
Nuclear Envelope Remnants: Fluid Membranes Enriched in STEROLS and Polyphosphoinositides

1993 82: 3580-3591
N Falla, Vlasselaer Van, J Bierkens, B Borremans, G Schoeters and U Van Gorp population of the murine bone marrowCharacterization of a 5-fluorouracil-enriched osteoprogenitor
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
reserved.Copyright 2011 by The American Society of Hematology; all rights900, Washington DC 20036.weekly by the American Society of Hematology, 2021 L St, NW, Suite Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom

Characterization of a 5-Fluorouracil-Enriched Osteoprogenitor Population of the Murine Bone Marrow
By N. Falla, P. Van Vlasselaer, J. Bierkens, B. Borremans, G. Schoeters, and U. Van Gorp
In the presence of j3-glycerophosphate and vitamin C, cul- tures of normal mouse bone marrow cells form three-di- mensional structures that stain positive with the Von Kossa technique and express alkaline phosphatase (ALP), collagen type I, and osteocalcin. Little is known about the characteristics and frequency of the cells that contribute to this phenomenon. Most likely, mature osteoblastic cells do not contribute to the nodule formation because no osteo- calcin expressing cells are detected in the flushed marrow by in situ hybridization. Limiting dilution analysis shows that, in normal bone marrow, 1 of 2.2 X 1 O5 cells has the potency to form a bone nodule and to express ALP, colla- gen, and osteocalcin in a temporal fashion. Upon in vivo treatment with 5-fluorouracil (5-FU), this frequency in- creases 1 2-fold, eg, 1 in 1.75 X 1 O4 cells shows osteo- genic activity. In comparison, fibroblast colony forming
ONE MARROW (BM) stroma forms a network of fi- B brbblasts, adipocytes, endothelial cells, and macro- phageP that supports and regulates hematopoiesis and is believed to harbor the osteogenic lineage."' The fact that BM differentiates into bone ossicles when transplanted under the kidney capsule or when cultured in intraperitone- ally implanted diffusion chambers supports this idea.'-" Al- though BM represents a unique tissue for studying the ontog- eny of osteogenic cells, little is known about the frequency, morphology, phenotype, and functional characteristics of these cells. This is partly caused by the low frequency of stromal cells in the marrow and the absence of markers for each stage of progression through the lineage.
It is clear that the delineation of differentiation steps in the osteogenic lineage is crucial in determining the restric- tion points in the differentiation of pluripotent cells12 and in defining the target populations for the activity of onco- g e n e ~ ~ ~ and growth factors.I4 Therefore, the need exists for the identification of defined lineage markers and the estab- lishment of cell separation and enrichment techniques that help in the purification of differentiation stages throughout the bone lineage. A step in that direction was the develop- ment of in vitro long-term stroma cultures in which the
From Vlaamse Instelling voor Technologisch Onderzoek (VITO), the Department of Environment, Division of Biology, Boeretang Mol Belgium, Geel. Belgium.
Submitted October 26, 1992; accepted August 10, 1993. N.F. was supported by a grant of the Vlaams Actieprogramma
Biotechnologie (VLAB/034), a programme of the Flemish Commu- nity.
Address reprint requests to Peter Van Vlasselaer, MD, VITO, Department of Environment, Division of Biology, Steenweg op Re- tie, 2440 Geel, Belgium.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 0 1993 by The American Society of Hematology. 0006-4971/93/8212-0014$3.00/0
cells occur at a frequency of 1 of 2.5 X 1 O4 or 1 of 5 X 1 O3 plated cells in normal or 5-FU-treated marrow, respec- tively. Using density centrifugation, the majority of the os- teoprogenitor cells in 5-FU marrow are found in the low- density (1.066 to 1.067 g/mL) fractions. In addition, these cells bind to nylon wool but not to plastic and aggregate in the presence of wheat germ agglutinin and soybean agglu- tinin. Scanning and transmission electron microscopy shows that the bone nodules in 5-FU marrow cultures are composed of fibroblastoid cells embedded in a mineralized collagen matrix. In conclusion, our results show that a qui- escent cell population in the murine bone marrow with fi- broblastoid characteristics contributes to the formation of bone-like nodules in vitro. 0 1993 by The American Society of Hematology.
osteoprogenitor population could be expanded.12*'5s20 How- ever, so far, few attempts were made to fractionate the BM to determine and characterize the cell population responsi- ble for in vitro osteogenic activity.
For the purification of hematopoietic stem cells from the BM, in vivo treatment with 5-fluorouracil (5-FU) has been widely used as an initial enrichment procedure. This drug drastically reduces the number of cycling cells but spares the quiescent cells of the marrow. This nucleotide analogue ex- erts its debilitating effect via incorporation into DNA dur- ing the S-phase of the cell cycle, which ultimately results in cell death of cycling ~ e l l s . ~ ' - ~ ~
In analogy with the purification of hematopoietic stem cells from the BM, an enrichment procedure for osteopro- genitor cells was developed based on in vivo treatment with 5-FU. The intention of this procedure was to determine the frequency, biophysical characteristics (eg, density, adhe- sion, and lectin binding), and the morphology of osteopro- genitor cells of the BM. In comparison, the frequency of fibroblast colony-forming cells was determined in normal and 5-FU-treated BM.
MATERIALS AND METHODS
Mice and BM cell preparation. Eight- to 10-week-old BALB/c mice were obtained from the animal facility of the Vlaamse Instel- ling voor Technologisch Onderzoek (VITO). The mice were killed and the femora removed and placed in phosphate-buffered saline (PBS). The ends of each femur were excised and the marrow ex- pelled by ejecting a stream of PBS into the medullary cavity. The marrow was collected and dispersed until a single-cell suspension was obtained. Debris and remaining aggregates were removed by passing the cell suspension over a 100-mesh stainless steel. Red blood cells were removed by density centrifugation on 70% Percoll (Pharmacia, Uppsala, Sweden).
The animals were weighed and 5-FU (Roche) was administered at a dose of 150 mg/kg body weight by tail vein injection. Five days later, the animals were killed by cervical dislo- cation and their BM was processed as described above.
Cell culture. BM cells were resuspended in Iscove's medium (GIBCO, Grand Island, NY) supplemented with 10% fetal calf serum (FCS), L-Glu, penicillin and streptomycin, ascorbic acid
5-FU treatment.
3580 Blood, Vol82. No 12 (December 15). 1993: pp 3580-3591
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom

OSTEOPROGENITOR CELLS OF MOUSE BONE MARROW 3581
(100 pg/mL), and 8-glycerophosphate (0.6% wt/wt) and plated in flat-bottom 96-well plates at different cell numbers as stated in the text. Every 3 days, 50% of the medium was replaced with fresh medium.
Histologic procedures. Cultures were fixed in neutral buffered formalin and selected areas of the cell layer were removed, decalci- fied with EDTA, embedded in glycol methacrylate, and sectioned in 3 pm. The sections were stained with hematoxylin and eosin. Undecalcified femur shafts were fixed in glycol methacrylate (JB-4 Polysciences) and sectioned in 3 pm. These sections were used for in situ hybridization and were counter-stained with Giemsa.
Determination ofjbroblast colony-forming units (CFU-F). The number of CFU-F was scored as described elsewhere.24 Briefly, 5- FU-treated BM cells were prepared as described above and plated in 35-mm culture dishes (Falcon) at 5 X IO4 cells per dish. Replicate cultures were incubated at 37°C for I O days and fed once by total replacement of the medium. Adherent cells were fixed with metha- nol and stained with May-Grunwald-Giemsa. Colonies of adher- ent, fibroblastoid cells were scored under a binocular (20X).
Plastic and nylon wool adherence. Adherence to tissue culture plastic was performed as described elsewhere?’ Briefly, BM cells, starting at a density of IO’ cells, were allowed to adhere twice to the bottom of T-75 plastic cell culture flasks (Falcon) for 1 hour at 37°C. The nonadherent cells were collected and washed with Iscove’s medium. Nylon wool adherence was performed as de- scribed elsewhere?6
Density centrifugation. Isotonic percoll was prepared according to the vendor’s guidelines and diluted with PBS to final concentra- tions of 7O%, 60%, 55%, 50%, 45%, 40%, and 30%. A discontinuous density gradient was established. The cells were slowly layered on top and subsequently centrifuged for 5 minutes at 1,000 rpm, fol- lowed by 15 minutes at 2,000 rpm. The cells were collected from the interfaces and washed twice with Iscove’s medium.
Wheat germ agglutinin (WGA) and soybean agglutinin (SBA) agglutination. Marrow cells ( 2 X lo6 cells/ml; 0.5 mL) were incu- bated with either SBA or WGA as described el~ewhere.~’.~’ Briefly, the cells were incubated in SBA (2 mg/mL in PBS) or WGA ( I mg/mL in PBS) for 30 minutes at room temperature and then gently layered on top of bovine serum albumin (BSA; 5% wt/vol in PBS). After 30 minutes of incubation at room temperature, the agglutinated cells were sedimented and the unagglutinated cells re- mained at the interface with the BSA. Top and bottom fractions were collected separately and the SBA- and WGA-agglutinated cells were resuspended in D-galactose (0.2 mol/L in PBS) or N-acetyl glucosamine (0.2 mol/L in PBS), respectively. After 15 minutes at 3 7 T , the remaining aggregates were dissociated by pipetting until a single-cell suspension was obtained. The cells were washed with PBS before being put in culture.
