
Sweat gland differentiation in mammary adenoid cystic carcinoma
Upload
independentCategory
view
1download
0
DEREGULATION OF STAT5 EXPRESSION AND ACTIVATION CAUSESMAMMARY TUMORS IN TRANSGENIC MICEElena IAVNILOVITCH
1, Robert D. CARDIFF2, Bernd GRONER
3 and Itamar BARASH1*
1Institute of Animal Science, Agricultural Research Organization, Volcani Center, Bet-Dagan, Israel2Center of Comparative Medicine, University of California at Davis, Davis, CA, USA3Georg Speyer Haus, Institute for Biomedical Research, Frankfurt am Main, Germany
Members of the signal transducers and activators of tran-scription (Stat) family regulate essential cellular growth andsurvival functions in normal cells and have also been impli-cated in tumorigenesis. We have studied the potential role ofStat5 in mammary tumorigenesis by targeting Stat5 variantsto the mammary gland of transgenic mice using regulatorysequences of the �-lactoglobulin gene. Mammary-directedexpression of the wild-type Stat5, constitutively activatedStat5 and carboxyl-terminally truncated dominant negativeStat5 forms resulted in mammary tumors with incidencerates of up to 22% and latency periods of 8–12 months.Undifferentiated carcinomas most frequently occurred inmice expressing the carboxyl-terminally truncated Stat5.The more differentiated papillary and micropapillary adeno-carcinomas were primarily found in mice overexpressing thenative and constitutively active transgenes. Higher levels oftranslation initiation factor 4E (eIF4E) and cyclin D1 expres-sion but lower levels of activated Stat3 were found in tumorsof mice expressing the constitutively active Stat5 when com-pared to mice expressing the wild-type or truncated forms. Ahigher expression of the estrogen receptor (ER�) was ob-served in carcinomas compared to other phenotypes. Theability of both forms of Stat5, the transactivating form andthe dominant negative form, to participate in oncogenesisindicates that there is more than one mechanism by whichStat5 contributes to this process. The transactivation func-tion of Stat5 is involved in the determination of tumors witha more differentiated phenotype.© 2004 Wiley-Liss, Inc.
Key words: transgenic mice; mammary gland; Stat5; cancer; onco-gene
The mammary gland is a highly dynamic organ in which cellproliferation and cell death establish a delicate balance to maintainfunction and differentiation. The mouse mammary gland developsin the embryo from an epithelial bud, which penetrates the under-lying mesenchyme. It forms a rudimentary branched ductal sys-tem, invading the fat pad under the hormonal stimulus providedduring puberty. In the adult mammary gland, massive proliferationof the epithelial cells occurs during pregnancy. The cells form theside branches of the main ducts and organize into secretory alveoli,which function upon lactation. The basic mammary units are theterminal ductal lobular unit (TDLU) and the lobuloalveolar unit(LA). Both share similarities in terms of cell composition anddynamic activities in response to steroid and polypeptide hor-mones. These units are also the sites of origin for most mammarycancer.1,2
Breast cancer is the second leading cause of cancer-relateddeaths in women in the Western world. It is a multistep diseaseinitiated and progressing through genetic changes in oncogenesand tumor suppressor genes.3,4 To investigate the molecular mech-anisms involved in mammary tumorigenesis, candidate genes sus-pected to be involved in tumor development have been targetedand expressed in the mammary gland of transgenic mice. Tumorshave been observed, some of which resemble the pathology ofhuman breast cancer. In some cases, a characteristic transgene-specific phenotype, a signature tumor phenotype, has been found.The pathologic characteristics of a specific genetic alteration mighttherefore be reflected in a distinct tumor phenotype.5
Signal transducer and activator of transcription (Stat) proteinsare latent transcription factors, activated by tyrosine phosphoryla-tion downstream of cytokine and growth factor receptors. Theyregulate cell proliferation, differentiation and survival.6 Conservedregions in each of the 7 members of the Stat family mediate theiractivation. This is initiated by ligand-induced phosphorylation of atyrosine residue located near the carboxyl terminus. Subsequently,2 Stat molecules dimerize through phosphotyrosine and SH2 do-main interactions. Stat dimers translocate into the nucleus, and theamino terminal region mediates their oligomerization and stabili-zation. Stats bind to specific gene promoters via their centralDNA-binding region. Interactions with coactivators are requiredfor the initiation of transcriptional activity. These molecules inter-act with the carboxyl terminal transactivation domain (TAD). Stat5not only is able to induce transcription, but can also act as atranscriptional repressor. This function requires the interaction ofa nuclear corepressor with the coiled-coil domain of Stat5 and therecruitment of histone deacetylases to the protein complex.7
Two of the Stat family members, Stat3 and Stat5, regulatefunctionality and survival of mammary epithelial cells. Stat3 isactivated at the onset of involution.8–10 Constitutive activity ofStat3 has been detected in a human breast cancer cell lines andtumors.11–13 Programmed breast cancer cell death was induced bya dominant negative variant of the molecule.14 Stat5a and Stat5bare nearly identical genes located on mouse chromosome 11 ad-jacent to the Stat3 locus.15 Stat5a is mandatory for terminal mam-mary development and cell survival.16,17 Stat5a exerts its activityon milk protein gene promoters by binding to specific DNAelements (5�-TTCNNNGAA-3�).18–21 Truncated Stat5 molecules,lacking the carboxyl terminal TAD, have been shown to exertstrong dominant negative effects on milk promoter activity intransfected cells.10,22
Abbreviations: BLG, �-lactoglobulin; DMEM, Dulbecco’s modified Ea-gle’s medium; eIF4E, translation initiation factor 4E; ER, estrogen recep-tor; FCS, fetal calf serum; H&E, hematoxylin and eosin; Jak, janus kinase;LA, lobuloalveolar unit; PA�, poly A RNA; PBS, phosphate-bufferedsaline; PCNA, proliferating cell nuclear antigen; PRL, prolactin; RT-PCR,reverse transcription-PCR; SMA, smooth muscle actin; Stat, signal trans-ducer and activator of transcription; STAT5, transgenic Stat5; STAT5ca,constitutively active Stat5 transgene; STAT5�750, dominant negativeStat5 transgene; TAD, transactivation domain; TDLU, terminal ductallobular unit; TGF, transforming growth factor.
Grant sponsor: the German-Israeli Research Fund; Grant sponsor: IsraeliMinistry of Agriculture; Grant sponsor: the National Institutes of Health,National Regional Resource Centers; Grant number: U42RR14905.
*Correspondence to: Institute of Animal Science, ARO, Volcani Center,P.O. Box 6, Bet-Dagan 50250, Israel. Fax: �972-8-947-5075.E-mail: [email protected]
Received 9 February 2004; Accepted after revision 26 May 2004
DOI 10.1002/ijc.20484Published online 30 July 2004 in Wiley InterScience (www.interscience.
wiley.com).
Int. J. Cancer: 112, 607–619 (2004)© 2004 Wiley-Liss, Inc.
Publication of the International Union Against Cancer
Deregulated Stat5 activity has been observed in a variety ofblood-derived malignancies23 and Stat5a activity has been associ-ated with mammary tumorigenesis in transgenic mice expressingtransforming growth factor (TGF) � or the SV40 T antigen. Inthese mice, inactivation of Stat5a delayed hyperplasia17 and mam-mary cancer progression.24 We investigated the expression ofdifferent Stat5 variants and the deregulation they impose on Stat5wild-type function in the process of mammary cell transformation.For this purpose, we generated transgenic mice expressing thenative, constitutively active and carboxyl-terminally truncated, i.e.,dominant negative, variants of Stat5. Our data suggest that thederegulated expression of Stat5 and enhanced activation as well assuppressed transcriptional activation by Stat5 contribute to mam-mary tumorigenesis in transgenic mice.
MATERIAL AND METHODS
Plasmids and constructsThree Stat5-based constructs were employed: the wild-type ver-
sion of sheep Stat5a, termed STAT5, linked to an influenza virusepitope for detection by specific antibodies;25 constitutively acti-vated STAT5, termed STAT5ca, comprising sequences from 3genes: amino acids 1–750 from sheep Stat5, 677–847 from humanStat6 and 757–1129 from mouse janus kinase (Jak) 2;26 a deletedconstruct, STAT5�750, prepared by introducing a stop codon atthe respective site of the native Stat5 DNA sequence.22 Theseconstructs were inserted into the �-lactoglobulin (BLG) multiple-cloning site as previously described.10 To avoid biphasic expres-sion, the natural BLG ATG translation-initiation codon, as well asa second potential initiation codon in BLG exon 1, was convertedinto noninitiating ATT and ATC sequences. BLG sequences be-tween the first part of BLG exon 2 and the middle of BLG exon 6(including the TAG termination codon) were deleted and replacedwith a multiple cloning site.
Generation and identification of transgenic miceMice used in this study were of the FVB/N strain. Plasmids
containing sequences of BLG/STAT5, BLG/STAT5ca and BLG/STAT5�750 were digested by SalI and the appropriate fragmentswere prepared and microinjected into fertilized mouse oocytes.Transgenic animals were identified by Southern blot analysis ofgenomic DNA prepared from tail biopsies using a probe encom-passing 3 kb of the BLG promoter. All animals were visuallyinspected for tumors at least once a week and allowed to live forup to 2 years.
Messenger RNA measurement by reverse transcription-PCRTotal RNA was isolated from mammary glands of lactating
mice with TRIzol (Gibco-BRL, Gaithersburg, MD) according tothe manufacturer’s protocol, and 150 mg aliquots were treated withDNase (Roche, Mannheim, Germany). Poly A (pA�) RNA wasseparated on oligo(dT) columns with an mRNA isolation kit(Boehringer, Mannheim, Germany) and 250 �g aliquots werereverse-transcribed at 37°C for 50 min as previously described.10
PCRs for STAT5, STAT5ca and STAT5�750 were performed in50 �l of 750 mM Tris-HCl buffer, pH 9.0, containing 1.5 �l of thereverse-transcribed reaction, 200 mM (NH4)SO4, 0.1% (w/v)Tween, 1.25 mM MgCl2, 1 U of Taq polymerase (Gibco-BRL) and1.5 �l of primers, 10 mM each.
