
Enhanced Tolerance to Multiple Abiotic Stresses in Transgenic Alfalfa Accumulating Trehalose
9
CROP SCIENCE, VOL. 49, SEPTEMBER– OCTOBER 2009 1791 RESEARCH A biotic-stress tolerance is a complex trait that involves tol- erances to drought, flooding, extreme temperatures, soil salin- ity or limited mineral nutrients, and excess or deficit in light. The combinations and severity of these stresses that crops must tolerate vary with environment and year (Bray et al., 2000). Climate change and limited water supply in several regions throughout the world emphasize the need to design different strategies to improve stress tolerance in cultivated plants. Genetic engineering of stress toler- ance has become one of the major goals of agricultural research. At the molecular level, plants adapt to abiotic stresses by the expression of different genes. Their encoded products comprise lipid desaturases, water channels, stress proteins such as LEA (late embryogenesis abundant) proteins and HSP (heat-shock proteins), and enzymes catalyzing the synthesis of antioxidants, ion trans- porters, or osmoprotectants (Bartels and Sunkar, 2005). One of the best-characterized osmoprotectants is the nonreducing disac- charide trehalose that is very efficient at maintaining lipids in a fluid phase in the absence of water, thus avoiding phase separation, leakage, and membrane fusion (Crowe et al., 1992). Trehalose Enhanced Tolerance to Multiple Abiotic Stresses in Transgenic Alfalfa Accumulating Trehalose Ramón Suárez, Cecilia Calderón, and Gabriel Iturriaga* ABSTRACT Alfalfa (Medicago sativa L.) is an important for- age crop in many countries with high biomass production and the third-most-cultivated crop in the United States, with high protein content and potential use as biofuel. Here we describe the use of a chimeric translational fusion of yeast trehalose-6-phosphate synthase (EC 2.4.1.15) and trehalose-6-phosphate phosphatase (EC 3.1.3.12) genes to engineer stress tolerance in alfalfa. The gene fusion was overexpressed using either the constitutive 35S promoter or the stress- regulated rd29A promoter. Transgene insertion in the genome was checked by polymerase chain reaction and transcript expression by reverse transcription polymerase chain reaction. Several independent transgenic plants were selected in the presence of kanamycin and further analyzed. Yeast gene encoding trehalose-6-phosphate syn- thase (TPS1)–yeast gene encoding trehalose-6- phosphate phosphatase (TPS2) gene expression under the 35S promoter led to plants with stunted growth and less biomass, whereas TPS1-TPS2 expression driven by the rd29A promoter improved growth and provoked a significant increase in plant biomass at the foliage level in the different transgenic lines. Trehalose accumulated in all the different lines at similar levels under stress con- ditions. Transgenic plants displayed a significant increase in drought, freezing, salt, and heat toler- ance. This is the first time that genetic engineering of trehalose metabolism is reported in alfalfa and demonstrates that it can improve multiple stress protection in this crop. Thus, the yeast TPS-TPP fusion protein represents a great potential for gen- erating stress-tolerant crop plants for agriculture. Centro de Investigación en Biotecnología, Universidad Autónoma del Estado de Morelos, Av. Universidad 1001, Col. Chamilpa, Cuernavaca Mor. 62209, Mexico. Received 30 Sept. 2008. *Corresponding author ([email protected]). Abbreviations: HPLC, high-performance liquid chromatography; MS, Murashige and Skoog; PCR, polymerase chain reaction; RWC, relative water content; SGWC, soil gravimetric water content; T6P, trehalose-6-phosphate; TPP, trehalose-6-phosphate phosphatase; TPS, trehalose-6-phosphate synthase; TPS1, yeast gene encoding TPS; TPS2, yeast gene encoding TPP. Published in Crop Sci. 49:1791–1799 (2009). doi: 10.2135/cropsci2008.09.0573 © Crop Science Society of America 677 S. Segoe Rd., Madison, WI 53711 USA All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Enhanced Tolerance to Multiple Abiotic Stresses in Transgenic Alfalfa Accumulating Trehalose

CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009 1791
RESEARCH
Abiotic-stress tolerance is a complex trait that involves tol-erances to drought, fl ooding, extreme temperatures, soil salin-
ity or limited mineral nutrients, and excess or defi cit in light. The combinations and severity of these stresses that crops must tolerate vary with environment and year (Bray et al., 2000). Climate change and limited water supply in several regions throughout the world emphasize the need to design diff erent strategies to improve stress tolerance in cultivated plants. Genetic engineering of stress toler-ance has become one of the major goals of agricultural research.
At the molecular level, plants adapt to abiotic stresses by the expression of diff erent genes. Their encoded products comprise lipid desaturases, water channels, stress proteins such as LEA (late embryogenesis abundant) proteins and HSP (heat-shock proteins), and enzymes catalyzing the synthesis of antioxidants, ion trans-porters, or osmoprotectants (Bartels and Sunkar, 2005). One of the best-characterized osmoprotectants is the nonreducing disac-charide trehalose that is very effi cient at maintaining lipids in a fl uid phase in the absence of water, thus avoiding phase separation, leakage, and membrane fusion (Crowe et al., 1992). Trehalose
Enhanced Tolerance to Multiple Abiotic Stresses in Transgenic Alfalfa Accumulating Trehalose
Ramón Suárez, Cecilia Calderón, and Gabriel Iturriaga*
ABSTRACT
Alfalfa (Medicago sativa L.) is an important for-
age crop in many countries with high biomass
production and the third-most-cultivated crop in
the United States, with high protein content and
potential use as biofuel. Here we describe the
use of a chimeric translational fusion of yeast
trehalose-6-phosphate synthase (EC 2.4.1.15)
and trehalose-6-phosphate phosphatase (EC
3.1.3.12) genes to engineer stress tolerance in
alfalfa. The gene fusion was overexpressed using
either the constitutive 35S promoter or the stress-
regulated rd29A promoter. Transgene insertion in
the genome was checked by polymerase chain
reaction and transcript expression by reverse
transcription polymerase chain reaction. Several
independent transgenic plants were selected in
the presence of kanamycin and further analyzed.
Yeast gene encoding trehalose-6-phosphate syn-
thase (TPS1)–yeast gene encoding trehalose-6-
phosphate phosphatase (TPS2) gene expression
under the 35S promoter led to plants with stunted
growth and less biomass, whereas TPS1-TPS2
expression driven by the rd29A promoter improved
growth and provoked a signifi cant increase in
plant biomass at the foliage level in the different
transgenic lines. Trehalose accumulated in all the
different lines at similar levels under stress con-
ditions. Transgenic plants displayed a signifi cant
increase in drought, freezing, salt, and heat toler-
ance. This is the fi rst time that genetic engineering
of trehalose metabolism is reported in alfalfa and
demonstrates that it can improve multiple stress
protection in this crop. Thus, the yeast TPS-TPP
fusion protein represents a great potential for gen-
erating stress-tolerant crop plants for agriculture.
