
PEPTIDE PATTERN OF AMNIOTIC FLUID AND ITS ... - LSMU
122
1 LITHUANIAN UNIVERSITY OF HEALTH SCIENCES MEDICAL ACADEMY Egl Machtejevien PEPTIDE PATTERN OF AMNIOTIC FLUID AND ITS CORRELATION WITH PROTEIN COMPOSITION OF FETAL MEMBRANES: THE SEARCH FOR NEW POTENTIAL BIOMARKERS TO PREDICT PRETERM PREMATURE RUPTURE OF MEMBRANES Doctoral dissertation Biomedical Sciences, Medicine (06B) Kaunas, 2013
-
Upload
khangminh22 -
Category
Documents
-
view
7 -
download
0
Transcript of PEPTIDE PATTERN OF AMNIOTIC FLUID AND ITS ... - LSMU

1
LITHUANIAN UNIVERSITY OF HEALTH SCIENCES MEDICAL ACADEMY
Egl! Machtejevien!
PEPTIDE PATTERN OF AMNIOTIC
FLUID AND ITS CORRELATION
WITH PROTEIN COMPOSITION
OF FETAL MEMBRANES:
THE SEARCH FOR NEW POTENTIAL
BIOMARKERS TO PREDICT
PRETERM PREMATURE RUPTURE
OF MEMBRANES
Doctoral dissertation Biomedical Sciences,
Medicine (06B)
Kaunas, 2013

2
This doctoral dissertation was carried out at the Lithuanian University of Health Sciences in 2008–2012. Scientific Supervisor
!"#$%& '"%& ()*+& ,#-+.*+& /+012+34516.7& 891*:3+.1+.&;.1<6"41*=& #$& >6+-*:&
Sciences, Biomedical Sciences, Medicine – 06B)

3
TABLE OF CONTENTS
ABBREVIATIONS ....................................................................................... 5
1. INTRODUCTION ................................................................................... 7
2. AIM AND TASKS OF THE STUDY ...................................................... 9 2.1. Aim of the study ........................................................................... 9 2.2. Tasks of the study ......................................................................... 9
3. NOVELTY OF THE RESEARCH ........................................................... 9
4. REVIEW OF THE LITERATURE ......................................................... 10 4.1. “Omics” technologies ...................................................................... 10
4.1.1. Human genome ........................................................................ 10 4.1.2. Transcriptome .......................................................................... 11 4.1.3. Proteome ................................................................................... 12
4.2. Fetal membranes ............................................................................. 20 4.2.1. The amniochorionic extracellular matrix proteins ................... 20 4.2.2. Matrix metalloproteinases ....................................................... 21 4.2.3. Tissue inhibitors of metalloproteinases ................................... 22
4.3. Physiology of amniotic fluid ........................................................... 22 4.3.1. Composition of amniotic fluid ................................................. 22 4.3.2. Regulation of amniotic fluid volume ....................................... 23
4.4. Amniotic fluid proteome ................................................................. 24 4.4.1. The normal human amniotic fluid proteome ........................... 24 4.4.2. Gestational age-dependent changes ......................................... 26 4.4.3. Clinical applications of amniotic fluid proteomics .................. 28
4.5. Human plasma proteome ................................................................. 33 4.6. Methods of proteomic analysis of amniotic fluid ............................ 34
4.6.1. Gel electrophoresis .................................................................. 35 4.6.2. Protein arrays ........................................................................... 35 4.6.3. Liquid chromatography ........................................................... 37
4.7. Preterm premature rupture of the membranes ................................. 46 4.7.1. Definitions and incidence ...................................................... 46 4.7.2. Risk factors ............................................................................ 47 4.7.3. Etiology and pathophysiology ............................................... 50 4.7.4. Diagnosis ............................................................................... 53 4.7.5. Prediction of preterm premature rupture of membranes ....... 56
5. PATIENTS, BIOLOGICAL SAMPLES AND METHODOLOGY ....... 58 5.1. Patients ............................................................................................ 58

4
5.2. Biological samples ........................................................................... 60 5.3. The analytical system ...................................................................... 62 5.4. Database searching and confidence of proteins identification ........ 65 5.5. Ethical aspects ................................................................................. 66
6. RESULTS AND DISCUSSION ............................................................. 67 6.1. Peptides and proteins identified in amniotic fluid ........................... 70 6.2. Characterization of amniotic fluid proteome ................................... 80 6.3. Comparison of the amniotic fluid proteome with previous publications ............................................................................................. 82 6.4. Gestational age-dependent changes in the amniotic fluid proteome ................................................................................................. 84 6.5. Fetal membranes proteome .............................................................. 88
6.5.1. Comparison of proteome from fetal membranes of patients with PPROM and term labor with intact membranes ........................ 89 6.5.2. Comparison of proteome from the fetal membranes and amniotic fluid of patients with PPROM and term labor with intact membranes ......................................................................................... 97
6.6. Comparison of biological samples peptidome and proteome from patients with preterm premature rupture of membranes ................ 98
7. CONCLUSIONS ................................................................................... 101
8. BENEFITS OF THE RESEARCH AND POTENTIAL FOR THE FUTURE .................................................................................. 102
9. LIST OF ORIGINAL PUBLICATIONS .............................................. 104
10. REFERENCES .................................................................................... 106
ACKNOWLEGMENTS ........................................................................... 120

5
ABBREVIATIONS 2DE two dimensional electrophoresis 2D LC/MS two dimensional liquid chromatography mass spectrometry 2D PAGE two dimensional polyacrylamide gel electrophoresis AC amniocentesis AF amniotic fluid AFI amniotic fluid index AFP alpha-fetoprotein CVF cervico-vaginal fluid Da dalton (the unified atomic mass unit) DNA deoxyribonucleic acid ECM extracellular matrix ELISA enzyme-linked immuno sorbent assay ESI electrospray ionization fFN fetal fibronectin FN false negative FP false positive G group group of amniotic fluid samples from patients at 16–18 week of gestation with intact membranes (obtained during prenatal genetic amniocentesis) HLUHS Hospital of the Lithuanian University of Health Sciences HPLC high performance liquid chromatography HPPP Human Plasma Proteome Project HUPO Human Proteome Organization IAI intraamniotic infection IEC ion exchange chromatography IUI intrauterine infection Ig immunogloblin IGFBP-1 insulin-like growth factor-binding protein 1 IL interleukins LC liquid chromatography MALDI-TOF matrix assisted laser desorption/ionization time-of-flight MD multidimensional MDG Millennium development goal MMP matrix metalloproteinases MR mass restricted MS mass spectrometry m/z mass-to-charge ratio

6
PCR polymerase chain reaction pI isoelectric point (the pH at which a protein carries no net electrical charge or net charge is zero) ppm parts per million PPROM preterm premature rupture of membranes PROM premature rupture of membranes PTD preterm delivery PTL preterm labor Q-TOF quadrupole time-of-flight R group group of amniotic fluid samples from patients with preterm premature rupture of membranes RAM restricted access material RNR ribonucleic acid RP reverse phase SD standard deviation SELDI-TOF surface-enhanced laser desorption/ionization time-of-flight SCX strong cation exchange SPE solid phase extraction TIMP tissue inhibitor of metalloproteinase Z group group of amniotic fluid samples from patients with term pregnancies and intact membranes (obtained during cesarean section)

7
1. INTRODUCTION
The Millennium Development Goals (MDGs) were set in 2000 by the
United Nations. MDG-4 targets a two-thirds reduction of under 5 years children deaths between 1990 and 2015 [1]. While under age of 5 mortality rates are improving in many countries worldwide, neonatal mortality rates (deaths in the first 28 days of life) have shown much less progress. Neonatal deaths now account for more than 42% of under-5 deaths [2,3]. Com-plications of preterm birth are the leading direct cause of neonatal mortality, accounting for an estimated 27% of the almost four million neonatal deaths every year, and act as a risk factor for many neonatal deaths due to multiple causes, particularly infections [3]. Therefore, achievement of MDG-4 is strongly influenced by progress in reducing neonatal deaths. Since preterm birth is the leading cause of these deaths, therefore progress is dependent on achieving of evidence-based interventions to prevent preterm delivery; also to improve survival for preterm newborns.
Preterm premature rupture of membranes (abbreviated as PPROM) is defined as the rupture of fetal membranes before the onset of labor before 37 weeks of gestation. PPROM occurs in 2% to 5% of pregnancies and is the most common cause of preterm birth, present in 25% to 40% of cases, and can result in significant neonatal morbidity and mortality [4,5,6]. According to the Lithuanian Medical Data of Births 1616 (5.6%) newborns were born prematurely in Lithuania in 2011 [7].
Epidemiological and clinical studies have identified a number of factors associated with increased risk for PPROM; however, the current methods of predicting women are non-specific to be of real clinical value. It is likely that biological markers exists that could be of clinical value as the evident increase that many of the events leading to PPROM are more likely to be chronic than acute. Therefore, there is a need to develop and to apply highly advanced methodologies to search and identify these biomarkers. Once the biomarkers are confirmed, efficient and selective biological tests could be developed which would help to achieve accurate prognosis, diagnosis and maybe medicine to prevent that process.
The etiology of PPROM is thought to be multifactorial and the actual mechanisms involved are still unknown. Intrauterine infections caused by bacteria are considered to be a predominant cause/risk factor. It has been estimated that about 40% of all PPROM occur in mothers with intrauterine infection. Once the pathogen reaches the uterine cavity through an ascending route, the fetal membrane is the last barrier for pathogens to break

8
before accessing the amniotic cavity. In this pathological scenario, the rupture of the amniochorion results from a degrading process that is potentiated by an exacerbated immunological response. Several scientific groups are working in order to understand the mechanisms related to infection and PPROM and have been focused on infection in the amniotic fluid. They have already identified a number of biomarkers specific to infection. The prognosis in PPROM is linked to maternal inflammatory markers that might predict perinatal infection, and therefore might be helpful to decide the timing of the delivery. However, the etiology of other 60% of PPROM cases, when the reason in not bacterial, remains unclear. Those cases are of the eminent interest.
Proteomic analysis, which combines two-dimensional liquid chroma-tography (2D LC) and mass spectrometry (MS), has found wide applications in protein screening in tissues obtained from healthy and diseased states for the discovery of novel diagnostic markers. The application of this new technology seems to have an important impact on the recognition of PPROM mechanisms. Improved understanding of these mechanisms should allow clinicians to design appropriate interventions so that the incidence of preterm birth and related fetal and neonatal morbidity and mortality will be significantly reduced.
Most of the time, the diagnosis of PPROM is done during physical examination. However, in 10–20% of equivocal cases, biological markers are needed to confirm the diagnosis, especially when the leakage of fluid is low or intermittent. In these cases, a quick and reliable diagnosis is neces-sary for applying the appropriate measures to reduce perinatal compli-cations.
In conclusion, detailed research is needed to identify potential bio-markers those could be used to predict patients at risk for PPROM and diagnose equivocal cases of PPROM.

9
2. AIM AND TASKS OF THE STUDY
2.1. Aim of the study
The aim of the study was to analyze the peptide composition of amniotic fluid and fetal membranes and link it to corresponding proteins in order to reveal the finding of new potential biomarkers of preterm premature rupture of membranes.
2.2. Tasks of the study
1. To investigate the peptide composition of amniotic fluid and link it to
corresponding proteins. 2. To compare peptidomes of amniotic fluid depending on the gestational
age. 3. To analyze proteomes of the digested fetal membranes and differentiate
between preterm premature rupture of membranes and term pregnancies with intact membranes.
4. To analyze the peptidome and proteome of amniotic fluid from patients
with preterm premature rupture of membranes and to search for new potential biomarkers for preterm premature rupture of membranes.
5. To compare peptidomes and proteomes of amniotic fluid, fetal
membranes and maternal plasma from patients with preterm premature rupture of membranes.

10
3. NOVELTY OF THE RESEARCH
This multidiscipline peptidome and proteome research incorporates obstetrics, biochemistry, analytical chemistry and highly advanced sepa-ration technology. For the first time in a single study peptidome and pro-teome of fetal membranes is directly compared to amniotic fluid proteome in PPROM and term pregnancies with intact membranes. Gestational age-dependent changes in peptidome of amniotic fluid during the second, early and late third trimesters of pregnancy are compared. The obtained dif-ferential peptide displays revealed new potential biomarkers of PPROM.

11
4. REVIEW OF THE LITERATURE
4.1. “Omics” technologies
4.1.1. Human genome
In 1944 deoxyribonucleic acid (DNA) as a molecule was discovered
by Avery and coworkers, the determination of its structure to be a double helix was resolved by Watson and Crick in 1953 [8,9]. The exploration of the mechanisms of expression of genetic information has opened new op-portunities for further studies of human diseases and high technologies.
In humans, each cell normally contains 23 pairs of chromosomes, for a total of 46. The DNA in each chromosome constitutes many genes. A gene is a segment of DNA containing the code used to synthesize a protein. An organism’s complete set of DNA, including all of the genes, makes up the genome. According to recent estimates, the human genome appears is made up of approximately 25,000 genes. The genome includes both the genes and the non-coding sequences of the DNA [10]. Each genome contains all of the information needed to build and maintain the organism. The gene encodes instructions that allow a cell to produce a specific protein. Proteins control the phenotype of the cell by determining its structure and by carrying out all required functions in the cell. An accurate catalog of the protein-coding genes encoded in the human genome is fundamental to the study of human biology and medicine.
For a long time, complete characterization of the genome of various species has been an aim of the scientific community. The first genome to be sequenced was that of Haemophilus influenzae in 1995. Since then, several other genomes have been entirely sequenced. Begun formally in 1990, the Human Genome Project was a 13-year effort to identify all the genes in human DNA which has been completed in 2003 [11]. Landmark papers detailing sequence and analysis of the human genome were published in February 2001 and April 2003 issues of Nature and Science [12–13]. The three most widely used human gene catalogs (Ensembl, RefSeq, and Vega) *#?6*:6"&@#.*+1.&+&*#*+-&#$&A 24, 500 protein-coding genes [14–15].
The genome of an individual determines its potential for protein expression. However, it does not specify which proteins are expressed in the cells. The complete genome of an organism gives only a relatively static overview of the functional potential of an organism and does not describe the immense dynamic process which occurs in a living organism. For

12
example, every somatic cell of butterfly and its caterpillar contains identical genetic information [16]. The conversion of this genetic information that is, the expression of the different genes into proteins takes place, however, during the different development stages of an organism as well as in dif-ferent cell types and under different environmental conditions. The absence of gene eliminates the possibility to synthesize certain protein what might directly lead to a disease; however the presence of the gene by itself does not guarantee the synthesis of the corresponding protein. This leads to an enormous individual phenotypic diversity.
Structural aberrations of the genome such as changes in the chro-mosome number and structure, changes in gene copy number, and mutations play a causal role in a number of diseases. Structural changes often result in functional genomics abnormalities, namely, changes in the gene expression patterns of individual cells. Genome-wide profiling of diseased tissues for gene copy number abnormalities has already been proven to be a fruitful strategy in cancer investigation. It is thought, that the information on the entire body of deregulated genes can be used to identify causal events in the disease and lead to the development of the personalized medicine. Therefore only the identification of coding sequences is insufficient to solve the future of personalized medicine and diagnosis.
Genomics is the study of genes or gene products in an organism. ! Structural genomics is the production and study of three-dimensional
structures of proteins. The structure of a protein is very important in determining its function.
! Functional genomics is the study of dynamic cellular processes such as gene transcription, translation, and gene product interactions that define an organism.
4.1.2. Transcriptome
After the age of groundbreaking scientific advances that took place in the middle of the twenties century, the past decades were devoted to clarification of the genotype – phenotype relationship. The basic mechanism how the genetic information contained in DNA is translated into proteins was deeply investigated. When a gene is expressed in a cell, the DNA sequence is copied by specialized enzymes – RNA polymerases – into RNA molecules during transcription. RNA molecules are processed through splicing into a messenger RNA (mRNA) which is then translated into a protein (Fig. 4.1.2.1).

13
Fig. 4.1.2.1. Illustration of DNA, mRNA, proteins and metabolites processing within the cell
Profiling of the mRNAs expressed by the genome is called transcript-
tomics [17]. This gives us an idea of the genome’s plans for possible and probable protein synthesis at that moment. Transcriptomics has the advantage over proteomics that the technology is simpler. Transcriptomics can therefore give important biological information about what genes are turned on (expressed) or turned off (repressed), and when. A major disad-vantage is that, although the snapshot provides the genome’s plans for protein synthesis, it does not represent the realization of those plans. The correlation between mRNA and protein levels is poor because many pro-teins are modified after they have been translated, so that one mRNA can give rise to more than one protein. Furthermore, the location and concen-tration of synthesized protein as well as post-translational complexation with other proteins might play an enormous role to the protein activity and function.
4.1.3. Proteome
After the Human Genome Project was finished, an increasing demand
for a functional analysis of gene products in order to understand the physiology has made proteomics, peptidomics and metabolomics highly valuable and promising technologies. The state of the organism is reflected to the key process in the living body – protein metabolism. One of the greatest challenges facing researchers in the post-genomic era is to identify, quantify and localize all expressed proteins and peptides. Proteomics is one of the most important approaches to understand gene function, because proteins expressed by genes are ultimately responsible for our phenotype. Proteins are synthesized by the translation of mRNA into polypeptides on ribosomes. Proteins are found in different cell compartments (cytoplasm, a

14
range of intracellular organelles) or as secreted extracellular proteins in various body fluids.
The term proteome originates from the words PROTEins expressed by the genOME and it was first used by Marc Wilkins at the first proteomic conference in 1994 [18,19]. It describes the expressed protein complement of a cell or a tissue at a given time. Proteomics is the large-scale study of gene expression at the protein level, which will ultimately provide direct measurement of protein expression levels and insight into the activity state of all relevant proteins [20]. The goal of proteomic research is the com-prehensive, qualitative, and quantitative analysis of all proteins expressed by genes as well as the description of changes occurring at the protein level under the influence of biological stimuli such as diseases or drug treatment. The aim of clinical proteomics is to find the function of every protein.
The proteome, unlike the genome, is not a fixed status in the organism and it changes. One gene leads to many gene products – proteins, peptides and they became different depending on site and time. Thus, every organism has one genome, but many time dependant proteomes.
The analysis of the proteome is by far much more complicated than in genomics. While the human genome sequencing endeavour was dealing with a static system composed of only four building blocks, the following battle of conquering the human proteome was of significantly higher com-plexity, namely dealing with a dynamically changing system of 20 amino acids (in humans) with a substantial range of post-translational modifica-tions (which are about 100 variations) and huge concentration differences. The dynamic range of protein expression, which stretched over the several orders of magnitude (zeptomole or yactomole per liter) with highly abun-dant proteins such as albumin in plasma or very low protein concentrations, makes the identification of the entire proteome a far more difficult and more complex challenge than the sequencing of the genome. An additional chal-lenge rises from the fact that in proteomics so far we do not have an am-plification technique, similar to polymerase chain reaction (PCR) technique which enables to create a desired number of DNA chains from a single copy. Therefore, proteins at lower concentrations detected with a higher error ratio, low protein concentrations cannot be detected at all. In con-clusion, proteomics bears a much higher degree of complexity than geno-mics, a fact which was totally underestimated when starting proteomics. Any protein, though a product of a single gene, may exist in multiple forms that vary within a particular cell or between different cells. In addition, most proteins exist in several modified forms because of different post-trans-

15
lational modifications such as phosphorylation or glycosylation. These and other post-translational modifications are crucial for the protein function, because they affect protein structure, localization and function [21].
In summary of most characteristic features in the field of proteomics are: 1) chemical constituents of widely different structure (peptides, proteins,
and different modifications: sugars, carbohydrates, nucleosides, etc.), 2) extremely large number of constituents (> 1 million), 3) high diversity in the abundance ratio (1:109), 4) large range of molecular weight (from 100 to several million Da), 5) constituents with relatively small differences in chemical structure for
example post-translational modifications of proteins: glycosylated, phosphorylated etc.; chemically small differences, however biologi-cally highly relevant,
6) number of detected constituents increase exponentially with decreas-ing concentration i.e. increasing the sensitivity.
The Human Proteome Project was launched in September 2010 with the goal of characterizing at least one protein product from each protein-coding gene [22]. In 2013 Farrah et al. assessed how much of the proteome has been detected to date via tandem mass spectrometry by analyzing PeptideAtlas, a compendium of human derived LC/MS/MS proteomics data from many laboratories around the world. It was found that this latest PeptideAtlas includes at least one peptide for each of !12500 Swiss-Prot entries, leaving !7500 gene products yet to be confidently cataloged.
4.1.3.1. Peptidome
Proteomics is a highly valuable technology for a functional analysis of gene products in oder to understand the physiology, but is restricted to proteins that are larger than 10 kDa [23]. The subproject of proteomics, namely the study of all peptides expressed by a certain cell, organ or orga-nism, is termed peptidomics. The term peptidomics was introduced in 2001 [24]. In analogy with proteomics, the aim of peptidomics is to identify all peptides. Peptides and small proteins like hormones, cytokines or growth factors act like messengers and play an essential role in the living systems with the high impact on human health. Peptidomics comprises not only peptides, originally synthesized by an organism to perform a certain task, but also degradation products of proteins (degradome). Peptidome is

16
mechanistically linked to the proteome as the cycle of protein maturation, activation, and degradation as well as the distribution between is regulated by proteases and counter-regulated by protease inhibitors. Therefore, pro-teolytic cleavage of proteins leads to peptides as indicators of protease activity and degradation. Degradome is a very important part of the protein metabolism, and thus also reflects the organism state. Peptidomics through differential peptide displays, peptide composition and amino acid sequence information delivers one insights into biological processes. However, pep-tidomics is far more challenging compared with genomics and proteomics.
The proteome/peptidome analysis usually includes the following stra-tegies: native protein pre-separation, then digestion followed by separation and identification, or alternatively straight digestion, separation and identi-fication by mass spectrometry. Therefore, starting with one protein, after digestion will end up with approximately 30 to 70 short peptide fragments. Identification of only very few of them will provide sufficient information which protein was present in the sample. Peptidomics does not possess such feature: from the beginning of the analysis to the end we have only one peptide at a certain concentration and we have to identify it. However, when peptides come from the degradome of proteins, then, naturally, peptidomics is in similar situation as proteomics. Then bioinformatics and statistics allow the assignment of the peptide fragments to the original protein.
Several promising attempts have been made to analyze the peptidome by Richter et al. [25,26]. They constructed the human circulating peptide database between tissues. To establish a mass database, all 480 fractions of a peptide bank generated from human hemofiltrate were analyzed by matrix assisted laser ionization time of flight (MALDI-TOF) mass spectrometry. Using this method, over 20,000 molecular masses representing native, cir-culating peptides were detected. Estimation of repeatedly detected masses suggests that approximately 5,000 different peptides were recorded. More than 95% of the detected masses were smaller than 15,000, indicating that the human hemofiltrate predominantly contains peptides.
4.1.3.2. Biomarkers
The general definition of a biomarker is an indicator of a biological state [27]. In medicine it is used to evaluate and measure normal biological processes, the presence or progress of disease or the effects of therapeutic intervention [28]. Proteomic and peptidomic analysis offers a powerful approach to identify disease-associated proteins and peptides that can be used as biomarkers for diagnosis and as drug targets for treatment.

17
Alterations in proteins abundance, structure, or function, act as useful indicators of pathological abnormalities prior to development of clinical symptoms and as such are often useful diagnostic and prognostic biomar-kers of a particular disease. In addition, proteins are the primary targets of most drugs and are the main basis for the development of new drugs. Also peptides hold great promise to be found as biomarkers.
Differential analysis between diseased and healthy material, treated and control is a powerful approach to detect statistically significant changes in protein expression levels. In practice it is very rare that a protein is either present or completely absent. In most cases only partly up- and down-regulations of certain proteins are observed. These quantitative changes should be determined with precision and confidence, but it is a very la-borious task.
In a biomarker discovery the main task is to find and identify novel, disease associated proteins and peptides through the systematic differential protein displays. The value of such biomarkers is based on their combi-nation of high specificity and selectivity for the biological process in which it takes place. The functions of the detected biomarkers must be proven by a second entirely independent analysis method. In most cases Western Blotting is employed for this task [29]. Identifying disease markers, proteins or peptides that appear or disappear during the course of a disease, do not necessarily require that all expressed proteins/peptides in a clinical sample should be identified – although the more complete the proteome, the more complete will be any set of markers. Therefore, there is a mutual need of highly efficient techniques to perform human proteomic/peptidomic maping to accelerate findings of biomarkers providing an answer about the health state of the patients.
Because of possible diseased location heterogeneity and other biases that might be related with biomarker identification and evaluation processes, it is essential that the identification of biomarkers should proceed in a systematic manner. In 2002, the National Cancer Institute’s ‘Early Detection Research Network’ developed a five-phase approach to systematic disco-very and evaluation of biomarkers which could be used in other fields of medicine as well. In general, biomarker development should follow an orderly process wherein one proceeds to the next phase only after meeting pre-specified criteria for the current phase [30]. In the phase 1 the focus is set on studies of preclinical exploration. Biomarkers are discovered through knowledge accumulation on gene expression profiling or protein profiling to distinguish diseased and normal samples. Identified markers are prioritized

18
based on their diagnostic/prognostic/therapeutic (predictive) value that could suggest their evolution into routine clinical use. Upon successful com-pletion of phase 1 requirements, an assay is established with a clear intended clinical use. The clinical assay could be a protein, peptide, RNA or DNA based technique, including enzyme-linked immuno sorbent assay (ELISA), protein profiles from MS, phenotypic expression profiles, gene arrays, antibody arrays or quantitative PCR. In order to ensure documented clinical usefulness, such techniques firstly need to be validated for reproducibility and also proven to be transferable among different laboratories. Then, the assays should be evaluated for their clinical performance in terms of ‘sensitivity’ and ‘specificity’ within thresholds determined by the intended clinical use. In phase 3 an investigator evaluates the sensitivity and spe-cificity of the test for the detection of diseases that have yet to be detected clinically. Samples analyzed in this evaluation phase are taken from study patients before the onset of clinical symptoms, with active follow-up to ascertain disease occurrence. Usually this phase is very time-consuming and expensive to perform; therefore, phase 3 should consist of large number of intervention trials whenever possible. This is there biomarker will become ready for clinical use once validation studies will end. Phase 4 evaluates the sensitivity and specificity of the test. An investigator can estimate the false referral rate based on tested biomarkers and describe the extent and characteristics of the disease detected. These studies are difficult to perform specifically for rare diseases. Phase 5 seeks to evaluate the overall benefits and risks of the new diagnostic test on the screened population group. The cost per life saved is one example of an endpoint for such a study. Again most probably this will require a large-scale study over a long time period and could also be prohibitively expensive. At the end of 5 phases de-termined biomarker should meet the following criteria: it should be safe and easy to measure, the cost should be relatively low, and it should be con-sistent across genders and ethnic groups. Despite intensive global efforts, most recent biomarker publications, have largely reported the inability to validate the biomarker for clinical use, rather than successful validation [31]. In fact, no new major cancer biomarker has been approved for clinical use for at least 25 years, despite the availability of highly sophisticated and powerful technologies and major advances in other areas of biomedical science. Although there have been over 10,000 publications on biomarker discovery with proteomics, a single proteomics-based diagnostic test approved by the Food and Drug Administration was HE4 protein for ovarian cancer in 2009; it was approved for monitoring recurrence. It measures five

19
different proteins in the blood, and is not used to screen for ovarian cancer, but to help evaluate whether an ovarian mass is benign or malignant prior to surgery [32].
Proteomics based identification of biomarkers for fetal abnormalities and pregnancy complications in amniotic fluid have made significant progress in the past five years. This is attributed mainly to advances in mass spectrometry-based proteomic technologies that enable new strategies for discovering biomarkers from complex biological fluids. These markers, although they still need to be verified, are diagnostic and may in the future provide targets for therapeutic intervention [33]. It is expected that peptides themselves can be used as biomarkers to monitor effectiveness and safety of drugs and to identify disease in very early stages.
4.1.3.3. Biological samples
Physiological and pathological changes are reflected in the production and the metabolism of proteins and peptides. Peptides, specifically derived by distinct proteolytic processing of specific tissue proteins, yield biomarker information. Readily released from tissue, they are detectable in extra-cellular body fluids, including blood plasma, amniotic fluid, cervicovaginal fluid, cerebrospinal fluid, synovial fluid, breast milk, urine, etc. [34]. The analysis of human body fluids constitutes one of the most important app-roaches to the diagnosis of disease and in following therapeutic inter-ventions. Human body fluids carry information about the status of the organism that may help in the recognition of physiological misbalances when overt pathological symptoms are not yet present. Analyzing the con-stituents of body fluids presents a number of challenges, the most difficult being the discrimination between variability in composition caused by an ongoing disease process and natural variability. This variability is most obvious when one is analyzing samples from different persons (cross-sec-tional studies) but is also present, albeit to a lesser extent, when one is ana-lyzing samples from the same person over a given time period (longitudinal studies) [35]. Variability cannot be avoided but may be reduced by careful selection of the study population. At any rate, the discovery of disease-related changes in the composition of body fluids requires the study of a significant number of samples from patients and controls and necessitates a careful statistical interpretation of the results.