The set of primers used for STAT5 and STAT5�750 amplifi-cation were 5�-CTCTTTACTCCTGCCAGAGGCTC-3� and 5�-GTCTGTCCCCAGAGGAGTCCAAG-3� originated from the 3�sequences of Stat5 and 5� sequences of BLG exon 6, and ampli-fying a 537 bp fragment, mainly of the Stat5 sequence. The set ofprimers used for STAT5ca amplification were 5�-TAGCTGCGT-GAAAGAGATGG-3� and 5�-TGCCTTTATTGGTGAAGGAGA-3�originated from the 3� sequences of Jak2 and 5� sequences of BLGexon 7, and amplifying a 726 bp fragment, mainly of the Jak2
TABLE I – LIST OF THE ANTIBODIES APPLIED IN THE IMMUNOBLOT ANALYSES
ProteinAntigen Primary antibody Secondary antibody
Mr (kDa) Source Host/dilution Manufacturer Host/dilution Manufacturer
p27 27 Human Rabbit 1:1,000 Chemicon,Temecula,CA
Goat 1:3,000 Zymed Laboratories,South San Francisco,CA
CK 18 45 Human Mouse 1:300 Progen,Heidelberg,Germany
Goat 1:3,000 Zymed
�-casein 29 Mouse Rabbit 1:250 Barash et al.54 Donkey 1:20,000 Amersham,Buckinghamshire U.K.
CyclinD1
36 Human, aa 151–170
Mouse 1:3,000 Cell SignalingTechnology,Beverly,MA
Goat 1:3,000 Zymed
elF4E 25 Rabbit Mouse 1:500 BDBiosciences,San Jose,CA
Goat 1:3,000 Zymed
AcidicCKs
40–56 Human Mouse 1:250 (AE1) Serotec,Oxford,U.K.
Goat 1:3,000 Zymed
Stat5a 95 Mouse, N-terminus Mouse 1:500 TransductionLaboratories,Lexington,KY
Goat 1:3,000 Zymed
Stat5 (p) 97 Mouse, around Tyr694
Mouse 1:500 Cell SignalingTechnology
Goat 1:3,000 Zymed
Stat3 (p) 97 Mouse, around Tyr705
Rabbit 1:1,000 Cell SignalingTechnology
Goat 1:3,000 Zymed
ER� 66 Rat, last 15 aa Rabbit 1:1,000 UpstateBiotechnology,LakePlacid, NY
Goat 1:3,000 Zymed
Mucin 1 16–230 Mouse, N-terminus Goat 1:500 Santa CruzBiotechnology,CA
Rabbit 1:20,000 Jackson ImmunoresearchLaboratories, WestGlove, PA
608 IAVNILOVITCH ET AL.

sequences. The set of primers used for mouse S16 amplificationwere 5�-GTAGAGGTGAAAAGCCTAACGAG-3� and 5�-CTTC-TACACCATTGGGATGTC-3� originated from the S16 gene, andamplifying a 1,055 bp fragment of this gene. S16 determinationserved as a control for the efficiency of the reverse transcription-PCR (RT-PCR). In all cases, the DNA was blotted onto a mem-brane and hybridized with labeled DNA probe encompassingrespective fragments of the Stat5-based transgenic variant or theS16. A sample that was not reverse-transcribed served as a controlfor transgenic DNA contamination.
Histopathology and immunohistochemistryTumor biopsies were fixed with Bouin, dehydrated in a series of
50–100% ethanol, cleared in xylene and embedded in paraffin.Sections of 4 �m were processed for hematoxylin and eosin(H&E) staining. For smooth muscle actin (SMA) and proliferatingcell nuclear antigen (PCNA) analyses, Bouin-fixed paraffin-em-bedded sections were dewaxed, dehydrated through graded eth-anols, treated with 0.3% hydrogen peroxidase for 30 min, boiled in
0.1 M sodium citrate buffer for 10 min, blocked and reacted withbiotinylated anti-PCNA (Zymed, San Francisco, CA) or SMA(Dako, Glostrup, Denmark) antibodies for 60 min. Sections werethen washed in phosphate-buffered saline (PBS), incubated for 10min with streptavidinated horse radish peroxidase (HRP) andreacted with the diaminobenzidine (DAB) substrate (Zymed). Forthe detection of ER�, sections were reacted with the respectiveantibody (Table I). Signal was generated with EnVision reagent(Dako) and DAB substrate. Cell nuclei were counterstained withhematoxylin.
Immunoblot analysisProteins from intact tissue or tumors were extracted with ex-
traction buffer composed of 10 mM Tris-HCl, pH 7.4, 50 mMNaCl, 5 mM EDTA, 0.1% (w/v) BSA, 1% Triton, 50 mM NaF, 1mM Na3V04, 1 mM EGTA, 1 mM PMSF, 1 mM aprotinin andsubjected to SDS-PAGE analysis. Fractionated proteins weretransferred onto nitrocellulose filters and equal amounts of proteincontent/sample were confirmed by staining the blot with PonceauS (Sigma, St. Louis, MO). The expression levels of selectedproteins were determined according to the signals generated withtheir respective antibodies (Table I). All second antibodies werecoupled to HRP, and signals were generated with Super Signal,West Pico Chemiluminescent Substrate (Pierce, Rockford, IL)according to the manufacturer’s protocol.
KaryologyExponentially growing cultures of tumor cells were incubated
with 0.4 �g/ml colchicine for 2 hr. Cells were trypsinized andtreated with hypotonic medium for 10 min. Metaphase chromo-some spreads were prepared as described previously27 and Giemsastaining was performed for chromosomal analyses. Chromosomesof at least 20 metaphase cells were counted at cell passages 3–4.
Derivation of tumor cell lines and transplantation experimentsTumors were cut into small pieces and finely minced. Epithelial
cells were recovered by trypsin-collagenase digestion and differ-ential centrifugation as previously described.28 Cells were platedon a 100 mm plastic culture dish (Corning, Corning, NY) inDulbecco’s modified Eagle’s medium (DMEM)/F-12 (Gibco-BRL) supplemented with 10% fetal calf serum (FCS; Sigma),insulin (5 �g/ml; Sigma), hydrocortisone (1 �g/ml; Sigma), mouseepidermal growth hormone (EGF; 10 ng/ml; Sigma), gentamycin
FIGURE 1 – BLG/STAT5-based gene constructs and fusion proteins.(a) Hybrid genes: the following gene sequences were inserted betweenthe 2nd and 6th exons of the BLG gene. Line 1, BLG/ STAT5. Anative oStat5 molecule (homologue to the mouse Stat5a) linked to ashort epitope of the influenza virus. Line 2, BLG/STAT5ca. A re-stricted oStat5 gene lacking the coding region to its TAD was linkedto sequences encoding the kinase domain of Jak2 via sequences of theStat6 TAD. Line 3, BLG/STAT5�750. A restricted oSTAT5 genelacking the coding region to its TAD. Restriction enzymes used toclone the genes into the BLG expression cassette and to excise insertsfor microinjection into the nuclei of mouse embryos are indicated.BLG introns are numbered. (b) Fusion proteins. Line 1, STAT5. TheSH2 domain and Tyr 694, the phosphorylation of which is essential foractivation, are indicated. Line 2, STAT5ca. The TAD of Stat5 wasremoved and replaced by the TAD of Stat6, linked to the kinasedomain of Jak2. Line 3, STAT5�750. A C-terminally truncated Stat5composed of 750 amino acids, including Tyr 694.
FIGURE 2 – Detection of BLG/STAT5�750 transgene expression inthe mammary glands of transgenic mice by RT-PCR analysis. PA�
RNA (300 ng) was extracted from mammary glands of lactatingtransgenic mice of the indicated lines and reverse-transcribed. PCR (35cycles) was performed with primers located at the 3� end of the Stat5sequences and 5� end of the adjacent BLG sequences, amplifying theSTAT5�750/BLG junction region. 16S sequences were amplified (20cycles) with the appropriate primers using the same procedure. DNAwas blotted onto a filter and reacted with 32P-labeled probe, excisedfrom the BLG/STAT5�750 gene or 16S probe. An RNA sample,which was not reversed-transcribed, served as a negative control fortransgenic DNA contamination.
609STAT5 AND MAMMARY TUMORS
(50 �g/ml; Sigma), penicillin (100 U/ml; BioLabs, Jerusalem,Israel) and streptomycin (0.1 mg/ml; BioLabs). For transplanta-tions, cells were removed from the plates by trypsinization, col-lected by centrifugation and resuspended in PBS. Cell suspensions(0.3 � 106 cells/2 �l medium) were injected into the fat pad ofmammary gland 4 of 11–13 g virgin mice.
Statistical analysisProtein expression levels in tumor extracts were compared by
one-way analysis of variance using JUMP software (SAS Institute,Pacific Grove. CA). p � 0.05 was considered significant.
