
PEGylated-thymoquinone-nanoparticle mediated retardation of breast cancer cell migration by...
17
PEGylated-thymoquinone-nanoparticle mediated retardation of breast cancer cell migration by deregulation of cytoskeletal actin polymerization through miR-34a Saurav Bhattacharya a , Manisha Ahir a , Prasun Patra a , Sudeshna Mukherjee b , Swatilekha Ghosh c , Minakshi Mazumdar c , Sreya Chattopadhyay b , Tanya Das c , Dhrubajyoti Chattopadhyay a , Arghya Adhikary a, * a Centre for Research in Nanoscience and Nanotechnology, University of Calcutta, JD-2, Sector III, Salt Lake, Kolkata 700098, West Bengal, India b Department of Physiology, University of Calcutta, 92 Acharya Prafulla Chandra Road, Kolkata 700009, West Bengal, India c Division of Molecular Medicine, Bose Institute, P-1/12, Calcutta Improvement Trust Scheme VII M, Kolkata 700054, West Bengal, India article info Article history: Received 29 August 2014 Accepted 20 January 2015 Available online Keywords: Thymoquinone Cell migration p53 miR-34a Rac1 Actin abstract Thymoquinone (TQ), a major active constituent of black seeds of Nigella sativa, has potential medical applications including spectrum of therapeutic properties against different cancers. However, little is known about their effect on breast cancer cell migration, which is the cause of over 90% of deaths worldwide. Herein, we have synthesized TQ-encapsulated nanoparticles using biodegradable, hydro- philic polymers like polyvinylpyrrolidone (PVP) and polyethyleneglycol (PEG) to overcome TQ's poor aqueous solubility, thermal and light sensitivity as well as consequently, minimal systemic bioavailability which can greatly improve the cancer treatment efficiency. Sizes of synthesized TQ-Nps were found to be below 50 nm and they were mostly spherical in shape with smooth surface texture. Estimation of the zeta potential also revealed that all the three TQ-Nps were negatively charged which also facilitated their cellular uptake. In the present investigation, we provide direct evidence that TQ-Nps showed more ef- ficiency in killing cancer cells as well as proved to be less toxic to normal cells at a significantly lower dose than TQ. Interestingly, evaluation of the anti-migratory effect of the TQ-Nps, revealed that PEG 4000 - TQ-Nps showed much potent anti-migratory properties than the other types. Further studies indicated that PEG 4000 -TQ-Nps could significantly increase the expression of miR-34a through p53. Moreover, NPs mediated miR-34a up-regulation directly down-regulated Rac1 expression followed by actin depoly- merisation thereby disrupting the actin cytoskeleton which leads to significant reduction in the lamel- lipodia and filopodia formation on cell surfaces thus retarding cell migration. Considering the biodegradability, non-toxicity and effectivity of PEG 4000 -TQ-Nps against cancer cell migration, TQ-Nps may provide new insights into specific therapeutic approach for cancer treatment. © 2015 Elsevier Ltd. All rights reserved. 1. Introduction Breast cancer, a severe life threatening cancer is the most common malignancy with the highest incidence rates among women worldwide. Despite the advancements in chemo- therapeutic treatment, significant metastatic property of the breast cancer cells still poses a major challenge to the available therapeutic regimes. Thus alternative therapies that can alter the metastatic potential of breast cancer cells are now drawing atten- tion of researchers worldwide. Thymoquinone (TQ; 2-methyl-5-isopropyl-1,4- benzoquinone; C 10 H 12 O 2 ; molecular weight:164.2), the principle active ingredient isolated from the volatile oil of black seed is known to exhibit prominent anti-neoplastic [1,2] activities along with its multiple pharmacological abilities like anti-oxidant [3], anti-inflammatory [4], anti-diabetic [5], as well as immunomodulatory properties [6]. TQ exerts its anti-cancerous effects through different modes of action, that is, by inhibition of cell proliferation, induction of apoptosis, cell cycle arrest, generation of reactive oxygen species (ROS), and inhibition of metastasis and angiogenesis [7]. Though * Corresponding author. Tel.: þ91 9830428550. E-mail address: [email protected] (A. Adhikary). Contents lists available at ScienceDirect Biomaterials journal homepage: www.elsevier.com/locate/biomaterials http://dx.doi.org/10.1016/j.biomaterials.2015.01.007 0142-9612/© 2015 Elsevier Ltd. All rights reserved. Biomaterials 51 (2015) 91e107
Transcript of PEGylated-thymoquinone-nanoparticle mediated retardation of breast cancer cell migration by...

lable at ScienceDirect
Biomaterials 51 (2015) 91e107
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
PEGylated-thymoquinone-nanoparticle mediated retardation ofbreast cancer cell migration by deregulation of cytoskeletal actinpolymerization through miR-34a
Saurav Bhattacharya a, Manisha Ahir a, Prasun Patra a, Sudeshna Mukherjee b,Swatilekha Ghosh c, Minakshi Mazumdar c, Sreya Chattopadhyay b, Tanya Das c,Dhrubajyoti Chattopadhyay a, Arghya Adhikary a, *
a Centre for Research in Nanoscience and Nanotechnology, University of Calcutta, JD-2, Sector III, Salt Lake, Kolkata 700098, West Bengal, Indiab Department of Physiology, University of Calcutta, 92 Acharya Prafulla Chandra Road, Kolkata 700009, West Bengal, Indiac Division of Molecular Medicine, Bose Institute, P-1/12, Calcutta Improvement Trust Scheme VII M, Kolkata 700054, West Bengal, India
a r t i c l e i n f o
Article history:Received 29 August 2014Accepted 20 January 2015Available online
Keywords:ThymoquinoneCell migrationp53miR-34aRac1Actin
* Corresponding author. Tel.: þ91 9830428550.E-mail address: [email protected] (A. Ad
http://dx.doi.org/10.1016/j.biomaterials.2015.01.0070142-9612/© 2015 Elsevier Ltd. All rights reserved.
a b s t r a c t
Thymoquinone (TQ), a major active constituent of black seeds of Nigella sativa, has potential medicalapplications including spectrum of therapeutic properties against different cancers. However, little isknown about their effect on breast cancer cell migration, which is the cause of over 90% of deathsworldwide. Herein, we have synthesized TQ-encapsulated nanoparticles using biodegradable, hydro-philic polymers like polyvinylpyrrolidone (PVP) and polyethyleneglycol (PEG) to overcome TQ's pooraqueous solubility, thermal and light sensitivity as well as consequently, minimal systemic bioavailabilitywhich can greatly improve the cancer treatment efficiency. Sizes of synthesized TQ-Nps were found to bebelow 50 nm and they were mostly spherical in shape with smooth surface texture. Estimation of thezeta potential also revealed that all the three TQ-Nps were negatively charged which also facilitated theircellular uptake. In the present investigation, we provide direct evidence that TQ-Nps showed more ef-ficiency in killing cancer cells as well as proved to be less toxic to normal cells at a significantly lowerdose than TQ. Interestingly, evaluation of the anti-migratory effect of the TQ-Nps, revealed that PEG4000-TQ-Nps showed much potent anti-migratory properties than the other types. Further studies indicatedthat PEG4000-TQ-Nps could significantly increase the expression of miR-34a through p53. Moreover, NPsmediated miR-34a up-regulation directly down-regulated Rac1 expression followed by actin depoly-merisation thereby disrupting the actin cytoskeleton which leads to significant reduction in the lamel-lipodia and filopodia formation on cell surfaces thus retarding cell migration. Considering thebiodegradability, non-toxicity and effectivity of PEG4000-TQ-Nps against cancer cell migration, TQ-Npsmay provide new insights into specific therapeutic approach for cancer treatment.
© 2015 Elsevier Ltd. All rights reserved.
1. Introduction
Breast cancer, a severe life threatening cancer is the mostcommon malignancy with the highest incidence rates amongwomen worldwide. Despite the advancements in chemo-therapeutic treatment, significant metastatic property of thebreast cancer cells still poses a major challenge to the availabletherapeutic regimes. Thus alternative therapies that can alter the
hikary).
metastatic potential of breast cancer cells are now drawing atten-tion of researchers worldwide.
Thymoquinone (TQ; 2-methyl-5-isopropyl-1,4- benzoquinone;C10H12O2; molecular weight:164.2), the principle active ingredientisolated from the volatile oil of black seed is known to exhibitprominent anti-neoplastic [1,2] activities along with its multiplepharmacological abilities like anti-oxidant [3], anti-inflammatory[4], anti-diabetic [5], as well as immunomodulatory properties[6]. TQ exerts its anti-cancerous effects through different modes ofaction, that is, by inhibition of cell proliferation, induction ofapoptosis, cell cycle arrest, generation of reactive oxygen species(ROS), and inhibition of metastasis and angiogenesis [7]. Though