Osteocalcin cell enzyme-linked immunosorbent assay (ELISA). The cultures were rinsed with PBS, fixed with 4% cold formalde- hyde for 30 minutes at 4”C, and then washed twice with 0.05 mol/L Tris/HCI, pH 7.6. Endogenous peroxidase activity was blocked by 5 minutes of incubation at room temperature with 3% H202. The samples were rinsed with Tris/HCl, pH 7.6, and blocked with nor- mal goat serum (l/5 diluted in Tris/HCl, pH 7.6) for 1 hour at 37°C. Rabbit-antimouse osteocalcin (kindly provided by Dr R. Bouillon, Katholieke Universiteit, Leuven, Belgium) was added at a dilution of 1/5,000 and incubated for 2 hours at 4°C. Nonbound serum was removed and the cultures were washed five times with Tris/HCl buffer. Horseradish peroxidase-conjugated goat antirab- bit Ig serum (1/2,000 diluted in Tris/HCI, pH 7.6; Tago, Burlin- game, CA) was added for 30 minutes at 4°C. After intensive rinsing, the cultures were incubated at 37°C in the presence of substrate solution (ABTS [Sigma, St Louis, MO] 1 mg/mL + H202 0.1 pL/
mL in citric acid/Na2HP0,, 10.5 g/14.2 g/500 mL H20) until color- ation occurred. Absorbance was read in a microtiter reader at 450 nm, with a reference of 650 nm. As a control for the nonspecific binding of the antimouse osteocalcin antiserum, nonimmune rab- bit serum was used under the same conditions. The amount of osteocalcin incorporated in the culture was determined in compari- son with standard ELISA of purified mouse osteocalcin (kindly provided by Dr R. Bouillon). The sensitivity of this assay was 0.3 ng/mL for osteocalcin, whereas no reactivity was observed with FCS.
DNA measurement. Cultures were washed with PBS and frozen and thawed in the presence of H,O. A sample was mixed with 2 mL of TNE buffer (0.1 mol/L NaCI, 10 mmol/L Tris-HCI, 1 mmol/L EDTA, pH 7.4) supplemented with Hoechst dye 33258 and incu- bated for 1 hour at room temperature. DNA was measured accord- ing to manufacturer’s guidelines in a Hoefler TKO 100 DNA Fluo- rometer. Calibration of the fluorometer was performed using a standard of mouse thymus DNA. A sensitivity of 5 ng/mL DNA was obtained.
Alkaline phosphatase (ALP) activity. ALP enzyme activity was determined as described elsewhere.29 The cultures were incubated with 0.1 mol/L sodium acetate solution supplemented with 0.1 % Triton X-100 and 5 mmol/L p-nitrophenol phosphate (Sigma 104), pH 9.6, for 1 hour at 37°C. Absorbance was determined at 405 nm and compared with a pnitrophenol standard titration curve. ALP activity was expressed as nanomoles of pnitrophenol formed per minute in relation with the DNA contents per well.
Von Kossa staining and calcium determination. Cultures were fixed with methanol and stained following the method of Von Kossam with 5% AgN03. Calcium levels were determined as de- scribed elsewhere.” The cell cultures were washed three times with Ca2+- and Mg2’-free PBS and incubated overnight at room tempera- ture in 0.6 N HCl. The extract was transferred to a 96-well plate and calcium was complexed with o-cresol-phthalein-complexon (Test Combination Calcium; Boehringer Mannheim, Mannheim, Ger- many). The colorimetric reaction was read at 570 nm in a spectro- photometer. The absolute calcium concentration was determined in comparison with a calibrated standard provided by the vendor and related to the DNA content per well.
Collagen synthesis. Collagen synthesis was measured as the in- corporation of (’H)-proline (Amersham, Amersham, UK) into col- lagenase-digestible protein (CDP).32 The collagenase was purchased from Worthington (NJ) and was substantially free of nonspecific protease activity. The cell cultures were exposed to (’H)-proline ( 1 pCi/well) for 18 hours at 37°C and then washed three times with PBS. Collagenase was added (0.1 mg/mL in PBS) for 1 hour and the CDP was measured in a liquid scintillation counter. The amount of CDP was related to the DNA content per well.
Scanning and transmission electron microscopy. For scanning electron microscopy, the cultures were rinsed with PBS and fixed for 1 hour with 2.5% sodium cacodylate in 0.1 mol/L phosphate buffer (pH 7.2). After rinsing, the cultures were postfixed for 1 hour with 1% osmium tetroxide in the same buffer. The cultures were subsequently rinsed and progressively dehydrated with alcohol. They were processed for critical point drying (Balzers Union, Liech- tenstein) in C02 and then coated with gold (50 nm; Balzers Union). The cultures were observed in a JEOL JSM-Fl5 microscope (Ja- pan). For transmission electron microscopy, the cultures were washed once with PBS and then fixed for 2 hours in a glutaralde- hyde (3%), paraformaldehyde ( I % ) , sucrose (5%) solution in so- dium-cacodylate buffer (0. l mol/L, pH 7.2) at 4°C. The cultures were subsequently washed twice in the same buffer and then post- fixed for I hour in 1% osmium tetroxide in the same buffer. After dehydration of the cultures with alcohol, the samples were embed-
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
3582 FALLA ET AL
,
ded in Epon. Sections were prepared on an LKB ultramicrotome, stained with uranyl acetate and lead citrate, and analyzed on a JEOL I200 EX microscope.
RNA preparafion and Northern blot. Total cellular RNA was prepared by acid guanidine thiocyanate/phenol/chloroform extrac- tion as described else~here.~’The RNA was fractionated by electro- phoresis in 1.5% agarose containing I O mmol/L sodium phosphate after denaturation with glyoxal and dimethyl sulfoxide. RNA was transferred to Biodyne filters (Pall) and hybridized to ”P-labeled ALP, collagen type I, and osteocalcin cDNA probes, kindly pro- vided by Dr Beatrice Mintz,” Dr Mon-li Chu;’ and the Genetics Institute;6 respectively. Hybridization and washings were per- formed according to the manufacturer’s guidelines. In sifu hybridizafion. Antisense oligodeoxynucleotides specific
for osteocalcin (5’-TCTGGGCTGGGGACTGAGGCT-T) were prepared on a DNA synthesizer (Biosearch, Cyclone DNA Synthe- sizers) by means of fi-cyanoethyl-phosphorymidite chemistry, as described elsewhere.37 The oligonucleotides were labeled with the DIG oligotide labeling kit (Boehringer Mannheim) according to the vendors instructions. In situ hybridization was performed on unde- calcified femur shafts embedded in glycol methacrylate. Sections were deproteinated using proteinase K at a concentration of I pg/ mL for 30 minutes at 37OC. Successively, the sections were incu- bated in 0.2% glycine and then prehybridized at 42°C for 2 hours in 50% deionized formamide supplemented with 5X SSC (1 X SSC = I50 mmol/L NaCl and I5 mmol/L sodium citrate), Denhardt’s solution, 1 mmol/L EDTA, and 500 pg/mL tRNA. Hybridization was performed overnight at 42°C in hybridization solution includ- ing all prehybridization reagents supplemented with approximately IO ng antisense or sense osteocalcin oligoprobes. The slides were then washed three times for 1 hour each in 50% formamide 2x SSC and 0. I X SSC, respectively. Hybridized osteocalcin oligoprobes were detected using goat-antidioxygenin monoclonal antibodies (Boehringer Mannheim).
Limifing dilufion analysis. Limiting dilution cultures were per- formed as described else~here.’~ Briefly, cell cultures were set up at different plating densities and stained with the Von Kossa tech- nique after 30 days. The percentage of wells showing no Von Kossa positive nodules was calculated for each cell plating density and was plotted against the number of BM cells plated per well. The number of cells required to form one bone nodule, which reflected the pro- portion of osteoprogenitor cells in the entire BM population, was then determined from the point at which the line crossed the 37% level, ie, F, = e-”, where F, is the fraction of wells without bone nodules and x is the mean number of osteoprogenitors per well. Based upon a Poisson di~tr ibut ion~~ of progenitor cells, F, = 37% corresponds to the dilution at which statistically there is one pro- genitor cell per well. In addition, the cultures were stained with May-Grlinwald-Giemsa and the frequency of fibroblast colony- forming cells was determined using identical statistical analysis.