Centro de Investigación en Biotecnología, Universidad Autónoma del
Estado de Morelos, Av. Universidad 1001, Col. Chamilpa, Cuernavaca
Mor. 62209, Mexico. Received 30 Sept. 2008. *Corresponding author
Abbreviations: HPLC, high-performance liquid chromatography;
MS, Murashige and Skoog; PCR, polymerase chain reaction; RWC,
relative water content; SGWC, soil gravimetric water content; T6P,
trehalose-6-phosphate; TPP, trehalose-6-phosphate phosphatase; TPS,
trehalose-6-phosphate synthase; TPS1, yeast gene encoding TPS; TPS2,
yeast gene encoding TPP.
Published in Crop Sci. 49:1791–1799 (2009).doi: 10.2135/cropsci2008.09.0573© Crop Science Society of America677 S. Segoe Rd., Madison, WI 53711 USA
All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.

1792 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009
(α-D-glucopyranosyl-1, 1-α-D-glucopyranoside) also plays a key role in the structural and functional stabili-zation of proteins in the anhydrous state, apparently by means of water replacement of osmoprotectant molecules or formation of a glassy state (Elbein et al., 2003).
Among the fi ve known routes for trehalose biosynthe-sis, the most widely distributed is the trehalose-6-phos-phate synthase (TPS)–trehalose-6-phosphate phosphatase (TPP) pathway that has been reported in several species of bacteria including E. coli, yeast, insects, and “resurrection plants” such as Selaginella lepidophylla (Avonce et al., 2006). Arabidopsis thaliana, rice (Oryza sativa L.), and other higher plants also possess TPS and TPP homologs comprising multigene families, although they do not accumulate sig-nifi cant amounts of trehalose. In recent years a new role has emerged for the intermediate trehalose-6-phosphate (T6P) as a signaling molecule for coordination of carbohydrate metabolism with plant development (Paul et al., 2008).
Trehalose metabolism has been studied in most detail in Saccharomyces cerevisiae. In yeast, trehalose is synthesized by a multienzyme complex formed by TPS encoded by TPS1, TPP encoded by TPS2, and two regulatory sub-units encoded by the TSL1 and TPS3 genes. Although the latter genes do not play a catalytic role, they share sig-nifi cant homology with TPS and TPP (Bell et al., 1998). Trehalose-6-phosphate is synthesized from uridine diphos-phate glucose and glucose-6-phosphate substrates by TPS and dephosphorylated to trehalose by TPP (Gancedo and Flores, 2004). Deletion of the TPS1 or TPS2 gene caused an impairment of stress tolerance (De Virgilio et al., 1994).
The overexpression of trehalose biosynthetic genes in several plants such as tobacco (Nicotiana tabacum L.), potato (Solanum tuberosum L.), rice, Arabidopsis, and tomato (Lycopersicum esculentum Mill.) has led to an improvement of stress tolerance (Holmström et al., 1996; Yeo et al., 2000; Garg et al., 2002; Avonce et al., 2004; Cortina and Culiáñez-Macià, 2005). Recently, we have shown that a chimeric yeast TPS1-TPS2 gene expressed in Arabidopsis confers tolerance to several abiotic stresses (Miranda et al., 2007).
Alfalfa (Medicago sativa L.) is a perennial forage legume of great agronomic interest around the world that reduces atmospheric nitrogen in association with the soil bacte-rium Rhizobium meliloti and is a high biomass crop with potential use as a biofuel ( Jung, 2006). Alfalfa is highly valued as an animal feed because of its high protein con-tent, high intake potential, and digestibility. Given its high nutritive value, alfalfa may serve as the sole plant compo-nent in many livestock feeding programs, which demand high-quality alfalfa (Barnes and Sheaff er, 1985).
Here we report the expression of a bifunctional yeast TPS1-TPS2 gene fusion in transgenic alfalfa plants that demonstrated enhanced tolerance to drought-, heat-, freezing-, and salt-stress conditions.
MATERIALS AND METHODS
Plant Material and Growth ConditionsAlfalfa (Medicago sativa L. cv. Regen SY27x) was used for over-
expressing the TPS1-TPS2 or TPS1 gene constructs. Plants were
routinely grown in a growth chamber on vermiculite soil at 22 to
20°C with a 16/8-h light/dark cycle with cool-white illumination
(80 μmol s–1 m–2) and a relative humidity of 40% and were propa-
gated clonally through stem cuttings that were rooted for around
2 wk in pots with soil and watered with nutrient solution.
Plant TransformationConstructs for plant transformation containing the TPS1-TPS2
gene fusion were reported recently (Miranda et al., 2007). These
constructs (Fig. 1) use as a backbone the pBin19 vector (Bevan,
1984) containing either the 0.8-kb 35S or the 1-kb rd29A pro-
moter (Yamaguchi-Shinozaki and Shinozaki, 1994) with its cor-
responding leader, and the 0.3-kb NOS polyadenylation site. The
constructs were introduced by electroporation in Agrobacterium
tumefaciens LBA4404 strain. The resulting bacteria were used to
transform alfalfa (Austin et al., 1995). Well-developed embryos
were recovered from callus culture on media containing 50 μg
mL−1 kanamycin (Sigma, St. Louis, MO). Five lines—in other
words, independent insertion events—from each construct were
used for the detailed analyses. Rooted plantlets were transferred
to pots with vermiculite soil and watered with 0.5X Murashige
and Skoog (MS) basal salt mixture medium (Sigma, St. Louis,
MO) supplemented with 7.5 mM KNO3.
Genomic and Reverse Transcription Polymerase Chain Reaction AnalysisGenomic DNA from alfalfa was isolated with a DNA isolation
kit (Puregene Gentra System, Minneapolis, MN) and 100 ng
were used for a polymerase chain reaction (PCR) using oli-
gonucleotides corresponding to neomycin phosphotransferase
(NPTII) gene (5′-GAACAAGATGGATTGTTGCACGC and
5′-GAAGAACTCGTCAAGAAGC). The program was one
cycle at 94°C for 3 min; 35 cycles at 94°C for 1 min, 59°C for
45 s, and 72°C for 1 min; and fi nally one cycle at 72°C for 5 min.
Reverse transcription polymerase chain reaction experiments
were performed using 2 μg of total RNA extracted from alfalfa
adult plants using TRIZOL reagent (Invitrogen, Carlsbad, CA)
according to manufacturer’s instructions and used for fi rst-
strand cDNA synthesis with SuperScript II reverse transcriptase
(Invitrogen, Carlsbad, CA) and oligo dT. Polymerase chain
reaction was conducted using oligonucleotides corresponding
to the 1.3-kb TPS2 region of TPS1-TPS2 (5′-CGGGATC-
CGCTAAATCTATTAACATGG3 and 5′-CGGGGTACCA-
TGGTGGGTTGAGAC) and the program was as described
above, except that annealing was at 57°C (1 min) and extension
at 72°C (1.5 min), and only 25 cycles were used, which corre-
sponded to the linearity phase of the exponential reaction after
comparison of the PCR products at diff erent cycles. Polymerase
chain reaction products were resolved in 1X TAE, 1% agarose
gels stained with ethidium bromide (EtBr).
