W/m2) at a temperature of 24◦C. In each ex-periment, 40 seeds were cultured in a 100-mm glasspetri dish, which contained 20 mL of the medium.
2.2. Stress agents used
tert-Butyl hydroperoxide (tBHP), paraquat (PQ),oxy-benzoic acid (salicylic acid, SA) and CuSO4were tested as stress agents. 2,3,5-Triiodobenzoic acid(TIBA) was tested because of its known morphogeniceffect. PQ and tBHP stock solutions were filter ster-ilized through 0.22�m Millipore membrane filters.SA was dissolved in ethanol and TIBA was dissolvedin DMSO. All agents were added to the autoclavedmedium after it had cooled down to 50◦C. To correctfor potential solvent effects, appropriate amounts ofethanol and DMSO were added to control medium. Atleast three replicates were carried out per treatment.
2.3. Determination of morphological parameters
Arabidopsis seedlings and plants were viewed un-der a Zeiss stereomicroscope. Photos were taken usingan Olympus digital camera under appropriate magni-fication (3×–20×). After 7 days of growth, all sec-ondary roots visible at 5× magnification were scoredas evolved lateral roots. Primary root, cotyledon andleaf sizes were determined using SPOT advanced soft-ware following imaging and processing. Results areexpressed as mean± standard error (S.E.;n = 20 ex-plants). Each measurement was performed at least inthree biologically replications (each comprising about40 seedlings). Cell sizes in petioles and leaves weremeasured under a Zeiss Standard25 microscope usingan ocular-micrometer.
2.4. Measurements of chlorophyll contents
Chlorophyll was extracted from individual leavesof 25 days old plants by grinding in 95% alcohol in a1.5 mL micro-centrifuge tube using a disposable pes-tle, in the dark. Measurements of the optical densityof supernatant were carried out on a Gilford-1500spectrophotometer. Chlorophyll concentration perfresh weight of leaf was calculated as described byVernon (Vernon, 1960).

T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314 301
All measurements represent the average of threeindependent experiments, each involving 10 seedlings.
3. Results
Stress exposure affects both morphological param-eters as well as developmental programmes. In thisstudy, we have recorded morphological changes inA.thaliana in the context of temporal or growth stageinformation according toBoyes et al. (2001)(Fig. 1).
3.1. Copper ions induce architectural anddevelopmental changes
Cu is an essential transition metal that causes ox-idative stress when given in excess. We exposedA.thaliana seeds to modest concentrations of CuSO4 (30and 50�M) and found that these promoted the earlystage of seed germination, i.e. testa rupture (Fig. 2A).Cu effects on the growth of seedlings were more com-plex. Cu (30 and 50�M) inhibited root elongation in
Fig. 1. Model of chronological progression through the principal growth stages ofArabidopsis on 1/2 MS medium in vitro, according toBoyes et al. (2001).
a dose dependent manner (Fig. 2B). Under these con-ditions, no macroscopic damage to root tip and/or rootmeristem itself could be observed. Yet, we did notethat the elongation zone, between the root apex and thepoint of emerging root hairs, was significantly short-ened in Cu-treated plants (Fig. 2D–F and I). This pointbecame particularly clear when plants were exposed toa high Cu concentration (100�M), which resulted inthe formation of root hairs adjacent to the root tip (notshown). Root hair density was significantly increasedin the Cu-treated plants (Fig. 2G, H and J). Misshapen(short and obtuse) root hairs were only observed inthe case of a high Cu concentration (100�M).
A second marked effect of Cu on the root systemis the acceleration of the emergence of lateral roots.Under the growth conditions used, lateral root for-mation could first be observed in the control plantsafter 10 days of culture. Cu significantly acceleratedthis process. For example, in 26% of the plants ex-posed to 50�M Cu, lateral roots were seen to emer-gence following 7 days of culture (Fig. 2C). The dataon root elongation, root hair development and lateral

302 T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314
Fig. 2. Cu treatments alter root growth and morphology inArabidopsis. For all graphs – bars indicate standard error.∗P < 0.05, ∗∗P <
0.01, ∗∗∗P < 0.005, as compared to the control plants. (A) Testa rupture at 48 h after the start of the imbibition ofArabidopsis seeds indifferent CuSO4 concentrations, expressed as percentage of the total cell population. (B) Root (white bars) and cotyledon (black bars) sizeat 5 days after the start of the imbibition ofArabidopsis seeds in different CuSO4 concentrations. (C) Percentage of plants with lateralroot emergence at 7 days after imbibition, inArabidopsis seedlings treated with different Cu concentrations. (D–F) Root morphology, 7days after imbibition, in control medium (D), or in medium supplemented with 30�M (E) or 50�M (F) Cu. Bar= 200�m. (G, H) Roothair aspect, 7 days imbibition, in control medium (G) or in medium supplemented with 50�m Cu (H). Bar= 200�m. (I) Elongation zonelength, in�m, at 7 days after imbibition, ofArabidopsis roots cultivated in medium with different Cu concentrations. (J) Root hair density(number per 40�m of root length) at 7 days after imbibition, ofArabidopsis roots cultivated in medium with different Cu concentrations.

T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314 303
root formation indicate that Cu impacts mainly on cellelongation rather than cell division and cell differenti-ation (i.e. root hair formation). These data also clearlyshow that Cu, rather than causing an a-specific ces-sation of growth, induces a reorientation in the pat-tern of growth. Cell division was eventually halted inthe original root meristem. This is in accordance withthe report ofFrancis (1998), who demonstrated thatzinc prolongs G1 phase and thus effectively lowersmitotic activity in root tips. However, our data showthat new meristematic zones are simultaneously be-ing formed, thus emphasising the concept of stress in-duced re-orientation of growth.
In the early stages of the growth cycle (days 5–7),Cu has a relatively small inhibitory effect on the expan-sion of the cotyledons (Fig. 2B). Cu does not impacton the progression of the leaf forming reactions in theshoot apex. Even exposure to a Cu concentration ashigh as 100�M does not delay the timing of the emer-gence of the true leaves, rather, we observed that lowconcentrations of Cu promote leaf formation (Fig. 3A).However, the newly formed leaves are smaller, andthis is a dose-dependent effect (Fig. 3B–E). So, atthe end of the leaf formation period (days 12–14) allplants have at least the same number of true leaves asthe untreated control (ca. 6), but the leaf area of theCu-treated plants is significantly smaller than that ofcontrol seedlings.