20
Protein samples of biological origin are by nature highly complex and require sophisticated analytical tools to provide reliable analysis of the components.
4.1.3.4. Ethical issues in proteomics and peptidomics
Over the last two decades, medical research has begun to make extensive use of products of human origin in all “omics” studies. The phy-sical risks involved in donating human samples for research are usually minimal, but the risk that information from laboratory tests on a sample might harm the donor or their interests must not be forgotten. Informed consent is required from the donor and the biological sample should be treated as a gift [36].
So far, the ethical and regulatory framework for using human tissue in biomedical research is still vague, and varies between different countries. Biobanks are facilities or institutions storing materials generated from the human body. Body substances (e.g. blood, amniotic fluid) and tissue (e.g. fetal membranes) but also genetic data can be stored in a biobank.
There are different factors influencing proteomic research using hu-man samples, which are related to the patient (the supplier), to the user (the researcher), and to ethical, legal and economic framework conditions. Ethics committees usually deliver authorization for sampling a few millilitres of blood or plasma in patients or in normal volunteers easily. In this respect, proteomics researchers might benefit from favorable framework conditions. However, the problem of plasma analysis in biomedical proteomics research is complicated by the large quantities of plasma that can be necessary for identifying certain peptides present in a very low concentration, so that the plasma of hundreds of patients has to been pooled. Proteomics often requires repeated analyses over the course of disease. Of course, anony-mization of probes is not directly compatible with such follow-up studies. The problem can be solved by contracting a third party for anonymization and follow-up tasks, so that the biological information never comes in to contact with the patient’s identity [37]. Finally, another important specificity of proteomics studies is that they do not amplify genetic information, so that the data protection and privacy issues are less important than when performing, for example, genome-wide cDNA expression studies. However, researchers should not overestimate this difference since even a single, but significant piece of information gained from the protein pattern might imply significant privacy issues.

21
4.2. Fetal membranes
Although the significance of PROM has long been recognized, the
nature of the process leading to a failure of fetal membranes is poorly understood. Rupture processes are intrinsically mechanical, but the mechanisms by which biological and mechanical factors are associated with PROM are mandfatory to be investigated.
Fetal membranes are composed of two layers: the amnion and the chorion [38]. The amnion is composed of five layers: the epithelium, basement membrane, compact layer, fibroblast layer, and the spongy layer. Amnion is the innermost structure facing amniotic cavity and is lined by amnion epithelial cells. Amnion cells are purely fetal in origin and is biologically active during pregnancy. The chorion is composed of a reticular layer, basement membrane, and trophoblast layer. Although thicker than amnion, the chorion plays only a minor role in maintaining the tensile strength of the fetal membranes [39]. The amniochorion is composed of cells and extracellular matrix (ECM).
4.2.1. The amniochorionic extracellular matrix proteins
The amniochorionic ECM proteins synthesized by several cell types
within the amnion and chorion confer both strength and elasticity to fetal membranes [40]. The breakdown of these proteins is regulated by matrix metalloproteinases (MMPs) and their inhibitor ratio. The ECM is composed of an interlocking mesh of fibrous proteins and proteoglycans. ECM proteins:
1) Collagens. Interstitial collagens are located in the compact layer of amnion. Type I and III (and smaller amount of types V and VI) are the pri-mary regulators of tensile strength. The cellular source of interstitial colla-gens is unclear. Casey et al. found that both epithelial and mesenchymal cells produce interstitial collagens [41]. It seems as mesenchymal cells express significantly higher levels of collagens I and III than epithelial cells. The epithelial cell contribution is likely to be dependent upon gestational age, as the density of mesenchymal cells in the amnion decreases in the later pregnancy. Type IV and VII collagens also make important contributions to the integrity of the fetal membranes. Type IV collagen, a basement mem-brane protein, is produced of both epithelial cells of the amnion and chorion. It assists in adhesion of other components of the basement membrane, such as laminin or heparin sulfate proteoglycans [42]. Type VII is expressed by

22
epithelial cells of amnion. It stabilizes the fetal membranes by creating anchoring fibrils that link the basal lamina of the amnion to the ECM components [43].
2) Elastins are synthesized by fibroblasts, in contrast to collagens, give elasticity to tissues, allowing them to stretch when needed and then return to their original state.
3) Laminins interact with collagen VII to stabilize fetal membranes as mentioned above.
4) Fibronectins are glycoproteins that connect cells with collagen fibers in the ECM. It can be thought to be "trophoblast glue" and is found at the placental-uterine and decidual-fetal membrane interfaces. It is releases when the ECM of chorionic-decidual interface is disrupted. It is used as a biochemical marker for diagnosis and prediction of preterm labor.
A balance between the synthesis and the degradation of membranes components is physiologic throughout the gestation. Two main mechanisms are involved in the degradation process: apoptosis in the cellular compartment and ECM degradation by MMPs [44]. Regulation of MMP is depending on factors increasing their expression (cytokines) and factors decreasing their activity tissue inhibitor of metalloproteinases (TIMPs). Particular conditions can induce an unbalance between synthesis and degradation leading to the weakening of membranes. Different factors can be associated to induce this unbalance: infection, hormonal factors, and default in membranes fusion, oxidative stress and mechanic factors. The spontaneous rupture of the membranes is always occurring in regard of the uterine cervix after a process started several weeks before.
4.2.2. Matrix metalloproteinases
Matrix metalloproteinases are a large family of calcium-dependent
zinc-containing endopeptidases, which are responsible for the tissue remodeling and degradation of the extracellular matrix [45]. The human MMP family currently consists of 26 members and is classified according to substrate specificity into collagenases, gelatinases, stromelysines, matrily-sins, membrane type-MMPs and other MMPs. Aberrant ECM degradation by activation of the MMP cascade or an imbalance between MMPs and their tissue inhibitors have been implicated in the pathogenesis of preterm labor and rupture of membranes [46]. PROM is associated with increased levels of active, TIMP free forms of MMP 2 and 9, in the amniotic fluid. Intra-

23
amniotic infection was associated with a significant increase in amniotic fluid MMP-3 concentrations in both women with preterm labor and intact membranes, and women with preterm PROM [47]. The intrauterine infection (IUI) triggers MMP production via inflammatory mediators [48].
4.2.3. Tissue inhibitors of metalloproteinases
TIMPs are endogenous specific inhibitors that have been shown to regulate the proteolytic activity of MMPs in normal and pathological processes [49]. A family of protease inhibitors include: TIMP-1, TIMP-2, TIMP-3, and TIMP-4. A fully functional TIMP network has been demons-trated in human fetal membranes [50,51] in placenta and decidua, and in amniotic fluid during the second trimester [52]. The majority of studies focused on TIMP-1 and TIMP-2. TIMP-1 concentrations in AF were in-creased in the presence of IUI and in patients with rupture of the membranes either term or preterm. In contrast, TIMP-2 levels were decreased in women with IUI and rupture of membranes.
4.3. Physiology of amniotic fluid
Amniotic fluid is fundamental for the normal development of the
fetus. It protects the fetus physically and biochemically. AF resides in the amniotic cavity that is lined by the fetal membranes.
4.3.1. Composition of amniotic fluid
Amniotic fluid is constituted of about 98–99% of water. The amniotic sac, which contains the embryo, forms about 12 days after conception. AF immediately begins to fill the sac [53]. During embryogenesis, AF is ini-tially formed from maternal plasma that passed through fetal membranes. Because free diffusion occurs bidirectionally between the AF and the fetus across fetal skin, placenta, and umbilical cord from 10 to 20 weeks of gestation, AF composition becomes similar to that of fetal plasma during this period. Therefore, analysis of AF composition before skin keratini-zation, which occurs between 19 and 20 weeks of gestation, would reveal valuable information that may indicate physiological or pathological con-ditions of the fetus. Fetal urine first enters the amniotic sac at 8–11 weeks’ gestation and makes up most of the AF throughout the second half of

24
pregnancy. The biochemical composition of AF is complex and varies with the gestational age. AF contains proteins, amino acids, carbohydrates, hormones, lipids and electrolytes. In contrast to other biological fluids, like plasma and serum, AF has a low protein concentration and relatively high carbohydrate and lipid content. Many of the protein molecules present in AF are structurally glycoconjugates and are protected against proteolytic attack from the extracellular matrix proteases. The increasing levels of enzymes and electrolytes in the later part of gestation correlate with the formation of the fetal kidneys, lungs, and the gastrointestinal tract. Studies have revealed the occurrence of growth factors, innate immunity molecules, and serum components in AF that are believed to be involved in growth, development, and protection of the fetus from infection [54]. The concentration of each protein in the AF is governed not only by fetal, placental, or maternal synthesis and degradation, but also by exchanges between the mother and the fetus through the placenta. Fetomaternal transfer of proteins involves several different mechanisms. Consequently, the concentration of each AF protein results from a balance between opposing dynamic metabolic and physiological processes, which proceed simultaneously.
4.3.2. Regulation of amniotic fluid volume
The amniotic fluid volume is related to the gestational age. The quan-tity of AF is the balance of water exchange between the mother and fetus, and is maintained within a relatively narrow range during all stages of pregnancy [55]. The amount of AF is highly regulated, although the exact mechanisms for regulation are not entirely clear [56]. Initially AF produc-tion is attributed to the amniotic epithelium and later to fetal kidneys and lungs [57]. Therefore, the two main sources of amniotic fluid are fetal urine and lung fluid, with an additional small contribute due to secretions from the fetal oral-nasal-tracheal cavities. The capillary bed in the fetal skin is also utilizes as an exchange surface. Part of AF may be derived from water trans-port across the highly permeable skin of the fetus, at least until kerati-nization of the skin occurs (intramembranous pathway). Transfer through the umbilical vessels is also believed to play a role in AF production. The two primary routes of amniotic fluid removal are fetal swallowing and absorption into fetal blood perfusing the fetal surface of the placenta and membranes (transmembranous pathway). Approximately half of the daily fetal urine output is eliminated by fetal swallowing.

25
4.4. Amniotic fluid proteome
Proteomics allows simultaneous study of a multitude of proteins and is
of great importance to gain insight into the physiology of amniotic fluid. A proteome is never static; therefore a snapshot of a protein expression status without any biological context cannot contain valuable information. However, proteome maps are necessary for establishing databases for protein identification and characterization. Amniotic fluid contains large amount of proteins and peptides produced by the fetal membranes cells, fetal tissues, fetal excretions and placental tissues [58]. The amniotic fluid proteome is therefore composed of urine proteins, intestinal proteins, alveolar fluid proteins, and their degradation products. In addition, cellular proteins are produced, either by the skin of the fetus or directly by the amnion. Amniocentesis is a method used to obtain samples for amniotic fluid proteome analysis. In order to identify patterns within the complex proteomic profile that can discriminate between normal and disease states it is needed to know the normal amniotic fluid proteome. A systematic analysis of proteins present both in AF and maternal serum could lead to the development of new noninvasive diagnostic procedures.
4.4.1. The normal human amniotic fluid proteome
Amniotic fluid is a potential source of biomarkers for many disorders
that may occur during pregnancy or for embryonic abnormalities. The first detailed study of the amniotic fluid proteome was published in 1997 by Liberatori et al. [59]. The authors identified 31 human AF protein in AF supernatant by two dimensional electrophoresis followed by postseparation analysis techniques such as N-terminal sequencing of human AF obtained at the 17th week of gestation. At this time AF is most commonly obtained for prenatal diagnosis of chromosomal abnormalities by amniocentesis.
In 2004 Nilsson et al. identified 43 proteins in AF proteome [60]. The AF sample (n = 1) was taken at the 15th week of gestation from a healthy 36 year old woman. AF sample was digested and peptides were separated by gradient capillary LC followed by electrospray ionization and mass spectrometric detection with a 9.4T Fourier transform ion cyclotron reso-nance mass spectrometer. They were the first to use MS for profiling AF. They were also the first to deplete albumin from AF in order to identify more proteins. In addition, the study revealed that a combination of different

26
proteomic methods should be used to evaluate the proteome of amniotic fluid globally.
In 2006, three groups analyzed normal AF proteome. Park et al. reported 37 proteins (n = 8, term pregnancy) by using 2DE followed by MALDI-TOF-MS [61]. Michel et al. identified 69 proteins (n = 10, term pregnancy) from albumin-depleted AF by Off-Gel electrophoresis LC/MS/MS [62]. Tsangaris et al. identified 136 proteins (n = 16, 16–18 weeks of pregnancy) by 2DE followed by MALDI-MS/MS [63]. These different groups not only used different approaches as well as different protein databases, but also applied different levels of stringency for protein identification that makes difficult to assess the accuracy of each data set. As illustration, the group who identified the highest number of proteins also has identified many non-human proteins from human AF.
In 2007, Cho et al. reported on the most extensive protein profile of the second trimester normal human AF, which is comprised of 1026 unique gene products from 842 different genes [64]. AF samples (n = 16) between gestational ages of 16 and 18 weeks were taken from women carrying chromosomally normal fetuses. Three samples were fractionated by strong anion exchange LC, another three were fractionated by strong cation exchange LC, and 10 samples were pooled together and fractionated by LC 2D PAGE. The protein analysis followed by a common reverse phase LC/MS/MS. Mascot and The Global Proteome Machine engines were used to search the International Protein Index human database for peptide sequence identification. The list of proteins was generated by combining the results of both engines. All proteins that were previously reported were identified in this study. Assuming that all of the proteins identified in previous publications and in this study are correct, a total of 936 proteins have been identified from human AF so far. Despite interest regarding composition and functions of amniotic fluid, there have been limited attempts to generate an in-depth analysis of its proteome. Each identified protein was assigned a subcellular localization based on information from Swiss-Prot, Entrez Gene, and Gene Ontology databases. The cellular distribution of 558 identified proteins with known localization was reported. The majority were extracellular (42%) and membrane (26%) proteins. Tissue expression of each protein was searched from Swiss-Prot, Entrez Gene, and Gene Ontology databases. The tissue expression of 301 proteins was identified. Some of the organs to which many proteins were attributed include kidney, placenta, lung, liver, and heart. 24 proteins were specifically annotated as being expressed from embryonic organs/tissues. The authors

27
utilized Ingenuity (Ingenuity Systems) to retrieve known functions of each protein. 221 (of 842) were matched with functions to retrieve known functions of each protein. Major categories included cellular movement, development and function of organs, cellular growth and proliferation, cancer, and cell-to-cell signaling. The top 15 proteins with the largest number of unique peptides with the top 15 proteins from the human plasma proteome were compared. Among these 15 abundant protein4& $"#B&CDE&F-$6*#G"#*61.E& *"+.4$#"B1.?& ?"#H*:& $+@*#"& I-induced protein ig-h3 precursor, and periostin are found at relatively low concentrations in plasma and are not included in the top 15 plasma proteins. Conversely haptoglobin, which is one of the most abundant proteins in plasma, was found at low concentration in AF, and apolipoprotein B was identified in AF by only one group, indicating its low abundance in AF.
The major challenge and the bottleneck in proteomics lie between protein discovery and target validation. Proteins with potential as bio-markers should be selected for further comparative analysis of expression and structural modifications with samples from normal and abnormal pregnancies. Currently used markers have insufficient individual detection rate and specificity; therefore the use of multiple markers is necessary.
4.4.2. Gestational age-dependent changes
Many proteins detected in AF are already present at a very early stage
of gestation, whereas other proteins are detected only at the end of the pregnancy [65]. The protein composition and concentration of AF varies throughout pregnancy.
Michaels et al. analyzed AF samples from three trimesters of preg-nancy [66]. The authors used differential dye labeling of proteins resolved by 2D PAGE to determine the general differences in AF protein com-position over time. Totally 219 AF proteins were identified. The largest protein abundance changes appeared to be between the first and second trimesters. The changes seen in expression of the proteins are indicative of the role the molecules have in fetal development. Among the proteins that show increased level are Apo A1, Apo A2, IGFBPs, gamma glutamyl transferase 4, and pigment epithelial derived factor. The increase of apoli-poproteins between the first and second trimesters may contribute to the fetal lung development that occurs at this stage. The increase in the level of gamma glutamyl transferase 4 is showing increases in metabolic enzymes during pregnancy. The relative amounts of kininogen, ceruloplasmin, angio-

28
tensinogen, alpha 2 HS glycoprotein, orosomucoid, and ubiquitin are decreased between the first and second trimesters. The significance of this change is not clear, as the function of these molecules is not understood. It is likely that the immediate expression of these proteins is most important during early fetal development that occurs during the first trimester. The authors also identified a set of proteins consistently expressed in all three trimesters. Transthyretin and several IGFBPs were present at high levels throughout gestation. These proteins may have utility as controls for the normalization of differentially abundant biomarkers in diagnostic multi-analyte assays.
Queloz et al. compared the proteomic profiles of normal AF obtained at 17 and 40 weeks of pregnancy using 2DE and silver staining, as well as two-dimensional difference in gel electrophoresis [67]. Results showed that some proteins were more abundant in early pregnancy, while others were over expressed at term, suggesting that the protein profile of AF is dynamic and changes occur during development. These observations are particularly important when biomarkers for a specific condition are identified.
4.4.3. Clinical applications of amniotic fluid proteomics
Researches focus on the emergence of proteomics as a major platform technology in studying AF and developing biomarkers for fetal aneuploidies [68] and pregnancy related disorders. Pregnancy related disorders such as preterm premature rupture of membranes, preterm labor and intraamniotic infection (IAI), intrauterine growth restriction, preeclampsia [69] contribute significantly to maternal and fetal mortality. Although several pathways for the pathogenesis of pregnancy complications have been proposed, the basic molecular mechanisms that modulate these events remain incompletely understood. Discovery of clinically and biologically relevant biomarkers able to reveal key pathogenic pathways and predict pregnancies at risk for antenatal fetal damage is a priority. Proteomics provides a unique oppor-tunity to fill this gap [35].
4.4.3.1. Premature rupture of membranes
In 2003 Vuadens et al. identified new potential biomarkers for premature rupture of membranes [70]. This result was achieved by research group of !"#$%& ,%'%& J144#*& from Switzerland. Proteomic studies were per-formed on samples collected from women at term (pairs of maternal plasma

29
and AF) as well as on samples of AF collected at the 17th week of gestation. Their study was based on two-dimensional gel electrophoresis differential display between AF and plasma samples, followed by micro-sequencing analysis of specific spots. Two „amniotic fluid-specific“ proteins were identified. These peptides were fragments of proteins which are present only in AF but absent in normal human plasma of corresponding mother. These two peptides were identified as COOH-terminus fragments of agrin (appa-rent molecular weight: 19.1 kDa, pI: 5.3; SwissProt: O00468) and perlecan (apparent molecular weight: 19.6 kDa, pI: 5.62; SwissProt: Q9H3V5), respectively, both of which are heparan sulfate proteoglycans. These results were further confirmed by Thadikkaran et al. [71]. Their physiological roles in amniotic fluid remain unknown. However, they are thought to mediate the action of growth factors and be involved in developmental processes. The origin of these peptides and possible roles are reviewed by Cretazz et al. [67].
Michel et al. applied the Off-Gel isoelectric focusing technique followed by tryptic digestion of the proteins and by LC/MS/MS to analyze the plasma and AF sample from a woman at term pregnancy. Totally 73 and 69 proteins were identified in maternal plasma and AF samples. Proteins, found in AF have been compared to those identified in the mother plasma as well as to the reference human plasma protein list [72]. Systematic com-parison revealed that nineteen proteins were specifically present in the AF and absent in maternal plasma and may be considered as potential markers of PROM (Table 4.4.3.1.1). Among these proteins, amiloride-sensitive amine oxidase [73,74] and perlecan have already been recognized as potential biomarkers of PROM by other researchers previously [72].

30
Table 4.4.3.1.1. List of the proteins identified in AF and absent in maternal plasma Swiss-Prot
accession
number
Protein name
O75363 Breast carcinoma amplified sequence 1
O94832 Myosin Id
P07093 Glia derived nexin precursor
P10451 Osteopontin precursor
P13535 Myosin heavy chain, skeletal muscle perinatal
P13987 CD59 glycoprotein precursor
P19801 Amiloride-sensitive amine oxidase [coppercontaining] precursor (EC1.4.3.6)
P35527 Keratin, type I cytoskeletal 9
P46100 Transcriptional regulator ATRX
P98160
Basement membrane-specific heparan sulphate proteoglycan core protein precursor (perlecan)
Q03519 Antigen peptide transporter 2
Q12841 Follistatin-related protein 1 precursor
Q13421 Mesothelin precursor
Q14204 Dynein heavy chain, cytosolic
Q14644 Ras GTPase-activating protein 3
Q8N3R9 MAGUK p55 subfamily member 5
KLM,KN& Protocadherin 16 precursor
Q9NTG1 Polycystic kidney disease and receptor for egg jelly related protein precursor
Q9Y4C8 Probable RNA-binding protein KIAA0682
The role of these proteins in amniotic fluid is still unknown. Accord-
ing to Michael et al., among the proteins identified as being specifically present in AF several are linked to pregnancy. Transcriptional regulator ATRX could be a global transcriptional regulator. It modifies gene expres-sion by affecting chromatin, and it may be involved in brain development and facial morphogenesis which is very likely to occur during the preg-nancy. Follistatin related protein 1 precursor may modulate the action of some growth factors on cell proliferation and differentiation. Probable

31
RNA-binding protein KIAA0682 has already been identified in the uterus. Dynein heavy chain cytosolic and osteopontin have already been detected in placenta. Keratin type I cytoskeletal 9 has already been found in human placenta. Mesothelin precursor is linked to ovarian activity, and myosin heavy chain, skeletal muscle perinatal is also linked to pregnancy.
In 2007 Cho et al. analyzed the AF proteome and compared the proteins present in AF with proteins from human plasma [64]. By searching the Plasma Proteome Database found that 304 (36%) of the 842 proteins have also been found in plasma and concluded that this does not mean that the remaining 538 proteins are exclusive to AF because the plasma pro-teome list is still growing. Also, several putative markers as well as cur-rently used markers for PROM were found in AF including prolactin, alpha-fetoprotein (AFP), IGFBP-1, fibronectin, agrin, plasma retinol-binding protein precursor, apolipoprotein A-I, B-factor. These proteins have been already mentioned as potential markers in literature. This study identified the new marker candidate for PROM - :OP&I chain.
However, the validation of these proteins as biomarkers of PROM was not done. In order to confirm proteins or peptides as biomarkers of PROM, they should be present in AF, absent in maternal plasma, and should be easily detected in vaginal fluid of patients with PROM. The proteome of normal human cervico-vaginal fluid (CVF) is already established and described in the literature [75,76]. Wang et al. used a protein array to screen amniotic fluid samples and cervical-vaginal fluid collected from normal and PROM pregnant women. Enzyme-linked immunosorbent assay was used to quantify two novel and potentially useful analytes, soluble intercellular adhesion molecule-1 (sICAM-1) and Axl receptor tyrosine kinase (Axl). Comparing 110 CVF samples of PROM/PPROM with 110 CVF samples of normal pregnancies, the diagnostic value for PROM was demonstrated by their high sensitivity and specificity (96.4 and 92.7%, respectively, for sICAM-1, and 92.4% and 90.4%, respectively, for Axl) [77]. sICAM-1 or Axl can be developed into a rapid strip test for bedside use.
No significant relationship, however regarding the sensitivity, speci-ficity and predictive value for the accurate detection of women at risk for PROM has been demonstrated up to now. Though, the analysis shows that proteomics is a valuable approach to gain insight into the physiology of amniotic fluid and to identify new potential biological markers for PROM diagnosis. The data further support the view that quantitative shotgun proteomics analysis of AF may be a feasible and effective method to screen multiple pathologies in the future.

32
4.4.3.2. Amniotic fluid infection
Intraamniotic infection (IAI) is linked with PPROM and preterm birth. Infection of amniotic fluid and membranes has been implicated as the major known cause of PPROM. The presence of IAI strongly associated with the adverse neonatal outcome. Early diagnosis of IAI is problematic, however, because clinical signs and symptoms (including preterm labor or rupture of membranes) tend to be late manifestations of this condition. Furthermore, the available noninvasive diagnostic tests of IAI have limited predictive value. The goal is to discover the sensitive and specific noninvasive test to predict clinically undetectable IAI by proteomic profiling methods.
By using surface-enhanced laser desorption/ionization time of flight MS (SELDI-TOF/MS), gel electrophoresis, and tandem mass spectrometry, Gravett et al. were able to characterize several amniotic fluid peptides in a model of animal infection [78,79]. Markers such as calgranulin B (S100A9), azurocidin, vitamin D binding protein, and insulin-like growth factor binding protein 1 were tested in a cohort of 11 women with occult IAI by liquid chromatography – tandem mass spectrometry. Based on the results of this study, calgranulin B and a proteolytic fragment IGFBP-1 have been proposed as candidate biomarkers for IAI. Similarly, Ruetschi et al. reported that human neutrophil defensins 1-3 and calgranulins A and B are part of the AF fingerprint characteristic of intraamniotic inflammation and/or infection [80]. Park et al. found altered expression of Calgranulin A and B in human amnion and AF samples obtained from pregnant women infected with Ureaplasma urealyticum. The results showed that specific biomarkers in amniotic fluid might have application in the early detection of intraamniotic infection. It is of special interest that the same differences in protein expression were also found in maternal serum, allowing for noninvasive detection of IAI. The role of IGFBP-1 biological activity in IAI was confirmed by Bujold et al. using a combination of techniques involving 2D chromatography, MS and immunoassays too [81]. All data together suggest that the total amount of IGFBP-1 does not change, but that IAI leads to increased proteolytic degradation of IGFBP-1 resulting in an increased con-centration of a fragment of IGFBP-1 (at about 13.5 kDa) [82]. The same study reported protein profile that was over-expressed in amniotic fluid of women with IAI who delivered preterm. Fibrinopeptide B, transferrin, ma-jor histocompatibility complex (MHC) class 1 chainrelated A antigen frag-ment, transcription elongation factor A, sex-determining region Y (SRY) box 5 protein, DSCR2, and HP8 were linked with IAI.

33
From 2005 two groups lead by C. Buhimschi and R. Romero mainly investigated the presence of biomarkers of IAI and the relationship with preterm labor (PTL) by proteomics tools. Buhimschi et al. also used surface-enhanced laser desorption/ionization time of flight MS and profiled specific proteins for infection in AF and reported on the existence of four bio-markers, including neutrophil defensins-1 and 2, calgranulins A (A100A8) and C (S100A12). Based on the presence or absence of these biomarkers, they devised the called the „mass restricted (MR) score“ ranging from 0 (all biomarkers absent) to 4 (all biomarkers present) [83]. A MR score >2 was associated with imminent preterm delivery. If no biomarkers are present, then the pregnancy is considered to be uncomplicated while an „MR score“ of three or four is highly predictive of adverse pregnancy outcome. Proteo-mic analysis of amniotic fluid was shown to be the most accurate test for diagnosis of intraamniotic inflammation, whereas addition of the MR score to the Gram stain provides the best combination of tests to rapidly predict infection [84]. The authors observed a sequential appearance of the bio-markers as the process of intraamniotic inflammation developed from acute to chronic, with S100A12 and S100A8 appearing last. In the later studies Buhimchi et al. confirm that high MR scores are associated early onset neonatal sepsis [85,86]. Presence of S100A12 and S100A8 in AF is pre-dictive of early-onset neonatal sepsis and poor neuro-developmental out-come [87]. This methodology shown that proteomic profiling of the AF can rapidly and accurately diagnose IAI, and can identify the subgroup of patients that might benefit most from interventions to prevent fetal damage in utero [88]. The biomarkers comprising the MR score have a unique ability to predict in utero clinically relevant histological chorioamnionitis [89] and funisitis [90], which are known risk factors for sepsis and poor neonatal outcome [91]. Presence in the AF of S100A12 (EN-RAGE, ligand for the advanced glycation end products (RAGE) receptor) [92,93] had the strongest correlation with chorioamnionitis and funisitis.
Romero et al. using liquid chromatography tandem mass spectrometry with isobaric labeling of the AF proteome of women with PTL identified proteins differentially regulated in women with IAI and in those without IAI who delivered preterm [94]. Importantly, many novel proteins were found to be up-regulated in the AF of patients with PTL and IAI including leukocyte elastase precursor, Thymosin-like 3, and 14-3-3 protein isoforms.