RESULTS
BLG/STAT5, BLG/STAT5ca and BLG/STAT5�750 fusion genesand their expression in mammary gland of transgenic mice
The regulatory sequences from the BLG milk-protein gene wereused to direct the expression of constructs encompassing 3 Stat5
variants to the mammary gland of transgenic mice (Fig. 1). Wepreviously showed that introduction of the wild-type BLG/STAT5and the constitutively active BLG/STAT5ca transgenes into thehost genome results in a developmentally dependent expression ofthese Stat5 variants in the mouse mammary gland.10 The wild-typeoStat5 molecule (which is homologous to the mouse Stat5a andtermed STAT5) was tagged with a hemagglutinin fragment fromthe human influenza virus (HA) sequence. This provides anepitope that allows the distinction from the endogenous protein.The constitutively active variant comprises the amino acid se-quences from position 1 to 750 of the Stat5 molecule, the trans-activation domain, TAD (position 677–857) of Stat6, and thekinase domain (positions 757–1129) of Jak2. This molecule isactivated constitutively in a receptor-independent fashion. TheBLG/STAT5�750 construct encodes a truncated Stat5 proteinlacking the TAD. This construct has previously been shown tofunction as a dominant negative variant in transfected cells.10 Italso is able to induce detectable phenotypes in transgenic mice. We
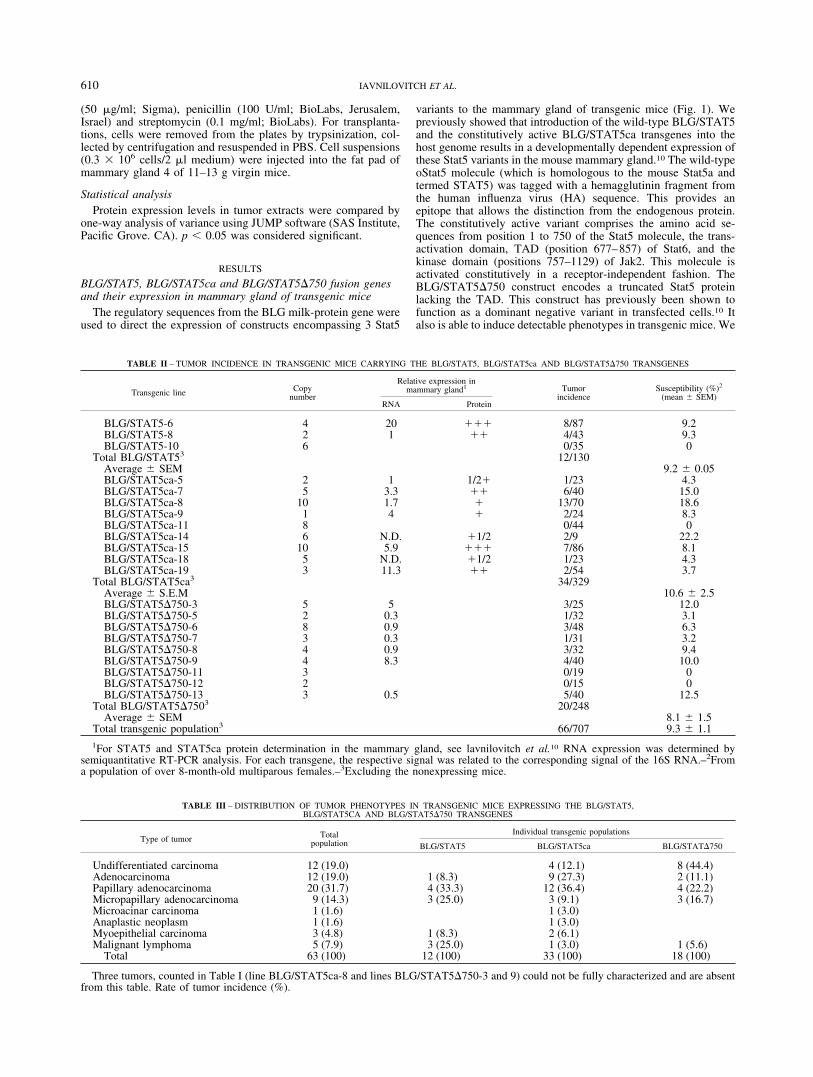
TABLE II – TUMOR INCIDENCE IN TRANSGENIC MICE CARRYING THE BLG/STAT5, BLG/STAT5ca AND BLG/STAT5�750 TRANSGENES
Transgenic line Copynumber
Relative expression inmammary gland1 Tumor
incidenceSusceptibility (%)2
(mean SEM)RNA Protein
BLG/STAT5-6 4 20 ��� 8/87 9.2BLG/STAT5-8 2 1 �� 4/43 9.3BLG/STAT5-10 6 0/35 0
Total BLG/STAT53 12/130Average SEM 9.2 0.05BLG/STAT5ca-5 2 1 1/2� 1/23 4.3BLG/STAT5ca-7 5 3.3 �� 6/40 15.0BLG/STAT5ca-8 10 1.7 � 13/70 18.6BLG/STAT5ca-9 1 4 � 2/24 8.3BLG/STAT5ca-11 8 0/44 0BLG/STAT5ca-14 6 N.D. �1/2 2/9 22.2BLG/STAT5ca-15 10 5.9 ��� 7/86 8.1BLG/STAT5ca-18 5 N.D. �1/2 1/23 4.3BLG/STAT5ca-19 3 11.3 �� 2/54 3.7
Total BLG/STAT5ca3 34/329Average S.E.M 10.6 2.5BLG/STAT5�750-3 5 5 3/25 12.0BLG/STAT5�750-5 2 0.3 1/32 3.1BLG/STAT5�750-6 8 0.9 3/48 6.3BLG/STAT5�750-7 3 0.3 1/31 3.2BLG/STAT5�750-8 4 0.9 3/32 9.4BLG/STAT5�750-9 4 8.3 4/40 10.0BLG/STAT5�750-11 3 0/19 0BLG/STAT5�750-12 2 0/15 0BLG/STAT5�750-13 3 0.5 5/40 12.5
Total BLG/STAT5�7503 20/248Average SEM 8.1 1.5
Total transgenic population3 66/707 9.3 1.11For STAT5 and STAT5ca protein determination in the mammary gland, see lavnilovitch et al.10 RNA expression was determined by
semiquantitative RT-PCR analysis. For each transgene, the respective signal was related to the corresponding signal of the 16S RNA.–2Froma population of over 8-month-old multiparous females.–3Excluding the nonexpressing mice.
TABLE III – DISTRIBUTION OF TUMOR PHENOTYPES IN TRANSGENIC MICE EXPRESSING THE BLG/STAT5,BLG/STAT5CA AND BLG/STAT5�750 TRANSGENES
Type of tumor Totalpopulation
Individual transgenic populations
BLG/STAT5 BLG/STAT5ca BLG/STAT�750
Undifferentiated carcinoma 12 (19.0) 4 (12.1) 8 (44.4)Adenocarcinoma 12 (19.0) 1 (8.3) 9 (27.3) 2 (11.1)Papillary adenocarcinoma 20 (31.7) 4 (33.3) 12 (36.4) 4 (22.2)Micropapillary adenocarcinoma 9 (14.3) 3 (25.0) 3 (9.1) 3 (16.7)Microacinar carcinoma 1 (1.6) 1 (3.0)Anaplastic neoplasm 1 (1.6) 1 (3.0)Myoepithelial carcinoma 3 (4.8) 1 (8.3) 2 (6.1)Malignant lymphoma 5 (7.9) 3 (25.0) 1 (3.0) 1 (5.6)
Total 63 (100) 12 (100) 33 (100) 18 (100)
Three tumors, counted in Table I (line BLG/STAT5ca-8 and lines BLG/STAT5�750-3 and 9) could not be fully characterized and are absentfrom this table. Rate of tumor incidence (%).
610 IAVNILOVITCH ET AL.
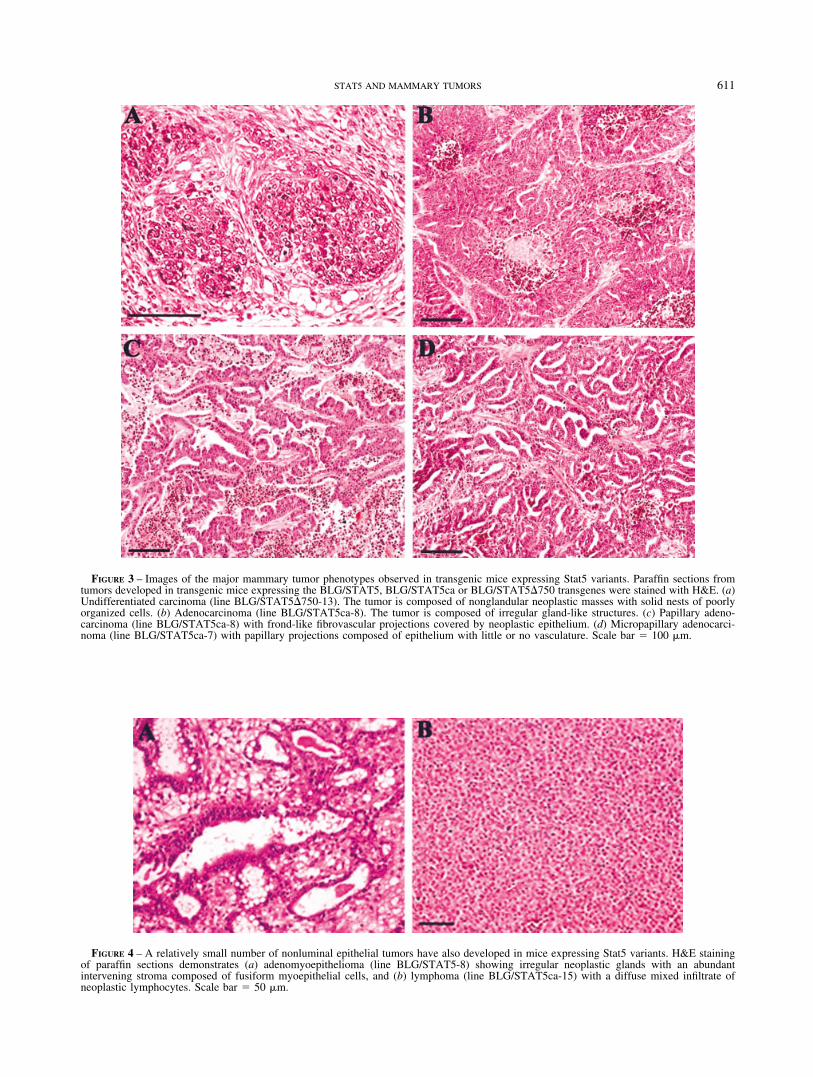
FIGURE 3 – Images of the major mammary tumor phenotypes observed in transgenic mice expressing Stat5 variants. Paraffin sections fromtumors developed in transgenic mice expressing the BLG/STAT5, BLG/STAT5ca or BLG/STAT5�750 transgenes were stained with H&E. (a)Undifferentiated carcinoma (line BLG/STAT5�750-13). The tumor is composed of nonglandular neoplastic masses with solid nests of poorlyorganized cells. (b) Adenocarcinoma (line BLG/STAT5ca-8). The tumor is composed of irregular gland-like structures. (c) Papillary adeno-carcinoma (line BLG/STAT5ca-8) with frond-like fibrovascular projections covered by neoplastic epithelium. (d) Micropapillary adenocarci-noma (line BLG/STAT5ca-7) with papillary projections composed of epithelium with little or no vasculature. Scale bar 100 �m.
FIGURE 4 – A relatively small number of nonluminal epithelial tumors have also developed in mice expressing Stat5 variants. H&E stainingof paraffin sections demonstrates (a) adenomyoepithelioma (line BLG/STAT5-8) showing irregular neoplastic glands with an abundantintervening stroma composed of fusiform myoepithelial cells, and (b) lymphoma (line BLG/STAT5ca-15) with a diffuse mixed infiltrate ofneoplastic lymphocytes. Scale bar 50 �m.