The next stage of theA. thaliana growth cycle(days 12–25) is mainly characterized by expansionof leaves that were already present. At the end of therosette expansion period (approximately day 24), con-trol plants had an average rosette diameter of 17 mm,plants treated with 30�M Cu of 9 mm and plantstreated with 100�M Cu of just 3 mm (Fig. 3E). Amajor component of rosette diameter is the length ofthe petiole. To distinguish between Cu effects on cellexpansion and cell division, we determined the sizesof petiole/vascular and mesophyll cells. Petiole cellsof plants exposed to 100�M Cu were on average 2.4times smaller than control ones. Similarly, mesophyllcells of Cu-treated plants were significantly smallerthan those of control plants (Fig. 3F). This resultsin a significant size reduction in leaves, petioles andcotyledons (Fig. 3G). Thus, we conclude that Cu ef-fects on leaf and rosette diameter are largely due toeffects on cell expansion. The observation that there isno clear Cu-effect on either the timing of leaf forma-
tion or the number of leaves formed (six true leavesby day 24 irrespective of Cu), strongly suggests thatCu does not significantly impact on cell division inthe shoot.
The next principal stage in theArabidopsis lifecycle is the formation and elongation of the inflores-cence stem. In the control plants, the inflorescencebecame visible at 21 days after seed soaking. How-ever, in plants treated with 30�M Cu the appearanceof the inflorescence was delayed and it became firstvisible at day 35 (not shown).
Thus, Cu induced changes in plant architecture arecomplex, comprising both stimulatory (germination,root/leaf formation and lateral root formation) and in-hibitory (root elongation, leaf expansion) effects. Itis not clear whether these morphogenic effects area direct consequence of molecular damage, or ratherpart of the physiological acclimation response of thestressed plant. To investigate this, we have exposedA.thaliana plants to salicylic acid, a signaling interme-diate involved in the responses of plants to pathogens(Delaney et al., 1994), wounding, and heat shock (Datet al., 1998).
3.2. Salicylic acid induce architectural anddevelopmental changes
We found that low concentrations of SA (0.05–0.2 mM) promoted the early stage of seed germination,i.e. testa rupture (Fig. 4A). SA effects on the growthof seedlings were more complex. Roots of SA-treatedplants were found to be relatively short (Fig. 4B),with the root hairs being localized close to the roottip. Lateral root formation was also accelerated inSA-treated plants (Fig. 4C). In a particularly extremecase, 0.2 mM SA was found to block root elongationwhile numerous short lateral roots were being formeddirectly behind the root tip (Fig. 4D). This pheno-type is very similar to that observed in Cu-treatedseedlings.
SA did not impact on the progression of the leafforming reactions in the shoot apex. The first trueleaves emerged at 5 days after germination irrespectiveof SA exposure. Also, SA did not impede the num-ber of leaves formed, rather we noted a small enhanc-ing effect in the early stages of growth. For example,at 9 days there are on average 3.4 leaves on controlplants and 3.8 leaves on plants treated with 0.1 mM
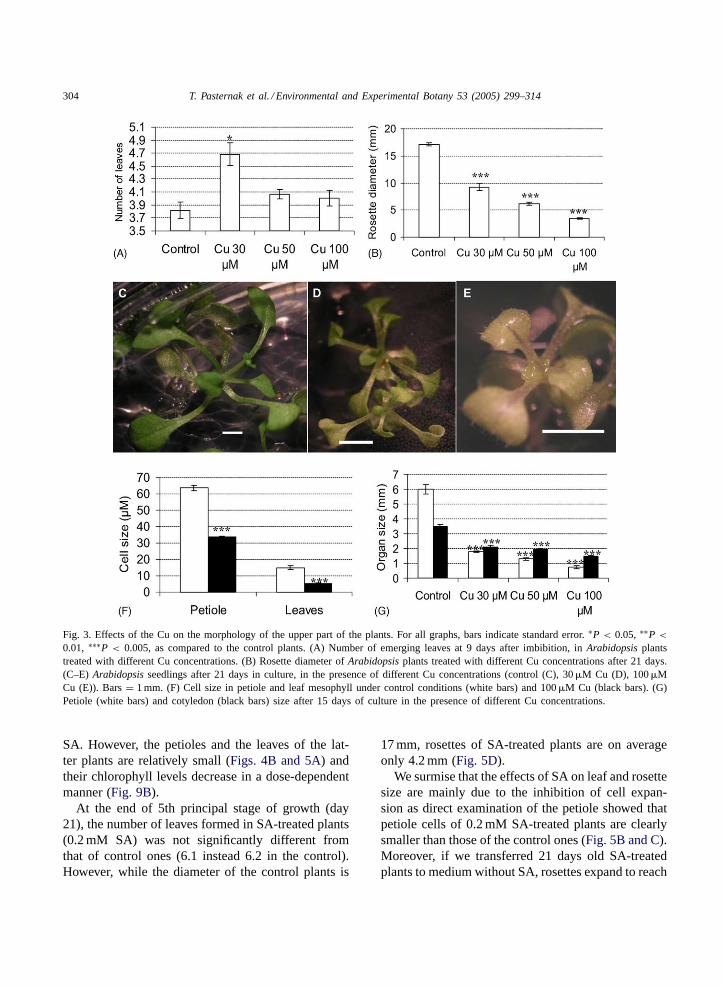
304 T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314
Fig. 3. Effects of the Cu on the morphology of the upper part of the plants. For all graphs, bars indicate standard error.∗P < 0.05, ∗∗P <
0.01, ∗∗∗P < 0.005, as compared to the control plants. (A) Number of emerging leaves at 9 days after imbibition, inArabidopsis plantstreated with different Cu concentrations. (B) Rosette diameter ofArabidopsis plants treated with different Cu concentrations after 21 days.(C–E) Arabidopsis seedlings after 21 days in culture, in the presence of different Cu concentrations (control (C), 30�M Cu (D), 100�MCu (E)). Bars= 1 mm. (F) Cell size in petiole and leaf mesophyll under control conditions (white bars) and 100�M Cu (black bars). (G)Petiole (white bars) and cotyledon (black bars) size after 15 days of culture in the presence of different Cu concentrations.