RESULTS
BM cells cultured in medium containing /3-glycerophos- phate and ascorbic acid form distinct three-dimensional
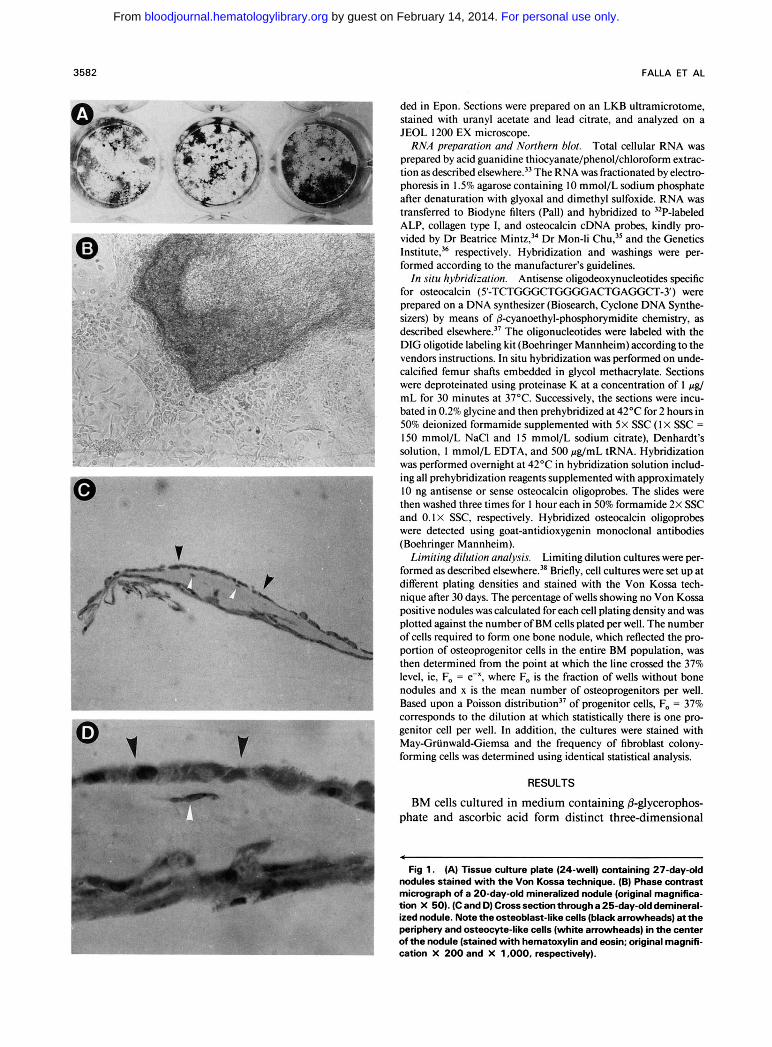
Fig 1 . (A) Tissue culture plate (24-well) containing 27-day-old nodules stained with the Von Kossa technique. (B) Phase Contrast micrograph of a 20-day-old mineralized nodule (original magnifica- tion X 50). (C and D) Cross section through a 25-day-old demineral- ized nodule. Note the osteoblast-like cells (black arrowheads) at the periphery and osteocyte-like cells (white arrowheads) in the center of the nodule (stained with hematoxylin and eosin; original magnifi- cation X 200 and X 1,000, respectively).
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
OSTEOPROGENITOR CELLS OF MOUSE BONE MARROW 3583
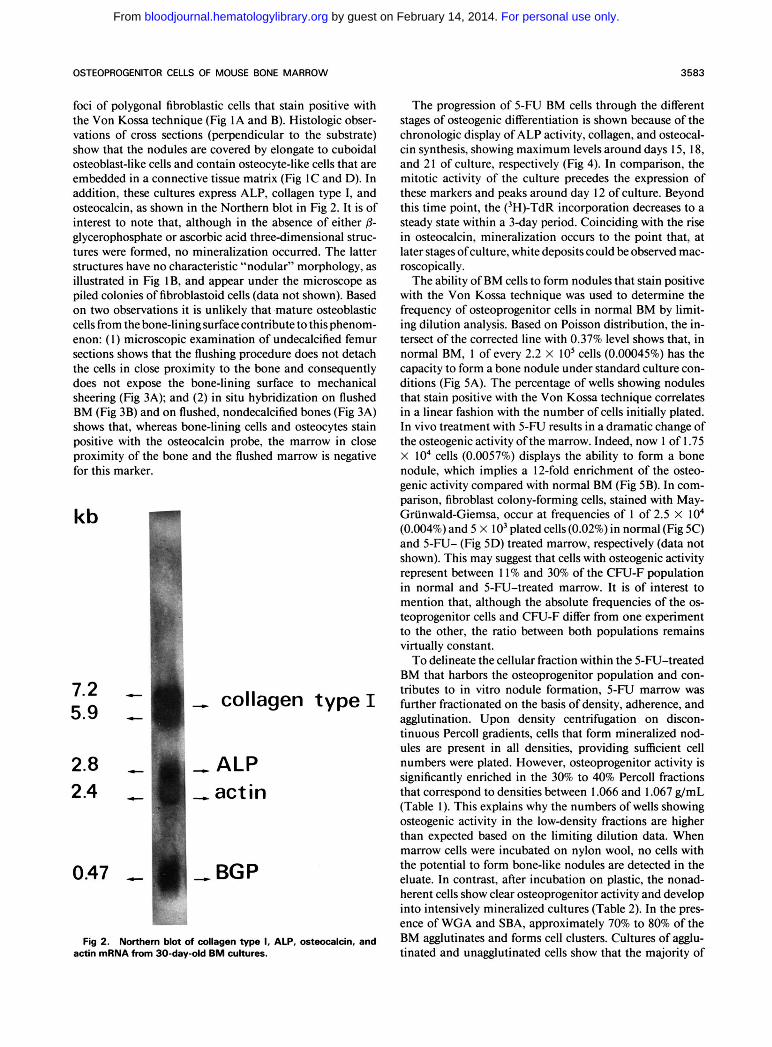
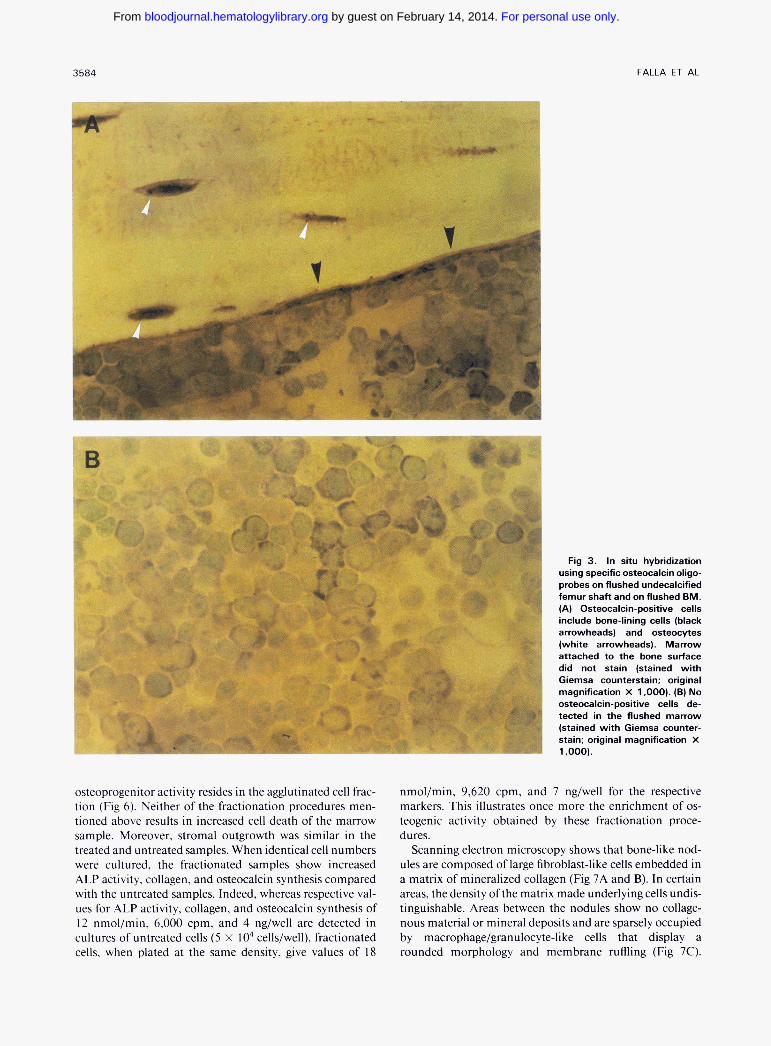
foci of polygonal fibroblastic cells that stain positive with the Von Kossa technique (Fig IA and B). Histologic obser- vations of cross sections (perpendicular to the substrate) show that the nodules are covered by elongate to cuboidal osteoblast-like cells and contain osteocyte-like cells that are embedded in a connective tissue matrix (Fig IC and D). In addition, these cultures express ALP, collagen type I, and osteocalcin, as shown in the Northern blot in Fig 2. It is of interest to note that, although in the absence of either 8- glycerophosphate or ascorbic acid three-dimensional struc- tures were formed, no mineralization occurred. The latter structures have no characteristic “nodular” morphology, as illustrated in Fig IB, and appear under the microscope as piled colonies of fibroblastoid cells (data not shown). Based on two observations it is unlikely that .mature osteoblastic cells from the bone-lining surface contribute to this phenom- enon: (1) microscopic examination of undecalcified femur sections shows that the flushing procedure does not detach the cells in close proximity to the bone and consequently does not expose the bone-lining surface to mechanical sheering (Fig 3A); and (2) in situ hybridization on flushed BM (Fig 3B) and on flushed, nondecalcified bones (Fig 3A) shows that, whereas bone-lining cells and osteocytes stain positive with the osteocalcin probe, the marrow in close proximity of the bone and the flushed marrow is negative for this marker.
kb
7.2 5.9
2.8 2.4
0.47
c - ALP c - act in
- - BGP !
Fig 2. Notthem blot of collagen type 1, ALP, osteocalcin, and actin mRNA from 30-day-old BM cultures.