Trehalose DeterminationMature leaves only were randomly selected for trehalose deter-
mination from all the transgenic lines described before, taking

CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009 WWW.CROPS.ORG 1793
2007). Eighteen 35S::TPS1-TPS2 and forty rd29A::TPS1-TPS2 independent alfalfa transgenic lines were obtained after transformation with the A. tumefaciens system.
The integration event from each transgenic line was determined by genomic PCR using NPTII gene as a target. Five independent lines from each construct (lines 19 to 101 for 35S::TPS1-TPS2 and lines 4 to 49 for rd29A::TPS1-TPS2 construct) were randomly selected and used for PCR analy-sis (Fig. 2A). All transgene insertions were of the expected 0.8-kb size. To assay for gene expression of TPS1-TPS2 in transgenic plants, we used reverse transcription polymerase chain reaction. Four 35S::TPS1-TPS2 lines except line 39 expressed the TPS1-TPS2 transgene within a 1.3- to 2-fold range among the diff erent lines (Fig. 2, B and C). The APT1 (adenine ribosyl phosphotransferase) gene was used as a con-stitutive control and showed comparable levels of expression also in the wild-type plants (data not shown). To examine the inducible expression of rd29A::TPS1-TPS2, plants were untreated or dehydrated for 1 h. All fi ve rd29A::TPS1-TPS2 lines expressed the transgene only after drought stress treat-ment and showed a twofold variation among them (Fig. 2, D and E). The endogenous rd29A gene was used as a stress-inducible control (data not shown).
Transgenic TPS1-TPS2 Lines Have Altered BiomassSeveral reports have documented that overexpression of TPS genes of diff erent origin in plants provoke growth and morphological abnormalities with variable severity (Holm-ström et al., 1996; Romero et al., 1997; Goddijn et al., 1997; Yeo et al., 2000; Avonce et al., 2004; Cortina and Culiáñez-Macià, 2005). Therefore, we analyzed possible growth and shape abnormalities in transgenic plants overexpressing yeast TPS-TPP enzyme. Five 35S::TPS1-TPS2 indepen-dent lines showed a slower vegetative growth, although leaf shape and color were similar to untransformed plants (Fig. 3A). In contrast, the rd29A::TPS1-TPS2 plants had similar leaf color and shape but grew faster after comparison with the wild-type plants (Fig. 3A). To further substantiate these morphological observations, the plant biomass was deter-mined. We focused on the aerial part, since it is the agro-nomically important part of the plant. Therefore, foliage
triplicate samples. Extracts were prepared as described before
(Avonce et al., 2004), except that samples were analyzed by high-
performance liquid chromatography (HPLC) using a ZOR-
BAX carbohydrate analysis column (Agilent Technologies, Santa
Clara, CA) eluted with acetonitrile:water (65:35, v/v). Trehalose
was used as a standard to determine the concentration.
Biomass and Water Content AnalysesTo determine plant biomass, foliage from all the transgenic
lines used in the present study were taken from 10 individu-
als and weighed immediately after detaching (fresh weight).
Relative water content (RWC) and soil gravimetric water con-
tent (SGWC) were determined as reported before (Gaxiola et
al., 2001) using triplicate samples. Briefl y, leaves were excised
and their fresh weight was determined. After fl oating them in
deionized water at 4°C overnight, their rehydrated weight was
determined. The dried weight was determined after incubating
leaves in an oven at 70°C overnight. Plant RWC was calculated
by measuring (fresh weight — dry weight)/(rehydrated weight
— dry weight). Soil gravimetric water content was determined
in fully watered soil and after 10 d without watering.
Plant Stress ToleranceTen individuals from 1-mo-old clonally propagated alfalfa
plants, which were randomly selected, were used for all the dif-
ferent stress tolerance experiments performed under controlled
conditions in a growth chamber as described before. For drought
tolerance, watering (always with 50 mL per pot) was withheld
for 5, 10, 20, and 30 d, and then plants were rewatered and
allowed to recover for 2 d before being photographed. Survival
was assessed when wilting reversed after rewatering. Intolerant
plants did not recover turgor by naked eye inspection. For salin-
ity tests, plants were irrigated with MS media supplemented
with diff erent concentrations of NaCl from 50 mM to 300 mM
and left 2 wk without NaCl for recovery. Freezing stress was
conducted on plants preadapted 5 d at 4°C before subjected to
freezing stress at –5, –10, and –15°C for 6, 12, 24, 48, and 72 h.
Plants were left at 24°C during 10 d for recovery. Heat-shock
experiments were conducted either without prior heat pread-
aptation or by subjecting plants to a preadaptation treatment at
37°C during 4 h, before a heat-shock treatment at 40, 45, 50,
and 55°C during 1 h, and allowing 10 d of recovery.
Statistical AnalysisAll experiments were repeated at least two times independently
with their corresponding repeats unless otherwise stated in the
fi gure caption. The data were processed by analysis of variance
by t student test followed by Duncan-Waller mean analysis.
RESULTS
Transformation and Molecular Analysis of Transgenic Alfalfa PlantsTo determine the eff ect of overexpressing the yeast TPS-TPP bifunctional enzyme in plants, the 2.9-kb TPS1-TPS2 gene fusion was subcloned in the pBin19 vector down-stream of the constitutive 35S promoter or under the con-trol of the stress-inducible rd29A promoter (Miranda et al.,
Figure 1. Gene constructs for plant transformation. Cassette maps of
the TPS1–TPS2 translational gene fusion under the control of either
35S or rd29A promoter and the nopaline synthase polyadenylation
site (P). The gene cassettes are downstream of the kanamycin
selectable marker (35S-NPTII) and between the left (LB) and right
(RB) borders of Agrobacterium tumefaciens T-DNA in pBin19 vector.

1794 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009
from greenhouse-grown plants was removed and the fresh weight determined. For the 35S::TPS1-TPS2 lines the bio-mass decreased 21%, whereas for transgenics expressing the gene fusion under the stress-inducible promoter there was
signifi cant increase in biomass of 62% compared with the wild-type plants (Fig. 3B).
