
p27kip1 (Cyclin-Dependent Kinase Inhibitor 1B) Controls Ovarian Development by Suppressing Follicle...
14
p27 kip1 (Cyclin-Dependent Kinase Inhibitor 1B) Controls Ovarian Development by Suppressing Follicle Endowment and Activation and Promoting Follicle Atresia in Mice Singareddy Rajareddy, Pradeep Reddy, Chun Du, Lian Liu, Krishna Jagarlamudi, Wenli Tang, Yan Shen, Cyril Berthet, Stanford L. Peng, Philipp Kaldis, and Kui Liu Department of Medical Biochemistry and Biophysics (S.R., P.R., C.D., L.L., K.J., W.T., Y.S., K.L.), Umeå University, SE-901 87 Umeå, Sweden; Qilu Hospital (L.L.), Shandong University, Jinan, 250012 Shandong, China; National Cancer Institute (C.B., P.K.), Mouse Cancer Genetics Program, National Cancer Institute-Frederick, Frederick, Maryland 21702-1201; and Department of Inflammation (S.L.P.), Autoimmunity and Transplantation Research, Roche Palo Alto, LLC, Palo Alto, California 94304 In humans, the molecular mechanisms underlying ovarian follicle endowment and activation, which are closely related to the control of female repro- duction, occurrence of menopause, and related diseases such as premature ovarian failure, are poorly understood. In the current study, we provide several lines of genetic evidence that the cyclin- dependent kinase (Cdk) inhibitor 1B (commonly known as p27 kip1 or p27) controls ovarian develop- ment in mice by suppressing follicle endowment and activation, and by promoting follicle death. In p27-deficient (p27 / ) mice, postnatal follicle as- sembly was accelerated, and the number of en- dowed follicles was doubled as compared with p27 / mice. Moreover, in p27 / ovaries the pri- mordial follicle pool was prematurely activated once it was endowed, and at the same time the massive follicular death that occurs before sexual maturity was rescued by loss of p27. In early adult- hood, however, the overactivated follicular pool in p27 / ovaries was largely depleted, causing pre- mature ovarian failure. Furthermore, we have ex- tensively studied the molecular mechanisms un- derlying the above-mentioned phenotypes seen in p27 / ovaries and have found that p27 controls follicular development by several distinct mecha- nisms at different stages of development of the ovary. For example, p27 controls oocyte growth by suppressing the functions of Cdk2/Cdc2-cyclin A/E1 in oocytes that are arrested at the diplotene stage of meiosis I. This function of p27 is distinct from its well-known role as a suppressor of cell cycle progression. In addition, we have found that p27 activates the caspase-9-caspase-3-caspase- 7-poly (ADP-ribose) polymerase apoptotic cascade by inhibiting Cdk2/Cdc2-cyclin A/B1 kinase activi- ties in follicles, thereby inducing follicle atresia. Our results suggest that the p27 gene is important in determining mammalian ovarian development. This study therefore provides insight into ovary- borne genetic aberrations that cause defects in folliculogenesis and infertility in humans. (Molecu- lar Endocrinology 21: 2189–2202, 2007) F EMALE GERM CELLS in prenatal mouse ovaries are found in the form of clusters (syncytia) (see schematic diagram in Fig. 1A), which are connected by intercellular bridges as a result of incomplete cytoki- nesis (1). By the time mice are a few days old, individ- ual oocytes are enclosed in primordial follicles with a few flattened pregranulosa cells and become arrested at the diplotene stage of meiosis I (Fig. 1A). This pro- cess is accompanied by widespread apoptosis of oo- cytes, leading to the endowment of finite numbers of primordial follicles in the ovaries (Fig. 1A) (2). The molecular mechanisms that control ovarian primordial follicle assembly have not been thoroughly investi- gated. Studies in mice and sheep have revealed that during follicle formation, somatic cells invade the clus- ters of germ cells, and syncytial breakdown occurs just before primordial follicles are assembled (Fig. 1A) (3, 4), suggesting that pregranulosa cells have an ac- tive role in the formation of follicles (5). Once formed, the pool of primordial follicles serves as a source of developing follicles and oocytes (6–8). Follicular activation is characterized by rapid oocyte growth and proliferation of a few flattened pregranu- losa cells into cuboidal granulosa cells (9) (Fig. 1B) and First Published Online June 12, 2007 Abbreviations: BrdU, Bromodeoxyuridine; cdk, cyclin-de- pendent kinase; DKO, double knockout; E14.5, embryonic d 14.5; PARP, poly (ADP-ribose) polymerase; PD1, postnatal d 1; PI3K, phosphatidylinositol 3 kinase; POF, premature ovar- ian failure. Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving the endocrine community. 0888-8809/07/$15.00/0 Molecular Endocrinology 21(9):2189–2202 Printed in U.S.A. Copyright © 2007 by The Endocrine Society doi: 10.1210/me.2007-0172 2189
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of p27kip1 (Cyclin-Dependent Kinase Inhibitor 1B) Controls Ovarian Development by Suppressing Follicle...

p27kip1 (Cyclin-Dependent Kinase Inhibitor 1B)Controls Ovarian Development by SuppressingFollicle Endowment and Activation and PromotingFollicle Atresia in Mice
Singareddy Rajareddy, Pradeep Reddy, Chun Du, Lian Liu, Krishna Jagarlamudi, Wenli Tang,Yan Shen, Cyril Berthet, Stanford L. Peng, Philipp Kaldis, and Kui Liu
Department of Medical Biochemistry and Biophysics (S.R., P.R., C.D., L.L., K.J., W.T., Y.S., K.L.),Umeå University, SE-901 87 Umeå, Sweden; Qilu Hospital (L.L.), Shandong University, Jinan, 250012Shandong, China; National Cancer Institute (C.B., P.K.), Mouse Cancer Genetics Program, NationalCancer Institute-Frederick, Frederick, Maryland 21702-1201; and Department of Inflammation(S.L.P.), Autoimmunity and Transplantation Research, Roche Palo Alto, LLC,Palo Alto, California 94304
In humans, the molecular mechanisms underlyingovarian follicle endowment and activation, whichare closely related to the control of female repro-duction, occurrence of menopause, and relateddiseases such as premature ovarian failure, arepoorly understood. In the current study, we provideseveral lines of genetic evidence that the cyclin-dependent kinase (Cdk) inhibitor 1B (commonlyknown as p27kip1 or p27) controls ovarian develop-ment in mice by suppressing follicle endowmentand activation, and by promoting follicle death. Inp27-deficient (p27�/�) mice, postnatal follicle as-sembly was accelerated, and the number of en-dowed follicles was doubled as compared withp27�/� mice. Moreover, in p27�/� ovaries the pri-mordial follicle pool was prematurely activatedonce it was endowed, and at the same time themassive follicular death that occurs before sexualmaturity was rescued by loss of p27. In early adult-hood, however, the overactivated follicular pool inp27�/� ovaries was largely depleted, causing pre-mature ovarian failure. Furthermore, we have ex-
tensively studied the molecular mechanisms un-derlying the above-mentioned phenotypes seen inp27�/� ovaries and have found that p27 controlsfollicular development by several distinct mecha-nisms at different stages of development of theovary. For example, p27 controls oocyte growth bysuppressing the functions of Cdk2/Cdc2-cyclinA/E1 in oocytes that are arrested at the diplotenestage of meiosis I. This function of p27 is distinctfrom its well-known role as a suppressor of cellcycle progression. In addition, we have found thatp27 activates the caspase-9-caspase-3-caspase-7-poly (ADP-ribose) polymerase apoptotic cascadeby inhibiting Cdk2/Cdc2-cyclin A/B1 kinase activi-ties in follicles, thereby inducing follicle atresia.Our results suggest that the p27 gene is importantin determining mammalian ovarian development.This study therefore provides insight into ovary-borne genetic aberrations that cause defects infolliculogenesis and infertility in humans. (Molecu-lar Endocrinology 21: 2189–2202, 2007)
FEMALE GERM CELLS in prenatal mouse ovariesare found in the form of clusters (syncytia) (see
schematic diagram in Fig. 1A), which are connected byintercellular bridges as a result of incomplete cytoki-nesis (1). By the time mice are a few days old, individ-ual oocytes are enclosed in primordial follicles with afew flattened pregranulosa cells and become arrested
at the diplotene stage of meiosis I (Fig. 1A). This pro-cess is accompanied by widespread apoptosis of oo-cytes, leading to the endowment of finite numbers ofprimordial follicles in the ovaries (Fig. 1A) (2). Themolecular mechanisms that control ovarian primordialfollicle assembly have not been thoroughly investi-gated. Studies in mice and sheep have revealed thatduring follicle formation, somatic cells invade the clus-ters of germ cells, and syncytial breakdown occursjust before primordial follicles are assembled (Fig. 1A)(3, 4), suggesting that pregranulosa cells have an ac-tive role in the formation of follicles (5).
Once formed, the pool of primordial follicles servesas a source of developing follicles and oocytes (6–8).Follicular activation is characterized by rapid oocytegrowth and proliferation of a few flattened pregranu-losa cells into cuboidal granulosa cells (9) (Fig. 1B) and
First Published Online June 12, 2007Abbreviations: BrdU, Bromodeoxyuridine; cdk, cyclin-de-
pendent kinase; DKO, double knockout; E14.5, embryonic d14.5; PARP, poly (ADP-ribose) polymerase; PD1, postnatal d1; PI3K, phosphatidylinositol 3 kinase; POF, premature ovar-ian failure.