SA. However, the petioles and the leaves of the lat-ter plants are relatively small (Figs. 4B and 5A) andtheir chlorophyll levels decrease in a dose-dependentmanner (Fig. 9B).
At the end of 5th principal stage of growth (day21), the number of leaves formed in SA-treated plants(0.2 mM SA) was not significantly different fromthat of control ones (6.1 instead 6.2 in the control).However, while the diameter of the control plants is
17 mm, rosettes of SA-treated plants are on averageonly 4.2 mm (Fig. 5D).
We surmise that the effects of SA on leaf and rosettesize are mainly due to the inhibition of cell expan-sion as direct examination of the petiole showed thatpetiole cells of 0.2 mM SA-treated plants are clearlysmaller than those of the control ones (Fig. 5B and C).Moreover, if we transferred 21 days old SA-treatedplants to medium without SA, rosettes expand to reach
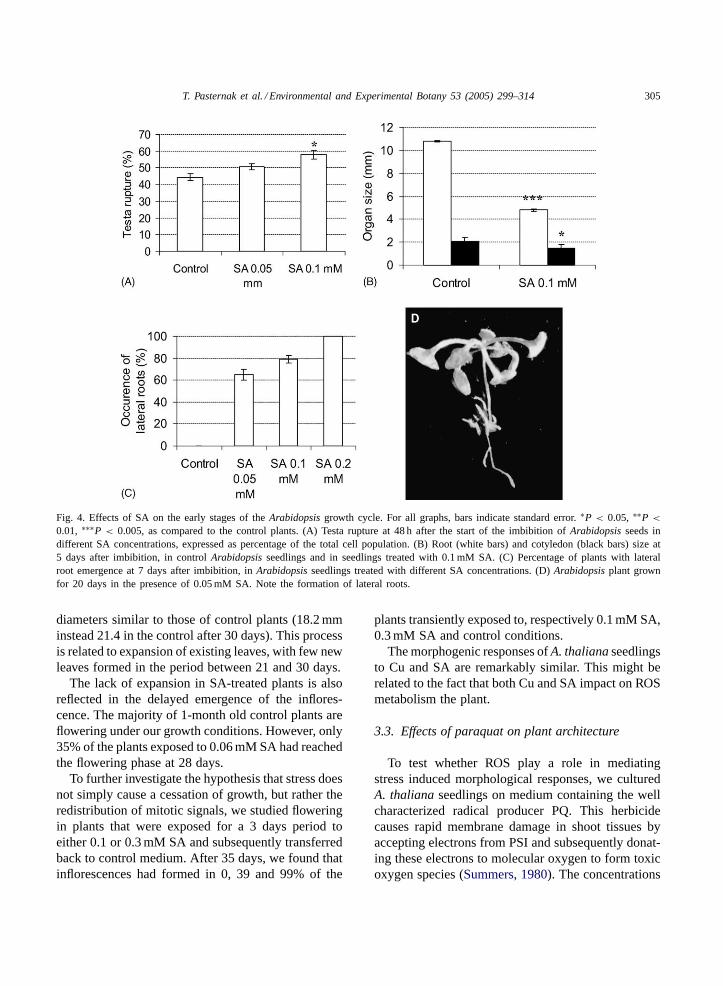
T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314 305
Fig. 4. Effects of SA on the early stages of theArabidopsis growth cycle. For all graphs, bars indicate standard error.∗P < 0.05, ∗∗P <
0.01, ∗∗∗P < 0.005, as compared to the control plants. (A) Testa rupture at 48 h after the start of the imbibition ofArabidopsis seeds indifferent SA concentrations, expressed as percentage of the total cell population. (B) Root (white bars) and cotyledon (black bars) size at5 days after imbibition, in controlArabidopsis seedlings and in seedlings treated with 0.1 mM SA. (C) Percentage of plants with lateralroot emergence at 7 days after imbibition, inArabidopsis seedlings treated with different SA concentrations. (D)Arabidopsis plant grownfor 20 days in the presence of 0.05 mM SA. Note the formation of lateral roots.
diameters similar to those of control plants (18.2 mminstead 21.4 in the control after 30 days). This processis related to expansion of existing leaves, with few newleaves formed in the period between 21 and 30 days.
The lack of expansion in SA-treated plants is alsoreflected in the delayed emergence of the inflores-cence. The majority of 1-month old control plants areflowering under our growth conditions. However, only35% of the plants exposed to 0.06 mM SA had reachedthe flowering phase at 28 days.
To further investigate the hypothesis that stress doesnot simply cause a cessation of growth, but rather theredistribution of mitotic signals, we studied floweringin plants that were exposed for a 3 days period toeither 0.1 or 0.3 mM SA and subsequently transferredback to control medium. After 35 days, we found thatinflorescences had formed in 0, 39 and 99% of the
plants transiently exposed to, respectively 0.1 mM SA,0.3 mM SA and control conditions.
The morphogenic responses ofA. thaliana seedlingsto Cu and SA are remarkably similar. This might berelated to the fact that both Cu and SA impact on ROSmetabolism the plant.
3.3. Effects of paraquat on plant architecture
To test whether ROS play a role in mediatingstress induced morphological responses, we culturedA. thaliana seedlings on medium containing the wellcharacterized radical producer PQ. This herbicidecauses rapid membrane damage in shoot tissues byaccepting electrons from PSI and subsequently donat-ing these electrons to molecular oxygen to form toxicoxygen species (Summers, 1980). The concentrations

306 T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314
Fig. 5. Effects of SA on the later stages of theArabidopsis growth cycle. For all graphs, bars indicate standard error.∗P < 0.05, ∗∗P <
0.01, ∗∗∗P < 0.005, as compared to the control plants. (A) Size ofArabidopsis petioles (black bars) and leaves (white bars) after 25 daysof cultivation in control medium or medium supplemented with 0.2 mM SA. (B) Petiole cells elongation at 25 days after seeds soaking.(C) Arabidopsis plantlet size after 25 days; left: control, right: treated with 0.2 mM SA. Bar= 1 mm. (D) Arabidopsis rosette diameter infunction of SA treatment, at 21 days after imbibition.
we used either slightly enhance chlorophyll levels(0.01–0.1�M) or induce modest bleaching (Fig. 9A).This gives an indication for the non-lethal characterof the treatment used here. PQ does not promotethe early stage of seed germination i.e. testa rupture(Fig. 6A), probably because PQ acts on the electrontransport in working chloroplasts. Root elongationand cotyledon expansion are significantly impededin PQ-treated plants (Fig. 6B), while root hairs areformed in a zone directly adjacent to the root tip. Lat-eral root formation is significantly increased in thePQ-treated seedlings. For example, after 6 days inculture 65% plants exposed to 0.2�M PQ had lateralroots (Fig. 6C).