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e10792
certain studies have already reported the anti-migratory effects ofTQ on colorectal [8], glioblastoma [9], melanoma [10], lung [11] yeta detail investigation of the molecular mechanisms are still lacking.
In spite of its significant anti-cancer properties, utilization of TQas a promising anti-cancer drug is often hindered due to its pooraqueous solubility thereby causing serious bioavailability issues.Various nano-carriers are therefore being developed to enhancethe solubility and increase the bioavailability of TQ in the cells. Thisprocess of nano-encapsulation of water-insoluble drugs havegained major interest as it protects the drugs from exposure toexternal factors, limits their diffusion to normal tissues and alsoprevents their rapid metabolism. Because of the leaky nature oftumor vasculature along with poor lymphatic drainage, nano-particles often accumulate near the tumors which results in bettertargeting and reduced exposure of non-specific sites to the drug. TQnanoparticles are therefore believed to hold greater promise thanfree TQ for two main reasons: (1) their enhanced activity due tobetter targeting of the cancer in vitro, and (2) their improved in vivobioavailability and distribution [12].
In recent years, there is significant interest in the developmentof nanoparticle-mediated drug delivery approach [13e15] whichhas been studied and proved a challenging advantageous tool tominimize the adverse effects andmaximize therapeutic outcome ofdrugs with the specificity and selectivity. Formulation of anti-can-cer agents by nanoparticles of biodegradable polymers alleviatesside effects of the drugs by targeting tumor via enhanced perme-ability and retention (EPR) effect. However, obtaining high anti-tumor efficiency and also delineating the mechanism of tumortargeting of this approach still remains a significant challenge. Inthis context in the present study we have attempted to encapsulateTQ in nanoparticles (referred herein as TQ-Np) with Poly-ethyleneglycol (PEG) and Polyvinylpyrrolidone (PVP) as encapsu-lating agent. PVP, which is soluble in water and ethanol has longbeen used in medicine, food, beverage, as a blood plasma substi-tute, a detoxification material, a binding agent, for a large numberof pharmacological preparations, thickeners for liquid drugs andalso as a shell for the targeted delivery of drugs [16]. The water-soluble PEG is a non-toxic, non-immunogenic, and commercially-available FDA-approved polymer. PEG, on the other hand, is alsoan amphiphilic polymer showing excellent biocompatibility and isfrequently used in biomedical applications. Furthermore, conjuga-tion with PEG restricts the passage through the blood brain barrierthus abrogating neurotoxicity associated with free drug or phyto-chemical and also prolongs the circulating half-life of the free drugsor phytochemicals [17]. Several small organic molecules, such ascamptothecin [17], doxorubicin [18], and paclitaxel [19], havealready been conjugated to PEGs and these conjugates especiallycamptothecin are in clinical trials [17]. As these properties may beeffective towards the clinical application of the drug we thereforepreferred to use PVP and PEG over other polymers for nano-encapsulation of TQ.
Following the nano-encapsulation of TQ, our work was furtherextended in exploring the molecular mechanisms of TQ-Np-mediated retardation of migration in breast cancer cells. The ma-jor reason behind 90% death due to breast cancer is metastasisinstead of primary neoplasias [20]. In order to metastasize, the cellinitially acquires the motile phenotype which helps it to cross thetissue boundaries [21]. The regulation of cellular morphology dur-ing cell migration is largely mediated by rearrangements of theactin cytoskeleton [22,23]. Lamellipodia which is a broad, flat cellprotrusion that dominates at the edges of cultured fibroblasts andmany other motile cells are one of these structures. The protrusionof the lamellipodium is provided by continuous growth of actinfilaments towards the leading edge of the cell [24]. This actinpolymerization is efficiently regulated by Rho GTPases [24]. Rho
GTPases belong to the Ras superfamily of low molecular mass(~21 kDa) proteins and are widely expressed in mammalian cells[25]. RhoA, Rac1 and Cdc42 are the best-characterized members ofRho family GTPases. They act as molecular switches to controlsignal transduction pathways by cycling between a GDP-bound,inactive form and a GTP-bound, active form. In their GTP-boundstate, they interact with downstream targets (effectors) to elicit avariety of intracellular responses [26]. Among these three, activa-tion of Rac1 (Ras-related C3 botulinum toxin substrate1) inducesactin polymerization to form lamellipodia under the influence ofgrowth factors, cytokines and extracellular matrix components[26]. Several reports indicate that aberrant Rac1 activity is mainlyresponsible for invasive and malignant behavior of hep-atocarcinoma [27], breast carcinoma [28], and melanoma [29].However, breast tissue sample analysis has demonstrated that thisaberrant functioning of Rac1 is not due to its genetic mutation.There are some upstream regulators whose activity controls itsproper functioning [30]. MicroRNAs (miRNAs) could be one of thoseregulators. These small, endogenous non-coding RNAs elicit theirregulatory effects by imperfectly binding to the 30-untranslatedregions (UTRs) of target mRNAs, either preventing their translationor causing target degradation. Through down-regulating theirtarget gene expression, miRNAs can direct a wide repertoire ofbiological mechanisms [31]. In this study we include several miR-NAs such as miR-34a, -10b, -155, -16-1,-142-3p and -146a that werepreviously reported for association with cancer cell migration[32e37]. We were interested to observe if TQ-Nps could regulatethe expression of these miRNAs to modulate the migratorymorphology of breast cancer cells and thus able to inhibit the cellmigration effectively.
Therefore, the goal of the present study was set to synthesizethymoquinone loaded nanoparticles to target breast cancer cellmigration by TQ-Nps, thus reducing TQ's side effects, improving itspharmacokinetics, modifying its biodistribution and enhancing itsefficacy and also to delineate the exact mechanisms by which TQ-Nps potentiates accumulation of depolymerized actin by down-regulation of Rac1 through miR-34a in a p53 dependent mannerwhich thereby decreases the lamellipodial extensions, ultimatelyrestricting breast cancer cell migration. This work thereby providesnew hopes in treatment of metastatic breast cancer by use ofnanoparticles with better therapeutic applications.
2. Materials and methods
2.1. Chemicals
Thymoquinone (2-isopropyl-5-methyl-1,4-benzoquinone) 99%, Pluronic P123,was purchased from Sigma (USA). Poly-(Ethylene Glycol)-200 [PEG200; solution;density 1.124e1.126] and Poly-(Ethylene Glycol)-4000 [PEG4000; powder; MW3500e4500], Polyvinylpyrrolidone (K-30) [powder; MW ~ 40,000] were purchasedfrom Merck (Darmstadt, Germany). Acetonitrile, Dichloromethane, Dime-thylformamide (DMF), Dimethyl sulfoxide (DMSO), Ethanol, Methanol were ofsynthesis grade and purchased from Merck (Darmstadt, Germany). Triton X-100,Ammonium persulphate (APS), TEMED and N,N0-methylene bis acrylamide wereobtained from Merck (Darmstadt, Germany). Culture media Dulbecco's ModifiedEagle's Medium (DMEM), RPMI-1640, penicillin, streptomycin, gentamycin,L-glutamine, non-essential amino acids, amphotericin B were purchased fromHIMEDIA (Mumbai, India). Chlorpromazine and Histopaque-1077 was purchasedfrom Sigma, USA. Fetal bovine serum (FBS) was purchased from Invitrogen (Carls-bad, USA). Antibodies against p53 and Rac1 were obtained from Santa CruzBiotechnology (Santa Cruz, USA). Alexa Fluor 488-Phalloidin and Prolong GoldAntifade Reagent, (40 ,6-diamidino-2-phenylindole) (DAPI) and Hoechest 33342were purchased from Invitrogen (Carlsbad, USA).
2.2. Cell culture and (peripheral blood mononuclear cells) PBMC isolation
The human mammary carcinoma cell lines (MCF-7, HBL-100) were obtainedfrom National Centre for Cell Science (Pune, India) and maintained in completeDMEM. Viable cell numbers were determined by Trypan blue exclusion test.
Human whole blood was collected in heparinized vacutainer blood collectiontubes (BD, Franklin Lakes, NJ) from adult healthy volunteers with prior consent.

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107 93
Whole blood (100 ml) was diluted with 150 ml of RPMI-1640 and then layered incentrifuge tubes onto 120 ml of Histopaque-1077 gradient. After centrifugation theperipheral blood mononuclear cells (PBMC) layer was washed twice with PBS andresuspended followed by cultured in RPMI-1640 with 10% FBS.
2.3. Synthesis of polymer-encapsulated thymoquinone nanoparticles (TQ-Nps)
Three nanopraticles of thymoquinone (TQ-Np) were synthesized by using threepolymers. Both Poly-(Ethylene Glycol200) [PEG200]- and Poly-(Ethylene Glycol4000)[PEG4000]-encapsulated TQ-Nps were synthesized according to solvent evaporationtechnique while Polyvinylpyrrolidone (PVP)-coated TQ-Nps were synthesized innanoprecipitation technique. The synthesis procedures of these three TQ-Nps areillustrated below.
2.3.1. Synthesis of Poly-(Ethylene Glycol)200-nanoThymoquinone (PEG200- TQ-Nps)In order to prepare a solution of bioactive lipophilic TQ, 5 mg was dissolved in
15 ml of acetonitrile. 10 mg of surfactant P123 was added to mixture. This mixturewas injected drop-wise into 50 ml Poly-(Ethylene Glycol)200 (PEG200) under ultra-sonic condition for 20e30 min. When addition was complete, the mixture was keptin ultrasonic condition for another 30 min. Then it was kept in stirring condition forovernight. Next day, the organic solvent present in suspension was evaporated byrotary evaporation (Eyela, USA) and PEG200-TQ-Nps were obtained.
2.3.2. Synthesis of Poly-(Ethylene Glycol4000)-nanoThymoquinone (PEG4000 - TQ-Nps)
For PEG4000-TQ-Np, first 0.821 g of TQwasmixed in 50ml of acetonitrile. 100mgof surfactant P123 was added to the mixture and stirred. Powdered 4 g of PEG4000
(50 mM) was mixed in 20 ml of deionized water and homogenized using bathsonicator for 20e30 min. When PEG4000 was completely dispersed in water, the TQ/Acetonitrile mixture was drop-wise injected under ultrasonic condition. Whenadditionwas completed, the mixturewas once again kept in ultrasonic condition foranother 30minwhich was followed by overnight stirring at room temperature. Nextday, the organic solvent present in suspension was evaporated by rotary evaporatorand PEG4000TQ-Nps were extracted.
2.3.3. Synthesis of Polyvinylpyrrolidone-nanoThymoquinone (PVP-TQ-Nps)Nanoprecipitation technique was used to prepare PVP-TQ-Np. Briefly, 164.2 mg
of TQ (100 mM) was dispersed in 10 ml of acetonitrile. Then 1 g of Poly-vinylpyrrolidone (PVP) was mixed in 5 ml Dimethylformamide (DMF) and vigor-ously stirred at 160 �C. The TQ/Acetonitrile mixture was added into PVP/DMFmixture by hot-injection method under stirring condition. After addition, themixture was kept in stirring condition continuously at 160 �C for 5 h. The organicsolvent present in final suspension was evaporated by rotary vacuum evaporationusing rotary evaporator and PVP-TQ-Np was obtained.
2.4. Physico-chemical characterization of TQ-Nps
To ensure the presence of TQ inside the encapsulated nanoparticles the absor-bance of nanoparticles were measured by UV/Vis Spectrophotometer (Perkin Elmer,LAMBDA 1050,USA) at 330 nm [38]. The hydrodynamic size and zeta potential weremeasured by DLS analysis (Delsa™ Nano C, Beckman Coulter, USA). The morphologyof the nanoparticles was observed by field emission scanning electron microscopy(FESEM, JSM-7600F, JEOL, Japan); size and shape was analysed by transmissionelectronmicroscopy (JEM-2100HR-TEM, JEOL, Japan). The chemical properties of TQ-Nps were characterized by Fourier transform infrared spectroscopy (FT-IR, JASCO,Japan). Powder X-ray diffraction (XRD) measurements were performed with aPANalytical X0 PERT Pro diffractometer at a scanning rate of 4�/min in the 2q rangefrom 5� to 70� using graphite monochromated Cu-ka radiation (l¼ 0.15405 nm) andFor 1H NMR analysis, PEG4000-TQ-Nps were dissolved in CDCl3, and analyzed by aNMR Spectrometer (Bruker AC 400 superconducting, Switzerland). Thermogravi-metric analysis (TGA) of the synthesized nanoparticles were done on TG/DTAanalyzer (Perkin Elmer, USA) under nitrogen atmosphere at a heating rate of20 �C min�1.
2.5. Entrapment efficiency
The entrapment efficiency (EE) of TQ-Nps was determined by ultracentrifuga-tion at 15,000 rpm, 4 �C for 30 min. The amount of free TQ in the supernatant wasdissolved in ethanol and sonicated for 10 min after which its absorbance at 330 nmwas measured. The amount of drug was estimated from the standard curve of TQ inethanol. The EE of Nps were calculated by the following equations reported previ-ously [39] and all measurements were performed in triplicate (n ¼ 3).
%EE ¼ ðTotal drug� Free drugÞTotal drug
� 100
2.6. In-vitro drug release
50 mg of TQ-Nps were dispersed in 5 ml phosphate buffer separately (both pH7.4 and pH 5.5) and then placed into dialysis membrane (MWCO-12KDa, HiMedia,
India) against 100ml of respective phosphate buffer in a glass bowl placed at stirringrate of 150 rpm. At various time intervals, 3 mL of the dialysis buffer was withdrawnand the absorbance at 330 nm was measured to determine the quantity of the TQreleased and 3 mL of fresh phosphate buffer was added to the dialysis solution tomaintain fixed volume. All the operations were carried out in triplicate and pre-cautions were taken to protect the drug from heat and light.
2.7. In vitro cellular uptake of TQ-Nps
The in vitro cellular uptake was measured according to minute modifications ofGupta et al. (2009) [40]. Briefly, MCF-7 and HBL-100 cells were seeded in 96-wellplates (2000 cells/well) and incubated overnight. The different doses of PVP-TQ-Np, PEG200-TQ-Np and PEG4000-TQ-Np solution were added to the plate in tripli-cate manner and incubated for 24 h. The nanoparticles contained in the mediumwere removed from the wells next day. Cells were lysed in 100 ml of dimethylsulfoxide (DMSO) and cell lysis suspensions were reserved for absorbance mea-surement at 330 nm. In order to study the involvement of the clathrin mediatedendocytosis, MCF-7 cells were pre-incubated with 25 mM of chlorpromazine (aspecific clathrin mediated endocytosis inhibitor) for 30 min following treatmentwith all the three different TQ-Nps.
2.8. Determination of cell cycle by flowcytometry
Cell viability under different doses of TQ and TQ-Nps were analyzed on flowcytometer (FACS Calibur, Beckton Dickinson, San Jose). Cell cycle phase distributionof nuclear DNA was determined using a fluorescence detector equipped with488 nm argon laser light source and 623 nm band pass filter (linear scale) andCellQuest software (BectoneDickinson, San Jose). Ten thousand total events wereacquired and histogram display of DNA content (x-axis, PI fluorescence) versuscounts (y-axis) has been displayed.
2.9. Cell viability assay
Cell viability was determined using the 3(4,5-dimethylthiazolyl-2)2,5-diphenyltetrazolium bromide assay (MTT, Sigma) according to the manufacturer's protocol.In brief, cells were seeded onto 96-well plates and subjected to treatments (TQ-Nps)and cultured for 24 h followed by MTT addition. After 6 h, 100 mL of detergent re-agent was added to each well and absorbance intensity was recorded at 570 nm on aBio-Rad (model 550) microplate reader. Cell viability is defined as the percent livecells compared with untreated controls.
2.10. Determination of cellular apoptosis by Annexin V
To determine the presence of apoptosis and necrosis, cell death was analyzed byAnnexin V and PI staining. For staining, cells (2 � 105 cells/well in 12-well plates)were treated with nanoparticles for 24 h. Cells were washed with PBS, centrifuged,and suspended in Annexin V binding buffer containing 5 ml/100 ml of Annexin V-FITC and PI (final concentration 5 mg/ml). Cells were incubated for 15 min at 37 �Cand analyzed using the flow cytometer (BD Biosciences, San Diego, CA). The Fluo-rochromewas excited using the 488 nm line of argon ion laser, and Annexin V and PIemissions were monitored at 525 and 620 nm, respectively. A total of at least1 � 104 cells were analyzed per sample.
2.11. Wound healing assay
The migration of breast cancer cells under different conditions were determinedby bidirectional wound healing assay. Briefly, cells were grown to 80e90% con-fluency in 12 well plates after which a sterile pipette tip was used to scratch themonolayer of cells to form a bidirectional wound. Migration was quantitated by asemi-automated, computer-assisted procedure by a person blinded with respect tothe experimental treatment. The data from triplicate wells were calculated as themeans ± S.E.M., the migration rate of control cells was taken as 100% and healingrate of other plates were compared with respect to control cells.
2.12. Transwell migration assay
Migration assay was done using cell culture inserts (BD Biosciences, Sparks,USA). Cell culture inserts with pores (8 mm) were placed in the 12-well companionplate containing 100 ml of DMEM Media without chemo-attractant i.e. serum. In theupper half of the insert 2 � 105 cells was placed inside the chamber. The serumcontaining DMEM (10% FBS) was then added to the lower chamber of the 12-wellplate. Next day the cells in the insert were washed with PBS, fixed with 3.7%formaldehyde andwere permealized usingmethanol. At last, cells were stainedwithGiemsa stain for 30 min. Cells present in the lower part of the inserts were deter-mined by counting cells in five microscopic fields per well, and the extent ofmigration will be expressed as an average number of cells per microscopic field.
2.13. Scanning electron micrographs to determine the morphology of cells
The morphology of breast carcinoma cells under different conditions wereobserved under Scanning ElectronMicroscope (SEM). The cells were grown on poly-L-Lysine (1:10) coated glass slide (1 cm2) overnight at 37 �C in 5% CO2. Next day, cellswere treated with specific dose of TQ-Nps and kept in the CO2 incubator for

