
Subpopulations of marrow stromal cells share a variety of osteoblastic markers
Upload
independentCategory
view
0download
0
Serum from Postmenopausal Women Directs Differentiation of Human
Clonal Osteoprogenitor Cells from an Osteoblastic toward an Adipocytic
Phenotype
Bradley Stringer,1 Rachel Waddington,2 Adam Houghton,3 Mike Stone,4 Graham Russell,5 George Foster6
1Department of Oral Surgery, Medicine and Pathology, Dental School, Cardiff University, Heath Park, Cardiff, CF14 4XY, UK2Department of Dental Health and Biological Sciences, Dental School, Cardiff University, Cardiff, CF14 4XY, UK3Health Care Research Center, Proctor and Gamble, Box 1057, Mason, OH, 45040-9462, USA4Llandough Hospital, Medical School, Cardiff University, Cardiff, CF64 2XX, UK5Nuffield Department of Orthopedic Surgery, University of Oxford, Willmill Rd. Headington, Oxford, OX37LD, UK6School of Biosciences, Cardiff University, P.O.Box 911, Cardiff, CF10 3US, UK
Received: 7 August 2006 / Accepted: 13 January 2007 / Online publication: 4 April 2007
Abstract. A consistent observation in osteoporosis isbone volume reduction accompanied by increasedmarrow adipose tissue. No single cause linking the twophenomena has yet been identified. In a human pro-genitor cell clone (hOP 7) derived from bone marrow,however, we have demonstrated that rabbit serum candirect differentiation away from an osteoblast lineage toone of adipocytes. We now report on whether humanserum has a similar effect. Serum was collected from 10pre- and 10 postmenopausal women and from the 10postmenopausal women before and following 6-weekhormone replacement therapy (HRT). hOP 7 cells werecultured with the various sera, and after 7-14 daysadipocytogenesis was determined by oil red O stainingand lipoprotein lipase (LPL) and glycerol 3-phosphatedehydrogenase (G3PDH) expression. Incubation with10% premenopausal serum led to labeling of 10.9% ofcells (P < 0.05) with oil red O, whereas application of10% postmenopausal serum led to a much larger effect,43.5% labeling (P < 0.001 with respect to premeno-pausal serum). Oil red O positivity was accompanied byloss of type I collagen expression and increased LPL andG3PDH expression. HRT did not reverse the adipocy-togenic effect of postmenopausal serum. In conclusion,serum from postmenopausal women contains factorsthat steer hOP 7 bone progenitor cells toward an adip-ocytic phenotype, irrespective of HRT. The study sug-gests a role for serum factors in the development of fattymarrow in postmenopausal osteoporosis.
Key words: Adipocyte — Differentiation — Menopause— Osteoblast — Serum
Although the pathogenesis of osteoporosis is multifac-torial, one consistent finding is of a reciprocal relation-ship between trabecular bone loss and marrow fat gain[1�7]. To explain this connection, it has been suggested
that the balance between adipocyte and osteoblast pro-duction by progenitor cells within the marrow is some-how disturbed, leading to increased adipocyte numbersat the expense of bone-forming osteoblasts during thepathogenesis of the disease. Such a hypothesis is con-sistent with our understanding of marrow stromal celldifferentiation. Bone-forming osteoblasts and marrowadipocytes are both derived from a common mesen-chymal stem cell, which can also give rise to chondro-blasts, fibroblasts, myocytes, and tendon cells [forreview, see 8]. Furthermore, we [9] and others [10, 11]have shown a closer cellular relationship betweenosteoblast and adipocyte formation than was firstthought, whereby bipotential osteoblastic/adipocyticlate progenitor cells have been identified and clonedfrom human marrow. Interestingly, the adipocytic orosteoblastic differentiation of these cell populations canbe directed in vitro by modifying the cell culture condi-tions.
The knowledge gained from in vitro cell differentia-tion studies combined with reports describing themolecular events involved in adipocyte-specific tran-scriptional activation [12] gives us the confidence topredict a likely mechanism to account for this reciprocalrelationship [13, 14]. However, despite these and addi-tional in vivo studies using knockout mice, e.g., to ex-plore the cellular and molecular ways in which such areciprocal interaction might occur [15�20], no in vivocausal relationship has yet been identified to explain theclinical presentation of bone loss linked to marrow fatgain in osteoporosis [21�24].
During the process of selecting culture conditions forgrowing mesenchymal stem/progenitor cells, it wasshown that serum derived from various sources couldhave differing effects on mesenchymal stem/progenitorcell differentiation. For example, the murine stromal cell
Correspondence to: B. M. J. Stringer; E-mail: [email protected]
Calcif Tissue Int (2007) 80:233–243
DOI: 10.1007/s00223-007-9016-2

lines BMS2 and +/+2.4 undergo adipocytogenesis inthe presence of fetal bovine serum (FBS) [25], but FBShas no identified adipogenic effect on human osteo-blastic/adipocytic clonal cell lines [9�11]. However,rabbit serum is a potent stimulator of adipocytogenesisof both human and rodent mesenchymal stem/progeni-tor cells, as determined by adipocyte-specific transcrip-tional activation as well as cytochemical localization oflipid product [9, 11, 26].
The recognition of a reciprocal relationship betweenmarrow adipocytes and osteoblasts and that differentanimal sera can influence the differentiation programs ofmesenchymal progenitor cells derived from human andanimal sources have led us and others [27] to postulatethere may be agents present also in human serum thatinfluence mesenchymal stem/progenitor cell differentia-tion. Further, we suggest that perturbation in theseagents might be responsible for the concomitant increasein marrow fat as bone is lost during the development ofosteoporosis.
In view of the above, this study was performed todetermine whether human serum, taken from pre-menopausal and postmenopausal women, could affectthe differentiation of the human clonal bipotentialadipocytogenic/osteoblastic cell line hOP 7. It was alsointended to discover whether any effects of postmeno-pausal serum might be modified by hormone replace-ment therapy (HRT).
Materials and Methods
Materials
Costar (High Wycombe, UK) tissue culture plastics were usedfor all cell cultures. a-Minimal essential medium (a-MEM)tissue culture medium, glutamine, penicillin/streptomycin,TRIzol reagent, and Taq DNA polymerase were purchasedfrom Life Technologies (Paisley, UK). FBS, normal rabbitserum, naphthol-AS-MX-phosphate substrate, 3-isobutyl-1-methylxanthine (IBMX), human recombinent-insulin, 1,25-dihydroxyvitamin D3 (1,25[OH]2D3), vitamin K, hematoxylin,and oil red O staining solutions were obtained from Sigma(Dorset, UK). The human osteocalcin radioimmunoassay kitwas purchased from Immunotopics (La Jolla, CA). The mur-ine Moloney leukemia virus reverse transcriptase (MMLV-RT), deoxynucleotide triphosphates (dNTPs), the pGEM-Tcloning vector kit, the Wizard plus SV minipreps, 100 bppGEM and /X174HaeIII restricted DNA ladders, agarose forgel electrophoresis, and restriction enzymes were purchasedfrom Promega (Southampton, UK). Type 1a(I) procollagen,glycerol 3-phosphate dehydrogenase (G3PDH), lipoproteinlipase (LPL), and b2-microglobulin primer sequences usedwere as described previously [7] and obtained from Invitrogen(Paisley, UK). All other biochemical and molecular biologicalreagents were purchased from Sigma.