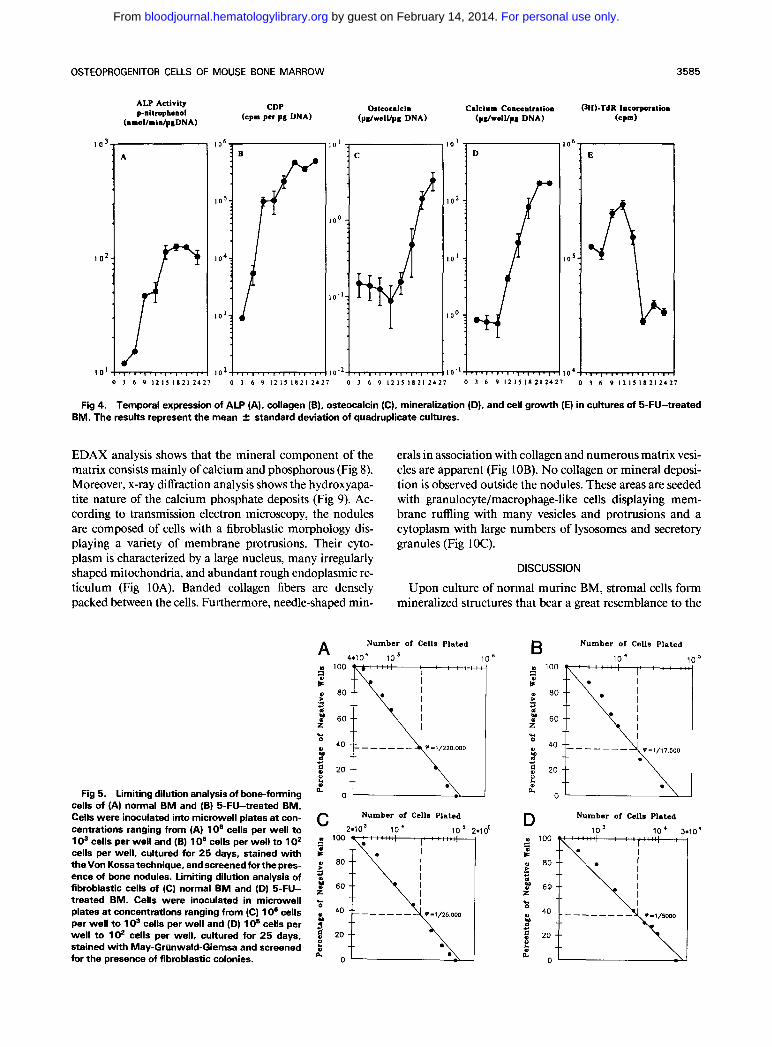
The progression of 5-FU BM cells through the different stages of osteogenic differentiation is shown because of the chronologic display of ALP activity, collagen, and osteocal- cin synthesis, showing maximum levels around days 15, 18, and 2 I of culture, respectively (Fig 4). In comparison, the mitotic activity of the culture precedes the expression of these markers and peaks around day 12 of culture. Beyond this time point, the (3H)-TdR incorporation decreases to a steady state within a 3-day period. Coinciding with the rise in osteocalcin, mineralization occurs to the point that, at later stages ofculture, white deposits could be observed mac- roscopically.
The ability of BM cells to form nodules that stain positive with the Von Kossa technique was used to determine the frequency of osteoprogenitor cells in normal BM by limit- ing dilution analysis. Based on Poisson distribution, the in- tersect of the corrected line with 0.37% level shows that, in normal BM, 1 of every 2.2 x IO5 cells (0.00045%) has the capacity to form a bone nodule under standard culture con- ditions (Fig 5A). The percentage of wells showing nodules that stain positive with the Von Kossa technique correlates in a linear fashion with the number of cells initially plated. In vivo treatment with 5-FU results in a dramatic change of the osteogenic activity ofthe marrow. Indeed, now 1 of 1.75 X IO4 cells (0.0057%) displays the ability to form a bone nodule, which implies a 12-fold enrichment of the osteo- genic activity compared with normal BM (Fig 5B). In com- parison, fibroblast colony-forming cells, stained with May- Griinwald-Giemsa, occur at frequencies of 1 of 2.5 X lo4 (0.004%) and 5 x IO3 plated cells (0.02%) in normal (Fig 5C) and 5-FU- (Fig 5D) treated marrow, respectively (data not shown). This may suggest that cells with osteogenic activity represent between 1 1% and 30% of the CFU-F population in normal and 5-FU-treated marrow. It is of interest to mention that, although the absolute frequencies of the os- teoprogenitor cells and CFU-F differ from one experiment to the other, the ratio between both populations remains virtually constant.
To delineate the cellular fraction within the 5-FU-treated BM that harbors the osteoprogenitor population and con- tributes to in vitro nodule formation, 5-FU marrow was further fractionated on the basis of density, adherence, and agglutination. Upon density centrifugation on discon- tinuous Percoll gradients, cells that form mineralized nod- ules are present in all densities, providing sufficient cell numbers were plated. However, osteoprogenitor activity is significantly enriched in the 30% to 40% Percoll fractions that correspond to densities between 1.066 and 1.067 g/mL (Table 1). This explains why the numbers of wells showing osteogenic activity in the low-density fractions are higher than expected based on the limiting dilution data. When marrow cells were incubated on nylon wool, no cells with the potential to form bone-like nodules are detected in the eluate. In contrast, after incubation on plastic, the nonad- herent cells show clear osteoprogenitor activity and develop into intensively mineralized cultures (Table 2). In the pres- ence of WGA and SBA, approximately 70% to 80% of the BM agglutinates and forms cell clusters. Cultures of agglu- tinated and unagglutinated cells show that the majority of
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
3584 FALLA ET AL
I
Fig 3. In situ hybridization using specific osteocalcin oligo- probes on flushed undecalcified femur shaft and on flushed BM. (A) Osteocalcin-positive cells include bone-lining cells (black arrowheads) and osteocytes (white arrowheads). Marrow attached to the bone surface did not stain (stained with Giemsa counterstain; original magnification X 1.000). (B) No osteocalcin-positive cells de- tected in the flushed marrow (stained with Giemsa counter- stain; original magnification X 1.000).
osteoprogenitor activity resides in the agglutinated cell frac- nmol/min, 9,620 cpm, and 7 ng/well for the respective tion (Fig 6). Neither of the fractionation procedures men- markers. This illustrates once more the enrichment of os- tioned above results in increased cell death of the marrow teogenic activity obtained by these fractionation proce- sample. Moreover. stromal outgrowth was similar in the dures. treated and untreated samples. When identical cell numbers Scanning electron microscopy shows that bone-like nod- were cultured, the fractionated samples show increased ules are composed of large fibroblast-like cells embedded in ALP activity, collagen, and osteocalcin synthesis compared a matrix of mineralized collagen (Fig 7A and B). In certain with the untreated samples. Indeed, whereas respective Val- areas, the density of the matrix made underlying cells undis- ues for ALP activity. collagen, and osteocalcin synthesis of tinguishable. Areas between the nodules show no collage- 12 nmol/min, 6,000 cpm, and 4 ng/well are detected in nous material or mineral deposits and are sparsely occupied cultures of untreated cells ( 5 X I O4 cells/well), fractionated by macrophage/granulocyte-like cells that display a cells, when plated at the same density. give values of 18 rounded morphology and membrane ruffling (Fig 7C).
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
OSTEOPROGENITOR CELLS OF MOUSE BONE MARROW 3585
ALP Activity p-nitrophenol
(naol lmidpg DNA)
Calcium Concentration (pdwelVpg DNA)
' O l i " 101 1 0 . 2 1
0 1 6 9 1 2 1 5 1 8 2 1 2 4 2 1 0 3 6 9 1 2 1 5 1 8 2 1 2 4 2 7 0 3 6 9 1 2 1 5 1 8 2 1 2 4
I
I O 5 - ?
l o 4 0 3 6 9 1 2 1 5 1 8 2 1 2 4 2 1 0 3 6 9 1 2 1 5 1 8 2 1 2 4
Fig 4. Temporal expression of ALP (A). collagen (B). osteocalcin (C), mineralization (D), and cell growth (E) in cultures of 5-FU-treated BM. The results represent the mean 2 standard deviation of quadruplicate cultures.
EDAX analysis shows that the mineral component of the matrix consists mainly of calcium and phosphorous (Fig 8). Moreover, x-ray diffraction analysis shows the hydroxyapa- tite nature of the calcium phosphate deposits (Fig 9). Ac- cording to transmission electron microscopy, the nodules are composed of cells with a fibroblastic morphology dis- playing a variety of membrane protrusions. Their cyto- plasm is characterized by a large nucleus, many irregularly shaped mitochondria, and abundant rough endoplasmic re- ticulum (Fig IOA). Banded collagen fibers are densely packed between the cells. Furthermore, needle-shaped min-
erals in association with collagen and numerous matrix vesi- cles are apparent (Fig IOB). No collagen or mineral deposi- tion is observed outside the nodules. These areas are seeded with granulocyte/macrophage-like cells displaying mem- brane ruffling with many vesicles and protrusions and a cytoplasm with large numbers of lysosomes and secretory granules (Fig IOC).
DISCUSSION
Upon culture of normal murine BM, stromal cells form mineralized structures that bear a great resemblance to the
Fig 5. Limiting dilution analysis of bone-forming cells of (A) normal BM and (B) 5-FU-treated BM. Cells were inoculated into microwell plates at con- centrations ranging from (A) 10' cells per well to 1 O3 cells per well and (B) 10' cells per well to 1 O2 cells per well, cultured for 25 days, stained with the Von Kossa technique, and screened for the pres- ence of bone nodules. Limiting dilution analysis of fibroblastic cells of (C) normal BM and (D) 5-FU- treated BM. Cells were inoculated in microwell plates at concentrations ranging from (C) 10' cells per well to 1 O3 cells per well and (D) 1 O5 cells per well to 10' cells per well, cultured for 25 days, stained with May-GrUnwald-Giemsa and screened for the presence of fibroblastic colonies.
