
The expression and role of c-Myc in mouse hair follicle morphogenesis and cycling
8
Acta Histochemica 114 (2012) 199–206 Contents lists available at ScienceDirect Acta Histochemica jou rnal h o mepage: www.elsevier.de/acthis The expression and role of c-Myc in mouse hair follicle morphogenesis and cycling Ning Wang a , Tian Yang a , Jin Li b , Mingxing Lei b , Jiazhong Shi a , Weiming Qiu a , Xiaohua Lian a,∗ a Department of Cell Biology, Third Military Medical University, Gao Tanyan 30, Sha Pingba, Chongqing, 400038, PR China b Department of Bioengineering, Chongqing University, Chongqing, 400044, PR China a r t i c l e i n f o Article history: Received 7 March 2011 Received in revised form 24 April 2011 Accepted 27 April 2011 Keywords: Hair follicle morphogenesis Hair follicle cycle c-Myc Keratinocytes Mice a b s t r a c t Although the function of c-Myc has been clarified in many tissues, until now its expression and role in hair follicle morphogenesis and the hair cycle remains unknown. In this study we detected c-Myc expression pattern in the process of mouse hair follicle development and normal cycle. We found that during hair follicle morphogenesis, the stage-specific expression of c-Myc was detected in mouse skin and was predominantly localized to the hair follicle epithelium. c-Myc expression was also consistently found in mouse skin throughout the hair follicle cycle. Through the in vivo injection of c-Myc inhibitory peptide and c-Myc expression plasmid, we also investigated the direct effects of c-Myc on the hair follicle structures during the hair follicle cycle. Our results showed that c-Myc inhibitory peptide significantly restrained the development of anagen hair follicles, while the injection of plasmid DNA encoding c-Myc in vivo clearly promoted anagen development. Our data indicate that c-Myc may play an important role in the proliferation and differentiation of the hair follicle keratinocytes during hair follicle development. c-Myc also was shown to participate in the regulation of the mouse hair growth cycle and could promote the proliferation of the hair matrix keratinocytes as well as the differentiation of the inner root sheath. © 2011 Elsevier GmbH. All rights reserved. Introduction The hair follicle is a cup shaped structure that consists of many concentric layers with different functions. Hair follicle morphogen- esis is initiated by cross-talk between ectodermal epithelium and mesenchyme at embryonic day 13.5 (E13.5) and continues until after birth. The aggregation of mesenchymal fibroblasts induces downward growth of the epithelium by proliferation and invagi- nation and successively leads to the formation of the hair placode, hair germ and hair peg. When the dermal condensate, referred to as the dermal papilla (DP), is engulfed by the epithelium, the epithe- lial cells proximal to the DP, which is known as the matrix, begin to grow and differentiate, and finally generate the hair shaft and inner root sheath (IRS). Surrounding the IRS, a stratified epithe- lium, known as the outer root sheath (ORS), is continuous with the epidermis. The hair follicle stem cells are located in the bulge region Abbreviations: DP, dermal papilla; EGFP, enhanced green fluorescence pro- tein; IRS, inner root sheath; ORS, outer root sheath; c-Myc-ER TM , fusion protein between c-Myc and the tamoxifen responsive hormone-binding domain of the estrogen receptor; PBS, phosphate buffered saline; HE, hematoxylin and eosin; PVDF, polyvinylidene difluoride; RT-PCR, reverse transcription polymerase chain reaction; SDS, sodium dodecyl sulphate. ∗ Corresponding author. E-mail address: [email protected] (X. Lian). of the ORS (Barinov and Sulaeva, 2004; Krause and Foitzik, 2006; Schneider et al., 2009). Mature hair follicles undergo repeated cycles of growth. The normal hair cycle can be divided into three phases: anagen (grow- ing phase), catagen (intermediate phase) and telogen (resting phase). In the anagen stage, the follicles, regenerating from the active hair follicle stem cells, grow actively and then attain their maximum length. Catagen is a transitional period of hair follicle from growing to resting. During this stage, the hair follicles undergo degradation and shrink in length owing to the destruction of the lower part of the hair follicle. Telogen is the resting phase follow- ing the catagen. During telogen the hair follicles cease to grow and the dermal papilla, which previously was enveloped by surround- ing epithelial cells, now becomes a small ball of cells located at the base of the follicle (Botchkarev and Paus, 2003; Alonso and Fuchs, 2006; Gorpinich and Nozdrin, 2007). The morphogenesis and growth cycle of hair follicles is maintained by a controlled balance between cell proliferation, dif- ferentiation and death. Revealing the control mechanisms of hair morphogenesis and cycle is essential for understanding normal hair growth, hair disorders as well as some hair follicle-related skin tumors. Numerous cytokines, transcription factors and adhesion molecules have been shown to be involved in the control of hair fol- licle development and cycling (Stenn and Paus, 2001; Botchkarev and Paus, 2003; Rogers, 2004). 0065-1281/$ – see front matter © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.acthis.2011.04.009