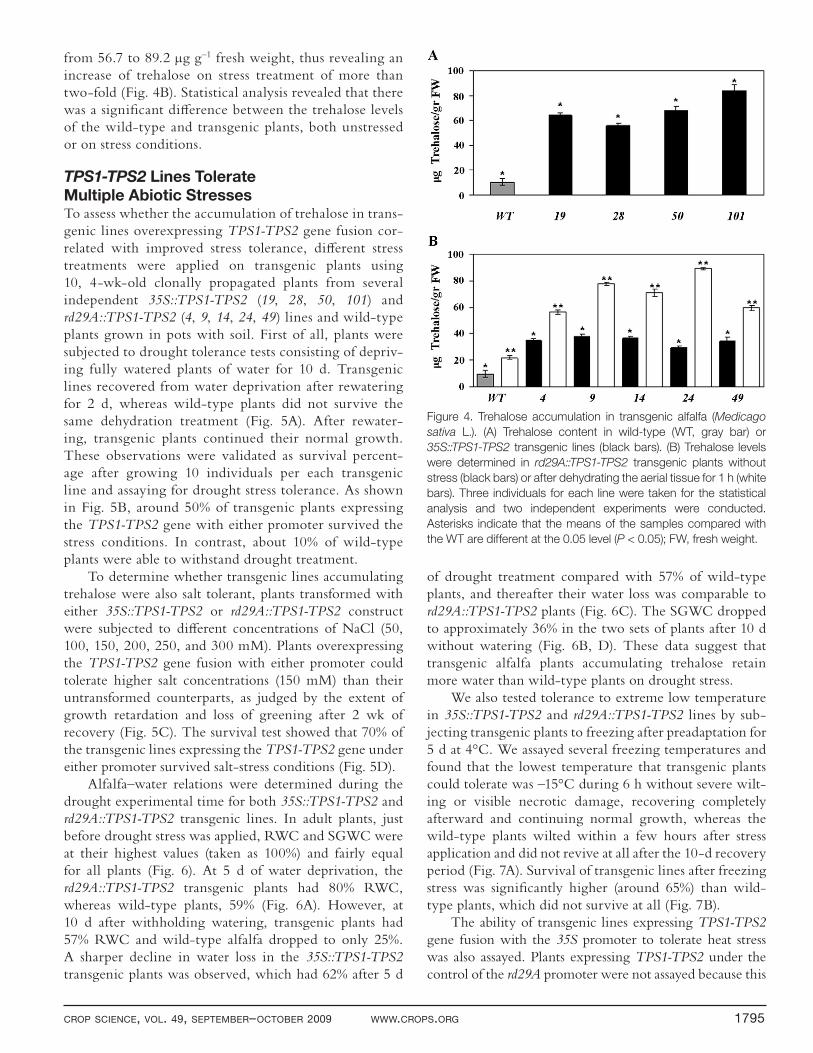
Alfalfa TPS1-TPS2 Lines Accumulate TrehaloseTo analyze the accumulation of trehalose in transgenic plants, carbohydrate analysis was performed by HPLC. Trehalose was detected in the diff erent 35S::TPS1-TPS2 transgenic lines at levels of 55.5 to 83.5 μg g–1 fresh weight, compared with 10.2 μg g–1 fresh weight in the wild-type (Fig. 4A). Interestingly, the transgenic rd29A::TPS1-TPS2 lines without stress accumulated trehalose at three-times higher levels compared with the wild-type plants, sug-gesting a basal and signifi cant expression of the rd29A promoter. Upon desiccation treatment, the rd29A::TPS1-TPS2 lines accumulated trehalose at values varying
Figure 3. Morphology of transgenic alfalfa (Medicago sativa L.)
and biomass determination. (A) One-mo-old clonally propagated
plants transformed with 35S::TPS1-TPS2 (35S-Bif, line 50) or
rd29A::TPS1-TPS2 (RD-Bif, line 9) construct and untransformed
wild-type (WT) plants. (B) Foliage fresh weight (FW) of 1-mo-
old clonally propagated alfalfa plants was determined in three
independent experiments. Percentages were calculated taking the
value of the WT strain as 100%. The white bar denotes wild-type
plants, black bars correspond to 35S::TPS1-TPS2 lines, and gray
bars correspond to rd29A::TPS1-TPS2 plants. Ten individuals for
each line were taken for the statistical analysis. Asterisks indicate
that the means of the samples compared with the WT are different
at the 0.05 level (P < 0.05).
Figure 2. Molecular analyses of transgenic plants. (A) Genomic
polymerase chain reaction (PCR) of alfalfa (Medicago sativa L.) lines
transformed with the 35S::TPS1-TPS2 (lines 19 to 101) or rd29A::TPS1-
TPS2 constructs (lines 4 to 49). The DNA band corresponds to a 0.8-kb
PCR fragment of NPTII selectable marker gene. (+) Is an amplifi cation
control from pBin19 containing NPTII. WT, untransformed wild-type
plants. Gene-expression analysis by reverse transcription PCR of
alfalfa lines transformed with the (B) 35S::TPS1-TPS2 (lines 19 to 101) or
(D) rd29A::TPS1-TPS2 construct (lines 4 to 49) and their corresponding
densitometric analysis (C and E). Total RNA extracted from untreated
plants (B) or plants dehydrated for 1 h (D) was used. The DNA band
corresponds to a 1.3-kb PCR fragment of TPS2 gene.
CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009 WWW.CROPS.ORG 1795
from 56.7 to 89.2 μg g–1 fresh weight, thus revealing an increase of trehalose on stress treatment of more than two-fold (Fig. 4B). Statistical analysis revealed that there was a signifi cant diff erence between the trehalose levels of the wild-type and transgenic plants, both unstressed or on stress conditions.