Human Serum
Permission was obtained from the South Glamorgan HealthAuthority Division of Medicine Ethics Committee and fromthe South East Wales Local Research Ethics Committee for the
collection of blood samples from patient volunteers undergoingbone-density scanning. At the time of the study, no volunteersexhibited detectable signs of osteoporosis, i.e., bone mineraldensity (BMD) more than 2.5 standard deviations below theyoung adult mean. The volunteers comprised premenopausalwomen aged 19�24 (mean 20.7) years and postmenopausalwomen aged 48�74 (mean 57.4) years. Menopausal status wasdetermined by lack of menstrual period for at least 12 months,follicle-stimulating hormone levels >40 IU/L, and serumestradiol <30 pg/mL. A regimen of up to 2 mg/day 17b-estradiol was used in HRT. The established dose of 17b-estradiol 0.5�2.0 mg was used daily, which was effective atameliorating some of the climacteric and other symptoms of thepostmenopausal syndrome including increasing trabecularbone or BMD and reducing the risk of fracture [28�31]. Thesedoses have been previously shown to increase serum concen-trations [17b-estrodiol] from <10�20 pg/mL in untreatedpostmenopausal women [32, 33] to a mean postestradiolreplacement therapy of 114 pg/mL [34]. Furthermore, 30�35pg/mL circulating 17b-estradiol is sufficient to relieve acutemenopausal symptoms [35], while 40�50 pg/mL is required forslowing osteoporosis [36]. Samples (20 mL) of fresh bloodobtained by venepuncture were allowed to clot overnight at4�C, followed by centrifugation at 400 · g. The serum layer wasremoved and separated into 2 mL aliquots prior to being frozenat )80�C for later studies. Serum samples were also taken andtreated in the same way, following 6 weeks of HRT.
Cell Culture
The conditionally immortalized human bipotential osteopro-genitor cell line hOP 7 was maintained as described previously[9] in basal medium comprising a-MEM, glutamine (2 mM),nonessential amino acids, penicillin (50 U/mL)/streptomycin(50 lg/mL) antibiotic, and 10% heat-inactivated FBS (HI-FBS). hOP 7 cells were incubated at 33�C, the permissivetemperature of the SV40-T immortalizing oncogene withwhich they had been transduced, which enabled cell replicationand passaging twice per week. In some experiments, cell rep-lication was attenuated by incubation at 39�C, the nonper-missive temperature of the immortalizing SV40-T oncogene. Insome experiments, FBS was replaced with 10% human serumor 10% normal rabbit serum (NRS).
Cell Counts
For analysis of the adipocytogenic effects of human sera, eachserum sample was assayed using procedures that followed thehistomorphometric principles of Weibel [37]. The study wasperformed on three separate occasions, and single, represen-tative experiments are provided. A single trained observer,who was blinded to the culture conditions, performed the cellcounting. To obtain a random field, standard stereologicalstrategies were followed [38, 39]. Multiple counting was per-formed on a series of selected fields by the same observer toascertain intra-assay observer variation, which was consis-tently <5%. No significant variation (i.e., >5%) in cellcounting between trained observers could be distinguished.
Alkaline Phosphatase Activity
Alkaline phosphatase (AP) was localized histochemically usingfreshly prepared 0.25% w/v of naphthol-AS-MX-phosphatesubstrate. Cells were first washed with phosphate-bufferedsaline (PBS), then fixed in 95% v/v ethanol for 10 minutes atroom temperature. The cells were then washed in deionizedwater for 1 minute, and sufficient substrate solution was addedto the cultures to cover all cells in the staining solution. Plateswere wrapped in foil and incubated at room temperature for 15minutes, followed by three washes in deionized water. The cellswere then counterstained in hematoxylin for 2 minutes and
234 B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation

washed three times in deionized water, and plates were allowedto air-dry prior to examination.
In addition, AP activity was measured quantitatively using amodification of the colorimetric method described first by Bes-sey et al. in 1946 [40]. In short, cell lysates were prepared bywashing the cells in PBS (three times), transferring the cell layerinto 200 lL of ice-cold 0.1% Triton X-100, and subjecting thesamples to mild sonication. Aliquots of the samples were thenincubated with prewarmed substrate solution (0.1 M dietha-nolamine, 1 mM MgCl2, and 2 mM p-nitrophenylphosphate[p-NP], pH 10.5) for 1 hour. The reaction was halted by theaddition of 0.1 M NaOH, and samples were assayed spectro-photometrically at 410 nm. The final p-NP concentration wasestimated by reference to a p-NP standard curve. Specific APactivity was calculated for each sample by standardizing the APvalues per unit protein each hour. The protein content of the celllayers was measured using the Bio-Rad (Richmond, CA) pro-tein assay, based on the Bradford dye-binding procedure [41].
Osteocalcin Assay
hOP 7 cells were seeded in six-well plates at a density of 104/cm2
and grown to confluence at 33�C prior to treatment with either1,25(OH)2D3 and 1 lM vitamin K2 or vehicle (ethanol, 0.1% v/v) at 39�C. Osteocalcin was assayed by an immunoradiometricassay, detecting intact osteocalcin and the large N-terminal andthe mid-fragments. Conditioned media were harvested fromhOP 7 cells after incubation with active vitamin D3 or vehiclefor 8 days. Medium was incubated with the cells for a 48-hourperiod prior to its collection and storage. Samples of mediumwere removed and stored at )80�C prior to assay.
Induction and Localization of Mineralized Matrix
Postconfluent hOP 7 cells were incubated in basal mediumsupplemented with 50 lg/mL ascorbic acid and 1 mM b-glycerophosphate. Cells were incubated in the presence orabsence of 10)7 M dexamethasone at 39�C for a period of 28days. Cultured cells were then washed in PBS and fixed in 10%(v/v) phosphate-buffered formalin in saline for 10 minutes atroom temperature and stained using the von Kossa method[42]. In short, cultures were immersed in a silver nitrate solu-tion and exposed to ultraviolet light for 1 hour. The excesssilver nitrate was washed away following exposure, and thecells were treated with 5% sodium thiosulfate for 5 minutes atroom temperature. The cells were subsequently washed withdistilled water and the plates allowed to dry in air.
Induction of Differentiation into an Adipocyte-like Phenotype
hOP 7 cells were seeded at 104/cm2 and cultured in basalmedium at the permissive temperature of 33�C until theyreached 60% confluence. The medium was then replaced with amodified basal medium containing 10% adult NRS, a knowninducer of adipocytic differentiation in marrow-derived stro-mal cell cultures [43], in place of HI-FBS. The cells were thenincubated in the serum-modified basal medium at 39�C forperiods up to 7 days. Cells were also incubated in basal med-ium where the HI-FBS was replaced by human serum derivedfrom various sources (see below).
In addition to NRS stimulation of adipocyte differentiation,cells were incubated for 14 days in basal medium containing 100lg/mL IBMX, 10 lg/mL insulin, and 10)7 M dexamethasone,which had been shown in previous studies to induce adipocy-togenesis of cloned marrow progenitor cells [10, 11, 26, 44].
Identification of Lipid Accumulation
Following culture as outlined above, the cells were washedtwice with PBS and fixed with 10% (v/v) phosphate-buffered
formalin in saline. The standard oil red O histological methodwas used to identify cells with lipid inclusions [42] as describedpreviously [9, 10, 11, 25, 26].