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The expression and role of c-Myc in mouse hair follicle morphogenesis and cycling

Ta
Na
b
a
ARRA
KHHcKM
I
cemadnhtltile
tbePr
0d
Acta Histochemica 114 (2012) 199– 206
Contents lists available at ScienceDirect
Acta Histochemica
jou rna l h o mepage: www.elsev ier .de /ac th is
he expression and role of c-Myc in mouse hair follicle morphogenesisnd cycling
ing Wanga, Tian Yanga, Jin Lib, Mingxing Leib, Jiazhong Shia, Weiming Qiua, Xiaohua Liana,∗
Department of Cell Biology, Third Military Medical University, Gao Tanyan 30, Sha Pingba, Chongqing, 400038, PR ChinaDepartment of Bioengineering, Chongqing University, Chongqing, 400044, PR China
r t i c l e i n f o
rticle history:eceived 7 March 2011eceived in revised form 24 April 2011ccepted 27 April 2011
eywords:air follicle morphogenesis
a b s t r a c t
Although the function of c-Myc has been clarified in many tissues, until now its expression and rolein hair follicle morphogenesis and the hair cycle remains unknown. In this study we detected c-Mycexpression pattern in the process of mouse hair follicle development and normal cycle. We found thatduring hair follicle morphogenesis, the stage-specific expression of c-Myc was detected in mouse skinand was predominantly localized to the hair follicle epithelium. c-Myc expression was also consistentlyfound in mouse skin throughout the hair follicle cycle. Through the in vivo injection of c-Myc inhibitorypeptide and c-Myc expression plasmid, we also investigated the direct effects of c-Myc on the hair follicle
air follicle cycle-Myceratinocytesice
structures during the hair follicle cycle. Our results showed that c-Myc inhibitory peptide significantlyrestrained the development of anagen hair follicles, while the injection of plasmid DNA encoding c-Mycin vivo clearly promoted anagen development. Our data indicate that c-Myc may play an important rolein the proliferation and differentiation of the hair follicle keratinocytes during hair follicle development.c-Myc also was shown to participate in the regulation of the mouse hair growth cycle and could promotethe proliferation of the hair matrix keratinocytes as well as the differentiation of the inner root sheath.
© 2011 Elsevier GmbH. All rights reserved.
ntroduction
The hair follicle is a cup shaped structure that consists of manyoncentric layers with different functions. Hair follicle morphogen-sis is initiated by cross-talk between ectodermal epithelium andesenchyme at embryonic day 13.5 (E13.5) and continues until
fter birth. The aggregation of mesenchymal fibroblasts inducesownward growth of the epithelium by proliferation and invagi-ation and successively leads to the formation of the hair placode,air germ and hair peg. When the dermal condensate, referred to ashe dermal papilla (DP), is engulfed by the epithelium, the epithe-ial cells proximal to the DP, which is known as the matrix, begino grow and differentiate, and finally generate the hair shaft and
nner root sheath (IRS). Surrounding the IRS, a stratified epithe-ium, known as the outer root sheath (ORS), is continuous with thepidermis. The hair follicle stem cells are located in the bulge regionAbbreviations: DP, dermal papilla; EGFP, enhanced green fluorescence pro-ein; IRS, inner root sheath; ORS, outer root sheath; c-Myc-ERTM, fusion proteinetween c-Myc and the tamoxifen responsive hormone-binding domain of thestrogen receptor; PBS, phosphate buffered saline; HE, hematoxylin and eosin;VDF, polyvinylidene difluoride; RT-PCR, reverse transcription polymerase chaineaction; SDS, sodium dodecyl sulphate.∗ Corresponding author.
E-mail address: [email protected] (X. Lian).
065-1281/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.acthis.2011.04.009
of the ORS (Barinov and Sulaeva, 2004; Krause and Foitzik, 2006;Schneider et al., 2009).
Mature hair follicles undergo repeated cycles of growth. Thenormal hair cycle can be divided into three phases: anagen (grow-ing phase), catagen (intermediate phase) and telogen (restingphase). In the anagen stage, the follicles, regenerating from theactive hair follicle stem cells, grow actively and then attain theirmaximum length. Catagen is a transitional period of hair folliclefrom growing to resting. During this stage, the hair follicles undergodegradation and shrink in length owing to the destruction of thelower part of the hair follicle. Telogen is the resting phase follow-ing the catagen. During telogen the hair follicles cease to grow andthe dermal papilla, which previously was enveloped by surround-ing epithelial cells, now becomes a small ball of cells located at thebase of the follicle (Botchkarev and Paus, 2003; Alonso and Fuchs,2006; Gorpinich and Nozdrin, 2007).