Number of Cella Plated
0
P
C
2
," 40
3 20
.I 100
u 80 c
60 z a.
Y
0
0 z
Number of Cells Plated
v -1/25.000 - _ _ _ _ _
2.1 o5 I B n 100
L ; EO
60
.I 4
c
40 M e B 20 B a
Number of Cells Plated
10'
Number of Cella Plated
10'
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
3586 FALLA ET AL
Table 1. Density Distribution of Marrow Osteoprogenitor Cells
Percentage of Von Kossa-Positive Cultures
Density in % Percoll No. of Cells
Plated per Well Control >70 60-70 55-60 50-55 45-50 40-45 30-40
z x 105 100 0 100 100 100 100 100 100 105 100 0 55 60 60 100 100 100
5 x 104 100 0 0 30 0 100 100 100 2.5 x 104 100 0 0 0 0 100 100 100 1.2 x 104 100 0 0 0 0 60 60 100
6 X lo3 - 0 0 0 0 0 30 60
Marrow cells of 5-FU-treated animals were layered on a discontinuous gradient of 30%, 40%. 45%. 50%. 55%. 60%, and 70% Percoll fractions. The cells were separated on density by centrifugation, collected from the interfaces, and washed. For example, 60-70 signifies that cells were collected from the interface between the 60% and 70% Percoll layer. After 25 days, the cultures were stained with the Von Kossa technique and the number of positive cultures was scored. The results represent the percentage of wells showing mineralized nodules. A representative experiment is shown
bone-like nodules described in human and rat.'7,39-41 This phenomenon does not require the addition of exogenous hormones to the system. This is of interest because gluco- corticoids, more precisely dexamethasone (Dex), have been shown to induce terminal differentiation of osteogenic cells in tissue culture. Mass population studies and limiting dilu- tion analysis showed that in vitro bone nodule formation is enhanced in the presence of glucocorticoids, more precisely Dex. However, according to the same studies, Dex appeared not to be a requirement for in vitro os t e~genes i s .~~~ ' In an attempt to verify whether Dex has similar effects on murine BM cells, we found that concentrations up to lo-'' mol/L significantly reduce bone nodule formation and bone pro- tein (ALP, collagen, and osteocalcin) synthesis by the latter cells (data not shown). Possibly, the discrepancy in results can be explained by differential responsiveness of different species to glucocorticoids. Along this line, it was shown that glucocorticoids have growth stimulatory effects on rat osteo- blasts, whereas identical concentrations block mouse osteo- blast proliferation.& However, at this point, the role of naturally occumng glucocorticoids during osteogenic dif- ferentiation of mouse marrow cells cannot be ruled out be-
Table 2. Nylon Wool and Plastic Adherence of Marrow Osteoprogenitor Cells
Calcium Concentration (pg/well/rg DNA)
No. of Cells Nylon Wool Plastic Plated Der Well Control Nonadherent Nonadherent
5 x 104 40 k 3.2 ND 39 k 2.2 2.5 x io4 5 0 k 7.1 ND 48 k 5.7 1.2 x 104 36 k 3 ND 40 k 4.5
6 X lo3 2o.t 1.1 ND 21 k3.1 3 x 103 ND ND ND
The calcium determination assay does not detect values less than 0.01 5 mmol (0.6 pg/mL). Complete and nonadherent, 5-FU-treated marrow was cultured for 25 days, and calcium levels were determined and related to the DNA content per well. One representative experiment is shown. The results represent the mean & SD of quadruplicate CUI- tures.
Abbreviation: ND, not detectable.
cause hormone levels in the culture medium may be suffi- cient to drive nodule formation.
Because there is no single definitive marker for bone, the nodules were identified as being "bone-like'' based on their microscopic characterization and their expression of alka- line phosphatase:' collagen type 1,48,49 and osteocalcin, a noncollagenous protein that is currently believed to be ex- clusively synthetized by cells associated with b ~ n e . ~ ' , ~ ' It can be assumed that the osteogenic activity in these cultures results from mature osteoblastic cells detached from the bone-lining surface by sheering forces of the flushing proce- dure. However microscopic examination of flushed bones shows that a thick layer of marrow remains attached after
60
T
.... P c1 2 501 T
40
30
20
10
0 I Control WGA- WGA+ SBA- SBA+
Fig 6. Osteogenic activity of WGA' and SBA+ cells. 5-FU- treated BM was incubated with WGA or SBA and the agglutinated and unagglutinated fractions were plated at 5 X 1 O4 cells per well and cultured for 25 days. The cultures were washed and calcium levels were determined. The results represent the mean 2 the standard deviation of quadruplicate cultures.
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
OSTEOPROGENITOR CELLS OF MOUSE BONE MARROW 3587
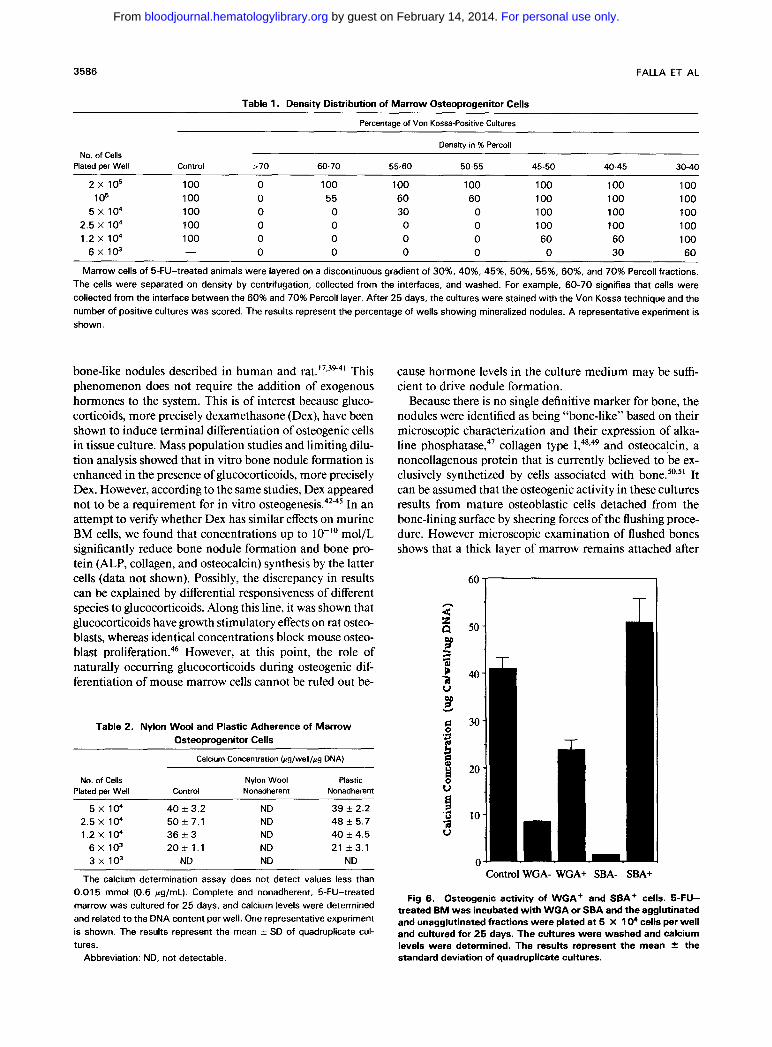
Fig 7. Scanning electron microscopy of 25-day-old cultures of 5- FU-treated BM. (A) Fibroblast-like cells (black arrows) showing cyto- plasmic blebs and filopods (black arrowheads) embedded in irregu- larly organized, mineralized collagen matrix (white arrows); (6) extracellular collagen matrix with mineralized particles (black arrow- heads); (C) cells of the nonmineralized spaces are devoid of collagen and mineral deposition.
flushing and covers the complete bone surface. Further- more, in situ hybridization shows that, whereas the osteo- cytes and the bone-lining cells stained bright with an osteo- calcin probe, the marrow in the vicinity of the bone and the flushed marrow were negative for this marker. Although the presence of a minor number of osteoblastic cells in the
flushed marrow cannot be excluded, the temporal expres- sion of ALP, collagen, and osteocalcin and the negligible levels of these markers at early explanation times makes it rather unlikely that contaminating mature osteoblast would be solely responsible for the in vitro bone formation in this model.