Molecular Endocrinology is published monthly by TheEndocrine Society (http://www.endo-society.org), theforemost professional society serving the endocrinecommunity.
0888-8809/07/$15.00/0 Molecular Endocrinology 21(9):2189–2202Printed in U.S.A. Copyright © 2007 by The Endocrine Society
doi: 10.1210/me.2007-0172
2189

is believed to be a continuous process that starts justafter follicle formation, long before the onset of pu-berty (6). It is thought that unknown intraovarian fac-tors stimulate some primordial follicles to initiategrowth at chosen times, whereas the rest of the folli-cles remain quiescent. On the other hand, it is alsopossible that follicular activation is caused by the re-lease from inhibitory mechanisms that maintain theprimordial follicles in a resting state (6).
In addition, during the initial wave of postnatal ovar-ian development in mice, large quantities of oocytes/follicles are depleted from the nongrowing folliclereservoir (6, 10, 11) (Fig. 1C). Compared with the nu-merous primordial follicles that have died and disap-peared, the number of primordial follicles recruitedinto the growing population is much lower (10, 11).This phenomenon is similar to the rapid loss of oo-cytes/follicles before puberty in human ovaries (6). Thereproductive life of the female is terminated when thepool of primordial follicles is exhausted (2, 6). How-ever, the mechanisms that govern the activation ofprimordial follicles and the rapid death of nongrowingfollicles have not been fully elucidated.
Previous studies from our and other researchgroups have demonstrated that a component of thephosphatidylinositol 3 kinase (PI3K) pathway, Foxo3a,serves as a suppressor of follicular activation andgrowth (12, 13). We have shown that when Foxo3a isoverexpressed in oocytes of primary follicles, oocytegrowth and follicular development are retarded (13).We presumed that one of the causes might be theretained expression of the cyclin-dependent kinase(Cdk) inhibitor 1B, commonly known as p27kip1 or p27,in the nuclei of oocytes (13). This has led us to hy-pothesize that p27 may suppress early follicular de-velopment. p27 itself is a negative regulator of cellcycle progression and a tumor suppressor (14). Inaddition, p27 has been shown to be essential for ovu-lation and luteinization in mice (15–17). However, therole and related mechanisms of p27 in controlling earlyfollicular development and oocyte growth have notbeen investigated. In the current study, we have usedwild-type and p27-deficient (p27�/�) mice and provideseveral lines of evidence to demonstrate that 1) p27suppresses follicle endowment/formation and activa-tion; 2) p27 induces follicle atresia that occurs before
BFollicle formation andendowment
Follicle activationA
C Rapid follicle loss prior to sexual maturity
Fig. 1. Schematic Diagram of Follicle Formation and Endowment, Primordial Follicle Activation, and Follicle Atresia during theInitial Wave of Postnatal Ovarian Development in Mice
A, Female germ cells in prenatal mouse ovaries are in the form of clusters (syncytia), which are surrounded by somatic cells.By the time mice are a few days old, somatic cells invade the clusters of germ cells, and syncytial breakdown occurs just beforeprimordial follicles are assembled, which is accompanied by widespread apoptosis of oocytes leading to the endowment of finitenumbers of primordial follicles in the ovaries. B, Once formed, the pool of primordial follicles serves as the source of developingfollicles. During primordial follicle activation in mammals, the rapid growth of oocytes is a major event. During the phase of oocytegrowth, granulosa cells proliferate from one layer of a few flattened pregranulosa cells in primordial follicles to three layers ofcuboidal granulosa cells by the time oocyte growth is almost complete. Most of the granulosa cell proliferation and differentiationthat is stimulated by FSH occurs after the oocyte has almost stopped growing, however, which is not shown in this schematicdiagram. C, During the initial wave of postnatal ovarian development in mice, large quantities of oocytes/follicles are removed fromthe nongrowing follicle reservoir, a phenomenon similar to the rapid loss of oocytes/follicles before puberty in human ovaries. Thediagram has been adapted from published diagrams of Bristol-Gould et al. (36) and Liu et al. (24).
2190 Mol Endocrinol, September 2007, 21(9):2189–2202 Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice

sexual maturity; and 3) the overactivated follicles inp27�/� ovaries are depleted in early adulthood, caus-ing premature ovarian failure (POF). The molecularmechanisms underlying these phenotypes in p27�/�
ovaries were also extensively investigated. We pro-pose that p27 is a key molecule in manipulation ofovarian development in mammals.
RESULTS
Dynamic Expression Patterns of p27 Protein inPerinatal Mouse Ovaries
Unexpectedly, we found that in perinatal mouseovaries that precede follicle formation, p27 is ex-pressed in somatic cells but not in oogonia/oocytes.At embryonic d 14.5 (E14.5) (Fig. 2A), E18.5 (datanot shown), and postnatal day (PD) 1 (Fig. 2B), p27expression was not detected in dividing oogonia oroocytes by immunostaining (Fig. 2, A and B, yellowarrows), but could be seen in somatic cells sur-rounding germ cells (Fig. 2, A and B; red arrows). Asshown in the inset of Fig. 2B (white arrow), germcells were identified by staining of germ cell nuclearantigen 1 (GCNA1). At PD4, when the primordial(type 2) follicles had already been formed, p27 ex-pression was initiated in the oocyte nuclei of primor-dial follicles (Fig. 2C, yellow arrow) and growingtransient (type 3a) follicles (Fig. 2C, yellow arrow-
head, and inset). Expression of p27 was also ob-served in pregranulosa cells (Fig. 2C). In growingprimary (type 3b) follicles (Fig. 2C, white arrow) andsecondary (type 4) follicles (Fig. 2D, white arrows),p27 was found to be expressed in both oocyte nu-clei and in granulosa cells. In further developedfollicles from 23-d-old mice, where oocytes are par-tially grown, however, p27 was no longer found tobe expressed in oocyte nuclei (Fig. 2D, yellow arrow)but was still expressed in granulosa cells (Fig. 2D,white arrowhead). The specificity of p27 immuno-staining was verified on ovarian sections fromp27�/� mice (15), where only minimal backgroundstaining was seen in granulosa cells (Fig. 2E, whitearrows), but not in oocytes (Fig. 2E, yellow arrows).
In addition, we have corroborated the p27 expres-sion pattern in mouse oocytes of primordial, primaryand further developed follicles as seen by immuno-staining in western blot. Using ovaries from 15- to17-d-old mice, which contain oocytes from primordialto preantral follicles, we isolated and separated oo-cytes into two groups that were larger or smaller than25 �m, respectively, with a cell-dispersing screen with25-�m opening (18). We found that p27 was highlyexpressed in oocytes that were smaller than 25 �m,but its expression was dramatically reduced in oo-cytes that were larger than 25 �m (Fig. 2F). This resultconfirms the data obtained by immunostaining (Fig. 2,C and D).
50 µm
β-actin
p27
> 25 µm < 25 µm
Oocytes
Western blot
A E14.5 B PD1
50 µm 25 µm
C PD4 D PD23
GCNA125 µm 25 m
p27 immunostaining
F
E PD8 p27-/-
25 µm
Fig. 2. Expression of p27 in Embryonic and Postnatal Mouse OvariesA–D, Ovaries from E14.5, 1-, 4-, and 23-d-old wild-type mice were isolated, fixed in 4% paraformaldehyde, dehydrated, and
embedded in paraffin. Sections of 5-�m thickness (E14.5 and PD1) or 8-�m thickness (PD4 and 23) were prepared andimmunostained for the presence of p27 as described in Materials and Methods. Signals appear as reddish to reddish-brown colorusing AEC as substrate. The ages of the mice are stated individually for each ovarian section. Germ cells were identified usingan antibody against GCNA1 (B, arrow in inset). E, Immunostaining of p27 in ovarian sections from p27�/� mice as a negativecontrol. F, Expression of p27 in oocytes that were larger or smaller than 25 �m. Oocytes were isolated from 17- to 20-d-old femaleC57/BL6J mice, sorted using a cell dispersing screen with 25-�m opening, and lysed as described in Materials and Methods.Levels of �-actin were used as an internal control. The experiments were repeated three times, and representative results areshown. For each experiment and each genotype in panel F, 15–20 mice were used.
Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice Mol Endocrinol, September 2007, 21(9):2189–2202 2191

Elevated Postnatal Ovarian Follicle Endowment inp27�/� Mice
To determine the functional roles of p27 that is ex-pressed by somatic cells surrounding oocytes in peri-natal ovaries during follicular formation and endow-ment, we used p27�/� mice. We first quantified theoocyte numbers at PD1 and follicle numbers at PD8 bycounting oocytes in serial sections of whole ovariesfrom p27�/� and p27�/� mice, to determine whetherthe lack of p27 in embryonic and postnatal ovariesinfluences the endowment of finite numbers of folliclesin mice. As shown in Table 1, at PD1 the GCNA1-positive oocytes in p27�/� ovaries were approximatelydouble compared with those in p27�/�ovaries. Thefollicles in PD8 p27�/� ovaries were also approxi-mately double compared with those in p27�/�ovaries(Table 1 and Fig. 3). These data indicate that morefollicles are endowed in p27�/� ovaries. When thenumber of follicles at PD8 was compared with thenumber of GCNA1-positive oocytes at PD1, a similarrate of oocyte survival (57%) was determined inp27�/� and p27�/� ovaries. Thus, we presume thatthe increased number of follicles in p27�/� mice maybe caused by elevated mitosis in female germ cells.The numbers of follicles in p27�/� ovaries counted inthis study are comparable to those in a previous reportby Pepling and Spradling (4).