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e10794
overnight. Next day, the cells were washed by sodium cacodylate buffer, and thenfixed with 2.5% glutaraldehyde buffered in sodium cacodylate for 1 h. The sampleswere washed with sodium cacodylate buffer for 3 times, and dehydrated through aseries of alcohol concentrations (10%, 30%, 50%, 70%, 90% and 100%), and subjected toair drying. The samples were then visualized by EVO-18 special edition SEM (ZEISS,Germany).
2.14. Immunoblotting
The cells were seeded (2.6 � 105 cells/well) in a 6-well microtiter plate followedby overnight incubation. After appropriate treatment for specific time points, thecells were trypsinized followed by whole cell lysate extraction. For whole cell ly-sates, cells were homogenized in buffer (20 mM HEPES pH 7.5, 10 mM KCl, 1.5 mM
MgCl2, 1 mM Na-EDTA, 1 mM Na-EGTA, and 1 mM DTT). All buffers were supple-mented with protease and phosphatase inhibitor mixtures [41]. Protein estimationwas done using bicinchoninic acid (BCA) assay Kit (Merck, Germany). An equalamount of proteins were separated by SDS-PAGE and transferred onto PVDFmembrane (Millipore) and then incubated with primary antibodies like, anti-p53,Rac1 and a-actin (purchased from Santa Cruz Biotechnologies, USA) and ALP-conjugated secondary antibodies. The blots were developed with NBT/BCIP (1:1).The results were expressed as fold change relative to the control after normalizationto a-actin.
2.15. RNA isolation and quantitative real-time PCR
Total RNAswere isolated from cells usingmiRVANA kit (Invitrogen, Carlsbad, CA)according to the manufacturer's instructions. The integrity and purity of total RNAsand small RNAs was verified by ND1000 spectrophotometer (Thermo) and gel-electrophoresis on urea denaturation gel. 1 mg of total RNAs were reverse tran-scribed using Transcriptor First Strand cDNA Synthesis Kit and oligo(dT)18 primer(Roche, Switzerland). In case of miRNA detection, the miR-specific stem-loopprimers (Supplementary Tables 1 and 2) were used along with same cDNA synthesiskit. qRT-PCR was performed with 50 mg sample using Light cycler 96 (Roche) andFastStart Universal SYBR Green Master (Roche) and specific primers (SupplementaryTables 1 and 2) snRNAU6 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH)were used as internal controls for miRNA and mRNA quantification, respectively.Gene and miR expression was defined from the threshold cycle (Cq), and relativeexpression levels were calculated by using the 2�DCq method after normalizationwith reference to expression of housekeeping genes (GAPDH or U6).
2.16. Fluorescence imaging
For fluorescence imaging, the cells were fixed with 3.7% formaldehyde andpermeabilized with 0.1% Triton X-100. Cells were then either incubated with anti-p53 antibody followed by FITC-conjugated secondary antibody (Sigma) or stainedwith Alexa Fluor 488-conjugated phalloidin (Invitrogen, Carlsbad, CA). The nucleuswas stained with Hoechst 33342 (Invitrogen, Carlsbad, USA). The samples wasmounted on clean glass slides using Prolong Gold Antifade Reagent (Invitrogen,Carlsbad, USA) and visualized under fluorescence microscope (Olympus.).
2.17. Plasmids and transfections
miR 34a antistrand expression plasmid were used for miR 34a downregulationstudies. The psiCHECK2-AS34a (procured from ADDGENE; Plasmid no. 37099)clones were introduced in the cell line (MCF-7) using Lipofectamine 2000 (Invi-trogen, Carlsbad, CA). Stably expressing clones were isolated by limiting dilution andselectionwith G418 sulphate (Cellgro, USA) at a concentration of 400 mg/ml and cellssurviving this treatment were cloned and screened by qRT PCR analysis with specificprimers.
2.18. In vivo evaluation of activity of PEG4000-TQ-Nps
We have established an Ehrlich's ascites carcinoma (EAC) solid tumor model inSwiss Albino mice. The tumor cells were withdrawn from the peritoneal cavity ofliquid EAC bearing mice, diluted in saline, counted and re-injected (1 � 106 cells/animal) to right thigh tissue of experimental animals for developing solid tumor.In vivo studies of systemic toxicity and anti-tumor activities of PEG4000-TQ-Nps wereconducted using EAC solid tumor mice model in compliance with the principles andprocedures approved by the local Institutional Animal Care and Use Committee.
2.18.1. Anti-tumor efficacySwiss albinomice, 6e8weeks old, weighing 18e22 gweremaintained as per the
principles and guidelines of the ethical committee for animal care of University ofCalcutta. The animals were housed in polypropylene cages and provided standardpellet diet (Agro Corporation Pvt. Ltd., India) and water ad libitum. The standardpellet diet composed of 21% protein, 5% lipids, 4% crude fiber, 8% ash, 1% calcium,0.6% phosphorus, 3.4% glucose, 2% vitamin, and 55% nitrogen-free extract (carbo-hydrates). The mice were maintained under controlled conditions of temperatureand humidity with a 12 h light/dark cycle.
The anti-cancer potential of PEG4000-TQ nanoparticles was determined with theEAC solid tumor mouse model. To study and compare the anti-tumor activity of Nps,
24 Swiss albino mice were used in the present study, (four batches of 6 animalseach). Out of 24 mice, six were used as untreated (normal) control. Rest of the micewere injected with EAC to induce solid tumor, and divided into three groups, eachcontaining six animals (n ¼ 6): (1) Tumor Control (No Treatment) (2) PEG4000-TQ-Nps (2.5mg/kg bodyweight); (3) PEG4000-TQ-Nps (5mg/kg bodyweight); Group twoand three among tumor bearers received intravenous (tail vein) administration ofPEG4000-TQ-NPs on every alternative day from 12th day of injection of tumor cells.Tumor induction was done as per institutional animal ethical guidelines whichstates that the tumor weight should not be more than 10% of the total bodyweight ofrodents involved in therapeutic experiments. In the present study, the tumor weight(<2.5 g) of the untreated tumor-bearing animals was much less than 10% of theirbody weights (~35 g average) such that the total tumor burden of the animals werewell within the permissible limits.
The tumor size was measured using a digital Vernier caliper. The tumor volumewas calculated using the ellipsoid volume equation: tumor volume (cm3) ¼ p/6 � L � W � H, wherein L is length, W is width, and H is height. The mice wereeuthanized after 45 days following treatment. Tumors were excised, collected, andfixed with 10% formalin for immunohistochemistry to determine cellular and mo-lecular changes that occurred within the tumors. Tumor tissues were also snap-freezed in liquid nitrogen and stored in �80 �C freezer for further western blotand qRT-PCR analysis.
2.18.2. Estimation of serum specific toxicity markers (ALT, AST, LDH, ALP)Serum specific toxicity markers, Alanine transaminase (AST; EC 2.6.1.1), Aspar-
tate transaminase (ALT; EC 2.6.1.2), Lactate dehydrogenase (LDH; EC 1.1.1.27) andAlkaline phosphatase (ALP; EC 3.1.3.1) activities weremeasured using commercially-available standard kits purchased from Span Diagnostics, India following manu-facturer's protocol.
2.18.3. Preparation of 10% post mitochondrial supernatantApproximately, 100 mg of liver, collected from the animals of different groups,
were homogenized in 1 ml of chilled phosphate buffer (0.1 M, pH 7.4) containing KCl(1.17% w/v). The homogenate was centrifuged at 800 g for 5 min at 4 �C to separatethe nuclear debris. The supernatant was collected and was again centrifuged at10,500 g for 20 min at 4 �C to obtain PMS (Kaur et al., 2006).
2.18.4. Catalase activityCatalase (CAT; EC 1.11.1.6) activity was measured spectrophotometricaly at
240 nm according to Claiborne (1985) [42] based on the decomposition of H2O2 toH2O and O2 by catalase present in hepatic PMS.
2.18.5. Estimation of lipid peroxidation (LPO)Level of lipid peroxidation in hepatic PMS was performed by the method of
Wright et al. (1981) [43] and was measured by measuring the amount of malo-naldehyde (MDA) formed as end product of lipid peroxidation.
2.18.6. Superoxide dismutase (SOD) activitySuperoxide dismutase (SOD, EC 1.15.1.1) enzyme activity in hepatic PMS was
assayed spectrophotometrically following the method of Marklund (1974) [44] SODactivity was expressed as units/mg of protein.
2.19. Immunohistochemistry
Immunohistochemistry was used to determine the expression pattern of p53using formalin-fixed, paraffin-embedded (FFPE) tissues of control and treated solidtumor bearing EAC mice. The sections were deparaffinized with xylene and rehy-drated through graded alcohols to water. The slides were incubated with anti-p53primary antibody for overnight at 4 �C followed by FITC-tagged secondary anti-body. The slides were counter stained with DAPI. The sections were visualized underBD Pathway 855 microscope. (BD Biosciences, Sparks, MD).
2.20. Statistical analysis
All the data in this study were analyzed by Graph Pad Prism 5.0 (GraphPadSoftware Inc., USA). Data are expressed as mean ± SD. For values that were normallydistributed, direct comparison between two groups was conducted by IndependentSample's T test. P value of <0.05 was considered statistically significant.
3. Results and discussion
3.1. Preparation and characterization of polymer-encapsulatedthymoquinone nanoparticles (TQ-Nps)
PEG200, PEG4000 and PVP-coated thymoquinone nanoparticles(TQ-Nps) were synthesized according to the procedure described in‘Materials and methods’ section (Fig. 1A). The presence of the TQ inthese polymer encapsulated nanoparticles was evaluated by UV-
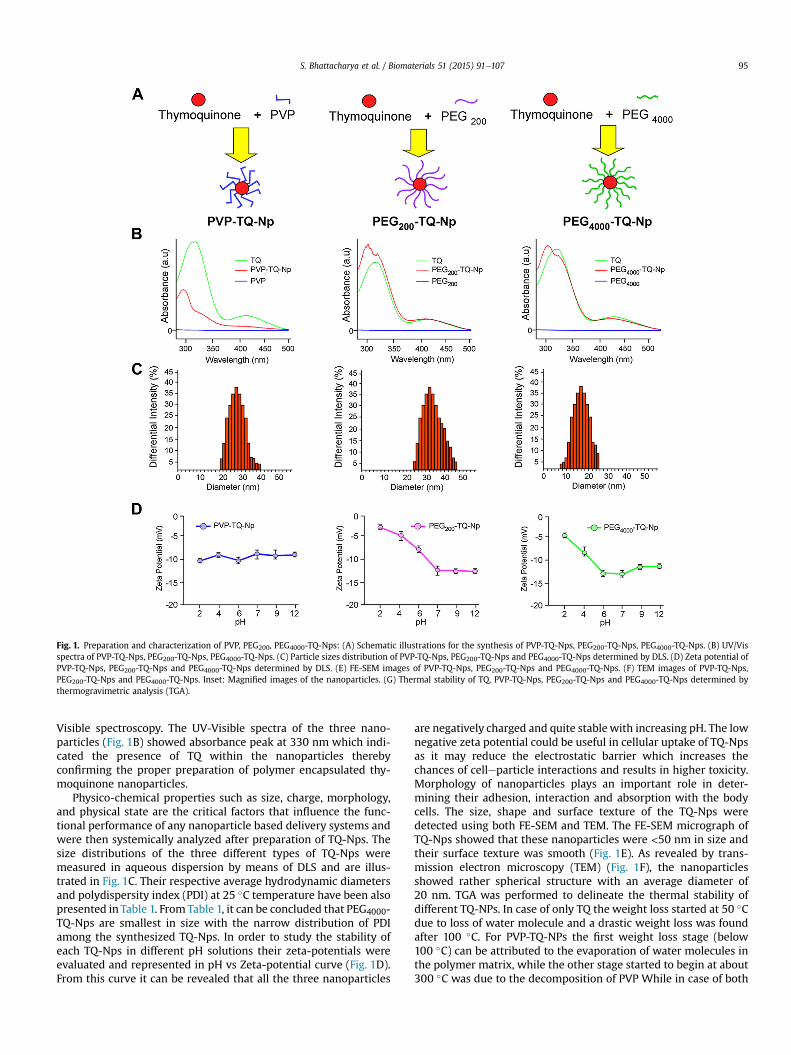
Fig. 1. Preparation and characterization of PVP, PEG200, PEG4000-TQ-Nps: (A) Schematic illustrations for the synthesis of PVP-TQ-Nps, PEG200-TQ-Nps, PEG4000-TQ-Nps. (B) UV/Visspectra of PVP-TQ-Nps, PEG200-TQ-Nps, PEG4000-TQ-Nps. (C) Particle sizes distribution of PVP-TQ-Nps, PEG200-TQ-Nps and PEG4000-TQ-Nps determined by DLS. (D) Zeta potential ofPVP-TQ-Nps, PEG200-TQ-Nps and PEG4000-TQ-Nps determined by DLS. (E) FE-SEM images of PVP-TQ-Nps, PEG200-TQ-Nps and PEG4000-TQ-Nps. (F) TEM images of PVP-TQ-Nps,PEG200-TQ-Nps and PEG4000-TQ-Nps. Inset: Magnified images of the nanoparticles. (G) Thermal stability of TQ, PVP-TQ-Nps, PEG200-TQ-Nps and PEG4000-TQ-Nps determined bythermogravimetric analysis (TGA).
S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107 95
Visible spectroscopy. The UV-Visible spectra of the three nano-particles (Fig. 1B) showed absorbance peak at 330 nm which indi-cated the presence of TQ within the nanoparticles therebyconfirming the proper preparation of polymer encapsulated thy-moquinone nanoparticles.
Physico-chemical properties such as size, charge, morphology,and physical state are the critical factors that influence the func-tional performance of any nanoparticle based delivery systems andwere then systemically analyzed after preparation of TQ-Nps. Thesize distributions of the three different types of TQ-Nps weremeasured in aqueous dispersion by means of DLS and are illus-trated in Fig. 1C. Their respective average hydrodynamic diametersand polydispersity index (PDI) at 25 �C temperature have been alsopresented in Table 1. From Table 1, it can be concluded that PEG4000-TQ-Nps are smallest in size with the narrow distribution of PDIamong the synthesized TQ-Nps. In order to study the stability ofeach TQ-Nps in different pH solutions their zeta-potentials wereevaluated and represented in pH vs Zeta-potential curve (Fig. 1D).From this curve it can be revealed that all the three nanoparticles
are negatively charged and quite stable with increasing pH. The lownegative zeta potential could be useful in cellular uptake of TQ-Npsas it may reduce the electrostatic barrier which increases thechances of celleparticle interactions and results in higher toxicity.Morphology of nanoparticles plays an important role in deter-mining their adhesion, interaction and absorption with the bodycells. The size, shape and surface texture of the TQ-Nps weredetected using both FE-SEM and TEM. The FE-SEM micrograph ofTQ-Nps showed that these nanoparticles were <50 nm in size andtheir surface texture was smooth (Fig. 1E). As revealed by trans-mission electron microscopy (TEM) (Fig. 1F), the nanoparticlesshowed rather spherical structure with an average diameter of20 nm. TGA was performed to delineate the thermal stability ofdifferent TQ-NPs. In case of only TQ the weight loss started at 50 �Cdue to loss of water molecule and a drastic weight loss was foundafter 100 �C. For PVP-TQ-NPs the first weight loss stage (below100 �C) can be attributed to the evaporation of water molecules inthe polymer matrix, while the other stage started to begin at about300 �C was due to the decomposition of PVP While in case of both
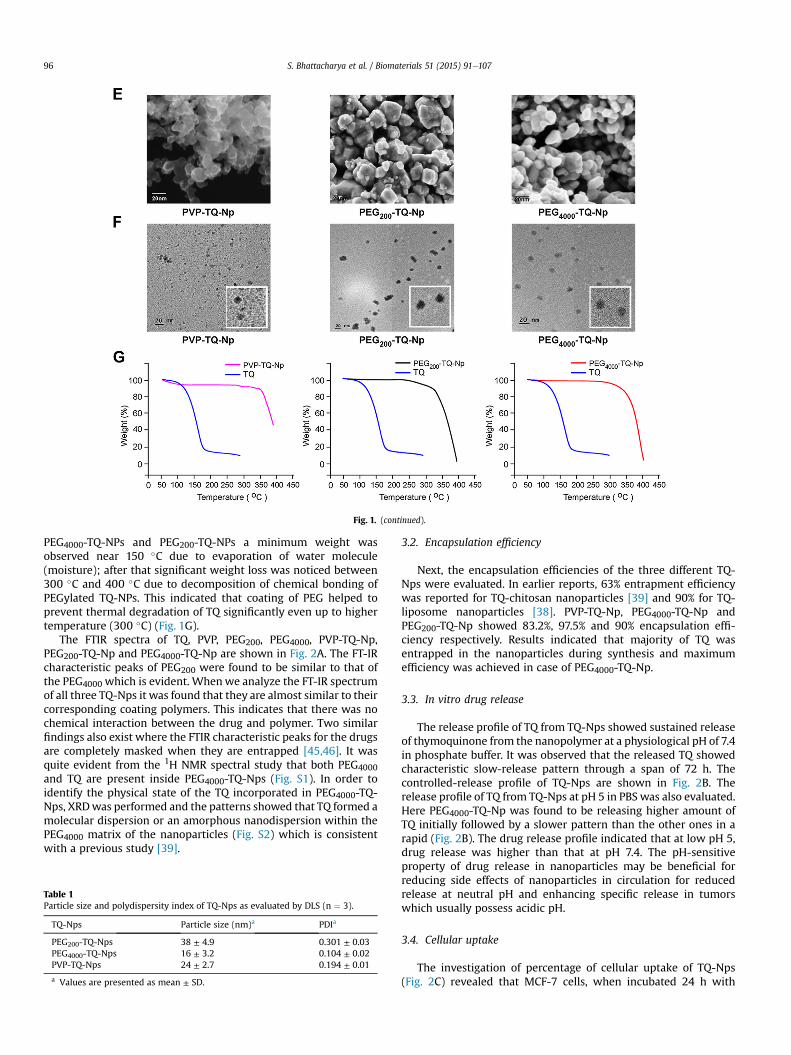
Fig. 1. (continued).
S. Bhattacharya et al. / Biomaterials 51 (2015) 91e10796
PEG4000-TQ-NPs and PEG200-TQ-NPs a minimum weight wasobserved near 150 �C due to evaporation of water molecule(moisture); after that significant weight loss was noticed between300 �C and 400 �C due to decomposition of chemical bonding ofPEGylated TQ-NPs. This indicated that coating of PEG helped toprevent thermal degradation of TQ significantly even up to highertemperature (300 �C) (Fig. 1G).
The FTIR spectra of TQ, PVP, PEG200, PEG4000, PVP-TQ-Np,PEG200-TQ-Np and PEG4000-TQ-Np are shown in Fig. 2A. The FT-IRcharacteristic peaks of PEG200 were found to be similar to that ofthe PEG4000 which is evident. Whenwe analyze the FT-IR spectrumof all three TQ-Nps it was found that they are almost similar to theircorresponding coating polymers. This indicates that there was nochemical interaction between the drug and polymer. Two similarfindings also exist where the FTIR characteristic peaks for the drugsare completely masked when they are entrapped [45,46]. It wasquite evident from the 1H NMR spectral study that both PEG4000and TQ are present inside PEG4000-TQ-Nps (Fig. S1). In order toidentify the physical state of the TQ incorporated in PEG4000-TQ-Nps, XRDwas performed and the patterns showed that TQ formed amolecular dispersion or an amorphous nanodispersion within thePEG4000 matrix of the nanoparticles (Fig. S2) which is consistentwith a previous study [39].
Table 1Particle size and polydispersity index of TQ-Nps as evaluated by DLS (n ¼ 3).
TQ-Nps Particle size (nm)a PDIa
PEG200-TQ-Nps 38 ± 4.9 0.301 ± 0.03PEG4000-TQ-Nps 16 ± 3.2 0.104 ± 0.02PVP-TQ-Nps 24 ± 2.7 0.194 ± 0.01
a Values are presented as mean ± SD.
3.2. Encapsulation efficiency
Next, the encapsulation efficiencies of the three different TQ-Nps were evaluated. In earlier reports, 63% entrapment efficiencywas reported for TQ-chitosan nanoparticles [39] and 90% for TQ-liposome nanoparticles [38]. PVP-TQ-Np, PEG4000-TQ-Np andPEG200-TQ-Np showed 83.2%, 97.5% and 90% encapsulation effi-ciency respectively. Results indicated that majority of TQ wasentrapped in the nanoparticles during synthesis and maximumefficiency was achieved in case of PEG4000-TQ-Np.
3.3. In vitro drug release
The release profile of TQ from TQ-Nps showed sustained releaseof thymoquinone from the nanopolymer at a physiological pH of 7.4in phosphate buffer. It was observed that the released TQ showedcharacteristic slow-release pattern through a span of 72 h. Thecontrolled-release profile of TQ-Nps are shown in Fig. 2B. Therelease profile of TQ from TQ-Nps at pH 5 in PBS was also evaluated.Here PEG4000-TQ-Np was found to be releasing higher amount ofTQ initially followed by a slower pattern than the other ones in arapid (Fig. 2B). The drug release profile indicated that at low pH 5,drug release was higher than that at pH 7.4. The pH-sensitiveproperty of drug release in nanoparticles may be beneficial forreducing side effects of nanoparticles in circulation for reducedrelease at neutral pH and enhancing specific release in tumorswhich usually possess acidic pH.
3.4. Cellular uptake
The investigation of percentage of cellular uptake of TQ-Nps(Fig. 2C) revealed that MCF-7 cells, when incubated 24 h with

