
Induction of basic fibroblast growth factor (bFGF) expression following focal cerebral ischemia
11
Ž . Molecular Brain Research 49 1997 255–265 Research report ž / Induction of basic fibroblast growth factor bFGF expression following focal cerebral ischemia Teng N. Lin a, ) , Jeannie Te a , Melody Lee a , Grace Y. Sun b , Chung Y. Hsu c a DiÕision of Neuroscience, Institute of Biomedical Sciences, Academia Sinica, Taipei 11529, Taiwan, ROC b Department of Biochemistry, UniÕersity of Missouri, Columbia, MO, USA c Department of Neurology, Washington UniÕersity School of Medicine, St. Louis, MO, USA Accepted 8 April 1997 Abstract Ž . Basic fibroblast growth factor bFGF is a biologically active polypeptide with mitogenic, angiogenic, and neurotrophic properties. In the present study, we examined the temporal and spatial expression profiles of bFGF mRNA and protein concentration in a focal cerebral Ž . Ž . ischemia model induced by transient occlusion of the right middle cerebral artery MCA and both common carotid arteries CCAs . Results of Northern blot analysis shows a transient 2.5-fold increase in the 6.0 kb transcript of bFGF mRNA within the ischemic cortex of rats subjected to 60 min ischemic insult followed by 12 h of reperfusion. Although enhanced expression of bFGF mRNA was also noted in the ipsilateral hippocampus, the temporal induction profile appeared to be different from that of the ischemic cortex. A significant increase in bFGF mRNA was observed as early as 60 min following ischemia and remained elevated for up to 2 weeks after the onset of reperfusion. In situ hybridization studies revealed constitutive expression of bFGF mRNA in discrete brain regions of sham-operated animals. Following 60 min ischemia and 12 h reperfusion, increased expression of bFGF mRNA was observed in the ischemic cortex Ž . both peri-infarct and infarct area . Increased expression of bFGF mRNA within the infarcted area is largely confined rostrally to the outer cortical layers of the infarct, an area with increased density of blood vessels. bFGF-like immunoreactivity was also detected in areas expressing bFGF mRNA. Furthermore, a striking increase in bFGF-like immunoreactivity was observed in the ipsilateral hippocampus. Double-staining with anti-GFAP antibody indicated that the majority of the bFGF-like immunoreactivity was localized in the astrocytes, however, not all astrocytes showed bFGF-like immunoreactivity. Some GFAP negative cell also showed bFGF-like immunoreactivity. In summary, increased expression of both bFGF mRNA and immunoreactivity following ischemia were located in the same brain regions. An increase in bFGF-like immunoreactivity after ischemic insult is likely due to an increase in the expression of its 6.0 kb bFGF mRNA transcripts. Although increased bFGF mRNA was observed in both ischemic cortex and ipsilateral hippocampus after ischemic insult, the temporal expression profiles differed. Results from the present study raise the possibility that increased expression of bFGF in the peri-infarcted area may limit the spread of ischemic injury. q 1997 Elsevier Science B.V. Keywords: Focal ischemia; bFGF; Rat; Hybridization, in situ; Immunohistochemistry 1. Introduction Cerebral ischemia is known to cause brain injury lead- ing to neuronal cell death. In mammalian brain, functional recovery may take place after stroke, even though neural w x regeneration may not occur 31 . The exact mechanisms leading to the recovery are not well defined. One of the potential mechanisms related to this event is the induction w x and synthesis of growth factors 18,52 . ) Ž. Corresponding author. Fax: q886 2 785-3569; E-mail: [email protected] Ž . Fibroblast growth factors FGFs are biologically active mitogenic polypeptides which belong to the family of w x heparin binding growth factors 6,26 . Two forms of FGF, acidic and basic, are widely distributed throughout the Ž . immature and adult central nervous system CNS w x 24,46,56,57 . Both molecules share a sequence homology of about 55% and their activities seem to be mediated w x through the same receptor localized in the CNS 28,51,71 . Other members of the FGF family have been recently w x identified 4,27,63 . In the rat brain, multiple species of Ž . basic FGF bFGF mRNA transcripts have been identified w x in the hippocampus, hypothalamus and cortex 57 . The regional expression of these bFGF mRNA transcripts also 0169-328Xr97r$17.00 q 1997 Elsevier Science B.V. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Induction of basic fibroblast growth factor (bFGF) expression following focal cerebral ischemia

Ž .Molecular Brain Research 49 1997 255–265
Research report
ž /Induction of basic fibroblast growth factor bFGF expression followingfocal cerebral ischemia
Teng N. Lin a,), Jeannie Te a, Melody Lee a, Grace Y. Sun b, Chung Y. Hsu c
a DiÕision of Neuroscience, Institute of Biomedical Sciences, Academia Sinica, Taipei 11529, Taiwan, ROCb Department of Biochemistry, UniÕersity of Missouri, Columbia, MO, USA
c Department of Neurology, Washington UniÕersity School of Medicine, St. Louis, MO, USA
Accepted 8 April 1997
Abstract
Ž .Basic fibroblast growth factor bFGF is a biologically active polypeptide with mitogenic, angiogenic, and neurotrophic properties. Inthe present study, we examined the temporal and spatial expression profiles of bFGF mRNA and protein concentration in a focal cerebral
Ž . Ž .ischemia model induced by transient occlusion of the right middle cerebral artery MCA and both common carotid arteries CCAs .Results of Northern blot analysis shows a transient 2.5-fold increase in the 6.0 kb transcript of bFGF mRNA within the ischemic cortex ofrats subjected to 60 min ischemic insult followed by 12 h of reperfusion. Although enhanced expression of bFGF mRNA was also notedin the ipsilateral hippocampus, the temporal induction profile appeared to be different from that of the ischemic cortex. A significantincrease in bFGF mRNA was observed as early as 60 min following ischemia and remained elevated for up to 2 weeks after the onset ofreperfusion. In situ hybridization studies revealed constitutive expression of bFGF mRNA in discrete brain regions of sham-operatedanimals. Following 60 min ischemia and 12 h reperfusion, increased expression of bFGF mRNA was observed in the ischemic cortexŽ .both peri-infarct and infarct area . Increased expression of bFGF mRNA within the infarcted area is largely confined rostrally to the outercortical layers of the infarct, an area with increased density of blood vessels. bFGF-like immunoreactivity was also detected in areasexpressing bFGF mRNA. Furthermore, a striking increase in bFGF-like immunoreactivity was observed in the ipsilateral hippocampus.Double-staining with anti-GFAP antibody indicated that the majority of the bFGF-like immunoreactivity was localized in the astrocytes,however, not all astrocytes showed bFGF-like immunoreactivity. Some GFAP negative cell also showed bFGF-like immunoreactivity. Insummary, increased expression of both bFGF mRNA and immunoreactivity following ischemia were located in the same brain regions.An increase in bFGF-like immunoreactivity after ischemic insult is likely due to an increase in the expression of its 6.0 kb bFGF mRNAtranscripts. Although increased bFGF mRNA was observed in both ischemic cortex and ipsilateral hippocampus after ischemic insult, thetemporal expression profiles differed. Results from the present study raise the possibility that increased expression of bFGF in theperi-infarcted area may limit the spread of ischemic injury. q 1997 Elsevier Science B.V.