Quantification of Adipocytic Cells
A trained operator, who was blind to the culture conditions,performed the study. The number of oil red O-positive cells perfield was determined, as was the total number of cells (positiveand negative) per field viewed. For each serum sample studied,four random fields (actual field size 0.09 mm2), delineated withan eyepiece graticule, were selected from each of four wellsfrom a 12-well tissue culture plate. The study was separatelyperformed three times, and representative data are provided.
The assessment of cells stained positively for oil red O, astandard marker for adipocytogenesis, was as follows. An in-verted microscope, with ·20 objective and ·10 eyepiece withgraticule covering 0.09 mm2 area, was used to scan each cho-sen field. Fields were selected randomly by choosing the firstcomplete field from the top left corner of each well. Four fieldsscanned from each well were selected by choosing the third,fifth, and seventh complete fields in turn from each well,respectively, as described previously [38, 39]. The total cellspositive and negative for oil red O were counted for each field.Four fields from each of four wells were quantified for eachserum sample.
Reverse Transcription Polymerase Chain Reaction
hOP 7 cells were seeded in flasks at 10,000/cm2 and cultured atthe permissive temperature of 33�C in basal medium until theyreached confluence. Cells were then incubated at 39�C either inbasal medium, in modified basal medium where 10% NRS wassubstituted for the FBS, or in basal medium where 10% pre- orpostmenopausal human serum was substituted for FBS. After7 days, the medium was removed and cells were washed twicewith PBS, TRIzol reagent was added (1 mL/10 cm2), and RNAwas extracted as recommended by Life Technologies. First-strand cDNA was synthesized from 5 lg of RNA, with 0.2 lgof oligo(dT) primer in the presence of dNTP mix (0.5 mM ofeach) by MMLV-RT (100 units) in a final volume of 20 lL.The prepared RNA and primer were then incubated togetherat 70�C for 10 minutes and cooled rapidly on ice before addingthe other reagents. Reverse transcription (RT) reaction wasallowed to take place for 1 hour at 37�C. This was followed byheat treatment (95�C) for 5 minutes to inactivate the MMLV-RT enzyme.
The cDNA produced by the method described above wasthen amplified by polymerase chain reaction (PCR) using the‘‘hot-start’’ method [45] in a 50 lL reaction volume, containinga mix of dNTP (0.2 mM of each), specific primers (0.5 mM ofeach), Taq DNA polymerase (1 unit), 20 mM Tris-HCl (pH8.4), 50 mM KCl, and 1.5 mM MgCl2.
RT-PCR primers used in the studies were as follows: b2-microglobulin (237 bp), forward 5¢-ggAgACAgCACTCAAAgTAg and reverse 5¢-ggTgTgAACCATgTgACTTT; collagentype 1a(I) chain (235 bp), forward 5¢-CTTggTCTCgTCACA-gATC and reverse 5¢-CTTTTAACgTTggAggATgTgC-TATTTggC; LPL (324 bp), forward 5¢-gAAAgTgTCTACTTTgCAgAAAggAAAAggC and reverse 5¢-CTTTTAACgTTggAggATgTgCTATTTggC; G3PDH (568 bp), forward 5¢-TTgCAACTgTTTTAggACTTT and reverse 5¢-AgCATTgggAAATgTTCAAgg; peroxisome proliferator-activated receptor(PPAR) c2 (310 bp), forward 5¢-TgAAACTCTgggAgATTCTCCTATTgACCC and reverse 5¢-TCAgAATAATAAggTggAgATgCAggCTCC.
PCR amplifications were performed using an Autogene IIthermal cycler (Grant, Dorset UK), whereby 30 cycles werecompleted after an initial denaturation at 94�C for 4 minutes.The same amplification cycles (94�C for 1 minute, 60�C for 2minutes, and 72�C for 2 minutes) were used for all PCRamplifications performed in this study. PCR products were
B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation 235

analyzed by electrophoresis of 16 lL samples in 1.5% agarose(w/v) gels, and DNA fragments were visualized by stainingwith ethidium bromide.
Sequencing of PCR Products
The Promega pGEM-T vector system was used to clone PCRproducts according to the manufacturer�s instructions. Inshort, 1 lL of PCR product was ligated with 50 ng pGEM-Tvector in the presence of 3 Weiss units of T4 DNA ligase. TheDNA ligations were used to transform competent JM109bacteria. Transformation-competent cells were isolated bystreaking Luria Bertani (LB) Agar plates overlaid with ampi-cillin/isopropylthiogalactoside/X-Gal (b-D-galactopyranoside)solution, and incubating them at 37�C overnight. White bac-terial colonies were then picked and grown overnight in LBbroth containing ampicillin. Plasmid DNA was prepared fromthe overnight cultures, using the Promega Wizard plus SVminipreps purification system per the manufacturer�s instruc-tions, from 3 mL of culture medium.
Following plasmid preparation, 0.5 lg of plasmid DNAwas restricted with 20 U Sal-1 and 20 U Nco-1 in Promegabuffer D for 2 hours. The samples were then electrophoresed in1.5% agarose gel to confirm the presence of plasmid insert.Clones with inserts of the appropriate size were then grownovernight in LB broth containing ampicillin. Plasmid stocksderived from these overnight incubations were then autose-quenced using a Licor 4000L near-infrared fluorescence auto-sequencer (Li-Cor Biosciences, Nebraska, USA) as describedpreviously [9].
Statistical Analyses
Statistical analyses were performed using either parametric(analysis of variance, Student�s t-test) or nonparametric(Kruskal-Wallis, Mann-Whitney U, QQ plot) methods andlinear correlation, where appropriate.
Results
Localization of the ts-SV40-T Antigen
Figure 1a demonstrates the immunolocalization of thetemperature-sensitive form of the simian virus-derivedlarge tumor antigen, the expression of which induces thesustained replication of SV40-T-transduced mammaliancells. As expected, the T antigen in its active form islocalized to the nucleus of the cells.
Induction of the Osteoblast Phenotype of Human ImmortalizedOsteoprogenitor Cells
Under basal culture conditions, incubation at 33�C inmedium containing 10% HI-FBS, hOP 7 cells constitu-tively expressed low levels of AP activity (0.24 ± 0.1[mean ± standard error of the mean, SEM] lmol p-NP/mg protein/h) (Fig. 2a). However, AP activity increased(Fig. 2a) (0.99 ± 0.2 lmol p-NP/mg protein/h) signifi-cantly (P < 0.05) after raising the incubation tempera-ture to 39�C to inactivate the temperature-sensitiveimmortalizing oncogene ts-SV40-T (Fig. 1b). Further-more, addition of dexamethasone, a known driver ofosteoblastic differentiation, caused a highly significant(P < 0.01) further increase in AP activity (7.44 ± 2.0
lmol p-NP/mg protein/h) (Figs. 1c, 2a). The basal levelsof AP activity and the increased activity induced bydexamethasone were similar to those seen in primarycultures of human marrow stromal cells [46] (Fig. 2a).Incubation of cells in basal medium containing 1 mM b-glycerophosphate, 100 mg/mL ascorbate, and 10)7 Mdexamethasone for 28 days at 39�C induced elaborationand mineralization of extracellular matrix (Fig. 1d).