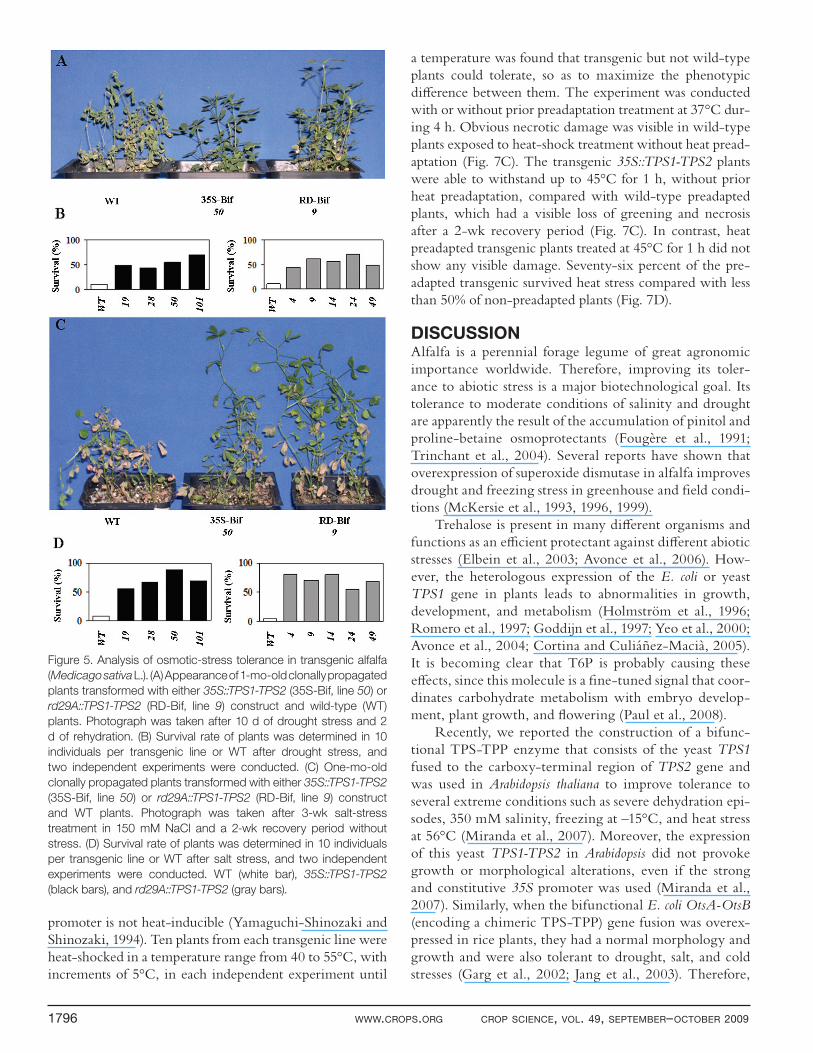
TPS1-TPS2 Lines Tolerate Multiple Abiotic StressesTo assess whether the accumulation of trehalose in trans-genic lines overexpressing TPS1-TPS2 gene fusion cor-related with improved stress tolerance, diff erent stress treatments were applied on transgenic plants using 10, 4-wk-old clonally propagated plants from several independent 35S::TPS1-TPS2 (19, 28, 50, 101) and rd29A::TPS1-TPS2 (4, 9, 14, 24, 49) lines and wild-type plants grown in pots with soil. First of all, plants were subjected to drought tolerance tests consisting of depriv-ing fully watered plants of water for 10 d. Transgenic lines recovered from water deprivation after rewatering for 2 d, whereas wild-type plants did not survive the same dehydration treatment (Fig. 5A). After rewater-ing, transgenic plants continued their normal growth. These observations were validated as survival percent-age after growing 10 individuals per each transgenic line and assaying for drought stress tolerance. As shown in Fig. 5B, around 50% of transgenic plants expressing the TPS1-TPS2 gene with either promoter survived the stress conditions. In contrast, about 10% of wild-type plants were able to withstand drought treatment.
To determine whether transgenic lines accumulating trehalose were also salt tolerant, plants transformed with either 35S::TPS1-TPS2 or rd29A::TPS1-TPS2 construct were subjected to diff erent concentrations of NaCl (50, 100, 150, 200, 250, and 300 mM). Plants overexpressing the TPS1-TPS2 gene fusion with either promoter could tolerate higher salt concentrations (150 mM) than their untransformed counterparts, as judged by the extent of growth retardation and loss of greening after 2 wk of recovery (Fig. 5C). The survival test showed that 70% of the transgenic lines expressing the TPS1-TPS2 gene under either promoter survived salt-stress conditions (Fig. 5D).
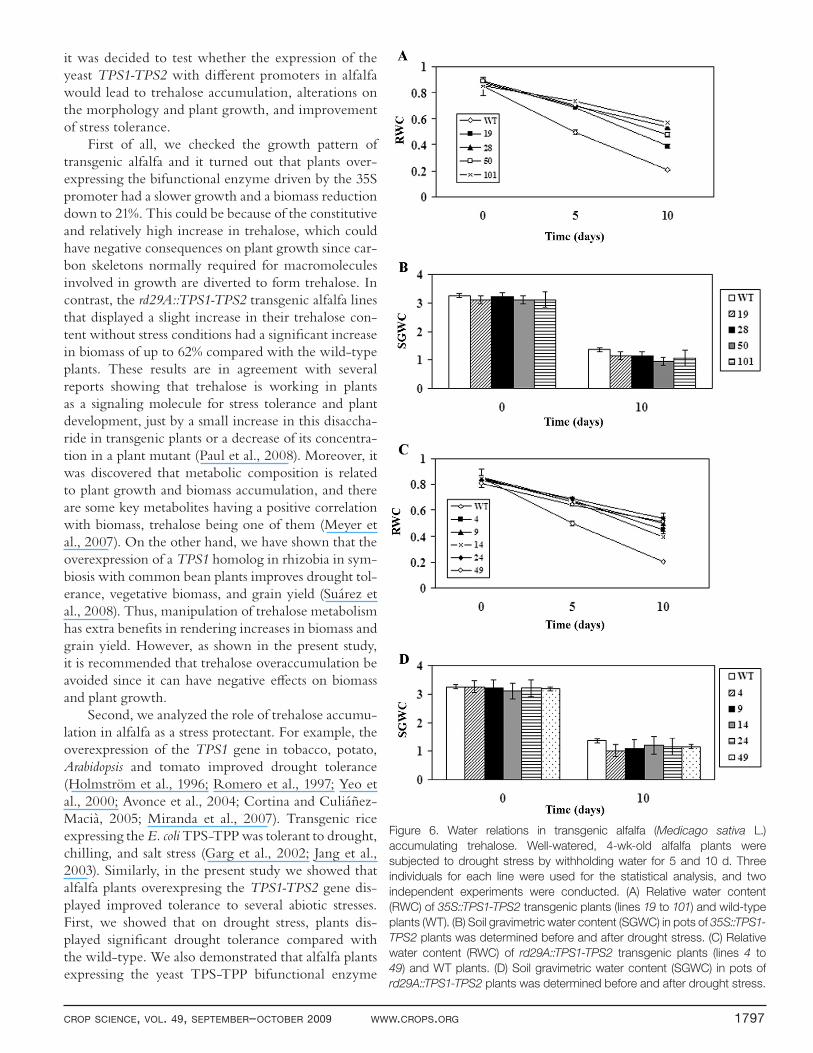
Alfalfa–water relations were determined during the drought experimental time for both 35S::TPS1-TPS2 and rd29A::TPS1-TPS2 transgenic lines. In adult plants, just before drought stress was applied, RWC and SGWC were at their highest values (taken as 100%) and fairly equal for all plants (Fig. 6). At 5 d of water deprivation, the rd29A::TPS1-TPS2 transgenic plants had 80% RWC, whereas wild-type plants, 59% (Fig. 6A). However, at 10 d after withholding watering, transgenic plants had 57% RWC and wild-type alfalfa dropped to only 25%. A sharper decline in water loss in the 35S::TPS1-TPS2 transgenic plants was observed, which had 62% after 5 d
of drought treatment compared with 57% of wild-type plants, and thereafter their water loss was comparable to rd29A::TPS1-TPS2 plants (Fig. 6C). The SGWC dropped to approximately 36% in the two sets of plants after 10 d without watering (Fig. 6B, D). These data suggest that transgenic alfalfa plants accumulating trehalose retain more water than wild-type plants on drought stress.
We also tested tolerance to extreme low temperature in 35S::TPS1-TPS2 and rd29A::TPS1-TPS2 lines by sub-jecting transgenic plants to freezing after preadaptation for 5 d at 4°C. We assayed several freezing temperatures and found that the lowest temperature that transgenic plants could tolerate was –15°C during 6 h without severe wilt-ing or visible necrotic damage, recovering completely afterward and continuing normal growth, whereas the wild-type plants wilted within a few hours after stress application and did not revive at all after the 10-d recovery period (Fig. 7A). Survival of transgenic lines after freezing stress was signifi cantly higher (around 65%) than wild-type plants, which did not survive at all (Fig. 7B).