Keywords: Focal ischemia; bFGF; Rat; Hybridization, in situ; Immunohistochemistry
1. Introduction
Cerebral ischemia is known to cause brain injury lead-ing to neuronal cell death. In mammalian brain, functionalrecovery may take place after stroke, even though neural
w xregeneration may not occur 31 . The exact mechanismsleading to the recovery are not well defined. One of thepotential mechanisms related to this event is the induction
w xand synthesis of growth factors 18,52 .
) Ž .Corresponding author. Fax: q886 2 785-3569; E-mail:[email protected]
Ž .Fibroblast growth factors FGFs are biologically activemitogenic polypeptides which belong to the family of
w xheparin binding growth factors 6,26 . Two forms of FGF,acidic and basic, are widely distributed throughout the
Ž .immature and adult central nervous system CNSw x24,46,56,57 . Both molecules share a sequence homologyof about 55% and their activities seem to be mediated
w xthrough the same receptor localized in the CNS 28,51,71 .Other members of the FGF family have been recently
w xidentified 4,27,63 . In the rat brain, multiple species ofŽ .basic FGF bFGF mRNA transcripts have been identified
w xin the hippocampus, hypothalamus and cortex 57 . Theregional expression of these bFGF mRNA transcripts also
0169-328Xr97r$17.00 q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0169-328X 97 00152-6

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265256
shows temporal changes during brain developmentw x7,14,57,59,72 . For example, levels of the 1.8 kb bFGFmRNA transcripts are high in the embryonic stage, whereas
the 6.0 kb bFGF mRNA transcripts predominate in theadult brain.
Besides acting as mitogenic and angiogenic factors in
Ž .Fig. 1. Time-course of bFGF mRNA induction in the cerebral cortex following focal cerebral ischemia-reperfusion. Total RNA 15 mg extracted from theŽ . Ž . 32ischemic A and contralateral cortex C of rats subjected to 60 min ischemia and various reperfusion periods was hybridized with P-labelled bFGF
Ž .probe as described in the text. The same membrane was subsequently stripped and rehybridized with GAPDH B,D to serve as internal control. Thus,Ž . Ž . Ž . Ž .panels A and B represent bFGF and GAPDH mRNA signal, respectively, from the ischemic cortex and panels C and D were from the contralateral
Ž . Ž .or non-ischemic cortex. The time points for ischemic and contralateral non-ischemic cortices are displayed below panels B and D , respectively. Note theŽ . Ž . Ž .loading order of 12 and 24 h samples was out of sequence for panels A and B . Panel E shows fold-increase in bFGF mRNA signal in the ischemic
cortex in reference to sham-operated control over time. The radioactive bands of bFGF mRNA were quantified and normalized with those derived fromcorresponding GAPDH mRNA. Signal obtained from the sham-operated control was arbitrarily defined as 1. The fold-increase refers to the ratio of thenormalized ischemic cortex to the normalized sham-operated cortex. Data are mean"SD from 4 animals. w and ww indicate P-0.05 and P-0.01,respectively, as compared with the sham-operated control. Sh denotes sham-operated control; y0.5 represents 30 min into ischemic period or 30 minbefore reperfusion; 0, 0.5, 1.5, 4, 12, 24, 72, 168, 336 and 504 represent 60 min ischemia with 0, 0.5, 1.5, 4, 12, 24, 72, 168, 336 and 504 h of reperfusion,respectively.

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265 257
endothelial cells, FGFs have been shown to have neu-rotrophic properties for neurons in the CNSw x3,48,55,69,70,74 . Recent data indicate that levels of bFGF
w x w ximmunoreactivity 17,32,36,41 and mRNA 15,42,65 in-crease in response to cerebral ischemic insult. These re-sults raise the possibility that bFGF expression may be arestorative neural response to ischemic brain injury. Fur-thermore, bFGF has been shown to offer protection against
w xhypoxia-ischemic insult in vitro 45 and in vivow x19,30,35,37,44,53,54,66 . bFGF also enhances recovery of
w xbehavior in rats following traumatic brain injury 13,33 .The exact mechanisms leading to this beneficial effect arenot presently clear. Moreover, the sources of the bFGFbearing cells, the mRNA transcripts which are responsiblefor the increase in bFGF protein, and the mechanisms ofaction of bFGF in the injured brain have yet to be eluci-dated. In the present study, Northern blot analysis and insitu hybridization were used to examine the temporal andspatial expression profiles of bFGF mRNA in relation tothe expression of bFGF-like immunoreactivity in the ratusing a focal cerebral ischemia-reperfusion model.
2. Materials and methods
2.1. Stroke model
The focal cerebral ischemia-reperfusion model in the ratw xhas been described previously 9,40,76 . Briefly, male
Long–Evans rats weighing 250–300 g were anesthetizedŽ .with 100 mgrkg ketamine i.p. and 6 mgrkg xylazine
Ž . Ž .i.m. . The trunk of the right middle cerebral artery MCAabove the rhinal fissure was identified under a stereomicro-scope and ligated with a 10-0 suture. Complete attenuationof blood flow was confirmed under the microscope. Both
Ž .common carotid arteries CCAs were then occluded usingnontraumatic aneurysm clips. After a predetermined dura-tion of ischemia, the aneurysm clips and the suture wereremoved and restoration of blood flow in all three arterieswas verified. During the operation, rectal temperature wasmonitored and maintained at 37.0 " 0.58C using a
Ž .homeothermic blanket Harvard, USA . Under these condi-tions, ischemia for 60 min results in a consistently largearea of infarction which is confined to the right MCA
Ž .Fig. 2. Time course of bFGF mRNA induction in the hippocampus following focal cerebral ischemia-reperfusion. Total RNA 15 mg extracted from theŽ . 32ipsilateral hippocampus panel A of rats subjected to 60 min ischemia and various reperfusion periods was hybridized with P-labelled bFGF probes as
Ž .described in the text. Panel B displays corresponding GAPDH mRNA signals as internal standards. Signal obtained from the sham-operated control wasŽ .arbitrarily defined as 1. Panel C shows fold-increase in bFGF mRNA signal in the ipsilateral hippocampus in reference to sham-operated control over
time. Data are mean"SD from 3 animals. w and ww indicate P-0.05 and P-0.01, respectively, as compared with the sham-operated controls. Otherinformation is as detailed in Fig. 1.

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265258
cortex, whereas small or no infarction is observed follow-w xing mild ischemia induced for 15 min 43 . Following the
ischemic insult, rats were kept in an air-ventilated incuba-Žtor at 24.0"0.58C for various reperfusion periods y30,
.0, 30, 60, 90 min, 4, 12 h, 1, 3 days, and 1, 2, 3, 4 weeksand were provided with water and lab chow ad libitum.Sample size for each time point in the individual experi-mental group is further indicated in figure legends. At theend of the experiments, rats were sacrificed either bytranscardial perfusion with cold 4% paraformaldehyde andthe brain removed and cryo-protected in 30% sucrose at48C overnight, or by decapitation followed by dissection ofthe cortex and hippocampus which were quick frozen inliquid nitrogen. Samples were stored at y708C until fur-ther processing. Rats were housed in Animal Facilities of
Ž .Institute of Biomedical Sciences IBMS according to theguidelines established in the Guide for the Care and Useof Laboratory Animals prepared by the Committee on Careand Use of Laboratory Animals of Institute of LaboratoryAnimal Resources Commission on Life Sciences, National
Ž .Research Council, USA 1985 .