34
4.5. Human plasma proteome
In proteomics the term „plasma“ is used to embrace all the proteins and protein components of the blood soluble phase (excluding cells) [95]. Human blood plasma can be obtained relatively non-invasively and contains proteins from most, if not all, tissues of the body and provides a window into an individual's state of health. Therefore, an extensive, quantitative catalog of plasma proteins is an important starting point for the discovery of disease biomarkers. Blood plasma has an exceptional proteome in many respects. It is the most complex human-derived proteome, containing other tissue proteomes as subsets. It is the most difficult protein-containing sample to characterize on account of the large proportion of albumin (55%), the wide dynamic range in abundance of other proteins, and the tremendous heterogeneity of its predominant glyco-proteins. Plasma represents the largest and deepest version of the human proteome present in any sample: in addition to the classical “plasma proteins” as those that carry out their functions in the circulation, it contains proteins that, for example, serve as messengers between tissues (e.g. peptide hormones) or all tissue proteins (as leakage markers that leak into the blood as a result of tissue damage) plus very numerous distinct immunoglobulin sequences. It has an extraordinary dynamic range in that more than 10 orders of magnitude in concentration therefore, it is often difficult to observe low-abundance proteins of interest among high abundance proteins.
As the era of proteomics started, the widespread adoption of liquid chromatography-tandem MS (LC/MS/MS) techniques resulted in a rapid increase in plasma proteome-related data sets that needed to be similarly integrated to form a next-generation comprehensive human plasma proteo-me reference set. In 2002, the Human Proteome Organization (HUPO) launched Human Plasma Proteome Project (HPPP). The HPPP stimulated access to emerging technologies and generated substantial datasets and inte-grated databases for proteins detectable and identifiable in human plasma.
In 2005 The PeptideAtlas Project was started [96,97]. It comprises a growing, publicly accessible database of peptides identified in many tandem mass spectrometry proteomics studies and software tools that allow the building of PeptideAtlas [98], as well as its use by the research community [99]. Using the first PeptideAtlas 6929 peptides were identified at a peptide false discovery rate of 12%, mapping to about 960 distinct proteins. Comparison of protein identifiers with those from different human proteome studies showed quite limited overlap. In 2011 Farrah et al. [100] tried to

35
compile a larger human plasma proteome reference set of similar high confidence by creating a new release of the Human Plasma PeptideAtlas incorporating more data than earlier and interpretate the data using more stringent criteria. The result was 20,433 distinct peptides and a plasma proteome reference set containing 1929 highly non-redundant protein sequences at a false discovery rate of 1%. Still, it is believed it is far from a complete catalog of the human plasma proteome. Now PeptideAtlas is an integral part of the ProteomeXchange infrastructure for HUPO initiatives and other worldwide data submissions.
4.6. Methods of proteomic analysis of amniotic fluid
Proteomic analysis of a biological sample usually consists of four steps: extraction of the proteins from the sample, their separation, detection and finally identification/analysis of the individual separated proteins or peptides. Analyzing body fluids sample collection and consecutive va-riations in treatment procedures play a major role on sample quality. The group of Schulz-Knappe concluded that specimen collection is a crucial step for successful peptide biomarker discovery in human samples [101]. Initial sample treatment is the major step which ensures how representative the data are and which kind of component losses could be acceptable. In general, sample preparation protocols that limit the number of preparation steps, circumvent the loss or dilution of the sample and purify and con-centrate the sample. Therefore, the most desirable sample pretreatment methods are those which are totally automated. Automation eliminates human type errors and also drastically increases the throughput. Another important issue while working with patient biofluids is safety of the re-searcher who is at higher risk for catching an illness. Fully automated sample treatment reduces that risk significantly.
In general only three methods are used for biofluid proteome analysis: 1) two dimensional gel electrophoresis (2DE), 2) affinity arrays and 3) liquid chromatography (LC). Each of these methods has their strengths and weaknesses depending
on the task, molecular weight range of the proteins, sample complexity, etc.

36
4.6.1. Gel electrophoresis
Gel electrophoresis for protein separations was pioneered by Arne
Tiselius in early 30s then later, in 1975, developed into 2D PAGE by Farrell [102,103]. This technique offered the separation capacity of 10,000 proteins, therefore quickly gained enormous attention. Since then, multiple pieces of work using this approach in order to tackle various biological questions have appeared in the literature. The main advantages of method are high separation power (~10,000 proteins) and simplicity. However 2D PAGE fails to analyze <15 kDa proteins (peptides) (proteins ~10 kDa does not focus well in isoelectric focusing step, proteins <15 kDa could be easily lost as they migrate too fast), hardly can cope with concentration differences of several orders of magnitude and lack of automation. Figure 4.6.2.1, column A shows the standard procedure using 2D PAGE: biological sample is pretreated during sample preparation step, applied protein mixture is separated by isoelectric focusing in the first dimension, and then proteins are separated by size exclusion in the second dimension. When analysis is finished the gel is stained, the desired spots are cut and digested into peptides by sequence-specific proteases (usually Trypsine). The resulting peptides are separated using reverse phase chromatography coupled to the mass spectrometry. Matrix-assisted laser desorption/ionization is performed to produce a mass spectrum or “peptide mass fingerprint”. The second step in protein identification relies on the fragmentation of individual peptides in the mixture to gain sequence information. Such approach usually provides very high confidence of identified proteins. It is through the integration of 2DE and MS that proteomics achieves its greatest power. Both, mass spectrum and sequence information can be searched against databases to identify proteins.
4.6.2. Protein arrays
Protein arrays are solid-phase ligand-binding assay systems using
immobilized proteins on surfaces such as glass, cellulose membranes, mass spectrometer plates, micro-beads, or micro/nano-particles [104]. The assays are highly parallel and often miniaturized. The main advantages of protein arrays include high-throughput, automation, relatively simple, exquisite sensitivity, and minute sample required for analysis (Fig. 4.6.2.1, E). Protein arrays can be also coupled to mass spectrometry (for example surface-enhanced laser desorption/ionization mass spectrometry). However, the

37
expression and purification of capture proteins, especially antibodies, is cumbersome. The design of capture arrays, particularly when screening against complex samples, also needs to take into consideration the problem of cross-reactivity. A technical difficulty in fabrication of peptide arrays is that peptides, usually with small molecular mass, are not easily accessible when adsorbed onto solid supports. Peptides also lack a well-defined 3D structure, and therefore a correct orientation is essential to promote the interaction between peptides and their targets. Incorporation of elongated spacer molecules or modification of the solid substrate can significantly improve the accessibility of a target molecule. This method is mainly used for the monitoring of known proteins, and as a second method verifying speculated biomarkers.
Fig. 4.6.2.1. Liquid chromatography workflow strategy options in proteomics
A – 2D PAGE followed by LC/MS, B – “bottom-up” approach, C – “top-down approach”, D – selective sample clean-up directly combined with chromatographic separation,
E – peptide capture with affinity type material.
DigestionDigestion Sample prepLC
MD LC
MS or MS/MSMS or MS/MS MS or MS/MS
LC or MD LC
Sample prep
LC
Sample prep
LC or MD LC
Pro
tein
sP
epti
des
B D
Sample prepAffinity
MS or MS/MS
E
Digestion
MD LC
MS or MS/MS
Sample prep
A
2 D PAGE
C

38
4.6.3. Liquid chromatography
Liquid chromatography (LC) has several advantages features for the
proteomic analysis. First of all, LC offers superior automation possibilities and high flexibility. One can vary the selectivity, and every different selectivity (or selectivity influenced by chromatographic conditions) will allow catching different proteinaceous substances. Sample volumes in proteomics could vary drastically from micro liter range for example blood or plasma to milliliter range as for example analyzing urine. Column size adjustments (mainly inner diameter) could handle the sample capacity (loadability) of the system. The sensitivity is boosted by reducing the column internal diameter. The combination of selectivities in multidimen-sional (MD) LC helps to obtain high peak separation capacities. Quan-titation is often problematic dealing with high complexity samples. Also in LC based approaches the handling of extreme abundance differences is quite complex and requires large experience. Automated sample processing in combination with adjusted column dimensions and elution sequences provides an optimum solution. The limitations of the approach are related to biological sample stability at the conditions of the optimum LC analysis. Proteins are prone to aggregation and precipitation especially when the pH, concentration and ion strength are altered. Chromatographic set up contains many tiny capillary connections, column filters/frits – all could be easily clogged if any precipitation occurs. Time of the analysis, also the tempe-rature influence the stability of the sample components. Therefore, LC copes better with the peptides than with the proteins. Despite its pros and cons LC already has been proven in several cases for automated sample clean up, depletion and enrichment and qualitative analysis. Summarizing last sentences it should be stated, that chromatographic separation methods require thoroughly selected sample preparation procedures. In general, sample preparation protocols that limit the number of preparation steps, circumvent the loss or dilution of the sample, preserve purity and con-centrate the samples are mostly wanted. Automation eliminates human type errors and also drastically increases throughput.
While MD LC/MS has found widespread use in the analysis of peptides from natural sources or generated by proteolytic digestion of larger proteins, the method is not suitable for analyzing proteins directly. First, proteins tend to denature under reversed-phase conditions either by stationary phase or mobile phase induced effects (strongly hydrophobic surfaces, low pH and high organic solvent concentrations) making their

39
quantitative elution rather difficult. Also, measuring the molecular mass of a protein by MS is not sufficient for its unambiguous identification. To circumvent these obstacles the proteins are digested and the separation is performed at the peptide level. One can distinguish two approaches: (1) proteins are separated and then digested (“top-down” proteomics, see Fig. 4.6.2.1, B)); (2) in “shotgun” proteomics a complex protein mixture is first digested and peptides are then chromatographically resolved (“bottom-up” proteomics, see Figure 4.6.2.1, C) [105]. In both cases, separation technologies play a critical role in protein identification and analysis.
Mass spectrometry is the main detector in proteomic analysis. Despite the recent breakthroughs in this technology it has still have limitations with respect to sensitivity. Therefore, a certain number of analyte molecules should be injected in order to be identified. Thus, higher amounts of the sample should be applied. Knowing the target analyte concentration in the sample provides the answer to the question: how much we should inject? According to Geigy scientific tables [106] for example human plasma con-tains only 0.03% peptides (dry mass). It might be estimated that in plasma several ten thousands different peptides could be observed with vast concentration differences. Therefore, huge injection volumes might be required. For example Tatemoto et al. extracted 0.6 mg of Peptide YY from four tons of porcine intestine [107]. Dart et al. obtained 47 µg of trans-forming growth factor-I& $"#B& Q%Q kg of human placenta [108]. Another important prerequisite for the suitability of a separation system for proteo-mic analysis is the ability to handle very small amounts of biological ma-terial [109]. These methods allow one to detect low concentrations of pep-tides from complex mixtures with a high degree of automation.
Biological, individual, and variations between individuals (such as gender, age and nutrition) affect peptidomes and require careful conside-ration in order to find valid biomarkers. A few, equally important factors for successful proteomic biomarker research are high sample quality, high sensitivity, and reproducibility which depend on proper selection of the high quality samples.
4.6.3.1. Sample preparation
In general HPLC analysis sample preparation is considered as the most essential step having a direct impact on the analytical results. Sample preparation including direct injection techniques serves the following purposes:

40
! eliminates undesired components to enhance resolution, ! removes interferences that affect the accuracy of quantitation, ! concentrates the analyte of interest to increase sensitivity.
Chromatographic separation techniques are well suited for the analysis
of high complexity samples. However, the analysis of biological materials requires well designed sample preparation procedures. In the early period of chromatography attempts were made to inject biological samples directly onto the LC column [110]. It was realized that this approach let to a rapid loss of column performance and selectivity, the column back pressure increased, as well as the irreversible adsorption of matrix compounds. Some useful means of sample preparation are required to reduce the sample complexity and to remove components which tend to bound irreversibly. Protein precipitation with 70% methanol is often used in order to remove most of the proteins [111]. It is important to note that the temperature and time are playing an important role in protein precipitation. Therefore, precision in sample handling time and temperatures should be monitored. Another concern is the mobile-phase pH, with precipitation occurring in the protein's pI. Classical methods of sample preparation include precipitation of proteins and/or extraction of analytes into organic phases (liquid-liquid extraction). Liquid-liquid extraction is the most common method for sample preparation which serves the dual function of sample clean-up (to eliminate interfering compounds) and to remove the analyte from the protein matrix which is mandatory in peptidome analysis. Solid phase extraction cartridges in proteome research were used with moderate success as these methods are labor intensive and time consuming. They also require a large volume of sample (at least 1 mL), as well as the addition of an internal standard (due to analyte loss resulting from the multiple sample manipulations). Losses of sample components might happen whenever the sample molecules touching 01$$6"6.*&43"$+@64%&>%&,#:.&6*%&+-%&0641?.60&+&41BG-6&R3+.*1*+*ive experiment consisting of seven consecutive steps for the transfer of a peptide solution from one polypropylene reaction vial (0.5 mL) to another followed by LC/MS quantification [112]. The experiment demonstrated that sample losses could be minimized by adding of acetonitrile. Losses become negli-gible at higher analyte concentrations. However, relative recovery might drop to 90% already after four transfer steps. Therefore, adsorption phe-nomena must be considered when developing quantitative procedures also identifying low concentration compounds.

41
Some of the biological fluids contain low protein/peptide concen-tration, for example urine, saliva, tears, etc. Therefore, preconcentration and removal of non proteinaceous solid materials and ions prior to the separation of proteins is essentially required. Several preconcentration methods have been reported, among the most common methods being dialysis and lyophilisation, filtration, ultracentrifugation, and precipitation. Protein losses were much higher in ultrafiltration and in the dialysis-lyophilisation method as compared to a precipitation method.
In this study, visual inspection of AF samples was done, and cen-trifugation at least at 2000 rpm was performed for 10 minutes only if a particulate matter was noted. Centrifugation is better than filtration, because there is minimal loss of sample, and it is cost saving (since disposable supplies are not used). Filtration through a 0.22 µm membrane filter eli-minates bacteria’s, therefore increasing sample stability and prevents from accidental pollution with microorganism proteome. However, the right filter must be chosen as filter holder also the filter membrane might exhibit some adsorption capability with certain capacity towards some analyte molecules. As adsorption might vary depending on molecules, after careless filtration the analyte components relative abundances could be shifted. This effect cannot be fully eliminated, but after some trial and error the most acceptable filters could be selected. In general borosilicate and cellulose containing filters should be avoided as they possess high protein binding properties. Polysulphonate and Durapore (EMD Millipore, USA) membranes are useful for many sample clean-up applications. Practically we advise to direct a few first droplets or up to 0.5 ml of the filtrated solution to waste as first portions of filtrate will face the highest losses at protein level even at minimal adsorption. The sample volume should match the filter size in order to keep surface adsorption capacity at minimum.
4.6.3.2. Sample extraction
Proteins are found in different cell compartments (cytoplasm, a range of intracellular organelles) or as secreted extracellular proteins in various body fluids. Furthermore, proteins range from highly soluble hydrophilic proteins, to membrane associated and trans-membrane proteins containing multiple hydrophobic domains. Moreover, proteins often exist as multi-subunit complexes or can form large macroscopic complexes with other proteins. No optimized conditions for sample clean-up exist so far to suit a wide range of physical and chemical properties of proteins. It would be

42
naive to believe that all cellular proteins can be solubilized and extracted efficiently and that such extraction can be reproducibly repeated for many different samples. This means that protein composition of two different tissues (for example liver and brain) cannot be compared even if suitable affinity assays were available, since inherent variability at the protein extraction step will make it impossible.
4.6.3.3. Most abundant component depletion
Most biofluids contain large amounts of well-known proteins such as albumin and IgGs, which overwhelm the separation system and make the detection of the low abundant proteins and peptides very difficult. It is therefore advantageous to remove these proteins prior to digestion and separation by specific adsorption of albumin and IgG to affinity matrices. While usually an affinity matrix is generally highly specific, in high content samples the affinity ligand is limited to exhibit its specificity. There are degrees of specificity between highly selective immunoaffinity matrices and less selective but more robust affinity supports using synthetic ligands. In an effort to reduce the amount of albumin from human serum, a number of affinity matrices have been evaluated based on antibodies or dye ligands. Antibody-mediated albumin removal was efficient and selective. Dye ligand chromatography, a technique that is extensively used in protein chroma-tography was surprisingly effective in particular with regard to high binding capacities and a long column lifetime, however, at the expense of selectivity [113]. The problem related to the depletion is the nature of proteins and peptides: they often form complexes. As the most abundant species are adsorbed as complexes, many interesting and wanted components are lost or their concentrations are reduced with different ratios. Also this heavily depends on the experimental conditions (buffers, pH, temperature etc.).
4.6.3.4. Affinity enrichment approaches
Often some classes of target molecules are present in very small amounts and need to be selectively isolated or enriched before identifica-tion. Affinity chromatography, which selectively retains proteins or peptides based on biospecific interactions, has been employed in several multidi-mensional separation methods for selective trapping of proteins of interest. Immobilized metal-affinity chromatography (IMAC) have reported as on-line combination of IMAC-Fe3+ and RP chromatography for identifying

43
phosphorylated peptides from 2D peptide maps or synthetic peptide mixtures [114].
One of the most published multidimensional separation techniques that employ an affinity chromatography step is based on the use of isotope-coded affinity tags (ICAT) [115]. In this strategy, cystein residues within proteins are modified with a thiol-reactive reagent that contains a biotin moiety. The proteins are enzymatically digested and the modified peptides recovered using immobilized avidin chromatography. The main purpose of affinity chromatography isolating only the cystein containing peptides is to reduce the complexity of the sample. However, since the ICAT-based strategy is designed for global proteomic studies, the post-affinity chromatography sample is still quite complex.
4.6.3.5. On line sample clean up
Direct injection techniques are generally preferable, since problems involved in off-line sample pretreatments, such as time consuming proce-dures, errors and risk for low recoveries can be readily avoided. Direct analysis of biofluids (Fig. 4.6.2.1, D) without prior digestion is a definitive option in biomarker discovery peptidomics. Those routes could be accom-plished by employing restricted access materials (RAM). RAM columns possess a dual function: firstly, they operate as size-exclusion columns to remove high molecular weight proteins and other undesired constituents. The term RAM is a general term for a packing material having a hydro-phobic interior covered by a hydrophilic barrier. The hydrophilic barrier allows passage of small molecules to the hydrophobic inner core of the stationary phase, while sterically preventing large molecules, such as pro-teins, from interacting with this part of the stationary phase. Macromo-lecules are excluded and may interact only with the outer surface of the particle support coated with hydrophilic groups, which minimizes the adsorption of matrix proteins. The size characteristics of proteins in pure size exclusion liquid chromatography (SEC) are known to be highly dependent on eluent composition such as pH, ionic strength (I) of the buffer (which includes salt type and concentration) and on the flow-rate [116]. Ionic strength and pH, however, can vary significantly among biofluids such as plasma and urine. The consequence will be that the sample cleanup procedures have to be adjusted individually with respect to each type of biological sample and standardized protocols have to be worked out.

44
Secondly, the RAM column serves as trap or capture column to selectively enrich target compounds in a reversed phase mode or in an ion-exchange mode. By regulating the pore size of the particles, the molecular weight exclusion can be varied as well as the molecular weight fractionation range, which allows certain analytes to be trapped at the internal surface. In this case, only proteins and peptides below a certain molecular shape and size have access to the inner pore surface of the RAM, are thus retained while the larger proteins encounter only the hydrophilic, non-adsorptive outer surface, and will be flushed out in the following washing step. Of the RAM, the strong cation exchanger with sulphonic ligands (RAM-SCX) was preferably employed in the sample clean up of proteins, which proved to show an acceptable capacity towards positively charged peptides and proteins. The features described above, when elegantly combined with column switching, become a powerful tool for direct analysis in the pro-filing of endogeneous peptides in a fully automated, multidimensional LC platform. RAM supports were able to withstand several hundred plasma or serum injections (total volume of 6–7 ml) without losing performance. The concept and the methodology were successfully used for the sample clean-up of peptides and proteins out of biofluids by extending the range of available materials employing cation and anion-exchanger RAM [117].
Mass loadability of solid-phase extraction (SPE) and RAM columns play a key role in executing the sample clean-up. It is advisable to work below the overload regime of the column. Otherwise, displacement effects and other phenomena such as secondary interaction by adsorbed species might take place, which will lead to non-reproducible results [118]. Last statement is particularly important when the task is to monitor medium to low abundant proteins. Therefore, usually large sample volumes in the millilitre range are applied. As the column lifetime is known to be limited a control measure has to be applied to check the condition of the RAM-SCX column and, if necessary, replace it by a new one.
RAM columns are currently used in many on-line solid phase ex-traction applications. Although the compatibility of these stationary phases with direct biological sample injection is high, one still has to keep in mind that samples have to be filtrated or centrifuged prior to injection to remove the solid contaminants and precipitations. Even so, some components tend to agglomerate/ precipitate with the time while samples queue up in auto sampler. Therefore, an additional in-line filter is highly recommended.

45
4.6.3.6. Designing a multidimensional LC system
Multidimensional (MD) chromatography had been discovered early as a powerful tool to separate complex mixtures. Two of the protagonists were ,%O%& P1001.?4& +.0& ,%D%S%& >3T6" [119]. MD LC is based on coupling co-lumns in an on-line or off-line mode, which are operated in an orthogonal mode, i.e. separate the sample mixture by different separation mechanisms. The sample separated on the first column (first dimension) is separated into fractions which can then be further treated independently of each other. The practical consequence is an enormous gain in peak capacity (number of peaks resolved at a given resolution) and the potential of independent optimization of the separation conditions for each fraction. Simultaneously, there is the option of relative enrichment/depletion and peak compression by fractionation. Multidimensional LC separation typically relies on utilizing two or more independent physical properties of the peptides to fractionate the mixture into individual components. Physical properties commonly exploited include are size, shape, charge, hydrophobicity and biomimetic or affinity interactions. These processes are the underlying phenomena for peptide/protein separations using different chromatographic modes, such as size exclusion, reversed phase, cation/anion exchange and hydrophobic in-teraction columns.
The primary criteria for the choice of a separation phase system are selectivity and orthogonality, mass loadability, and biocompatibility (in case of quantitation). As a rule of thumb, the first dimension should possess a high mass loadability (sample capacity) combined with sufficient selectivity and maintenance of bioactivity. Ion exchange chromatography (IEC) therefore is the method of choice offering charge selectivity. In principle, there are two options in IEC, either to employ a cation or anion exchanger, which in return influences the pH working range. Note that either cationic or anionic species are resolved, i.e. only a limited number of species from the whole spectrum. The IEC columns are operated via salt gradients with in-creasing ionic strength. Consequently, the salt load must be removed before the fractions are transferred to the second dimension column. It is most common to use reversed phase chromatography as the second dimension. The term RP stands for a number of columns with different degrees of hydrophobicity. The most commonly applied phases are n-octadecyl bonded silicas (RP-18 columns). An intrinsic feature of RP columns is their desalting property. Salts are eluted at the front of the chromatogram, when running a gradient elution with an acidic buffer/acetonitrile mobile phase

46
with increasing acetonitrile content. The hydrophobic surface of the RP packing and the hydrophobic eluent are not favorable with respect to providing a biocompatible environment for proteins: they may change their conformation or denature which may be seen by the appearance of broad peaks, splitting of peaks etc. RP columns possess a much lower mass loada-bility than IEC columns (10 mg of protein per gram of packing as compared to 100 mg in IEC). An advantage of RP is the fact that the eluents are compatible to MS, provided volatile buffers such as ammonium acetate are employed. In case of an on-line MD LC system, the speed of analysis in the second dimension should be as high as possible. This, however, conflicts with the requirement of high resolution or high peak capacity. The highest peak capacity in gradient elution RPC is obtained with a shallow gradient at relatively low flow-rate. Thus, a compromise between the desired peak capacity and the gradient time is inevitable. Often gradient times of several hours are applied for the analysis of peptides from protein digests. A question often arising is: how many dimensions do we need in MD LC? It becomes obvious that as the number of dimensions increases, the peak capacity will increase. In an ideal case the total peak capacity of the MD LC system is equal to the product of the individual orthogonal dimensions. At the same time, above two dimensions an on-line MD LC system becomes very sophisticated in its instrumental setup and may be difficult to control. The major goal in proteomics for the common user is to design a highly efficient, error minimizing and easy-to-handle system. Reduction of the system complexity is the major demand. It is essential to select a minimum number of dimensions to handle complex separations.
4.6.3.7. Mass spectrometry
Mass spectrometry (MS) is a high-throughput and reliable method for protein identification. It provides the ability to accurately measure the mass of almost any molecule that can be ionized in the gas phase. A mass spec-trometer consists of three essential components: an ion source, a mass analyzer and a detector. An ion source converts molecules into gas-phase ions. Once these ions are created, they are separated in the mass analyzer by their mass (m) to charge (z) ratio and detected by an electron multiplier. MS data are recorded as 'spectra' which display ion intensities versus their m/z value. MS allows ionization of macromolecules without destroying their chemical entity.

47
4.7. Preterm premature rupture of membranes
4.7.1. Definitions and incidence
Premature rupture of membranes (PROM) – is defined as spontaneous
membrane rupture that occurs at least one hour before the onset of labor. When spontaneous rupture of membranes occurs before 37 weeks of gestation, it is termed as preterm PROM (PPROM).
According to the Lithuanian Medical Data of Births 1616 (5.6%) new-borns were born prematurely in Lithuania in 2011; 557 of them were born in Hospital of Lithuanian University of Health Sciences [120]. PPROM is the one of the leading causes of premature birth and is responsible for approxi-mately one third of all preterm births. About 30–35% of preterm births are indicated, 40–45% follow spontaneous preterm labor, and 25–30% follow PPROM [121]. Births that follow spontaneous labor and PPROM are together designated spontaneous preterm births [122]. Over the past several decades, for reasons that are not fully understood, the preterm delivery (PTD) rate has raised worldwide. Much of the increase in the singleton PTD rate is explained by rising numbers of indicated preterm births [123]. A high number of preterm multiple gestations associated with assisted reproductive technologies is also an important contributor to the overall increase in preterm births. Singleton pregnancies after in vitro fertilisation are also at increased risk of preterm birth [124]. However, in Lithuania, the rate of PTD has stayed stable and has not increased since 1995, being 5.3–5.9% in recent years.
Most women with PPROM begin labour spontaneously within several days, but a small proportion of women remains undelivered for weeks or months. The term “latency” refers to the time from membrane rupture to delivery. „Prolonged PROM“ refers to PROM greater than 24 hours and „conservative” management is defined as treatment directed at continuing the pregnancy. Since the membranes generally form a barrier to ascending infection, a common complication of PPROM is development of intraute-rine infection and preterm labour. The risk of these complications increases with decreasing gestational age at membrane rupture. The major cause of perinatal morbidity and mortality associated with PPROM is prematurity. It is often convenient to divide preterm delivery into subgroups according to gestational age. Extreme prematurity occurs at 22 weeks to 28 weeks, appearing in about 5% of PTD, about 15% represent severe prematurity at 28–31 weeks, about 20% represent moderate prematurity at 32–33 weeks

48
and 60–70% represent late or near term prematurity at 34–37 weeks [122]. There is no global lower limit for PTD, but 22+0 completed weeks of gestation is generally accepted, corresponding to an average fetal weight of 500 g [125]. The same limit is used in Lithuania. Morbidities related to prematurity include respiratory distress syndrome, necrotizing enterocolitis, interventricular hemorrhage, cerebral palsy, and sepsis [126]. Other com-plications include in utero umbilical cord compression, cord prolapse and fetal distress, fetal malpresentation, placental abruption, chorioamnionitis with subsequent endometritis, and risk of operative delivery from this multitude of factors. Maternal sepsis is a rare but life-threatening compli-cation reported in nearly 1% of cases.
4.7.2. Risk factors
Risk factors for PPROM are generally similar to those for PTD with intact membranes, although infections and tobacco exposure play important parts [127]. There are many maternal or fetal risk factors that have been associated with PTD and PPROM, including maternal demographic cha-racteristics, nutritional status, pregnancy history, present pregnancy charac-teristics, psychological characteristics, adverse behaviours, infection, uterine contractions and cervical length, biological and genetic markers. Although many risk factors are well known, the cause of membrane rupture in most cases remains often unexplained; therefore, it is thought the etiology of PPROM is multifactorial. More than half of all preterm deliveries and PPROM occur in apparently low risk pregnancies with no major risk factor and some women are more prone than others.
4.7.2.1. Genetic factors
Women with PTD have higher risk to subsequent PTD. Mercer and colleagues reported that women with previous preterm deliveries had an 3.3 – fold increased risk in their next pregnancy [128]. The risk of another preterm birth is inversely related to the gestional age of the previous preterm birth. Those with a history of PPROM have a 13.5% risk of preterm birth due to PPROM in a subsequent gestation (versus 4.1%, relative risk 3.3, p < 0.01) and a 14 – fold higher risk of PPROM at less than 28 weeks in the subsequent gestation (1.8% versus 0.13 %, p < 0.01) [129]. There is also a tendency for repeated PTD to occur at the same gestational age as the previous pregnancies. Genetics contributes to individual variations in the

49
length of gestation and the timing of birth. Studies in twins have noted that maternal genetics may account from 15 to 40% of preterm deliveries [130]. The familial link in rates of preterm birth reflects genetic influences too. Woman with a sister who has given birth to a preterm infant have an 80% higher risk to have a PTD [131]. Genetic differences between individuals i. e. gene polymorphisms, can result in variation to the timing of birth and in production and activity of proinflammatory proteins, causing inadequate immunologic response to infectious stimuli. Each mechanism of disease responsible for preterm labour and PPROM has the potential for a genetic component. Preterm premature rupture of membranes may soon be consi-dered a disease of the genome that is influenced by environmental and nutritional factors. The PPROM phenotype may be the result of interactions of environmental and genetic elements both of which are potentially identifiable in high risk patients. Although available data support the idea of exploring the human genome to identify genes that may influence the timing of birth, the challenge is to determine whose genome should be explored. Does the maternal or fetal genome influence the timing of birth? Does variation in the maternal or fetal genome, or the interaction between the two genomes, contribute to the risk of prematurity? The questions should be answered in future.
PTD rates among black women is twice as high, and the rate of recurrent preterm birth four times as high, as the rate among white women in the United States and United Kingdom [132]. Black women are also three to four times more likely to have a very early preterm birth than women from other racial or ethnic groups. The beginning of PTD differs by ethnic groups. Spontaneous preterm birth is most commonly caused by preterm labour in white women, but by PPROM in black women [133]. The lowest rates of PTD exist in East-Asians and Hispanics.
4.7.2.2. Maternal risk factors
Maternal demographic characteristics associated with preterm birth include low socioeconomic and educational status, low and high maternal ages, and single marital status [134]. The mechanisms by which the ma-ternal demographic characteristics are related to preterm birth are unknown. Smoking, alcohol or drugs abuse are independent risk factors of PTD. Smoking increases the risk of PTD 2 – fold. Nutritional status during pregnancy can be described by body mass index, nutritional intake. A low prepregnancy BMI is associated with a high risk of PTD, being overweight

50
is associated with PPROM. Persistent inflammation may be an explanation for reported association between PPROM and overweight [135]. Morever, overweight women are more likely to develop preeclampsia and diabetes, which increase the rate of induced PTD. Psychological and social stress or depression (2 – fold) increase the risk of PTD, whereas physical activity is not consistently related to the rate of PTD.
4.7.2.3. Pregancy associated risk factors
Infection is the most common single risk factor of PTD and PPROM. It is known that the relation between infection and PTD is not consistent throughout gestation. Spontaneous preterm deliveries that occur before the 30th week of gestation, have been strongly associated to intrauterine infection. The earlier the preterm birth the stronger the association. Infection is rare in late preterm deliveries.
Bacterial vaginosis, a condition characterized by a decrease in number of lactobacilli and overgrowth of several anaerobic or facultative bacteria, including Gardnerella vaginalis, Mycoplasma hominis and Ureaplasma urealyticum, has been shown to be present in up to 15–20% of pregnant women. It is asymptomatic in approximately 50% of women and can resolve spontaneously. However, bacterial vaginosis has been shown to increase the risk for spontaneous PTD and PPROM 2 – to 3 – fold.
Any systemic infections, such as asymptomatic bacteriuria, pyelo-nephfritis, appendicitis and pneumonia can trigger uteroplacental response leading to PPROM and PTD. Even periodontitis has been linked to PTD. Human immunodeficiency virus and tuberculosis infections globally pre-dispose women to spontaneous PTD and PPROM.
Multiple gestations, account for about 1.1 – 1.3% of pregnancies in Lithuania, carry a substantial risk of PTD and PPROM. Over the past years there was a great increase in the incidence of multiple deliveries, largely as a result of the use of assisted reprodution technologies. Most of them have spontaneous labour or PPROM before 37 weeks of gestation, others usully are indicated PTD because of maternal or fetal disorders.
In vitro fertilization (IVF) is strongly associated to moderately pre-term births. The preterm rate for singleton pregnancies after IVF is approxi-mately 12% in other countries, but we don‘t have accurate data in Lithuania yet.