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107 97
different doses of three TQ-Nps, able to internalize PEG4000-TQ-Npsbetter than other two at almost every dose. Probably the smallersize of PEG4000-TQ-Nps is helping them to get internalized. It hasbeen acknowledged that clathrin mediated endocytosis serves as amajor process of internalization of negatively charged nano-particles [47]. In order to study the involvement of the said processin our system, we pre-incubated the MCF-7 cells with 25 mM ofchlorpromazine (a specific clathrinmediated endocytosis inhibitor)for 30 min following treatment with all the three different TQ-Nps.Interestingly our observations revealed that chlorpromazine pre-
Fig. 2. (A) FTIR spectra of TQ, PVP, PEG200, PEG4000, PVP-TQ, PEG200-TQ and PEG4000-TQ NaPEG4000-TQ Nanoparticles at pH 7.4 and pH 5. (C) In vivo percentage of cellular uptake of Pafter cell lysis. (D) In vivo percentage of cellular uptake of PVP-TQ, PEG200-TQ and PEG4000-spectral analysis after cell lysis. (E) Analysis of percentage of cell death or cellular toxicity byTQ. (F) Analysis of percentage of cell survival by MTT assay on PBMC at different doses of PVPAnalysis of percentage of cell death by annexin-V-FITC on MCF-7 (left panel) and HBL-100 (r24 h of incubation with the NPs. **P < 0.01, and ***P < 0.001.
treated MCF-7 cells showed prominent decline in the cellular up-take of all the TQ-Nps thereby suggesting that clathrin-mediatedendocytosis is one of the prime processes involved in the cellularinternalization of the TQ-nanoparticles (Fig 2D).
3.5. In vitro cytotoxicity
In order to assess the cytotoxicity of TQ on both peripheral bloodmononuclear cells (PBMC) and breast cancer cells we performedthe flow-cytometric cell cycle phase distribution analysis.
noparticles. (B) In vitro cumulative release kinetics of TQ from PVP-TQ, PEG200-TQ andVP-TQ, PEG200-TQ and PEG4000-TQ Nanoparticles measured by UV/Vis spectral analysisTQ Nanoparticles in the presence and absence of chlorpromazine measured by UV/Visflowcytometric cell cycle analysis on PBMC, MCF-7, HBL-100 cells at different doses of-TQ, PEG200-TQ and PEG4000-TQ Nanoparticles after 24 h of incubation with the NPs. (G)ight panel) at different doses of PVP-TQ, PEG200-TQ and PEG4000-TQ Nanoparticles after