Expression of osteocalcin, a late marker for differ-entiated osteoblasts, was also determined in these stud-ies. Osteocalcin protein expression was undetectable inhOP 7 cells incubated at 39�C, in either the presence orabsence of dexamethasone, for up to 3 weeks. However,there was a highly significant, dose-dependent increasein osteocalcin expression when cells were incubated with1,25(OH)2D3 in the presence of vitamin K compared tocontrols (P < 0.01 for 10)11 M and P < 0.001 for 10)7
M 1,25[OH]2D3). Normalization of osteocalcin expres-sion into nanograms/milligram cell protein/48-hourperiod showed an increase from 50 to 533 ng/mg cellprotein/48 hours. As with AP activity, osteocalcin pro-tein expression induced by low levels of 1,25(OH)2D3
approximated the osteocalcin expression seen in humanmarrow stromal cells in primary culture, 50 ng osteo-calcin/mg cell protein/72 hours [46] (see Fig. 2b). Also,extracellular matrix elaboration and mineralization wasachieved after 28-day incubation in the presence of 50lg/mL ascorbate and 1 mM b-glycerophosphate (seeFig. 1d).
However, where HI-FBS was substituted by serumfrom postmenopausal women, no significant increase inAP activity was seen above baseline, even after incuba-tion of cells at 39�C. Furthermore, osteocalcin remainedundetectable, and neither matrix elaboration nor min-eralization was apparent.
Induction of the Adipocyte Phenotype of Human ImmortalizedOsteoprogenitor Cells
No lipid accumulation could be detected in hOP 7 cellscultured at 39�C for 7 days in basal medium containingHI-FBS (Fig. 3a), in either the presence or absence ofdexamethasone, as determined histologically by stainingwith oil red O. However, replacement of FBS with NRSresulted in a morphological change in cells from afibroblastic to a more rounded shape. Also, detectablelipid accumulation occurred within 3 days, with the vastmajority of hOP 7 cells staining positive for oil red O by7 days (see Fig 3b). Quantification of the adipogenicresponse (Fig. 3c) demonstrated a maximal response of90% ± 4% with 10% NRS and an almost half-maximalresponse of 49% ± 14% with 3% NRS. The adipocy-togenic effect of NRS on hOP 7 cells was unaffected byaddition of 10)7 M 17b-estradiol (Fig. 3d). Adipocyto-genic morphological changes were also seen when hOP 7cells were incubated with 100 lg/mL IBMX combined
236 B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation

with 10 lg/mL insulin and 10)7 M dexamethasone, al-though significant levels of cell death were apparentalong with a much reduced level of cell oil red O posi-tivity (12.0 ± 4%) compared to NRS treatment alone.
In addition, incubation of hOP 7 cells with NRSinduced LPL mRNA expression, an early marker ofadipocyte differentiation, after 7 days (Fig. 4a, lane 2).NRS also induced the expression of mRNA forthe late-stage adipogenic marker G3PDH following14-day incubation (Fig. 4b, lane 3). A loss of type1a(I) procollagen mRNA was concomitant with theexpression of these adipogenic markers (Fig. 4c, lane3). LPL, G3PDH, and type 1a(I) procollagen RT-PCR products were cloned and sequenced to confirmtheir expression.
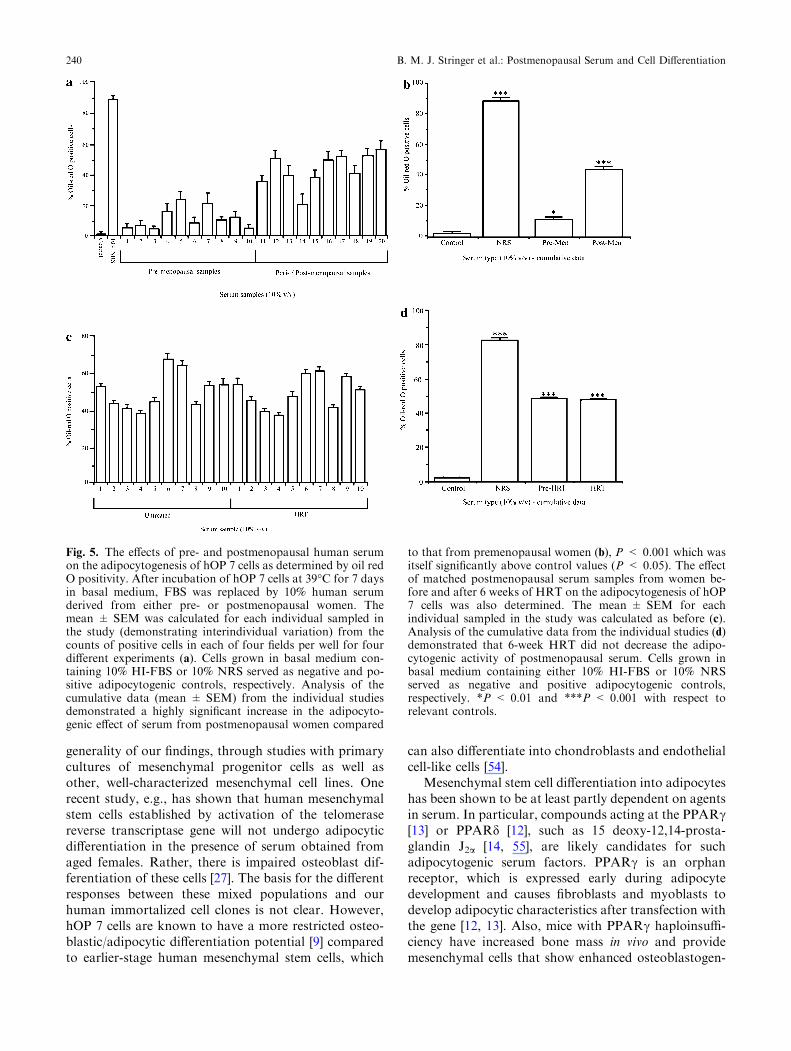
Premenopausal serum caused only a small increase inthe proportion of oil red O-positive cells (10.9%)(P < 0.05) with respect to cultures without humanserum. However, postmenopausal serum caused a highlysignificant increase in oil red O positivity compared topremenopausal serum (43.5%) (P < 0.001) with respectto cultures incubated with premenopausal serum(Fig. 5a, b). The total cell number was unaffected byincubation with either rabbit or human serum. Incuba-tion of cells with postmenopausal serum (final concen-tration 10% v/v) taken from the same patients following6 weeks of HRT also resulted in markedly enhanced oilred O staining to 42.8% of cells (Fig. 5c, d), there beingno significant difference in adipocytogenic activity ofpostmenopausal serum before or after HRT.
Fig. 1. ts-SV-40T immunofluorescence inthe human marrow stroma-derived clonalcell line hOP 7, immortalized with thetemperature-sensitive form of the simianvirus-40-derived large tumor antigen, inbasal medium at 33�C, the permissivetemperature of the oncogene (a). Incubationof the clone at 39�C, the oncogene�snonpermissive temperature, on its ownresulted in low AP activity (b), whereasaddition of 10)7 M dexamethasone inducedmuch greater AP activity (c, red cells) andelaboration and mineralization of anextracellular matrix (d, black stain). Scalebar = 25 lm.