The morphogenesis and growth cycle of hair follicles ismaintained by a controlled balance between cell proliferation, dif-ferentiation and death. Revealing the control mechanisms of hairmorphogenesis and cycle is essential for understanding normal hairgrowth, hair disorders as well as some hair follicle-related skin
tumors. Numerous cytokines, transcription factors and adhesionmolecules have been shown to be involved in the control of hair fol-licle development and cycling (Stenn and Paus, 2001; Botchkarevand Paus, 2003; Rogers, 2004).
2 ochem
fedmNetpcadrms(MogiItwegcmindK
siuapsWhdg
M
A
oiMlospces
H
gta
00 N. Wang et al. / Acta Hist
c-Myc, as a basic helix-loop helix leucine zipper transcriptionactor that plays extensive roles in cell cycle control, cell differ-ntiation and apoptosis. c-Myc activation has been observed in theevelopment of many tissues, such as the pancreas, liver and mam-ary glands (Blakely et al., 2005; Gruppuso and Sanders, 2006;akhai et al., 2008), but there is no information about the dynamicxpression of c-Myc during hair follicle morphogenesis. Moreover,he expression and the role of c-Myc in the hair follicle cycle isoorly understood. In humans, there are only a few studies on-Myc in hair follicle cycle, but the results from different reportsre somewhat conflicting. c-Myc immunoreactivity has never beenetected during anagen and telogen, and the location and possibleoles of c-Myc are disputed. In some studies, c-Myc expression wasainly found in the inner root sheath, which implied a relation-
hip between c-Myc and the differentiation of the inner root sheathRumio et al., 2000; Barajon et al., 2001). While in other reports, c-
yc protein was preferentially located in the follicle bulb and theuter root sheath, especially in the bulge region, and it was sug-ested that in the hair follicle, c-Myc may have important functionsn regulating cell proliferation and stem cell action (Bull et al., 2001).n mice, there are only limited reports on the expression of c-Myc inhe normal hair follicle cycle. Only in c-Myc-ERTM transgenic mice,as the ectopic expression of c-Myc detected in hair follicles (Bull
t al., 2005).AE15 is a monoclonal antibody against trichohyalinranules. Because trichohyalin granules could promote the cornifi-ation of the inner root sheath, they were regarded as an importantarker of the inner root sheath and AE15 was usually used in the
dentification of the inner root sheath differentiation. Ki-67 is auclear protein, which is especially expressed in proliferating cellsuring the cell cycle and necessary for cellular proliferation, so thei-67 antigen is a reliable marker for cellular proliferation.
In this study, we initially investigated the presence and expres-ion of c-Myc during mouse hair follicle morphogenesis and thendentified the stage and location of c-Myc expression in the nat-ral hair follicle cycle. We also used a c-Myc inhibitory peptidend c-Myc expression plasmid to interrupt the function of c-Mycrotein in vivo, and then traced the alteration of the hair follicletructure during the hair cycle by using AE15 and Ki-67 antibodies.
e aimed to explore the precise expression pattern of c-Myc inair follicle development and normal cycle, and also to reveal moreetails about the regulated effects of c-Myc on cell proliferation,rowth, and differentiation of hair follicles.
aterials and methods
nimals
C57BL/6 mice were purchased from experimental animal centref the Third Military Medical University, Chongqing, China. Accord-ng to the classical method described by Paus et al. (1999) and
üller-Röver et al. (2001), skin specimens were respectively col-ected on embryonic days 13.5, 15.5, 19.5 (E13.5, E15.5, E19.5) andn postnatal days 4, 8 (P4, P8) for the assessment c-Myc expres-ion in hair follicle morphogenesis. Dorsal skin was harvested onostnatal days 17, 25 and 33 (P17, P25, P33) for the analysis of-Myc expression in the hair follicle cycle. 5 mice were used atach time point. 8–9 weeks old mice were selected for induction ofynchronized hair cycle.
ematoxylin and eosin (HE) staining
Paraffin sections were routinely dewaxed and hydrated inraded ethanols. Sections were firstly stained with the alum hema-oxylin, and then washed in running tap water. After differentiatingnd rewashing, sections were counterstained with eosin for 3 s,
ica 114 (2012) 199– 206
then dehydrated with graded ethanols and mounted with mount-ing medium.