Fig. 2. (continued).
S. Bhattacharya et al. / Biomaterials 51 (2015) 91e10798
Interestingly our observations revealed significant death of theMCF-7 and HBL-100 cells with increasing concentrations of TQ(Fig. 2E), however TQ also generated prominent cytotoxicity in thePBMCs in a dose dependent manner (Fig. 2E). Contrastingly on theother hand, the percentage of PBMC cell survival as assessed byMTT assay was significantly higher when subjected to PVP-TQ-Np,PEG200-TQ-Np and PEG4000-TQ-Np respectively (Fig. 2F). Further-more, the effects of these TQ-Nps were also assessed on breastcarcinoma cell lines like MCF-7 and HBL-100. The results of the cellcycle analysis revealed that all the three TQ-NPs produced signifi-cant percentage of cell death (about 40e45 %) in a dose-dependentmanner in both MCF-7 (Fig. 2G, left panel) and HBL-100 (Fig 2G,right panel) cells, with PEG4000-TQ-Nps producing a prominentlyhigher percentage of cell death in comparison to the two other TQ-Nps. These findings thereby established the fact that the synthe-sized TQ-Nps showed prominent cytotoxicity in breast cancer cellsat a dose significantly lower than the dose inwhich they developedtoxicity in normal PBMC cells. Additionally, our findings alsohighlighted that PEG4000-TQ-Nps create toxicity in cancer cell linesmuch efficiently than the other two, levying the normal PBMC.
3.6. In vitro effect of TQ-Nps on retardation of breast cancer cellmigration
With the prime objective of investigating the anti-migratoryproperty of TQ-Nps on breast carcinoma cells, we first elucidatedthe role of TQ in inhibiting breast cancer cell migration usingin vitro bidirectional wound healing assay in MCF-7 and HBL-100cells. As illustrated in Fig. 3A and B, TQ significantly (p < 0.001)inhibited migration of both MCF-7 and HBL-100 cells respectivelyeven at a very low dose of 2 mg/ml. Next, we evaluated the indi-vidual anti-migratory properties of the three newly synthesizedTQ-Nps using the bidirectional wound healing assay. Results of theassay performed with the above mentioned cell lines highlightedthat all the three TQ-Nps were capable of retarding migration inboth MCF7 and HBL100 cells (Fig. 3C and D). However, furtheranalysis revealed that in comparison to the other two TQ-Nps,PEG4000-TQ-Np was much more effective in retarding migration
of both MCF-7 and HBL-100 even at a much lower dose of 1 mg/ml(Fig. 3C and D). These findings therefore highlighted the fact thateach of the newly synthesized TQ-Nps could prevent the in vitromotility of breast cancer cells at significantly lower doses than TQ,with PEG4000-TQ-Nps being themost effective of the all three. Thus,we were prompted to delineate specifically the effect of PEG4000-TQ-Nps on retardation of in vitro breast cancer cell migration incomparison with original TQ. To further confirm our above-mentioned findings we next performed the transwell migrationassay using both MCF-7 and HBL-100 cell lines. Both the cell lineswere treated with 2 mg/ml dose of TQ and with 1 mg/ml of PEG4000-TQ-Nps in a separate set. Interestingly, as illustrated in Fig. 3E,lesser number of cells were found to have migrated to the undersurface of the transwell insert when treated with PEG4000-TQ-Npsin comparison to a significant high number of migratory cells in thecase of TQ treatment. These observations discussed so far clearlyreinstate our findings that PEG4000-TQ-Nps display more potentanti-migratory properties than TQ and also allured us to discern thecomplete molecular mechanism underneath such anti-migratoryeffect of PEG4000-TQ-Nps on breast cancer cells.
3.7. Anti-migratory effect of PEG4000-TQ-Np on lamellipodiaformation through miR-34a
Lamellipodium is a broad, flat cell protrusion that dominates atthe edges of cultured fibroblasts and many other motile cells.Lamellipodia extensions determine generally the migratory abilityof cancer cells [24]. Henceforth we monitored the ultrastructure ofbothMCF-7 and HBL-100 cells under SEM to determine the changesin cellular morphology in presence and absence of PEG4000-TQ-Nps(at 1 mg/ml dose). Our findings interestingly revealed that incontrast to the numerous protrusions present on the surface of boththe MCF-7 and HBL-100 control cells (Fig. 4A) both the treated cellsshowed a smooth surface with significant less number of pro-trusions (Fig. 4A). The protrusion of the lamellipodium is known tobe provided by continuous growth of actin filaments towards theleading edge of the cells [22]. Therefore to further elucidate thepossible regulatory mechanism of PEG4000-TQ-Nps on cytoskeletal