Accelerated Assembly of Primordial Follicles inthe Absence of p27
To study in greater detail how p27 expressed by so-matic cells in perinatal ovaries may influence postnatalfollicle formation, we analyzed primordial follicle for-mation in p27�/� and p27�/�ovaries. As shown insupplemental Fig. S1, A–D (published as supplementaldata on The Endocrine Society’s Journals Online website at http://mend.endojournals.org), ovaries of p27�/�
and p27�/� mice at E17.5 and E18.5 displayed similarmorphologies. The only difference observed was thatat E18.5, more germ cells in the p27�/� ovary werelocated in the medullary part (supplemental Fig. S1,C—D; arrows in circles). At PD1, however, when most
of the oocytes in p27�/� mice were in clusters andlocated in the peripheral region of the ovary (Fig. 4A,arrows; inset), some oocytes in the p27�/�ovary hadalready been enclosed by somatic cells into primordialfollicles (Fig. 4B, arrows; inset). In PD2 p27�/� ovaries,the majority of oocytes were still in clusters (Fig. 4C,arrows) despite the fact that a limited number of pri-mordial follicles were identified. In PD2 p27�/� ova-ries, however, most of the oocytes were separatedand enclosed in primordial follicles (Fig. 4D, yellowarrows), which were shown by staining with an anti-VASA antibody that specifically stains the cytoplasmof oocytes, because anti-GCNA1 antibody does notstain the oocytes nuclei once oocytes are enclosed infollicles (19). Some primordial follicles in PD2 p27�/�
ovaries had even been activated into primary follicleswith enlarged oocytes, which were surrounded by in-creased numbers of cuboidal granulosa cells (Fig. 4D,inset; white arrow). These results indicate that in theabsence of p27, follicular assembly occurs earlier.
Premature Activation of the Primordial FolliclePool and Follicle Depletion in Early Adulthood inp27�/� Mice
As mentioned above, in PD2 p27�/� ovaries someactivated follicles already existed (Fig. 4D, inset).This phenomenon was further confirmed by the find-ing that at PD4, p27�/� ovaries were larger thanp27�/� ovaries due to an increase in primary folli-cles (supplemental Fig. S1, E and F; arrows). At PD8(Fig. 4, E and F) and at PD13 (supplemental Fig. S1,G and H), p27�/� ovaries appeared much largerthan p27�/� ovaries because of noticeably moreprimary, secondary, and even some further devel-oped follicles. By quantifying the follicles at variousdevelopmental stages at PD8 (Table 2), we foundthat the percentage of primordial follicles in p27�/�
mice was 46%, which was significantly lower (P �0.0001) than the proportion of primordial follicles inp27�/� ovaries (71%). Accordingly, the proportionof activated follicles in PD8 p27�/� ovaries (includ-ing primary, secondary, and more developed folli-cles) was 54%, which was significantly higher (P �
Table 1. Numbers of Oocytes or Follicles in 1-, 8-, 18-, 23-, and 35-Day-Old p27�/� and p27�/� Mice
Day No. of Oocytes perp27�/� Ovary
No. of Oocytes perp27�/� Ovary P Value
PD1 (GCNA1-positive) 3278 � 469a (n � 3) 7030 � 501 (n�3) 0.0055PD8 1863 � 444 (n � 6) 3984 � 627 (n�6) �0.0001PD18 1550 � 51 (n � 3) 3851 � 340 (n�3) �0.0001PD23 1450 � 221 (n � 3) 3814 � 130 (n�3) �0.0001PD35 706 � 37 (n � 4) 3979 � 348 (n�3) �0.0001Follicle survival rate (PD8/PD1) 56.8% 56.7%Follicle survival rate (PD35/PD8) 37.9% 99.9%
Ovaries from PD1, PD8, PD18, PD23, and PD35 p27�/� mice and their p27�/� littermates were isolated, and total numbers ofgerm cells or oocytes per ovary (mean � SD) were calculated as described in Materials and Methods. For each genotype and eachage, one ovary from each individual mouse was included, and three to six mice were used. n, Number of mice.a Mean � SD.
2192 Mol Endocrinol, September 2007, 21(9):2189–2202 Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice
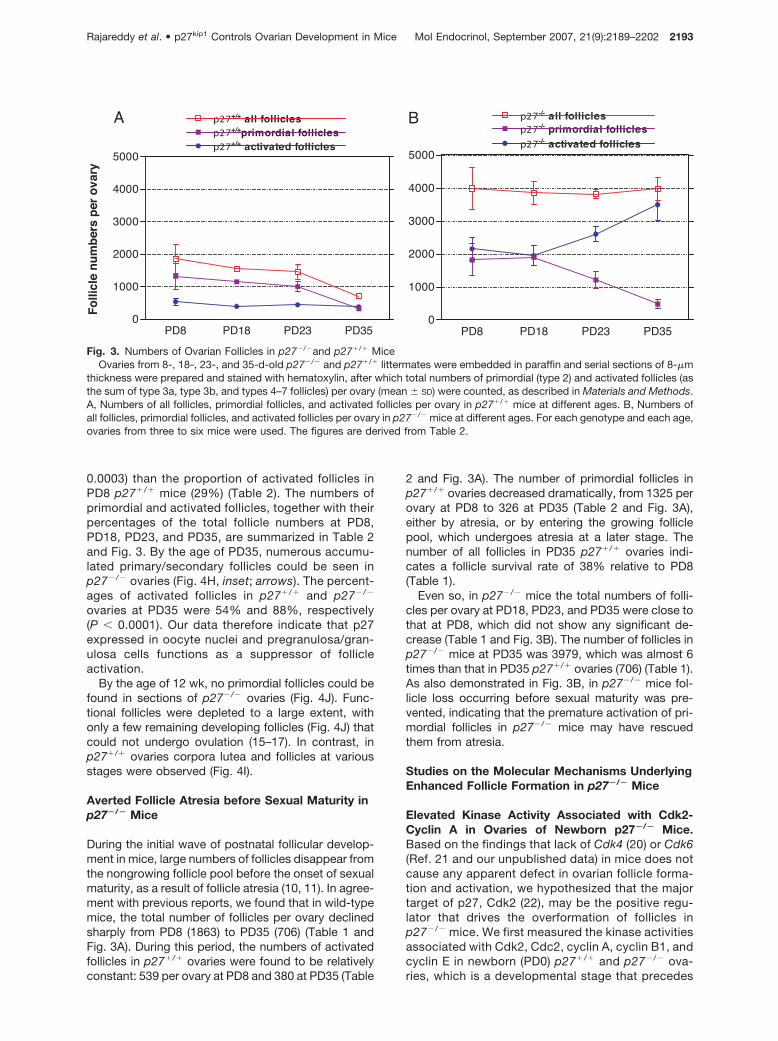
0.0003) than the proportion of activated follicles inPD8 p27�/� mice (29%) (Table 2). The numbers ofprimordial and activated follicles, together with theirpercentages of the total follicle numbers at PD8,PD18, PD23, and PD35, are summarized in Table 2and Fig. 3. By the age of PD35, numerous accumu-lated primary/secondary follicles could be seen inp27�/� ovaries (Fig. 4H, inset; arrows). The percent-ages of activated follicles in p27�/� and p27�/�
ovaries at PD35 were 54% and 88%, respectively(P � 0.0001). Our data therefore indicate that p27expressed in oocyte nuclei and pregranulosa/gran-ulosa cells functions as a suppressor of follicleactivation.
By the age of 12 wk, no primordial follicles could befound in sections of p27�/� ovaries (Fig. 4J). Func-tional follicles were depleted to a large extent, withonly a few remaining developing follicles (Fig. 4J) thatcould not undergo ovulation (15–17). In contrast, inp27�/� ovaries corpora lutea and follicles at variousstages were observed (Fig. 4I).
Averted Follicle Atresia before Sexual Maturity inp27�/� Mice
During the initial wave of postnatal follicular develop-ment in mice, large numbers of follicles disappear fromthe nongrowing follicle pool before the onset of sexualmaturity, as a result of follicle atresia (10, 11). In agree-ment with previous reports, we found that in wild-typemice, the total number of follicles per ovary declinedsharply from PD8 (1863) to PD35 (706) (Table 1 andFig. 3A). During this period, the numbers of activatedfollicles in p27�/� ovaries were found to be relativelyconstant: 539 per ovary at PD8 and 380 at PD35 (Table
2 and Fig. 3A). The number of primordial follicles inp27�/� ovaries decreased dramatically, from 1325 perovary at PD8 to 326 at PD35 (Table 2 and Fig. 3A),either by atresia, or by entering the growing folliclepool, which undergoes atresia at a later stage. Thenumber of all follicles in PD35 p27�/� ovaries indi-cates a follicle survival rate of 38% relative to PD8(Table 1).
Even so, in p27�/� mice the total numbers of folli-cles per ovary at PD18, PD23, and PD35 were close tothat at PD8, which did not show any significant de-crease (Table 1 and Fig. 3B). The number of follicles inp27�/� mice at PD35 was 3979, which was almost 6times than that in PD35 p27�/� ovaries (706) (Table 1).As also demonstrated in Fig. 3B, in p27�/� mice fol-licle loss occurring before sexual maturity was pre-vented, indicating that the premature activation of pri-mordial follicles in p27�/� mice may have rescuedthem from atresia.