2.2. RNA isolation and northern blot analysis
Total RNA was isolated from frozen tissue using thesingle-step acid guanidinium thiocyanate-phenol-chloro-
w xform extraction method 11 . For Northern blot analysis,Ž .RNA samples 15 mgrlane were applied on 1.2% agarose
gel in the presence of 2.2 M formaldehyde. After elec-trophoresis, gel was transblotted onto Nytrane membranesŽ . w xGene Screen Plus, DuPont, Boston, Mass 67 . Mem-branes were prehybridized at 608C in a solution containing1% SDS, 1 M NaCl, 10% dextran sulfate and 100 mgrmlof sheared salmon sperm DNA. A rat bFGF cDNA was
Žused to probe bFGF mRNA a gift from Dr. Seth P.. w xFinklestein 38,57 . An 832 bp ECoRI fragment of the
plasmid l FR-1 containing the entire coding region ofbFGF with 50 bp of 3X untranslated region and 200 bp of5X untranslated region was isolated as the initial bFGFcDNA probe. Since this probe is homologous to the 28Sribosomal RNA, a 350 bp sub-fragment was obtained afterKpnI–ECoRI digestion. This fragment corresponded to thethird exon coding region and the 3X untranslated region ofthe rat bFGF cDNA. Radioactive probes were prepared by
Ža random-primer labeling method Amersham, Arlington. 32 Ž 6 .Heights, IL . P-labelled bFGF probe 1=10 cpmrml
was added directly to the prehybridization solution. Fol-lowing hybridization, which ranged from 24 to 48 h at608C, the membranes were washed twice in 2= SSC atroom temperature for 5 min each, followed by two 30-minwashes at 608C in 2= SSCr1% SDS and two 30-min
w xwashes at 608C in 0.1= SSC 1 . Membranes were thenŽexposed to Hyperfilm-MP Amersham, Arlington Heights,
.IL . The radioactive bands in the film were quantified by aŽ .300S Computing Densitometer Molecular Dynamics, CA .
2.3. In situ hybridization
Rats were perfused with cold 4% paraformaldehyde,sacrificed, and the extracted brain tissue placed in cryo-protective 30% sucrose overnight. Frozen brains weresectioned at 25 mm and mounted onto poly-L-lysine coatedslides. Brain sections were subjected to 0.001% proteinaseK digestion at 378C for 30 min, then immersed in 0.1 M
Ž .triethanolamine TEA with 0.25% acetic acid anhydride atroom temperature for 10 min and subsequently dehydrated
Ž .in 50, 70, 95 and 100% ethanol 3 min each . Hybridiza-tion was carried out in a solution containing 12.5 Mformamide, 10% dextran sulfate, 0.3 M NaCl, 1= Den-hardt’s solution, 10 mM Tris-Cl 8.0, 500 mgrml ssDNA,100 mgrml tRNA, 20 mM DTT and 106 cpmrml of
Fig. 3. In situ hybridization of bFGF mRNA in the rat brain. The regionaldistribution of bFGF mRNA is shown in a rat subjected to 60 minischemia followed by 12 h reperfusion and in a sham-operated control.
Ž . 35Brain sections 25 mm were hybridized with S-labelled bFGF probe asdescribed in the text. bFGF message was increased in the ischemic right
Ž .MCA cortex and adjacent regions A ;C . Note the peri-infarcted en-Ž .hancement of bFGF message A ;C . Increase in bFGF signal was also
Ž .noted in the outer layers of the infarcted cortex A,B . High basal levelsŽ .of bFGF mRNA was observed in the CA2 and fasciola cinereum D in a
sham-operated control. Similar results were duplicated in two other setsof ischemic and sham-operated rats.
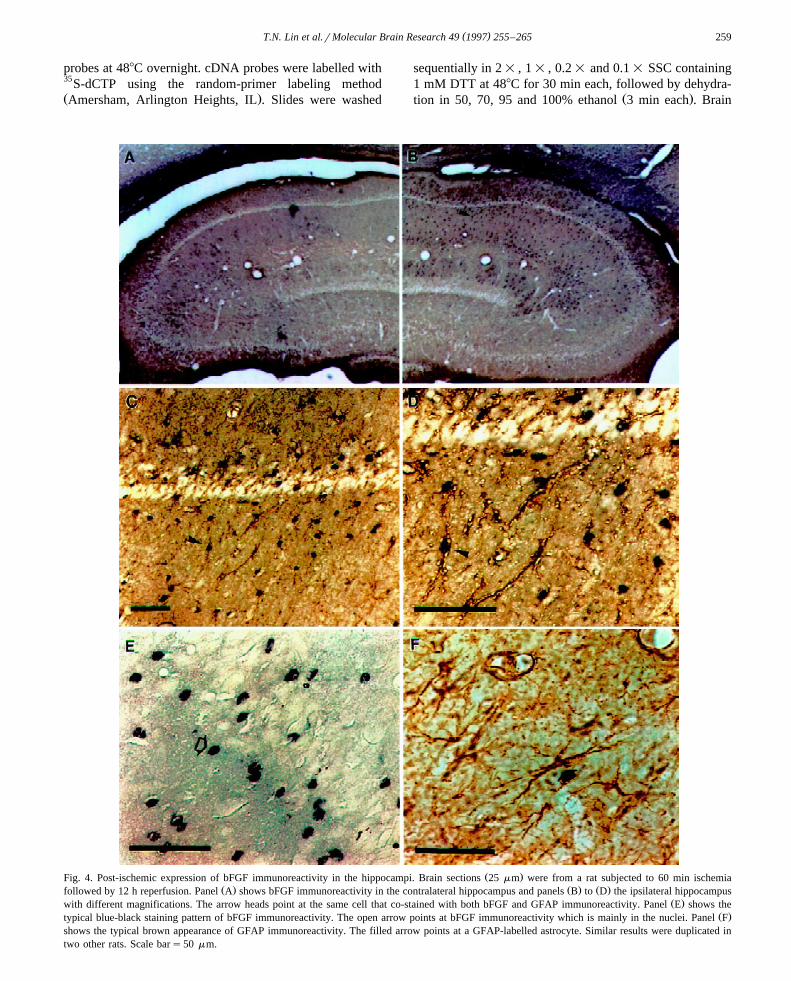
( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265 259
probes at 488C overnight. cDNA probes were labelled with35S-dCTP using the random-primer labeling methodŽ .Amersham, Arlington Heights, IL . Slides were washed
sequentially in 2= , 1= , 0.2= and 0.1= SSC containing1 mM DTT at 488C for 30 min each, followed by dehydra-
Ž .tion in 50, 70, 95 and 100% ethanol 3 min each . Brain
Ž .Fig. 4. Post-ischemic expression of bFGF immunoreactivity in the hippocampi. Brain sections 25 mm were from a rat subjected to 60 min ischemiaŽ . Ž . Ž .followed by 12 h reperfusion. Panel A shows bFGF immunoreactivity in the contralateral hippocampus and panels B to D the ipsilateral hippocampus
Ž .with different magnifications. The arrow heads point at the same cell that co-stained with both bFGF and GFAP immunoreactivity. Panel E shows theŽ .typical blue-black staining pattern of bFGF immunoreactivity. The open arrow points at bFGF immunoreactivity which is mainly in the nuclei. Panel F
shows the typical brown appearance of GFAP immunoreactivity. The filled arrow points at a GFAP-labelled astrocyte. Similar results were duplicated intwo other rats. Scale bars50 mm.