51
Cervical insufficiency and uterine anomalies are strongly associated with mid-trimester abortion and PTD. Cervical loop electrosurgical excision procedure (LEEP) secondary to premalignant cervical disease can be assosiated to PTD, but the risk is not very high. An interpregnancy interval of less than 6 months confers greater than 2 – fold risk of PTD. A short interval decreases the opportunity to replenish the essential maternal nutrient stores which pregnancy consumes and to resolute the inflammatory status with the previous pregnancy. In some studies the induced abortions increase the risk of PTD due to cervical damage during termination of pregnancy but the data are contradictory.
4.7.3. Etiology and pathophysiology
Rupture of amniochorionic membrane may have several underlying
causes, although in most PPROM cases a precise mechanism cannot be established. The pathophysiology leading to PROM at term has been shown to be different from the pathophysiology leading to PPROM. At term, weakening of the membranes may result from physiologic changes com-bined with shearing forces induced by contractions [136]. Histologic studies of the site of membrane rupture at term have demonstrated a zone of altered morphology characterized by thickening of the connective tissue compo-nents of the membranes, thinning of the cytotrophoblast layer and decidua, and disruption of the connections between amnion and chorion. These normal physiologic changes accompany cervical ripening in preparation for labor at term, and result in focal weakening of the fetal membranes in the region of the internal cervical os that predisposes to rupture at that site. At a cellular level, these changes result from the release of phospholipases, eicosanoids (especially prostaglandin E2), cytokines, elastases, matrix metalloproteinases, and/or other proteases. Although the downstream cellular changes may be similar, the inciting etiologies in PPROM are likely different from term PROM [137].
PPROM etiology factors are similar to those for spontaneous preterm labor with intact membranes. Four different main pathways have been described which may lead to PPROM and PTD: infection and inflammation, decidual haemorrhage, activation of the maternal or fetal hypothalamic-pituitary-adrenal (HPA) axis, and uterine over-distension [138] (Fig. 4.7.3.1).

52
Fig. 4.7.3.1. Main pathways leading to PPROM and PTD
4.7.3.1. Infection and inflammation
Intrauterine infection is the most important mechanism leading to preterm delivery and PPROM [139]. It has been estimated that about 40% of all PPROMs occur in mothers with intrauterine infection (IUI). PPROM or preterm labor may be the first symptoms of IUI. Microorganisms have been isolated from the amniotic fluid of women who experienced preterm labour with or without PPROM, although the rates of positive cultures are higher in women who have PPROM (approximately 32.4%) than in those with preterm labour and intact membranes (approximately 12.8%) [140]. In women with PPROM, the most common microorganism isolated from AF is Ureaplasma urealyticum [141]. Other less commonly identified microor-ganisms are: Mycoplasma hominis, Chlamydia trachomatis, Streptococcus agalactiae, Streptococcus !"haemolyticus, Streptococcus pneumoniae, coa-gulase-negative Staphylococcus epidermidis, Staphylococcus auricularis, Staphylococcus hominis, Enterococcus spp., Enterococcus faecalis, Haemo-phylus influenzae, Fusobacterium, Lactobacillus spp., Bacillus spp., Esche-richia coli and Candida albicans [141]. In some cases, a polymicrobial infection in the amniotic fluid is detected. Microorganisms can gain access to the amniotic cavity by: ascending from the vagina and the cervix; haematogenous dissemination through the placenta; accidental introduction at the time of invasive procedures; and by retrograde spread through the fallopian tubes [142].

53
Infection can be associated with PPROM as either a cause or a consequence. Infection preceding PPROM is often subclinical and thought to ascend from the lower genital tract. Following rupture of the membranes, ascending bacterial invasion can lead to IUI in up to 60% of cases in the absence of antibacterial therapy [142].
Mechanisms of PPROM Due to Infection
The mechanisms by which intrauterine infections lead to preterm labour are related to activation of the innate immune system. The putative mechanism underlying infection and PPROM requires intrauterine bacterial invasion. Microorganisms in the vagina produce enzymes such as proteases and mucinases, which allow penetration of the cervical mucus plug, fa-vouring the ascent of bacteria. Cervical mucus, containing antimicrobial proteins and peptides such us lysozyme, lactoferrin, defensin and immu-noglobulins, acts like a mechanical and immunological barrier to ascending infection from the vagina. IUI activates the decidua and fetal membranes to produce pro-inflammatory cytokines, such as interleukins-1 (IL-UF&+.0& V9-UIWE&1.*6"-6351.-6 and 8 (IL-6, IL-8) and tumour necrosis factor-F&8J/D-FW%&This in turn leads to the release of prostaglandins, metalloproteases, and other bioactive substances. The prostaglandins stimulate uterine contrac-tions, and matrix metalloproteases cause proteolysis in the extracellular matrix and target the amniochorionic membranes by weakening them, leading to rupture. In addition, matrix metalloproteinases cause tissue dis-ruption in the choriodecidual space and facilitate separation of the chorion from the decidual layer in the lower uterine segment as well as ripening of the cervix by remodelling the collagen fibres. Tissue disruption in the lower segment of the uterus due to either uterine contractions or infection-induced proteolysis, may cause leakage of choriodecidual products, such as insulin-like growth factor-binding protein-1, fetal fibronectin, and others into the cervix and vagina, where they become detectable.
4.7.3.2. Activation of the maternal or fetal hypothalamic-pituitary-adrenal axis
Stress, psychological or physical, is one of the main independent risk
factors leading to PTD, accounting for about 30% of cases. This pathway is typically associated with PTL after 32–34 weeks. Rare in PPROM cases.

54
4.7.3.3. Decidual haemorrhage (abruption)
This pathway is involved in about 20% of PTD, especially those related to PPROM. Vaginal bleeding caused by decidual haemorrhage or placental abruption is associated with a 3 – fold increased adjusted relative risk of PTD. Harger et al. observed that vaginal bleeding in more than one trimester carried the highest identifiable risk of PPROM with an odds ratio of 7.4 [143]. The biochemical pathway from decidual bleeding to PTD is closely related to thrombin generation. Tissue factor is the primary initiator of hemostasis and is found in large quantities in the decidua. After hemor-rhage, tissue factor forms a complex with activated factor VII to activate factor X, which then generates thrombin production. Thrombin acts to stimulate coagulation and clot formation; however, it also stimulates the production of proteases (plasminogen activators) capable of ripening the cervix and damaging fetal membranes, leading to PPROM. Thrombin may also exert an indirect uterotonic effect on the myometrium and stimulate contractions.
4.7.3.4. Uterine over-distension
Intraamniotic pressure remains relatively constant throughout gesta-
tion despite the growth of the fetus and placenta. Premature delivery may be precipitated by mechanical stretching of the myometrium caused by an increase in uterine size that exceeds the ability of the uterus to compensate. Uterine over-distension is caused by multi-fetal pregnancy, polyhydram-nion, or structural uterine anomalies. Over-distension or decreased expan-sive capacity of the uterus leads to activation of the cytokine network and/or activation of the myometrium, leading to uterine contractions, cervical changes and/or rupture of the fetal membranes.
4.7.4. Diagnosis
Preterm PROM is largely a clinical diagnosis. It is typically suggested
by a history of watery vaginal discharge and confirmed on sterile speculum examination. The traditional minimally invasive gold standard for the diagnosis of rupture of membranes relies on ability to document clinical signs on sterile speculum examination: ! visual pooling of clear fluid in the posterior fornix of the vagina or
leakage of fluid from the cervical os;

55
! the nitrazine test – an alkaline pH of the cervicovaginal secretions (the pH of the vaginal secretions is generally 4.5–6.0, whereas amniotic fluid usually has a pH of 7.1–7.3);
! the fern test – microscopic crystallization of amniotic fluid on drying from the cervicovaginal discharge;
! evidence of diminished amniotic fluid volume by ultrasound alone cannot confirm the diagnosis, but may help to suggest it.
The nitrazine test is the most common test used to diagnose PPROM.
It is associated with high false-positive rates related to cervicitis, vaginitis (bacterial vaginosis), and contamination with blood, urine, semen, or antiseptic agents. The sensitivity and specificity of this test in diagnosing rupture of membranes are 90.2% and 79.3%. The fern test may give false-positive results due to fingerprints or contamination with semen and cervical mucus as well as false negative results due to technical error (using a dry swab to collect the sample) or contamination with blood. Reported sensitivity and specificity for the fern test are 90.8% and 95.3%, respec-tively. Ultrasound amniotic fluid index has a sensitivity and specificity of 94.0% and 91.0% with 6.2% false negative and 9.0% false positive results [144]. Therefore, the diagnosis of PROM is difficult if there is a slow fluid leak or any bleeding, or when the classic “gush of fluid” does not occur. In addition, the relatively small amount of amniotic fluid present early in ges-tation further challenges the diagnosis of ruptured membranes. A patient’s history may suggest membrane rupture, but test results are non-confir-matory, creating an obstetrical dilemma. Early and accurate diagnosis of membrane rupture would allow for gestational age-specific interventions to optimize perinatal outcome and minimize serious complications [145].
Investigators have long been searching for an alternative and more objective test. Analysis of amniotic fluid and serum from women with PPROM has been undertaken to identify biomarkers for PPROM. Although initial studies may be encouraging, diagnostic techniques are usually found subsequently to be limited by inaccuracies from false positives and false negatives with poorer sensitivities and specificities than originally antici-pated (Table 4.7.4.1). The ideal test should be simple, rapid, inexpensive, and non-invasive. Optimally, the accuracy of the test should not be ham-pered by the presence of blood, semen, infected urine, or other contami-nants. An accurate biomarker for membrane rupture should have a high concentration in the amniotic fluid, a low concentration in maternal blood,

56
and an extremely low background concentration in cervico-vaginal dis-charge with intact membranes [146]. The review of proteomic ond other diagnostic methods of PROM is reported by El-Messidi et al. [147]. Table 4.7.4.1. The review of available proteomic tests for PROM diagnosis
Test
Sampl-
ing
site
Discrimi-
natory con-
centration
Sensitivity,%
Mean
(range)
Specificity,%
Mean
(range)
FN, %
Mean
(range)
FP,%
Mean
(range)
PAMG-1 (AmniSure)
vagina 5 ng/ml 98.8 98.7–98.9
93.8 87.5–100
1.2 1.1–1.3
6.3 0–12.5
IGFBP-1 (ActimProm)
vagina, cervix
35 X?Y- 84.9 74.4–100
92.8 71.4–98.2
14.7 0–39.2
7.3 1.8–28.6
fFN vagina 50 ng/ml 93.6 90–98.2
65.7 26.8–97.0
13.1 3.0–25.0
17.0 6.0–28.0
I:OP vagina 46.4 mg/ml 83.9 68.0–100
89.5 84.2–95.0
9.1 0–25.0
10.5 5.0–16.0
Prolactin vagina 2ng/mL 80 76.0–100
85 70.0–100
68.0 15.0 0–30.0
AFP vagina cervix
Z[%[&X?Y9 \\%Z&X?Y9
92.0 84.0–100
86.9 85.7–88.0
8.0 0–16.0
13.2 12.0–14.3
FN: false negative; FP: false positive; AFP, alpha-#$%&'(&%$)*+",-hCG: beta-subunit of human chorionic gonadotropin; IGFBP-1: insulin like growth factor binding protein 1; PAMG-1: placental alpha-microglobulin 1; fFN: fetal fibronectin.
Placental alpha-microglobulin 1 (PAMG-1) is the most accurate test in diagnosing of rupture of the membranes and have the highest sensitivity, specificity, positive and negative predictive values at the moment [148]. This 34 kDa placental glycoprotein is abundant in amniotic fluid (2000–25,000 ng/mL), with much lower concentrations in maternal blood (5–25 ng/mL). The protein is in even lower concentrations in cervicovaginal fluid in the absence of ruptured membranes (0.05–0.2 ng/mL). This 1000 – to 10,000-fold difference in concentration between amniotic fluid and cervi-covaginal secretions makes PAMG-1 a very attractive biomarker for PROM. Thus, proteomic techniques can be used to identify biomarkers in women with premature labour and PPROM.

57
4.7.5. Prediction of preterm premature rupture of membranes
Interventions to reduce the morbidity and mortality related to preterm
birth and PPROM can be classified as primary (directed to all women before or during pregnancy to prevent and reduce risk), secondary (aimed at eli-minating or reducing risk in women with known risk factors), or tertiary (initiated after PPROM has occurred, aimed at preventing neonatal compli-cations associated with prematurity) [149].
Most obstetric interventions to reduce the morbidity and mortality of PPROM are classified as tertiary – e.g., regionalized perinatal care, treat-ment with antenatal corticosteroids, tocolytic agents, and antibiotics. These measures are intended to reduce the burden of prematurity-related illness more than to reduce the rate of PPROM and PTD.
Secondary prevention requires identification and reduction of risk, both of which have proved difficult. Epidemiological and clinical studies have identified a number of factors associated with increased risk for PPROM. Women with a previous history of PPROM have a 3.3 – fold increased risk of preterm birth caused by PPROM and a 14 – fold higher risk of PPROM before 28 weeks in a subsequent pregnancy. Bacterial vagi-nosis has been established as being associated with preterm birth and PPROM, it is unclear if this is a cause – effect relationship or if bacterial vaginosis only identifies those at risk for infection and inflammation. A short cervix (< 25 mm) by transvaginal ultrasonography has been associated with PPROM in both nulliparas and multiparas [150], and a positive fetal fibronectin screen has also been associated with PPROM in multiparas [151]. Nulliparas with a positive cervicovaginal fetal fibronectin and a short cervix have been found to have a one-in-six risk (16.7%) of preterm birth caused by PPROM, whereas multiparas with a previous history, a short cervix, and a positive fetal fibronectin have a 31 – fold higher risk of PPROM with delivery before 35 weeks than those without risk factors (25 vs. 0.8%, p = 0.001). However, the current methods of predicting women are expensive, inconvenient and non-specific to be of real clinical value. The majority (> 50%) of those suffering preterm premature rupture of the membrane lack risk factors that might lead to preventative treatments.
Primary prevention is prevention of the onset of PPROM in asymp-tomatic women by cessation of smoking and/or alcohol use, by maintaining a healthy genitourinary tract and reducing body mass index to normal.
It is likely that biological markers exists that could be of clinical value as the evident increase that many of the events leading to PPROM are more

58
likely to be chronic than acute. Therefore, it would be of clinical importance if this condition could be predicted before it becomes manifest clinically, as spontaneous PPROM. Before systematic screening, effective prophylactic intervention or treatment for individuals with positive test results should be available. In addition, the test should be efficient, with high sensitivity, specificity and a high positive predictive value, and be of low cost. At present, despite all efforts, there are no accurate screening methods. Primary or secondary interventions to prevent PTD or PPROM have also been largely unsuccessful. If an accurate marker of threatened PPROM would be available, it could be used to tailor intervention among individual women at risk. Therefore there is a need to apply the newest methodologies to search and identify these biomarkers. Once the biomarkers are confirmed, new biological tests later could be developed which would help to achieve accurate prognosis, diagnosis and maybe treatment to prevent that process.

59
5. PATIENTS, BIOLOGICAL SAMPLES AND
METHODOLOGY
The case – control study was carried out at the Department of Obstetrics and Gynecology of Hospital of the Lithuanian University of Health Sciences (HLUHS), Kaunas, Lithuania. HLUHS is a referral tertiary care center where patients with preterm deliveries are transferred from about two thirds of Lithuania.
5.1. Patients
The enrollment of the participants in the study was carried out from *:6&T6?1..1.?&#$&,3.6&*#&*:6&6.0&#$&December, 2008.
The inclusion criteria were: 1) prenatal genetic amniocentesis at 16 - 18 weeks of gestation or 2) admission due to preterm premature rupture of membranes at 22–36 weeks of gestation or 3) cesarean section for breech presentation at term 8]&[\&H6654W.
The exclusion criteria were:
1) the refusal to participate in the study; 2) age < 18 years; 3) clinical and/or laboratory diagnosis of infection; 4) chronic or acute illness, usage of antibiotics or medication at any time during pregnancy (with exception of vitamins); 5) any pregnancy complications; 6) dysplasia or abnormal kariotype of the fetus/newborn; 7) multiply pregnancy; 8) stillbirth; 9) contamination of amniotic fluid sample with blood.
Information was obtained by interview and from the patient clinical
case reports. Maternal characteristics and obstetrical information were estimated (age, nationality, residence, education, smoking status, alcohol intake, history of chronic or acute diseases, and use of medications, parity, information about previous and ongoing pregnancy, delivery, maternal and neonatal outcome).

60
The gestational age was determined from the last menstrual period if it agreed with an ultrasound estimation within 7 days in the second trimester; otherwise, the pregnancy was dated based on ultrasound data.
The patients with PPROM admitted to the Department of Obstetrics and Gynecology of HLUHS were as a target group of participants in the study. Patients with early and term pregnancies were as controls, involved in the research because the fetal membranes were intact.
Patients with PPROM were included in the study when the diagnosis of PPROM was suggested by subjective complaints of loss of fluid from the vagina and confirmed on sterile speculum examination by visual pooling of clear fluid in the posterior fornix of the vagina or leakage of fluid from the cervical os, a positive nitrazine test (pH > 7.0 of the cervicovaginal fluid) and/or ferning test (microscopic crystallization of amniotic fluid on drying from the cervicovaginal discharge) and/or decreased amniotic fluid index (AFI) less than 80 mm on ultrasonography.
Whereas the goal was to investigate amniotic fluid peptidome and proteome of PPROM patients without infection, clinical and laboratory tests were performed to exclude infection under the local protocol: ! maternal temperature and heart rate, ! fetal heart rate on CTG, ! blood test (white blood cell count) and C-reactive protein (CRP), ! urinanalysis and urine microbiological culture, ! swab for group B streptococcus culture from lower vagina and peri-neal area, ! additionaly to protocol, an amniocentesis and amniotic fluid micro-biological culture, ! after delivery, histopathology of placenta and fetal membranes.
Patients with PPROM were followed after enrollment until delivery.
After amniocentesis and amniotic fluid sampling all patients were monitored and treated according to the local protocol (all patients recieved antibiotics – penicillin intravenously; an intramuscular dexamethason administration, if gestational age was <34 weeks; and nifedipine was used for tocolysis until the lung maturation, if it was needed). All patients were monitored with serial ultrasound examinations to evaluate growth of the fetus and AFI. Maternal (latency period, mode of delivery, infectious complications) and neonatal (Apgar scors in 1 and 5 min, umbilical arterial pH, wheight, height, admission to NICU, congenital infections) outcomes were estimated. All cases were excluded when infection was suspected or diagnosed.

61
5.2. Biological samples
For the research amniotic fluid, maternal plasma and fetal membranes samples were used.
Amniotic fluid samples
Amniotic fluid samples were retrieved by trans-abdominal amniocen-
tesis (AC) in the absence of maternal blood contamination. Amniocentesis is an invasive method that is used to obtain amniotic fluid samples. In this study, amniotic fluid was retrieved by maternal-fetal medicine specialists using trans-abdominal amniocentesis under ultrasound guidance following guidelines and applicable local regulatory requirements.
30 amniotic fluid samples were obtained during prenatal genetic counceling. Prenatal genetic amniocentesis is specific for gestational ages of 16–18 weeks. This period was chosen because most amniocenteses are done during this gestational age. By cytogenetic analysis it was shown that all women carried chromosomally normal fetuses, the pregnancies went on without complications and ended with deliveries of healthy newborns.
The same trans-abdominal amniocentesis approch was used for the patients with PPROM. 35 patients were admitted with this diagnosis and fulfilled the study criteria, but for 19 patients the amniocentesis was not performed. The predominant reasons for not performing an amniocentesis were patient refusal, technical difficulties because of small amount of am-niotic fluid and provider discomfort. 16 patients of those offered an amnio-centesis underwent the procedure. The amniotic fluid samples were taken at the time of inpatient admission from patients with spontaneous PPROM. The intraamniotic infection was ruled out by the negative microbiological amniotic fluid culture for aerobic and anaerobic bacteria. 16 amniotic fluid samples were retrieved from the largest amniotic fluid pocket. Six patients were excluded from the study because one amniotic fluid sample was con-taminated with maternal blood, one sample was with insufficient amniotic fluid amount, one patient had positive amniotic fluid culture for several microorganisms, one patient had positive urine culture for E. coli, placental samples from two patients were with histological chorioamnionitis.
At term 16 amniotic fluid samples were obtained by trans-amniotic puncture after extra-membranous hysterotomy from women with obstetrical indication of cesarean section.

62
9 ml of pure clear amniotic fluid sample has been withdrawn to empty tubes (BD Vacutainer® Tubes) each time. The samples were stored at –72°C until further processing.
The amniotic fluid samples were divided into three groups according to the gestational age (Table 5.2.1). Table 5.2.1. Groups of the study
The name of the
group
Gestational
age
Number of
samples
Characterization of the
group
1st group Geltonoji (G) – yellow
16–18 week 30 Early pregnancies with intact membranes
2nd group Raudonoji (R) – red
30–36 week 10 Preterm premature rupture of membranes
3rd group -./)&0)"(Z) – green 38–40 week 16 Term pregnancies with intact membranes
Maternal plasma samples
Blood samples as well as amniotic fluid samples were taken at the time of inpatient admission from patients with spontaneous PPROM. Within few hours the samples were centrifuged at 3000×g for 10 min and the supernatant was removed and stored at –72°C until further processing.
Fetal membranes samples
The amniochorionic membranes were obtained from a healthy woman with spontaneous PPROM at the 32nd week of gestation after spontaneous delivery. The intraamniotic infection was ruled out by negative microbio-logical amniotic fluid culture and normal placental histological examination. As control, it was used fetal membranes taken from a healthy patient who underwent cesarean section for breech presentation at 39 weeks of gestation with intact membranes and no signs of infection. Membrane samples were obtained within 10 min of delivery from the afterbirth and were cut (at the site of rupture in PPROM case) into 5×5 cm pieces and were stored at –72°C until further processing.