Studies on the Molecular Mechanisms UnderlyingEnhanced Follicle Formation in p27�/� Mice
Elevated Kinase Activity Associated with Cdk2-Cyclin A in Ovaries of Newborn p27�/� Mice.Based on the findings that lack of Cdk4 (20) or Cdk6(Ref. 21 and our unpublished data) in mice does notcause any apparent defect in ovarian follicle forma-tion and activation, we hypothesized that the majortarget of p27, Cdk2 (22), may be the positive regu-lator that drives the overformation of follicles inp27�/� mice. We first measured the kinase activitiesassociated with Cdk2, Cdc2, cyclin A, cyclin B1, andcyclin E in newborn (PD0) p27�/� and p27�/� ova-ries, which is a developmental stage that precedes
Fo
llicl
en
um
ber
sp
er o
vary
A B
PD8 PD18 PD23 PD350
1000
2000
3000
4000
5000
p27-/- primordial folliclesp27-/- activated follicles
p27-/- all follicles
PD8 PD18 PD23 PD350
1000
2000
3000
4000
5000
p27+/+primordial folliclesp27+/+ activated follicles
p27+/+ all follicles
Fig. 3. Numbers of Ovarian Follicles in p27�/�and p27�/� MiceOvaries from 8-, 18-, 23-, and 35-d-old p27�/� and p27�/� littermates were embedded in paraffin and serial sections of 8-�m
thickness were prepared and stained with hematoxylin, after which total numbers of primordial (type 2) and activated follicles (asthe sum of type 3a, type 3b, and types 4–7 follicles) per ovary (mean � SD) were counted, as described in Materials and Methods.A, Numbers of all follicles, primordial follicles, and activated follicles per ovary in p27�/� mice at different ages. B, Numbers ofall follicles, primordial follicles, and activated follicles per ovary in p27�/� mice at different ages. For each genotype and each age,ovaries from three to six mice were used. The figures are derived from Table 2.
Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice Mol Endocrinol, September 2007, 21(9):2189–2202 2193

follicle formation. As shown in Fig. 5, we found thatkinase activities associated with Cdk2 and cyclin Awere elevated in newborn p27�/� ovaries relative tothose in PD0 p27�/� ovaries. Kinase activities as-sociated with Cdc2 (Fig. 5), cyclin B1, and cyclin E1(data not shown) were at similar levels in PD0p27�/� and p27�/� ovaries. These results suggestthat the increased kinase activity of the Cdk2-cyclinA complex may be one of the forces that drive theaccelerated follicle formation and endowment. Thecellular localization of the elevated Cdk2-cyclin Akinase activity remains to be determined.
Mechanistic Studies of the Premature Activationof Primordial Follicles in p27�/� Mice
Elevated Kinase Activity Associated with Cdk2-Cy-clin A/E1 in p27�/� Oocytes. Rapid oocyte growthis a major event that takes place during primordialfollicle activation (9). To compare the kinase profiles inp27�/� and p27�/� oocytes during follicle activationand early development, we measured kinase activitiesassociated with Cdk2, Cdc2, cyclin A, cyclin B1, andcyclin E1 in oocytes isolated from ovaries of 8- to12-d-old p27�/� and p27�/� mice. As shown in Fig.6A, kinase activities associated with Cdk2 and cyclin Awere found to be apparently elevated in p27�/� oo-cytes. Kinase activity associated with cyclin E1 wasmoderately elevated in p27�/� oocytes (Fig. 6A). Ki-nase activities associated with Cdc2 and cyclin B1,however, showed no apparent change (Fig. 6A).Oocyte-Specific Knockout of Cdk2 Did Not AlterFollicular Activation in Mice. To study the func-tional roles of oocyte Cdk2 during follicular activation,we have deleted Cdk2 from oocytes of primordial fol-licles by crossing Cdk2 loxP mice (our unpublisheddata) with the growth differentiation factor 9 (Gdf-9)promoter-mediated improved Cre (iCre) transgenicmice, the effectiveness of which has been well proven(23). Surprisingly, oocyte-specific deletion of Cdk2 didnot lead to any apparent difference in primordial folli-cle activation and ovarian development in mice (datanot shown). These results indicate that there may beoverlap in the function of oocyte Cdk2 during follicleactivation with the functions of other related kinases.Follicle Activation Is Suppressed in p27�/� Ovariesupon Inhibition of Both Cdk2 and Cdc2. To eval-uate the importance of Cdk2 and Cdc2 in the overac-tivation of primordial follicles in p27�/� ovaries, wecultured 2-d-old p27�/� ovaries (where primordial fol-licles had formed) with Roscovitine (20 �M), a sub-stance that specifically inhibits both Cdk2 and Cdc2.We found that Roscovitine could largely block theactivation of primordial follicles in cultured PD2p27�/� ovaries (Fig. 6B, arrows). Therefore, it is likelythat in oocytes of p27�/� primordial follicles, althoughCdc2 kinase activity is not elevated, kinase activitiesassociated with both Cdk2-cyclin A/E1 and Cdc2-cyclin A/E1 complexes are important for the acceler-ated follicular activation.p27 and Foxo3a Suppress Primordial Follicle Acti-vation Independently of Each Other. Recently, ac-tivation of the oocyte PI3K pathway, including theactivation of Akt and suppression of Akt substrateFoxo3a, has been suggested to be important for initi-ation and further development of follicles (13, 18, 24).Foxo3a is an Akt substrate that can be phosphorylatedand suppressed by Akt (25, 26). Previous reports haveindicated that loss of Foxo3a in mice leads to earlyactivation of primordial follicles (12), and overexpres-sion of Foxo3a in mouse oocytes facilitates the nuclearlocalization of p27 in oocytes, which is thought to bepartly responsible for the retarded oocyte and follicular
p27+/+
PD1
A B
PD2
E F
PD8
PD35
G
12-wk
I
CL
D
p27-/-
100 µm 100 µm
250 µm 250 µm
H
J
C
150 µm 150 µm
150 µm150 µm
250 µm250 µm
Fig. 4. Histological Analysis of Postnatal Ovaries fromp27�/� and p27�/� Littermates
Ovarian sections of 5-�m thickness (PD1) or 8-�m thick-ness (PD2, PD8, PD35, and 12-wk) were prepared andstained with anti-GCNA1 antibody (A–C) or anti-VASA anti-body (D) to identify oocytes, or stained with hematoxylin formorphological observation (E–J), as described in Materialsand Methods. All experiments were repeated at least fourtimes, and representative images of ovaries are shown. CL (I,arrows): corpora lutea.
2194 Mol Endocrinol, September 2007, 21(9):2189–2202 Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice

development in the transgenic mice (13). Thus, weendeavored to study whether p27 and Foxo3a func-tions in an up- or downstream cascade that sup-presses follicle activation.
We found that the expression of p27 and Foxo3aappeared to be normal in ovaries of 8- and 13-d-oldFoxo3a�/� (27) and p27�/� mice, respectively, as de-tected by Western blot and immunohistochemistry(data not shown). Moreover, in oocytes isolated from8- to 12-d-old mice where overactivation of primordialfollicles has been seen in both Foxo3a�/� and p27�/�
ovaries, the expression/phosphorylation levels of Aktand Foxo3a in p27�/� oocytes were similar comparedwith those in p27�/� oocytes (Fig. 6C); and the p27expression in Foxo3a�/� oocytes was also at a similarlevel as compared with Foxo3a�/� oocytes (Fig. 6D).
We then crossed p27�/� mice with Foxo3a�/� miceto generate double knockout (DKO) mice that lack
both p27 and Foxo3a, and compared the ovarian phe-notypes of p27�/� mice, Foxo3a�/� mice, and p27�/�
Foxo3a�/� DKO mice. By looking at PD13, which is atime when p27�/� ovaries still have some primordialfollicles (Fig. 6E, arrows), we found that in Foxo3a�/�
ovaries, in agreement with the previous report (12),there were more activated transient follicles, whichappeared to have somewhat enlarged oocytes sur-rounded by flattened pregranulosa cells (Fig. 6F, ar-rowheads) that distinguish them from primordial folli-cles (Fig. 6F, arrows). In p27�/� ovaries, however,transient follicles with enlarged oocytes but with flat-tened pregranulosa cells were rarely seen: the en-larged oocytes in activated follicles were always ac-companied by elevated numbers of cuboidalgranulosa cells (Fig. 6E, arrowheads). These resultsindicate that p27 deficiency, but not Foxo3a deficiencyin pregranulosa cells, triggers pregranulosa cell prolif-eration and differentiation during follicle activation. Inovaries of PD13 p27�/�Foxo3a�/� DKO mice, the ma-jority of the follicles were activated, with apparentlyenlarged oocytes that were surrounded by mostlycuboidal granulosa cells (Fig. 6G, arrowheads); typicalprimordial follicles were rarely seen (Fig. 6G).
By counting follicle numbers in serially sectionedovaries, we found that in PD13 Foxo3a�/� ovaries,primordial follicles represented approximately 30% ofall follicles (Fig. 6K), which was a significantly lowerrate than the primordial follicle rate in wild-type orp27�/� ovaries (Fig. 6K). Moreover, in PD13 DKO ova-ries the proportion of primordial follicles was less than10%, which was significantly lower than that in p27�/�
and Foxo3a�/� mice (Fig. 6K). These data indicate thatthe simultaneous loss of p27 and Foxo3a synergisti-cally accelerates follicle initiation. Thus, we proposethat p27 and Foxo3a suppress follicle activation inde-pendently of each other.