611STAT5 AND MAMMARY TUMORS

observed a delay in alveolar development in transgenic mice priorto lactation and an enhancement of apoptosis at early stages ofinvolution. When BLG/STAT5�750 mice were crossed with micecarrying a BLG/luciferase transgene, luciferase expression wasmuch lower than in mice containing only the BLG/luciferasetransgene (data not shown). To distinguish the in vivo-expressedSTAT5�750 from the wild-type, mammary RNA samples wereanalyzed by RT-PCR. Sequences comprising the junction betweenthe Stat5 gene and the BLG 3� regulatory region were amplified.The detection of STAT5�750 expression in mammary glands oftransgenic mice is shown in Figure 2. Transgenic mice of linesBLG/STAT5�750-11 (Fig. 2) and -12 (not shown) carry the trans-gene but do not express the respective RNA.
Mammary tumors develop in mice expressing 3 transgenic Stat5variants: wild-type, constitutively active and carboxyl-terminallytruncated Stat5
Constitutive activity of Stat3 and Stat5 proteins is frequently asso-ciated with cell transformation. In contrast, the dominant negativevariant of Stat3 has been shown to act as a tumor suppressor.29
Previously, we have suggested that Stat5 transgene expression mightbe involved in mammary tumors.10 We extended these studies andcompared the roles of wild-type, constitutively activated and TAD-truncated Stat5 on mammary tumorigenesis in transgenic mice.
Two mouse lines express the wild-type Stat5 (BLG/STAT5) asa transgene in addition to the endogenous gene (Table II). About9% of the over 8-month-old multiparous mice developed mam-mary tumors. Tumors also developed in all 8 lines of transgenicmice expressing the BLG/STAT5ca transgene with frequenciesranging between 3.7% and 22%. Unexpectedly, 7 lines of miceoverexpressing the truncated form of Stat5, BLG/STAT5�750,developed mammary tumors, with frequencies of 3.1–12.5%.
The locations and latency periods for tumor development wereevaluated in the total population of tumor-bearing mice. Singletumors were identified in 92% of the animals in the thoracic(47.5%) or inguinal (52.5%) glands of multiparous transgenicfemales aged 8–12 months. Mice that have incorporated the trans-genes but fail to express them served as controls. No tumors wereobserved in these animals. Also, no spontaneous palpable mam-mary tumors were detected in the first year of life in a populationof at least 46 transgenic mice expressing other transgenes, mainlyBLG/luciferase, which are maintained at our disposal,30 or in atleast 40 wild-type FVB/N females aged 8–12 months. The absenceof tumor incidence in the wild-type FVB/N cohort at this age is inagreement with previous reports.31,32
A high variability in transgene copy number was observedamong the transgenic mice. The impact of copy number on tumordevelopment was examined for individual sets of mice, as well asfor the complete transgenic colony (Table II). Only 2 lines ex-pressed the BLG/STAT5 transgene. Thus, the correlation betweentumorigenesis and number of integrated transgenic copies, whichintegrated in a tandem repeats, was individually examined only formice expressing the BLG/STAT5ca and BLG/STAT5�750 trans-genes. In both sets of transgenic mice, r-values were found insig-nificant (r 0.3, n 7 for BLG/STAT5ca mice, and r 0.17, n 7 for BLG/STAT5�750 mice). Similarly, an insignificant r-valuewas determined for the correlation between transgenic copy num-ber and tumor incidence in the total population of transgenic micecarrying the Stat5 variants (r 0.45, n 16).
The extent of transgenic RNA expression was also not pertinent totumorigenesis. r-values calculated for the correlation between expres-sion of transgenic STAT5ca or STAT5�750 RNA and tumor inci-dence were found to be insignificant for both transgenes (r 0.36,n 7, and r 0.47, n 7, respectively). Analysis of the totaltransgenic population was not applicable due to the different primersused to detect the Stat5 variants. Note that the truncated STAT5�750protein could not be clearly distinguished from the endogenous Stat5.The authenticity of the semiquantitation of its RNA relies on the
positive correlation observed between levels of STAT5 and STAT5caRNA and their corresponding proteins.10
Distinct pathologies characterize tumors derived from miceexpressing 3 transgenic variants of Stat5
A comprehensive analysis was performed on 63 palpable tumorswith an average diameter of 1.0 cm with respect to their pathologicfeatures and possible correlations with a specific Stat5 variant(Table III). H&E staining of paraffin sections revealed 4 mainphenotypes: undifferentiated carcinoma, adenocarcinoma, papil-lary adenocarcinoma and micropapillary adenocarcinoma (TableIII, Fig. 3). Tumors of mammary epithelial origin, encompassing apoorly or undifferentiated nonglandular phenotype, were termedundifferentiated carcinomas. This tumor phenotype was relativelycommon, representing over 44% of the neoplasms observed inmice expressing the BLG/STAT5�750 transgene, which encodesthe truncated Stat5 (Table III). The carcinomas were distinguishedby a typical structure of sheets of neoplastic mass with solid nestsof poorly organized and dyscohesive cells (Fig. 3a). Higher mag-nification revealed cells with large and pleomorphic nuclei, prom-inent nucleoli, a vesicular chromatin pattern, frequent mitoticfigures and relatively abundant pale cytoplasm.
Tumors with glandular differentiation are termed adenocarcino-mas. This type of tumor was more frequent in mice expressing theconstitutively active BLG/STAT5ca transgene (27%) than in miceexpressing the native or the truncated variants of Stat5 as trans-genes (Table III). The tumor mass is composed of cords and nestsof cells that form irregular tubules and glands, separated by a denseand frequently hyalinized stroma (Fig. 3b). Pleomorphic nucleiwith a vesicular chromatin pattern and scanty, slightly amphophiliccytoplasm were observed.
Two types of better-differentiated papillary adenocarcinomasdeveloped at high frequency in mice expressing the BLG/STAT5and BLG/STAT5ca transgenes (Table III, Fig. 3c and d). The term“papillary” was used here to describe adenocarcinomas in whichthe microscopic pattern is composed predominantly of fibrovas-cular fronds covered by epithelial cells.33 Typically, the papillaryand micropapillary tumor phenotypes were moderately firm to thetouch, with no evidence of hemorrhaging. However, 67% of thepapillary and 75% of the micropapillary tumors were partiallynecrotic. In contrast, 23% and 37% of adenocarcinomas and car-cinomas were necrotic.
More than 36% of the tumors in BLG/STAT5ca mice weremicropapillary adenocarcinomas. They are composed of large nod-ules with a glandular pattern, separated by a dense connectivetissue. The glands are lined by small epithelial papillae with analmost complete absence of fibrovascularization. This vascularepithelial pattern distinguishes the micropapillary phenotype fromthe more fibrovascular structure found in the papillary counterpart.In both, the columnar neoplastic epithelial cells lining the gland-like spaces tend to be tightly packed and lack polarity. Large,pleomorphic and hyperchromatic nuclei with an open chromatinpattern were unevenly distributed in relation to the basementmembrane. A high mitotic rate was observed.
A single tumor with a predominantly microacinar phenotypewas also observed in a BLG/STAT5ca transgenic mice. This tumorhad small back-to-back nests of cells surrounding a small centrallumen. A tumor with an anaplastic population of dyscohesive cellshaving large pleomorphic and hyperchromatic nuclei was alsonoted (Table III).
Fewer tumors developed from nonluminal epithelial cells (TableIII). Myoepithelial cells are widely distributed in the mammarygland and are part of the normal structure of the lobules and ducts.Mammary neoplasms composed of myoepithelial cells are rare inhumans.33 In transgenic mice expressing the variants of Stat5, onlyabout 5% of the tumors were characterized by overgrowth ofmyoepithelial cells and defined myoepitheliomas (Fig. 4a).
Lymphomas represented about 8% of the tumors. Here, themammary lymph node was enlarged, effaced by an infiltrate of
612 IAVNILOVITCH ET AL.
neoplastic lymphocytes. The lymphoid cells vary in size and shape(Fig. 4b) and contain large pleomorphic nuclei with vesicularchromatin, abundant mitotic figures and a scanty pale cytoplasm.
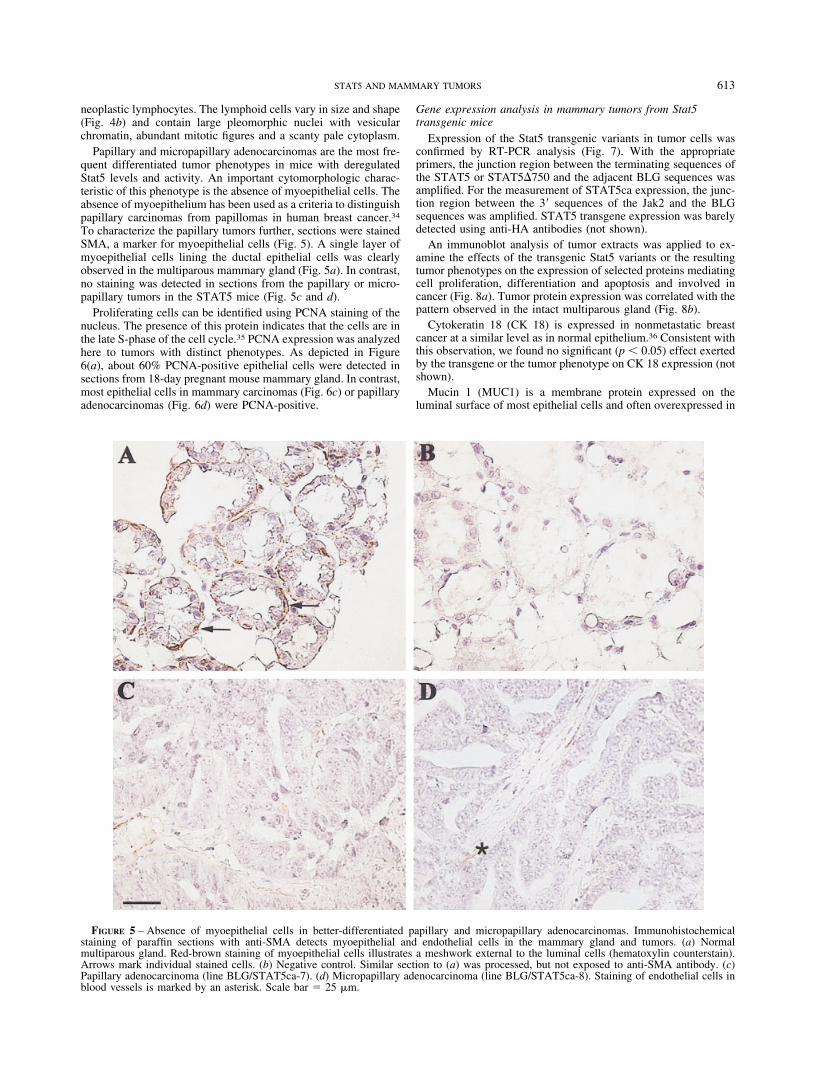
Papillary and micropapillary adenocarcinomas are the most fre-quent differentiated tumor phenotypes in mice with deregulatedStat5 levels and activity. An important cytomorphologic charac-teristic of this phenotype is the absence of myoepithelial cells. Theabsence of myoepithelium has been used as a criteria to distinguishpapillary carcinomas from papillomas in human breast cancer.34
To characterize the papillary tumors further, sections were stainedSMA, a marker for myoepithelial cells (Fig. 5). A single layer ofmyoepithelial cells lining the ductal epithelial cells was clearlyobserved in the multiparous mammary gland (Fig. 5a). In contrast,no staining was detected in sections from the papillary or micro-papillary tumors in the STAT5 mice (Fig. 5c and d).