63
Digestion of fetal membranes
The fetal membranes were thoroughly washed with Milli Q synthesis
water and cut into 3×3 mm pieces, three of which were then transferred into the digestion vial. 500 µl of 50 mM ammonium bicarbonate (Sigma) and 10 µl dithiothreitol (DTT) (200 mM in water) (DTT – reduction of disulfide bonds) solution were added. The mixture was incubated at 37°C for 10 min. After storage of 10 min at room temperature in dark 10 µl of iodoacetamide (IAA) (400 mM in water) (IAA – alkylates cysteines) was added. Trypsin and chymotrypsin solutions were prepared as directed by the supplier and added to the protein mixture at a molar ratio of 1:40. The reaction was incubated in a water bath at 30°C for 24 hours. The digest mixture was centrifuged at least at 2000 rpm for 10 min and frozen at –72°C. Before analysis samples were defrosted and filtered through a 0.22 µm membrane filter (Millipore).
5.3. The analytical system
Proteomic studies on samples collected from women with PPROM (pairs of maternal plasma and AF) as well as samples of AF collected at the 16–18 and 38–^N& H6654& #$& ?64*+*1#.& H6"6& 3.06"*+56.& +*& *:6& ,#:+..64&Gutenberg University, Mainz, Germany, Department of Inorganic and Analytical Chemistry at the laboratory of Prof. K.K. Unger. Biological samples of amniotic fluid, plasma and fetal membranes were transported to the laboratory in dry ice packages by the priority post. The analytical system was previously successfully used for human urine studies [152,153] and was modified for amniotic fluid, plasma and digested amniochorionic membranes analysis. Detailed descriptions concerning sample preparation using restricted access columns (RAM) and the usage of monolithic silica columns at peptide separation in 2D LC/MS were reported previously [154,155].
The total analysis concept was based on a sample preparation and fully automated two-dimensional liquid chromatography (2D LC) system (HP1100, Agilent, Waldbronn, Germany): pre-separating the majority of the matrix components from the analytes which retained on a restricted access strong cation-exchange (RAM-SCX) column for selective, < 5 kDa peptide extraction from biological samples (RAM-SCX, Merck KGaA, Darmstadt, Germany) followed by a fraction transfer onto a reverse-phase (RP)

64
capillary column (Chromolith CapRod, Merck KGaA, Darmstadt, Germany) by means of column switching in a way to obtain two dimensional ortho-gonal separations. For sample clean-up prior to the loading step the RAM-SCX column was equilibrated with aqueous 20 mM potassium dihydrogen phosphate/phosphoric acid (pH 2.5) buffer. This eluent was also employed for the second dimension as a low ionic strength buffer. The loading of the RAM-SCX column was performed at a total flow rate of 0.4 ml/min, which consisted of the sample delivery flow and the main flow performing ten time sample dilution on-line. A step gradient was performed in two steps by mixing 20 mM potassium dihydrogen phosphate/phosphoric acid (pH 2.5) (eluent A1) and A1 with a 1.5 M sodium chloride (eluent B1) at the following proportions: 85/15 (V/V); 0/100 (V/V). A constant flow rate of 0.4 ml/min was applied after the switching to the second dimension where the fractions were trapped on two RP trap columns to fulfill an orthogonal separation. Next, the gradient micro HPLC system was started to deliver a gradient at a constant flow rate of 4 µl/min starting from 5% acetonitrile in water (V/V) with 0.2% formic acid (V/V) (eluent A2) adding to 50% of 5% water in acetonitrile (V/V) with 0.2% formic acid (V/V) (eluent B2) in 40 min. Proteinaceous constituents were washed from the trap columns and separated on a RP monolithic silica capillary column (Fig. 5.3.1).
On-line mass spectrometric detection of amniotic fluid and plasma samples was performed using an Esquire Series 3000 PLUS electrospray ionization (ESI) ion-trap MS (Bruker Daltonics, Bremen, Germany) and amniochorionic membranes were analyzed using Agilent 6500 Series accu-rate-mass quadrupole time-of-flight (Q-TOF) MS (Waldbronn, Germany). The settings of the mass spectrometer were as follows: scan mode: normal 13,000 m/z/s; mode: positive; dry temperature: 200°C; nebulizer: 5 psi; dry gas: 4 l/min, high voltage capillary: 4,000 V; trap target: 20,000, trap max accumulation time 200 ms; scan range 200–3,000 m/z; averages: 5, rolling averages: 2; auto MS/MS: 2; number of precursor ions: 2; double charge ions: preferred.
Before use the 2D LC/MS/MS system was validated. The Pearson correlation of 49 consecutive runs for a signal intensity of 24 selected peptides from digested bovine serum albumin eluting 24–26 min was 0.88. The retention time variability for the 582, 675, 720 and 1230 fragments were ± 25 seconds. For amniotic fluid samples the mean m/z value for 975 signals was 0.12 (SD ± 0.04), the mean retention time value for 975 signals was 0.30 (SD ± 0.05).
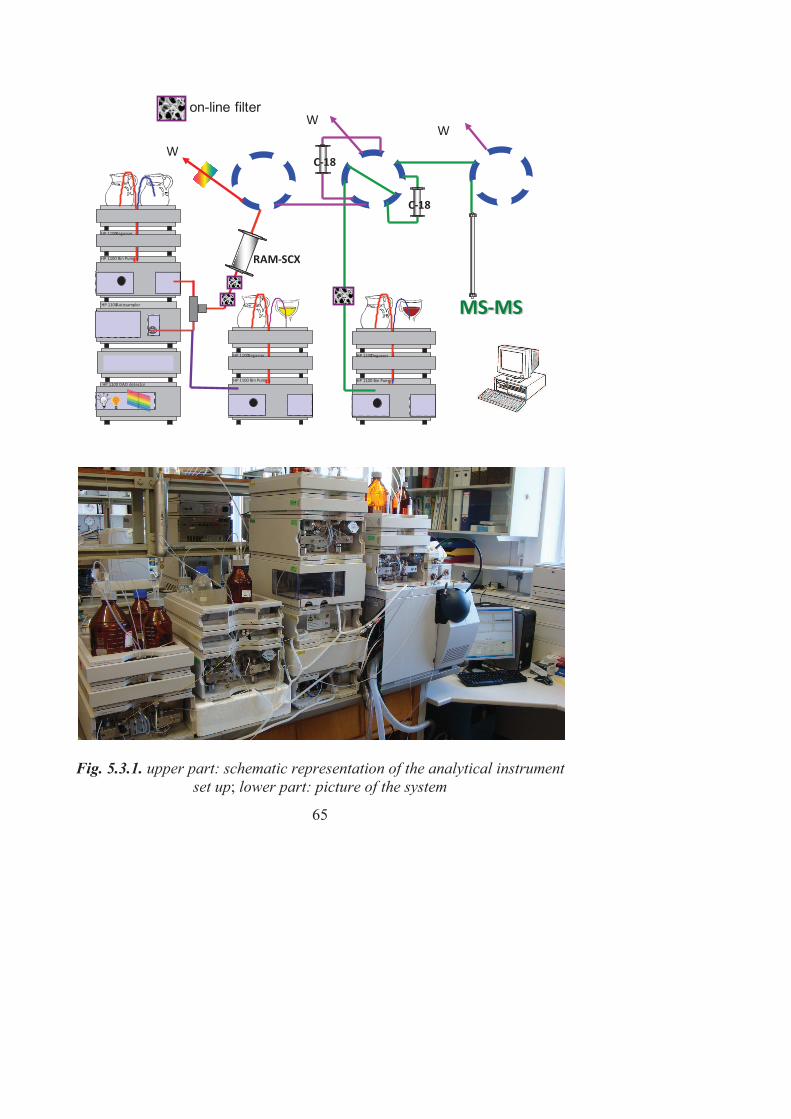
65
Fig. 5.3.1. upper part: schematic representation of the analytical instrument
set up; lower part: picture of the system
HP 1100 DAD detector
HP 1100 Degasser
HP 1100 Bin Pump
HP 1100 Autosampler
HP 1100 Degasser
HP 1100 Bin Pump
MS-MS
W
W
RAM-SCX
HP 1100 Degasser
HP 1100 Bin Pump
C-18
C-18
W
on-line filter

66
An automated peptide search was performed for all analyses using the following parameters: mz Radius = 0.3, TRadius = 0.6, Fraction = 0.25, mz Start = 500, rt Start = 50 min, mz End = 3,000, rt End =100 min.
A manual peptide search was performed for some selected analyses using the following parameters: typical peptidic signals (> triplets, mass difference: 1 – for single charged species, 0.5 – for double charged species, 0.3 – for triple charges spiecies), signal intensity min 5 times higher than the surrounding background noice, mz Start = 500, rt Start = 50 min, mz End = 3,000, rt End = 100 min.
5.4. Database searching and confidence of proteins identification
The resulting mass spectra from each sample were processed with a DataAnalysis 3.5 program (Bruker Daltonics) and submitted to a database search using BioTools (Bruker Daltonics). The Mascot engine was used to search the International Protein Index human database for peptide sequence identification. The proteins were identified from the UniProtKB/Swiss-Prot database.
The search of proteins from amniotic fluid was performed using the following parameters: (a) enzyme – none; (b) one missed cleavage allowed; (c) variable modification, oxidation of methionine; (d) peptide mass tolerance – 10 ppm; and (e) fragment mass tolerance – 0.3 Da.
The search of proteins from amniochorionic membranes was per-formed using the following parameters: (a) enzyme – trypsin, chymotrypsin; (b) one missed cleavage allowed; (c) variable modification, oxidation of methionine; (d) peptide mass tolerance – 10 ppm; and (e) fragment mass tolerance – 0.3 Da.
A Mascot score above 67 was used for positive protein identification (p < 0.05). Mascot compares the experimental data spectra to a data base of theoretical spectra and scores how well they match. The protein identifi-cation reports contain a list of proteins that have been positively identified by a significant Mascot score. The Mascot score for a protein is the summed score for all peptides matching a given protein. Mascot uses probability based scoring. This enables a simple rule to be used to judge whether a result is significant or not. A commonly accepted threshold is that an event is significant if it would be expected to occur at random with a frequency of less than 5%. This is the default value that is reported on the Mascot results summary page. The Protein Summary page for typical peptide mass finger-print search reports that “scores greater than 67 are significant” [156].

67
5.5. Ethical aspects
The research was approved by the Regional Biomedical Research
Ethics Committee of Kaunas (No. BE-2-3), Lithuania. The participants were provided with information about the aims, plan of the study and methods to be applied, which was made easily understandable to women. The study started after a written informed consent was obtained.
Conflicts of Interest statement
The author declares that there are no conflicts of interest.

68
6. RESULTS AND DISCUSSION
A total number of 56 women with singleton pregnancies were enrolled in the study. All women were healthy, free of any medical history and under no medication. Maternal demographics and characteristics are listed in the Table 6.1.
Table 6.1. Maternal demographics and characterictics (p value >0.05 for all variables)
G group
(n=30)
R group
(n=10)
Z group
(n=16)
Maternal age (years) Mean Median (range)
36.6 SD±2.28
36 (32-42)
30.7 SD±7.53
32 (19-43)
29.6 SD±4.33
29 (24-39)
Nationality* Lithuanian Other
28 (93.3%)
2 (6.7%)
10 (100%)
0
15 (93.8%)
1 (6.3%)
Residence* City Countryside
22 (73.3%) 8 (26.7%)
7 (70.0%) 3 (30.0%)
11 (68.8%) 5 (31.2%)
Marital status* Married Living with partner Single (never married, widowed, separated or divorced)
24 (80.0%) 4 (13.3%) 2 (6.7%)
7 (70.0%) 2 (20.0%) 1 (10.0%)
12 (75.05) 2 (12.5%) 2 (12.5%)
Education* University College Secondary Basic
15 (50.0%) 12 (40.0%)
2 (6.7%) 1 (3.3)
4 (40.0%) 1 (10.0%) 2 (20.0%) 3 (30.0%)
7 (43.8%) 7(43.8%) 1 (6.3%) 1 (6.3%)
Cigarette smoking* Non-smoking Smoking**
25 (83.3%) 5 (16.7%)
8 (80.0%) 2 (20.0%)
14 (87.5%) 2 (12.5%)
Alcohol ingestion during pregnancy* yes no
0 0
1 (10%)
0
0 0
Number of current delivery* 1 2 ][
7 (23.3%)
16 (53.3%) 7 (23.3%)
4 (40.0%) 3 (30.0%) 3 (30.0%)
11 (68.8%) 4 (25.0%) 1 (6.3%)

69
Table 6.1 continued
G group
(n=30)
R group
(n=10)
Z group
(n=16)
Previously having* Preterm delivery Miscarriage Abortion
2 (6.7%)
10 (33.3%) 5 (16.7%)
1 (10.0%)
0 5 (50.0%)
1 (6.3%) 4 (25%)
2 (12.5%)
Gestational age at amniocentesis (weeks) Mean Median (range)
17.2 SD±0.76
17 (16-18)
33.5 SD±1.8 33 (30-36)
38.7 SD±0.6 39 (38-40)
Gestational age at delivery (weeks) Mean Median (range)
39.1 SD±1.08
39 (37-41)
34.3 SD±1.16
34 (33-36)
38.7 SD±0.6 38 (38-40)
*Data presented in n (%). **All smoking patients smoked less than five cigarettes per day during pregnancy.
Maternal age mean varied among the G, R and Z groups. Maternal age mean in the group of women with prenatal genetic AC was 36.6 years, median 36 years (the range from 32 to 42 years); in the group of women with PPROM maternal age mean was 30.7 years, median 32 years (the range from 19 to 43 years); and in the group of women with cesarean section at term with intact membranes maternal age mean was 29.6 years, median 29 years (the range from 24 to 39 years). Most women were lithuanians (93.3–100%), living in the cities (68.8–73.3%), married (70.0–80.0%), and with the university education (43.8–50.0%) in all groups. During pregnancy about one fifth of the women (12.5–20.0%) smoked, all of them smoked less than 5 cigarettes per day. Only one women in the group with PPROM stated about alkohol ingestion during pregnancy. There was a difference in the number of current pregnancy among the groups. In the group of women with term secarean section and in the group with PPROM most pregnant women were primiparas (68.8% and 40.0%). In the group with prenatal genetic AC most of women were pregnant with the second child (53.3%). In the group of women with PPROM was increased rate of previous abortions (50.0%).
The mean total gestational age at delivery was 34.3, SD ± 1.16 in PPROM group; 39.1, SD ± 1.08 in prenatal genetic AC group and 38.7 weeks, SD ± 0.6 weeks in group of cesarean section at term with intact membranes. Mean total birth weight was 2572 g, SD ± 369 g in the PPROM group; 3522 g, SD ± 331g in prenatal genetic AC group and 3573 g, SD ± 337 g in group of cesarean section at term with intact membranes.

70
Table 6.2. Characteristics of the PPROM study
Case
GA at time of
PPROM and
amniocentesis
(weeks)
Latency
period
(weeks)
GA at
delivery
(weeks)
Labor status
Birth weight
(g)
1 33 1 34 Sp labor 1990
2 35 0 35 Sp labor 2490
3 35 0 35 Sp labor 2282
4 35 1 36 Induced 2446
5 30 4 34 Induced 2520
6 32 1 33 Sp labor 2530
7 33 0 33 Sp labor 2490
8 33 1 34 Sp labor 2690
9 36 0 36 Induced 3380
10 33 0 33 Sp labor 2898
Table 6.2 describes the characteristics of the PPROM study popula-
tion. The median (range) gestational age at the amniocenteses was 33 (30–36) (mean 33.5, SD ± 1.78) weeks. All patients had spontaneous or induced labor. The onstet of labor was spontaneous in 70% of cases, 30% were induced because of anhydramnion or late PPROM (36 weeks). The latency period mean (range) to delivery was 0.8, SD ± 1.23 (0–4) weeks. Most of the women delivered in few days after rupture of membranes, few prolonged pregnancy for more than one week, and only one patient was able to prolong the latency period more than two weeks. Patients were followed after enrollment until delivery. The median (range) gestational age at deli-very was 34 (33–36), mean 34.3, SD ± 1.16 weeks of gestation. All patients delivered vaginally without complications.
Characteristics of newborns in cases of PPROM patients All newborns survived, mean total birth weight was 2572 g, SD ± 369
g, mean height was 45.7 cm, SD ± 3.8; Apgar score mean at 1 min 8.1, SD ± 0.57 (range 7–9) and at 5 min 8.9, SD ± 0.74 (range 8–10); umbilical cord arterial blood pH 7.32 SD ± 0.06 (range 7.21–7.40). All newborns were non-admitted to neonatal intesive care unit (NICU) and there were no signs of congenital infection.

71
The cause of membrane rupture in most cases is unknown, but asymptomatic intrauterine infection is a frequent precursor in early PPROM cases. The research indirectly confirms this affirmation because all PPROM cases before 30 weeks were found to be with asymptomic infection; therefore were excluded from the study.
6.1. Peptides and proteins identified in amniotic fluid
A fully automated 2D LS/MS/MS system identified 836 peptides in amniotic fluid. Processed MS/MS data were submitted to the Mascot search engine by using BioTools (Bucker Daltonics) for protein identification. The chromatograms of amniotic fluid samples from yellow, red and green groups were relatively similar (Fig. 6.1.1). Low to high intensity peak (> 2000 counts) appearance reproducibility from sample to sample was in the range of usually obtained in proteomics using LC/MS: a) yellow group 214, SD ± 49.4 (n = 30); b) red group 260, SD ± 12.5 (n = 10); c) green group 263, SD ± 18.6 (n = 16) (Figure 6.1.2). Thus, from each group were selected five samples for LC/MS/MS analysis and submitted to the Mascot search engine for protein identification.
Fig. 6.1.1. Mass spectra of AF samples Yellow – G group, red – R group, and green – Z group.

72
Fig. 6.1.2. Box charts for high intensity peaks appearance
in chromatograms of AF samples a – G group, b – R group, and c – Z group of AF samples.
In total, 502 proteins were identified. To select only distinct proteins,
the entries were sorted by their names and molecular weights and removed identical ones. A Mascot score above 30 was used for positive protein identification. The non-redundant list was generated. Different matches of one gene may reflect biologically significant different gene products such as splice variants, sequence variants, and cleavage products. In the method applied such a distinction is often difficult to perform because protein prediction is based on peptide sequence searches. Therefore, a more strin-gent method was used to include only non-redundant proteins. Proteins that were identified with two or more unique peptides were searched. All the proteins with at least two independent peptide identifications were considered to be highly likely to be present in the sample (probability ] 80%). The redunducies were removed all, but one entry with the highest number of unique sequences left. In total, 85 proteins were identified with high confidence. All proteins identified from amniotic fluid are listed in Table 6.1.1.

73
Table 6.1.1. Proteins identified from the amniotic fluid
No. Group* Protein
(human) Protein name
Function
(from UniProtKB/Swiss-Prot and Gene Ontology databases)
1 2 3 4 5
1 Z RS7 40S ribosomal protein S7 Required for rRNA maturation, RNR binding
2 Z RL28 60S ribosomal protein L28 RNA binding
3 GRZ Q64FX6 AT-hook-containing transcription factor
Transcription factor
4 GR Q0PVP5 Alpha-1 antitrypsin variant
5 GRZ A1AT Alpha-1-antitrypsin Serine protease inhibitor. Its primary target is elastase, but it also has a moderate affinity for plasmin and thrombin (plasma).
6 G Q9NQC9 Ankyrin repeat domain 2 (Fragment)
Muscle contraction, muscle organ development
7 G ANR43 Ankyrin repeat domain-containing protein SOWAHA
8 GRZ ANXA2 Annexin A2 Calcium-regulated membrane-binding protein
9 R Q13789 Apolipoprotein B (Fragment) Innate immunity
10 GR Q6FGT0 Aquaporin 9, isoform CRA_a (Fragment)
Transport
11 GRZ Q4TWB7 Beta globin chain (Fragment) Oxygen transport
12 GR Q9P129 Calcium-binding mitochondrial carrier protein SCaMC-1 (Fragment)
Transmembrane transport
13 GRZ Q9NW12 La-related protein 1B RNA binding
14 R Q9H9G5 @'/C&D9,U_\ML&$14E&@-#.6&
NT2RP2001581 C-terminal protein lipidation
15 GRZ Q6ZNS5 @'/C&D9,_\_[^&$is, clone SYN07051
Small GTPase mediated signal transduction
16 Z Q6ZTZ1 Myb/SANT-like DNA-binding domain-containing protein 1
17 GRZ Q542Y5 '.+,&:#B#-#?&43T$+B1-=&`&
member 11 Activation of signaling protein activity, involved in unfolded protein response
18 GRZ Q7Z7Q2 Chromodomain-helicase-DNA-binding protein 7 (Fragment)
In uterus embryonic development, probable transcription regulator
19 GRZ CLIC4 Chloride intracellular channel protein 4
Transport
20 R CO1A1 Collagen alpha-1(I) chain Type I collagen is a member of group I collagen (fibrillar forming collagen). Extracellular matrix structural constituent

74
Table 6.1.1 continued
1 2 3 4 5
21 Z CO3A1 Collagen alpha-1(III) chain Collagen type III occurs in most soft connective tissues along with type I collagen. Extracellular matrix struc-tural constituent
22 GRZ CO1A2 Collagen alpha-2(I) chain Type I collagen is a member of group I collagen (fibrillar forming collagen). Extracellular matrix structural constituent
23 G CFAB Complement factor B Immunity, innate imunity
24 GZ Q53TY4 CSMD2 protein
25 Z AMPL Cytosol aminopeptidase Proteolysis Presumably involved in the processing and regular turnover of intracellular proteins. Catalyzes the removal of unsubstituted N-terminal amino acids from various peptides.
26 GRZ KM!,'M DVL1 protein (Fragment) Signal transducer activity
27 R Q32M70 F-box only protein 40 Heart and skeletal muscle cell differentiation
28 GRZ Q5VWW3 Myoferlin Muscle contraction. Calcium/phospholipid-binding protein that plays a role in the plasmalemma repair mechanism of endothelial cells that permits rapid resealing of mem-branes disrupted by mechanical stress.
29 GRZ Q8WW76 FGA protein Fibrinogen complex
30 GRZ FIBA Fibrinogen alpha chain Blood coagulation, hemostasis
31 GRZ FIBB Fibrinogen beta chain Blood coagulation, hemostasis
32 G FINC Fibronectin Fibronectins bind cell surfaces and various compounds including collagen, fibrin, heparin, DNA, and actin. Fibronectins are involved in cell adhesion, cell motility, opsonization, wound healing, and maintenance of cell shape. Extracellular matrix structural constituent
33 GRZ A1L468 Fibronectin type III domain-containing protein 7
34 GRZ FREA Forkhead box protein S1 Development: artery morphogenesis, embryonic heart tube development, ossification, positive regulation of multicellular organism growth, regu-lation of organ growth, somitogenesis, vasculogenesis, tissue morphogenesis

75
Table 6.1.1 continued
1 2 3 4 5
35 GR Q53XB8 Full-length cDNA clone CS0DE007YP21 of Placenta of Homo sapiens (human)
36 RZ G3P Glyceraldehyde-3-phosphate dehydrogenase
Apoptosis, glycolysis, translation regulation
37 GR Q9H2Q8 GREB1a (Fragment) Play a role in estrogen-stimulated cell proliferation
38 R Q6GTG1 Vitamin D binding protein Vitamin D binding
39 G Q2TU31 Growth-inhibiting protein 21 Development
40 GR MYO15 Unconventional myosin-XV Inner ear morphogenesis
41 Z Q9H342 Olfactory recept#"&ZU,U Olfaction, sensory transduction
42 G Q9UF27 Fibrocystin-L (Fragment) Receptor activity
43 GRZ Q6MZU6 Putative uncharacterized protein DKFZp686C15213
44 GR Q68CN4 Putative uncharacterized protein DKFZp686E23209
45 G Q7Z3N9 Putative uncharacterized protein DKFZ50p686G16 (Translin-associated factor X, isoform CRA_a)
Sequence-specific DNA binding
46 GRZ Q6N097 Putative uncharacterized protein DKFZp686H20196
47 GRZ Q6N093 Putative uncharacterized protein DKFZp686I04196 (Fragment)
48 GRZ Q6N030 Putative uncharacterized protein DKFZp686I15212
49 GRZ Q6MZX7 Putative uncharacterized protein DKFZp686M24218
50 GRZ Q7Z351 Putative uncharacterized protein DKFZp686N02209
51 GRZ Q6N094 Putative uncharacterized protein DKFZp686O01196
52 RG Q53SU3 Putative uncharacterized protein DLX1
Embryonic skeletal system develop-ment, hippocampus development, negative regulation of oligodendrocyte differentiation, odontogenesis of dentin-containing tooth
53 GRZ IGHG1 Ig gamma-1 chain C region Innate immune response, antigen binding
54 GRZ IGHG2 Ig gamma-2 chain C region Innate immune response 55 G Q9BZS0 Transcription factor HIVEP3 Immunity

76
Table 6.1.1 continued
1 2 3 4 5 56 GR Q5XKE5 Keratin, type II cytoskeletal
79 Expressed in skeletal muscle, skin and scalp, but not in any other tissues or organs examined
57 Z K1C14 Keratin, type I cytoskeletal 14 Structural constituent of cytoskeleton
58 R K1C18 Keratin, type I cytoskeletal 18 Structural constituent of cytoskeleton Expressed in colon, placenta, liver and very weakly in exocervix
59 R K1C19 Keratin, type I cytoskeletal 19 Structural constituent of cytoskeleton
60 GR K2C7 Keratin, type II cytoskeletal 7 Structural constituent of cytoskeleton
61 R Q86SV6 K-glypican GPI-anchored heparan sulfate proteoglycan that is highly expressed in developing brain and kidney, may play a role in the control of cell division and growth regulation
62 Z LRIG3 Leucine-rich repeats and immunoglobulin-like domains protein 3
May play a role in craniofacial and inner ear morphogenesis during embryonic development. May act within the otic vesicle epithelium to control formation of the lateral semicircular canal in the inner ear.
63 GRZ LUM Lumican (Keratan sulfate proteoglycan lumican)
Cartilage development
64 GRZ Q53FV4 Lumican variant (Fragment)
65 GRZ Q2LHB3 NADH-ubiquinone oxidoreductase chain 2
Nervous system development, may be required for neuronal cell differentiation.
66 Z AHN Neuroblast differentiation-associated protein AHNAK
Nervous system development. May be required for neuronal cell differentiation.
67 G ORCT2 Solute carrier family 22 member 18
68 ZR KZ,`^[ Proliferation-inducing protein 10 or 9
69 GR PCSK5 Proprotein convertase subtilisin/kexin type 5
Found in pregnancy, embrionic system development
70 R Q5F2G0 Serine/threonine-protein phosphatase 2B catalytic subunit beta isoform (Fragment)
Calcium-dependent, calmodulin-stimulated protein phosphatase. This subunit may have a role in the calmodulin activation of calcineurin
71 Z Q96P56 Cation channel sperm-associated protein 2
Voltage-gated calcium channel that plays a central role in calcium-dependent physiological responses

77
Table 6.1.1 continued
1 2 3 4 5
72 GR KPYM Pyruvate kinase isozymes M1/M2
Glycolysis
73 GRZ CHK2 Serine/threonine-protein kinase Chk2
74 GRZ TRFE Serotransferrin Ferric iron binding
75 GRZ ALBU Serum albumin
76 Z AT1A1 Sodium/potassium-transporting ATPase subunit alpha-1
77 R SYTL2 Synaptotagmin-like protein 2 Exocytosis
78 GR Q06AH7 Transferrin Ferric iron binding
79 Z Q197M4 Protein-glutamine gamma-glutamyltransferase K
Keratinization
80 GRZ Q8NCT0 Transketolase-like protein 2 (Fragment)
81 GRZ O15417 Trinucleotide repeat-containing gene 18 protein
82 GRZ SNUT1 U4/U6.U5 tri-snRNP-associated protein 1
83 GRZ VIME Vimentin Structural constituent of cytoskeleton. Vimentins are class-III intermediate filaments found in various non-epithelial cells, especially mesenchymal cells. Involved with LARP6 in the stabilization of type I collagen mRNAs for CO1A1 and CO1A2.
84 GRZ KZ,;\Z WW domain containing adaptor with coiled-coil (Fragment)
85 GRZ KNZ`,^ ZNF690 protein
*G-yellow group, R-red group, Z-green group.
Some authors use the depletion of high abundance proteins from the sample before proteomics analysis to avoid the masking effect. They eli-minate major serum proteins (albumin, imunoglobulins, anti-trypsin, trans-ferrin, haptoglobin) which constitute about 70% of all proteins composition in AF. This approach has a pronounsed disadvantage. Albumin and immu-noglobulins or other high abundance proteins tend to bind small proteins or peptides, a phenomenon called the “sponge” effect. Removal of these pro-teins may eliminate some small proteins or peptides which are the field of interest. Therefore, the depletion of high abundance proteins was not done in this work. In the table of proteins from amniotic fluid also peptides from

78
albumin and immunoglobulins were found. IgGs in AF may have a protective function.
The levels of proteins like AFP in maternal serum are highly related to pregnancy. These proteins are produced and secreted by either the fetus or the placenta as pregnancy progresses. AFP was not detected in this study, probably because the AFP level is the highest at 10–13 weeks and reduces with the gestational age. All found keratins (cytokeratins 7, 14, 18, 19 and 79) show keratinization of the fetus skin, which starts after 19–20 week of gestation. Transferrin, serotransferrin, calcium binding and vitamin D binding proteins are transport proteins that are relevant to fetal growth and development: they transport Vitamin E, Vitamin D, Vitamin A, copper and iron. The aquaporins are a family of water-selective membrane channels. Aquaporin 9 allows passage of a wide variety of non charged solutes. It stimulates the transport of urea and the osmotic water permeability. Aqua-porin 9 might play also some role in specialized leukocyte functions such as immunological response and bactericidal activity. AF contributes to fetal lung development, lipids that compose fetal lung surfactant are derived from apolipoproteins [157]. Apolipoprotein B was found in AF. Chloride intra-cellular channel-4 (CLIC4) protein regulates fundamental cellular processes including stabilization of the cell membrane potential, the transepithelial transport, the maintenance of intracellular pH, and the regulation of cell volume. The CLIC4 protein, encoded by the CLIC4 gene, is a member of the p64 family. The gene is expressed in many tissues and binds to dynamin VE& F-tubulinE& I-actin, creatine kinase and two 14-3-3 isoforms [158]. The growth-inhibiting protein 21 (FGF21) causes growth inhibition by acting directly at the long bones' growth plate. The expression of FGF21, FGFR1 +.0&DPD([&8*H#&"6@6G*#"4&5.#H.&*#&T6&+@*1<+*60&T=&DPD_UW&+.0&I-klotho (a co-receptor required for the FGF21-mediated receptor binding and activation) in fetal and 3-week-old mouse growth plate chondrocytes was demonstrated [159]. High concentrations of FGF21 might directly suppress the growth plate chondrocyte proliferation and differentiation. The FGA protein and fibrinogens are complement and coagulation proteins. Comple-ment factor B, is a component of the alternative pathway of complement activation. Factor B circulates as a single chain polypeptide. Upon active-tion of the alternative pathway, it is cleaved by the complement factor D yielding the noncatalytic chain Ba and the catalytic subunit Bb. Since all of the complement pathway proteins are known to be part of the acute-phase response, they may have a protective function for the developing fetus. As many of the proteins of the complement cascade are present in AF, it is

79
possible that they may prevent intraamniotic infection and the subsequent complications that can result. Several of the coagulation proteins we have identified are also involved in hemostasis and blood pressure regulation. A subset of metabolic proteins of AF have implications for the fetal develop-ment. These enzymes might protect the developing fetus. Type I and III are the primary regulators of the tensile strength. Interstitial collagens are located in the compact layer of amnion. The cellular source of interstitial collagens is unclear. Casey et al. found that both epithelial and mesen-chymal cells produce interstitial collagens. It seems as mesenchymal cells express significantly higher levels of collagens I and III than epithelial cells [160]. Vimentin is a structural constituent of cytoskeleton. Vimentins are class-III intermediate filaments found in various non-epithelial cells, espe-cially mesenchymal cells. They are involved in the stabilization of type I collagen. Proprotein convertase subtilisin/kexin type 5 is an enzyme. Many enzymes are inactive when they are first synthesized, because they have a section of peptide chains that blocks their activity. Proprotein convertases remove that section to activate the enzyme. Annexin A2 is a member of the annexin family. Members of this calcium-dependent phospholipid-binding protein family play a role in the regulation of cellular growth and in signal transduction pathways. This protein functions as an autocrine factor which heightens osteoclast formation and bone resorption.
There are still quite a few proteins found in the amniotic fluid protein whose function remains undiagnosed.
Due to the complexity of the amniotic fluid sample it appears difficult
to choose the appropriate method for proteome investigation. Scientific groups working in this field use various methods of analysis. Therefore the identified proteins and their number varies depending on the method employed. Most of the proteins were identified using several analytical methods, but due to the intricacy of the analysis the number of tested samples is usually small. These studies are expensive and time-consuming, and require substantial manual work. Thus, an automated system is pre-ferred, which was developed for this study. In order to create a larger amniotic fluid proteome map different analytical methods are justified for usage. In the future proteins that could be used as specific disease biomar-kers should be validated. According to the literature in this study so far the largest number of amniotic fluid samples was investigated using one automated 2D LS/MS/MS system and samples were from three periods of pregnancy (second trimester, early and late third trimester of pregnancy).