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265260
w xsections were exposed to Hyperfilm-MP 1 and autoradio-Žgraphs analyzed with a Bio-imaging analyzer system BAS
.1500rPictrography 3000, Fuji, Japan .
2.4. Immunohistochemical staining
Brain slices of 25 mm were frozen sectioned and incu-bated by free floatation. Endogenous peroxidase wasblocked with 0.3% hydrogen peroxide for 30 min at roomtemperature. Preincubation with 3% normal goat serumŽ .NGS containing 0.2% Triton X-100 was carried out atroom temperature for 60 min to block non-specific binding
Ž .of immunoglobin G IgG . Two commercially availableŽ .antibodies were used in the present study: 1 A DEAE
Žpurified mouse monoclonal anti-bFGF antibody UBI, New.York, 5 mgrml which recognizes the molecular confor-
mation related to the biological activity of bFGF and doesw xnot cross-react with aFGF 21,47 . The immunogen is
Ž .purified bovine brain basic FGF. 2 Rabbit anti-cow GFAPŽ .antibody DAKO, Kyoto, Japan, 20.5 mgrml which re-
acts strongly with human GFAP and with GFAP in almostall animal species tested. This antibody is particularlyuseful for distinguishing neoplasms of astrocyte origin inthe central nervous system. Sections were incubatedovernight with each antibody diluted in 1% NGS at 48C,rinsed with PBS for 30 min, and incubated in biotinylated
Ž .goat anti-mouse IgG bFGF immunostaining or biotinyl-
Ž .ated goat anti-rabbit IgG GFAP staining for 1 h. Afterseveral rinses with PBS, sections were incubated in avidinhorseradish peroxidase complex for 1 h, incubated in
X Ž .3,3 -diaminobenzidine DAB; 0.5 mgrml in the presenceof 0.003% H O and then mounted on gelatinized slides.2 2
The specificity of each antiserum was demonstrated by theabsence of stain when diluted primary antiserum waspreabsorbed with the respective antigen or was replaced bynormal serum. For negative controls, sections were incu-bated with heat denatured primary antibody. The results ofthese immunocytochemistry controls were consistentlynegative.
2.5. Double-staining immunohistochemistry
In order to ascertain the cellular localization of bFGF,sections were co-stained with a second antibody, anti-
Ž .GFAP, to detect the GFAP q astrocytes. The color reac-Ž .tion to detect the first antibody bFGF was developed
Ž .with nickel chloride 2 mgrml combined with DAB,which yielded a blue–black reaction product in lieu of thetypical brown reaction. After thorough rinsing in PBS,sections were incubated at 48C overnight with anti-GFAPantibody. The remaining process was the same as de-scribed above, except that the second antibody was de-tected with DAB alone, immunostaining astrocytes with a
w xbrown reaction product 25 .
Ž .Fig. 5. Post-ischemic expression of bFGF immunoreactivity in the ischemic cortex. Brain sections 25 mm were from a rat subjected to 60 min ischemiaŽ . Ž .and 12 h reperfusion. Panels a and b show the bFGF immunoreactivity in cortical area medial to the infarcted zone with different magnifications. Panels
Ž . Ž .c and d show bFGF immunoreactivity in cortical area inferior to the infarcted zone with different magnifications. I denotes the infarct zone and arrowheads point at cells expressing bFGF immunoreactivity. Similar results were observed in two additional rats. Scale bars50 mm.

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265 261
2.6. Chemicals
All chemicals were of reagent purity and purchased,Žunless otherwise indicated, either from E. Merck Darms-
. Ž .tadt, Denmark or Sigma Chem. Co. St. Louis, MO .
2.7. Statistics
Ž .One-way analysis of variance ANOVA was used tocompare the temporal expression of bFGF mRNA. Thelevel of significance for differences between two groupswas further analyzed with post-hoc Tukey’s protected t
Fig. 6. Effect of focal cerebral ischemia-reperfusion on the regional distribution of blood vessels in the cerebral cortex. Rats were subjected to 60 minischemia followed by 12 h of reperfusion. The dorsal view of the blood vessels in the cerebral cortex is shown in A, and the lateral views of the same
Ž . Ž .brain: the right ischemic side and the left control side are shown in B and C, respectively. Note increased density of blood vessels in the ischemic righthemisphere compared to the left non-ischemic hemisphere. Similar results were observed in two other rats.

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265262
Žtests using statistical software GB-STAT 2.1, Dynamic.Microsystem Inc., Silver Springs, Md . A probability value
of less than 0.05 was considered significant.
3. Results
Although there are at least 7 species of bFGF mRNAŽpresent in the adult rat brain, only one mRNA species 6.0
.kb from the total RNA extracted from both cerebralcortex and hippocampus hybridized to this truncated ratbFGF cDNA probe. Results from Northern blot analysisshowed basal expression of bFGF mRNA in the cerebral
Ž .cortex of sham-operated control Fig. 1, line 1, Sh . A 60min ischemic insult resulted in a transient induction of
Ž .bFGF mRNA in the ischemic cerebral cortex Fig. 1A butŽ .not on the contralateral side Fig. 1C . Levels of bFGF
mRNA were gradually increased and became significantlyelevated at 12 and 24 h after reperfusion and followed by agradual decline. Quantitative analysis of the blot indicateda 2.5-fold increase at the peak level at 12 h as compared to
Ž .the sham-operated control Fig. 1E . An increase in thelevels of bFGF mRNA was also observed in the ipsilateral
Ž .hippocampus Fig. 2 . However, in this brain region, asignificant increase in the levels of bFGF mRNA wasobserved as early as 60 min following ischemia and re-
Ž .mained elevated for up to 2 weeks of reperfusion Fig. 2 .Quantitative analysis also showed a similar 2.5-fold peakincrease as compared to the sham-operated control.