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
3588
511 FS m 251 5ml
b
0 . 0 2 . 0 I
0 6 . 0 8 . 0 10 .0
Lev
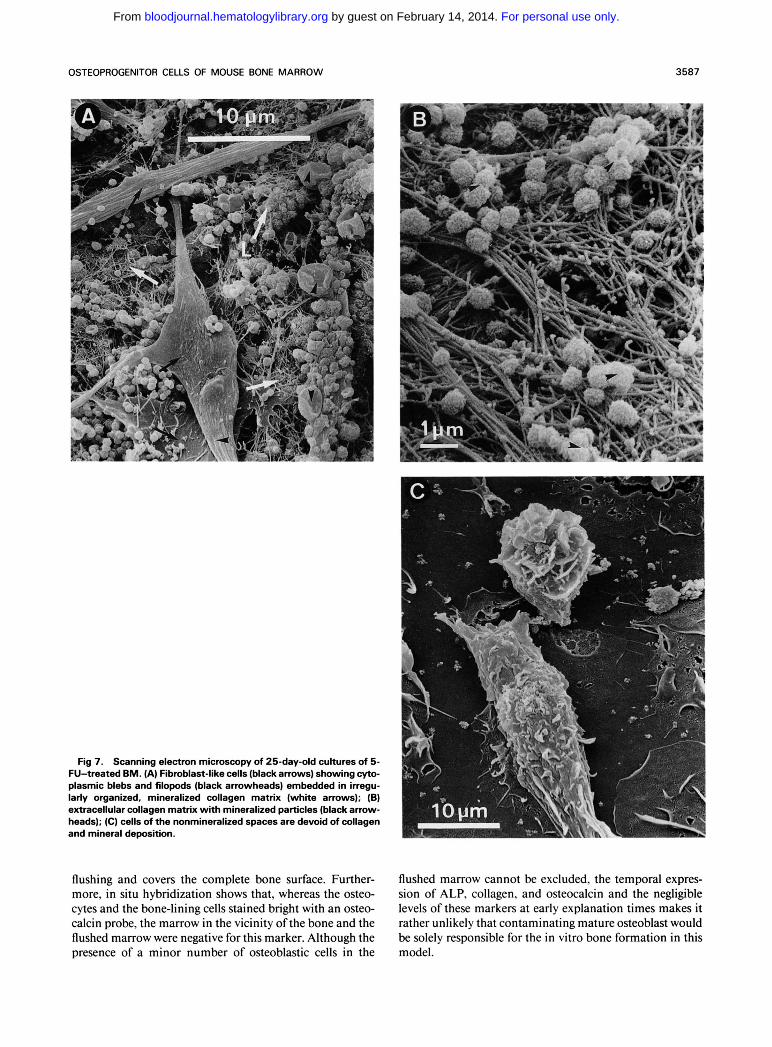
Fig 8. Energy dispersive x-ray spectrometry (EDAX) analysis of mineral-like material, similar to that shown in Fig 7. Demonstration of major peaks for calcium (Ca) and phosphorus (P).
The kinetic study of bone-associated markers in cultures of 5-FU-treated BM shows the temporal expression of ALP, collagen, and osteocalcin after a decrease in mitotic activity and preceding final mineralization. This resembles the situa- tion in cultures of rat calvaria osteoblasts, which also showed a temporal sequence of expression of markers en- coding the osteoblast phenotype characterized by three dis- tinct periods: proliferation, extracellular matrix matura- tion, and mineralization.'* BM cultures contain a variety of cell types. Therefore, part of the thymidine incorporation, ALP activity, and collagen synthesis in these cultures has to be ascribed to cells that do not belong to the osteogenic population. Although we cannot exclude the latter fact, we like to emphasize that the increase in thymidine incorpora- tion, ALP activity, and collagen synthesis coincides espe- cially with the formation and outgrowth of bone nodules, as observed in the microscope. No correlation with the occur- rence and increase in size of other fibroblastoid colonies was observed (data not shown).
To quantify the osteoprogenitor cells of the BM, the abil- ity of this tissue to form bone-like nodules in vitro was used to determine the clonal frequency of these cells by limiting dilution analysis. Approximately 0.00045% of the normal BM cells have the potency to form a bone nodule. Upon 5-FU treatment, this frequency is 12-fold increased, eg, 0.0057% cells displayed osteogenic activity. This drug was documented to have cytotoxic effects on cells in active mi-
FALLA ET AL
t o~ i s* ' -~~ and was previously used for the enrichment and purification of hematopoietic stem cells from the BM.S3 The 5-FU resistance of marrow osteoprogenitor cells suggests their nonproliferative nature in vivo. This correlates with the data on the quiescent status of the fibroblast compart- ment of the marrow stroma,'"56 which is believed to harbor these cell^."^"^^" In support, our data show that 5-FU treat- ment results in an enrichment of the fibroblast colony- forming cells as well. In normal marrow, they represent 0.004% of the cells, a frequency that is fivefold increased in 5-FU-treated marrow, eg, 0.02%. A linear relationship be- tween the number of cells plated and the fraction of the wells without nodules was obtained, which suggests the sin- gle-hit kinetic of the system. Therefore, cooperation be- tween different categories ofcells in the isolate was probably not required. Although the data strongly suggest that the bone nodules arise from an individual osteoprogenitor cell without participation from other cell types within the initial cell isolate, it cannot be excluded that homotypic cell-cell interactions between progeny of such an individual osteo- progenitor cell may be necessary for expression ofbone nod- u le~ .~" Based on the in situ hybridization data, it is evident that the frequency of osteogenic cells shown in this report may be underestimated, because it is unlikely that all pro- genitor cells were flushed from the marrow. Although the absolute frequencies of the CFU-F and osteoprogenitor cells in normal and 5-FU BM differed from one experiment to the other, the ratio between both populations remained vir- tually constant. This suggests that I I % to 30% ofthe CFU-F have osteogenic potential. This ratio is similar with in vivo transplantation data that indicated that 15% of in vitro de- rived fibroblastic clones yielded bone tissue when trans- planted under the renal cap~ule.'~
To better characterize the stroma subpopulation of the 5-FU BM that comprises the osteogenic lineage, marrow cells were fractionated on the basis oftheir biophysical char- acteristics. Density centrifugation on discontinuous percoll gradients showed that osteogenic activity was present over a wide range of densities, but was enriched in the lower densi-
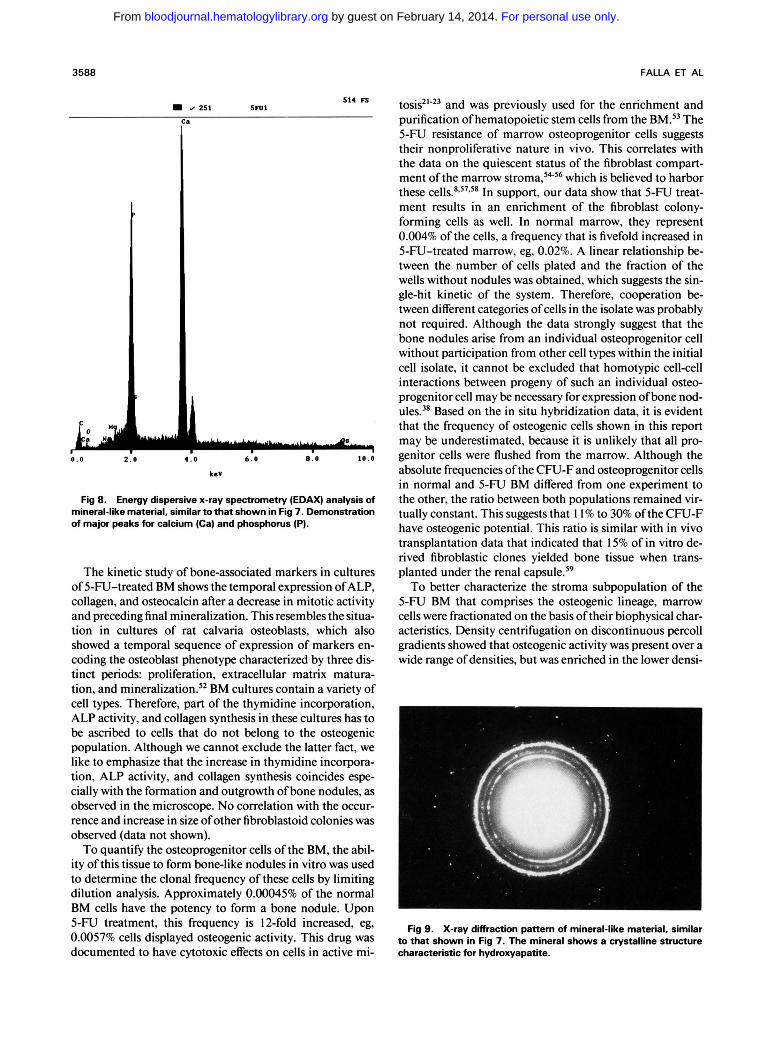
Fig 9. X-ray diffraction pattern of mineral-like material, similar to that shown in Fig 7. The mineral shows a crystalline structure characteristic for hydroxyapatite.