Immunohistochemical staining
Skin specimens were fixed in 4% paraformaldehyde solutionand embedded in paraffin wax. Sections of skin specimens weredewaxed, rehydrated, and washed three times with phosphate-buffered saline (PBS). After treatment with citrate-buffered salinein a microwave oven for 10 min (92–100 ◦C), sections were treatedwith H2O2 for 10 min at room temperature, then washed threetimes with PBS and incubated with the following primary anti-body overnight at 4 ◦C: mouse anti-c-myc monoclonal antibody(eBioscience, San Diego, CA, USA), goat polyclonal antibody to Ki67(Santa Cruz Biotechnol., Santa Cruz, CA, USA), AE15 monoclonalantibody (a kind gift from Dr. Tung-Tien Sun, Professor of New YorkUniversity). Sections were then labelled with HRP-conjugated sec-ondary antibodies (Zhongshan Goldbridge Biotechnology, Beijing,China) and visualized with DAB peroxidase substrate (ZhongshanGoldbridge Biotechnology). Finally, sections were counterstainedwith hematoxylin. A negative control was performed by omittingthe first antibody from the procedure.
RT-PCR
Total RNA was isolated from skin specimens using Trizol reagent(Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s rec-ommended protocols. 0.6 �g of total RNAs were reverse transcribedwith ReverTra Ace-�-reverse transcription kit (Toyobo, Osaka,Japan). Products of reverse transcriptase reaction were subjectedto PCR cycles with specific primer set. The primer pairs used in thestudy were as follows: c-Myc, 5′-TCACCAGCACAACTACGCCG-3′ and 5′-CAGGATGTAGGCGGTGGCTT-3′; GAPDH, 5′-ACCACAGTCCATGCCATCAC-3′ and 5′-TCCACCACCCTGTTGCTGTA-3′. Samples were denatured at 94 ◦C for 3 min and followed by 29cycles of PCR amplification (94 ◦C, 30 s for denaturation; 60 ◦C, 30 sfor annealing; 72 ◦C, 30 s for extension). The PCR products wereelectrophoresed on agarose gels and their amount was evaluatedby ethidium bromide staining.
Western blot
Skin specimens harvested at selected time points were lysed inRIPA buffer (150 mM NaCl, 50 mM Tris, pH 7.4, 1 mM EDTA, 0.1%NP-40 and 0.2 mM PMSF). After vigorous pipetting, lysates werecentrifuged for 15 min at 13,000 rpm. Total protein was measuredby a Bradford protein assay kit (Bioteke, Beijing, China). Sam-ples were run on SDS-polyacrylamide gels and transferred ontoPVDF membranes, then blocked in 5% defatted milk for 30 min andincubated with c-myc monoclonal antibody for overnight at 4 ◦Cwith gentle agitation. Blots were then incubated with peroxidase-conjugated secondary antibodies for 60 min at room temperature,and visualized by enhanced chemiluminescence (Thermo FisherScientific, Hudson, NH, USA).
Hair cycle synchronization
Female C57 BL/6 mice at 8–9 weeks of age were purchased.Only mice with pink skin color and all hair follicles of the dorsalskin in telogen were used. Induction of synchronized anagen phasewas performed by depilation of the hair shafts under anesthesia asdescribed by Paus (Paus et al., 1994). Briefly, female C57 BL/6 mice