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107 99
reorganization, we used phalloidin staining to observe the actindistribution pattern in both treated and untreated MCF7 cells. Ourresults significantly revealed less detectable F-actin and onlycortical actin in the PEG4000-TQ-Np-treated cells (Fig. 4B). Thesedata are therefore indicative of the fact that PEG4000-TQ-Nps wereable to prevent the formation of cellular protrusions on the surfaceby disrupting actin organization.
miRNAs are known to modulate a variety of cellular pathwaysrelated to cell development and differentiation [48,49] and therebyhave gained considerable interest in the study of cancer cellmigration. Thus we next focused our aim to identify the miRNAswhose differential expression in PEG4000-TQ-Np-treated cells inrespect to the control regulates the migratory potential of the cellsby controlling the cytoskeletal reorganization through deregulated
Fig. 3. The rate of migration of breast cancer cells; (A) MCF-7 and (B) HBL-100 on treatmenbidirectional wound healing assay (left panel). Graphical representation of the percentage oconcentration of TQ (2, 4, 6 mg/mL) (right panel). Percentage of migration of (C) MCF-7 anddifferent concentrations (0.5, 1, 2, 4 mg/mL) of PVP-TQ-Np, PEG200-TQ-Np and PEG4000-TQ-NpHBL-100 cells under the above mentioned conditions (right panel). (E) Phase contrast imagesPEG4000-TQ-Np (1 mg/ml) treated conditions (left panel). Percentage of cell migration of M(right panel). Values are mean ± SEM of three independent experiments in each case or re
actin polymerization. For this purpose, we performed a miRNAarray screening using qRT PCR. Six mature miRNAs related to cellmigration was therefore selected. On analysis of the array results,we observed that the expression of miR-34a was substantially up-regulated by 7 folds in MCF-7 cells on treatment with PEG4000-TQ-Np (Fig. 4C). In fact the expression of miR-34a was also found to beup-regulated in a time dependent fashion for a period of 24 h(Fig. 4D). To further reinstate the key role of mir 34a in PEG4000-TQ-Np induced retardation of breast cancer cell migration, cells werethen transfectedwithmiR-34a antisense strand synthesizing vector(psi-CHECK2-AS34a) and followed by treatment with PEG4000-TQ-Np. PEG4000-TQ-Np failed to show its anti-migratory effect in thesetransfected cells as determined from both bidirectional woundhealing as well as transwell migration assay (Fig. 4E). These results
t with different concentration of TQ (2, 4, 6 mg/mL) at 0 and 24 h as determined fromf migration of (A) MCF-7 (B) HBL-100 cells at 0 and 24 h on treatment with different(D) HBL-100 cells represented graphically at 0 and 24 h time point on treatment with(left panel). Scratch wound pictures showing the rate of migration of (C) MCF-7 and (D)showing the stained MCF-7 and HBL-100 migratory cells in untreated, TQ (2 mg/ml) andCF-7 and HBL-100 cells represented graphically under the aforementioned conditionspresentative of typical experiment. **P < 0.01, and ***P < 0.001.

Fig. 3. (continued).
S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107100
thereby highlight the importance of miR-34a in regulation of breastcancer cell migration and prompted us to further elucidate theintricate mechanisms involved in this process.
3.8. Effect of PEG4000-TQ-Np in regulation of miR-34a expression byp53
It has been acknowledged that miR-34a expression is chieflyregulated by p53 and it is also well known that TQ has the ability toup-regulate the expression of p53 [50]. So, it can be hypothesizedthat PEG4000-TQ-Nps might also up-regulate p53 which in turn,induced the expression of miR-34a in the breast cancer cell lines. Inorder to verify this hypothesis we performed the western blotanalysis to compare the p53 expression in PEG4000-TQ-Np-treatedand untreated MCF-7 cells which were also followed by immuno-fluorescences imaging. Our western blot analysis clearly high-lighted that p53 expressionwas prominently up-regulated in a timedependent manner in the MCF-7 cells on treatment with PEG4000-TQ-Np (Fig. 5A). Additionally, our immunofluorescence imagingalso confirmed the elevation in p53 level on PEG4000-TQ-Np treat-ment (Fig. 5B). Moreover, a time dependent study of the changes inthe mRNA expression pattern of p53 and miR-34a in treated anduntreated cells revealed that there exist a strong relationship be-tween p53 expression and miR-34a up-regulation. In fact ourfindings revealed that p53 expression was gradually elevated from4 h and continued up to 24 h, whereas miR-34awas found to be up-regulated, starting from 8 h onwards (Fig. 5C), thereby suggestingthe dependency of miR-34a expression on p53. To further validatethe importance of p53 in regulation of miR-34a expression, wetreated the cells with pifithrin-a (p53 activity inhibitor) followed bytreatment with PEG4000-TQ-Nps. Results of the experiment clearlyhighlighted that PEG4000-TQ-Nps could not up-regulate miR-34aexpression in presence of pifithrin-a (Fig 5D) andwas also unable toretard MCF-7 cell migration as evident from both wound healing
and transwell migration assay (Fig. 5E), thereby establishing thefact that blocking the activity of p53 impaired the process ofPEG4000-TQ-Nps mediated miR-34a up-regulation. All these evi-dences thereby indulged us to hypothesize that PEG4000-TQ-Npsmust be regulating the expression of p53 which induces itsdownstream effector miR-34a.
3.9. Modulation of rearrangement of actin cytoskeleton throughmir-34a by targeting Rac1
It has been well acknowledged that the protrusions, present onthe cell surface, play a significant role in regulating cell motility.Lamellipodium is one of the cell protrusions which are frequentlyfound at the leading edge of migratory cells [22]. Accumulatingevidences suggest that Rac1 actively induces actin polymerizationto form lamellipodia at the leading edge during cell migration [26].So, we next attempted to investigate whether PEG4000-TQ-Nps aretargeting Rac1 through p53/miR34a axis to regulate actin poly-merization thereby controlling breast cancer cell migration. Toevaluate this, MCF-7 cells were treated with 1 mg/ml of PEG4000-TQ-Nps and examined for both protein and mRNA expression of Rac1.Decreased Rac1 expression was observed from western blotting(Fig. 6A, left panel), in treated cells with respect to the control. Theexpression levels of the Rac1 transcripts was also found to besignificantly decreased in the PEG4000-TQ-Np treated sets asdetermined by qRT-PCR (Fig. 6A, middle panel) and by reversetranscriptase PCR (Fig. 6A, right panel). Thus all our findingscumulatively indicate that PEG4000-TQ-Nps down-regulates theexpression of Rac1 at both protein and mRNA levels therebyinterrupting the lamellipodia formation which is necessary for thecells to migrate. At this juncture, when it is evident that Rac1 isdown-regulated by PEG4000-TQ-Nps in breast cancer cells, now thequestion arises as to how PEG4000-TQ-Nps actually exert its

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107 101
negative regulation on Rac1. To reveal the answer, we have questedfor the miRNA-mediated regulatory mechanism of PEG4000-TQ-Np.
It has recently been established that miR-34a can target Rac1protein. Kim et al. (2012) verified their finding by 30-UTR assay [32].To further justify that whether in our system PEG4000-TQ-Npsinduced miR-34a up-regulation could suppress Rac1 inducedbreast cancer cell migration in vitro, we transfected the MCF-7 cellswith psi-CHECK2-AS34a. After transfection, cells were treated withPEG4000-TQ-Nps for stipulated time and qRTePCR as well aswestern blot analysis was performed. The results obtained signifi-cantly displayed that only PEG4000-TQ-Nps treated cells showedprominent down-regulation of Rac1 protein (Fig. 6B) as well asmRNA (Fig. 6C) whereas PEG4000-TQ-Np treatment in psi-CHECK2-AS34a transfected cells failed to down-regulate the expression ofRac1 protein (Fig. 6B) as well as mRNA (Fig. 6C) confirming thedirect regulation of Rac1 by miR-34a. To examine the possibleinvolvement of miR-34a in cytoskeletal reorganization by Rac1, weused phalloidin staining to observe actin polymerization in both
Fig. 4. (A) SEM photographs of MCF-7 (left panel) and HBL-100 (right panel) cells showing chindicating protrusion like lammelipodia and filopodia formation and yellow arrow indicatinofluorescence images of phalloidin staining in MCF-7 cells from untreated and PEG4000-TQ-Nexpression of six miRNAs in MCF-7 cells as determined by qRT-PCR from untreated and PEG4
in MCF-7 cells represented graphically in untreated and PEG4000-TQ-Np treated sets. (E) Petreated, miR-34a-psicheck2 and PEG4000-TQ-Np þ miR-34a-psicheck2 treated sets as deterimages of wound healing and transwell migration assay of MCF-7 cells at 0 and 24 h in unmean ± SEM of three independent experiments in each case or representative of typical expthis figure legend, the reader is referred to the web version of this article.)
plasmid transfected and non-transfected MCF-7 with furthertreatment of PEG4000-TQ-Np in transfected cells. Results of theexperiment revealed that PEG4000-TQ-Np treatment in psi-CHECK2-AS34a transfected cells failed to disorganize the F-actinformation and thereby maintained cell morphology and F-actinorganization (Fig. 6D).
Lamelliopodia formation studied by SEM was also found to beunaffected in, psi-CHECK2-AS34a transfected treatedwith PEG4000-TQ-Nps cells in respect to the control (Fig 6E). So, the above findingsfurther provoked us to hypothesize that over-expression of miR-34a by PEG4000-TQ-Nps suppresses cell migration via the down-regulation of Rac1. All these findings therefore are an indicativeof the fact that PEG4000-TQ-Nps are affecting the breast cancer cellmigration by disrupting the actin polymerization and lamellipodiaformation through suppressing Rac1 by miR-34a. So, we proposedone probable mode-of-action of PEG4000-coated-TQ-Nps in thisstudy. Notably, this is the first report which elucidates TQ's inhib-itory effect on Rac1 via miR-34a.
anges in cellular morphology in untreated and PEG4000-TQ-Np treated sets. Blue arrowsng smooth surfaces of cell with no lammelipodia and filopodia formation. (B) Immu-p treated sets. Arrows indicating deregulated actin or cortical actin. (C) Relative miRNA
000-TQ-Np treated sets. (D) Relative expression of miR-34A in a time dependent mannerrcentage of MCF-7 cell migration represented graphically in untreated, PEG4000-TQ-Npmined from wound-healing and transwell migration assay (left panel). Phase contrasttreated and PEG4000-TQ-Np þ miR-34a-psicheck2 treated sets (right panel). Values areeriment. **P < 0.01, and ***P < 0.001. (For interpretation of the references to colour in

Fig. 4. (continued).
Fig. 5. (A) Western blot showing p53 expression levels in MCF-7 cells treated with PEG4000-TQ-Np in at different time points (0,4,8,12,18 and 24 h). (B) Nuclear localization of p53 inMCF-7 cells on treatment with PEG4000-TQ-Np determined using fluorescence microscopy. Arrows indicating colocalization of DAPI and FITC. The Pearson's coefficient determiningthe level of overlap of DAPI and FITC on treatment PEG4000-TQ-Np with has been represented graphically. (C) Changes in the relative m/miRNA expression pattern of p53 and miR-34A in a time dependent manner as represented graphically. (D) Changes in the relative miR-34A expression represented graphically in untreated, PEG4000-TQ-Np, Pifithrin a,PEG4000-TQ-Np þ Pifithrin a treated set. (E) Graphical representations of the percentage of MCF-7 cell migration as determined fromwound-healing and transwell migration assayin the aforementioned sets. a-actin/GAPDH was used as internal loading control. Values are mean ± SEM of three independent experiments in each case or representative of typicalexperiment. **P < 0.01, and ***P < 0.001.
S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107102