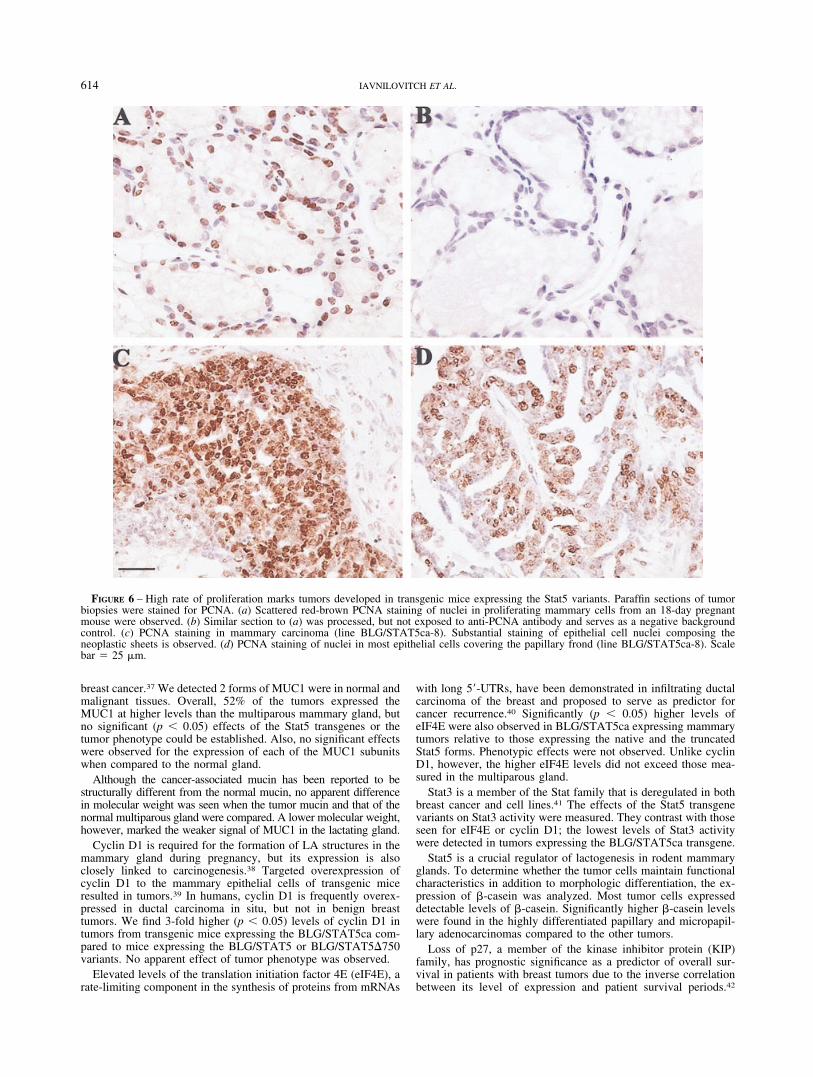
Proliferating cells can be identified using PCNA staining of thenucleus. The presence of this protein indicates that the cells are inthe late S-phase of the cell cycle.35 PCNA expression was analyzedhere to tumors with distinct phenotypes. As depicted in Figure6(a), about 60% PCNA-positive epithelial cells were detected insections from 18-day pregnant mouse mammary gland. In contrast,most epithelial cells in mammary carcinomas (Fig. 6c) or papillaryadenocarcinomas (Fig. 6d) were PCNA-positive.
Gene expression analysis in mammary tumors from Stat5transgenic mice
Expression of the Stat5 transgenic variants in tumor cells wasconfirmed by RT-PCR analysis (Fig. 7). With the appropriateprimers, the junction region between the terminating sequences ofthe STAT5 or STAT5�750 and the adjacent BLG sequences wasamplified. For the measurement of STAT5ca expression, the junc-tion region between the 3� sequences of the Jak2 and the BLGsequences was amplified. STAT5 transgene expression was barelydetected using anti-HA antibodies (not shown).
An immunoblot analysis of tumor extracts was applied to ex-amine the effects of the transgenic Stat5 variants or the resultingtumor phenotypes on the expression of selected proteins mediatingcell proliferation, differentiation and apoptosis and involved incancer (Fig. 8a). Tumor protein expression was correlated with thepattern observed in the intact multiparous gland (Fig. 8b).
Cytokeratin 18 (CK 18) is expressed in nonmetastatic breastcancer at a similar level as in normal epithelium.36 Consistent withthis observation, we found no significant (p � 0.05) effect exertedby the transgene or the tumor phenotype on CK 18 expression (notshown).
Mucin 1 (MUC1) is a membrane protein expressed on theluminal surface of most epithelial cells and often overexpressed in
FIGURE 5 – Absence of myoepithelial cells in better-differentiated papillary and micropapillary adenocarcinomas. Immunohistochemicalstaining of paraffin sections with anti-SMA detects myoepithelial and endothelial cells in the mammary gland and tumors. (a) Normalmultiparous gland. Red-brown staining of myoepithelial cells illustrates a meshwork external to the luminal cells (hematoxylin counterstain).Arrows mark individual stained cells. (b) Negative control. Similar section to (a) was processed, but not exposed to anti-SMA antibody. (c)Papillary adenocarcinoma (line BLG/STAT5ca-7). (d) Micropapillary adenocarcinoma (line BLG/STAT5ca-8). Staining of endothelial cells inblood vessels is marked by an asterisk. Scale bar 25 �m.
613STAT5 AND MAMMARY TUMORS
breast cancer.37 We detected 2 forms of MUC1 were in normal andmalignant tissues. Overall, 52% of the tumors expressed theMUC1 at higher levels than the multiparous mammary gland, butno significant (p � 0.05) effects of the Stat5 transgenes or thetumor phenotype could be established. Also, no significant effectswere observed for the expression of each of the MUC1 subunitswhen compared to the normal gland.
Although the cancer-associated mucin has been reported to bestructurally different from the normal mucin, no apparent differencein molecular weight was seen when the tumor mucin and that of thenormal multiparous gland were compared. A lower molecular weight,however, marked the weaker signal of MUC1 in the lactating gland.
Cyclin D1 is required for the formation of LA structures in themammary gland during pregnancy, but its expression is alsoclosely linked to carcinogenesis.38 Targeted overexpression ofcyclin D1 to the mammary epithelial cells of transgenic miceresulted in tumors.39 In humans, cyclin D1 is frequently overex-pressed in ductal carcinoma in situ, but not in benign breasttumors. We find 3-fold higher (p � 0.05) levels of cyclin D1 intumors from transgenic mice expressing the BLG/STAT5ca com-pared to mice expressing the BLG/STAT5 or BLG/STAT5�750variants. No apparent effect of tumor phenotype was observed.
Elevated levels of the translation initiation factor 4E (eIF4E), arate-limiting component in the synthesis of proteins from mRNAs
with long 5�-UTRs, have been demonstrated in infiltrating ductalcarcinoma of the breast and proposed to serve as predictor forcancer recurrence.40 Significantly (p � 0.05) higher levels ofeIF4E were also observed in BLG/STAT5ca expressing mammarytumors relative to those expressing the native and the truncatedStat5 forms. Phenotypic effects were not observed. Unlike cyclinD1, however, the higher eIF4E levels did not exceed those mea-sured in the multiparous gland.
Stat3 is a member of the Stat family that is deregulated in bothbreast cancer and cell lines.41 The effects of the Stat5 transgenevariants on Stat3 activity were measured. They contrast with thoseseen for eIF4E or cyclin D1; the lowest levels of Stat3 activitywere detected in tumors expressing the BLG/STAT5ca transgene.
Stat5 is a crucial regulator of lactogenesis in rodent mammaryglands. To determine whether the tumor cells maintain functionalcharacteristics in addition to morphologic differentiation, the ex-pression of �-casein was analyzed. Most tumor cells expresseddetectable levels of �-casein. Significantly higher �-casein levelswere found in the highly differentiated papillary and micropapil-lary adenocarcinomas compared to the other tumors.
Loss of p27, a member of the kinase inhibitor protein (KIP)family, has prognostic significance as a predictor of overall sur-vival in patients with breast tumors due to the inverse correlationbetween its level of expression and patient survival periods.42
FIGURE 6 – High rate of proliferation marks tumors developed in transgenic mice expressing the Stat5 variants. Paraffin sections of tumorbiopsies were stained for PCNA. (a) Scattered red-brown PCNA staining of nuclei in proliferating mammary cells from an 18-day pregnantmouse were observed. (b) Similar section to (a) was processed, but not exposed to anti-PCNA antibody and serves as a negative backgroundcontrol. (c) PCNA staining in mammary carcinoma (line BLG/STAT5ca-8). Substantial staining of epithelial cell nuclei composing theneoplastic sheets is observed. (d) PCNA staining of nuclei in most epithelial cells covering the papillary frond (line BLG/STAT5ca-8). Scalebar 25 �m.