in telogen of hair follicle cycle (8–9 weeks old), as judged by thepink color of their back skin, were chosen and anesthetized by 1%pentobarbital sodium. The mixture of molten wax/rosin (1:1) wasapplied on the mice dorsal skin. After coagulation, the wax/rosin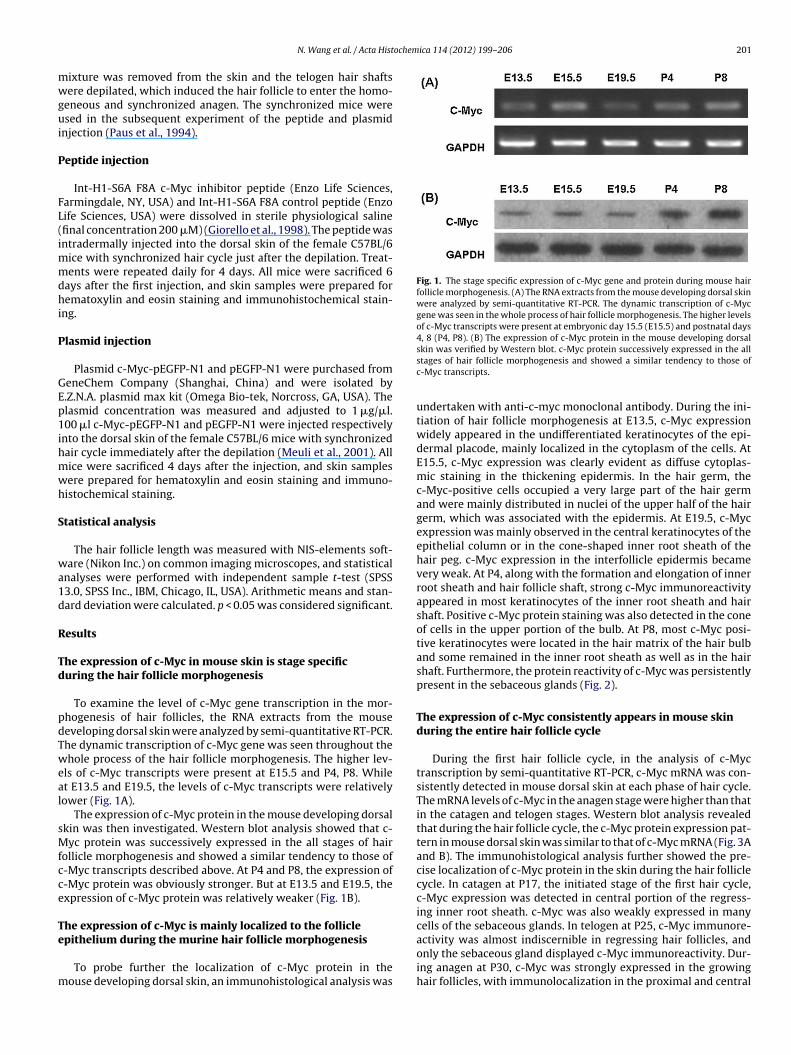
ochemica 114 (2012) 199– 206 201
mwgui
P
FL(immdhi
P
GEp1ihmwh
S
wa1d
R
Td
pdTweal
sMfcce
Te
m
Fig. 1. The stage specific expression of c-Myc gene and protein during mouse hairfollicle morphogenesis. (A) The RNA extracts from the mouse developing dorsal skinwere analyzed by semi-quantitative RT-PCR. The dynamic transcription of c-Mycgene was seen in the whole process of hair follicle morphogenesis. The higher levelsof c-Myc transcripts were present at embryonic day 15.5 (E15.5) and postnatal days4, 8 (P4, P8). (B) The expression of c-Myc protein in the mouse developing dorsal
N. Wang et al. / Acta Hist
ixture was removed from the skin and the telogen hair shaftsere depilated, which induced the hair follicle to enter the homo-
eneous and synchronized anagen. The synchronized mice weresed in the subsequent experiment of the peptide and plasmid
njection (Paus et al., 1994).
eptide injection
Int-H1-S6A F8A c-Myc inhibitor peptide (Enzo Life Sciences,armingdale, NY, USA) and Int-H1-S6A F8A control peptide (Enzoife Sciences, USA) were dissolved in sterile physiological salinefinal concentration 200 �M) (Giorello et al., 1998). The peptide wasntradermally injected into the dorsal skin of the female C57BL/6
ice with synchronized hair cycle just after the depilation. Treat-ents were repeated daily for 4 days. All mice were sacrificed 6
ays after the first injection, and skin samples were prepared forematoxylin and eosin staining and immunohistochemical stain-
ng.
lasmid injection
Plasmid c-Myc-pEGFP-N1 and pEGFP-N1 were purchased fromeneChem Company (Shanghai, China) and were isolated by.Z.N.A. plasmid max kit (Omega Bio-tek, Norcross, GA, USA). Thelasmid concentration was measured and adjusted to 1 �g/�l.00 �l c-Myc-pEGFP-N1 and pEGFP-N1 were injected respectively
nto the dorsal skin of the female C57BL/6 mice with synchronizedair cycle immediately after the depilation (Meuli et al., 2001). Allice were sacrificed 4 days after the injection, and skin samplesere prepared for hematoxylin and eosin staining and immuno-istochemical staining.
tatistical analysis
The hair follicle length was measured with NIS-elements soft-are (Nikon Inc.) on common imaging microscopes, and statistical
nalyses were performed with independent sample t-test (SPSS3.0, SPSS Inc., IBM, Chicago, IL, USA). Arithmetic means and stan-ard deviation were calculated. p < 0.05 was considered significant.
esults
he expression of c-Myc in mouse skin is stage specificuring the hair follicle morphogenesis
To examine the level of c-Myc gene transcription in the mor-hogenesis of hair follicles, the RNA extracts from the mouseeveloping dorsal skin were analyzed by semi-quantitative RT-PCR.he dynamic transcription of c-Myc gene was seen throughout thehole process of the hair follicle morphogenesis. The higher lev-
ls of c-Myc transcripts were present at E15.5 and P4, P8. Whilet E13.5 and E19.5, the levels of c-Myc transcripts were relativelyower (Fig. 1A).