B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation 237

The effects of time since menopause on adipocyto-genic activity of postmenopausal serum were analyzedby linear correlation. No significant correlation existed(0.05 < P < 0.711) within the sample tested.
In further experiments, the increase in AP activity(1.03 ± 0.03 lmol p-NP/mg protein/h) induced byincubation of hOP 7 cells for 7 days with 10% FBS atthe nonpermissive temperature of 39�C was reversed
Fig. 2. AP activity (lmol p-NP/mg protein/hour) of hOP 7cells was measured in response to 10)7 M dexamethasonetreatment. Cells were incubated in basal medium at 33�C or at39�C (respectively, the permissive and nonpermissive temper-atures of the oncogene) in either the presence or absence ofdexamethasone (a). A highly significant increase in AP activitywas seen in response to addition of dexamethasone. Further-
more, osteocalcin expression (b) also exhibited a highly sig-nificant, concentration-dependent increase in response toaddition of 1,25(OH)2D3 in the presence of vitamin K at 39�C.Error bars represent the SEM of six samples per point.*P < 0.05, **P < 0.01, and ***P < 0.001 with respect torelevant controls.
Fig. 3. Effects of NRS onhOP 7 cells. hOP 7 cells wereincubated for 7 days at 39�Cin basal medium containing10% HI-FBS (a) and then inmodified basal medium wherethe FBS was replaced with10% NRS (b), followed byfixation and oil red O staining.Most cells became positiveafter addition of NRS. Scalebar = 25 lm. The response ofhOP 7 cells first showed asignificant increase in oil redO positivity andadipocytogenesis in thepresence of 1% NRS (c), witha half-maximal effect atapproximately 3% NRS. Theadipocytogenic effect of NRSwas not reversed byconcurrent incubation with10)7 M 17b-estradiol (d).**P < 0.01 and***P < 0.001 with respect torelevant controls.
238 B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation

(0.38 ± 0.1 lmol p-NP/mg protein/h, P < 0.001) bysubstitution of FBS with NRS for 7 days. However, ifhOP 7 cells were first incubated with 10% FBS and 10)7
M dexamethasone for 14 days, to induce more fully theirosteoblastic phenotype, the hOP 7 cell capacity forNRS-induced adipocyte differentiation was removedirreversibly (1.83 ± 0.62% oil red O positivity).
Discussion
The demonstration of human bipotential adipocytic/osteoblast progenitor cells in marrow is now welldocumented, as is the ability of rabbit serum to directtheir differentiation to the adipocyte phenotype [9�11,26]. The hOP 7 clonal cell is such a human bipotentialadipocytic/osteoblastic progenitor and remains undif-ferentiated in basal medium [9]. Addition of dexa-methasone and/or vitamin D3 causes this progenitor toexpress late osteoblast markers, whereas incubationwith rabbit serum or adipogenic agents such as IBMXcombined with insulin and dexamethasone elicitsexpression of adipocyte characteristics. Interestingly,maintenance of hOP 7 cells for 14 days in culturemedia that induce full osteoblastic differentiation,including extracellular matrix elaboration and miner-alization, removes their ability to become adipocytes.We have yet to identify, though, any effect of post-menopausal serum on restricting the hOP 7 cell po-tential for osteoblastic differentiation, once drivendown an adipocytic route.
Rabbit serum-induced adipocytic differentiation isnot a consequential feature inherent to cell lines createdby a cell immortalization process. We have immortalizedtrabecular bone-derived osteoblastic, articular chondr-ocytic, hypertrophic chondrocytic, and neural cells byretroviral temperature-sensitive oncogene transduction[47�51]; but none of these has the capacity to differen-tiate into adipocytes in the presence of rabbit serum.
In the present study, serum from postmenopausalwomen induced hOP 7 cells to follow an adipocyticdifferentiation route, away from the osteoblastic lineage.This suggests there exists in human serum one or morefactors which can shift the balance between humanmarrow stromal progenitor cells becoming osteoblastsor developing instead into adipocytes.
There is a striking similarity between the presentin vitro observations and the reciprocal changes in theabundance of bone marrow adipocytes and osteoblastsoccurring in postmenopausal women in vivo. Theimportance of this phenomenon is underlined by thesmaller but significant effect of premenopausal serum onthe fate of the adipocyte/osteoblast progenitor cell. Boneturnover is known to increase with estrogen deficiency(due to an increase in all elements of bone remodeling[52]), but trabecular bone formation during eachremodeling cycle does not match the volume of boneresorbed. The reduction in bone formation may resultfrom a proportionately smaller increase in the numberof osteoblasts. However, if so, this is not due to a loss incapacity for the marrow cells to differentiate into os-teoblasts with age as the osteoblastic/adipocytic differ-entiation potential of stromal cells is maintained withage and/or the onset of osteoporosis [53].
The age at which there is an increase in the adipo-genic activity of female serum is not clear from ourstudy. The lack of correlation between adipogenicactivity and time since menopause suggests strongly thatthe event is largely complete by menopause. However,the low sample numbers and the rather narrow spread ofages in the premenopausal group in this early reportprevent us from delineating the timing more accuratelythan somewhere between the mid-twenties and peri-menopausal age. A larger study, comprising womensampled preferably from all age groups, is clearly re-quired to resolve this issue.
Moreover, to evaluate further the significance of thiswork, additional research is needed to determine the
Fig. 4. Effects of NRS on the expression of adipocytic andosteoblastic cell markers in hOP 7 cells. hOP 7 cells of 55-60population doublings were grown to confluence at 33�C inbasal medium containing 10% HI-FBS. FBS was then re-placed with 10% NRS, and cells were incubated at 39�C forup to 14 days. NRS induced the early adipocytic marker LPL(a, 324 bp band, lane 2) after 7 days. LPL is not demon-strable in the absence of NRS (lane 3). The late marker ofadipocytogenesis G3PDH was induced by 14-day NRStreatment (b, 568 bp band, lane 3) but could not be dem-onstrated when cells were incubated for 14 days in FBS in-
stead (lane 2). Furthermore, type a1(I) procollagen mRNAexpression (c, 235 bp band, lane 2), an early marker of cellsof the osteoblastic lineage, is lost following incubation withNRS for 7 days. A 100 bp ladder is used as a molecularweight size marker (a-c, lane 1). For loading control,expression of the housekeeping gene b2-microglobulin (d, 237bp band, lanes 2�4) demonstrates equal loading of the RT-PCR products derived from hOP 7 cells without (lane 2) andwith NRS treatment for 7 (lane 3) and 14 (lane 4) days. Allproduct identities were confirmed by subsequent cloning andsequencing of RT-PCR products.
B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation 239
generality of our findings, through studies with primarycultures of mesenchymal progenitor cells as well asother, well-characterized mesenchymal cell lines. Onerecent study, e.g., has shown that human mesenchymalstem cells established by activation of the telomerasereverse transcriptase gene will not undergo adipocyticdifferentiation in the presence of serum obtained fromaged females. Rather, there is impaired osteoblast dif-ferentiation of these cells [27]. The basis for the differentresponses between these mixed populations and ourhuman immortalized cell clones is not clear. However,hOP 7 cells are known to have a more restricted osteo-blastic/adipocytic differentiation potential [9] comparedto earlier-stage human mesenchymal stem cells, which
can also differentiate into chondroblasts and endothelialcell-like cells [54].