Fig. 6. (A) Changes in the expression level of protein and mRNA of Rac1 in untreated and PEG4000-TQ-Np treated sets. (B) Changes in the Rac1 expression level in untreated,PEG4000-TQ-Np, miR-34Apsicheck2 and miR-34Apsicheck2 þ PEG4000-TQ-Np treated sets as determined from western blotting. (C) Graphical representation of the changes in themRNA expression of Rac1 in the aforementioned sets. (D) Immuno fluorescence images of phalloidin staining in MCF-7 cells from untreated and PEG4000-TQ-Np þ miR-34Apsi-check2 treated sets. Arrows indicating polymerized actin. (E) SEM photographs of MCF-7 cells showing changes in cellular morphology in untreated and PEG4000-TQ-Np þ miR-34Apsicheck2 treated sets. Blue arrows indicating protrusion like lammelipodia and filopodia formation. a-actin was used as internal loading control. Values are mean ± SEM ofthree independent experiments in each case or representative of typical experiment. **P < 0.01, and ***P < 0.001. (For interpretation of the references to colour in this figure legend,the reader is referred to the web version of this article.)
S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107 103
3.10. In vivo validation of the effect of PEG4000-TQ-Nps in Rac1downregulation through p53 -miR-34a expression
In an attempt to validate the role of PEG4000-TQ-Nps in regu-lating Rac1 expression through p53/miR-34a axis in breast cancer
cells in vivo, we next assessed the effect of nanoparticles on solidtumors induced by EAC injection in the thigh muscle of Swiss al-bino mice. Based on pilot studies, 2.5 mg/kg and 5 mg/kg bodyweight of PEG4000-TQ-Nps was used for treatment in animalsbearing tumors. After 12th day of EAC injection (small size tumor

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107104
was visible), the animals were treated with specific doses everyalternate day. We found that treatment with PEG4000-TQ-Nps onanimals bearing tumor resulted in significant reduction of tumorsize compared to that of untreated (Fig. 7A). The gross appearance
Fig. 7. (A) Typical photographs of EAC solid tumor-bearing mice and individual excised tumofrom control and PEG4000-TQ-Np treated sets were measured, analyzed and represented in twere measured weekly, analyzed and plotted graphically. (C) Photographs of spleen excised(D) Tabular representation showing the changes in spleenic mononuclear cell number on trthe control and treated sets were analyzed histopathologically using eosin haematoxylin stathe excised tumors from the control and PEG4000-TQ-Np treated sets using anti-p53 antiboRac1 in the control and PEG4000-TQ-Np treated tumors. (H) Graphical representation of p53actin was used as internal loading control. Values are mean ± SEM of three independen***P < 0.001.
of thigh tissue in the tumor bearing and PEG4000-TQ-Nps treatedtumor bearing mice showed a proportional morphological differ-ence (Fig 7A). More importantly, we also found that upon treatmentwith PEG4000-TQ-Nps, animals with tumor showed a significant
r from untreated, 2.5 mg/kg, and 5 mg/kg treated sets. (B) Weight of the excised tumorshe form of bar graph; Tumor volumes of the control and PEG4000-TQ-Np treated tumorsfrom control, tumor-bearing and PEG4000-TQ-Np (5 mg/kg body weight) treated mice.eatment with different doses of PEG4000-TQ-Np for six weeks. (E) Tumors excised fromining and phase contrast images were captured. (F) Immuno-histochemical analysis ofdy. (G) Western blot analysis depicting the changes in the expression level of p53 andand Rac1 mRNA expression and also miR-34A expression in control and treated sets. a-t experiments in each case or representative of typical experiment. **P < 0.01, and

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107 105
difference in the survival rate compared to the untreated tumorcontrol (data not shown). While control animals survived for only amaximum of 60 days after tumor development, majority of micetreated with PEG4000-TQ-Nps survived for more than 150 daysindicating a,2-fold increase in life span (data not shown). Therefore,our results demonstrate that PEG4000-TQ-Nps treatment signifi-cantly reduced the tumor load and increased the lifespan of theanimals. Our further experiments were carried out using the 5 mg/kg dose of PEG4000-TQ-Nps.Our findings also clearly elucidated thatPEG4000-TQ-Nps treatment resulted in significant decline in tumorweight (Fig. 7B) and tumor volume (Fig. 7B). On the other hand, itwas also observed that the tumor-induced splenomegaly wasrestored in the Np-treated sets (Fig. 7C) The nanoparticle exhibitedmost prominent effect at the above mentioned dose and also thedepletion in the splenic mononuclear cell (MNC) number producedby the growing tumor was restored close to the normal value ontreatment with PEG4000-TQ-Np (Fig. 7D). Histological evaluationwas also performed at specific time point (45th day) of treatment.Tissue sections from the control tumors of thigh showed higherinfiltration and poorly differentiated structures in comparison tothose treated with PEG4000-TQ-Nps (Fig. 7E).
In order to know the status of p53, tumormasses were sectionedfor western blotting as well as immuno-histochemical staining. Ourresult clearly illustrated an increased expression status of p53 inPEG4000-TQ-Np-treated tumors as also confirmed by immuno-histochemical analysis (Fig. 7F) and from western blotting(Fig. 7G) and which re-confirms the p53-induced anti-migratoryeffects of the nanoparticles on tumor cells. Moreover, Rac1 pro-tein expression was also found to be significantly down-regulatedin PEG4000-TQ-Np-treated tumors in comparison to the untreatedones. The mRNA and miRNAs were isolated to further verify theeffect of PEG4000-TQ-Nps in the in vivo models. Results of qRT PCRanalysis revealed significant up-regulation of p53 and miR-34a andprominent down-regulation of Rac1 in PEG4000-TQ-Np-treated
Fig. 8. (A) Blood was collected after sacrifice from the mice from control and PEG4000-TQ-Npand represented graphically. (B) Levels of (i) malonaldehyde, (ii) catalase activity and (iii) Smean ± SEM of three independent experiments in each case or representative of typical ex
tumors compared to the control (Fig 7H). Thus, all these findingsare clearly indicative of the fact that PEG4000-TQ-Nps are capable ofdown-regulating Rac1 through miR-34a/p53 axis in both in vitroand in vivo systems.
3.11. Protection of tumor-bearing mice from cancer inducedsystemic toxicity by PEG4000-TQ-Nps
It has been well documented that cancer causes systemictoxicity and hepatic damage which can be evidenced by increase inthe levels of serum toxicity markers e lactate dehydrogenase(LDH), alkaline phosphatase (ALP), aspartate transaminase (AST)and alanine transaminase (ALT) [51]. Our experimental findingsalso showed that LDH level was increased 15-fold in the tumorbearing mice as compared to normal counterparts. However,treatment with the PEG4000-TQ-Nps decreased the LDH level byapproximately 7 and 9 folds in a dose dependent manner (Fig. 8Ai).Tumor bearingmice also showed a significant increase in the serumlevels of ALT, AST and ALP when compared to non treated controlsets. PEG4000-TQ-Nps treatment however significantly restored thevalues of ALT (20% and 52% decrease) (Fig. 8Aii), AST (20% and 52%decrease) (Fig. 8Aiii) and ALP (26.8% and 45.2% decrease) (Fig. 8Aiv)towards normal in a dose dependent manner.
In the present study it was observed that the anti-oxidant de-fense system of the liver of the tumor bearing mice was disturbedas compared to their normal counterparts. Our results indicatedthat in the tumor bearing mice the hepatic level of malonaldehyde,an index of lipid peroxidation, significantly increased 3 folds(Fig. 8Bi) while the activities of the anti-oxidant enzymes catalase(Fig. 8Bii) and SOD (Fig. 8Biii) was declined by 70% and 65% ascompared to normal mice indicating the presence of oxidativestress in the liver of the tumor bearing mice. Catalase is an anti-oxidant enzyme that catalyses the detoxification of hydrogenperoxide to oxygen and water. The catalase activity in the tumor-
treated sets and the serum levels of (i) LDH, (ii) ALT, (iii) AST and (iv) ALP was measuredOD activity were also measured subsequently in the aforementioned sets. Values areperiment. **P < 0.01, and ***P < 0.001.

Fig. 9. Schematic illustration depicting the molecular mechanisms of PEG4000-TQ-Np mediated retardation of breast cancer cell migration.
S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107106
bearing group decreased about ~70% compared with the controlgroup. PEG4000-TQ-Np treatment in tumor bearing mice showedsubstantial increase in catalase activity (Fig. 8Bii). Interestingly, ourfindings also revealed that PEG4000-TQ-Np treatment was able torestore the levels of both SOD in tumor-bearing mice (Fig. 8Biii).Modulation of these enzymes by PEG4000-TQ-Np in the liver of thetumor-bearing mice prevented oxidative stress which in turnameliorated the overall systemic toxicity (see Fig. 9).
All the above-described results cumulatively illustrate the suc-cessful synthesis of PEG4000-coated TQ-nanoparticles and alsojustify its potent anti-metastatic activity on breast carcinoma cellswith least systemic toxicity.
4. Conclusion
It has been demonstrated in the present study that the water-soluble PEG4000-TQ-Nanoparticles exhibited strong specificity tocancer cell migration, showing less toxicity towards the normalcells in comparison to that of TQ alone. Our findings also clearlyhighlighted that PEG4000-TQ-NPs were able to up-regulate miR-34ain a p53-dependent manner thereby retarding cell migrationwhichis the initial event of themetastatic cascade. Subsequently, miR-34aup-regulation also inhibited Rac1 mRNA and protein expressionwhich ultimately hindered cytoskeletal actin polymerizationthereby preventing the formation of membrane protrusion likelamellipodia and filopodia. Our in vivo result also supported thein vitro findings that Rac1 down-regulation was mediated throughthe p53/miR-34a axis on treatment with PEG4000-TQ-NPs. ThusPEG4000-TQ-NPsmay act as a novel nano-preparationwhichmay be
extremely effective in targeting migratory breast cancer cells andmay also be explored for treatment of other metastatic cancers.Hence we expect that these explorations can provide a new strat-egy for the development of nano-medicine and suggest PEG4000-TQ-NPs as a strong candidate for a newgeneration of chemotherapywith reduced side effects for treatment of cancer patients in the notso distant future.
Acknowledgment
The work has been supported by Department of Science andTechnology, DST INSPIRE Faculty Scheme (IFA12-LSBM-40) Govt. ofIndia and Center for Research in Nanoscience and Nanotechnology[Sanction no: Conv/126/Nano Pr. (2014)], University of Calcutta. Wealso acknowledge the central instrumental facility of Center forResearch in Nanoscience and Nanotechnology,University of Cal-cutta. Authors would also like to thank Prof. Gaurishankar Sa, AbirKumar Panda and Sayantan Bose (Division of Molecular Medicine,Bose Institute, Kolkata), Dr. Ankita Ghatak (S.N. Bose Center forBasic Science, Kolkata), Dr. Govinda Gopal Khan (Centre forResearch in Nanoscience and Nanotechnology, University of Cal-cutta, Kolkata) for technical help. P.P acknowledge UGC, GOI forDSK-PDF.
Appendix A. Supplementary data
Supplementary data related to this article can be found online athttp://dx.doi.org/10.1016/j.biomaterials.2015.01.007.