614 IAVNILOVITCH ET AL.

Significantly (p � 0.05) higher levels of p27 expression weredetected in the nondifferentiated carcinoma tumors compared todata pooled from all other types. An effect for individual trans-genic Stat5 variant could not be distinguished.
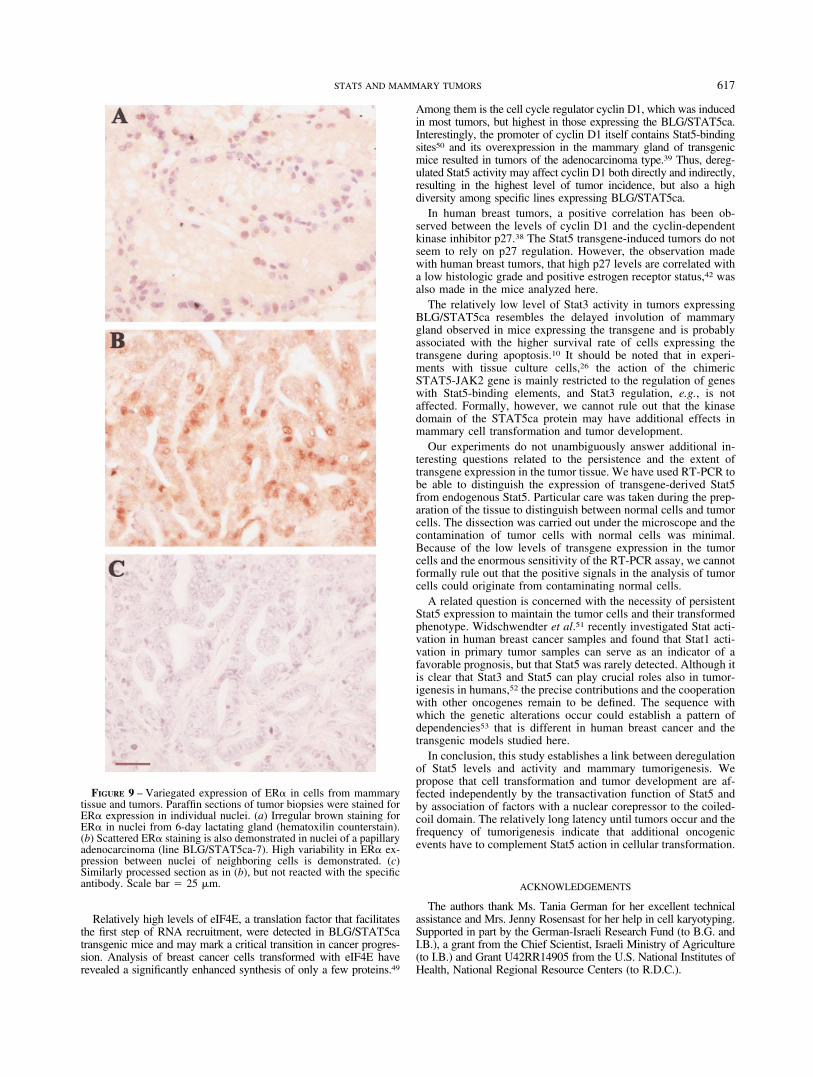
Estrogens are potent mitogens in the mammary gland. Endog-enous estrogens directly affect the growth of breast cancer cellsand inhibitors of estrogen receptor (ER) action, e.g., tamoxifen,have been successfully applied in breast cancer therapy.43 Most ofthe tumors that developed in mice carrying the Stat5 variantsexpressed detectable, albeit variable, levels of ER�. No significantdifference between the effects of the individual transgenes wasfound. This may be obscured by the high heterogeneity among thetumor cells in ER� expression (Fig. 9). Among the tumor pheno-types, the less-differentiated carcinomas maintained higher levelsof ER� compared to the rest of the tumors, but a significant effect(p � 0.05) was obtained only when compared to data pooled fromall other tumors or to lymphomas. Finally, it should be noted thatall these transgenic and phenotypic effects on tumor protein ex-pression did not involve significant alterations in the level oractivity of endogenous Stat5a.
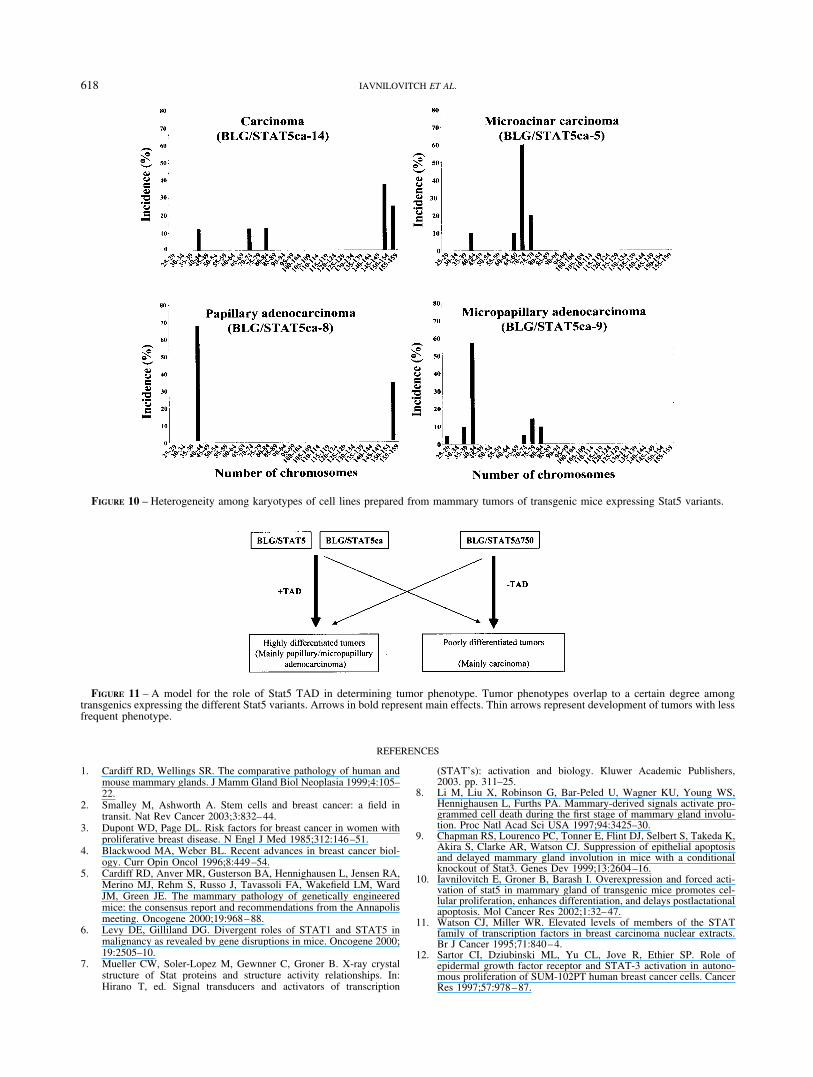
Derivation of tumor cell lines, karyotyping and transplantabilityTo investigate the karyotype of tumor cells, cell lines were
derived from primary tumors of mouse lines BLG/STAT5ca-14, 5and 8 and BLG/STAT5�750-9. These tumors were histologicallyclassified as carcinoma, microacinar carcinoma, papillary adeno-carcinoma and micropapillary adenocarcinoma, respectively.
Normal mouse cells contain 40 chromosomes. FACS analysisperformed on the respective cell lines suggested irregularities inthe number of chromosomes (not shown). The karyotypes weredetermined and aberrations were found in the number of chromo-somes (Fig. 10). An average of 30 chromosome spreads werecounted and aneuploidies were found in the cell lines originatingfrom tumors of the carcinoma and microacinar type. Nearly normalprofiles with about 30% duplication were observed for cells orig-inated from the papillary and micropapillary adenocarcinomas.This rate of duplication was shown for the mouse mammaryepithelial cell line CID-9.27
Cells from passages 3–4 from the line BLG/STAT5ca-5 weretransplanted into the fat pad of a nontransgenic recipient. Twelvedays after implantation, tumors of 20 mm in diameter developed in9 out of 10 implanted hosts. A further increase of about 150% intumor size was observed 7 days later.
DISCUSSION
The investigation into the activity of Stat5 throughout preg-nancy, lactation and involution suggests distinguishable roles forthis molecule at individual stages of mammary development, dif-ferentiation and involution. Early ablation of Stat5 function inconditional knockout animals has shown that epithelial cell differ-entiation and normal lobuloalveolar outgrowth are impeded. Stat5is required for milk protein expression at later stages in fullydifferentiated secretory alveolar cells and plays a role in theinhibition of apoptosis during the involution period. Since Stat5affects crucial growth and cell death processes, it is reasonable topresume that its inappropriate expression and activation might alsobe involved in tumorigenesis.
We have used variants of the Stat5 gene as transgenes to addressthis question and provide evidence for the potential of deregulatedStat5 levels and activity to initiate mammary cell transformation.Three variants of Stat5 were expressed in the mammary gland oftransgenic mice using regulatory sequences from the BLG milkprotein gene: wild-type Stat5, a truncated form of Stat5 that is ableto bind to specific DNA response sequences but does not activatethe transcription, and a constitutively activated Stat5 that com-prises a strong TAD and the Jak2 kinase region. Multiparousfemales from all transgenic lines expressing Stat5 variants devel-oped tumors at a frequency of up to 22% within 8–12 months. Thismight be complementary to the observation that loss of Stat5adelays mammary cancer progression.24
Models for breast cancer have been established, which make useof targeted oncogene expression in the mammary gland of genet-ically engineered mice. Some of the tumors exhibit unique phe-notypes characteristic for their causal genetic lesions. For example,the ErbB/ras signal-induced tumors can be distinguished from
FIGURE 7 – Detection of transgenic Stat5 variants in mammary tumors by RT-PCR analysis: a representative demonstration. PA� RNA wasextracted from tumors and reverse-transcribed. Specific sequences of the BLG/STAT5 and BLG/STAT5�750 genes were amplified by PCR withprimers located at the 3� end of the Stat5 gene and 5� end of the adjacent BLG regulatory sequences. BLG/STAT5ca-specific sequences wereamplified with primers located at the 3� end of the Jak2 gene and 5� end of the adjacent BLG regulatory sequences.