In situ hybridization was used to further investigate theŽ .regional expression of bFGF mRNA Fig. 3 . In this study,
Ž .brain slices 25 mm were obtained from rats subjected toŽ .60 min ischemia followed by 12 h of reperfusion A;C
Ž .and sham-operated control D . bFGF mRNA was ex-pressed in the indusium griseum and septohippocampal
Ž . Ž .nucleus A , fasciola cinereum and CA2 D regions in thesham controls, in agreement with findings reported by
w xEmoto et al. 14 . Following 60 min ischemia, enhancedexpression of bFGF mRNA was observed in the cortical
Ž . Ž .area surrounding the infarct peri-infarct area A;C andŽ .area within the infarct A,B . bFGF mRNA was also
slightly increased in the ipsilateral caudate putamen, amyg-Ž .daloid, pyriform cortex and hippocampus A;C . It is
noteworthy that the increased expression of bFGF mRNAŽ .within the infarct the demarcated area is confined mainly
to the rostral outer cortical layers of the infarct, an areaŽ .with increased density of blood vessels Fig. 6 . There wasŽ .no increase in the contralateral cortex A or sham-oper-
Ž .ated controls D .We further examined whether the increase in bFGF
mRNA resulted in an increase in the protein levels. bFGF-like immunoreactivity in the contralateral and ipsilateralhippocampus of rats subjected to 60 min ischemia fol-lowed by 12 h of reperfusion is shown in Fig. 4. bFGF-like
Ž .immunoreactivity appeared mainly in the nuclei E . Se-vere ischemia led to a striking increase in bFGF-like
Ž .immunoreactivity in the ipsilateral hippocampus B;D
as compared to the low basal level found in the contralat-Ž .eral side A . Double-staining of brain sections with anti-
GFAP and anti-bFGF antibodies indicated that the major-ity of the bFGF-like immunoreactivity was in astrocytesŽ .B;D , but was also noted in other GFAP negative cells.Although most of the bFGF positive cells were of astro-cyte origin, not all astrocytes showed bFGF-like immuno-
Ž .reactivity C and D . Typical appearance of GFAP-labeledŽ .astrocytes is shown F . Under the same conditions, the
bFGF-like immunoreactivity in the ischemic cortex isshown in Fig. 5. Enhanced expression of bFGF-like im-munoreactivity was found in the cortical area surroundingthe infarct. No bFGF-like immunoreactivity was found inthe contralateral areas. Selected neurons appeared to ex-hibit bFGF-like immunoreactivity, particular in corticallayers 5 and 6, and the retrosplenial agranular cortex.
Fig. 6 shows the regional distribution of blood vesselsin the brain of rats subjected to 60 min ischemia followedby 12 h of reperfusion. Results indicated that blood vessels
Ž .in the ischemic hemisphere b are more prominent andŽ .denser compared to those in the contralateral side c .
4. Discussion
Depending upon the severity of insult, prolonged cere-bral ischemia is known to cause neuronal dysfunction, celldeath and structural damage. Altered levels of severalpolypeptide growth factors have been reported to play arole in mediating growth and repair after CNS injuryw x18,52 . Among these factors, bFGF has been studied mostextensively. Although multiple species of bFGF mRNAtranscripts have been reported in adult rat brain, thephysiological role and regulatory mechanisms of thesedifferent mRNA transcripts in response to ischemic insulthave not been examined in detail. Results from Northernblot analysis clearly showed that only a single mRNAspecies within the total RNA extracted from both cerebralcortex and hippocampus hybridized to this truncated ratbFGF cDNA probe. This mRNA species corresponds tothe 6.0 kb transcripts, which is most abundant in adult rat
w xbrain 57 . Severe ischemic insult led to a transient in-crease in bFGF mRNA in the ischemic cortex. This resultis further supported by the in situ hybridization and im-munohistochemical data. Enhanced expression of bFGFmRNA was observed in the cortical area surrounding the
Ž .infarct Fig. 3 and increased bFGF-like immunoreactivityŽ .was also detected in the same brain regions Fig. 5 . Taken
together, these results suggest that increased bFGF-likeimmunoreactivity following ischemic insult is most likelydue to the increased expression of its 6.0 kb mRNAtranscripts. Double-staining of brain sections with anti-GFAP antibody further showed that most of the bFGFpositive cells were astrocytes. However, not all astrocytesshowed bFGF-like immunoreactivity and other GFAP neg-ative cells also showed bFGF-like immunoreactivity. Thisfinding is in agreement with the notion that in addition to

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265 263
astrocytes, bFGF is also expressed in activatedmacrophages, microglia, endothelial cells, neurons, and
w xextracellular matrix 12 . Although not significant, therewas a trend of second increase of bFGF mRNA at subse-
Ž .quent periods Fig. 1E, 762, ;4 weeks , and bFGF-likeimmunoreactivity was indeed increased exclusively in the
Žcortical area immediately adjacent to area of necrosis data.not shown .
Besides the ischemic cerebral cortex, enhanced expres-sion of this 6.0 kb bFGF mRNA was also noted in theipsilateral hippocampal region, in spite of the fact that no
w xobvious cellular damage was found 43 . This lack ofcongruence regarding the anatomical distribution of is-chemic infarct and that of bFGF expression was also noted
w x w xin the expression of fos and jun 2,49 , krox 20 1 ,w x w xNGFI-B 39 and heat shock protein 70 34,73 . These
profiles of gene expression are likely related to spreadingŽ . w xdepression repeated cortical depolarizations 22,29,50
Žandror diaschisis effects expressed in brain regions re-. w xmote from the initiation site 23,62 in response to the
ischemic insult. However, the exact mechanism andphysiological significance leading to this type of inductionremains to be determined. Unlike the ischemic cortex,which showed a transient induction of bFGF mRNA, asignificant increase in the levels of bFGF mRNA in theipsilateral hippocampus was observed as early as 60 minfollowing ischemia and remained elevated throughout the
Ž .entire ischemia-reperfusion periods studied Fig. 2 . Thus,w xunlike that of c-fos and c-jun 2 , bFGF shows a strikingly
different temporal induction profile between ischemic cor-tex and ipsilateral hippocampus. The mechanism andphysiological significance leading to this prolonged ex-pression in not known. However, it is likely that followingischemia prolonged expression of bFGF may contribute tocell survival, since recent studies have demonstrated theability of bFGF to regulate c-fos and Zifr268 expressionw x w x16 , stabilize calcium homeostasis 10 , and prevent
w x w xNMDA neurotoxicity 20 and nitric oxide toxicity 45 .Furthermore, systemic administration of bFGF has beenshown to reduce the infarct volume after ischemic insultw x30,35,37,44,53,54,66 .
Similar results have been reported by Speliotes et al.w x64 . Nevertheless, a number of findings reported herecertainly provide additional data to the current knowledgeof bFGF. First, we have shown a more complete time
Ž .course up to 4 weeks and report that bFGF mRNAexpression peaks at 12 h after the onset of reperfusion in
w xthe ischemic cortex. Since Speliotes et al. 64 did not havea 12 h time point, it is difficult to judge if their peak leveloccurred at day 1 or 12 h after ischemia in their studies.Second, we note a striking selective upregulation of bFGFin the ipsilateral hippocampus. A significant increase inlevels of bFGF mRNA was noted immediately following60 min of ischemia, whereas no significant increase was
w x w xobserved by Speliotes et al. 64 . Third, Speliotes et al. 64showed glial upregulation of bFGF in regions adjacent to
the infarct, except in the ipsilateral piriform cortex, whereneuronal upregulation of bFGF was noted. In our studies,the intense expression of bFGF mRNA in the outer layersof the ischemic right MCA cortex and adjacent cortex wasconsistently noted. It is possible that these differences arerelated to the application of different experimental strokemodels. The right MCA was permanently occluded in the
w xstudy by Speliotes et al. 64 . In our study, the right MCAand both CCAs were transiently occluded for 60 min.
In situ hybridization studies also revealed that increasedexpression of bFGF mRNA within the infarct area isconfined mainly to the rostral outer cortical layers of the
Ž .infarct and peri-infarct zones Fig. 3B , where the MCAŽ .trunk and the area showed prominent vascularity Fig. 6 .