The ability of transgenic lines expressing TPS1-TPS2 gene fusion with the 35S promoter to tolerate heat stress was also assayed. Plants expressing TPS1-TPS2 under the control of the rd29A promoter were not assayed because this
Figure 4. Trehalose accumulation in transgenic alfalfa (Medicago
sativa L.). (A) Trehalose content in wild-type (WT, gray bar) or
35S::TPS1-TPS2 transgenic lines (black bars). (B) Trehalose levels
were determined in rd29A::TPS1-TPS2 transgenic plants without
stress (black bars) or after dehydrating the aerial tissue for 1 h (white
bars). Three individuals for each line were taken for the statistical
analysis and two independent experiments were conducted.
Asterisks indicate that the means of the samples compared with
the WT are different at the 0.05 level (P < 0.05); FW, fresh weight.
1796 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009
promoter is not heat-inducible (Yamaguchi-Shinozaki and Shinozaki, 1994). Ten plants from each transgenic line were heat-shocked in a temperature range from 40 to 55°C, with increments of 5°C, in each independent experiment until
a temperature was found that transgenic but not wild-type plants could tolerate, so as to maximize the phenotypic diff erence between them. The experiment was conducted with or without prior preadaptation treatment at 37°C dur-ing 4 h. Obvious necrotic damage was visible in wild-type plants exposed to heat-shock treatment without heat pread-aptation (Fig. 7C). The transgenic 35S::TPS1-TPS2 plants were able to withstand up to 45°C for 1 h, without prior heat preadaptation, compared with wild-type preadapted plants, which had a visible loss of greening and necrosis after a 2-wk recovery period (Fig. 7C). In contrast, heat preadapted transgenic plants treated at 45°C for 1 h did not show any visible damage. Seventy-six percent of the pre-adapted transgenic survived heat stress compared with less than 50% of non-preadapted plants (Fig. 7D).
DISCUSSIONAlfalfa is a perennial forage legume of great agronomic importance worldwide. Therefore, improving its toler-ance to abiotic stress is a major biotechnological goal. Its tolerance to moderate conditions of salinity and drought are apparently the result of the accumulation of pinitol and proline-betaine osmoprotectants (Fougère et al., 1991; Trinchant et al., 2004). Several reports have shown that overexpression of superoxide dismutase in alfalfa improves drought and freezing stress in greenhouse and fi eld condi-tions (McKersie et al., 1993, 1996, 1999).
Trehalose is present in many diff erent organisms and functions as an effi cient protectant against diff erent abiotic stresses (Elbein et al., 2003; Avonce et al., 2006). How-ever, the heterologous expression of the E. coli or yeast TPS1 gene in plants leads to abnormalities in growth, development, and metabolism (Holmström et al., 1996; Romero et al., 1997; Goddijn et al., 1997; Yeo et al., 2000; Avonce et al., 2004; Cortina and Culiáñez-Macià, 2005). It is becoming clear that T6P is probably causing these eff ects, since this molecule is a fi ne-tuned signal that coor-dinates carbohydrate metabolism with embryo develop-ment, plant growth, and fl owering (Paul et al., 2008).
Recently, we reported the construction of a bifunc-tional TPS-TPP enzyme that consists of the yeast TPS1 fused to the carboxy-terminal region of TPS2 gene and was used in Arabidopsis thaliana to improve tolerance to several extreme conditions such as severe dehydration epi-sodes, 350 mM salinity, freezing at –15°C, and heat stress at 56°C (Miranda et al., 2007). Moreover, the expression of this yeast TPS1-TPS2 in Arabidopsis did not provoke growth or morphological alterations, even if the strong and constitutive 35S promoter was used (Miranda et al., 2007). Similarly, when the bifunctional E. coli OtsA-OtsB (encoding a chimeric TPS-TPP) gene fusion was overex-pressed in rice plants, they had a normal morphology and growth and were also tolerant to drought, salt, and cold stresses (Garg et al., 2002; Jang et al., 2003). Therefore,
Figure 5. Analysis of osmotic-stress tolerance in transgenic alfalfa
(Medicago sativa L.). (A) Appearance of 1-mo-old clonally propagated
plants transformed with either 35S::TPS1-TPS2 (35S-Bif, line 50) or
rd29A::TPS1-TPS2 (RD-Bif, line 9) construct and wild-type (WT)
plants. Photograph was taken after 10 d of drought stress and 2
d of rehydration. (B) Survival rate of plants was determined in 10
individuals per transgenic line or WT after drought stress, and
two independent experiments were conducted. (C) One-mo-old
clonally propagated plants transformed with either 35S::TPS1-TPS2
(35S-Bif, line 50) or rd29A::TPS1-TPS2 (RD-Bif, line 9) construct
and WT plants. Photograph was taken after 3-wk salt-stress
treatment in 150 mM NaCl and a 2-wk recovery period without
stress. (D) Survival rate of plants was determined in 10 individuals
per transgenic line or WT after salt stress, and two independent
experiments were conducted. WT (white bar), 35S::TPS1-TPS2
(black bars), and rd29A::TPS1-TPS2 (gray bars).
CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009 WWW.CROPS.ORG 1797
it was decided to test whether the expression of the yeast TPS1-TPS2 with diff erent promoters in alfalfa would lead to trehalose accumulation, alterations on the morphology and plant growth, and improvement of stress tolerance.
First of all, we checked the growth pattern of transgenic alfalfa and it turned out that plants over-expressing the bifunctional enzyme driven by the 35S promoter had a slower growth and a biomass reduction down to 21%. This could be because of the constitutive and relatively high increase in trehalose, which could have negative consequences on plant growth since car-bon skeletons normally required for macromolecules involved in growth are diverted to form trehalose. In contrast, the rd29A::TPS1-TPS2 transgenic alfalfa lines that displayed a slight increase in their trehalose con-tent without stress conditions had a signifi cant increase in biomass of up to 62% compared with the wild-type plants. These results are in agreement with several reports showing that trehalose is working in plants as a signaling molecule for stress tolerance and plant development, just by a small increase in this disaccha-ride in transgenic plants or a decrease of its concentra-tion in a plant mutant (Paul et al., 2008). Moreover, it was discovered that metabolic composition is related to plant growth and biomass accumulation, and there are some key metabolites having a positive correlation with biomass, trehalose being one of them (Meyer et al., 2007). On the other hand, we have shown that the overexpression of a TPS1 homolog in rhizobia in sym-biosis with common bean plants improves drought tol-erance, vegetative biomass, and grain yield (Suárez et al., 2008). Thus, manipulation of trehalose metabolism has extra benefi ts in rendering increases in biomass and grain yield. However, as shown in the present study, it is recommended that trehalose overaccumulation be avoided since it can have negative eff ects on biomass and plant growth.