The expression of c-Myc protein in the mouse developing dorsalkin was then investigated. Western blot analysis showed that c-yc protein was successively expressed in the all stages of hair
ollicle morphogenesis and showed a similar tendency to those of-Myc transcripts described above. At P4 and P8, the expression of-Myc protein was obviously stronger. But at E13.5 and E19.5, thexpression of c-Myc protein was relatively weaker (Fig. 1B).
he expression of c-Myc is mainly localized to the follicle
pithelium during the murine hair follicle morphogenesisTo probe further the localization of c-Myc protein in theouse developing dorsal skin, an immunohistological analysis was
skin was verified by Western blot. c-Myc protein successively expressed in the allstages of hair follicle morphogenesis and showed a similar tendency to those ofc-Myc transcripts.
undertaken with anti-c-myc monoclonal antibody. During the ini-tiation of hair follicle morphogenesis at E13.5, c-Myc expressionwidely appeared in the undifferentiated keratinocytes of the epi-dermal placode, mainly localized in the cytoplasm of the cells. AtE15.5, c-Myc expression was clearly evident as diffuse cytoplas-mic staining in the thickening epidermis. In the hair germ, thec-Myc-positive cells occupied a very large part of the hair germand were mainly distributed in nuclei of the upper half of the hairgerm, which was associated with the epidermis. At E19.5, c-Mycexpression was mainly observed in the central keratinocytes of theepithelial column or in the cone-shaped inner root sheath of thehair peg. c-Myc expression in the interfollicle epidermis becamevery weak. At P4, along with the formation and elongation of innerroot sheath and hair follicle shaft, strong c-Myc immunoreactivityappeared in most keratinocytes of the inner root sheath and hairshaft. Positive c-Myc protein staining was also detected in the coneof cells in the upper portion of the bulb. At P8, most c-Myc posi-tive keratinocytes were located in the hair matrix of the hair bulband some remained in the inner root sheath as well as in the hairshaft. Furthermore, the protein reactivity of c-Myc was persistentlypresent in the sebaceous glands (Fig. 2).
The expression of c-Myc consistently appears in mouse skinduring the entire hair follicle cycle
During the first hair follicle cycle, in the analysis of c-Myctranscription by semi-quantitative RT-PCR, c-Myc mRNA was con-sistently detected in mouse dorsal skin at each phase of hair cycle.The mRNA levels of c-Myc in the anagen stage were higher than thatin the catagen and telogen stages. Western blot analysis revealedthat during the hair follicle cycle, the c-Myc protein expression pat-tern in mouse dorsal skin was similar to that of c-Myc mRNA (Fig. 3Aand B). The immunohistological analysis further showed the pre-cise localization of c-Myc protein in the skin during the hair folliclecycle. In catagen at P17, the initiated stage of the first hair cycle,c-Myc expression was detected in central portion of the regress-ing inner root sheath. c-Myc was also weakly expressed in manycells of the sebaceous glands. In telogen at P25, c-Myc immunore-