80
Large-scale studies were reported only by research groups investigating amniotic fluid proteome at intrauterine infection.
In this study a set of 836 amniotic fluid peptides was identified. This can be considered, as a good result in comparison with previous publications. The biggest challenge in proteomics, which is also faced by other scientific groups too, is protein identification. Modern databases, such as the UniProtKB/Swiss-Prot and others, need digested protein samples for high score protein identification. The goal was to find original peptides (synthesized by organism as peptides, or digested by the organism) in amniotic fluid. Therefore, the undigested samples were analyzed. It was difficult to identify the proteins from peptides as protein prediction is based on digested protein peptide sequence. Therefore, relatively small amounts of proteins were identified from a large number of peptides seen in mass spectra.
6.2. Characterization of amniotic fluid proteome
Subcellular localization According to data obtained from the UniProtKB/Swiss-Prot and Gene
Ontology databases each protein was classified by subcellular localization. When one protein is known to be localized in more than one cellular compartment, all of the categories were counted non exclusively. Figure 6.2.1 shows the cellular distribution of 69 identified proteins with known localization. The cellular distribution of identified proteins was reported as membrane 22% (33 proteins), extracellular 13% (20 proteins), nuclear 13% (19 proteins), secreted 13% (19 proteins), cytoplasmic 11% (17 proteins), cytoskeletal 9% (14 proteins), goldi 5% (7 protein), mitochondrial 3% (5 proteins), endoplasmic reticulum 3% (4 protein), centrosome 3% (4 pro-teins), lysosomal 1% (2 protein) and others 5% (7 proteins) (Fig. 6.2.1). The majority were membrane proteins.

81
Fig. 6.2.1. Subcellular localization of proteins from amniotic fluid
Assignments of functions
The function of each protein was searched from the UniProtKB/ Swiss-Prot and Gene Ontology databases. 72 of total 85 proteins were matched with functions. Functions were categorized as shown in figure 6.2.2. Because one protein may have multiple functions, all of the categories were counted nonexclusively. The functional analysis indicated that 25 of the identified proteins were regulatory (27%), 16 structural (17%), 14 development (15%), 13 binding proteins (14%), 8 enzymes (9%), 10 were related to transportation (11%) and 7 were responsible for immune response (7%) (Fig. 6.2.2).
13%
22%
13%
11%
9%
3%
5%
3%
1%
3%
5%
13%
Extracellular
Membrane
Nuclear
Cytoplasmic
Cytoskeletal
Endoplasmic reticulum
Golgi
Mitochondrial
Lysosomal
Centrosome
Others
Secreted
0 5 10 15 20 25 30 35
Lo
cali
zati
on
Number of proteins

82
Fig. 6.2.2. Functional analysis of proteins from amniotic fluid
Despite of the interest regarding localization and functions of proteins in amniotic fluid, there have been limited opportunities to generate an in-depth analysis. 7.1% of the identified proteins are yet with unknown functions, 10.6% with unkown subcellular localization and 8.2% with unrevealed both funtion and localization in AF. Some of the top functions of AF proteins, as shown in Figure 6.2.2, including regulatory, structural, development and binding. The list for functions of proteins provide ideas on how different proteins act in AF to support the normal fetal development. This also allows one to explore novel biomarkers based on known func-tional pathways.
6.3. Comparison of the amniotic fluid proteome with previous
publications
The list was generated to combine human AF proteome entries from
nine previous publications. A list of overlapping proteins known at the moment to be present in AF was analyzed. From all proteins identified in this study 18 were reported by other research groups at least one time (Table 6.3.1).
Regulatory 27%
Structural 17%
Enzyme 9%
Development 15%
Transportation 11%
Immune response 7%
Binding 14%

83
Table 6.3.1. Overlapping proteins of this study with previously reported proteins by other research groups (proteins identified in this study with confidence are in bold)
No Protein name Protein
code
Ma
chte
je"#$%!
Lib
era
tori
[59
]
Vu
ad
ens
[70
]
Gra
vet
t [7
8]
Nil
sso
n [
60
]
Pa
rk [
61
]
Mic
ha
el [
62
]
Tsa
nga
ris
[63
]
Ch
o [
64
]
1 Alpha-1-antitrypsin Q0PVP5 Y* Y Y Y
2 Angiotensinogen variant (Fragment)
Q59EP2 Y Y Y
3 Antigen NY-CO-41 (Fragment) O60535 Y Y
4 Apolipoprotein B (Fragment) Q13789 Y Y
5 ATP-binding cassette sub-family B member 5 (P-glycoprotein ABCB5) (ABCB5 P-gp)
ABCB5 Y Y Y
6 Beta globin chain (Fragment) Q4TWB7 Y Y YY Y
7 Collagen, type XVII, alpha 1 KZ,a[M Y Y Y
8 Complement factor B
precursor CFAB Y Y Y Y Y Y
9 Cystatin B (Cystatin B (Stefin B), isoform CRA_a)
Q76LA1 Y Y Y
10 Fibrinogen alpha chain Q4QQH7 Y Y Y
11 Fibronectin type III domain
containing 7
A1L468 Y Y Y Y
12 Formin binding protein 2 (Fragment)
Q5VZB5 Y
13 Gelsolin precursor (Actin-depolymerizing factor)
GELS Y Y Y
14 Leucine-rich repeats and
immunoglobulin-like domains
protein 3 precursor
LRIG3 Y Y Y Y
15 Lumican (Keratan sulfate
proteoglycan lumican)
LUM Y Y Y Y
16 Protocadherin protein (Fragment) Q6V1P8 Y Y
17 Serum albumin Q56G89 Y Y Y Y Y Y Y
18 Transferrin Q06AH7 Y Y Y Y Y Y
*Y – yes.

84
13 proteins identified in this study were previously reported in at least two publications. Five proteins were reported only in one study, five proteins in two and four proteins in three previuos publications (Fig. 6.3.1). Four proteins were identified by at least four research groups previously.
Fig. 6.3.1. Number of overlapping proteins of previous publications
6.4. Gestational age-dependent changes in the amniotic
fluid proteome
Relatively little is known about the detailed amniotic fluid proteome
despite the interest in amniotic fluid composition and function. This study was designed to analyze the amniotic fluid proteins and peptides in three groups at different gestational period. A fully automated 2D LS/MS system identified 836 peptides distributed between the groups depending on the gestational age (Table 6.4.1 and Fig. 6.4.1).
Table 6.4.1. Number of peptides in the groups according to gestational age
G – yellow group, R – red group, Z – green group.
0 1 2 3 4 5 6
1
2
3
4
5
6
No. of proteins
No.
of
prev
iuos
pub
licat
ions
Groups of AF samples No. of peptides after analysis of signals
G group (16–18 weeks) 223
R group (30–36 weeks) 282
Z group (38–40 weeks) 331

85
Fig. 6.4.1. Differential peptide map for the 1st fraction 1"2(&3'"456, R 2(&3'476, Z 2(&3'"486, RT – retension time. Signal intensity is coded by the
colour.
118 (42.3%) of the peptides were present from the beginning and remained during pregnancy. It is likely that those proteins and peptides are important for the development of the fetus and other pregnancy related processes. Figure 6.4.2 shows the unique and shared peptides among groups depending on the gestational age. As expected, the yellow group G, shows substantially less similarity to group R (16 peptides) and Z (27 peptides) as it is more distant from other groups by pregnancy time.

86
Fig. 6.4.2. Similarities and differencies between peptides according to the gestational age
G – yellow group, R – red group, Z – green group.
From the identified peptides, the Mascot database was able to link dependence to 85 proteins. Figure 6.4.3 and Table 6.1.1 shows the unique and shared proteins among the groups depending on the gestational age.
Fig. 6.4.3. Similarities and differencies between proteins according
to the gestational age G – yellow group, R – red group, Z – green group.
Out of 85 AF proteins, nine were found only in early pregnancy: the
complement factor B; fibronectin; ankyrin repeat domain-containing protein SOWAHA; fibrocystin-L; transcription factor HIVEP3; growth-inhibiting

87
protein 21; ankyrin repeat domain-containing protein 2; solute carrier family 22 member 18; translin-associated factor X, isoform CRA_a.
Some proteins and peptides occur only in the second half of preg-nancy. 14 proteins were observed in both G and R goups: keratin, type II cytoskeletal 7; hypothetical protein DKFZp686E23209; serum albumin; alpha-1 antitrypsin variant; alpha-1-antitrypsin; aransferrin; gene regulated in breast cancer 1 protein; putative uncharacterized protein DLX1; aqua-porin 9, isoform CRA_a; proprotein convertase subtilisin/kexin type 5; pyruvate kinase isozymes M1/M2; keratin, type II cytoskeletal 79; unconventional myosin-XV; calcium-binding mitochondrial carrier protein SCaMC-1.
Ten proteins were found in AF only during PPROM: vitamin D-bin-ding protein; synaptotagmin-like protein 2; F-box only protein 40; apoli-G#G"#*61.&`b&O'/C&D9,U_\ML&$14E&@-#.6&/J_(!_NNUZQUb&serine/threonine-protein phosphatase 2B catalytic subunit beta isoform; collagen alpha-1(I) chain; keratin, type I cytoskeletal 18; K-glypican; keratin, type I cytoske-letal 19.
Two proteins were shared in the G and Z groups: glyceraldehyde-3-phosphate dehydrogenase; proliferation-inducing protein 10 or 9.
12 proteins were found only in term pregnancies with intact mem-branes: collagen alpha-1(III) chain; 40S ribosomal protein S7; olfactory "6@6G*#"& ZU,Ub& MNc& "1T#4#B+-& G"#*61.& 9_Qb&d=TYcC/J-like DNA-binding domain-containing protein 1; keratin, type I cytoskeletal 14; protein-glutamine gamma-glutamyltransferase K; sodium/potassium-transporting ATPase subunit alpha-1; leucine-rich repeats and immunoglobulin-like domains protein 3; neuroblast differentiation-associated protein AHNAK; cation channel sperm-associated protein 2; cytosol aminopeptidase.
The Z and G group shared only one - CSMD2 - protein. This approach also identified a set of proteins consistently expressed
throughout the pregnancy. Among the 85 proteins identified in this study from AF, 37 (43.5%) proteins were found in all groups (GRZ) indepen-dently on gestational age (Table 6.1.1). It is highly likely that the function of those proteins is closely associated with the molecular and physiological pathways ralated to pregnancy.
Since the amniotic fluid protein and peptide composition is dynamic and depends on the gestational age, the discovery the new biomarkers of diseases in the pregnancy should take into account the physiological characteristics. Michaels and al. reported the biggest difference in amniotic fluid proteome between the first and second trimester of pregnancy

88
[66].This was confirmed in this study too. Comparing the amniotic fluid taken at the 16–18th and the 30–36th weeks of gestation only 16 shared peptides were found, and comparing the second group with a term amniotic fluid 66 common peptides were found. The significance of these changes is not really clear. It should be noted, however, that AF is a very dynamic mixture, the composition of which is known to differ depending on the gestational age. These data further support the view that proteomics analysis of AF may be a feasible and become an effective method to screen multiple pathologies in the future.
6.5. Fetal membranes proteome
The fetal membranes are essential for maintenance of pregnancy. The
amniochorion can secrete and release peptides and proteins into the amniotic cavity (Fig. 6.5.1). In contrast to its significant role, only a limited protein profile of human amniochorion is found in literature. Proteomics allows a simultaneous study of a multitude of proteins and is of great importance to gain insight into the physiology of PPROM. The differences in the proteome of fetal membranes in women with PPROM and at term pregnancies with intact membranes could reveal potential biomarkers for PPROM diagnosis. This study is the first proteome analysis of amnio-chorion.
Fig. 6.5.1. Components of amniochorionic membrane Microscopic images, magnification ×20.

89
The ideal way to explore processes in the fetal membranes is to analyze intact proteins at the native location – inside the amniochorion. However, this is so far impossible as analytical instruments which would be able to detect a single protein is still absent. Therefore, in order to analyze the membrane composition it is needed to crash the membrane – into the proteins or digest it to its components - peptides. Proteins could be separated by gel electrophoresis, digested into peptides and analyzed with MS. However, 2D PAGE fails to analyze < 15 kDa proteins (peptides) hardly can cope with concentration differences of several orders of magnitude and lack of automation. Proteins of ~10 kDa do not focus well in isoelectric focusing step. Proteins of < 15 kDa could be easily lost as they migrate too fast. Where 2D PAGE has weaknesses, chromatographic techniques have their strengths. On the other hand, chromatography lacks the ability to separate proteins, therefore proteins have to be digested prior to analysis, or originally existing peptides should be selectively cached and analysed. Once membranes are digested (the digestion procedure is described in section 5). we lose valuable information about the precise protein location and only construct the overall picture of membranes general composition. It needs to be pointed out that this method is not able to reveal proteolytic processes in membranes itself. Protein identification is accomplished by matching peptides and those peptides could be originated from the processes in membrane in vivo or could be products of the membranes digestion in vitro.
6.5.1. Comparison of proteome from fetal membranes of patients
with PPROM and term labor with intact membranes
A fully automated 2D liquid chromatographic system with online sample cleans up and directly coupled to mass spectrometry (see section 5) was used. The obtained MS spectra were manually analyzed (procedure and rules are described in section 5). Initially, 220 peptide-like signals were found in PPROM membranes and 270 peptide-like signals in term pregnancy membranes. Then the data were additionally matched, and only those signals were selected which were identified multiple times in the row, in order to increase the peptide identification probability. After such data reprocessing 195 peptide signals were identified and submitted to a Mascot search, for protein matching. It was noticed that the digestion of PPROM membrane was substantially faster, approximately 24 hours, instead of 48 hours needed for the complete digestion of the control of fetal membranes.

90
A total of 43 proteins from 195 peptides were identified from the digested fetal membranes (Table 6.5.1.1). To avoid false positive identi-$1@+*1#.E&G"#*61.4&H6"6& 106.*1$160&T=&]&_&matching peptides within a single protein.
Table 6.5.1.1. Proteins identified from the digested fetal membranes and also found in amniotic fluid (proteins characteristic to PPROM membranes are in bold). For positive protein identification was used Mascot score above 67 (p < 0.05).
No.
Protein
code
(all
human)
Protein
Found in
membra-
nes
Found
in AF
Function
(from UniProtKB/ Swiss-Prot and Gene Ontology databases)
1 2 3 4 5 6 1 RS7 40S ribosomal
protein S7
R Z Required for rRNA maturation, RNR binding
2 RL28 60S ribosomal
protein L28
R Z RNA binding
3 PPB1 Alkaline phosphatase, placental type
RZ Alkaline phosphatase activity, hydrolase
4 A1AT Alpha-1-antitrypsin
R RZ Serine protease inhibitor. Its
primary target is elastase, but it also has a moderate affinity for plasmin and thrombin (plasma).
5 ENOA Alpha-enolase RZ Multifunctional enzyme that, as well as its role in glycolysis, plays a part in various processes such as growth control, hypoxia tolerance and allergic responses.
6 ANXA2 Annexin A2 RZ RZ Calcium-regulated membrane-binding protein

91
Table 6.5.1.1 continued 1 2 3 4 5 6 7 PGBM Basement
membrane-specific
heparan sulfate
proteoglycan core
protein (perlecan,
endorepellin)
R Integral component of basement membranes. Required for avascular cartilage development. It interacts with other basement membrane components such as laminin, prolargin and collagen type IV. Interacts with COL13A1, FGFBP1 and VWA1. Interacts (via C-terminus) with ECM1 (via C-terminus).
8 CO1A1 Collagen alpha-1(I) chain
RZ R Type I collagen is a member of group I collagen (fibrillar forming collagen). Extracellular matrix structural constituent.
9 CO3A1 Collagen alpha-1(III) chain
RZ Z Collagen type III occurs in most soft connective tissues along with type I collagen. Extracellular matrix structural constituent.
10 CO1A2 Collagen alpha-2(I) chain
RZ RZ Type I collagen is a member of group I collagen (fibrillar forming collagen). Extracellular matrix structural constituent.
11 CO6A2 Collagen alpha-2(VI) chain
RZ Collagen VI acts as a cell-binding protein (cell adhaesion). Extracellular matrix structural constituent.
12 AMPL Cytosol aminopeptidase
RZ Z Presumably involved in the processing and regular turnover of intracellular proteins. Catalyzes the removal of unsubstituted N-terminal amino acids from various peptides.
13 DPOLA DNA polymerase
alpha catalytic
subunit
R Initiation of DNA replication

92
Table 6.5.1.1 continued 1 2 3 4 5 6 14 FIBA Fibrinogen alpha
chain RZ RZ Blood coagulation,
hemostasis 15 FIBB Fibrinogen beta
chain RZ RZ Blood coagulation,
hemostasis 16 FINC Fibronectin R Fibronectins bind cell surfa-
ces and various compounds including collagen, fibrin, heparin, DNA, and actin. Fibronectins are involved in cell adhesion, cell motility, opsonization, wound healing, and maintenance of cell shape. Extracellular matrix structural constituent.
17 G3P Glyceraldehyde-3-phosphate dehydrogenase
RZ RZ Apoptosis, glycolysis, translation regulation
18 H2A1B Histone H2A type 1-B/E
RZ Transcription regulation, DNA repair, DNA replication and chromosomal stability
19 IGHG1 Ig gamma-1 chain C region
RZ RZ Antigen bindig
20 K1C14 Keratin, type I cytoskeletal 14
RZ Z Structural constituent of cytoskeleton
21 K1C16 Keratin, type I
cytoskeletal 16
R Structural constituent of cytoskeleton
22 K1C17 Keratin, type I
cytoskeletal 17
R Structural constituent of cytoskeleton
23 K1C18 Keratin, type I cytoskeletal 18
RZ R Structural constituent of cytoskeleton
24 K1C19 Keratin, type I cytoskeletal 19
RZ R Structural constituent of cytoskeleton
25 K1C24 Keratin, type I cytoskeletal 24
RZ Structural constituent of cytoskeleton
26 K2C5 Keratin, type II cytoskeletal 5
RZ Structural constituent of cytoskeleton
27 K2C6A Keratin, type II cytoskeletal 6A
RZ Structural constituent of cytoskeleton
28 K2C7 Keratin, type II cytoskeletal 7
RZ R Structural constituent of cytoskeleton
29 K2C8 Keratin, type II cytoskeletal 8
RZ Structural constituent of cytoskeleton

93
Table 6.5.1.1 continued 1 2 3 4 5 6 30 MIF Macrophage
migration inhibitory factor
RZ Pro-inflammatory cytokine. Involved in the innate immune response.
31 AHNK Neuroblast differentiation-associated protein AHNAK
RZ Z Nervous system development. May be required for neuronal cell differentiation.
32 PRDX6 Peroxiredoxin-6 RZ Lipid metabolism 33 TGM2 Protein-glutamine
gamma-
glutamyltransferase
2
R R Catalyzes the cross-linking of proteins and the conjugation of polyamines to proteins
34 KPYM Pyruvate kinase isozymes M1/M2
RZ R Glycolysis
35 ALBU Serum albumin RZ RZ 36 SMD1 Small nuclear
ribonucleoprotein
Sm D1
R mRNA processing, mRNA splicing
37 S26A1 Sulfate anion
transporter 1
R Transport
38 QSOX1 Sulfhydryl oxidase
1
R Catalyzes the oxidation
39 SYTL2 Synaptotagmin-like
protein 2
R R Exocytosis
40 TENA Tenascin R Cell adhaesion. Extracellular matrix protein implicated in guidance of migrating neurons as well as axons during development, synaptic plasticity as well as neuronal regeneration. Promotes neurite outgrowth from cortical neurons grown on a monolayer of astrocytes. Ligand for integrins alpha-8/beta-1, alpha-9/beta-1, alpha-V/beta-3 and alpha-V/beta-6.

94
Table 6.5.1.1 continued 1 2 3 4 5 6 41 BGH3 Transforming
growth factor-beta-induced protein ig-h3
RZ Binds to type I, II, and IV collagens. This adhesion protein may play an important role in cell-collagen interactions. In cartilage, may be involved in endochondral bone formation.
42 TPIS Triosephosphate isomerase
RZ Triose-phosphate isomerase activity
43 VIME Vimentin RZ RZ Structural constituent of cytoskeleton. Vimentins are class-III intermediate filaments found in various non-epithelial cells, especially mesenchymal cells. Involved with LARP6 in the stabilization of type I collagen mRNAs for CO1A1 and CO1A2.
Legend: R – red group, Z – green group.
Table 6.5.1.2. Top five proteins in PPROM fetal membranes according to number of peptide
No. of
peptides
Mascot
score Protein code Protein name (Swiss-Prot)
13 433 G3P_HUMAN Glyceraldehyde-3-phosphate dehydrogenase
10 618 FINC_HUMAN Fibronectin
10 127 CO1A1_HUMAN Collagen alpha-1(I) chain
9 384 CO1A2_HUMAN Collagen alpha-2(I) chain
9 356 K1C19_HUMAN Keratin, type I cytoskeletal 19
According to the data obtained from UniProtKB/Swiss-Prot and Gene
Ontology databases each protein was classified by subcellular localization. When one protein is known to be localized in more than one cellular compartment, all of the categories were counted non-exclusively. The cellular distribution of identified proteins was reported as extracellular 18% (13 proteins), membrane 14% (10 proteins), nuclear 17% (12 proteins), cytoplasmic 22% (16 proteins), cytoskeletal 16% (12 proteins), endoplasmic
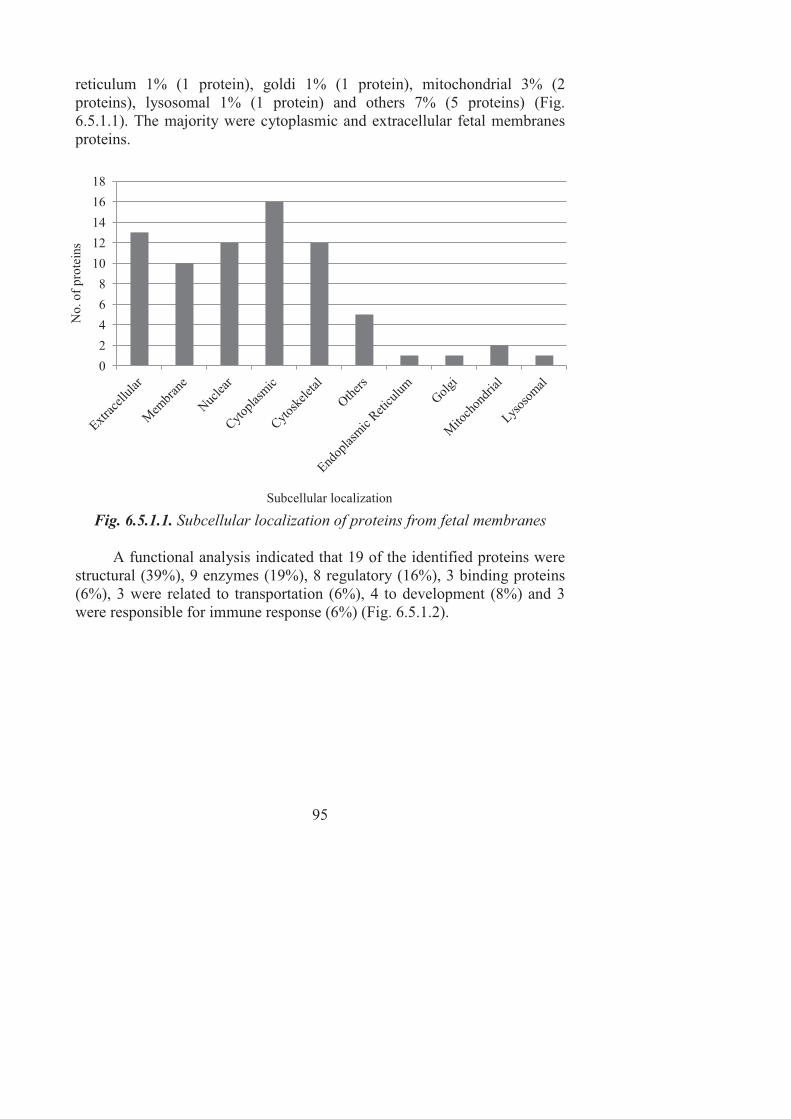
95
reticulum 1% (1 protein), goldi 1% (1 protein), mitochondrial 3% (2 proteins), lysosomal 1% (1 protein) and others 7% (5 proteins) (Fig. 6.5.1.1). The majority were cytoplasmic and extracellular fetal membranes proteins.
Fig. 6.5.1.1. Subcellular localization of proteins from fetal membranes
A functional analysis indicated that 19 of the identified proteins were
structural (39%), 9 enzymes (19%), 8 regulatory (16%), 3 binding proteins (6%), 3 were related to transportation (6%), 4 to development (8%) and 3 were responsible for immune response (6%) (Fig. 6.5.1.2).
0
2
4
6
8
10
12
14
16
18
No.
of
prot
eins
Subcellular localization

96
Fig. 6.5.1.2. Functional analysis of proteins from the fetal membranes
The comparison showed that 29 (67.4%) proteins were identical in both groups. 16 of them were reported earlier by Soo-,1.& !+"5& +.0& +-%e&alpha-enolase, annexin A2, collagen alpha-2(VI) chain, cytosol aminopep-tidase, glyceraldehydes-3-phosphate dehydrogenase, cytokeratin 5, cyto-keratin 8, cytokeratin 19, cytokeratin 24, macrophage migration inhibitory factor, peroxiredofin -6, pyruvate kinase isozymes M1, serum albumin, transforming growth factor-beta-induced protein ig-h3, triosephosphate isomerise, vimentin [61]. The authors performed proteome analysis of amnion at term using different system by 2D PAGE and MALDI-TOF/MS.
14 proteins were present only in the amniochorionic membrane during PPROM and not at term pregnancy: basement membrane-specific heparan sulfate proteoglycan core protein (perlecan), fibronectin, 40S ribosomal protein S7, 60S ribosomal protein L28, alpha-1-antitrypsin, DNA poly-merase alpha catalytic subunit, keratin, type I cytoskeletal 16, keratin, type I cytoskeletal 17, protein-glutamine gamma-glutamyltransferase, small nuclear ribonucleoprotein Sm D1, sulfate anion transporter, sulfhydryl oxi-dase, synaptotagmin-like protein and tenascin. Some proteins such as base-ment membrane-specific heparan sulfate proteoglycan core protein (perle-can) has already been recognized as potential biomarker of PPROM in am-niotic fluid by Vuadens and al. was also found in this study in fetal mem-branes with PPROM [70]. The presence of fetal fibronectin in cervico-vaginal fluid as indicator for the preterm labor was reported by other researchers previously [161]. Fibronectins are glycoproteins that connect cells with collagen fibres in the ECM. It is thought to be released through mechanical or inflammatory mediated damage to the membranes before
Structural; 39%
Enzymes; 19%
Regulatory; 16%
Development; 8%
Transportation; 6%
Immune responce;6%
Binding; 6%

97
preterm birth. Other proteins identified in this study have not yet been reported present in PPROM. The differences revealed in the fetal mem-branes proteome after proteins were classified by their function (Fig. 6.5.1.3).
Fig. 6.5.1.3. Differences of protein functions from the fetal membranes
As shown in Figure 6.5.1.3, significant differences were found in proteins with regulatory functions. Almost a 2 – fold increase in regulatory proteins, a 25% increase in number of structural proteins and a 30% increase in enzymes and were calculated in amniochorionic membranes with PPROM. This finding is not surprising as PPROM is expected to undergo a substantial structural changes regulated by enzymatic activity in the membranes.
6.5.2. Comparison of proteome from the fetal membranes and
amniotic fluid of patients with PPROM and term labor
with intact membranes
Manually analyzing amniotic fluid MS spectra 365 peptide like ave-
rage to high intensity signals were found in term delivery AF sample, and 347 peptide like average to high intensity signals in AF with PPROM
0 2 4 6 8 10 12 14 16 18 20
Binding
Immune responce
Transportation
Development
Regulatory
Enzymes
Structural
No. of proteins
Func
tions
PPROM
PPROM and term

98
sample. For further comparison only those peptides were selected which were identified more than two times in a consecutive runs. Identifying similarities between the AF peptides elution time was also used for matching in order to increase data confidence. Results are summarized in Table 6.5.2.1. Most of the identified peptides in amniotic fluid were shared between those two groups, however in case of the fetal membranes relatively more peptides were unique. Table 6.5.2.1 . Comparison of peptides numbers in AF and fetal membranes
Peptides in total Shared peptides Unique peptides
AF - Z 149 87 62
AF - R 146 59
M - Z 104 33 71
M - R 91 58
AF – amniotic fluid, M – amniochorionic membrane, R - red group, Z - green group.
The task was not only to match membranes compositions like in section 6.5.1, but also to find similarities between the PPROM membranes and amniotic fluid on the peptide level. Peptide comparison analysis revealed that the AF sample at term pregnancy has very little in common not only with PPROM membranes, but also with term membranes (Fig. 6.5.2.1). We expected such an observation as once the membranes are intact, they should not release or release very little peptides into the amniotic fluid. We found eight proteins matching peptides (two peptides unmatched) which were common for all AF and amniochorionic membranes samples (also see table 6.5.1.1): annexin A2, collagen alpha-2(I) chain, fibrinogen alpha chain, fibrinogen beta chain, glyceraldehyde-3-phosphate dehydrogenase, Ig gamma-1 chain C region, serum albumin and vimentin. The main focus was to found the same peptides in PPROM AF as in PPROM membranes, as those peptides could be as indicators of PPROM in AF. Unfortunately, out of eight peptides found in both PPROM AF and membranes we were able to identify only two proteins: protein-glutamine gamma-glutamyl transferase 2 and synaptotagmin-like protein 2.