The hypothesis that Foxo3a and p27 may functionthrough distinct pathways to suppress follicle activa-tion and growth was also supported by the finding thatin Foxo3a�/� ovaries, the activated follicles (Fig. 6F,arrowheads) did not appear to grow as rapidly asthose in p27�/� ovaries: at PD23, the Foxo3a�/� ova-ries did not appear to be apparently enlarged (Fig. 6I)
p27+/+ p27-/-
H1
H1
IP Cyc A
IP Cdk2
p27 blot
PD0 kinase activity assay
H1IP Cdc2
Fig. 5. Studies on the Molecular Mechanisms of Follicle For-mation in p27�/� Mice
Kinase activities associated with Cdk2, cyclin A, and Cdc2in ovaries of newborn (PD0) p27�/� and p27�/� mice. Ovariesfrom PD0 p27�/� and p27�/� mice were collected and ho-mogenized, and ovarian lysates (150 �g) were used for im-munoprecipitation with indicated antibodies. Kinase activityassay was performed using histone H1 as substrate, as de-scribed in Materials and Methods. The experiments wererepeated three times, and representative images are shown.For each experiment and each genotype, 20–30 mice wereused. IP, Immunoprecipitation.
Table 2. Numbers of Primordial and Activated Follicles in 8-, 18-, 23-, and 35-Day-Old p27�/� and p27�/� Mice
No. of Follicles per p27�/� Ovary No. of Follicles per p27�/� Ovary
Primordial (%) Activated (%) Primordial (%) Activated (%)
PD8 1325 � 403a (71.1) 539 � 107 (28.9) 1825 � 486 (45.8) 2155 � 345 (54.1)PD18 1155 � 58 (74.5) 395 � 33 (25.5) 1897 � 47 (49.3) 1954 � 297 (50.7)PD23 1005 � 152 (69.3) 445 � 69 (30.7) 1214 � 249 (31.8) 2600 � 229 (68.2)PD35 326 � 46 (46.2) 380 � 39 (53.8) 484 � 141 (12.2) 3494 � 489 (87.8)
Mean numbers of total primordial and activated follicles per ovary in PD8, PD18, PD23, and PD35 ovaries of p27�� mice and theirp27�/� littermates were counted as described in Materials and Methods. Average percentages of primordial and activated folliclesat each age are also shown in parentheses. For each genotype and each age, one ovary from each individual mouse was included,and three to six mice were used, as indicated in Table 1.a Mean � SD.
Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice Mol Endocrinol, September 2007, 21(9):2189–2202 2195

p27+/+ p27-/-
H1IP Cdk2
H1IP Cdc2
H1IP Cyc B1
Kinase activities in oocytes (PD8-12)
Control
50 µm
50 µm
Roscovitine
3-day culture of PD2 p27-/- ovaries
H1IP Cyc A
A B
H1IP Cyc E1
β-actin
p-Foxo3a (T32)
p27
+/+
Foxo3a
C
p27
-/-
Akt
p-Akt (S473)
β-actin
p27
Foxo
3a+/
+
D
Foxo
3a-/-
Oocytes (PD 8-12)Oocytes
(PD 8-12)
PD13 PD23
K
E H
I
J
250 µm
250 µm50 µm50 µm
50 µm
p27-/-
Foxo3a-/-
Foxo3a-/-
p27-/-
a
b
c
d
Wild-type p27-/- Foxo3a-/- DKO0
20
40
60
80
100
Pri
mo
rdia
l fo
llic
les
(%)
G
50 µm50 µm 50 µm
F
50 µm 50 µm
250 µm
Fig. 6. Studies of Follicle Activation in p27�/� MiceA, Kinase activities associated with Cdk2, cyclin A, cyclin E1, Cdc2, and cyclin B1 in isolated oocytes of p27�/� and p27�/�
mice. Oocytes from ovaries of 8- to 12-d-old p27�/� and p27�/� mice were isolated and lysed, and 50 �g of oocyte lysates wereused for immunoprecipitation with indicated antibodies. Kinase activity assay was performed using histone H1 as substrate, asdescribed in Materials and Methods. The experiments were repeated three times, and representative images are shown. For eachexperiment, 10–15 p27�/� mice and 20–25 p27�/� mice were used. B, Suppression of follicle activation by Roscovitine in culturedPD2 ovaries of p27�/� mice. Ovaries of PD2 p27�/� mice (where primordial follicles have formed) were cultured without or withtreatment of the Cdk2/Cdc2 inhibitor Roscovitine (20 �M), as described in Materials and Methods. After the 3-d culture period,
2196 Mol Endocrinol, September 2007, 21(9):2189–2202 Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice

as the p27�/�ovaries (Fig. 6H). The PD23 DKO ova-ries, however, were found to have numerous develop-ing preantral follicles and were larger than bothFoxo3a�/� and p27�/� ovaries (Fig. 6, H–J), confirm-ing the notion that concurrent loss of Foxo3a and p27synergistically accelerates follicle initiation andgrowth. At PD35, clusters of activated primordial fol-licles with apparently enlarged oocytes were seen inFoxo3a�/� ovaries (supplemental Fig. S2 published assupplemental data on The Endocrine Society’s Jour-nals Online web site; PD35, arrows), suggesting thatthe overactivated oocytes in Foxo3a�/� ovaries un-dergo rapid growth at this stage. Ovaries of bothp27�/� and p27�/�Foxo3a�/� DKO mice also showednumerous developing follicles at PD35 (supplementalFig. S2). At the age of 12 wk, however, Foxo3a�/�
ovaries were depleted of follicle structures (supple-mental Fig. S2), which is a similar phenomenon to thatof the p27�/� ovaries (Fig. 4J).
Studies on the Molecular Mechanisms UnderlyingAverted Follicle Atresia in p27�/� Mice
Elevated Cdk2/Cdc2-Cyclin A/B1 Kinase Activitiesin p27�/� Ovaries Prevent Follicle Death throughSuppression of the Caspase Pathway. It was no-table that as compared with p27�/� ovaries, granulosacells in p27�/� ovaries did not undergo hyperprolif-eration, as shown by in vivo bromodeoxyuridine (BrdU)incorporation assay (Fig. 7A, arrows). This is distinctfrom the prolonged proliferation of luteinizing granu-losa cells of p27�/� ovaries and p21�/�; p27�/� dou-ble null ovaries (28), and is also distinct from the casein hyperplastic thymus and spleen of p27�/� mice (15),indicating that p27 does not play a major role in con-trolling granulosa cell proliferation. To determine themolecular mechanism that prevents follicle atresia be-fore sexual maturity in p27�/� mice, we used 18-d-oldp27�/� and p27�/� ovaries in which no dramatic fol-licle loss has occurred (Fig. 3). Our data showed that inp27�/� ovaries, there was an overall increase in kinaseactivities associated with Cdk2, Cdc2, cyclin A, andcyclin B1, but not with cyclin E1 (Fig. 7B), implying thatthe kinase activities of Cdk2/Cdc2-cyclin A/B1 com-
plexes may be involved in preventing the death ofovarian follicles in p27�/� ovaries. It seems to us thatat different developmental stages and cell populationsof the ovary, Cdc2, cyclin B1, and cyclin E1 activitiesare differently regulated by the loss of p27.
Previous reports have shown that activation ofcaspases is involved in follicle atresia (29–32). By us-ing ovaries from 18-d-old p27�/� mice and p27�/�
mice, we found that the levels of cleaved caspase-9,cleaved caspase-3, cleaved caspase-7, and cleavedpoly (ADP-ribose) polymerase (PARP) were lower inp27�/� ovaries than those in p27�/� ovaries (Fig. 7C).The levels of cleaved caspase-6 and cleaved lamin Awere, however, similar in p27�/� and p27�/� ovaries(data not shown). Our data therefore imply thatthe caspase-9-caspase-3-caspase-7-PARP apoptosispathway is suppressed in p27�/� ovaries, which leadsto an elevated follicle survival rate. Thus, p27 mayinduce follicle atresia before sexual maturity through ac-tivation of the caspase-dependent apoptotic pathways.
DISCUSSION
The endowment of a definite number of primordial ovar-ian follicles and the activation of some of these follicles atchosen times, leading to further development and even-tual release of mature oocytes via ovulation, are essentialfor the propagation of all mammalian species. The end offemale reproductive life occurs when the pool of primor-dial follicles is exhausted (2, 6). In the current study, wehave provided several lines of evidence that implicate theCdk inhibitor p27 as an essential molecule for controllingovarian development via suppression of follicle endow-ment and activation, and provoking follicle death in mice.
Somatic Cell-Expressed p27 in Perinatal OvariesSuppresses Follicle Formation and Endowmentin Mice
In this study, we have provided evidence that p27expressed in somatic cells surrounding oocytes inperinatal ovaries suppresses follicular formation and
the ovaries were fixed in 4% paraformaldehyde, dehydrated, and embedded in paraffin. Sections of 8-�m thickness were stained withhematoxylin for morphological observation of follicular activation. All experiments were repeated at least four times, and representativeimages are shown. C and D, Normal expression of p27 and Foxo3a in Foxo3a�/�and p27�/� oocytes, respectively. Oocytes fromovaries of 8- to 12-d-old Foxo3a�/� or p27�/� mice and their wild-type control mice were isolated. Western blots were performed tomeasure protein levels of Akt, p-Akt (serine 473), Foxo3a, and p-Foxo3a (threonine 32) in p27�/� and p27�/� oocytes, and levels ofp27 expression in Foxo3a�/� and Foxo3a�/� oocytes, as described in Materials and Methods. Levels of �-actin are shown as internalcontrols. The experiments were repeated three times. For each experiment and each genotype, five to 10 mice were used. Repre-sentative images are shown. E–K, Comparison of primordial follicle activation in ovaries of p27�/�, Foxo3a�/�, and p27�/�Foxo3a�/�
DKO mice. Ovaries from 13- and 23-d-old p27�/�, Foxo3a�/�, and p27�/�Foxo3a�/� DKO mice were isolated, fixed in 4% parafor-maldehyde, dehydrated, and embedded in paraffin. Sections of 8-�m thickness were prepared and stained with hematoxylin formorphological observation, as described in Materials and Methods. Primordial follicles (E–G, arrows) and activated or transient follicles(E–G, arrowheads) are indicated. The genotypes and ages of mice are given in the figure. K, Percentages of primordial follicles per ovary(mean � SD) in ovaries of 13-d-old wild-type, p27�/�, Foxo3a�/�, and p27�/�Foxo3a�/� DKO mice. Follicles were counted asdescribed in Materials and Methods (three ovaries from three individual mice were used for each genotype). Lowercase letters (a, b,c, and d) indicate significant differences (P � 0.01). IP, Immunoprecipitation.
Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice Mol Endocrinol, September 2007, 21(9):2189–2202 2197

endowment (Fig. 8A). In perinatal ovaries before pri-mordial follicle formation, p27 is expressed mainly insomatic cells surrounding female germ cells, but not inoogonia/oocytes. The functional role of p27 was elu-cidated by the finding that primordial follicles wereformed earlier in p27�/� ovaries, and postnatal follicleendowment is doubled in p27�/� mice compared withthat in p27�/� mice. Based on our data that in wild-type ovaries before follicle formation, somatic cellssurrounding germ cells do not appear to be prolifer-ating as measured by in vivo BrdU incorporation assay(data not shown), we suggest that p27 may suppressthe proliferation of these somatic cells at this stageand may also regulate the migration of somatic cells
during their invasion into the germ cell clusters. Theloss of p27 in somatic cells may therefore lead toenhanced proliferation and/or increased mobility ofthese cells, which may result in accelerated formationof primordial follicles.
In addition, because the oocyte survival rates fromPD1 to PD8 in p27�/� and p27�/� ovaries appear tobe similar, we presume that the increased number offollicles in p27�/� mice may be caused by enhancedproliferation of female germ cells. Expression of p27 insomatic cells of embryonic and perinatal ovaries maycontribute to the negative regulation of germ cell mi-tosis. We presume that p27 in somatic cells may neg-atively regulate the paracrine and gap junction-medi-
IP Cyc A H1
IP Cyc B1 H1
IP Cdc2
IP Cdk2
H1
H1
p27+/+ p27-/-
Cleaved PARP
Cleaved caspase-3
p27
Cleaved caspase-9
Cleaved caspase-7
B Kinase activity assay (PD18 ovaries)
C Western blot (PD18 ovaries)
p27-/-
p27+/+
A In vivo BrdU incorporation(PD18 ovaries)
H1IP Cyc E1
p27+/+ p27-/-
β-actin
100 µm
100 µm
Fig. 7. Elevation of Kinase Activities Associated with Cdk2/Cdc2-cyclin A/B1 Complexes Prevent Follicle Atresia throughSuppression of the Caspase Pathway
A, BrdU incorporation assay indicated that proliferation of granulosa cells was not accelerated in p27�/� ovaries. In vivo BrdUincorporation assay was performed with 18-d-old p27�/� and p27�/� mice, as described in Materials and Methods. Reddish signalsindicate proliferating cells. Similar BrdU incorporation was seen in granulosa cells of ovaries from p27�/�and p27�/� mice. Theexperiments were repeated three times and representative images are shown. For each experiment and each genotype, one mousewas used. B, Kinase activities associated with Cdk2, Cdc2, cyclin A, cyclin B1, and cyclin E1 complexes in ovaries of p27�/� andp27�/� mice. Ovaries from 18-d-old p27�/� and p27�/� mice were homogenized and 300 �g of ovarian lysate was used forimmunoprecipitation with the antibodies indicated. Kinase activity assays were performed using histone H1 as substrate, as describedin Materials and Methods. All experiments were repeated three times and representative images are shown. For each experiment andeach genotype, five to 10 mice were used. C, Suppressed activation of caspase-9, -3, and -7, and PARP in p27�/�ovaries. Ovaries from18-d-old p27�/� mice and p27�/� mice were collected and used for Western blot, as described in Materials and Methods. Levels ofcleaved caspase-9, -3, and -7, and cleaved PARP, indicating apoptosis in p27�/�and p27�/� ovaries at PD18, are shown. Levels ofp27 and �-actin in the ovaries were used as internal controls. The experiments were repeated three times, and for each experiment,five to 10 mice were used. Representative images are shown. IP, Immunoprecipitation.
2198 Mol Endocrinol, September 2007, 21(9):2189–2202 Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice

ated communication between somatic cells and germcells, by which growth factors, ions, and nutrients, etc.are supplied to germ cells to ensure their survival andproliferation. We believe that the interactions betweengerm cells and somatic cells are important for themitosis/survival of oogonia and for primordial follicleformation, in both of which processes the somaticcell-expressed p27 is actively involved as a negativeregulator.
To identify the molecules that accelerate follicle for-mation in p27�/� mice, we measured the kinase ac-tivity of the major target of p27, Cdk2 (22). Our datasuggest that in neonatal p27�/� ovaries, the kinaseactivities associated with Cdk2-cyclin A are elevatedand may be involved in driving the accelerated follicleformation in these mice. Further experiments areneeded to determine whether the elevated Cdk2-cy-clin A kinase activity in newborn (PD0) p27�/� ovariesis located in oocytes or in somatic cells, or both.Equally importantly, the relationship between p27 andother molecules that have been suggested to haveroles in follicle formation, such as neurotrophins and
neurotrophin receptors (33, 34), synaptonemal com-plex protein-1 (35), activin (36), and factor in the germ-line � (37), is worthy of further study.
p27 Expressed by Oocyte Nuclei andPregranulosa Cells in Primordial FolliclesSuppresses Follicle Activation in Mice
As illustrated in Fig. 8B, in this study we have providedevidence that in primordial follicles, p27 molecules ex-pressed in oocyte nuclei and pregranulosa cells functionas suppressors of oocyte growth and pregranulosa cellproliferation/differentiation, respectively. In p27�/� mice,the pool of primordial follicles was prematurely activatedby early oocyte growth and proliferation of the few flat-tened pregranulosa cells into cuboidal granulosa cells.Thus, our data support the notion that follicular activationis caused by the release from the inhibitory mechanism,which involves p27, that maintains the primordial folliclesin a resting state, as previously hypothesized (6). More-over, we have shown that premature activation of theprimordial follicle pool in p27�/� mice results in depletion
Germ cell syncytia
p27 in somatic cells surrounding oocytes
Atretic follicles
p27 in oocyte nuclei and pregranulosa cells
p27 in oocyte nuclei and pregranulosa cells
p27 in oocyte nuclei and granulosa cells
suppression promotion
Primordial follicles
Activated follicles
A
B
C
C
Folli
cle
activ
atio
n
Follicle formatoin & endowment
Atresia Atresia
Fig. 8. Suggested Functions of p27 in Controlling Mouse Ovarian DevelopmentA, In mouse ovaries before follicle formation, somatic cell-expressed p27 may suppress follicular endowment and follicle
formation by inhibiting germ cell mitosis and also by suppressing migration/proliferation of somatic cells that will invade germ cellsyncytia to form primordial follicles. B, Once a definite number of primordial follicles have been endowed, p27 expressed in oocytenuclei and pregranulosa cells suppresses the activation of primordial follicles. C, Meanwhile, p27 expressed in oocyte nuclei,pregranulosa cells, and granulosa cells promotes follicle atresia and causes the rapid loss of follicles that occurs before sexualmaturity.
Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice Mol Endocrinol, September 2007, 21(9):2189–2202 2199

of functional follicles in early adulthood (i.e. in 12-wk-oldmice), which is a symptom similar to that of POF inhumans. Thus, the p27�/� mice may be a useful modelfor the study of ovarian failure, in which follicles aredepleted due to overactivation.
Previous studies have suggested that oocyte growth isa major event during primordial follicle activation in mam-mals (9). At the same time, p27 has always been con-sidered to be a suppressor of cell cycle progression (14).It is thus interesting to study how the p27-Cdk systemregulates cell growth, rather than cell division, usinggrowing oocytes as a model system. In the current study,we have shown that in oocytes of primordial and earlygrowing follicles in p27�/� mice, the kinase activitiesassociated with Cdk2-cyclin A/E1 appears to be ele-vated. Further in-depth studies are necessary, however,to uncover the mechanisms by which p27 functions as asuppressor of cell (oocyte) growth.