activity was almost indiscernible in regressing hair follicles, andonly the sebaceous gland displayed c-Myc immunoreactivity. Dur-ing anagen at P30, c-Myc was strongly expressed in the growinghair follicles, with immunolocalization in the proximal and central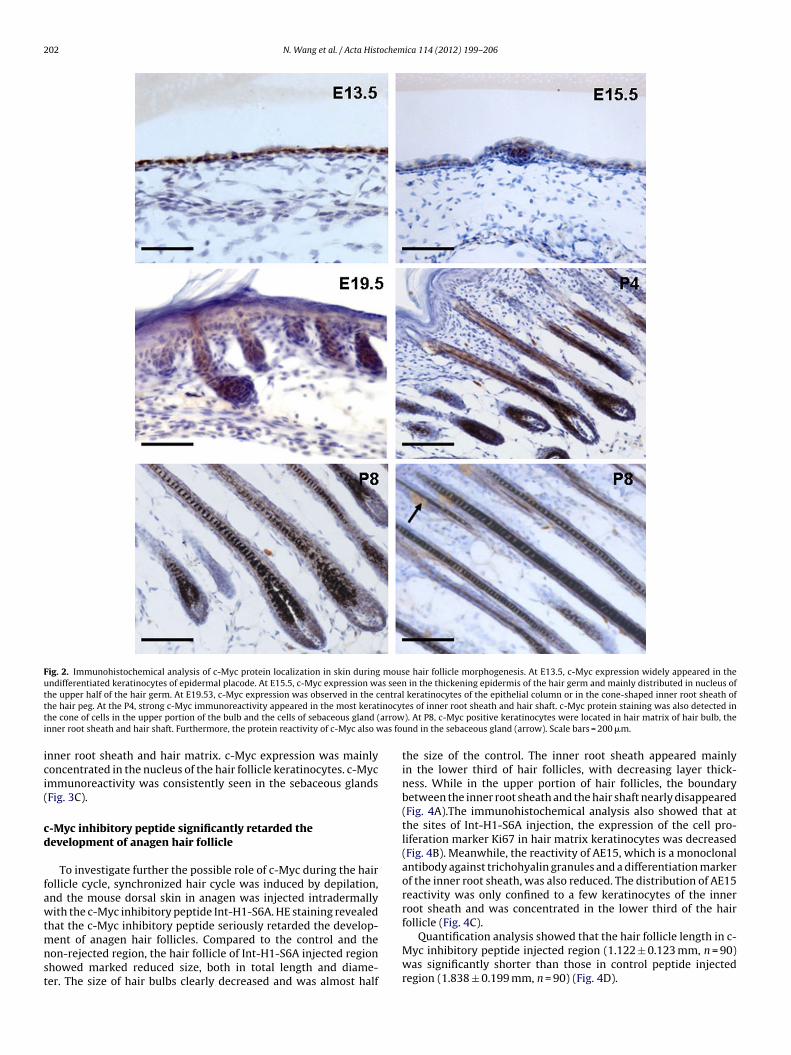
202 N. Wang et al. / Acta Histochemica 114 (2012) 199– 206
Fig. 2. Immunohistochemical analysis of c-Myc protein localization in skin during mouse hair follicle morphogenesis. At E13.5, c-Myc expression widely appeared in theundifferentiated keratinocytes of epidermal placode. At E15.5, c-Myc expression was seen in the thickening epidermis of the hair germ and mainly distributed in nucleus ofthe upper half of the hair germ. At E19.53, c-Myc expression was observed in the central keratinocytes of the epithelial column or in the cone-shaped inner root sheath oft inocyt arrowi as fo
ici(
cd
fawtmnst
he hair peg. At the P4, strong c-Myc immunoreactivity appeared in the most kerathe cone of cells in the upper portion of the bulb and the cells of sebaceous gland (nner root sheath and hair shaft. Furthermore, the protein reactivity of c-Myc also w
nner root sheath and hair matrix. c-Myc expression was mainlyoncentrated in the nucleus of the hair follicle keratinocytes. c-Mycmmunoreactivity was consistently seen in the sebaceous glandsFig. 3C).
-Myc inhibitory peptide significantly retarded theevelopment of anagen hair follicle
To investigate further the possible role of c-Myc during the hairollicle cycle, synchronized hair cycle was induced by depilation,nd the mouse dorsal skin in anagen was injected intradermallyith the c-Myc inhibitory peptide Int-H1-S6A. HE staining revealed
hat the c-Myc inhibitory peptide seriously retarded the develop-
ent of anagen hair follicles. Compared to the control and theon-rejected region, the hair follicle of Int-H1-S6A injected regionhowed marked reduced size, both in total length and diame-er. The size of hair bulbs clearly decreased and was almost half
tes of inner root sheath and hair shaft. c-Myc protein staining was also detected in). At P8, c-Myc positive keratinocytes were located in hair matrix of hair bulb, theund in the sebaceous gland (arrow). Scale bars = 200 �m.
the size of the control. The inner root sheath appeared mainlyin the lower third of hair follicles, with decreasing layer thick-ness. While in the upper portion of hair follicles, the boundarybetween the inner root sheath and the hair shaft nearly disappeared(Fig. 4A).The immunohistochemical analysis also showed that atthe sites of Int-H1-S6A injection, the expression of the cell pro-liferation marker Ki67 in hair matrix keratinocytes was decreased(Fig. 4B). Meanwhile, the reactivity of AE15, which is a monoclonalantibody against trichohyalin granules and a differentiation markerof the inner root sheath, was also reduced. The distribution of AE15reactivity was only confined to a few keratinocytes of the innerroot sheath and was concentrated in the lower third of the hairfollicle (Fig. 4C).
Quantification analysis showed that the hair follicle length in c-Myc inhibitory peptide injected region (1.122 ± 0.123 mm, n = 90)was significantly shorter than those in control peptide injectedregion (1.838 ± 0.199 mm, n = 90) (Fig. 4D).