The timing of the appearance of the first true leaveswas not affected by PQ (5 days after germination).However, the number of leaves formed is slightly, butnot significantly, enhanced by PQ. For example, at 12
days there are on average 4.25 leaves on control plantsand 4.31 leaves on plants treated with 0.15�M PQ.However, the leaves of PQ-treated plants are signifi-cantly smaller, than those of control ones (not shown).At the end of the expansion stage (day 18), the rosettediameter of PQ-treated plants is significantly smallerthan that of control plants (Fig. 6D). The appear-ance of the inflorescence was delayed by PQ treat-ment (Fig. 6E). Thus, the morphogenic responses ofA. thaliana seedlings to PQ are remarkably similar tothose induced by Cu and SA.
3.4. Effects of tert-butylhydroperoxide on seedlingdevelopment
The similarity of the morphogenic effects inducedby Cu, SA and PQ suggests a role for ROS in control-ling plant architecture. We have further tested this idea

T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314 307
Fig. 6. Effects of PQ onArabidopsis morphogenesis. For all graphs, bars indicate standard error.∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.005,as compared to the control plants. (A) Testa rupture (expressed as a percentage of the total population) at 48 h after imbibition of theseeds in control medium or medium supplemented with 0.2 mM PQ. (B) Root (white bars) and cotyledon (black bars) size at 5 days afterimbibition of the seeds in control medium or medium supplemented with 0.2 mM. (C)Arabidopsis plantlet after 7 days in culture. Notethe lateral root emergence. (D) Rosette diameter after 21 days in control plants and in plants treated with 0.2 mM PQ. (E) Inflorescenceoutgrowth (expressed as a percentage of the total population) after 25 days of treatment with different concentrations of PQ.
by exposing plants directly to ROS, which, in planta,are an intrinsic part of the response to various bioticand abiotic stresses. We have used both hydrogen per-oxide as well as its analogue, tBHP as direct-actingoxidative-stress inducing agents. Both compoundsinduce the same effects. The concentrations tBHPthat we used, either stimulate or decrease chlorophylllevels in seedlings (not shown), comparable to whatwas observed in PQ or SA-treated plants (Fig. 9).Concentrations of 0.3 and 0.6 mM tBHP promote
the early stage of seed germination, i.e. testa rupture(data not shown). We also observed significant tBHPeffects on root length and morphology. Roots of tBHPtreated plants are shorter, as are the hypocotyls andthe cotyledons (Fig. 7A). Root hairs are formed in azone relatively close to the root cap (Fig. 7D). Theformation of lateral roots is also significantly accel-erated in tBHP treated plants. We observed that 95%of the plants exposed to 0.6 mM tBHP had developedlateral roots after 6 days of culture. At that stage, no

308 T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314
Fig. 7. Addition of tBHP to the medium of developingArabidopsis plants affects their morphology. For all graphs, bars indicate standarderror. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.005, as compared to the control plants. (A) Root (white bars), hypocotyls (grey bar) or cotyledon(black bar) size in control plantlets or in plants treated wit 0.3 mM tBHP. (B) Rosette diameter ofArabidopsis plants treated with differenttBHP concentrations for 30 days. (C) Inflorescence outgrowth in plants treated with different tBHP concentrations, expressed as a percentageof the population. (D) Root tip of anArabidopsis plant treated with 0.2 mM tBHP for 7 days. Note the absence of a proper elongationzone between meristem and root hairs. (E) Root tip of anArabidopsis plant treated with 0.6 mM tBHP for 7 days. Instead of one elongatedprimary root, different lateral roots have formed, and neither primary nor lateral roots elongate.
lateral roots could be observed in control plantlets(Fig. 7E).
The timing of the de novo leaf formation was notaffected by peroxide exposure. Moreover, the numberof leaves formed is not affected by tBHP. However,leaves from peroxide treated plants were smallerthan those of controls. Following 18 days of growth,rosettes of tBHP treated plants were significantlysmaller (Fig. 7B). The emergence of the inflores-cence was delayed in tBHP treated plants (Fig. 7C).Thus, the morphogenic effects induced by a hydrogenperoxide analogue are very similar to those inducedby Cu, SA and PQ. These data strongly suggest arole for active oxygen species in the control of plantarchitecture.
3.5. The polar auxin transport inhibitor TIBAmimics the effects of stress
The alterations in plant architecture induced by Cu,SA and PQ resemble those induced by phytohormonesand which involve a re-distribution of mitogenic sig-nals. Auxin in particular has been implicated in theinhibition of cell expansion as well as the induction oflateral roots. We found that the synthetic polar auxintransport inhibitor TIBA does have similar effects onplant architecture as the aforementioned stress condi-tions. Thus, TIBA promoted germination (Fig. 8A),inhibited root elongation and cotyledon expansion(Fig. 8B), and root hairs were located relativelyclose to the root-tip (Fig. 8C). Moreover, rosettes

T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314 309
of TIBA-treated plants after 25 days in the culturewere relatively small (Fig. 8E). The number of leavesformed is apparently slightly but not significantly en-hanced by TIBA. For example, at 12 days there are on
Fig. 8. Effects of the TIBA onArabidopsis plants. For all graphs, bars indicate standard error.∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.005,as compared to the control plants. (A) Testa rupture in germinatingArabidopsis seeds treated with different TIBA concentrations at 48 hafter imbibition. (B) Root (white bars) and cotyledon (black bars) size ofArabidopsis plants cultivated in the presence of different TIBAconcentrations at 5 days after imbibition. (C) Root morphology in TIBA treated plant (30�M). Note the number and the localization oflateral roots and the excessive number of root hairs. (D) Inflorescence occurrence inArabidopsis plants after 30 days of cultivation in thepresence of different TIBA concentrations, expressed as a percentage of the total population. (E) Rosette diameter ofArabidopsis plantstreated with different TIBA concentrations for 21 days. (F) Size of the cells of petiole of leaf mesophyll of the second true leaf, in controlplants (white bars) plants treated with 30�M TIBA (black bars), at 30 days of cultivation.
average 4.25 leaves on control plants and 4.69 leaveson plants treated with 5�M TIBA. Flowering wasalso delayed in TIBA-treated plants; at 25 days theinflorescence was clearly visible in 90% of the control

310 T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314
plants, but only in 23% of the plants treated with15�M TIBA ( Fig. 8D). However, in sharp contrast toprevious treatments very few lateral roots were formedin plants treated with 10 or 20�M TIBA ( Fig. 8E) atthe basal part of the root, but they emerged closer tothe apical meristem (Fig. 8C). Instead we observeda proliferation of callus. TIBA concentrations used,slightly enhance chlorophyll levels (data not shown).