615STAT5 AND MAMMARY TUMORS

tumors originating from the deregulation of the Wnt pathway.5,44
To identify putative signature phenotypes for the 3 Stat5 variantsin transgenic mice, morphologic analyses were performed on 63tumors. In the 3 sets of mice, tumor phenotypes overlapped to acertain degree. However, mice expressing the native and constitu-tively activated Stat5 transgenes mainly developed tumors withhighly differentiated papillary/micropapillary phenotypes whencompared to the poorly differentiated carcinomas that occur pre-dominantly in STAT5�750 expressing tumors (Fig. 11).
The observation that transactivation of Stat5 target genes might notbe an absolute requirement for tumor formation is unexpected. Trun-cation of the carboxyl terminal results in a molecule that cannotinduce gene transcription due to the loss of its transactivation func-tion. This variant, however, might exert its function through itsassociation with nuclear corepressor proteins. The Stat5 coiled-coildomain, present in the carboxyl terminal truncation variant, binds toSMRT (silencing mediator for retinoic acid receptor and thyroidhormone receptor), a protein that recruits histone deacetylase activityto a promoter sequence and participates in gene silencing.45,46
Additional Stat5 interacting proteins might be of importance.Stat5 coiled-coil domain has been shown to bind to Nmi, a proteindiscovered as an N-MYC interactor, which augments IL-2-medi-ated transcription. High expression levels of Nmi were found inmyeloid leukemic cells, which also express high levels of c-MYC.47,48 Thus, it is possible that cell transformation and tumordevelopment involve balanced mechanism based on target geneactivation and suppression. The TAD activity is also associatedwith the development of distinct differentiated phenotypes, mainlypapillary and micropapillary adenocarcinomas.
As in human breast cancer, the mouse papillary and micropap-illary adenocarcinomas lack myoepithelial cells, a cytomorpho-logic parameter used to distinguish these malignant tumors frombenign papillomas. Interestingly, these highly differentiated tu-mors expressed high levels of �-casein compared to the other typesof tumors. Further insights into the mechanism involved in mam-mary cell transformation by Stat5 could come from analysis ofother cellular genes deregulated in tumors originating in miceexpressing the individual Stat5 variants.
FIGURE 8 – Genetic and phenotypic effects on the expression of proteins involved in cell proliferation, differentiation, apoptosis and oncogenesis intumors from transgenic mice harboring transgenic Stat5 variants. (a) Representative immunoblot analyses. Extracts from individual tumors or mammaryglands were analyzed by SDS-PAGE, blotted onto filters and reacted with antibodies for the indicated proteins. Ponceau S staining confirmed equallevels of protein/lane. The transgenic lines and tumor phenotypes are indicated. Mp, micropapillary adenocarcinoma; Ad, adenocarcinoma; Pap,papillary adenocarcinoma; L, lymphoma; Ca, carcinoma. (b) Statistical analysis for the effect of the transgene and the resulting tumor phenotype onprotein expression. Immunoblots were scanned and the intensity of the individual signals was measured and related to that averaged from glands of 3multiparous mice (6-day lactation for �-casein). Mean values were calculated for signals of tumors expressing the same transgene or from tumorsdisplaying a similar phenotype. One-way ANOVA test was performed for transgenic effect and phenotype effect. Values marked by different lettersare significant (p � 0.05). For the number and types of tumors, refer to Table II.
616 IAVNILOVITCH ET AL.
Relatively high levels of eIF4E, a translation factor that facilitatesthe first step of RNA recruitment, were detected in BLG/STAT5catransgenic mice and may mark a critical transition in cancer progres-sion. Analysis of breast cancer cells transformed with eIF4E haverevealed a significantly enhanced synthesis of only a few proteins.49
Among them is the cell cycle regulator cyclin D1, which was inducedin most tumors, but highest in those expressing the BLG/STAT5ca.Interestingly, the promoter of cyclin D1 itself contains Stat5-bindingsites50 and its overexpression in the mammary gland of transgenicmice resulted in tumors of the adenocarcinoma type.39 Thus, dereg-ulated Stat5 activity may affect cyclin D1 both directly and indirectly,resulting in the highest level of tumor incidence, but also a highdiversity among specific lines expressing BLG/STAT5ca.
In human breast tumors, a positive correlation has been ob-served between the levels of cyclin D1 and the cyclin-dependentkinase inhibitor p27.38 The Stat5 transgene-induced tumors do notseem to rely on p27 regulation. However, the observation madewith human breast tumors, that high p27 levels are correlated witha low histologic grade and positive estrogen receptor status,42 wasalso made in the mice analyzed here.
The relatively low level of Stat3 activity in tumors expressingBLG/STAT5ca resembles the delayed involution of mammarygland observed in mice expressing the transgene and is probablyassociated with the higher survival rate of cells expressing thetransgene during apoptosis.10 It should be noted that in experi-ments with tissue culture cells,26 the action of the chimericSTAT5-JAK2 gene is mainly restricted to the regulation of geneswith Stat5-binding elements, and Stat3 regulation, e.g., is notaffected. Formally, however, we cannot rule out that the kinasedomain of the STAT5ca protein may have additional effects inmammary cell transformation and tumor development.
Our experiments do not unambiguously answer additional in-teresting questions related to the persistence and the extent oftransgene expression in the tumor tissue. We have used RT-PCR tobe able to distinguish the expression of transgene-derived Stat5from endogenous Stat5. Particular care was taken during the prep-aration of the tissue to distinguish between normal cells and tumorcells. The dissection was carried out under the microscope and thecontamination of tumor cells with normal cells was minimal.Because of the low levels of transgene expression in the tumorcells and the enormous sensitivity of the RT-PCR assay, we cannotformally rule out that the positive signals in the analysis of tumorcells could originate from contaminating normal cells.
A related question is concerned with the necessity of persistentStat5 expression to maintain the tumor cells and their transformedphenotype. Widschwendter et al.51 recently investigated Stat acti-vation in human breast cancer samples and found that Stat1 acti-vation in primary tumor samples can serve as an indicator of afavorable prognosis, but that Stat5 was rarely detected. Although itis clear that Stat3 and Stat5 can play crucial roles also in tumor-igenesis in humans,52 the precise contributions and the cooperationwith other oncogenes remain to be defined. The sequence withwhich the genetic alterations occur could establish a pattern ofdependencies53 that is different in human breast cancer and thetransgenic models studied here.
In conclusion, this study establishes a link between deregulationof Stat5 levels and activity and mammary tumorigenesis. Wepropose that cell transformation and tumor development are af-fected independently by the transactivation function of Stat5 andby association of factors with a nuclear corepressor to the coiled-coil domain. The relatively long latency until tumors occur and thefrequency of tumorigenesis indicate that additional oncogenicevents have to complement Stat5 action in cellular transformation.
ACKNOWLEDGEMENTS
The authors thank Ms. Tania German for her excellent technicalassistance and Mrs. Jenny Rosensast for her help in cell karyotyping.Supported in part by the German-Israeli Research Fund (to B.G. andI.B.), a grant from the Chief Scientist, Israeli Ministry of Agriculture(to I.B.) and Grant U42RR14905 from the U.S. National Institutes ofHealth, National Regional Resource Centers (to R.D.C.).
FIGURE 9 – Variegated expression of ER� in cells from mammarytissue and tumors. Paraffin sections of tumor biopsies were stained forER� expression in individual nuclei. (a) Irregular brown staining forER� in nuclei from 6-day lactating gland (hematoxilin counterstain).(b) Scattered ER� staining is also demonstrated in nuclei of a papillaryadenocarcinoma (line BLG/STAT5ca-7). High variability in ER� ex-pression between nuclei of neighboring cells is demonstrated. (c)Similarly processed section as in (b), but not reacted with the specificantibody. Scale bar 25 �m.
617STAT5 AND MAMMARY TUMORS
REFERENCES
1. Cardiff RD, Wellings SR. The comparative pathology of human andmouse mammary glands. J Mamm Gland Biol Neoplasia 1999;4:105–22.
2. Smalley M, Ashworth A. Stem cells and breast cancer: a field intransit. Nat Rev Cancer 2003;3:832–44.
3. Dupont WD, Page DL. Risk factors for breast cancer in women withproliferative breast disease. N Engl J Med 1985;312:146–51.
4. Blackwood MA, Weber BL. Recent advances in breast cancer biol-ogy. Curr Opin Oncol 1996;8:449–54.
5. Cardiff RD, Anver MR, Gusterson BA, Hennighausen L, Jensen RA,Merino MJ, Rehm S, Russo J, Tavassoli FA, Wakefield LM, WardJM, Green JE. The mammary pathology of genetically engineeredmice: the consensus report and recommendations from the Annapolismeeting. Oncogene 2000;19:968–88.
6. Levy DE, Gilliland DG. Divergent roles of STAT1 and STAT5 inmalignancy as revealed by gene disruptions in mice. Oncogene 2000;19:2505–10.
7. Mueller CW, Soler-Lopez M, Gewnner C, Groner B. X-ray crystalstructure of Stat proteins and structure activity relationships. In:Hirano T, ed. Signal transducers and activators of transcription
(STAT’s): activation and biology. Kluwer Academic Publishers,2003. pp. 311–25.
8. Li M, Liu X, Robinson G, Bar-Peled U, Wagner KU, Young WS,Hennighausen L, Furths PA. Mammary-derived signals activate pro-grammed cell death during the first stage of mammary gland involu-tion. Proc Natl Acad Sci USA 1997;94:3425–30.
9. Chapman RS, Lourenco PC, Tonner E, Flint DJ, Selbert S, Takeda K,Akira S, Clarke AR, Watson CJ. Suppression of epithelial apoptosisand delayed mammary gland involution in mice with a conditionalknockout of Stat3. Genes Dev 1999;13:2604–16.
10. Iavnilovitch E, Groner B, Barash I. Overexpression and forced acti-vation of stat5 in mammary gland of transgenic mice promotes cel-lular proliferation, enhances differentiation, and delays postlactationalapoptosis. Mol Cancer Res 2002;1:32–47.
11. Watson CJ, Miller WR. Elevated levels of members of the STATfamily of transcription factors in breast carcinoma nuclear extracts.Br J Cancer 1995;71:840–4.