99
Fig. 6.5.2.1. Similarities of fetal membranes and amniotic fluid peptides in women with PPROM and at term pregnancy with intact membranes AF – amniotic fluid, M – amniochorionic membrane, R – red group, Z – green group.
Soo-,1.&!+"5&et. al. also reported a low overlapping between proteins
found in amnion and amniotic fluid samples [61]. Only nine proteins were observed: serum albumin, apolipoprotein, serotranferrin, hemoglobin alpha and beta chains, profilin I, peptidyl-prolyl cis-trans isomerase A, actin and cytokeratin 1. The difference in the protein profile may be a gestational age-dependent phenomenon or due to the difference in the sample preparation and analytical method.
Our observation of proteins in fetal membranes opens a new area of research, in order to better understand the role of proteins in fetal membranes biology, to study the mechanisms involved in proteolysis of these proteins as well as to assess their usefulness as biological markers of PPROM. Future studies are needed to focus on the validation of these proteins as potential biomarkers of PPROM.
6.6. Comparison of biological samples peptidome and proteome
from patients with preterm premature rupture of membranes
The top ten proteins from the AF, plasma and fetal membranes pro-
teome were compared based on the Mascot score and by the number of matched peptides (Table 6.6.1).
0
2
4
6
8
10MzrAFzr
MzrAFz
MzrAFr
MzAFzr
MrAFzrMzAFr
MzAFz
MrAFz
MrAFr

100
Table 6.6.1. Comparison of top ten proteins from AF, plasma and fetal membranes Top 10 proteins from AF Top 10 from plasma Top 10 from fetal membranes
Alpha-1-antitrypsin Albumin Glyceraldehyde-3-phosphate dehydrogenase
Fibrinogen alpha chain Alpha-1-antitrypsin Fibronectin
Beta globin chain Apoliprotein A-I Collagen type I, alpha 2 and 1 chains
Collagen alpha-2(I) chain Apolipoprotein C-III Keratin, type I cytoskeletal 19
Lumican variant Serotransferin Albumin
U4/U6.U5 tri-snRNP-associated protein 1
Haptoglobin Keratin, type I cytoskeletal 14
'.+,&:#B#-#?&43T$+B1-=&
B member 11 Lipoprotein Collagen type VI, alpha-2 chain
DVL1 protein (Fragment) Fibrinogen Keratin, type I cytoskeletal 17
Ig gamma-1 chain C region Complement component 4B preproprotein
Alpha-enolase
Complement factor B precursor
Human complement C3 peptide
Transforming growth factor-beta-induced protein ig-h3
Differences in the proteome of biological samples were observed.
Only 24.7% (21 protein) of the AF proteins were also found in the fetal membranes proteome and 14.1% (12 proteins) in the plasma proteome. The results suggest that AF has a significantly different protein composition than plasma or fetal membranes. These differences should be associated with different functions of the biological fluids.
Out of ten patients with PPROM differentially expressed proteins were identified by comparing the protein profile from AF, plasma and fetal membranes. In amniotic fluid 63 proteins were identified from patients with PPROM, 53 proteins were present in both PPROM and control groups and ten proteins were found only in AF from patients with PPROM: vitamin D-binding protein; collagen alpha-1(I) chain; F-box only protein 40; synapto-tagmin-like protein 2; +G#-1G#G"#*61.& `b& O'/C& D9,U_\ML& $14E& @-#.6&NT2RP2001581; serine/threonine-protein phosphatase 2B catalytic subunit beta isoform; keratin, type I cytoskeletal 18; keratin, type I cytoskeletal 19; K-glypican.

101
Pereira et. al. reported vitamin D-binding protein to be present in cervico-vaginal fluid and up-regulated by a > 3-fold in spontaneous preterm birth [162]. Vitamin D-binding protein is found at low concentration in plasma [163]. This protein is present in amniotic fluid and therefore could be used as a potential biomarker for PPROM diagnosis or PTL prediction marker in cervicovaginal fluid. A chain of collagen I was found to be present in amniotic fluid samples with PPROM. This is the extracellular matrix protein which is a fundamental component of the basement membrane of amniotic membranes. Type I and III are the primary regulators of tensile strength of fetal membranes. Changes in its metabolism can contribute to preterm premature rupture of membranes.
F-box proteins are proteins containing at least one F-box domain and mediates ubiquitination of proteins targeted for degradation by the proteosome [164]. In PPROM cases it could be that F-box protein 40 is involved in the degradation of amniochorionic membrane proteins. Those proteins have also been associated with cellular functions such as signal transduction and regulation of the cell cycle that functions as a site of protein-protein interaction. Synaptotagmin-like protein 2 (SYTL2) isoform 1 plays a role in cytotoxic granule exocytosis in lymphocytes [165]. It is required for cytotoxic granule docking at the immunologic synapse. Isoform 4 binds phosphatidylserine and phosphatidylinositol-4,5-bisphosphate and promotes the recruitment of glucagon-containing granules to the cell membrane. O'/C&D9,U_\ML&$14E&@-#.6&/J_(!_NNUZQU&G"#*61.&14&5.#H.&*#&be present in the endoplasmic reticulum membrane, but the function is still unknown. Serine/threonine-protein phosphatase is enzyme involed in phosphorylation process of proteins and dephosphorylate serine and threo-nine residues in proteins [166]. Phosphorylation of structural and regulatory proteins is a major intracellular control mechanism in eukaryotes. The phosphorylation state of a protein is a dynamic process controlled by both protein kinases and protein phosphatases. K-glypican is from a family of glycosylphosphatidylinositol-anchored cell surface heparan sulfate proteo-glycans, involved in embryonic development [167]. Calgranulins A and B were not detected in any of studied PPROM cases with negative AF cul-*3"64%& J:14& "643-*& @#.$1"B4& !+"5& c,& @#.@-341#.E& H:1@:& +$$1"B4& *:+*& *:646&
calgranulins are reliable indicators of Ureaplasma infection.

102
CONCLUSIONS 1. In total, 836 peptides which were linked to 85 proteins have been
identified in amniotic fluid using a fully automated 2D liquid chro-matographic system coupled to mass spectrometry.
2. The amniotic fluid peptide and protein composition is dynamic and varies depending on the gestational age. Only 118 (42.3%) of peptides and 37 (43.5%) of identified proteins were consistently expressed throughout the pregnancy.
3. A total of 43 proteins from 195 peptides were identified from the digested fetal membranes. A comparison of proteomes of amnio-chorionic membranes with preterm premature rupture and term intact membranes revealed that 14 (32.6%) proteins were present only in PPROM fetal membranes with prevalent regulatory functions proteins. These proteins could be potential biomarkers of PPROM from fetal membranes.
4. Ten proteins from amniotic fluid were identified as potential biomar-kers for PPROM: vitamin D-binding protein; collagen alpha-1(I) chain; F-box only protein 40; synaptotagmin-like protein 2; apolipoprotein B; O'/C& D9,U_\ML& $14E& @-#.6& /J_(!_NNUZQUb& serine/threonine-protein phosphatase 2B catalytic subunit beta isoform; keratin, type I cyto-skeletal 18; keratin, type I cytoskeletal 19; K-glypican. In oder to va-lidate and confirm these proteins as potential biomarkers of PPROM additional studies are needed.
5. Amniotic fluid has a different peptide and protein composition than maternal plasma or fetal membranes. Only 24.7% (21 protein) of the amniotic fluid proteins were also found in the fetal membranes pro-teome and 14.1% (12 proteins) in the maternal plasma proteome.

103
BENEFITS OF THE RESEARCH AND POTENTIAL
FOR THE FUTURE
In order to identify patterns within the complex proteomic profile that can discriminate between normal and disease states it is essential to know the normal amniotic fluid peptidome and proteome. The created map of amniotic fluid peptides and proteins depending on the gestational age is im-portant for proteomics-based identification of biomarkers for fetal abnor-malities and other pregnancy complications. These peptides and proteins can be used to distinguish complicated cases or women carrying chromo-somally abnormal fetuses in other studies.
The identification of novel candidate peptide and protein biomarkers of PPROM from amniotic fluid and fetal membranes represents an important step forward in advancing our understanding of the physiologic mechanisms that lead to preterm premature rupture of membranes and preterm birth. These potential markers, although they still need to be veri-fied, could be diagnostic markers and may provide targets for therapeutic intervention in the future. It should be noticed that the findings should be considered preliminary until they are thoroughly validated in a larger cohort and by different analytical methods.

104
LIST OF ORIGINAL PUBLICATIONS
The present doctoral thesis is based on the following original communi-
cations:
1. d+@:*6f6<16.7& gE& P1.*+3*+4& aE& /+012+34516.7& (E& ;.?6"& SS%& Ge-
stational age-dependent changes of amniotic fluid proteome (Article in Lithuanian). Lithuanian Obstetrics and Gynecology. 2013; 16(1):9-17.
2. Machtejevien7 gE&P1.*+3*+4&aE&/+012+34516.7&(E&d1-1#*14&JE&;.?6"&SS%&Changes in the fetal membranes proteome composition as an indicator for their preterm premature rupture. Medicine (Kaunas), 2013, an article in print.
Related publications to the research:
Chapters in the books:
1. Machtejeviene E, Machtejevas E. Profiling of endogenous peptides by multidimensional liguid chromatography. In: Khan TM, editor. www. intechopen.com; 2012, p.225-246.
2. Machtejevas E, Machtejeviene E. Monolithic silica columns in multi-dimesional LC-MS for proteomics and peptidomics. In: Unger KK, Tanaka N, Machtejevas E, editors. Monolithic silicas in separation science. Concepts, syntheses, characterization, modeling and appli-cations. Weinheim: WILEY-VCH Verlag GmbH&Co.; 2011, p. 297-317.
3. d+@:*6f6<16.7 EE& `-+3h0i1).16.7 O. Preterm premature rupture of membranes (Art1@-6&1.&(3441+.W%&V.e&/+014+34516.6&(,E&601*#"%&c6-6@*1<6&questions in perinatology. Kaunas: KTC; 2012, p.113-118.
4. d+@:*6f6<16.7 EE&/+012+34516.7 R. Preterm labor (Article in Russian) . V.e& /+014+34516.6& (,E& 601*#"%& Selective questions in perinatology. Kaunas: KTC; 2012, p.101-112.
Articles in journals: 1. P"152+1*7 ,E& !+*"1T+<1j1)*7 AE& d+@:*6f6<16.7 E. Antenatal cortico-
steroids for accelerating fetal lung maturation for women at risk for preterm birth (Article in Lithuanian). Lithuanian Obstetrics and Gynecology. 2010;13(4):352-359.

105
2. d+@:*6f6<16.7 E, Rovas LE&(+B+2+345+1*7 DE&c3j5#<+ I, Gintautas V. The news in diagnostics and treatment of preterm labor (Article in Lit-huanian). Lithuanian Obstetrics and Gynecology. 2010;13(4):382-385.
3. d+@:*6f6<16.7 E, Nadi2+34516.7 RE& ,+532#<+1*7 I. Using human samples in proteomics. Ethics, social and regulatory aspects (Article in Lithuanian). Lithuanian Obstetrics and Gynecology. 2009;12(1):27-35.
Conferences:
1. Machtejevien7 E, Gintautas V, Nadišauskien7 R, Unger KK. Changes in the amniotic fluid and membranes composition as an indicator for their preterm premature rupture. EBCOG, Tallinn, 2012 (Oral prie-sentation).
2. d+@:*6f6<16.7 E. Etiology and pathogenesis of preterm labor. 9th Con-gress of Lithuanian Obstetrics and Gynaecology. Kaunas, 2010 (Oral presentation).
3. d+@:*6f6<16.7&gE&P1.*+3*+4&aE&d+@:*6f6<+4&gE&/+012+34516.7&(E&;.?6"&KK. Proteomics of amniotic fluid: the search for potential biomarkers of preterm premature rupture of membranes and preterm labor. The Science for Human Health, Kaunas, 2010 (Oral presentation). The presentation awarded with a special price.
4. Machtejevien7 E, Gintautas V, Nadišauskien7 R, Unger KK. Search of peptides as biomarkers of preterm premature rupture of the membranes in amniotic fluid. FIGO, Cape Town, 2009 (Poster).

106
REFERENCES
1. The United Nations Millennium Development Goals. Available at: http://www.un.org/millenniumgoals/childhealth.shtml. Accessed April 20, 2013.
2. 9+H.&,g, Gravett MG, Nunes TM, Rubens CE, Stanton C, the GAPPS Review Group. Global report on preterm birth and stillbirth (1 of 7): definitions, description of the burden and opportunities to improve data. BMC Pregnancy and Childbirth 2010,10(Suppl 1):S1.
3. 9+H.&,gE&k1-@h=.45+-Ketende K, Cousens SN. Estimating the causes of 4 million neonatal deaths in the year 2000. I.*& ,&gG106B1#-&_NNME&35(3):706-18.
4. Royal College of Obstetricians and Gynaecologists. Green-Top Guideline No. 44: Preterm prelabour rupture of membranes. London, RCOG, 2010. Available at: http://www.rcog.org.uk/files/rcog-corp/ GTG44PPROM28022011.pdf. Accessed April 20, 2013.
5. ACOG Committee on Practice Bulletins-Obstetrics. ACOG Practice Bulletin no. 80: premature rupture of membranes. Clinical mana-gement guidelines for obstetrician-gynecologists. Obstet Gynecol. 2007;109:1007-19.
6. The Society of Obstetricians and Gynaecologists of Canada. SOGC Clinical Practice guideline No. 233: Antibiotic Therapy in Preterm Premature Rupture of the Membranes. SOGC Clinical practice guideline. SOGC, 2009. Available at: http://sogc.org/wp-content/ uploads/2013/02/gui233CPG0909.pdf. Accessed April 20, 2013
7. Medical Data of Births, 2011. Vilnius, Institute of Hygiene Health Information Centre, 2012. Available at: http://sic.hi.lt/data/l11_gim. pdf. Accessed April 20, 2013.
8. Avery OT, MacLeod CM, McCarty M. Studies on the chemical nature of the substance inducing transformation of pneumonococcus types: induction of transformation by a desoxyribonucleic acid fraction isolated from pneumococcus type III. ,&glG&d60&1944; 79(2):137-58.
9. Semizarov D. Introduction: genomics and personalized medicine. In: Semizarov D, Blomme E, editors. Genomics in Drug Discovery and Development. /6H&,6"46=e&k1-6=b&_NNL%&G% 1–12.
10. O-+BG& dE& D"=& `E& S+B+-& dE& m16& mE& O3$$& ,E& 91.& dDE& 6*& +-%&
Distinguishing protein-coding and noncoding genes in the human genome. Proc Natl Acad Sci USA 2007;104(49):19428-33.

107
11. The Human Genome Project Information. Available at: http://www. ornl.gov/sci/techresources/Human_Genome/project/hgp.shtml. Acces-sed April 20, 2013.
12. International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature 2001; 409 (6822): 860-921.
13. Collins FS, Morgan M, Patrinos A. The Human Genome Project: Les-sons from Large-Scale Biology. Science 2003; 3005 (5617):286-290.
14. >3TT+"0&J,E&C56.&`9E&`6+-&SE&`+--64*6"&`E&O+@@+B#&dE&O:6.&nE&6*&
al. Nucleic Acids Res. 2007;35:D610-D617. 15. C4:3"4*&,9E&O:6.&OSE&P1-T6"*&,PE&,65#4@:&SE&S66.+.&cE&d610-&!E&6*&
al. Nucleic Acids Res. 2005;33:D459-D465. 16. Lottspeich F. Proteome analysis: a pathway to the functional analysis
of proteins. Angew Chem Int Ed Engl. 1999;38(17):2476-92. 17. Grabherr MG, Haas `,E& Yassour M, Levin ,oE& & Thompson DA,
Amit I, et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nature Biotechnology 2011;29:644-52.
18. k1-51.4&d(E&c+.@:6h&,OE&P##-6=&CCE&CGG6-&('E&>3BG:6"=-Smith I, Hochstrasser DF, Williams KL. Progress with proteome projects: why all proteins expressed by a genome should be identified and how to do it. Biotechnol Genet Eng Rev. 1996;13:19-50.
19. Mishra NC, editor. Introduction to proteomics: principles and +GG-1@+*1#.4%&/6H&,6"46=e&k1-6=b&_NUN%
20. Pandey A, Mann M. Proteomics to study genes and genomes. Nature 2000;405:837-846.
21. Liebler DC. The proteome. In: Liebler DC, editor. Introduction to Proteomics. Tools for the new biology. Humana Press; 2002.p.15-24.
22. Farrah T, Deutsch EW, Hoopmann MR, >+--#H4& ,9, Sun Z, Huang CY, et al. ,&!"#*6#B6&(64% The state of the human proteome in 2012 as viewed through PeptideAtlas. 2013;12(1):162-71.
23. Al-Sayah MA. New strategies for proteomics and peptidomics using polymer liquid crystals for electrophoresis. 2004; electronic thesis, treatises and dissertations. Paper 25. Available at: http://diginole.lib. fsu.edu/cgi/viewcontent.cgi?article=1024&context=etd. Accessed April 23, 2013.
24. Chandramouli K, Qian PY. Proteomics: challenges, techniques and possibilities to overcome biological sample complexity. Hum Geno-mics Proteomics. 2009;2009:ID239204. Available at: http://www.

108
ncbi.nlm.nih.gov/pmc/articles/PMC2950283/pdf/HGP2009-239204. pdf. Acceseed April 23, 2013.
25. Richter R, Schulz-Knappe P, Schrader M, et al. Composition of the peptide fraction in human blood plasma: data base of circulating human peptides. ,%&O:"#B+*#?"%&` 1999,726:25-35.
26. Zhu P, Bowden P, Zhang D, d+"4:+--& ,P. Mass spectrometry of peptides and proteins from human blood. Mass Spectrom Rev. 2010 Nov 2.
27. Biomarker. Available at: http://en.wikipedia.org/wiki/Biomarker. Accessed April 23, 2013.
28. Biomarkers Definitions Working Group. Biomarkers and surrogate endpoints: preferred definitions and conceptual framework. Clin Pharmacol Therapeut 2001;69:89-95.
29. Westermeier R, Naven T, Höpker HR, editors. Proteomics in practice: a guide to successful experimental design. 2nd ed. Wiley; 2008.
30. Pepe MS, Etz1#.1&(E& D6.?& oE& !#**6"& ,'E& J:#BG4#.&d9E&J:#".R314*&d%&!:+464&#$&T1#B+"56"&06<6-#GB6.*& $#"&6+"-=&06*6@*1#.&#$&@+.@6"%&,&
Natl Cancer Inst 2001;93(14):1054-61. 31. Diamandis EP. Cancer biomarkers: can we turn recent failures into
success? ,&/+*-&O+.@6"&V.4*&_NUNb102(19):1462-7. 32. Montagnana M, Danese E, Giudici S, Franchi M, Guidi GC, Plebani
M, Lippi G. HE4 in ovarian cancer: from discovery to clinical application. Adv Clin Chem. 2011;55:1-20.
33. Tsangaris GT, Anagnostopoulos AK, Tounta G, Antsaklis A, Mavrou A, Kolialexi A. Application of proteomics for the identification of biomarkers in amniotic fluid: are we ready to provide a reliable prediction? g!dC&,% 2011;2(2):149-55.
34. Clynen E, De Loof A, Schoofs L. The use of peptidomics in endocrine research. Gen Comp Endocr. 2003;132:1-9.
35. Govorukhina N, Bischoff R. Sample preparation of body fluids for proteomics analysis. In: Thongboonkerd V, editor. Proteomics of human body fluids: principles, methods, and applications. Totowa, /6H&,6"46=e&>3B+.+&!"644b&_NN\%&G% 31–71.
36. Medical research council. Human Tissue and Biological Samples for use in Research - Operational and Ethical guidelines. Available at: http://www.mrc.ac.uk/Utilities/Documentrecord/index. htm?d=MRC002420. Accessed February 12, 2009.

109
37. Reymond MA, Steinert R, Eder F, Lippert H. Ethical and regulatory issues arising from proteomic research and technology. Proteomics 2003;3:1387-1396.
38. !+""=&cE&c*"+344&,D&["0%&!"6B+*3"6&"3G*3"6&#$&*:6&$6*+-&B6BT"+.64%&N g.?-&,&d60&1998;338(10): 663-70.
39. Oyen ML, Calvin SE, Landers DV. Premature rupture of the fetal membranes: is the amnion the major determinant? CB& ,& pT4*6*&Gynecol 2006;195(2):510-5.
40. Bryant-Greenwood GD: The extracellular matrix of the human fetal membranes: structure and function. Placenta 1998;19(1):1-11.
41. Casey ML, MacDonald PC. Interstitial collagen synthesis and process-ing in human amnion: a property of the mesenchymal cells. Biol Reprod 1996;55(6):1253-60.
42. Malak TM, Ockleford CD, Bell SC, Dalgleish R, Bright N, Macvicar ,%&Confocal immunofluorescence localization of collagen types I, III, IV, V and VI and their ultrastructural organization in term human fetal membranes. Placenta 1993;14(4):385-406.
43. Rousselle P, Keene DR, Ruggiero F, Champliaud MF, Rest M, Burgeson RE. Laminin 5 binds the NC-1 domain of type VII collagen. ,&O6--&`1#- 1997;138(3):719-28.
44. !+4R316"&,O, Doret M. [Fetal membranes: embryological development, structure and the physiopathology of the preterm premature rupture of membranes]. ,&P=.6@#-&pT4*6*&`1#-&(6G"#0& 8!+"14W% 2008;37(6):579-88.
45. Verma RP, Hansch C. Matrix metalloproteinases (MMPs): Chemical–biological functions and (Q)SARs. Bioorganic & Medicinal Chemistry 2007;15:2223-68.
46. D#"*3.+*#& c,, Menon R, 9#BT+"01& c,. MMP/TIMP imbalance in amniotic fluid during PROM: an indirect support for endogenous pathway to membrane rupture. ,&!6"1.+*&d60 1999;27(5):362-8.
47. !+"5& S>E& O:+1H#"+G#.?4+& JE& S1B& ndE& g4G1.#h+& ,E& n#4:1B+*43& ,, Edwin S, et al. Matrix metalloproteinase 3 in parturition, premature rupture of the membranes, and microbial invasion of the amniotic @+<1*=%&,#3".+-&#$&!6"1.+*+-&[email protected]&_NN[b[UeU_-22.
48. P#-06.T6"?&(9E&>+3*:& ,OE&C.0"6H4&kk%&d6@:+.14B4& #$& 0146+46& - intrau*6"1.6& 1.$6@*1#.& +.0& G"6*6"B& 06-1<6"=%&/6H&g.?-+.0& ,#3".+-& #$&Medicine 2000;342:1500-7.
49. Moore CS, O"#@56"&c,. An alternate perspective on the roles of TIMPs and MMPs in pathology. CB&,&!+*:#- 2012;180(1):12-6.

110
50. D#"*3.+*#& c,E& d6.#.& (E& 9#BT+"01& c,%& !"646.@6& #$& $#3"& *14436&
inhibitors of matrix metalloproteinases (TIMP-1, -2, -3 and -4) in :3B+.&$6*+-&B6BT"+.64%&CB&,&(6G"#0&VBB3.#-&ULLQb^Ne[LZ–400.
51. Riley SC, Leask R, Denison FC, Wisely K, Calder AA, et al. Secretion of tissue inhibitors of matrix metalloproteinases by human $6*+-& B6BT"+.64E& 06@103+& +.0& G-+@6.*+& +*& G+"*3"1*1#.%& ,#3".+-& #$&
Endocrinology 1999;162:351-9. 52. Riley SC, Leask R, Chard T, Wathen NC, Calder AA, Howe DC.
Secretion of matrix metalloproteinase-2, matrix metalloproteinase-9 and tissue inhibitor of metalloproteinases into the intrauterine com-partments during early pregnancy. Molecular human reproduction 1999;5:376-81.
53. Brace RA. Physiology of amniotic fluid volume regulation. Clin Obstet Gynecol. 1997;40:280-9.
54. Underwood MA, Gilbert WM, Sherman MP. Amniotic fluid: not just fetal urine anymore. ,&!erinatol 2005;25(5):341-8.
55. Fischer R. Amniotic fluid: physiology and assessment. The global library of women's medicine. Available at: http://www.glowm.com/ section_view/item/208. Accessed April 24, 2013.
56. Modena AB, Fieni S. Amniotic fluid dynamics. Acta Bio Medica Ateneo Parmense 2004;75 Suppl. 1:11-3.
57. Weber G, Merz E. Amniotic fluid. In: Merz E, editor. Ultrasound in obstetrics and gynecology Vol.1:Obstetrics. 2nd ed. Stuttgard:Thieme; 2005.
58. Perluigi M, Di Domenico F, Cini C, Coccia R, Giorlandino FR, Giorlandino M, Cignini P, Mesoraca A, Giorlandino C. Proteomic analysis for the study of amniotic fluid protein composition. ,&!"6.+*&Med. _NNL&,3-b[8[We[L-41.
59. Liberatori S, Bini L, De Felice C, Magi B, Marzocchi B, Raggiaschi R, et al. A two-dimensional protein map of human amniotic fluid at 17 weeks’ gestation. Electrophoresis 1997;18(15):2816-22.
60. /1-44#.& cE& (+B4*"#B& dE& !+-BT-+0& dE& Cl6-44#.& pE& `6"?R314*& ,%&
Explorative study of the protein composition of amniotic fluid by liquid chromatography electrospray ionization Fourier transform ion @=@-#*"#.& "64#.+.@6&B+44& 4G6@*"#B6*"=%& ,&!"#*6#B6&(64&_NN^b[eQQ^-9.
61. !+"5& c,E& n##.& kPE& c#.?& ,cE& ,3.?& >cE& S1B& O,E& p:& cnE& 6*& +-%&
Proteome analysis of human amnion and amniotic fluid by two-dimensional electrophoresis and matrix-assisted laser desorption/

111
ionization time-of-flight mass spectrometry. Proteomics 2006;6:349-63.
62. d1@:6-&!gE&O"6**+h&'E&d#"16"&!E&>6--6"&dE&P+--#*&'E&J144#*&,'E&6*&+-%&
Proteome analysis of human plasma and amniotic fluid by Off-Gel isoelectric focusing followed by nano-LC-MS/MS. Electrophoresis 2006;27:1169-81.
63. Tsangaris GT, Kolialexi A, Karamessinis PM, Anagnostopoulos AK, Antsaklis A, Fountoulakis M, et al. The normal human amniotic fluid supernatant proteome. In Vivo 2006;20:479-90.
64. O:#&OSE&c:+.&c,E&k1.4#"&g,E&'1+B+.014&g!%&!"#*6#B1@4&+.+-=414&#$&
human amniotic fluid. Mol Cell Proteomics 2007;6:1406-15. 65. O"6**+h&'E&J:+0155+"+.&9E&P+--#*&'E&K36-#h&!CE&c+G1.&aE&(#4416"&,cE&
et al. Proteomics of amniotic fluid. In: Thongboonkerd V, editor. Proteomics of human body fluids: principles, methods, and +GG-1@+*1#.4%&&J#*#H+E&/6H&,6"46=e&>3B+.+&!"644b&_NN\%&G% 415-436.
66. d1@:+6-4&,gE&'+4+"1&cE&!6"61"+&9E&(600=&C!E&9+G1034&,CE&93&mE&,+@#T&
JE&J:#B+4&CE&(#0-+.0&dE&(#T6"*4&OJ&,"E&Gravett MG, Nagalla SR. Comprehensive proteomic analysis of the human amniotic fluid proteome: gestational age-06G6.06.*&@:+.?64%&,&!"#*6#B6&(64&&_NN\b 6(4):1277-85.
67. K36-#h&!CE&O"6**+h&'E&J:+0155+"+.&9E&c+G1.&aE&P+--#*&'E&,+.1&,E&6*&+-%&
Proteomic analysis of amniotic fluid: potential applications in health +.0& 0146+464%& ,& O:"#B+*#?"& `& C.+-=*& J6@:.#-& `1#B60& 91$6& c@1&
2007;850:336-42. 68. Kolialexi A, Tsangaris GT, Mavrou A. Proteomics in prenatal
diagnosis. Expert Rev Proteomics 2009;6(2):111-3. 69. p:& S,E& !+"5& ,cE& /#"H1*h& g(E& S1B& cdE& S1B& `,E& !+"5& OkE& 6*& +-%&
Proteomic biomarkers in second trimester amniotic fluid that identify women who are destined to develop preeclampsia. Reproductive Sciences 2012;19:694-703.
70. Vuadens F, Benay C, Crettaz D, Gallot D, Sapin V, Schneider P, et al. Identification of biologic markers of the premature rupture of fetal membranes: proteomic approach. Proteomics 2003;3:1521-5.
71. Thadikkaran L, Crettaz D, Siegenthaler MA, Gallot D, Sapin V, Iozzo RV, et al. The role of proteomics in the assessment of premature rupture of fetal membranes. Clin Chim Acta 2005;360:27-36.
72. Anderson NL, Polanski M, Pieper R, Gatlin T, Tirumalai RS, Conrads TP, et al. The human plasma proteome: a nonredundant list developed