4. Discussion
4.1. Oxidative stress and morphogenesis
Plants resort to a range of distinct acclimationstrategies in response to abiotic environmental stressessuch as high salt, dehydration, cold, heat, and exces-sive osmotic pressure. Acclimation responses includechanges in physiological and biochemical processesas well as morphological and developmental patterns(Eerens et al., 1998; Malinowski and Belesky, 2000;Schikora and Schmidt, 2001; Bahaji et al., 2002).Stress treatments can modify root architectural traitssuch as root branching, total root length, and roothair formation. Despite the important implications foragriculture, it has remained an enigma what phys-iological and molecular events are responsible forarchitectural alternations. In this paper we have de-tailed how a set of architectural parameters is specifi-cally altered in response to oxidative stress inducingagents and the auxin transport inhibitor TIBA, inA.thaliana seedlings grown under in vitro conditions.All agents tested, triggered a redistribution of growthrather than a general retardation of growth. Thus,exposure to copper, PQ, SA or tBHP induced a fasttransition from a tap root system to a fibrous one.The very strong similarities between the architecturalchanges induced by Cu and SA (Figs. 1–4), suggesta common molecular-physiological response system.Exposure to Cu has been widely reported to resultin increasing in the levels of reactive oxygen species(ROS) (Hartley-Whitaker et al., 2001; Raeymaekerset al., 2003). Salicylic acid, which accumulates inresponse to abiotic or biotic stresses, interferes withROS metabolism via inhibitory effects on catalase andperoxidases activity (Chen et al., 1993). To test theinvolvement of ROS in stress induced morphogenesis,we interfered directly in plant ROS metabolism. PQ
directly mediates the formation of ROS via the inter-action with mitochondrial and chloroplastic electrontransfer systems (cf.Ye and Gressel, 1994), while hy-drogen peroxide is a ROS. The morphogenic effectsinduced by these two compounds are identical tothose induced by Cu and SA. Thus, taking into con-sideration the oxidative character of these stresses, weconsider it very likely that ROS are playing a key rolein controlling the architectural changes in the stressedA. thaliana seedlings. Consistent with our data, wenote that an ascorbate deficient mutant (vtc1) is char-acterized by slow growth, relatively small rosettes andretarded flowering (Veljovic-Jovanovic et al., 2001).Growth of the APX-1 mutant (cytoplasmic ascorbateperoxidase) is similarly affected, i.e. a retardationof growth and late flowering, although leaf numbersare not affected (Pnueli et al., 2003). It is commonlybelieved, that exposure to many abiotic and bioticstress causes enhanced ROS production, includingheat stress (Dat et al., 1998; Larkindale and Knight,2002), UV-radiation stress (Brosche and Strid, 2003),photoinhibition (Muller-Moule et al., 2003), heavymetal stress (Pinto et al., 2003) and anoxia (Blokhinaet al., 2001). Thus, while our hypothesis on the role ofROS in controlling plant architecture refers directlyto our experiments whereby seedlings were exposedto Cu, SA, tBHP or PQ applied to the medium, wethink it likely that similar control mechanisms wouldoperate when plants are exposed to other abiotic andbiotic stresses. Experiments withNicotiana tabacum,andMedicago sativa indicated that the stress inducedalterations observed inA. thaliana could be readilydetected in the case ofNicotiana andMedicago (datanot shown), emphasising the ubiquitous character ofthe response system. We surmise that the functionalimportance of the root-stress induced reorientation ofgrowth is related to stress avoidance. Soil is normallya highly heterogeneous matrix. Thus, the primarystress effect, the formation of lateral roots, may serveto redirect root development away from a local sourceof xenobiotics.
The molecular-physiological mechanism involvedin translating changes in oxidative stress in to changesin plant architecture might be hypothesized to be theoxidative cross linking of cell wall components. Suchcross-linking reactions would impinge directly on thecapacity of the cell to expand. However, we think itis unlikely that such a cross-linking mechanism is

T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314 311
a primary factor controlling stress induced morpho-genesis. In our studies the oxidative stress inducedphenotype is a result of a re-distribution of growthrather than a retardation of growth. This point is ex-emplified by the stress-induced acceleration of lateralroot formation (Figs. 2C, 4C, 6C and 7E), the ac-celeration of germination as well as by increases inshoot/root ratio. Additional evidence to support theconcept of growth re-distribution comes from ourlong-term growth experiments. Thus, after 2 monthsof cultivation, Cu-treated plants remained vegetativeand formed additional leaves, while control plants hadbolted, were setting seed and leaves were senescing(data not shown).