Mesenchymal stem cell differentiation into adipocyteshas been shown to be at least partly dependent on agentsin serum. In particular, compounds acting at the PPARc[13] or PPARd [12], such as 15 deoxy-12,14-prosta-glandin J2a [14, 55], are likely candidates for suchadipocytogenic serum factors. PPARc is an orphanreceptor, which is expressed early during adipocytedevelopment and causes fibroblasts and myoblasts todevelop adipocytic characteristics after transfection withthe gene [12, 13]. Also, mice with PPARc haploinsuffi-ciency have increased bone mass in vivo and providemesenchymal cells that show enhanced osteoblastogen-
Fig. 5. The effects of pre- and postmenopausal human serumon the adipocytogenesis of hOP 7 cells as determined by oil redO positivity. After incubation of hOP 7 cells at 39�C for 7 daysin basal medium, FBS was replaced by 10% human serumderived from either pre- or postmenopausal women. Themean ± SEM was calculated for each individual sampled inthe study (demonstrating interindividual variation) from thecounts of positive cells in each of four fields per well for fourdifferent experiments (a). Cells grown in basal medium con-taining 10% HI-FBS or 10% NRS served as negative and po-sitive adipocytogenic controls, respectively. Analysis of thecumulative data (mean ± SEM) from the individual studiesdemonstrated a highly significant increase in the adipocyto-genic effect of serum from postmenopausal women compared
to that from premenopausal women (b), P < 0.001 which wasitself significantly above control values (P < 0.05). The effectof matched postmenopausal serum samples from women be-fore and after 6 weeks of HRT on the adipocytogenesis of hOP7 cells was also determined. The mean ± SEM for eachindividual sampled in the study was calculated as before (c).Analysis of the cumulative data from the individual studies (d)demonstrated that 6-week HRT did not decrease the adipo-cytogenic activity of postmenopausal serum. Cells grown inbasal medium containing either 10% HI-FBS or 10% NRSserved as negative and positive adipocytogenic controls,respectively. *P < 0.01 and ***P < 0.001 with respect torelevant controls.
240 B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation

esis in vitro [20]. Furthermore, PPARc-deficientembryonic stem cells fail to differentiate into adipocytes[20]. It is clear, therefore, that the regulation of adipo-cyte differentiation from mesenchymal progenitor cells iscomplex and may involve a multiplicity of factors inhuman serum, with PPARc playing a central role in theprocess (for review, see [24]). Accordingly, to providesome insight into the possible mechanisms by which thedifferentiation of hOP 7 cells may be modulated byhuman serum factors, we looked for expression ofPPARc2, the adipocyte-specific isoform of PPARc, andthe ability of its natural ligands and associated ligandprecursors and metabolites to effect an adipocytic dif-ferentiation response in hOP 7 cells. Our studies showedexpression of PPARc2 in hOP 7 cells in basal medium,and this appeared to be enhanced during the inductionof adipocyte differentiation. Furthermore, incubationwith either the natural ligand of PPARc (15-deoxy-12,14-prostaglandin J2, prostaglandin J2, or prosta-glandin D2) induced an adipocytogenic differentiationresponse (Fig. 6), consistent with adipocyte differentia-tion being mediated by activation of PPARc2.
Several additional ligands for PPAR have also beenidentified [14, 55�57], including fatty acids and theirmetabolites [58]; and some of these are particularlyabundant in rabbit serum [26]. Interestingly, an imbal-ance between n-6 and n-3 serum fatty acids has beenlinked to the alveolar bone destruction seen in peri-odontal disease [59]. Further, both animal [60�63] and
human [64�66] studies have shown that administeringnonsteroidal anti-inflammatory agents, inhibitors ofarachidonic acid metabolism, will block periodontalbone loss. Therefore, fatty acids and/or their metabolitesmay well participate in the hOP 7 adipocytogenic re-sponse to incubation with postmenopausal serum.
Bone loss accelerates in women following menopause,but HRT in the form of 17b-estradiol is able to slow themenopause-induced decline in skeletal bone mass.Estrogen�s mechanism of action is thought to be throughinhibition of osteoclastic bone resorption and addi-tionally by reducing the frequency of bone remodelingcycles. To determine whether estrogen may also affectthe serum-induced adipocytogenic differentiation ofhuman osteoprogenitors, we incubated hOP 7 cells withrabbit serum plus 17b-estradiol and with postmeno-pausal serum from patients following 6 weeks of 17b-estradiol HRT. However, neither addition of 17b-estradiol to rabbit serum nor 6 weeks of HRT was ableto attenuate the adipocytogenic effect of these sera onhOP 7 cells.
In summary, the study demonstrates that factors inhuman serum, and postmenopausal human serum inparticular, are capable of inducing the differentiation ofthe human clonal adipocytic/osteoblastic cell line hOP 7away from an osteoblast phenotype to one of adipo-cytes. Further studies on primary bone marrow stromalcells and additional stromal cell lines are needed todemonstrate the generality of this serum effect and to
Fig. 6. Effects of 10% NRS on expression of PPARc2, theadipocyte-specific isoform of PPARc (a), and of various pro-staglandins on the adipocytogenesis of hOP 7 cells (b). Underbasal culture conditions (39�C in 10% FBS), PPARc2 mRNAis expressed by hOP 7 cells in their precursor state (lane 2).Following incubation with NRS for 7 days, expression ofPPARc2 mRNA appears to be enhanced (lane 3). Both RT-PCR products were the anticipated size of 310 bp followingamplification. Restriction of the 310 bp amplification product
with BglI produced two fragments of 110 and 200 bp in length,confirming the PPARc2 sequence. The molecular weight sizemarker is HaeIII restricted /X174 (lane 1). Putative prosta-glandin ligands for PPARc were analyzed for their adipocy-togenic activity on hOP 7 cells. Prostaglandin J2 caused ahighly significant increase in adipocytogenesis at concentra-tions as low as 10)7 M, while prostaglandin D2 and 15deoxy-prostaglandin J2 produced a less marked but nonethelesshighly significant response.
B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation 241

justify investigation into the potential regulatory mech-anisms involved. From a clinical viewpoint, the possi-bility of preventing/inhibiting the effects of adipogenicagents in postmenopausal serum may provide a newroute to enhancing bone formation in the treatment ofosteoporosis.
Acknowledgment. We are grateful to the Arthritis ResearchCampaign, the Nuffield Foundation, the University of Shef-field, and Cardiff University for supporting this work.