Several studies have shown that bFGF has angiogenicw xproperties in vivo 8,12,68 , and can increase the expres-
sion of collagenase, plasminogen activator and other pro-w x w xteases 58,61 , increase collateral circulation 5,44,66,75 ,
w xas well as act as a vasodilator in rat brain 60 . Unfortu-nately, we are unable to determine with certainty whetherthis prominent vascularity is due to new vessels or simplyby increased dilation of existing blood vessels. Neverthe-less, these results are in line with the notion that angiogen-esis and collateral circulation play an important role inischemic injury.
In summary, our data demonstrate that ischemia en-hanced the expression of the 6.0 kb bFGF mRNA tran-scripts and subsequently led to an increased expression ofbFGF protein. In situ hybridization and immunohistochem-ical studies show that both bFGF mRNA and protein areincreased in the area surrounding the infarction. Althoughthe mechanism leading to the prolonged induction of bFGFmRNA in the ipsilateral hippocampus is not presentlyclear, our findings suggest the possibility that bFGF mayplay a crucial role in cell survival following cerebralischemia. Double-labeling immunohistochemical methodsshowed that in addition to astrocytes, bFGF was alsoexpressed in other cell populations, most likely neuronsand endothelial cells. Further studies are needed to confirmthis preliminary observation.
Acknowledgements
This work was supported by grants from National Sci-Žence Council NSC-85-2331-B001-039-M10, NSC-84-
.2331-B001-001 and NSC-83-0203-B001-102 andAcademia Sinica, Taipei, Taiwan, ROC. We thank Ms.Kelly Treat for editorial assistance.
References
w x1 G. An, T.N. Lin, J.S. Liu, C.Y. Hsu, Induction of krox-20 expres-sion after focal cerebral ischemia, Biochem. Biophys. Res. Com-
Ž .mun. 188 1992 1104–1110.

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265264
w x2 G. An, T.N. Lin, J.S. Liu, J.J. Xue, Y.Y. He, C.Y. Hsu, Expressionof c-fos and c-jun family genes after focal cerebral ischemia, Ann.
Ž .Neurol. 33 1993 457–464.w x3 K.J. Anderson, D. Dan, S. Lee, C.W. Cotman, Basic fibroblast
growth factor prevents death of lesioned cholinergic neurons in vivo,Ž .Nature 332 1988 360–361.
w x4 A. Baird, Fibroblast growth factors: activities and significance ofnon-neurotrophin neurotrophic growth factors, Curr. Opin. Neurol. 4Ž .1994 78–86.
w x5 S. Bemotat-Danielowski, H.S. Sharma, R.J. Schott, W. Schaper,Generation and localization of monoclonal antibodies against fibrob-last growth factors in ischemic collateralized porcine myocardium,
Ž .Cardiovasc. Res. 27 1993 1220–1228.w x Ž .6 W.H. Burgess, T. Maciag, The heparin-binding fibroblast growth
Ž .factor family of proteins, Annu. Rev. Biochem. 58 1989 575–606.w x7 C.G. Caday, M. Klagsbrun, P.J. Fanning, A. Mirzabegian, S.P.
Ž .Finklestein, Fibroblast growth factor FGF levels in the developingŽ .rat brain, Dev. Brain Res. 52 1990 241–246.
w x8 H.H. Chen, C.H. Chien, H.M. Liu, Correlation between angiogenesisand basic fibroblast growth factor expression in experimental brain
Ž .infarct, Stroke 25 1994 1651–1657.w x9 S.T. Chen, C.Y. Hsu, E.L. Hogan, H. Macriq, J.D. Balentine, A
model of focal ischemic stroke in the rat: reproducible extensiveŽ .cortical infarction, Stroke 17 1986 738–743.
w x10 B. Cheng, M.P. Mattson, NGF and bFGF protect rat hippocampaland human cortical neurons against hypoglycemic damage by stabi-
Ž .lizing calcium homeostasis, Neuron 7 1991 1031–1041.w x11 P. Chomczynski, N. Sacchi, Single-step of RNA isolation by acid
guanidinium thiocyanate-phenol-chloroform extraction, Anal.Ž .Biochem. 162 1987 156–159.
w x12 P. Cuevas, F. Carceller, G. Gimenez-Gallego, Fibroblast growthŽ .factor and cerebral ischemia, Neurol. Res. 16 1994 181–183.
w x13 W.D. Dietrich, O. Alonso, R. Busto, S.P. Finklestein, PosttreatmentŽ .with intravenous basic fibroblast growth factor bFGF reduces
histopathological damage following fluid-percussion brain injury inŽ .rats, J. Neurotrauma 13 1996 309–316.
w x14 N. Emoto, A.M. Gonzalez, P.A. Walike, E. Wada, D.M. Simmons,Ž .S. Shimasaki, A. Baird, Basic fibroblast growth factor FGF in the
central nervous system: identification of specific loci of basic FGFŽ .expression in the rat brain, Growth Factors 2 1989 21–29.
w x15 M. Endoh, W.A. Pulsinelli, J.A. Wagner, Transient global ischemiainduces dynamic changes in the expression of bFGF and the bFGF
Ž .receptor, Mol. Brain Res. 22 1994 76–88.w x16 L. Ferhat, M. Khrestchatisky, M.P. Roisin, G. Barbin, Basic fibrob-
last growth factor-induced increase in Zifr268 and c-fos mRNAlevels is Ca2q dependent in primary cultures of hippocampal neu-
Ž .rons, J. Neurochem. 61 1993 1105–1112.w x17 S.P. Finklestein, P.J. Apostolides, C.G. Caday, J. Prosser, M.F.
Phillips, M. Klagsbrun, Increased basic fibroblast growth factorŽ .bFGF immunoreactivity at the site of focal brain wounds, Brain
Ž .Res. 460 1988 253–259.w x18 S.P. Finklestein, C.G. Caday, M. Kano, D.J. Berlove, C.Y. Hsu, M.
Moskowitz, M. Klagsbrun, Growth factor expression after stroke,Ž . Ž .Stroke 21 III 1990 122–124.
w x19 M. Fisher, M.E. Meadows, T. Do, J. Weise, V. Trubetskoy, M.Charette, S.P. Finklestein, Delayed treatment with intravenous basicfibroblast growth factor reduces infarct size following permanentfocal cerebral ischemia in rats, J. Cereb. Blood Flow Metab. 15Ž .1995 953–959.
w x20 A. Freese, S.P. Finklestein, M. DiFiglia, Basic fibroblast growthfactor protects striatal neurons in vitro from NMDA-receptor medi-
Ž .ated excitotoxicity, Brain Res. 575 1992 351–355.w x21 Y.M. Fu, P. Spirito, Z.X. Yu, S. Biro, J. Sasse, J. Lei, V.J. Ferrans,
S.E. Epstein, W. Casscells, Acidic fibroblast growth factor in theŽ .developing rat embryo, J. Cell Biol. 114 1991 1261–1273.
w x22 R. Gill, O. Andine, L. Hillered, L. Persson, H. Hagberg, The effectof MK-801 on cortical spreading depression in the penumbra zone
following ischemia in the rat, J. Cereb. Blood Flow Metab. 12Ž .1992 371–379.