4.2. Phytohormones and stress inducedmorphogenesis
Redistribution of growth does reflect changes inhormonal homeostasis. Several hormones, includ-ing cytokinin, auxin, ethylene and gibberellic acid,impinge on the elongation, cell division and re-orientation of plant growth. We have focused onauxin as this phytohormone (often in conjunction withethylene) plays a key role in both altering primaryroot growth and in promoting root hair and lateralroot formation (Torrey, 1976; Casimiro et al., 2001),which were all found to be altered in this study. In-deed, the general similarity in phenotype in termsof redistribution of growth, between stressed plants(Figs. 1–7), and TIBA-treated plants (Fig. 8), stronglysuggest that changes in auxin metabolism play a rolein controlling stress induced morphological changes.Similarities between stressed plants and TIBA-treatedplants include accelerated testa rupture, decreasedroot elongation, increased root hair density, decreasedleaf size, inhibition of mesophyll cell expansion, un-altered timing of leaf formation, delayed flowering,and fluctuations in chlorophyll content (Fig. 9). Nei-ther treatment influences the de novo leaf formationsignificantly. The overall similarity between stressand TIBA treatment is punctuated by a difference inlateral root formation. Stressed plants form lateralroots (Figs. 1–7), while TIBA-treated plants do not(Fig. 8). The plant hormone auxin plays a criticalrole in lateral root initiation (Bhalerao et al., 2002;Lopez-Bucio et al., 2002). Exogenous application ofauxin stimulates lateral root formation (Evans et al.,
Fig. 9. Effects of PQ (A) and SA (B) on chlorophyll contents inArabidopsis plants. For all graphs, bars indicate standard error.∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.005, as compared to thecontrol plants. Bars/left axis: chlorophyll contents, expressed as apercentage, compared to the chlorophyll level in the control plants(100%). Line plot/right axis: chlorophyll a to b ratio.
1994) and some auxin-resistant mutants have reducednumbers of lateral roots (Malamy and Benfey, 1997).Mutants that accumulate auxin to high levels pro-duce excess lateral roots (Boerjan et al., 1995; Kinget al., 1995; Delarue et al., 1998) and this is also thecase for some mutants with altered auxin distribution(Marchant et al., 2002). The capacity and pattern-ing for lateral organ formation is directly related toauxin patterning and/or distribution (Reindhart et al.,2003). Indeed, inhibition of polar auxin transport by1-naphthylphthalamic acid (NPA) and TIBA inhibitslateral root formation (Casimiro et al., 2001) (Fig. 8E)and this reflects the accumulation of auxin in the rootapex, and the depletion in the pericycle (Casimiroet al., 2001). Conversely, we surmise that the in-creased formation of lateral roots in stressed plantsreflects local auxin accumulation near the pericyclecells. We also note the formation of callus in root tipsin TIBA treated plants, rather than of cortical cells(data not shown). This reflects the continuation ofcell division activity of cortical cells in TIBA treated

312 T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314
plants and is fully consistent with the local accumu-lation of auxin in the cortex (Casimiro et al., 2003).No callus is formed in the roots of Cu, SA, tBHP orPQ stressed plants, where root growth has decreased,but where lateral root formation is increased. Thus,we surmise that differences in lateral root formationbetween stressed (Cu, SA, tBHP and PQ) plants andTIBA treated plants are likely to reflect spatial dif-ferences in stress- or TIBA-induced indoleacetic acidgradients.
The inhibitory effect of stress on cell and leaf expan-sion is consistent with an auxin accumulation effect,although contribution by other phytohormones can notbe excluded. InA. thaliana, leaf expansion is directlycorrelated with auxin concentrations, i.e. decreasingindoleacetic acid concentrations facilitate leaf expan-sion through cell expansion (Ljung et al., 2001a,b).This is also consistent with our earlier work in whichwe showed that the Fe (oxidative) stress treatment ofisolated alfalfa protoplasts did not inhibit cell divisionbut only cell expansion (Pasternak et al., 2002). In-deed, the oxidative stress exerted by an excess of Fe,induced changes in auxin metabolism which in turnaffected the developmental pathway that the proto-plasts followed (Pasternak et al., 2002). In alfalfa pro-toplast cultures, the effects of stress on developmentcould be substituted by high levels of exogenous auxin.However, the mere observation that oxidative stresscan impact on auxin controlled development, createsan avenue for adjustment of cellular metabolism tothe environment. Clearly, it is difficult to translatecellular events directly to whole plant responses. In-deed, at present we do not know how changes in ROSmetabolism are related to local changes in auxin gradi-ents. A survey of recent literature indicated a numberof possible scenarios.
1. Oxidative stress conditions impact directly, at thecellular level, on the auxin controlled cellular de-velopment programme (Pasternak et al., 2002).
2. Class III peroxidases are induced under abioticstress conditions and are capable to oxidativelydegrade indoleacetic acid with the help of H2O2(Østergaard et al., 1998; Jansen et al., 2001; Lee,2002).
3. Both biotic and abiotic stressors can induce pHchanges (Murphy et al., 1998; Raeymaekers et al.,2003). Since indoleacetic acid transport is directly
pH dependent, changes in pH may impact on polarauxin transport.
4. Various phenolics can accumulate in response tostress exposure and impact on polar auxin transportand plant morphology (Brown et al., 2001).
5. The microtubule network, which directly controlscell extension, responds to Al stress (Schwarzerovaet al., 2002) as well as to auxin signaling (Baluskaet al., 1996). Plants are capable of responding tooxidative stress and auxin signals by activating spe-cific MAP kinases (Kovtun et al., 1998; Kovtunet al., 2000) resulting in altered microtubule andcell elongation dynamics (Samaj et al., 2002).
5. Conclusion
We conclude that the morphogenic effects of abioticstress involve an active reorientation of growth. Ourdata suggest that this reorientation is a complex pro-cess that integrates changes in ROS metabolism withchanges in auxin distribution.
Acknowledgements
G.P. is post-doctoral researcher at the Fund for Sci-entific Research in Flanders (F.W.O. – Vlaanderen).Mrs. Inge Van Dyck is gratefully acknowledged forher help in preparing the figures.
References
Allen, R.D., 1995. Dissection of oxidative stress tolerance usingtransgenic plants. Plant Physiol. 107, 1049–1054.
Bahaji, A., Mateu, I., Sanz, A., Cornejo, M., 2002. Commonand distinctive responses of rice seedlings to saline- andosmotically-generated stress. Plant Growth Regul. 38, 83–94.
Baluska, F., Barlow, P.W., Volkmann, D., 1996. Completedisintegration of the microtubular cytoskeleton precedes itsauxin-mediated reconstruction in postmitotic maize root cells.Plant Cell Physiol. 37, 1013–1021.
Bhalerao, R.P., Eklof, J., Ljung, K., Marchant, A., Bennett, M.,Sandberg, G., 2002. Shoot derived auxin is essential for earlylateral root emergence in Arabidopsis seedlings. Plant J. 29,325–332.