Second, we analyzed the role of trehalose accumu-lation in alfalfa as a stress protectant. For example, the overexpression of the TPS1 gene in tobacco, potato, Arabidopsis and tomato improved drought tolerance (Holmström et al., 1996; Romero et al., 1997; Yeo et al., 2000; Avonce et al., 2004; Cortina and Culiáñez-Macià, 2005; Miranda et al., 2007). Transgenic rice expressing the E. coli TPS-TPP was tolerant to drought, chilling, and salt stress (Garg et al., 2002; Jang et al., 2003). Similarly, in the present study we showed that alfalfa plants overexpresing the TPS1-TPS2 gene dis-played improved tolerance to several abiotic stresses. First, we showed that on drought stress, plants dis-played signifi cant drought tolerance compared with the wild-type. We also demonstrated that alfalfa plants expressing the yeast TPS-TPP bifunctional enzyme
Figure 6. Water relations in transgenic alfalfa (Medicago sativa L.)
accumulating trehalose. Well-watered, 4-wk-old alfalfa plants were
subjected to drought stress by withholding water for 5 and 10 d. Three
individuals for each line were used for the statistical analysis, and two
independent experiments were conducted. (A) Relative water content
(RWC) of 35S::TPS1-TPS2 transgenic plants (lines 19 to 101) and wild-type
plants (WT). (B) Soil gravimetric water content (SGWC) in pots of 35S::TPS1-
TPS2 plants was determined before and after drought stress. (C) Relative
water content (RWC) of rd29A::TPS1-TPS2 transgenic plants (lines 4 to
49) and WT plants. (D) Soil gravimetric water content (SGWC) in pots of
rd29A::TPS1-TPS2 plants was determined before and after drought stress.

1798 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009
could tolerate up to 150 mM of NaCl. Overexpression in alfalfa of the Alfi n1 transcription factor gene allowed plant growth in up to 171 mM NaCl and also promoted root growth (Winicov, 2000).
Additionally, in the present study we found that 35S::TPS1-TPS2 and rd29A::TPS1-TPS2 lines could
tolerate 6 h at –15°C, after preadaptation for 5 d at 4°C, without severe wilting or visible necrotic damage and showed complete recovery and continued growth. Simi-larly, freezing treatments from –8 to –16°C to alfalfa plants overexpressing a superoxide dismutase increased twice the plant regrowth after stress (McKersie et al., 1993). Also, it has been shown that accumulation of trehalose in trans-genic rice allowed recovery of plants after 6 h of 4°C treat-ment ( Jang et al., 2003) or 3 d at 10°C (Garg et al., 2002). Finally, we found that transgenic lines expressing the TPS1-TPS2 gene fusion tolerated heat stress. They were able to withstand up to 45°C during 1 h, without prior heat pre-adaptation. As far as we know, this is the fi rst report of improving heat tolerance in alfalfa. Therefore, the use of the yeast TPS1-TPS2 gene fusion allows survival of alfalfa at extreme temperatures, drought, or salt stress.
In summary, we have shown that engineering tre-halose metabolism in alfalfa Regen SY27x by using a bifunctional TPS1-TPS2 gene fusion can increase its abil-ity to withstand several abiotic stresses in greenhouse tests. However, it would be important to demonstrate if this is the case in other alfalfa genotypes of agronomic impact and in fi eld trials before this technology can be applied.
AcknowledgmentsWe are indebted to Dr. Georgina Hernández (Centro de Cien-
cias Genómicas-UNAM, Cuernavaca, Mexico) for her kind
gift of alfalfa plants. Gabriel Iturriaga received funding from
CONACYT-Mexico (No. 2004-CO1-46078) and CYTED-
Spain (107PIC0312), and Ramón Suárez from PROMEP-
Mexico (No. PTC-137).
ReferencesAustin, S., E.T. Bingham, D.E. Matthews, M.N. Shahan, J. Will,
and R.R. Burgess. 1995. Production and fi eld performance of
transgenic alfalfa (Medicago sativa L.) expressing alpha-amylase and
manganese-dependent lignin peroxidase. Euphytica 85:381–393.
Avonce, N., B. Leyman, J.O. Mascorro-Gallardo, P. Van Dijck, J.
Thevelein, and G. Iturriaga. 2004. The Arabidopsis trehalose-
6P-synthase AtTPS1 gene is a regulator of glucose, ABA and
stress signaling. Plant Physiol. 136:3649–3659.
Avonce, N., A. Mendoza-Vargas, E. Morett, and G. Iturriaga.
2006. Insights on the evolution of trehalose biosynthesis.
BMC Evol. Biol. 6:109.
Barnes, D.K., and C.C. Sheaff er. 1985. Alfalfa. In M.E Heath, R.F
Barnes, and D.S. Metcalfe (ed.) Forages: The science of grass-
land agriculture. 4th ed. Iowa State Univ. Press, Ames.
Bartels, D., and R. Sunkar. 2005. Drought and salt tolerance in
plants. Crit. Rev. Plant Sci. 24:23–58.
Bell, W., W. Sun, S. Hohmann, S. Wera, A. Reinders, C. De Vir-
gilio, A. Wiemken, and J.M. Thevelein. 1998. Composition
and functional analysis of the Saccharomyces cerevisiae trehalose
synthase complex. J. Biol. Chem. 273:33311–33319.
Bevan, M. 1984. Binary Agrobacterium vectors for plant transforma-
tion. Nucleic Acids Res. 12:8711–8721.
Bray, E.A., J. Bailey-Serres, and E. Weretilnyk. 2000. Responses
to abiotic stresses. In W. Gruissem, B. Buchanan, and R. Jones
Figure 7. Tolerance to extreme temperatures in transgenic
alfalfa (Medicago sativa L.). (A) Appearance of 1-mo-old clonally
propagated plants transformed with either 35S::TPS1-TPS2
(35S-Bif, line 50) or rd29A::TPS1-TPS2 (RD-Bif, line 9) construct and
wild-type (WT) plants. Photograph was taken after preadapting 5
d at 4°C, freezing stress at –15°C during 6 h and 10 d of recovery.
(B) Survival rate of plants was determined in 10 individuals per
transgenic line or WT after freezing stress, and two independent
experiments were conducted. WT (white bar), 35S::TPS1-TPS2
(black bars), and rd29A::TPS1-TPS2 (gray bars). (C) One-mo-old
clonally propagated plants transformed with the 35S::TPS1-TPS2
(35S-Bif, line 50) construct and wild-type (WT) plants. Photograph
was taken after preadapting (*) at 37°C during 4 h or not, heat
stress at 45°C during 1 h, and a 10-d recovery period. (D) Survival
rate of plants was determined in 10 individuals per transgenic line
or WT after heat stress, and two independent experiments were
conducted. WT (white bar), 35S::TPS1-TPS2 preadapted (black
bars), and 35S::TPS1-TPS2 not preadapted (gray bars).