w x23 M.D. Ginsberg, M. Reivich, A. Giandomenico, J.H. Greenberg,Local glucose utilization in acute focal cerebral ischemia: local
Ž .dysmetabolism and diaschisis, Neurology 27 1977 1042–1048.w x24 F. Gomez-Pinilla, C.W. Cotman, Transient lesion-induced increase
Ž .of bFGF and its receptor in layer VI subplate cells of the adult ratbŽ .cerebral cortex, Neuroscience 49 1992 771–780.
w x25 F. Gomez-Pinilla, J.W. Lew, C.W. Cotman, Basic FGF in adult ratbrain: cellular distribution and response to entorhinal lesion and
Ž .fimbria-fornix transection, J. Neurosci. 12 1992 345–355.w x26 D. Gospodarowicz, Localization of a fibroblast growth factor and its
effect alone and with hydrocortisone on 3T3 cell growth, Nature 249Ž .1974 123–127.
w x27 D. Gospodarowicz, N. Ferrara, L. Schweigerer, G. Neufeld, Struc-tural characterization and Biological functions of FGF, Endocrine
Ž .Rev. 8 1987 95–114.w x28 J.G. Heuer, C.S. Von Bartheld, Y. Kinoshita, P.C. Evers, M. Both-
well, Alternating phases of FGF receptor and NGF receptor expres-Ž .sion in the developing chicken nervous system, Neuron 5 1990
283–296.w x29 R. Iijuma, G. Mies, K.A. Hossmann, Repeated negative DC deflec-
tions in rat cortex following middle cerebral artery occlusion areabolished by MK-801: effect on volume of ischemic injury, J. Cereb.
Ž .Blood Flow Metab. 12 1992 727–733.w x30 N. Jiang, S.P. Finklestein, T.Y. Do, C.G. Caday, M. Charette, M.
Chopp, Delayed intravenous administration of basic fibroblast growthŽ .factor bFGF reduces infarct volume in a model of focal cerebral
Ž .ischemiarreperfusion in the rat, J. Neurol. Sci. 139 1996 173–179.w x31 B.B. Johansson, M. Grabowski, Functional recovery after brain
infarction: plasticity and neural transplantation, Brain Pathol. 4Ž .1994 85–95.
w x32 T. Kato, S. Nakano, K. Kogure, H. Sasaki, K. Koiwai, Y. Yamasaki,T. Katagiri, H. Sasaki, The binding of basic fibroblast growth factorto ischemic neurons in the rat, Neuropathol. Appl. Neurobiol. 18Ž .1992 282–290.
w x33 T. Kawamata, N.E. Alexis, W.D. Dietrich, S.P. Finklestein, Intracis-Ž .ternal basic fibroblast growth factor bFGF enhances behavioral
recovery following focal cerebral infarction in the rat, J. Cereb.Ž .Blood Flow Metab. 16 1996 542–547.
w x34 H. Kinouchi, F.R. Sharp, J. Koistinaho, K. Hicks, H. Kamii, P.H.Chan, Induction of heat shock hsp70 mRNA and HSP70 KDaprotein in neurons in the penumbra following focal cerebral is-
Ž .chemia in the rat, Brain Res. 619 1993 334–338.w x35 P.B. Kirschner, R. Henshaw, J. Weise, V. Trubetskoy, S. Fin-
klestein, B.J. Schulz, M.F. Beal, Basic fibroblastic growth factorprotects against excitotoxicity and chemical hypoxia in both neona-
Ž .tal and adult rats, J. Cereb. Blood Flow Metab. 15 1995 619–623.w x36 Y. Kiyota, K. Takami, M. Iwane, A. Shino, M. Miyamoto, R.
Tsukuda, A. Nagaoka, Increase in basic fibroblast growth factor-likeimmunoreactivity in rat brain after forebrain ischemia, Brain Res.
Ž .545 1991 322–328.w x37 N. Koketsu, D.J. Berlove, M.A. Moskowitz, N.W. Kowall, C.G.
Caday, S.P. Finklestein, Pretreatment with intraventricular basicfibroblast growth factor decrease infarct size following focal cere-
Ž .bral ischemia in rats, Ann. Neurol. 35 1994 451–457.w x38 T. Kurokawa, M. Seno, K. Igarashi, Nucleotide sequence of rat basic
Ž .fibroblast growth factor cDNA, Nucleic Acid Res. 16 1988 5201.w x39 T.N. Lin, J.J. Chen, S.J. Wang, J.T. Cheng, S.I. Chi, A.B. Shyu,
G.Y. Sun, C.Y. Hsu, Expression of NGFI-B mRNA in a rat focalŽ .cerebral ischemia-reperfusion model, Mol. Brain Res. 43 1996
149–156.w x40 T.N. Lin, Y.Y. He, G. Wu, M. Khan, C.Y. Hsu, Effect of Brain
edema on infarct volume in a focal cerebral ischemia model in rat,Ž .Stroke 24 1993 117–121.
w x41 T.N. Lin, Y.P. Wong, J.J. Chen, J.T. Cheng, S.F. Yu, S.H. Sun, S.I.Chi, C.Y. Chai, Elevated basic fibroblast growth factor levels in

( )T.N. Lin et al.rMolecular Brain Research 49 1997 255–265 265
stroke-prone spontaneously hypertensive rats, Neuroscience 76Ž .1997 557–570.
w x42 A. Lippoldt, B. Andbjer, L. Rosen, E. Richter, D. Ganten, Y. Cao,R.F. Pettersson, K. Fuxe, Photochemically induced focal cerebralischemia in rat: time dependent and global increase in expression of
Ž .bFGF mRNA, Brain Res. 625 1993 45–56.w x43 T.H. Liu, J.S. Beckman, B.S. Freeman, E.L. Hogan, C.Y. Hsu,
Polyethylene glycol-conjugated superoxide dismutase and catalaseŽ .reduce ischemia brain injury, Am. J. Physiol. 256 1989 H589–
H593.w x44 M.K. Lyons, R.E. Anderson, F.B. Meyer, Basic fibroblast growth
factor promotes in vivo cerebral angiogenesis in chronic forebrainŽ .ischemia, Brain Res. 558 1991 315–320.
w x45 K. Maiese, I. Boniece, D. DeMeo, J.A. Wagner, Peptide growthfactors protect against ischemia in culture by preventing Nitric oxide
Ž .toxicity, J. Neurosci. 13 1993 3034–3040.w x46 A. Matsuyama, H. Iwata, N. Okumura, S. Yoshida, K. Imaizumi, Y.
Lee, S. Shiraishi, S. Shiosaka, Localization of bFGF-like immuno-Ž .reactivity in the rat brain, Brain Res. 587 1992 59–65.
w x47 K. Matsuzaki, Y. Yoshitake, Y. Matuo, H. Sasaki, K. Nishikawa,Monoclonal antibodies against heparin-binding growth factorIIrbasic fibroblast growth factor that block its biological activity: invalidity of the antibodies for tumor angiogenesis, Proc. Natl. Acad.