Blokhina, O.B., Chirkova, T.V., Fagerstedt, K.V., 2001. Anoxicstress leads to hydrogen peroxide formation in plant cells. J.Exp. Bot. 52, 1179–1190.

T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314 313
Boerjan, W., Cervera, M., Delarue, M., Beeckman, T., Dewitte, W.,Bellini, C., Caboche, M., Van Onckelen, H., Van Montagu, M.,Inzé, D., 1995. Superroot, a recessive mutation in Arabidopsis,confers auxin overproduction. Plant Cell 7, 1405–1419.
Boyes, D.C., Zayed, A.M., Ascenzi, R., McCaskill, A.J., Hoffman,N.E., Davis, K.R., Görlach, J., 2001. Growth stage-basedphenotypic analysis of Arabidopsis: a model for highthroughput functional genomics in plants. Plant Cell 13, 1499–1510.
Britt, A.B., Fiscus, E., 2003. Response of Arabidopsis DNA repairdefective mutants to solar radiation. Physiol. Plant 118, 183–192.
Brosche, N., Strid, A., 2003. Molecular events following perceptionof ultraviolet-B radiation by plants. Physiol. Plant 17, 1–10.
Brown, D.E., Rashotte, A.M., Murphy, A.S., Normanly, J., Tague,B.W., Peer, W.S., Taiz, L., Muday, G.K., 2001. Flavonoids actas negative regulators of auxin transport in vivo in Arabidopsis.Plant Physiol. 126, 524–535.
Casimiro, I., Beeckman, T., Graham, N., Bhalerao, R., Zhang,H., Casero, P., Sandberg, G., Bennett, M., 2003. DissectingArabidopsis lateral root development. Trends Plant Sci. 8, 165–171.
Casimiro, I., Marchant, A., Bhalerao, R., Beeckman, T., Dhooge,S., Swarup, R., Graham, N., Inzé, D., Sandberg, G., Casero,P., Bennett, M., 2001. Auxin transport promotes Arabidopsislateral root initiation. Plant Cell 13, 843–852.
Chen, Z.X., Silva, H., Klessing, D., 1993. Active oxygen species inthe induction of plant systemic acquired resistance by salicylicacid. Science 262, 1883–1886.
Dat, J.F., Foyer, C.H., Scott, I.M., 1998. Changes in salicylicacid and antioxidants during induction of thermotolerance inmustard seedlings. Plant Physiol. 118, 1455–1461.
Davison, P.A., Hunter, C.N., Horton, P., 2002. Overexpressionof beta-carotene hydroxylase enhances stress tolerance inArabidopsis. Nature 418, 203–206.
Delaney, T.P., Uknes, S., Vernooij, B., Friedrich, L., Weymann,K., Negrotto, D., Gaffney, T., Rella, M., Kessmann, H., Ward,E., Ryals, J., 1994. A central role of salicylic acid in plantdisease resistance. Science 266, 1247–1250.
Delarue, M., Prinsen, E., Van Onckelen, H., Caboche, M., Bellini,C., 1998. Sur2 mutations ofArabidopsis thaliana define a newlocus involved in the control of auxin homeostasis. Plant J.12, 603–611.
Eerens, J.P.J., Lucas, R.J., Easton, H.S., White, J.G.H., 1998.Influence of the ryegrass endophyte (Neotyphodium lolii) in acool-moist environment. III. Interaction with white clover NewZeal. J. Agric. Res. 41, 201–207.
Evans, M.L., Ishikawa, H., Estelle, M.A., 1994. Responsesof Arabidopsis roots to auxin studied with high temporalresolution: comparison of wild type and auxin responsemutants. Planta 194, 215–222.
Francis, D., 1998. Environmental contol of the cell cycle inhigher plants. In: Bryant, J.A., Chiatante, D. (Eds.), Plant CellProliferation and its Regulation in Growth and Development.Wiley, New York, pp. 79–97.
Hartley-Whitaker, J., Ainsworth, G., Meharg, A.A., 2001. Copper-and arsenate-induced oxidative stress inHolcus lanatus L.
clones with differential sensitivity. Plant Cell Environ. 24, 713–722.
Jansen, M.A.K., Malan, C., Shaaltiel, Y., Gressel, J., 1990. Mode ofevolved photooxidant resistance to herbicides and xenobiotics.Z. Naturforsch 45, 97–103.
Jansen, M.A.K., Van den Noort, R.E., Tan, A., Prinsen, E.,Lagrimini, M., Thorneley, R.N.F., 2001. Phenol-oxidisingperoxidases play a key role in protecting plants from UV-Bradiation: linking UV-tolerance to changes in leaf and plantarchitecture. Plant Physiol. 126, 1012–1023.
Jouili, H., El Ferjani, E., 2003. Changes in antioxidant andlignifying enzyme activities in sunflower roots (Helianthusannuus L.) stressed with copper excess. C. R. Biologies 326,639–644.
King, J.J., Stimart, D.P., Fisher, R.H., Bleecker, A.B., 1995. Amutation altering auxin homeostasis and plant morphology inArabidopsis. Plant Cell 7, 2023–2037.
Kovtun, Y., Chiu, W.L., Tena, G., Sheen, J., 2000. Functionalanalysis of oxidative stress-activated mitogen-activated proteinkinase cascade in plants. Proc. Natl. Acad. Sci. U.S.A. 97,2940–2945.
Kovtun, Y., Chiu, W.L., Zeng, W., Sheen, J., 1998. Suppressionof auxin signal transduction by a MAPK cascade in higherplants. Nature 395, 716–720.
Larkindale, J., Knight, M.R., 2002. Protection against heatstress-induced oxidative damage in Arabidopsis involvescalcium, abscisic acid, ethylene and salicylic acid. PlantPhysiol. 128, 682–695.
Lee, M.Y., 2002. Effects of Na2SO3 on the activities of antioxidantenzymes in geranium seedlings. Phytochemistry 59, 493–499.
Ljung, K., Bhalerao, R.P., Sandberg, G., 2001a. Sites andhomeostatic control of auxin biosynthesis in Arabidopsis duringvegetative growth. Plant J. 28, 465–474.
Ljung, K., Ostin, A., Lioussanne, L., Sandberg, G., 2001b.Developmental regulation of indole-3-acetic acid turnover inScots pine seedlings. Plant Physiol. 125, 464–475.