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
OSTEOPROGENITOR CELLS OF MOUSE BONE MARROW 3589
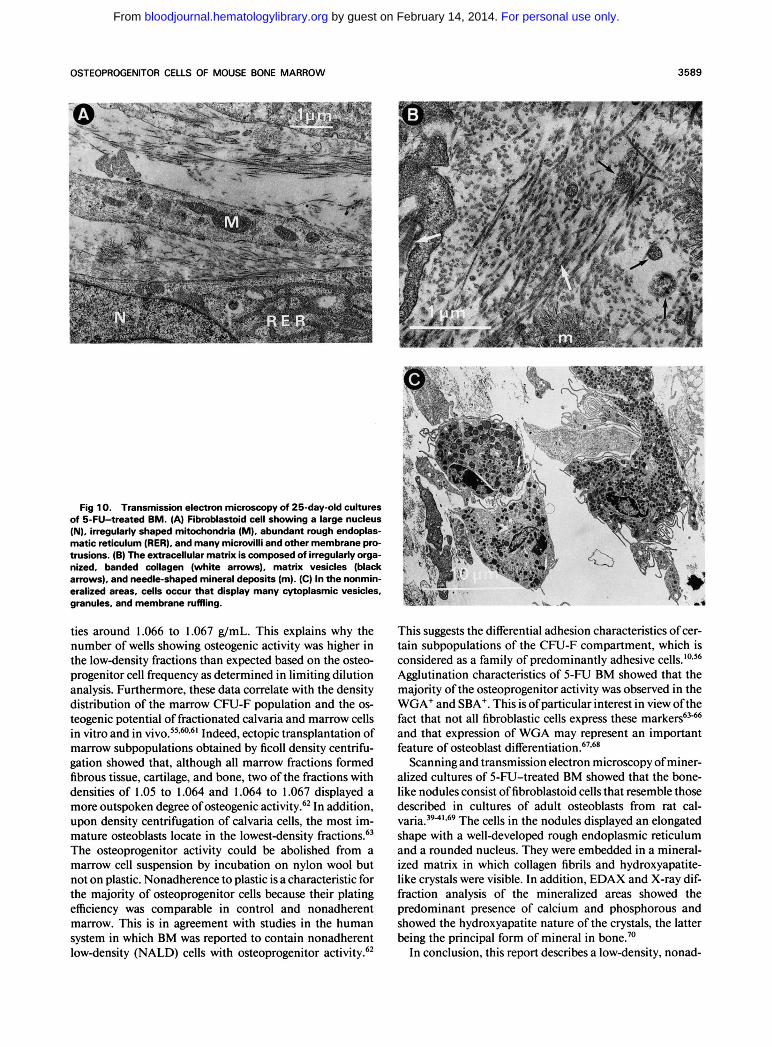
Fig 10. Transmission electron microscopy of 25-day-old cultures of 5-FU-treated BM. (A) Fibroblastoid cell showing a large nucleus (N), irregularly shaped mitochondria (M), abundant rough endoplas- matic reticulum (RER), and many microvilli and other membrane pro- trusions. (6) The extracellular matrix is composed of irregularly orga- nized, banded collagen (white arrows), matrix vesicles (black arrows), and needle-shaped mineral deposits (m). (C) In the nonmin- eralized areas, cells occur that display many cytoplasmic vesicles, granules, and membrane ruffling.
ties around 1.066 to 1.067 g/mL. This explains why the number of wells showing osteogenic activity was higher in the lowdensity fractions than expected based on the osteo- progenitor cell frequency as determined in limiting dilution analysis. Furthermore, these data correlate with the density distribution of the marrow CFU-F population and the os- teogenic potential of fractionated calvaria and marrow cells in vitro and in V ~ V O . ~ ~ * ~ - ~ ' Indeed, ectopic transplantation of marrow subpopulations obtained by ficoll density centrifu- gation showed that, although all marrow fractions formed fibrous tissue, cartilage, and bone, two of the fractions with densities of 1.05 to 1.064 and 1.064 to 1.067 displayed a more outspoken degree of osteogenic activity!' In addition, upon density centrifugation of calvaria cells, the most im- mature osteoblasts locate in the lowestdensity fra~tions.6~ The osteoprogenitor activity could be abolished from a marrow cell suspension by incubation on nylon wool but not on plastic. Nonadherence to plastic is a characteristic for the majority of osteoprogenitor cells because their plating efficiency was comparable in control and nonadherent marrow. This is in agreement with studies in the human system in which BM was reported to contain nonadherent lowdensity (NALD) cells with osteoprogenitor activity!*
This suggests the differential adhesion characteristics of cer- tain subpopulations of the CFU-F compartment, which is considered as a family of predominantly adhesive Agglutination characteristics of 5-FU BM showed that the majority of the osteoprogenitor activity was observed in the WGA+ and SBA+. This is of particular interest in view ofthe fact that not all fibroblastic cells express these marked- and that expression of WGA may represent an important feature of osteoblast differentiati~n!'*~*
Scanning and transmission electron microscopy of miner- alized cultures of 5-FU-treated BM showed that the bone- like nodules consist of fibroblastoid cells that resemble those described in cultures of adult osteoblasts from rat cal- varia.394'b9 The cells in the nodules displayed an elongated shape with a welldeveloped rough endoplasmic reticulum and a rounded nucleus. They were embedded in a mineral- ized matrix in which collagen fibrils and hydroxyapatite- like crystals were visible. In addition, EDAX and X-ray dif- fraction analysis of the mineralized areas showed the predominant presence of calcium and phosphorous and showed the hydroxyapatite nature of the crystals, the latter being the principal form of mineral in bone."
In conclusion, this report describes a lowdensity, nonad-
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom

3590 FALLA ET AL
herent osteogenic cell population from the BM that is 5-FU resistant and binds to WGA and SBA. It is evident that these characteristics will be of use in the further purification and identification of osteoprogenitor cells of the marrow.
ACKNOWLEDGMENT
The authors thank Dr R. Kemps for x-ray diffraction and for energy dispersive x-ray spectrometry (EDAX) analysis, Dr A. Van de Voorde for his critical review of the manuscript, and F. Vander Plaetse and L. Dierckx for technical and administrative support.
REFERENCES 1. Dexter TM: Stromal cell associated hematopoiesis. J Cell
Physiol87, 1982 (suppl I ) 2. Allen TD, Dexter TM: The essential cells of the haemopoietic
environment. Exp Haematol 12:5 17, I984 3. Strobe1 EA, Gay RE, Greenberg P L Characterization ofthe in
vitro stromal microenvironment of human bone marrow. Int J Cell Cloning 4:341, 1986
4. Perkins S, Fleischman RA: Stromal cell progeny of murine bone marrow fibroblast colony-forming units are clonal endothelial cells that express collagen type IV and laminin. Blood 75:620, 1990
5. Tsuji T, Hughes FJ, McCulloch CG, Melchers AH: Effects of donor age on osteogenic cells of rat bone marrow in vitro. Mech Ageing Dev 51:121, 1990
6. Dexter TM, Allen TD, Lajtha LG: Conditions controlling the proliferation for haemopoietic stem cells in vitro. J Cell Physiol 91:355, 1977
7. Sorrel JN, Weiss L Cell interactions between hemopoietic and stromal cells in the embryonic chick bone marrow. Anat Rec 197:1, 1980
8. Owen ME: Lineage of osteogenic cells and their relationship to the stromal system. Bone Mineral Res 3.1, 1980
9. Friedenstein AJ, Chailaklyan RK, Latsinik NV, Pansyuk AF, Keiliss-Borok 1 V Stromal cells responsible for transferring the mi- croenvironment of hemopoietic tissue. Transplantation 17:33 l , 1974
10. Friedenstein AJ: Precursor cells of mechanocytes. Int Rev Cytol47:327, 1976
1 I . Ashton BA, Allen TD, Howlett CR, Eaglesom CC, Hattori A, Owen ME: Formation of bone and cartilage by marrow stromal cells in diffusion chambers in vivo. Clin Orthop 15 1 :294, 1980
12. Grigordiadis AE, Heersche JNM, Aubin JE: Differentiation of muscle, fat, cartilage and bone from progenitor cells present in a bone derived clonal cell population: Effects of dexamethasone. J Cell Biol 106:2139, 1988
13. Ruther U, Garber C, Komitowski D, Muller R, Wagner EF: Deregulated c-fos expression interferes with normal bone develop ment in transgenic mice. Nature 325:415, 1987
14. Maxine G: Cytokines and Bone Metabolism. Boca Raton, FL, CRC, 1992
15. McCulloch CAG, Struguresco M, Hughes F, Melcher AH, Aubin JE: Osteogenic progenitor cells in rat bone marrow stromal populations exhibit self-renewal in culture. Blood 77: 1906, 1991
16. Howlett CR, Cave J, Williamson M, Farmer J, Ali SY, Bab I, Owen ME: Mineralization in in vitro cultures of rabbit marrow stromal cells. Clin Orthop 2 13:25 1, 1986
17. Haynesworth SE, Goshima J, Goldberg VM, Caplan AI: Characterization of cells in the osteogenic potential from human bone marrow. Bone 13:8 1, 1992
18. Maniatopoulos C, Sodek J, Melcher AH: Bone formation in vitro by stromal cells obtained from bone marrow of young adult rats. Cell Tissue Res 254:317, 1988
19. Schoeters GER, de Saint Georges L, Van Den Heuvel R, Leppens H, Vanderborght 0 Mineralization of adult mouse bone marrow cells in vitro. Cell Tissue Kinet 21:1, 1988
20. Kamalia N, McCulloch CAG, Tenebaum HC, Limeback H: Dexamethasone recruitment of self renewing osteoprogenitor cells in chick bone marrow stromal cell cultures. Blood 79:320, 1992
2 1. Yeager AM, Levin J, Levin M: The effects of 5-fluorouracil on hematopoeisis: Studies of murine megakaryocyte-cfc, granulo- cyte-macrophage, and peripheral blood levels. Exp Hematol 11:944, 1983
22. Bruce WR, Meeker BE Comparison of the sensitivity of he- matopoietic colony forming cells in different proliferative states to 5-fluorouracil. J Natl Cancer Inst 38:401, 1967
23. Roosendaal M, Hodgson GS, Bradley TR: Organization of haemopoietic stem cells: The generation-age hypothesis. Cell Tis- sue %net 12:17, 1979
24. Van Den Heuvel RL, Versele SRN, Schoeters GER, Vander- borght OLJ: Stromal stem cells (CFU-F) in yolk sac, liver, spleen and bone marrow of pre- and postnatal mice. Br J Haematol66: 15, 1987
25. Long MW, Hutchinson RJ, Gragowski LL, Heffner CH, Emerson SG: Synergistic regulation ofhuman megakaryocyte devel- opment. J Clin Invest 82:1779, 1990
26. Julius MH, Simpson E, Hertzenberg LA: A rapid method for the isolation of functional thymus derived murine lymphocytes. Eur J Immunol3:645, 1973