112
by combination of four separate sources. Mol. Cell. Proteomics 2004;3:311-26.
73. P+:-& kCE& S#h1.+& J,E& D3:"B+..& ''E& a+-6& Cd%& Obstet. Gynecol. 1982;60:297-304.
74. Gaucherand P, Guibaud S, Awada A, Rudigoz RC. Comparative study of three vaginal markers of the premature rupture of membranes. Insulin like growth factor binding protein, 1 diamineoxidase, and pH. Acta Obstet Gynecol Scand. 1997;76:536-40.
75. '+4+"1& cE& !6"61"+& 9E& (600=&C!E&d1@:+6-4& ,gE& 93&mE& ,+@#T& JE& 6*& +-% O#BG"6:6.41<6& G"#*6#B1@& +.+-=414& #$& :3B+.& @6"<1@+-q<+?1.+-& $-310%&
Proteome Res. 2007;6:1258-268. 76. c:+H& ,9aE& cB1*:&O(E&'1+B+.014& g!%& !"#*6#B1@& +.+-=414& #$& :3B+.&
cervico-<+?1.+-& $-310%& ,#3".+-& #$& !"#*6#B6& (646+"@:& _NN\bMe_QZL-2865.
77. Wang T, Zhou R, Zhang L, Wang Y, Song Cp, Lin W, et al. Proteins in leaked amniotic fluid as biomarkers diagnostic for prelabor rupture of membranes. Proteomics Clin Appl. 2011;5(7-8):415-21.
78. P"+<6**&dPE&/#<=&d,E&(#46.$6-0&(PE&(600=&C!E&,+@#T&JE&J3".6"&dE&
et al. Diagnosis of intra-amniotic infection by proteomic profiling and 106.*1$1@+*1#.&#$&.#<6-&T1#B+"56"4%&,CdC%&_NN^b_L_e^M_–9.
79. Gravett dPE&k1*51.& ccE&>+-345+&P,E& g0H+"04& ,9E& O##5&d,E&/#<=&d,%& C.& 6lG6"1B6.*+-&B#06-& $#"& 1.*"++B.1#*1@& 1.$6@*1#.& +.0& G"6*6"B&
-+T#"&1.&":6434&B#.56=4%&CB&,&pT4*6*&P=.6@#-&ULL^bU\U8MWeUMMN–7. 80. Ruetschi U, Rosen A, Karlsson G, Zetterberg H, Rymo L, Hagberg H,
et al. Proteomic analysis using protein chips to detect biomarkers in cervical and amniotic fluid in women with intra-amniotic inflame-B+*1#.%&,&!"#*6#B6&(64%&_NNZb&^e__[M-42.
81. `3f#-0& gE& (#B6"#& (E& S34+.#<1@& ,!E& g"6h& pE& P#*4@:& DE& O:+1H#"+-pongsa T, et al. Proteomic profiling of amniotic fluid in preterm labor using two-01B6.41#.+-& -1R310& 46G+"+*1#.& +.0& B+44& 4G6@*"#B6*"=%& ,&Matern Fetal Neonatal Med. 2008;21:697-713.
82. Lee SE, Han BD, Park IS, Romero R, Yoon BH. Evidence supporting proteolytic cleavage of insulin-like growth factor binding protein-1 (IGFBP-UW&G"#*61.&1.&+B.1#*1@&$-310%&,&!6"1.+*&d60&_NNQb[Me[UM–323.
83. Buhimschi IA, Christner R, Buhimschi CS. Proteomic biomarker analysis of amniotic fluid for identification of intra-amniotic 1.$-+BB+*1#.%&`,pP%&_NNZb&UU_e173-81.
84. Buhimschi CS, Bhandari V, Hamar BD, Bahtiyar MO, Zhao G, Sfakianaki AK, et al. Proteomic profiling of the amniotic fluid to

113
detect inflammation, infection, and neonatal sepsis. PLoS Med. 2007; 4:e18.
85. Buhimschi CS, Rosenberg VA, Dulay AT, Thung S, Sfakianaki AK, Bahtiyar MO, et al. Multidimensional system biology: genetic markers and proteomic biomarkers of adverse pregnancy outcome in preterm birth%&CB&,&!6"1.+*#-%&2008; 25:175-87.
86. Buhimschi CS, Bhandari V, Han YW, Dulay AT, Baumbusch MA, d+0"1&,CE&`3:1B4@:1&VC%&;41.?&G"#*6#B1@4&1.&G6"1.+*+-&+.0&.6#.+*+-&
sepsis. Hopes and challenges for the future. Curr. Opin. Infect. Dis. 2009;22:235-43.
87. Buhimschi CS, Dulay AT, Abdel-Razeq S, Zhao G, Lee S, Hodgson g,, et al. Fetal inflammatory response in women with proteomic biomarkers characteristic of intra-amniotic inflammation and preterm T1"*:%&`,pP%&_N09;116(2):257-67.
88. Buhimschi IA, Zhao G, Rosenberg VA, Abdel-Razeq S, Thung S, Buhimschi CS. Multidimensional proteomics analysis of amniotic fluid to provide insight into the mechanisms of idiopathic preterm birth. PLoS ONE. 2008;3:e2049.
89. Buhimschi IA, Zambrano E, Pettker CM, et al. Using proteomic analysis of the human amniotic fluid to identify histologic chorioam-nionitis. Obstet Gynecol. 2008; 111:403-12.
90. Buhimschi CS, Buhimschi IA, Abdel-Razeq S, Rosenberg VA, Thung SF, Zhao G, et al. Proteomic biomarkers of intra-amniotic inflame-mation: relationship with funisitis and early-onset sepsis in the pre-mature neonate. Pediatr Res. 2007;61:318-24.
91. Buhimschi IA, Buhimschi CS. Proteomics of the amniotic fluid in assessment of the placenta. Relevance for preterm birth. Placenta. 2008;29(Suppl A):S95-101.
92. Buhimschi IA, Zhao G, Pettker CM, Bahtiyar MO, Magloire LK, Thung S, et al. The receptor for advanced glycation end products (RAGE) system in women with intraamniotic infection and inflame-B+*1#.%&CB&,&pT4*6*&P=.6@#-%&_NN\b&ULMeUQU%6U-13.
93. Buhimschi CS, Baumbusch MA, Dulay AT, Oliver EA, Lee S, Zhao G, et al. Characterization of RAGE, HMGB1 and S100ß in inflame-B+*1#.& 1.03@60& G"6*6"B& T1"*:& +.0& $6*+-& *14436& 1.f3"=%& CB%& ,%& !+*:#-%&
2009;175:958-75. 94. (#B6"#& (E& S34+.#<1@& ,!E& P#*4@:& DE& g"6h& pE& a+14T3@:& gE& d+h+51-
Tovi S, et al. Isobaric labeling and tandem mass spectrometry: a novel approach for profiling and quantifying proteins differentially ex-

114
pressed in amniotic fluid in preterm labor with and without intraa-B.1#*1@& 1.$6@*1#.Y1.$-+BB+*1#.%& ,&d+*6".&D6*+-&/6#.+*+-&d60%&_NUNb 23:261-80.
95. Anderson NL, Anderson NG. The human plasma proteome: history, character, and diagnostic prospects. Mol. Cell. Proteomics 2002;1: 845-67.
96. Deutsch EW, g.?&,S, Zhang H, King NL, Nesvizhskii AI, Lin B, et al. Human Plasma PeptideAtlas. Proteomics. 2005;5(13):3497-500.
97. Desiere F, Deutsch EW, King NL, Nesvizhskii AI, Mallick P, g.?&,, et al. The PeptideAtlas project. Nucleic Acids Res. 2006;34(Database issue):D655-8.
98. Peptide Atlas. Available at: http://www.peptideatlas.org/. Accessed April 24, 2013.
99. Deutsch EW, Lam H, Aebersold R. PeptideAtlas: a resource for target selection for emerging targeted proteomics workflows. EMBO Rep. 2008;9(5):429-34.
100. D+""+:&JE&'63*4@:&gkE&pB6..&PcE&O+BGT6--&'cE&c3.&oE&`-6*h&,CE&6*&al. A high-confidence human plasma proteome reference set with estimated concentrations in PeptideAtlas. Mol. Cell. Proteomics 2011;10(9):M110.006353.
101. Tammen H, Schulte I, Hess R, Menzel C, Kellmann M, Mohring T, Schulz-Knappe P. Peptidomic analysis of human blood specimens: Comparison between plasma specimens and serum by differential peptide display. Proteomics 2005;5:3414-22.
102. Tiselius A. The moving-boundary method of studying the electrophoresis of proteins. Nova Acta Regiae Societatis Scientiarum Upsaliensis 1930;Ser. IV, Vol.7, No.4.
103. O’Farrell PH. High resolution two-dimensional electrophoresis of proteins. ,&`1#-&O:6B&1975, 250, 4007-21.
104. >3& cE& 9##& ,CE& k#.?& 'J. Human body fluid proteome analysis. Proteomics 2006;6:6326–6353
105. Machtejevas E, Machtejeviene E. Monolithic silica columns in multi-dimesional LC-MS for proteomics and peptidomics. In: Unger KK, Tanaka N, Machtejevas E, editors. Monolithic silicas in separation science. Concepts, syntheses, characterization, modeling and appli-cations. Weinheim: Wiley-VCH; 2011. p. 297-317.
106. Machtejevas E, Unger KK. Peptidomics. In: Cohen SA, Schure MR, editors. Multidimensional liquid chromatography: theory and appli-

115
cations in industrial chemistry and life sciences. New ,6"46=e&k1-6=-VCH; 2008. p. 207-219.
107. Tatemoto K. Neuropeptide Y: Complete amino acid sequence of the brain peptide. Proc. Natl. Acad. Sci. USA 1982;79:5485-9.
108. Dart LL, Smith DM, Meyers CA, Sporn MB, Frolik CA. Transform-ing growth factors from a human tumor cell: characterization of transforming growth factor ß and identification of high molecular weight transforming growth factor Ó. Biochem. 1985;24:5925-31.
109. Premstaller A, Oberacher H, Walcher W, Timperio AM, Zolla L, O:6"<6*& ,-P, et al. High-performance liquid chromatography-electrospray ionization mass spectrometry using monolithic capillary columns for proteomic studies. Anal. Chem. 2001;73:2390-6.
110. Westerlund D. Direct injection of plasma into column liquid chro-matographic systems. Chromatographia 1987;24:155-64.
111. Machtejevas E, Unger KK. Sample preparation for HPLC – based G"#*6#B6& +.+-=414%& V.e& <#.& >+?6.& ,E& 601*#"%& !"#*6#B1@4& 4+BG-6&
preparation. Weinheim:Wiley-VCH; 2008. p.245-64 112. ,#:.&>E&k+-06.&dE& c@:+$er S, Genz S, Forssmann WG. Analytical
procedures for quantification of peptides in pharmaceutical research by liquid chromatography–mass spectrometry. Anal Bioanal. Chem. 2004;378,883-97.
113. C.0"6@:*& cE& C.06"4& ,E& >6.0"154& (E& d+@:*6f6<+4& gE& ;.?6"& S%& g$$1-ziente Probenvorbereitung fuer die Proteomanalyse von Koerper-fluessigkeiten. Laborwelt 2004;5:4-7.
114. Li S, Dass C. Iron(III)-immobilized metal ion affinity chromatography and mass spectrometry for the purification and characterization of synthetic phosphopeptides. Anal. Biochem. 1999;270:9-14.
115. Gygi SP, Rist B, Griffin J,E&g.?&,E&C6T6"4#-0 R. Proteome analysis of low-abundance proteins using multidimensional chromatography and isotope-coded affinity tags. ,%&!"#*6#B6&(64%&_NN_bUe47-54.
116. Quaglia M, Machtejevas E, Hennessy T, Unger KK. Gel filtration/ size-exclusion chromatography (SEC) of biopolymers – optimization strategies and trouble shouting. In: Kromidas S, editor. HPLC made to measure – a practical handbook for optimization. 2006 Wiley-VCH, Weinheim, Germany, 383-403.
117. Wagner K, Miliotis T, Marko-Varga G, Bischoff R, Unger KK. An automated on-line multidimensional HPLC system for protein and peptide mapping with integrated sample preparation Anal. Chem. 2002;74:809-20.

116
118. Willemsen O, Machtejevas E, Unger KK. Enrichment of proteinceous materials on a strong cation-exchange diol silica restricted access material (RAM): protein-protein displacement and interaction effects. ,%&O:"#B+*#?"+G:=&C%&_NN^bUN_Ze_NL-216.
119. Machtejeviene E, Machtejevas E. Profiling of endogenous peptides by multidimensional liguid chromatography. In: Khan TM, editor. www.intechopen.com; 2012, p. 225-246 and references therein.
120. Medical Data of Births, 2011 (p.25-26). Vilnius, Institute of Hygiene Health Information Centre, 2012. Available at: http://sic.hi.lt/data/ l11_gim.pdf. Accessed April 20, 2013.
121. c-+**6"=&ddE&d#""14#.&,&,%&!"6*6"B&06-1<6"=%&9+.@6*&_NN_b[MNeU^QL-1497.
122. Goldenberg (9E&O3-:+.6&,DE&V+B4&,'E&(#B6"#&(%&gG106B1#-#?=&+.0&causes of preterm birth. Lancet 2008;371:75-84.
123. C.+.*:& OaE& ,#46G:& ScE& p=6-646& nE& '6B14416& SE& a1.*h1-6#4& Cd%&Trends in preterm birth and perinatal mortality among singletons: United States, 1989 through 2000. Obstet Gynecol 2005;105:1084-91.
124. ,+@54#.&(CE&P1T4#.&SCE&k3&nkE&O"#3?:+.&dc%&!6"1.+*+-*@#B64&in singletons following in vitro fertilization: a meta–analysis. Obstet Gynecol 2004;103:551-63.
125. EURO-PERISTAT. Available at: http://www.europeristat.com. Ac-cessed April 25, 2013.
126. Royal College of Obstetricians and Gynaecologists. Green-Top Guideline No. 7: Antenatal corticosteroids to reduce neonatal mor-bidity and mortality. London, RCOG, 2010. Available at: http:// www.rcog.org.uk/files/rcog-corp/GTG%207.pdf. Accessed April 20, 2013.
127. d6"@6"&`dE&P#-06.T6"?&(9E&d614&!,E&Moawad AH, Shellhaas C, Das A, et al. The preterm prediction study: prediction of preterm prie-mature rupture of membranes through clinical findings and ancillary testing. CB&,&pT4*6*&P=.6@#-&2000;183:738-45.
128. Mercer BM, Goldenber?& (9E&d#+H+0& C>E&d614& !,E& V+B4& ,'E& '+4&AF, et al. The preterm prediction study: effect of gestational age and @+346&#$&G"6*6"B&T1"*:&#.&43T46R36.*&#T4*6*"1@*@#B6%&CB&,&pT4*6*&
Gynecol 1999;181:1216-21. 129. Mercer B. Preterm premature rupture of the membranes. The Global
library of women's medicine. Available at: http://www.glowm.com/ section_view/item/120#5541. Accessed April 24, 2013.

117
130. Kistka ZA, DeFranco EA, Ligthart L, Willemsen G, !-3.56**& ,, d3?-1+& 9,, Boomsma DI. Heritability of parturition timing: an 6l*6.060& *H1.& 0641?.& +.+-=414%& CB& ,& pT4*6*& P=.6@#-& _NNQbULL8UWe 43.e1-43.e5.
131. Winkvist A, Mogren I, Hogberg U. Familial patterns in birth cha-racteristics: impact on individual and population risks. V.*&,&gG106B1#-&1998;27:248-54.
132. Kistka ZA-F, Palomar L, Lee KA, Boslaugh SE, Wangler MF, Cole FS, et al. Racial disparity in the frequency of recurrence of preterm birth. CB&,&pT4*6*&P=.6@#-&_NN\bU96(2):131.e1-131.e6.
133. Ananth CV, Vintzileos AM. Epidemiology of preterm birth and its @-1.1@+-&43T*=G64%&,&d+*6".&D6*+-&/6#.+*+-&d60&_NNMbULe\\[-82.
134. cB1*:& 9SE& '"+G6"& gcE& d+.5*6-#H& `/E& '#"-1.?& ,cE& D16-0& ',%&Socioeconomic inequalities in very preterm birth rates. Arch Dis Child Fetal Neonatal Ed 2007;92:F11-4.
135. Nohr EA, Bech BH, Vaeth M, Rasmussen KM, Henriksen TB, Olsen ,%&Obesity, gestational weight gain and preterm birth: a study within the danish national birth cohort. Paediatr Perinat Epidemiol 2007; 21:5-14.
136. El Khwad M, Stetzer B, Moore RM, Kumar D, Mercer B, Arikat S, et al. Term human fetal membranes have a weak zone overlying the lower uterine pole and cervix before onset of labor. Biol Reprod 2005;72:720-6.
137. !+""=&cE&c*"+344&,D%&!"6B+*3"6&"3G*3"6&#$&*:e fetal membranes. N Engl ,&d60&ULLQb&[[QeMM[-70.
138. (#B6"#&(E&g4G1.#h+&,E&S34+.#<1@&,!E&Gotsch F, Hassan S, Erez O, et +-%&J:6&G"6*6"B&G+"*3"1*1#.&4=.0"#B6%&`"&,&pT4*6*&P=.+6@#-&_NNMb113 Suppl 3:17-42.
139. P#-06.T6"?&(9E&>+3*:& ,OE&C.0"6H4&kk%& V.*"+3*6"1.6& 1.$6@*1#.& +.0&G"6*6"B&06-1<6"=%&/&g.?-&,&d60&_NNNb[^_eUZNN-7.
140. Goncalves LF, Chaiworapongsa T, Romero R. Intrauterine infection and prematurity. Ment. Retard. Dev. Disabil. Res. Rev. 2002;8: 3–13.
141. Cobo T, Kacerovsky M, Palacio M, Hornychova H, Hougaard DM, et al. (2012) Intra-Amniotic Inflammatory Response in Subgroups of Women with Preterm Prelabor Rupture of the Membranes. PLoS ONE 7(8): e43677. doi:10.1371/journal.pone.0043677.
142. Gomez R, Romero R, Mazor M, Ghezzi F, David C, Yoon BH. The role of infection in preterm labor and delivery. In: Elder M, Romero

118
R, Lamont R, editors. Preterm Labor. London: Churchill Livingstone, 1997:85-125.
143. >+"?6"& ,>E&>41.?&CkE& J3#B+-+&(gE&Gibbs RS, Mead PB, Eschen-bach DA, et al. Risk factors for preterm premature rupture of fetal membranes: a multicenter case-@#.*"#-& 4*30=%&CB&,&pT4*6*&P=.6@#-%&1990;163:130-7.
144. Erdemoglu E, Mungan T. Significance of detecting insulin-like growth factor binding protein-1 in cervicovaginal secretions: comparison with nitrazine test and amniotic fluid volume assessment. Acta Obstet Gynecol Scand. 2004;83:622-6.
145. !+"5&,cE&966&cgE&/#"H1*h&g(%&/#.-invasive testing for rupture of the fetal membranes. Touch Briefings: US Obstetrics and Gynecology 2007;Issue 1:13-6.
146. O:6.& DOSE& '306.:+346.& ,k%& O#BG+"14#.& #$& *H#& "+G10& 4*"1G& *64*4&based on IGFBP-1 and PAMG-1 for the detection of amniotic fluid. CB&,&!6"1.+*#-&_NNQb_Ze243-6.
147. El-Messidi A, Cameron A. Diagnosis of premature rupture of mem-T"+.64e& 1.4G1"+*1#.&$"#B&*:6&G+4*&+.0& 1.41?:*4& $#"& *:6&$3*3"6%&,&pT4*6*&
Gynaecol Can 2010;32(6):561-9. 148. Tagore S, Kwek K. Comparative analysis of insulin-like growth factor
binding protein-1 (IGFBP-1), placental alpha-microglobulin-1 (PAMG-1) and nitrazine test to diagnose premature rupture of membranes in pregnancy. ,&!6"1.+*&d60% 2010;38(6):609-12.
149. V+B4&,'E&(#B6"#&(E&O3-:+.6&,DE&Goldenberg RL. Primary, secondary, and tertiary interventions to reduce the morbidity and mortality of preterm birth. Lancet 2008;371:164-175.
150. Odibo AO, Talucci M, Berghella V. Prediction of preterm premature rupture of membranes by transvaginal ultrasound features and risk factors in a high-risk population. Ultrasound Obstet Gynecol. 2002;20(3):245-51.
151. d6"@6"&`dE&P#-06.T6"?&(9E&d614&!,E&Moawad AH, Shellhaas C, Das A, et al. The preterm prediction study: prediction of preterm premature rupture of the membranes using clinical findings and an-cillary testing. The national institute of child health and human deve-lopment maternal-fetal medicine units network. CB&,&pT4*6*&P=.6@#-% 2000 Sep;183(3):738-45.
152. Machtejevas E, Marko-Varga G, Lindberg C, Lubda D, Hendriks R, Unger KK. Profiling of endogenous peptides by multidimensional

119
liquid chromatography: on-line automated sample cleanup procedures for biomarker discovery in human urine. ,%&c6G%&c@1%&_NNLb[_e___[-32.
153. Unger KK, Machtejevas E. Peptid-Profilanalyse aus Körperflüs-sigkeiten. ChromChat 2009;4:44-47.
154. M+@:*6f6<+4&gE&'6.#=6-&(E&d6.6464& ,dE&S301"5+1*7&aE&P"1B64&`CE&Lubda D, Unger KK. Sulphonic acid strong cation-exchange restricted access columns in sample cleanup for profiling of endogeneous peptides in multidimensional liquid chromatography: structure and function of strong cation-exchange restricted access materials. ,%&Chromatogr. A 2006;1123:38-46.
155. Skudas R, Grimes BA, Machtejevas E, Kudirkaite V, Kornysova O, Hennessy TP, et al. Impact of pore structural parameters on column performance and resolution of reversed-phase monolithic silica @#-3B.4& $#"& G6G*1064& +.0& G"#*61.4%& ,%& O:"#B+*#?"%& C& _NN\bUU^^e\_-84.
156. Matrix Science. Available at: http://www.matrixscience.com/help/ scoring_help.html. Accessed April 24, 2013.
157. `-+@56**& !(E& d@O#.+*:=& k,%& O#BG+"14#.& #$& -1G104& +.0& +G#-1G#-proteins in amniotic fluid, neonatal urine, and cord serum. Ann. Clin. Lab. Sci. 1982;12(4):288-95.
158. Suginta W, Karoulias N, Aitken A, Ashley RH. Chloride intracellular channel protein CLIC4 (p64H1) binds directly to brain dynamin I in a complex containing actin, tubulin and 14-3-3 isoforms. `1#@:6B%& ,% 2001;359(Pt1):55-64.
159. Wu S, Levenson A, Kharitonenkov A, De Luca F. Fibroblast growth factor 21 (FGF21) inhibits chondrocyte function and growth hormone action directly at the growth plate. ,&`1#-&O:6B% 2012;287(31):26060-7.
160. Casey ML, MacDonald PC. Interstitial collagen synthesis and pro-cessing in human amnion: a property of the mesenchymal cells. Biol Reprod 1996;55(6):1253-60.
161. DeFranco EA, Lewis DF, Odibo AO. Improving the screening accuracy for preterm labor: is the combination of fetal fibronectin and cervical length in symptomatic patients a useful predictor of preterm birth? A systematic review. CB&,&pT4*6*&P=.6@#-. 2013 Mar;208(3): 233.e1-6.
162. !6"61"+&9E&(600=&C!E&,+@#T&JE&J:#B+4&CE&c@:.6106"&SCE Dasari S, et al. Identification of novel protein biomarkers of preterm birth in human cervical-vaginal fluid. ,&!"#*6#B6&(64. 2007;6(4):1269-76.

120
163. Tirumalai RS, Chan KC, Prieto DA, V44+R&>,E&O#."+04&J!E&a66.4*"+&TD. Characterization of the low molecular weight human serum proteome. Mol. Cell. Proteomics 2003;2:1096–1103.
164. Kipreos ET, Pagano M. The F-box protein family. Genome Biol. 2000;1(5): reviews3002-reviews3002.7. Available at: http://genome biology.com/2000/1/5/reviews/3002. Accessed April 25,2013.
165. Gálvez-Santisteban M, Rodriguez-Fraticelli AE, Bryant DM, Vergara-jauregui S, Yasuda T, Bañón-Rodríguez I, et al. Synaptotagmin-like proteins control the formation of a single apical membrane domain in epithelial cells. Nat Cell Biol. 2012;14(8):838-49.
166. Wera S, Hemmings BA. Serine/threonine protein phosphatases. `1#@:6B&,%&1995;311(Pt 1):17–29.
167. Watanabe K, Yamada H, Yamaguchi Y. K-glypican: a novel GPI-anchored heparan sulfate proteoglycan that is highly expressed in developing brain and kidney. ,&Cell Biol. 1995;130(5):1207-18.

121
ACKNOWLEGMENTS
I would like to express my sincere appreciation and gratitude to all who have contributed to this work.
Special thanks to my supervisor, Prof. 9:%.";.<)=.3>?)$*@, Head of the Department of Obstetrics and Gynecology, Lithuanian University of Health Sciences, for great inspiration to start research work and your guidance in real science, encouregement, support and belief in me. Thank you, for sharing your scientific and clinical knowledge. I have learned to organize my work and to find the most important things not only in science but in my life too.
My co-supervisor, Prof. Klaus K. Unger, I would never have started *:14& "646+"@:& H1*:#3*& =#3%& J:6& <141*& *#& d+1.hE& ,#:+..64& P3*6.T6"?&
University and meeting with you changed my life and the way of thinking. Thank you for the lessons not only in analytical science but for my carrier too. Thank you for the support throughout the years, for valuable advises and kind friendship. Because of you and your family Germany became my second home country.
Assoc. Prof. Vladas Gintautas, Head of the Department of Obstetrics, Hospital of Lithuanian University of Health Sciences, who believed in me from the beginning. Thank you for your help and all amniocentesis, this made it possible to start and carry out this research.
Assoc. Prof. A)*<.32.>" B/)3C)*>?.>, Head of the Department of Delivery, Hospital of Lithuanian University of Health Sciences, thanks you for your encouragement, finding a teacher’s thread in me and team work in Lithuania and abroard with ALSO and BLSO courses.
Assoc. Prof. 9&>)%." D*)3/)$*@, Administrator of Obstetrics and Gynecology Department, and colleagues at the Department of Women Consultation, for understanding and patience during my absence from clinical work.
Valdas Šarauskas, M.D. thank you for microscopic images.
The secretary 9:%."1(3&<@ and administrators E&F."B.>'$(.G)C):%@ and 1($%."1.(F3%@ – thank you for your always happy smiles and making difficult things possible.

122
All my colleagues at the Department of Obstetrics and Gynecology and Laboratory, Hospital of Lithuanian University of Health Sciences and ,#:+..64&P3*6.T6"?&;.1<6"41*=E&$#"&*:61"&43GG#"*&+.0&6.@#3"+?6B6.*%
My colleagues from Crisis Research Center thank you for harmonic working atmosphere and all impressive events.
Lina, Vilma, Jovita and Ligita my dear friends, for your friendship, loylty and long talks…
My mother, H)(3%@, thank you for your endless love, support and help. Thank you for everything you have learned and given me in my life…
In memoriam to my father, Stasys, his love to science inspired me to become a scientist.
My dear brother Evaldas, Edita and Paulius, thank you for your encourage.
I.*3%@".*<"J3()03>, thank you for your help during the years and for always being ready to take care of us.
Laura and Marius, thank you for your support.
My beloved husband and best friend Egidijus, thank you for your love and respect, constructive scientific discussions, guidance and help.
Ieva, my wonderful daughter, for being a charming and wise girl, understanding and forgiving when my mind was full of research and greeting me with your smile every day.
Finally, I would like to thank every woman who voluntarily parti-cipated in this study. This thesis would not exist without your contribution.




