S. Bhattacharya et al. / Biomaterials 51 (2015) 91e107 107
References
[1] Gali-Muhtasib H, Roessner A, Schneider-Stock R. Thymoquinone: a promisinganti-cancer drug from natural sources. Int J Biochem Cell Biol 2006;38:1249e53.
[2] Woo CC, Kumar AP, Sethi G, Tan KH. Thymoquinone: potential cure for in-flammatory disorders and cancer. Biochem Pharmacol 2012;83:443e51.
[3] Mansour MA, Nagi MN, El-Khatib AS, Al-Bekairi AM. Effects of thymoquinoneon antioxidant enzyme activities, lipid peroxidation and DT-diaphorase indifferent tissues of mice: a possible mechanism of action. Cell Biochem Funct2002;20:143e51.
[4] Umar S, Zargan J, Umar K, Ahmad S, Katiyar CK, Khan HA. Modulation of theoxidative stress and inflammatory cytokine response by thymoquinone in thecollagen induced arthritis in Wistar rats. Chem Biol Interact 2012;197:40e6.
[5] Pari L, Sankaranarayanan C. Beneficial effects of thymoquinone on hepatic keyenzymes in streptozotocin-nicotinamide induced diabetic rats. Life Sci2009;85:830e4.
[6] El-Mahmoudy A, Matsuyama H, Borgan MA, Shimizu Y, El-Sayed MG,Minamoto N, et al. Thymoquinone suppresses expression of inducible nitricoxide synthase in rat macrophages. Int Immunopharmacol 2002;2:1603e11.
[7] Banerjee S, Padhye S, Azmi A, Wang Z, Philip PA, Kucuk O, et al. Review onmolecular and therapeutic potential of thymoquinone in cancer. Nutr Cancer2010;62:938e46.
[8] Gali-Muhtasib H, Ocker M, Kuester D, Krueger S, El-Hajj Z, Diestel A, et al.Thymoquinone reduces mouse colon tumor cell invasion and inhibits tumorgrowth in murine colon cancer models. J Cell Mol Med 2008;12:330e42.
[9] Kolli-Bouhafs K, Boukhari A, Abusnina A, Velot E, Gies JP, Lugnier C, et al.Thymoquinone reduces migration and invasion of human glioblastoma cellsassociated with FAK, MMP-2 and MMP-9 down-regulation. Invest New Drugs2012;30:2121e31.
[10] Ahmad I, Muneer KM, Tamimi IA, Chang ME, Ata MO, Yusuf N. Thymoquinonesuppresses metastasis of melanoma cells by inhibition of NLRP3 inflamma-some. Toxicol Appl Pharmacol 2013;270:70e6.
[11] Jafri SH, Glass J, Shi R, Zhang S, Prince M, Kleiner-Hancock H. Thymoquinoneand cisplatin as a therapeutic combination in lung cancer: in vitro and in vivo.J Exp Clin Cancer Res 2010;29:87.
[12] Schneider-Stock R, Fakhoury IH, Zaki AM, El-Baba CO, Gali-Muhtasib HU.Thymoquinone: fifty years of success in the battle against cancer models. DrugDiscov Today 2014;19:18e30.
[13] Murphy EA, Majeti BK, Barnes LA, Makale M, Weis SM, Lutu-Fuga K, et al.Nanoparticle-mediated drug delivery to tumor vasculature suppressesmetastasis. Proc Natl Acad Sci U S A 2008;105:9343e8.
[14] Sultana S, Khan MR, Kumar M, Kumar S, Ali M. Nanoparticles-mediated drugdelivery approaches for cancer targeting: a review. J Drug Target 2013;21:107e25.
[15] Bai F, Wang C, Lu Q, Zhao M, Ban FQ, Yu DH, et al. Nanoparticle-mediated drugdelivery to tumor neovasculature to combat P-gp expressing multidrugresistant cancer. Biomaterials 2013;34:6163e74.
[16] Martindale W, Reynolds JEF, Royal Pharmaceutical Society of Great Britain.The extra pharmacopoeia. 29th ed. London: Department of PharmaceuticalSciences, Pharmaceutical Press; 1989.
[17] Greenwald RB, Zhao H, Xia J. Tripartate poly(ethylene glycol) prodrugs of theopen lactone form of camptothecin. Bioorg Med Chem 2003;11:2635e9.
[18] Sun H, Guo B, Cheng R, Meng F, Liu H, Zhong Z. Biodegradable micelles withsheddable poly(ethylene glycol) shells for triggered intracellular release ofdoxorubicin. Biomaterials 2009;30:6358e66.
[19] Wu L, Tang C, Yin C. Folate-mediated solid-liquid lipid nanoparticles forpaclitaxel-coated poly(ethylene glycol). Drug Dev Ind Pharm 2010;36:439e48.
[20] Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science2011;331:1559e64.
[21] Condeelis JS, Wyckoff JB, Bailly M, Pestell R, Lawrence D, Backer J, et al.Lamellipodia in invasion. Semin Cancer Biol 2001;11:119e28.
[22] Small JV. Lamellipodia architecture: actin filament turnover and the lateralflow of actin filaments during motility. Semin Cell Biol 1994;5:157e63.
[23] Zigmond SH. Signal transduction and actin filament organization. Curr OpinCell Biol 1996;8:66e73.
[24] Van Aelst L, D'Souza-Schorey C. Rho GTPases and signaling networks. GenesDev 1997;11:2295e322.
[25] DerMardirossian C, Bokoch GM. Regulation of cell function by Rho GTPases.Drug News Perspect 2001;14:389e95.
[26] Ridley AJ, Paterson HF, Johnston CL, Diekmann D, Hall A. The small GTP-binding protein rac regulates growth factor-induced membrane ruffling. Cell1992;70:401e10.
[27] Lee TK, Man K, Ho JW, Wang XH, Poon RT, Sun CK, et al. Significance of the Racsignaling pathway in HCC cell motility: implications for a new therapeutictarget. Carcinogenesis 2005;26:681e7.
[28] Bouzahzah B, Albanese C, Ahmed F, Pixley F, Lisanti MP, Segall JD, et al. Rhofamily GTPases regulate mammary epithelium cell growth and metastasisthrough distinguishable pathways. Mol Med 2001;7:816e30.
[29] Uhlenbrock K, Eberth A, Herbrand U, Daryab N, Stege P, Meier F, et al. TheRacGEF Tiam1 inhibits migration and invasion of metastatic melanoma via anovel adhesive mechanism. J Cell Sci 2004;117:4863e71.
[30] Fritz G, Brachetti C, Bahlmann F, Schmidt M, Kaina B. Rho GTPases in humanbreast tumours: expression and mutation analyses and correlation withclinical parameters. Br J Cancer 2002;87:635e44.
[31] Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell2009;136:215e33.
[32] Kim D, Song J, Kim S, Park HM, Chun CH, Sonn J, et al. MicroRNA-34a mod-ulates cytoskeletal dynamics through regulating RhoA/Rac1 cross-talk inchondroblasts. J Biol Chem 2012;287:12501e9.
[33] Sun XJ, Liu H, Zhang P, Zhang XD, Jiang ZW, Jiang CC. miR-10b promotesmigration and invasion in nasopharyngeal carcinoma cells. Asian Pac J CancerPrev 2013;14:5533e7.
[34] Huang C, Li H, Wu W, Jiang T, Qiu Z. Regulation of miR-155 affects pancreaticcancer cell invasiveness and migration by modulating the STAT3 signalingpathway through SOCS1. Oncol Rep 2013;30:1223e30.
[35] Wang D, Liu D, Gao J, Liu M, Liu S, Jiang M, et al. TRAIL-induced miR-146aexpression suppresses CXCR4-mediated human breast cancer migration.FEBS J 2013;280:3340e53.
[36] Wu L, Cai C, Wang X, Liu M, Li X, Tang H. MicroRNA-142-3p, a new regulatorof RAC1, suppresses the migration and invasion of hepatocellular carcinomacells. FEBS Lett 2011;585:1322e30.
[37] Li X, Ling N, Bai Y, Dong W, Hui GZ, Liu D, et al. MiR-16-1 plays a role inreducing migration and invasion of glioma cells. Anat Rec Hob 2013;296:427e32.
[38] Odeh F, Ismail SI, Abu-Dahab R, Mahmoud IS, Al Bawab A. Thymoquinone inliposomes: a study of loading efficiency and biological activity towards breastcancer. Drug Deliv 2012;19:371e7.
[39] Alam S, Khan ZI, Mustafa G, Kumar M, Islam F, Bhatnagar A, et al. Develop-ment and evaluation of thymoquinone-encapsulated chitosan nanoparticlesfor nose-to-brain targeting: a pharmacoscintigraphic study. Int J Nanomed2012;7:5705e18.
[40] Gupta V, Aseh A, Rios CN, Aggarwal BB, Mathur AB. Fabrication and charac-terization of silk fibroin-derived curcumin nanoparticles for cancer therapy.Int J Nanomed 2009;4:115e22.
[41] Saha B, Adhikary A, Ray P, Saha S, Chakraborty S, Mohanty S, et al. Restorationof tumor suppressor p53 by differentially regulating pro- and anti-p53 net-works in HPV-18-infected cervical cancer cells. Oncogene 2012;31:173e86.
[42] Claiborne AK. Catalase activity. In: Greenwald RA, editor. CRC handbook ofmethods for oxygen radical research. Boca Raton, Fla: CRC Press; 1985.p. 283e4.
[43] Wright JR, Colby HD, Miles PR. Cytosolic factors which affect microsomal lipidperoxidation in lung and liver. Arch Biochem Biophys 1981;206:296e304.
[44] Marklund SL. Superoxide dismutase isoenzymes in tissues and plasma fromNew Zealand black mice, nude mice and normal BALB/c mice. Mutat Res1985;148:129e34.
[45] Gaber NN, Darwis Y, Peh KK, Tan YT. Characterization of polymeric micellesfor pulmonary delivery of beclomethasone dipropionate. J Nanosci Nano-technol 2006;6:3095e101.
[46] Rastogi R, Sultana Y, Aqil M, Ali A, Kumar S, Chuttani K, et al. Alginate mi-crospheres of isoniazid for oral sustained drug delivery. Int J Pharm 2007;334:71e7.
[47] Vranic S, Boggetto N, Contremoulins V, Mornet S, Reinhardt N, Marano F, et al.Deciphering the mechanisms of cellular uptake of engineered nanoparticlesby accurate evaluation of internalization using imaging flow cytometry. PartFibre Toxicol 2013;10:2.
[48] Nakahara K, Carthew RW. Expanding roles for miRNAs and siRNAs in cellregulation. Curr Opin Cell Biol 2004;16:127e33.
[49] Fineberg SK, Kosik KS, Davidson BL. MicroRNAs potentiate neural develop-ment. Neuron 2009;64:303e9.
[50] Gali-Muhtasib H, Diab-Assaf M, Boltze C, Al-Hmaira J, Hartig R, Roessner A,et al. Thymoquinone extracted from black seed triggers apoptotic cell death inhuman colorectal cancer cells via a p53-dependent mechanism. Int J Oncol2004;25:857e66.
[51] Cao R, Wang LP. Serological diagnosis of liver metastasis in patients withbreast cancer. Cancer Biol Med 2012;9:57e62.




