27. Reisner Y, Kapoor N, Hodes MZ, OReilly RJ, Good RA: Enrichment of CFU-C from murine and human bone marrow us- ing soybean agglutinin. Blood 59:360, 1982
28. Visser JWM, Bauman JGJ, Mulder AH, Eliason JF, De Leeuw AM: Isolation of murine pluripotent hematopoietic stem cells. J Exp Med 50: 1576, 1984
29. Lowry OH, Roberts NR, Wu M, Hixen WS, Crawford D: The quantitative histochemistry of brain 11. Enzyme measure- ments. J Bioi Chem 207: 13, 1954
30. Von Kossa J: Uber die im organismus kunztlich erzeugen Verkalkungen. Beitr Anat 29:163, 1901
3 1. Gitelman HJ: An improved automated procedure for the determination of calcium in biological specimens. Anal Biochem 18:521, 1967
32. Peterkovsky B, Diegelmann R: Use of a mixture of protein- ase-free collagenase for the specific assay of radioactive collagen in the presence of the proteins. Biochemistry 10:988, 1985
33. Chomczynski P, Sacchi N: Single-step method of RNA isola- tion by acid guanidinium thiocyanate-phenol-chloroform extrac- tion. Anal Biochem 162:156, 1987
34. Terao M, Mintz B: Cloning and characterization of a cDNA coding for mouse placental alkaline phosphatase. Proc Natl Acad Sci USA 84:7051, 1987
35. Chu M-L, Meyers JC, Bernard MP, Ding J-F, Ramirez F: Cloning and characterization of five overlapping cDNAs specific for the human pro a collagen chain. Nucleic Acids Res 10:5924, 1982
36. Celeste AJ, Rosen V, Buecker JL, Kriz R, Wang EA, Woz- ney JM: Isolation of the human gene for bone @a protein using mouse and rat cDNA clones. EMBO J 5:1885, 1986
37. De Capri0 JA, Ludlow JW, Lynch D, Furukawa T, Griffin J, Piwnicka-Worms U, Huang CM, Livingston DM: The product of the retinoblastoma susceptibility gene has properties of a cell cycle regulatory element. Cell 58: 1085, 1989
38. Bellows CG, Aubin J E Determination of numbers of osteo- progenitors present in isolated rat calvaria cells in vitro. Dev Biol 133:8, 1989
39. Nefussi JR, Boy-Lefevre ML, Boulekbache H, Forest N: Min-
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom

OSTEOPROGENITOR CELLS OF MOUSE BONE MARROW 3591
eralization in vitro of matrix formed by osteoblasts isolated by colla- genase digestion. Differentiation 29: 160, 1985
40. Bellows CG, Aubin JE, Heersche JNM, Antosz ME: Mineral- ized bone nodules formed in vitro from enzymatically released rat calvaria cell populations. Calcif Tissue Int 38: 143, 1986
4 1. Bhargava U, Bar-Lev M, Bellows CG, Aubin JE: Ultrastruc- tural analysis of bone nodules formed in vitro by isolated fetal rat calvaria cells. Bone 9: 155, 1988
42. McCulloch CAG, Tenenbaum H C Dexamethasone induces proliferation and terminal differentiation of osteogenic cells in tis- sue culture. Anat Rec 215397, 1986
43. Bellows CG, Aubin JE, Heersche JNM: Physiological con- centrations of glucocorticoids stimulate formation of bone nodules from isolated rat calvaria cells in vitro. Endocrinology 12 1 : 1985, 1987
44. Bellows CG, Heersche JNM, Aubin JE: Determination of the capacity for proliferation and differentiation of osteoprogenitor cells in the presence and absence of dexamethasone. Dev Biol 140: 132, I990
45. Kamalia N, McCulloch CAG, Tenebaum HC, Limeback H: Dexamethasone recruitment of self-renewing osteoprogenitor cells in chick bone marrow stromal cell cultures. Blood 79:230, 1992
46. Chen TL, Cone CM, Feldman D Glucocorticoid modula- tion of cell proliferation in cultured osteoblast-like cells: Differ- ences between rat and mouse. Endocrinology 112: 1739, 1983
47. Rodan GA, Rodan SB: Expression ofthe osteoblastic pheno- type, in Peck WA (ed): Bone and Mineral Research. Annual 2. Amsterdam, The Netherlands, Elsevier, 1983, p 244
48. Miller EJ: A review ofbiochemical studies on the genetically distinct collagens of the skeletal system. Clin Orthop 92:260, 1973
49. Bornstein P, Sage H: Structurally distinct collagen types. Annu Rev Biochem 59:957, 1980
50. Lian JB, Friedman PA: The vitamin K dependent synthesis of gamma-carboxiglutamic acid by bone microsomes. J Biol Chem 253:6623, 1978
5 1. Nishimoto SK, Price PA: Proof that the gamma-carboxiglu- tamic acid containing protein is synthesized in calf bone: Compara- tive synthesis rate and effect of coumadin on synthesis. J Biol Chem 254:437, 1979
52. Owen TA, Aronow M, Shalhoub V, Barone LM, Wilming L, Tassinari MS, Kennedy MB, Pockwinse S, Lian JB, Stein GS Pro- gressive development ofthe rat osteoblast phenotype in vitro: Recip- rocal relationships in expression of genesassociated with osteoblast proliferation and differentiation during formation of the bone ex- tracellular matrix. J Cell Physiol 143:420, 1990
53. Hodgson GS, Bradley TR: Properties of hematopoietic stem cells surviving 5-fluorouracil treatment evidence for a pre-CFU-s cell? Nature 281:381, 1979
54. Mathieu E, Schoeters G, Vander Plaetse F, Merregaert J: Establishment of an osteogenic cell line derived from adult mouse bone marrow stroma by use of a recombinant retrovirus. Calcif Tissue Int 50362, 1992
55. Friendenstein YA, Latsinik V, Grosheva AG, Yu F, Gorskaya L Marrow microenvironment transfer by heterotopic transplantation of freshly isolated and cultured cells in porous sponges. Exp Hematol 10:217, 1982
56. Castro-Malaspina H, Gay RE, Resnick G, Kapoor N, Meyers P, Chiarieri D, McKenzie S, Broxmeyer HE, Moore MAS: Characterization of human bone marrow fibroblast colony-form- ing cells (CFU-f) and their progeny. Blood 56:289, 1980
57. Hass RJ, Bohne E, Fliedner TM: On the development of slowly turning over cell types in neonatal rat bone. Blood 34:79 1, 1969 58. Benayahn D, Kletter Y, Zipori D, Wientroub S Bone
marrow derived stromal cell line expressing osteoblastic phenotype in vitro and osteogenic capacity in vivo. J Cell Physiol 140: 1, 1989
59. Friedenstein AJ: Stromal mechanisms of bone marrow: Cloning in vitro and retransplantation in vivo, in Thienfelder S (ed): Immunobiology of Bone Marrow Transplantation. Berlin, Ger- many, Springer-Verlag, 1980, p 19
60. Piersma AH, Brockbank KGM, Ploemacher RE, van Vliet E, Brakel-van Peer KMJ, Visser PJ: Characterization of fibroblastic stromal cells from murine bone marrow. Exp Hematol 13:237, 1985
6 1. Budenz RA, Bernard GW Osteogenesis and leukopoiesis within diffusion chamber implants of isolated bone marrow sub- populations. Am J Anat 159:455, 1980
62. Wong GL, Ng MC, Calabrese DW, Swartzendruber DE: Characterization of bone cells isolated on discontinuous percoll gradients: Distribution in sequentially derived populations. J Bone Mineral Res 6:969, 199 1
63. Long MW, Williams JL, Mann KG: Expression of human bone-related proteins in the hematopoietic microenvironment. J Clin Invest 86:1387, 1990
64. Zipori S, Sasson T, Frenkel A: Myelopoiesis in the presence of stromal cells from mouse bone marrow. Exp Hematol 9:656, 1981
65. Gordon BB, Pena SDJ: The surface glycoproteins of human skin fibroblasts detected after electrophoresis by binding of peanut agglutinin and Ricinus communis agglutinin I. J Biochem 2089:351, 1982
66. Ebell W, Castro-Malaspina H, Moore MAS, O'Reilly RJ: Depletion of stromal elements in human marrow grafts separated by soybean agglutinin. Blood 65: 1105, 1985
67. Nakamura H, Ozawa H: Lectin cytochemical study on osteo- blasts and calcification nodules of bone matrix in rats. Bone Min- eral 17: 137, 1992 (abstr, suppl I )
68. Nakamura H, Ozawa H: Lectin cytochemistry on the stra- tum intermedium and the papillary layer in the rat incisor enamel. Arch Histol Cytol 53:351, 1990
69. Masquelier D, Herbert B, Hauser N, Mermillod P, Schonne E, Remacle C: Morphologic characterization of osteoblast-like cell cultures isolated from newborn rat calvariae. Calcif Tissue Int 47:92, 1990
70. Posner AS: The mineral of bone. Clin Orthop 200:87, 1985
For personal use only. by guest on February 14, 2014. bloodjournal.hematologylibrary.orgFrom
Copyright © 2022 FDOKUMEN




