Ž .Sci. USA 86 1989 9911–9915.w x48 R.S. Morrison, A. Sharma, J. devellis, R. Bradshaw, Basic fibroblast
growth factor supports survival of cerebral cortical neurons inŽ .primary cultures, Proc. Natl. Acad. Sci. USA 83 1986 7537–7541.
w x49 J.R. Naranjo, B. Mellstrom, M. Achaval, J.J. Lucas, J. Del Rio, P.Sassone-Corsi, Co-induction of jun B and c-fos in a subset of
Ž .neurons in the spinal cord, Oncogene 6 1991 223–227.w x50 M. Nedergaard, A.J. Hansen, Characterization of cortical depolariza-
tion evoked in focal cerebral ischemia, J. Cereb. Blood Flow Metab.Ž .13 1993 568–574.
w x51 G. Neufeld, D. Gospodzrowica, Basic and acidic fibroblast growthfactors react with the same surface receptors, J. Biol. Chem. 261Ž .1986 5631–5637.
w x52 M. Nieto-Sampedro, Growth factor induction and order of event inŽ .CNS repair, in: D.G. Stain, B.S. Sabel Eds. , Pharmacological
Approaches to the Treatment of Brain and Spinal Cord Injury,Plenum Press, New York, 1988, pp. 301-337.
w x53 K. Nozaki, S.P. Finklestein, M.F. Beal, Basic Fibroblast Growthfactor protects against Hypoxia-Ischemia and NMDA Neurotoxicity
Ž .in neonatal rats, J. Cereb. Blood Flow Metab. 13 1993 221–228.w x54 K. Nozaki, S.P. Finklestein, M.F. Beal, Delayed administration of
basic fibroblast growth factor protects against N-methyl-D-aspartateŽ .neurotoxicity in neonatal rats, Eur. J. Pharmacol. 232 1993 295–
297.w x55 D. Otto, K. Unsicker, Basic FGF reverses chemical and morphologi-
cal deficits in the nigro-striatal system of MPTP-treated mice, J.Ž .Neurosci. 10 1990 1912–1921.
w x56 B. Pettman, G. Labourdette, M. Weibel, M. Sensenbrenner, TheŽ .brain fibroblast growth factor FGF is localized in neurons, Neu-
Ž .rosci. Lett. 68 1986 175–180.w x57 P.P. Powell, S.P. Finklestein, C.A. Dionne, M. Jaye, M. Klagsborun,
Temporal, differential and regional expression of mRNA for basicfibroblast growth factor in the developing and adult rat brain, Mol.
Ž .Brain Res. 11 1991 71–77.w x58 M. Presta, D. Moscatelli, J. Joseph-Silverstein, D.B. Rifkin, Purifica-
tion from a human hepatoma cell line of a basic fibroblast growthfactor-like molecule that stimulates capillary endothelial plasmino-
gen activator production, DNA synthesis and migration, Mol. CellŽ .Biol. 6 1986 4060–4066.
w x59 M.A. Riva, K. Gale, I. Mocchetti, Basic FGF mRNA increase inspecific brain regions following convulsive seizures, Mol. Brain Res.
Ž .15 1992 311–318.w x60 S. Rosenbaltt, K. Irikura, C.G. Caday, S.P. Finklestein, M.A.
Moskowitz, Basic fibroblast growth factor dilates rat pial arterioles,Ž .J. Cereb. Blood Flow Metab. 14 1994 70–74.
w x61 G.A. Rosenberg, M. Navratil, F. Barone, G. Feuerstein, Proteolyticcascade enzymes increase in focal cerebral ischemia in rat, J. Cereb.
Ž .Blood Flow Metab. 16 1996 360–366.w x62 S.M. Sagar, The molecular biology of brain ischemia: trendy genes
Ž .in sick neurons, Ann. Neurol. 33 1993 437–438.w x63 L. Schweigerer, Fibroblast growth factor and angiogenesis, Z. Kar-
Ž . Ž .diol. 78 Suppl. 6 1989 12–15.w x64 E.K. Speliotes, C.G. Caday, T. Do, J. Weise, N.W. Kowall, S.P.
Finklestein, Increased expression of basic fibroblast growth factorŽ .bFGF following focal cerebral infarction on the rat, Mol. Brain
Ž .Res. 39 1996 31–42.w x65 K. Takami, M. Iwane, Y. Kiyota, M. Miyamoto, R. Tsukuda, S.
Shiosaka, Increase of bFGF immunoreactivity and its mRNA levelin rat brain following transient forebrain ischemia, Exp. Brain Res.
Ž .90 1992 1–10.w x66 R. Tanaka, Y. Miyasaka, K. Yada, T. Ohwada, T. Kameya, Basic
fibroblast growth factor increase regional cerebral blood flow andreduces infarct size after experimental ischemia in a rat model,
Ž .Stroke 26 1995 2154–2158.w x67 P.S. Thomas, Hybridization of denatured RNA and small DNA
fragments transferred to nitrocellulose, Proc. Natl. Acad. Sci. USAŽ .77 1980 5201–5205.
w x68 J.A. Thompson, C.C. Howdenschild, K.D. Anderson, J.M. DiPietro,W.F. Anderson, T. Maciag, Heparin-binding growth factor 1 inducesthe formation of organoid neovascular structures in vivo, Proc. Natl.
Ž .Acad. Sci. USA 86 1989 7928–7932.w x69 I. Torres-Aleman, F. Naftolin, R.J. Robbins, Trophic effects of basic
fibroblast growth factor on fetal rat hypothalamic cells: interactionsŽ .with insulin-like growth factor I, Dev. Brain Res. 52 1990 253–257.
w x70 D.A. Walicke, Basic and acidic fibroblast growth factors havetrophic effects on neurons from multiple CNS regions, J. Neurosci. 8Ž .1988 2618–2627.
w x71 A. Wanaka, E.M. Johnson Jr., J. Milbrandt, Localization of FGFreceptor mRNA in the adult rat central nervous system by in situ
Ž .hybridization, Neuron 5 1990 267–281.w x72 B. Weise, T. Janet, C. Grothe, Localization of bFGF and FGF-recep-
tor in the developing nervous system of the Embryonic and newŽ .born rat, J. Neurosci. Res. 34 1993 442–453.
w x73 F.A. Welsh, D.J. Moyer, V.A. Harris, Regional expression of heatshock protein 70 mRNA and c-fos mRNA following focal ischemia
Ž .in the rat brain, J. Cereb. Blood Flow Metab. 12 1992 204–212.w x74 K. Yamada, A. Kinoshita, E. Kohmura, T. Sakaguchi, J. Taguchi, K.
Kataoka, T. Hayakawa, Basic fibroblast growth factor preventsthalamic degeneration after cortical infarction, J. Cereb. Blood Flow
Ž .Metab. 11 1991 472–478.w x75 A. Yanagisawa-Miwa, Y. Uchida, F. Nakamura, T. Tomaru, H.
Kido, T. Kamijo, T. Sugimoto, K. Kaji, M. Utsugama, C. Kurashima,H. Ito, Salvage of infarcted myocardium by angiogenic action of
Ž .basic fibroblast growth factor, Science 257 1992 1401–1403.w x76 P.K. Yip, Y.Y. He, C.Y. Hsu, N. Garg, P. Marangos, E.L. Hogan,
Effect of plasma glucose on infarct size in focal cerebral ischemia-Ž .reperfusion, Neurology 41 1991 899–905.




