Although it is apparent that both Foxo3a and p27function as suppressors of primordial follicle activation,our data from the current study have indicated that,unlike in other cell types, p27 and Foxo3a do not appearto function in an upstream/downstream fashion in regu-lating follicle activation in mice. In DKO mice that lackboth p27 and Foxo3a, synergistically accelerated follicleactivation was seen, indicating that the mechanisms ofsuppression of follicle activation by p27 or Foxo3a areindependent of each other. Additionally, large numbersof transient follicles with enlarged oocytes that were sur-rounded by a few flattened pregranulosa cells were ob-served in Foxo3a�/� mice (Ref. 12 and the currentstudy). In p27�/� mice, however, enlarged oocytes inactivated follicles were always surrounded by increasednumbers of cuboidal granulosa cells. These results indi-cate that p27, but not Foxo3a, is the factor that sup-presses the proliferation and differentiation of pregranulosacells in primordial follicles. The in-depth mechanisms, in-cluding a possible cross talk between p27-mediated cas-cade and PI3K/Akt/Foxo3a signaling pathway in follicularactivation, are currently under investigation in our labora-tory. Moreover, our preliminary data showing that both p27and Foxo3a are expressed in oocyte nuclei of human pri-mordial and primary follicles suggest that these moleculesmay have a role in regulating follicle activation in humans.
p27 Promotes Postnatal Follicle Death via theCaspase-Dependent Pathway
In mice, a large proportion of follicles disappear fromthe nongrowing follicle pool during the first wave offollicle development (10, 11). However, in p27�/� ova-ries this rapid follicle loss is largely prevented, indicat-ing that p27 is a key molecule in the promotion offollicle atresia before sexual maturity, as illustrated inFig. 8C. The prevented follicle loss in p27�/� ovariesmay be achieved by the premature activation of theprimordial follicle pool. We there propose that p27expressed in primordial and early primary folliclesfunctions as a key regulator that determines the fate offollicles of either being activated and recruited into the
growing population at chosen times, or being removedby atresia.
By performing kinase assays, we showed that Cdk2/Cdc2-cyclin A/B1 complex-associated kinase activitieswere elevated in p27�/� ovaries, which may outweighthe death signals in p27�/� ovaries. Moreover, caspasesare known to be involved in mediating follicle atresia(29–32). Our data demonstrated that the activation of thecaspase-9-caspase-3-caspase-7-PARP cascade wassuppressed in p27�/� ovaries, suggesting that the ele-vated kinase activities associated with Cdk2/Cdc2-cy-clin A/B1 complexes that were caused by p27 loss mayprevent follicle atresia via suppressing the activation ofcaspases -9, -3, and -7. Thus, we can designate p27 asa possible upstream enhancer for caspase activation,which induces follicle atresia.
In conclusion, the current study has provided severallines of evidence that define the previously unknownfunction of p27 to be a suppressor of ovarian follicleendowment/formation and activation, and an enhancerof ovarian follicle atresia. Together with previous reportsthat p27 is important for luteal cell differentiation (15, 16,28), we propose that the deregulation, or malfunctioning,of the ovarian p27-mediated cascade may lead to de-fects in follicular development, which may cause dis-turbed ovarian function and pathological changes in theovary. The findings in this study may provide some usefulknowledge in the search for genetic aberrations of theovary that lead to defects in follicle development in hu-man diseases, such as POF.
MATERIALS AND METHODS
Mouse Lines
C57BL/6J mice were obtained from Charles River Laboratory(Sulzfeld, Germany). The p27�/� mice have been describedpreviously (15), and had been backcrossed to C57BL/6J for10 generations before we performed the experiments. Gen-eration of Foxo3a�/� mice was described previously (27). Inthe current study, the Foxo3a�/� mice have been back-crossed to C57BL/6J for seven generations. Breeding pairsof p27�/� or p27�/� male mice with p27�/� female mice, orFoxo3a�/� or Foxo3a�/� male mice with Foxo3a�/� femalemice were used to obtain littermates for comparison. Toobtain p27�/� Foxo3a�/� DKO mice, an initial breeding withp27�/� males � Foxo3a�/� females was set up to obtainpups for further breeding, and breeding pairs of p27�/�
Foxo3a�/� females � p27�/� Foxo3a�/� males or p27�/�
Foxo3a�/� females � p27�/� Foxo3a�/� males were used toobtain p27�/� Foxo3a�/� female pups. The mice werehoused under controlled environmental conditions with freeaccess to water and food. Illumination was on between 0600and 1800 h. E0.5 refers to the day that a vaginal plug wasfound. The ethics committee of Umeå University approved allexperimental protocols.
Reagents, Antibodies, Immunostaining, Western Blots,and Kinase Activity Assays
Rabbit polyclonal antibodies to p27, phospho-Akt (serine473), cleaved caspase-3, cleaved caspase-6, cleavedcaspase-7, cleaved caspase-9, cleaved PARP, and cleaved
2200 Mol Endocrinol, September 2007, 21(9):2189–2202 Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice

lamin A were obtained from Cell Signaling Technologies (Bev-erly, MA). Rabbit polyclonal antibodies against Foxo3a andphospho-Foxo3a (threonine 32) were from Upstate Biotech-nology, Inc. (Lake Placid, NY). BrdU and mouse monoclonalantibodies against �-actin and BrdU were purchased fromSigma-Aldrich Sweden AB (Stockholm, Sweden). A ratmonoclonal antibody to GCNA1 was kindly provided by Dr.George C. Enders (University of Kansas, Lawrence, KS). Goatpolyclonal antibody against human VASA was purchasedfrom R&D Systems (Minneapolis, MN). Cdk2/Cdc2-specificinhibitor Roscovitine was obtained from Calbiochem (SanDiego, CA). The ABC Staining System for immunohistochem-istry was obtained from Santa Cruz Biotechnology, Inc.(Santa Cruz, CA). Immunostaining was performed accordingto the instructions of the manufacturer. Western blot analysiswas carried out according to the instructions for differentantibodies from the suppliers and visualized using the ECLPlus Western Blotting Detection System (Amersham Bio-sciences, Uppsala, Sweden). For immunoprecipitations andkinase assays using histone H1, antibodies to Cdk2, Cdc2,cyclin A, cyclin B1, and cyclin E1, all conjugated to agarosebeads, were obtained from Santa Cruz Biotechnology, andkinase assays were performed as previously described (38).
Quantification of Oocytes and Follicles
Ovaries were fixed in 4% paraformaldehyde, dehydrated, andembedded in paraffin. For quantification of germ cells inovaries of PD1, serial sections of 5-�m thickness were pre-pared from an ovary, and all sections were stained for GCNA1to visualize germ cells (19). Thus, the germ cell numberscounted represent the GCNA1-positive oocytes. The totalgerm cell number of an ovary was calculated as the sum ofthe number of germ cells from each individual section.
To count the numbers of ovarian follicles in 8- to 35-d-oldmice, paraffin-embedded ovaries were serially sectioned at8-�m thickness and stained with hematoxylin for morpholog-ical analysis, as previously described (13). Ovarian follicles atdifferent stages of development, including primordial (type 2),transient (type 3a, i.e. follicles with enlarged oocytes butflattened pregranulosa cells), primary (type 3b), secondary(type 4), preantral, early tertiary, and tertiary follicles (types5–7) were counted in all sections of an ovary, based on thewell-accepted standards established by Pedersen and Pe-ters (39). In each section, follicles that contained oocytes withclearly visible nuclei were scored as previously described (13,40). The total number of follicles in an ovary was calculated asthe sum of the numbers of follicles from all sections. Judgingfrom careful morphological analysis, the incidence of count-ing the same oocyte or germ cell twice or missing an oocyte/germ cell is negligible. All quantification of oocytes and germcells and all morphological analyses were performed with aZeiss AX10 microscope (Carl Zeiss, Thornwood, NY).
Isolation of Oocytes from Postnatal Mouse Ovaries
Isolation of oocytes, separation of small oocytes from par-tially grown oocytes using a cell dispersing screen with25-�m opening, and lysis of oocytes were performed aspreviously described (18). Red blood cells were removedusing a hypotonic buffer containing 144 mM NH4Cl and 17 mM
Tris-HCl (pH 7.2).
Culture of Postnatal Ovaries
Ovarian cultures were performed as previously described(34), with minor modifications. PD2 p27�/� ovaries were re-moved aseptically, and the whole ovary was cultured in a CellStrainer (40 �m pore size) (BD Biosciences, Stockholm, Swe-den) in 1 ml �-MEM (Life Technologies, Inc., Gaithersburg,MD) supplemented with 28 �M ascorbic acid and 0.3% (wt/
vol) BSA, with or without kinase inhibitors. The cultured ova-ries were incubated in a humidified incubator (5% CO2, 37 C)with one third of the medium exchanged for fresh mediumevery day for the duration of the culture period. For fixation,the ovaries were washed once in PBS and fixed overnight in4% paraformaldehyde and embedded as mentioned abovefor morphological analysis.
Preparation of Ovarian Extracts
Ovaries were dissected free of fat and adhering tissues, andextracts were prepared on ice by homogenizing in a lysisbuffer containing 50 mM Tris-HCl (pH 8.0), 120 mM NaCl, 20mM NaF, 20 mM �-glycerophosphate, 1 mM EDTA, 6 mM
EGTA (pH 8.0), 1% Nonidet P-40, 1 mM dithiothreitol, 5 mM
benzamidine, 1 mM phenylmethylsulfonylfluoride, 250 �M so-dium orthovanadate, 10 �g/ml aprotinin, 10 �g/ml leupeptin,and 1 �g/ml pepstatin, followed by centrifugation at 14,000rpm for 20 min at 4 C. The supernatants were collected andprotein concentrations were determined using the bicincho-ninic acid protein assay.
Statistical Analysis
All experiments were repeated at least three times. For count-ing of oocytes and follicles, three to six mice per group wereused, and data were analyzed with Student’s t test. A differ-ence was considered to be significant when P � 0.01.
Acknowledgments
We thank Dr. Melissa Pepling for constructive discussions.
Received April 4, 2007. Accepted June 5, 2007.Address all correspondence and requests for reprints to:
Kui Liu, Department of Medical Biochemistry and Biophysics,Umeå University, SE-901 87 Umeå, Sweden. E-mail:[email protected].
This work was supported by the Swedish Research Coun-cil, the Swedish Cancer Foundation, and the Swedish LionsCancer Research Foundation in Norrland.