12. Sartor CI, Dziubinski ML, Yu CL, Jove R, Ethier SP. Role ofepidermal growth factor receptor and STAT-3 activation in autono-mous proliferation of SUM-102PT human breast cancer cells. CancerRes 1997;57:978–87.
FIGURE 10 – Heterogeneity among karyotypes of cell lines prepared from mammary tumors of transgenic mice expressing Stat5 variants.
FIGURE 11 – A model for the role of Stat5 TAD in determining tumor phenotype. Tumor phenotypes overlap to a certain degree amongtransgenics expressing the different Stat5 variants. Arrows in bold represent main effects. Thin arrows represent development of tumors with lessfrequent phenotype.
618 IAVNILOVITCH ET AL.

13. Garcia R, Yu CL, Hudnall A, Catlett R, Nelson KL, Smithgall T,Fujita DJ, Ethier SP, Jove R. Constitutive activation of Stat3 infibroblasts transformed by diverse oncoproteins and in breast carci-noma cells. Cell Growth Differ 1997;8:1267–76.
14. Garcia R, Bowman TL, Niu G, Yu H, Minton S, Muro-Cacho CA,Cox CE, Falcone R, Fairclough R, Parsons S, Laudano A, Gazit A,Levitski A, Kraker A Jove R. Constitutive activation of Stat3 by theSrc and JAK tyrosine kinases participates in growth regulation ofhuman breast carcinoma cells. Oncogene 2001;20:2499–513.
15. Miyoshi K, Cui Y, Riedlinger G, Robinson P, Lehoczky J, Zon L, OkaT, Dewar K, Hennighausen L. Structure of the mouse Stat 3/5 locus:evolution from Drosophila to zebrafish to mouse. Genomics 2001;71:150–5.
16. Liu X, Robinson GW, Wagner KU, Garrett L, Wynshaw-Boris A,Hennighausen L. Stat5a is mandatory for adult mammary gland de-velopment and lactogenesis. Genes Dev 1997;11:179–86.
17. Humphreys RC, Hennighausen L. Signal transducer and activator oftranscription 5a influences mammary epithelial cell survival and tu-morigenesis. Cell Growth Differ 1999;10:685–94.
18. Schmitt-Ney M, Doppler W, Ball RK, Groner B. Beta-casein genepromoter activity is regulated by the hormone-mediated relief oftranscriptional repression and a mammary-gland-specific nuclear fac-tor. Mol Cell Biol 1991;11:3745–55.
19. Pierre S, Jolivet G, Devinoy E, Theron MC, Malienou-N’Gassa R,Puissant C, Houdebine LM. A distal region enhances the prolactininduced promoter activity of the rabbit alpha s1-casein gene. Mol CellEndocrinol 1992;87:147–56.
20. Burdon TG, Maitland KA, Clark AJ, Wallace R, Watson CJ. Regu-lation of the sheep beta-lactoglobulin gene by lactogenic hormones ismediated by a transcription factor that binds an interferon-gammaactivation site-related element. Mol Endocrinol 1994;8:1528–36.
21. Liu X, Robinson GW, Gouilleux F, Groner B, Hennighausen L.Cloning and expression of Stat5 and an additional homologue (Stat5b)involved in prolactin signal transduction in mouse mammary tissue.Proc Natl Acad Sci USA 1995;92:8831–5.
22. Moriggl R, Gouilleux-Gruart V, Jahne R, Berchtold S, Gartmann C,Liu X, Hennighausen L, Sotiropoulos A, Groner B, Gouilleux F.Deletion of the carboxyl-terminal transactivation domain of MGF-Stat5 results in sustained DNA binding and a dominant negativephenotype. Mol Cell Biol 1996;16:5691–700.
23. Buettner R, Mora LB, Jove R. Activated STAT signaling in humantumors provides novel molecular targets for therapeutic intervention.Clin Cancer Res 2002;8:945–54.
24. Ren S, Cai HR, Li M, Furth PA. Loss of Stat5a delays mammarycancer progression in a mouse model. Oncogene 2002;21:4335–9.
25. Gouilleux F, Wakao H, Mundt M, Groner B. Prolactin induces phos-phorylation of Tyr694 of Stat5 (MGF), a prerequisite for DNA bind-ing and induction of transcription. EMBO J 1994;13:4361–9.
26. Berchtold S, Moriggl R, Gouilleux F, Silvennoinen O, Beisenherz C,Pfitzner E, Wissler M, Stocklin E, Groner B. Cytokine receptor-independent, constitutively active variants of STAT5. J Biol Chem1997;272:30237–43.
27. Danielson KG, Oborn CJ, Durban EM, Butel JS, Medina D. Epithelialmouse mammary cell line exhibiting normal morphogenesis in vivoand functional differentiation in vitro. Proc Natl Acad Sci USA1984;81:3756–60.
28. German T, Barash I. Characterization of an epithelial cell line frombovine mammary gland. In Vitro Cell Dev Biol Anim 2002;38:282–92.
29. Bromberg J, Darnell JE Jr. The role of STATs in transcriptionalcontrol and their impact on cellular function. Oncogene 2000;19:2468–73.
30. Reichenstein M, Gottlieb H, Damari GM, Iavnilovitch E, Barash I. Anew beta-lactoglobulin-based vector targets luciferase cDNA expres-sion to the mammary gland of transgenic mice. Transgenic Res2001;10:445–56.
31. Mahler JF, Stokes W, Mann PC, Takaoka M, Maronpot RR. Sponta-neous lesions in aging FVB/N mice. Toxicol Pathol 1996;24:710–6.
32. Landesman-Bollag E, Romieu-Mourez R, Song DH, Sonenshein GE,Cardiff RD, Seldin DC. Protein kinase CK2 in mammary glandtumorigenesis. Oncogene 2001;20:3247–57.
33. Rosen P. Rosen’s breast pathology, 2nd ed. Philadelphia: LippincottWilliams and Wilkins, 2001.
34. Tavassoli FA. Pathology of the breast, 2nd ed. New York: McGraw-Hill, 1999.
35. Saji S, Jensen EV, Nilsson S, Rylander T, Warner M, Gustafsson JA.Estrogen receptors alpha and beta in the rodent mammary gland. ProcNatl Acad Sci USA 2000;97:337–42.
36. Su L, Morgan PR, Lane EB. Expression of cytokeratin messengerRNA versus protein in the normal mammary gland and in breastcancer. Hum Pathol 1996;27:800–6.
37. Taylor-Papadimitriou J, Burchell JM, Plunkett T, Graham R, Correa I,Miles D, Smith M. MUC1 and the immunobiology of cancer. J MammGland Biol Neoplasia 2002;7:209–21.
38. Barnes DM, Gillett CE. Cyclin D1 in breast cancer. Breast Cancer ResTreat 1998;52:1–15.
39. Wang TC, Cardiff RD, Zukerberg L, Lees E, Arnold A, Schmidt EV.Mammary hyperplasia and carcinoma in MMTV-cyclin D1 transgenicmice. Nature 1994;369:669–71.
40. Li BD, Gruner JS, Abreo F, Johnson LW, Yu H, Nawas S, McDonaldJC, DeBenedetti A. Prospective study of eukaryotic initiation factor4E protein elevation and breast cancer outcome. Ann Surg 2002;235:732–8.
41. Bowman T, Garcia R, Turkson J, Jove R. STATs in oncogenesis.Oncogene 2000;19:2474–88.
42. Chiarle R, Pagano M, Inghirami G. The cyclin dependent kinaseinhibitor p27 and its prognostic role in breast cancer. Breast CancerRes 2001;3:91–4.
43. Poola I. Molecular assay to generate expression profile of eightestrogen receptor alpha isoform mRNA copy numbers in picogramamounts of total RNA from breast cancer tissues. Anal Biochem2003;314:217–26.
44. Rosner A, Miyoshi K, Landesman-Bollag E, Xu X, Seldin DC, MoserAR, MacLeod CL, Shyamala G, Gillgrass AE, Cardiff RD. Pathwaypathology: histological differences between ErbB/Ras and Wnt path-way transgenic mammary tumors. Am J Pathol 2002;161:1087–97.
45. Maurer AB, Wichmann C, Gross A, Kunkel H, Heinzel T, RuthardtM, Groner B, Grez M. The Stat5-RARalpha fusion protein repressestranscription and differentiation through interaction with a corepressorcomplex. Blood 2002;99:2647–52.
46. Nakajima H, Brindle PK, Handa M, Ihle JN. Functional interaction ofSTAT5 and nuclear receptor co-repressor SMRT: implications innegative regulation of STAT5-dependent transcription. EMBO J2001;20:6836–44.
47. Bao J, Zervos AS. Isolation and characterization of Nmi, a novelpartner of Myc proteins. Oncogene 1996;12:2171–6.
48. Zhu M, John S, Berg M, Leonard WJ. Functional association of Nmiwith Stat5 and Stat1 in IL-2- and IFNgamma-mediated signaling. Cell1999;96:121–30.
49. De Benedetti A, Harris AL. eIF4E expression in tumors: its possiblerole in progression of malignancies. Int J Biochem Cell Biol 1999;31:59–72.
50. Matsumura I, Kitamura T, Wakao H, Tanaka H, Hashimoto K, Al-banese C, Downward J, Pestell RG, Kanakura Y. Transcriptionalregulation of the cyclin D1 promoter by STAT5: its involvement incytokine-dependent growth of hematopoietic cells. EMBO J 1999;18:1367–77.
51. Widschwendter A, Tonko-Geymayer S, Welte T, Daxenbichler G,Marth C, Doppler W. Prognostic significance of signal transducer andactivator of transcription 1 activation in breast cancer. Clin CancerRes 2002;8:3065–74.
52. Yu H, Jove R. The STATs of cancer: new molecular targets come ofage. Nat Rev Cancer 2004;4:97–105.
53. Felsher DW. Cancer revoked: oncogenes as therapeutic targets. NatRev Cancer 2003;3:375–80.
54. Barash I, Faerman A, Puzis R, Peterson D, Shani M. Synthesis andsecretion of caseins by the mouse mammary gland: production andcharacterization of new polyclonal antibodies. Mol Cell Biochem1995;144:175–80.
619STAT5 AND MAMMARY TUMORS
Copyright © 2022 FDOKUMEN




