Lopez-Bucio, J., Hernandez-Abreu, E., Sanchez-Calderon, L.,Fernanda, M., Nieto-Jacobo, , Simpson, J., Herrera-Estrella,L., 2002. Phosphate availability alters architecture and causeschanges in hormone sensitivity in the Arabidopsis root system.Plant Physiol. 129, 244–256.
Malamy, J.E., Benfey, P.N., 1997. Down and out in Arabidopsis:the formation of lateral roots. Trends Plant Sci. 2, 390–396.
Malinowski, D.P., Belesky, D.P., 2000. Adaptations ofendophyte-infected cool-season grasses to environmentalstresses: mechanisms of drought and mineral stress tolerance.Crop Sci. 40, 923–940.
Marchant, A., Bhalerao, R.P., Casimiro, I., Eklof, J., Casero,P.J., Bennett, M., Sandberg, G., 2002. AUXl promotes lateralroot formation by facilitating indole-3-acetic acid distributionbetween sink and source tissues in the Arabidopsis seedling.Plant Cell 14, 589–597.
Mittler, R., 2002. Oxidative stress, antioxidants and stresstolerance. Trends Plant Sci. 7, 405–410.
Mittler, R., Hallak-Herr, E., Orvar, B.L., Van Camp, W., Willekens,H., Inze, D., Ellis, B., 1999. Transgenic tobacco plants with

314 T. Pasternak et al. / Environmental and Experimental Botany 53 (2005) 299–314
reduced capability to detoxify reactive oxygen intermediatesare hyper-responsive to pathogen infection. Proc. Natl. Acad.Sci. U.S.A. 96, 14165–14170.
Muller-Moule, P., Havaux, M., Niyogi, K.K., 2003. Zeaxanthindeficiency enhances the high light sensitivity of anascorbate-deficient mutant of Arabidopsis. Plant Physiol. 133,748–760.
Murashige, T., Skoog, F., 1962. A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol. Plant 15,473–497.
Murphy, T.M., Asard, H., Cross, A.R., 1998. Possible sources ofreactive oxygen during the oxidative burst in plants. In: Asard,H., Bérczi, A., Caubergs, R.J. (Eds.), Plasma Membrane RedoxSystems and Their Role in Biological Stress and Disease.Kluwer Academic Publishers, Dordrecht, The Netherlands,pp. 215–246.
Østergaard, L., Pedersen, A.G., Jespersen, H.M., Brunak, S.,Welinder, K.G., 1998. Computational analysis and annotationof the Arabidopsis peroxidase gene family. FEBS Lett. 433,98–102.
Pasternak, T., Prinsen, E., Ayaydin, F., Miskolczi, P., Potters, G.,Asard, H., Van Onckelen, H., Dudits, D., Fehér, A., 2002. Therole of auxin, pH and stress in the activation of embryogeniccell division in leaf protoplast-derived cells of alfalfa. PlantPhysiol. 129, 1807–1819.
Patakas, A., Nikolaou, N., Zioziou, E., Radoglou, K., Noitsakis,B., 2002. The role of organic solute and ion accumulation inosmotic adjustment in drought-stressed grapevines. Plant Sci.163, 361–367.
Pinto, E., Sigaud-Kutner, T.C.S., Leitao, M.A.S., Okamoto, O.K.,Morse, D., Colepicolo, P., 2003. Heavy metal-induced oxidativestress in algae. J. Phycol. 39, 1008–1018.
Pnueli, L., Liang, H., Rozenberg, M., Mittler, R., 2003.Growth suppression, altered stomatal response and augmentedinduction of heat shock proteins in cytosolic ascorbateperoxidase (Apx-1)-deficient Arabidopsis plants. Plant J. 34,187–203.
Raeymaekers, T., Potters, G., Asard, H., Guisez, Y., Horemans, N.,2003. Copper-mediated oxidative burst inNicotiana tabacum
L. cv. bright yellow 2 cell suspension cultures. Protoplasma221, 93–100.
Reindhart, D., Pesce, E., Stieger, P., Mansel, T., Baltensperger,K., Bennett, M., Traas, J., Friml, J., Kuhlemeier, C., 2003.Regulation of phyllotaxis by polar auxin transport. Nature 426,255–260.
Samaj, J., Ovecka, M., Hlavacka, A., Lecourieux, F., Meskiene,I., Lichtscheidl, I., Lenart, P., Salaj, J., Volkmann, D.,Bogre, L., Baluska, F., Hirt, H., 2002. Involvement of themitogen-activated protein kinase SIMK in regulation of roothair tip growth. EMBO J. 21, 3296–3306.
Sauge-Merle, S., Cuine, S., Carrier, P., Lecomte-Pradines, C.,Luu, D., Peltier, G., 2003. Enhanced toxic metal accumulationin engineered bacterial cells expressingArabidopsis thalianaphytochelatin synthase. Appl. Environ. Biol. 69, 490–494.
Schikora, A., Schmidt, W., 2001. Iron stress-induced changesin root epidermal cell fate are regulated independently fromphysiological responses to low iron availability. Plant Physiol.125, 1679–1687.
Schwarzerova, K., Zelenkova, S., Nick, P., Opatrny, Z.,2002. Aluminum-induced rapid changes in the microtubularcytoskeleton of tobacco cell lines. Plant Cell Physiol. 43, 207–216.
Summers, L.A., 1980. The Bipyridilium Herbicides. AcademicPress, London, pp. 1–449.
Torrey, J.G., 1976. Root hormones and plant growth. Annu. Rev.Plant Physiol. 27, 435–459.
Veljovic-Jovanovic, S.D., Pignocchi, C., Noctor, G., Foyer,C.H., 2001. Low ascorbic acid in the vtc-1 mutantof Arabidopsis is associated with decreased growth andintracellular redistribution of the antioxidant system. PlantPhysiol. 127, 426–435.
Vernon, L.P., 1960. Spectrophotometric determination ofchlorophyll and pheophytins in plant extracts. Anal. Chem. 32,1144–1146.
Ye, B., Gressel, J., 1994. Constitutive variation of ascorbateperoxidase activity during development parallels that ofsuperoxide dismutase and gluthatione reductase in paraquat-resistant conyza. Plant Sci. 102, 147–151.
Copyright © 2022 FDOKUMEN




