Present address for C.B.: Oncodesign, 20 rue Jean Mazen,BP 27627, 21076 Dijon Cedex, France.
Disclosure Statement: The authors have nothing todisclose.
REFERENCES
1. Pepling ME, Spradling AC 1998 Female mouse germcells form synchronously dividing cysts. Development125:3323–3328
2. Hirshfield AN 1991 Development of follicles in the mamma-lian ovary. Int Rev Cytol 124:43–101
3. McNatty KP, Fidler AE, Juengel JL, Quirke LD, Smith PR,Heath DA, Lundy T, O’Connell A, Tisdall DJ 2000 Growthand paracrine factors regulating follicular formation and cel-lular function. Mol Cell Endocrinol 163:11–20
4. Pepling ME, Spradling AC 2001 Mouse ovarian germ cellcysts undergo programmed breakdown to form primordialfollicles. Dev Biol 234:339–351
5. Epifano O, Dean J 2002 Genetic control of early folliculo-genesis in mice. Trends Endocrinol Metab 13:169–173
6. McGee EA, Hsueh AJ 2000 Initial and cyclic recruitment ofovarian follicles. Endocr Rev 21:200–214
7. Vanderhyden B 2002 Molecular basis of ovarian develop-ment and function. Front Biosci 7:d2006–d2022
8. Eppig JJ, Viveiros MM, Bivens CM, De La Fuente R 2004Regulation of mammalian oocyte maturation. In: Leung PC,
Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice Mol Endocrinol, September 2007, 21(9):2189–2202 2201

Adashi EY, eds. The ovary. 2nd ed. New York: ElsevierAcademic Press; 113–129
9. Wassarman PM, Albertini DF 1994 The mammalian ovum.In: Knobil E, Neill JD, eds. The physiology of reproduction.2nd ed. New York: Raven Press; 79–122
10. Faddy MJ, Telfer E, Gosden RG 1987 The kinetics of pre-antral follicle development in ovaries of CBA/Ca mice dur-ing the first 14 weeks of life. Cell Tissue Kinet 20:551–560
11. Bristol-Gould SK, Kreeger PK, Selkirk CG, Kilen SM, MayoKE, Shea LD, Woodruff TK 2006 Fate of the initial folliclepool: empirical and mathematical evidence supporting itssufficiency for adult fertility. Dev Biol 298:149–154
12. Castrillon DH, Miao L, Kollipara R, Horner JW, DePinho RA2003 Suppression of ovarian follicle activation in mice bythe transcription factor Foxo3a. Science 301:215–218
13. Liu L, Rajareddy S, Reddy P, Du C, Jagarlamudi K, Shen Y,Gunnarsson D, Selstam G, Boman K, Liu K 2007 Infertilitycaused by retardation of follicular development in mice withoocyte-specific expression of Foxo3a. Development 134:199–209
14. Kaldis P 2007 Another piece of the p27(Kip1) puzzle. Cell128:241–244
15. Fero ML, Rivkin M, Tasch M, Porter P, Carow CE, Firpo E,Polyak K, Tsai LH, Broudy V, Perlmutter RM, Kaushansky K,Roberts JM 1996 A syndrome of multiorgan hyperplasiawith features of gigantism, tumorigenesis, and female ste-rility in p27(Kip1)-deficient mice. Cell 85:733–744
16. Kiyokawa H, Kineman RD, Manova-Todorova KO, SoaresVC, Hoffman ES, Ono M, Khanam D, Hayday AC, FrohmanLA, Koff A 1996 Enhanced growth of mice lacking thecyclin-dependent kinase inhibitor function of p27(Kip1). Cell85:721–732
17. Nakayama K, Ishida N, Shirane M, Inomata A, Inoue T,Shishido N, Horii I, Loh DY, Nakayama K 1996 Mice lackingp27(Kip1) display increased body size, multiple organ hy-perplasia, retinal dysplasia, and pituitary tumors. Cell 85:707–720
18. Reddy P, Shen L, Ren C, Boman K, Lundin E, Ottander U,Lindgren P, Liu YX, Sun QY, Liu K 2005 Activation of Akt(PKB) and suppression of FKHRL1 in mouse and rat oo-cytes by stem cell factor during follicular activation anddevelopment. Dev Biol 281:160–170
19. Enders GC, May JJ 1994 Developmentally regulated ex-pression of a mouse germ cell nuclear antigen examinedfrom embryonic day 11 to adult in male and female mice.Dev Biol 163:331–340
20. Moons DS, Jirawatnotai S, Tsutsui T, Franks R, Parlow AF,Hales DB, Gibori G, Fazleabas AT, Kiyokawa H 2002 Intactfollicular maturation and defective luteal function in micedeficient for cyclin-dependent kinase-4. Endocrinology143:647–654
21. Malumbres M, Sotillo R, Santamaria D, Galan J, Cerezo A,Ortega S, Dubus P, Barbacid M 2004 Mammalian cellscycle without the D-type cyclin-dependent kinases Cdk4and Cdk6. Cell 118:493–504
22. Aleem E, Kiyokawa H, Kaldis P 2005 Cdc2-cyclin E com-plexes regulate the G1/S phase transition. Nat Cell Biol7:831–836
23. Lan ZJ, Xu X, Cooney AJ 2004 Differential oocyte-specificexpression of Cre recombinase activity in GDF-9-iCre,Zp3cre, and Msx2Cre transgenic mice. Biol Reprod 71:1469–1474
24. Liu K, Rajareddy S, Liu L, Jagarlamudi K, Boman K, SelstamG, Reddy P 2006 Control of mammalian oocyte growth andearly follicular development by the oocyte PI3 kinasepathway: new roles for an old timer. Dev Biol 299:1–11
25. Arden KC, Biggs WH, III 2002 Regulation of the FoxO familyof transcription factors by phosphatidylinositol-3 kinase-activated signaling. Arch Biochem Biophys 403:292–298
26. Accili D, Arden KC 2004 FoxOs at the crossroads of cellularmetabolism, differentiation, and transformation. Cell 117:421–426
27. Lin L, Hron JD, Peng SL 2004 Regulation of NF-�B, Thactivation, and autoinflammation by the forkhead transcrip-tion factor Foxo3a. Immunity 21:203–213
28. Jirawatnotai S, Moons DS, Stocco CO, Franks R, Hales DB,Gibori G, Kiyokawa H 2003 The cyclin-dependent kinaseinhibitors p27Kip1 and p21Cip1 cooperate to restrict pro-liferative life span in differentiating ovarian cells. J Biol Chem278:17021–17027
29. Robles R, Tao XJ, Trbovich AM, Maravel DV, Nahum R,Perez GI, Tilly KI, Tilly JL 1999 Localization, regulation andpossible consequences of apoptotic protease-activatingfactor-1 (Apaf-1) expression in granulosa cells of the mouseovary. Endocrinology 140:2641–2644
30. Moriyama T, Maravei DV, Kuida K, Tilly JL, Role ofcaspase-9 in programmed cell death execution in the ovary.Program of the 84th Annual Meeting of The Endocrine So-ciety, San Francisco, CA, 2002, pp 418–419
31. Matikainen T, Perez GI, Zheng TS, Kluzak TR, Rueda BR,Flavell RA, Tilly JL 2001 Caspase-3 gene knockout definescell lineage specificity for programmed cell death signalingin the ovary. Endocrinology 142:2468–2480
32. Johnson AL, Bridgham JT 2000 Caspase-3 and -6 expres-sion and enzyme activity in hen granulosa cells. Biol Reprod62:589–598
33. Dissen GA, Romero C, Hirshfield AN, Ojeda SR 2001 Nervegrowth factor is required for early follicular development inthe mammalian ovary. Endocrinology 142:2078–2086
34. Spears N, Molinek MD, Robinson LL, Fulton N, Cameron H,Shimoda K, Telfer EE, Anderson RA, Price DJ 2003 The roleof neurotrophin receptors in female germ-cell survival inmouse and human. Development 130:5481–5491
35. Paredes A, Garcia-Rudaz C, Kerr B, Tapia V, Dissen GA,Costa ME, Cornea A, Ojeda SR 2005 Loss of synaptonemalcomplex protein-1, a synaptonemal complex protein, con-tributes to the initiation of follicular assembly in the devel-oping rat ovary. Endocrinology 146:5267–5277
36. Bristol-Gould SK, Kreeger PK, Selkirk CG, Kilen SM, CookRW, Kipp JL, Shea LD, Mayo KE, Woodruff TK 2006 Post-natal regulation of germ cells by activin: the establishmentof the initial follicle pool. Dev Biol 298:132–148
37. Soyal SM, Amleh A, Dean J 2000 FIG�, a germ cell-specifictranscription factor required for ovarian follicle formation.Development 127:4645–4654
38. Berthet C, Aleem E, Coppola V, Tessarollo L, Kaldis P 2003Cdk2 knockout mice are viable. Curr Biol 13:1775–1785
39. Pedersen T, Peters H 1968 Proposal for a classification ofoocytes and follicles in the mouse ovary. J Reprod Fertil17:555–557
40. Johnson J, Canning J, Kaneko T, Pru JK, Tilly JL 2004Germline stem cells and follicular renewal in the postnatalmammalian ovary. Nature 428:145–150
Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremostprofessional society serving the endocrine community.
2202 Mol Endocrinol, September 2007, 21(9):2189–2202 Rajareddy et al. • p27kip1 Controls Ovarian Development in Mice




