CROP SCIENCE, VOL. 49, SEPTEMBER–OCTOBER 2009 WWW.CROPS.ORG 1799
(ed.) Biochemistry and molecular biology of plants. American
Society of Plant Physiologists, Rockville, MD.
Cortina, C., and F.A. Culiáñez-Macià. 2005. Tomato abiotic
stress enhanced tolerance by trehalose biosynthesis. Plant Sci.
169:75–82.
Crowe, J.H., F.A. Hoekstra, and L.M. Crowe. 1992. Anhydrobio-
sis. Annu. Rev. Physiol. 54:579–599.
De Virgilio, C., T. Hottiger, J. Domínguez, T. Boller, and A.
Wiemken. 1994. The role of trehalose synthesis for the acqui-
sition of thermotolerance in yeast: I. Genetic evidence that tre-
halose is a thermoprotectant. Eur. J. Biochem. 219:179–186.
Elbein, A.D., Y.T. Pan, I. Pastuszak, and D. Carroll. 2003. New
insights on trehalose: A multifunctional molecule. Glycobiol-
ogy 13:17R–27R.
Fougère, F., D. Le Rudulier, and J.G. Streeter. 1991. Eff ects of salt
stress on amino acid, organic acid, and carbohydrate composi-
tion of roots, bacteroids, and cytosol of alfalfa (Medicago sativa
L.). Plant Physiol. 96:1228–1236.
Gancedo, C., and C.L. Flores. 2004. The importance of a func-
tional trehalose biosynthetic pathway for the life of yeasts and
fungi. FEMS Yeast Research. 4:351–359.
Garg, A.K., J.-K. Kim, T.G. Owens, A.P. Ranwala, Y.D. Choi,
L.V. Kochian, and R. Wu. 2002. Trehalose accumulation in
rice plants confers high tolerance levels to diff erent abiotic
stresses. Proc. Natl. Acad. Sci. USA 99:15898–15903.
Gaxiola, R.A., J. Li, S. Undurraga, L.M. Dang, G.J. Allen, S.L.
Alper, and G.R. Fink. 2001. Drought- and salt-tolerant plants
result from overexpression of the AVP1 H+-pump. Proc. Natl.
Acad. Sci. USA 98:11444–11449.
Goddijn, O.J.M., T.C. Verwoerd, E. Voogd, R.W.H.H. Krutwa-
gen, P.T.H.M. de Graaf, J. Poels, K. van Dun, A.S. Ponstein,
B. Damm, and J. Pen. 1997. Inhibition of thehalase activity
enhances trehalose accumulation in transgenic plants. Plant
Physiol. 113:181–190.
Holmström, K.O., E. Mäntylä, B. Welin, A. Mandal, E.T. Palva,
O.E. Tunnela, and J. Londesborough. 1996. Drought toler-
ance in tobacco. Nature 379:683–684.
Jang, I.-C., S.-J. Oh, J.-S. Seo, W.-B. Choi, S.I. Song, C.H. Kim,
Y.S. Kim, H.-S. Seo, Y.D. Choi, B.H. Nahm, and J.-K. Kim.
2003. Expression of a bifunctional fusion of the Escherichia
coli genes for trehalose-6-phosphate synthase and trehalose-
6-phosphate phosphatase in transgenic rice plants increases
trehalose accumulation and abiotic stress tolerance without
stunting growth. Plant Physiol. 131:516–524.
Jung, H. 2006. Alfalfa: A sustainable crop for biomass energy pro-
duction. USDA–ARS, Plant Sci. Res. Unit, St. Paul, MN.
McKersie, B.D., S.R. Bowley, E. Harjanto, and O. Leprince. 1996.
Water-defi cit tolerance and fi eld performance of transgenic
alfalfa overexpressing superoxide dismutase. Plant Physiol.
111:1177–1181.
McKersie, B.D., S.R. Bowley, and K.S. Jones. 1999. Winter sur-
vival of transgenic alfalfa overexpressing superoxide dis-
mutase. Plant Physiol. 119:839–847.
McKersie, B.D., Y. Chen, M. de Beus, S.R. Bowley, C. Bowler,
D. Inze, K. D’Halluin, and J. Botterman. 1993. Superoxide
dismutase enhances tolerance of freezing stress in transgenic
alfalfa (Medicago sativa L.). Plant Physiol. 103:1155–1163.
Meyer, R.C., M. Steinfath, J. Lisec, M. Becher, H. Witucka-Wall,
O. Törjék, O. Fiehn, A. Eckardt, L. Willmitzer, J. Selbig, and
T. Altmann. 2007. The metabolic signature related to high
plant growth rate in Arabidopsis thaliana. Proc. Natl. Acad. Sci.
USA 104:4759–4764.
Miranda, J.A., N. Avonce, R. Suárez, J.M. Thevelein, P. Van Dijck,
and G. Iturriaga. 2007. A bifunctional TPS-TPP enzyme from
yeast confers tolerance to multiple and extreme abiotic-stress
conditions in transgenic Arabidopsis. Planta 226:1411–1421.
Paul, M.J., L.F. Primavesi, D. Jhurreea, and Y. Zhang. 2008. Trehalose
metabolism and signaling. Annu. Rev. Plant Biol. 59:417–441.
Romero, C., J.M. Belles, J.L. Vaya, R. Serrano, and F.A. Culia-
ñez-Macià. 1997. Expression of the yeast trehalose-6-phos-
phate synthase gene in transgenic tobacco plants: Pleiotropic
phenotypes include drought tolerance. Planta 201:293–297.
Suárez, R., A. Wong, M. Ramírez, A. Barraza, M. del C. Orozco,
M.A. Cevallos, M. Lara, G. Hernández, and G. Iturriaga.
2008. Improvement of drought tolerance and grain yield in
common bean by overexpressing trehalose-6-phosphate syn-
thase in rhizobia. Mol. Plant Microbe Interact. 21:958–966.
Trinchant, J.C., A. Boscari, G. Spennato, G. Van de Sype, and D. Le
Rudulier. 2004. Proline betaine accumulation and metabolism
in alfalfa plants under sodium chloride stress. Exploring its com-
partmentalization in nodules. Plant Physiol. 135:1583–1594.
Winicov, I. 2000. Alfi n1 transcription factor overexpression
enhances plant root growth under normal and saline conditions
and improves salt tolerance in alfalfa. Planta 210:416–422.
Yamaguchi-Shinozaki, K., and K. Shinozaki. 1994. A novel cis-
acting element in an Arabidopsis gene is involved in respon-
siveness to drought, low-temperature, or high-salt stress.
Plant Cell 6:251–264.
Yeo, E.-T., H.-B. Kwon, S.-E. Han, J.-T. Lee, J.-C. Ryu, and
M.-O. Byun. 2000. Genetic engineering of drought resistant
potato plants by introduction of the trehalose-6-phosphate
synthase (TPS1) gene from Saccharomyces cerevisiae. Mol. Cells
10:263–268.




