References
1. Meunier PJ, Aaron J, Edouard C, Vignon G (1971)Osteoporosis and the replacement of cell populations ofthe marrow by adipose tissue: a quantitative study of 84iliac bone biopsies. Clin Orthop Relat Res 80:147�154
2. Burkhardt R, Kettner G, BohmW, Schmidmeir M, SchlagR, Frisch B, MaIlman B, Eisenmenger W, Gilg TH (1987)Changes in trabecular bone, haematopoiesis and bonemarrow vessels in aplastic anaemia, primary osteoporosis,and old age: a comparative histomorphometric study.Bone 8:157�164
3. Parfitt AM (1990) Bone forming cells in clinical conditions.In: Hall BK (ed), Bone, A Treatise. The Osteoblast andOsteocyte, vol 1. Telford Press, Caldwell, NJ, pp 351�429
4. Justesen J, Stenderup K, Ebbeson EN, Mosekilde L,Steiniche T, Kassem M (2001) Adipocyte tissue volume inbone marrow is increased with aging and with osteopo-rosis. Biogerontology 2:165�171
5. Verma S, Rajaratnam JH, Denton J, Hoyland JA, ByersRJ (2002) Adipocytic proportion of bone marrow is in-versely related to bone formation in osteoporosis. J ClinPathol 55:693�698
6. Wronski TJ, Walsh CC, Ignaszewski LA (1986) Histo-logical evidence for osteopenia and increased bone turn-over in ovariectomized rats. Bone 7:119�123
7. Miniare E, Meunier PJ, Edouard C, Bernard J, CourpronJ, Bourret J (1974) Quantitative histological data on disuseosteoporosis. Calcif Tissue Res 17:57�73
8. Aubin JE, Turksen K, Heersche JNM (1993) Osteoblasticcell lineage. In: Noda M (ed), Cellular and MolecularBiology of Bone. Academic Press, London, pp 1�45
9. Houghton A, Oyajobi BO, Foster GA, Russell RGG,Stringer BMJ (1998) Immortalization of human bonemarrow stromal cells by retroviral transduction with atemperature sensitive oncogene: identification of bipoten-tial precursor cells capable of directed differentiation toeither an osteoblast or adipocyte phenotype. Bone22:7�16
10. Nuttall ME, Patton AJ, Olivera DL, Nadeau DP, GowenM (1998) Human trabecular bone cells are able to expressboth osteoblastic and adipocytic phenotype: implicationsfor osteopenic disorders. J Bone Miner Res 13:371�382
11. Hicok KC, Thomas T, Gori F, Rickard DJ, Spelsberg TC,Riggs BL (1998) Development and characterisation ofconditionally immortalised osteoblast precursor cell linesfrom human bone marrow stroma. J Bone Miner Res13:205�217
12. Tontonoz P, Hu E, Spiegelman BM (1994) Stimulation ofadipogenesis in fibroblasts by PPARc2, a lipid-activatedtranscription factor. Cell 79:1147�1156
13. Hu E, Tontonoz P, Spiegeleman BM (1995) Transdiffer-entiation of myoblasts by the adipogenic transcriptionfactors PPARc and C/EBPa. Proc Natl Acad Sci USA92:9856�9860
14. Forman BM, Tontonoz P, Chen J, Brun RP, SpiegelmanBM, Evans RM (1995) 15 Deoxy-12,14-prostaglandin J2 is aligand for the adipocyte determination factor PPARc. Cell83:803�812
15. Ducy P, Amling M, Takeda S, Priemel M, Schilling AF,Beil FT, Shen J, Vinson C, Reuger JM, Karsenty G(2000) Leptin inhibits bone formation through a hypo-thalamic relay: a central control of bone mass. Cell100:197�207
16. Jilka LR, Weinstein RS, Takahashi K, Parfitt AM, Ma-nolagas SC (1996) Linkage of decreased bone mass withimpaired osteoblastogenesis in a murine model of accel-erated senescence. J Clin Invest 97:1732�1740
17. Sims NA, Sbatakos G, Chen J-S, Kelz M, Amling M,Nestler E, Baron R (1999) Progressive and pronouncedincrease in bone mass, osteosclerosis and reduced adipo-cytes in mice overexpressing delta-fosB, a truncated fromof fosB. J Bone Miner Res 14(suppl 1):172
18. Kveiborg M, Sabatakos G, Chiusaroli R, WuM, PhilbrickHorne WM WC, Baron R (2004) DeltaFosB induces os-teosclerosis and decreases adipogenesis by two indepen-dent cell-autonomous mechanisms. Mol Cell Biol24:2820�2830
19. Kodama Y, Takeuchi Y, Suzawa M, Fukumoto S, Mu-rayama H, Yamato H, Fujita T, Kurokawa T, MatsumotoT (1998) Reduced expression of interleukin-11 in bonemarrow stromal cells of senescence-accelerated mice(SAMP6): relationship to osteopenia with enhanced adi-pogenesis. J Bone Miner Res 13:1370�1377
20. Akune T, Ohba S, Kamekura S, Yamaguchi M, ChungUI, Kubota N, Terauchi Y, Harada Y, Azuma Y, Na-kamura K, Kadowaki T, Kawaguchi H (2004) PPAR-gamma insufficiency enhances osteogenesis throughosteoblast formation from bone marrow progenitors. JClin Invest 113:846�855
21. Nuttall ME, Gimble JM (2000) Is there a therapeuticopportunity to either prevent or treat osteopenic disordersby inhibiting marrow adipogenesis. Bone 27:177�184
22. Gimble JM, Nuttall ME (2004) Bone and fat: old ques-tions, new insights. Endocrine 23:183�188
23. Nuttall ME, Gimble JM (2004) Controlling the balancebetween osteoblastogenesis and adipogenesis and theconsequent therapeutic implications. Curr Opin Pharma-col 4:290�294
24. Gimble JM, Zvonic S, Floyd ZE, Kassem M, Nuttall ME(2006) Playing with bone and fat. J Cell Biochem98:251�266
25. Dorheim M-A, Sullivan M, Dandapani V, Wu X, HudsonJ, Segerini PR, Rosen DM, Aulthouse AL, Gimble JM(1993) Osteoblastic gene expression during adipogenesis inhematopoietic supporting murine bone marrow stromalcells. J Cell Physiol 154:317�328
26. Diascro DD Jr, Vogel RL, Johnson TE, Witherup KM,Pitzenberger SM, Rutledge SJ, Prescott DJ, Rodan GA,Schmidt A (1998) High fatty acid content in rabbit serumis responsible for the differentiation of osteoblasts intoadipocyte-like cells. J Bone Miner Res 13:96�106
27. Abdallah B, Haack-Sorensen M, Fink T, Kassem M(2006) Inhibition of osteoblast differentiation but notadipocyte differentiation of mesenchymal stem cells bysera obtained from aged females. Bone 39:181�188
28. Ettinger B, Genant HK, Steiger P, Madvig P (1992) Low-dosage 17b-estradiol prevents bone loss in postmeno-pausal women. Am J Obstet Gynecol 66:479�488
29. Lindsay R, Hart DM, Clark DM (1984) The minimumeffective dose of estrogen for prevention of postmeno-pausal bone loss. Obstet Gynecol 63:759�763
30. Keil DP, Felson DT, Anderson JJ, Wilson PWF, Mo-skowitz MA (1987) Hip fracture and the use of estrogensin postmenopausal women: the Framingham Study. NEngl J Med 317:1169�1174
31. Weiss NS, Ure CL, Ballard JH, Williams AR, Daling JR(1980) Decreased risk of fractures of the hip and lowerforearm with postmenopausal use of estrogen. N Engl JMed 303:1195�1198
32. Weigratz I, Fink T, Rohr UD, Lang E, Leukel P, Kuhl H(2001) Cross-over comparison of the pharmacokinetics ofestradiol during hormone replacement therapy with
242 B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation

estradiol valerate or micronized estradiol. Zentralbl Gy-nakol 123:505�512
33. Jones KP (1992) Estrogens and progestins: what to useand how to use it. Clin Obstet Gynecol 35:871�873
34. Scott RT, Ross B, Anderson C, Archer DF (1991) Phar-macokinetics of percutaneous estradiol: a crossover studyusing a gel and a transdermal system in comparison withoral micronized estradiol. Obstet Gynecol 77:758�764
35. Nichols KC, Schenkel L, Benson H (1984) 17Beta-estra-diol for postmenopausal estrogen replacement therapy.Obstet Gynecol Surv 39:230�245
36. Consensus Development Conference (1987) Prophylaxisand treatment of osteoporosis. Br Med J (Clin Res Ed)295:914�915
37. Weibel ER (1979) Stereological Methods. PracticalMethods for Biological Morphometry, vol 1. AcademicPress, London
38. Weibel ER, Gonzague S, Kistler F, Scherle WF (1966)Practical stereological methods for morphometric cytol-ogy. J Cell Biol 30:23�38
39. Stringer BM, Wynford-Thomas D, Williams ED (1982)Physical randomisation of tissue architecture � an alter-native to systematic sampling. J Microsc 126:179�182
40. Bessey OA, Lowry OH, Brock MF (1946) A method forthe rapid determination of alkaline phosphatase with fivecubic millilitres of serum. J Biol Chem 164:321�329
41. Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilisingthe principle of protein-dye binding. Anal Biochem72:248�254
42. Cook HC (1974) Manual of Histological DemonstrationTechniques. Butterworths, London
43. Mackenzie CG, Mackenzie JB, Reiss OK, Wisneski JA(1970) Identification of albumin-bound fatty acids as themajor factor in serum-induced lipid accumulation in cul-tured cells. J Lipid Res 11:571�582
44. Sakaue H, Ogawa W, Nakamura T, Mori T, NakamuraK, Kasuga M (2004) Role of MAPK phosphatase-1(MKP-1) in adipocyte differentiation. J Biol Chem279:39951�39957
45. Horton RM, Hoppe BL, Conto Tronconi BM (1994)Ampligrease: Hot Start PCR using petroleum jelly. Bio-techniques 16:42�43
46. Cheng S, Yang JW, Rifas L, Zhang S, Avioli LV (1994)Differentiation of human marrow osteogenic cells in vitro:induction of osteoblast penotype by dexamethasone.Endocrinology 134:277�286
47. Stringer BMJ, Verhofstad AAJ, Foster GA (1994) Rapheneural cells immortalised with a temperature sensitiveoncogene: differentiation by default down an APUD celllineage. Dev Brain Res 79:267�274
48. Oyajobi BO, Houghton A, Hatton P, Frazer A, GraveleyR, Russell RGG, Stringer BMJ (1998) Expression of typeX collagen and matrix calcification in three-dimensionalcultures of immortalised temperature-sensitive chondro-cytes derived from adult human. J Bone Miner Res13:432�442
49. Stringer BMJ, Phillips I, Gangemi L, Gosal M, Foster GA(2002) Generation of hypertrophic cartilage for bone tis-sue engineering. Eur Cells Materials 4:5�6
50. Foster GA, Stringer BMJ (2004) Cell-based therapies forParkinson�s disease. Drug Discov Today Ther Strat1:43�49
51. Stringer BMJ, Waddington R, Sloan A, Phillips I, TelfordG, Hughes D, Craig G, Gangemi L, Brook I, Freeman C,Cao X, Gosal M, Smith S, Russel RGG, Foster GA
(2006) Bespoke human hypertrophic chondrocytic celllines provide the osteoinductive signals required for vas-cularised bone formation. Tissue Eng 13:133�145
52. Kanis J (1996) Estrogens, the menopause, and osteopo-rosis. Bone 19:185S�190S
53. Justesen J, Stenderup K, Eriksen EF, Kassem M (2002)Maintenance of osteoblastic and adipocytic differentiationpotential with age and osteoporosis in human marrowstromal cell cultures. Calcif Tissue Int 71:36�44
54. Abdallah BM, Haack-Sørensena M, Burnsa JS, ElsnabaB, Jakob F, Hoklandc P, Kassem M (2005) Maintenanceof differentiation potential of human bone marrow mes-enchymal stem cells immortalized by human telomerasereverse transcriptase gene in despite of extensive prolifer-ation. Biochem Biophys Res Commun 326:527�538
55. Kliewer SA, Lenhard JM, Willson TM, Patel I, MorrisDC, Lehmann JM (1995) A prostaglandin J2 metabolitebinds peroxisome proliferator-activated receptor c andpromotes adipocyte differentiation. Cell 83:813�819
56. Keller H, Dreyer C, Medin J, Mahfoudi A, Ozato K,Wahli W (1993) Fatty acids and retinoids control lipidmetabolism through activation of peroxisome proliferator-activated receptor-retinoid X receptor heterodimers. ProcNatl Acad Sci USA 90:2160�2164
57. Gulick T, Cresci S, Caira T, Moore DD, Kelly DP (1994)The peroxisome proliferator-activated receptor regulatesmitochondrial fatty acid oxidative enzyme gene expres-sion. Proc Natl Acad Sci USA 91:11012�11016
58. Schopfer FJ, Lin Y, Baker PR, Cui T, Garcia-Barrio M,Zhang J, Chen K, Chen YE, Freeman BA (2005) Nitro-linoleic acid: an endogenous peroxisome proliferator-activated receptor gamma ligand. Proc Natl Acad Sci USA102:2340�2345
59. Requirand P, Gibert P, Tramini P, Cristol JP, Descomps B(2000) Serum fatty acid imbalance in bone loss: examplewith periodontal disease. Clin Nutr 19:271�276
60. Weaks-Dybvig M, Sanavi F, Zender H, Rifkin BR (1982)The effect of indomethacin on alveolar bone loss inexperimental periodontitis. J Periodont Res 17:90�100
61. Jeffcoat MK, Williams RC, Wechter WJ, Johnson HG,Kaplan ML, Gandrup JS, Goldhaber P (1986) Flurbi-profen treatment of periodontal disease in beagles. JPeriodont Res 21:624�633
62. Kornman KS, Blodgett RF, Brunsvold M, Holt SC (1990)Effects of topical applications of meclofenamic acid andibuprofen on bone loss, subgingival microbiota and gin-gival PMN response in the Macaca fascicularis. J Peri-odont Res 25:300�307
63. Li KL, Vogel R, Jeffcoat MK, Alfano MC, Smith MA,Collins JG, Offenbacher S (1996) The effect of ketoprofencreams on periodontal disease in rhesus monkeys. J Peri-odont Res 31:525�532
64. Waite JM, Saxton CA, Young A, Wagg BJ, Corbett M(1981) The periodontal status of subjects receivingnon-steroidal antiinflammatory drugs. J Periodont Res16:100�108
65. Jeffcoat MK, Williams RC, Ready English MS R,Goldhaber P (1988) Flurbiprofen treatment of humanperiodontitis effect on alveolar bone height and metabo-lism. J Periodont Res 23:381�385
66. Jeffcoat MK, Williams RC, Reddy M, Wannawisute A,Waite P, Palcanis K, Cogen R, Williams RC, Basch C(1991) Use of digital radiography to demonstrate thepotential of naproxen as an adjunct in the treatmentof rapidly progressive periodontitis. J Periodont Res26:415�421
B. M. J. Stringer et al.: Postmenopausal Serum and Cell Differentiation 243
Copyright © 2022 FDOKUMEN




