N. Wang et al. / Acta Histochemica 114 (2012) 199– 206 203
Fig. 3. The expression of c-Myc gene and protein during the hair follicle cycle. (A) The RNA extracts from the mouse dorsal skin at three phases of hair cycle were analyzedby semi-quantitative RT-PCR. The mRNA levels of c-Myc in the anagen stage were much higher than that in catagen and telogen stages. (B) The protein extracts from themouse dorsal skin at three phases of hair cycle were analyzed by Western blot. c-Myc protein expression pattern in mouse dorsal skin was similar to that of c-Myc mRNA. (C)The immunohistochemical analysis of c-Myc protein in skin during the hair follicle cycle. In catagen at P17, the initiated stage of the first hair cycle, c-Myc expression wasd Myc imt ed in ts
Ia
doeicM(ef(c
D
Mco
pwtMoftsdmwssba
etected in central portion of the regressing inner root sheath. In telogen at P25, c-he sebaceous gland (arrow). During the anagen at P30, c-Myc was strongly expressebaceous gland (arrow). Scale bars = 150 �m.
njection of plasmid DNA encoding c-Myc in vivo promotednagen development
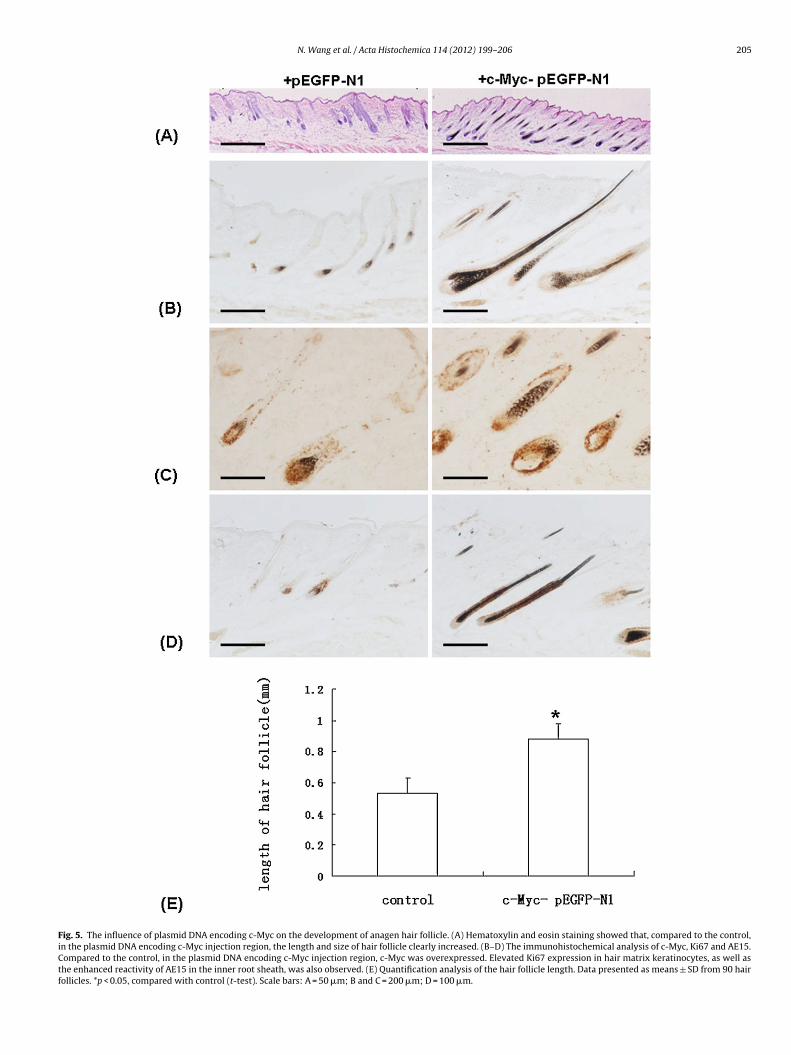
Using the synchronized hair cycle mouse model, we alsoetected the hair follicle growth in anagen phase under conditionsf c-Myc overexpression. After the injection of the plasmid DNAncoding c-Myc, the hair follicle in the injection region increasedn length and size and showed an elevated expression of c-Myc,ompared the control (Fig. 5A and B). The overexpression of c-yc promoted the Ki67 expression in hair matrix keratinocytes
Fig. 5C). The reactivity of AE15 in inner root sheath was alsonhanced (Fig. 5D). Quantification analysis showed that the hairollicle length in plasmid DNA encoding c-Myc injected region0.879 ± 0.123 mm, n = 90) was significantly longer than those inontrol (0.530 ± 0.072 mm, n = 90) (Fig. 5E).
iscussion
Here we report for the first time the expression pattern of c-yc during morphogenesis of mouse hair follicles and proved that
-Myc was an essential regulation factor for the hair follicle devel-pment.
We also found that during different stages of hair follicle mor-hogenesis, the diversity in c-Myc expression actually existed,hich suggested that c-Myc may have a distinct function during
he different stage of hair follicle development. Firstly, because c-yc expression was specifically detected in the most keratinocytes
f hair germ at E15.5, it is possible that at the early stages of hairollicle morphogenesis, c-Myc may be involved in the prolifera-ion regulation of progenitor cells in the hair germ. But at the latertages of hair follicle morphogenesis, c-Myc expression was notetected in the proliferating keratinocytes in the lower hair bulbatrix, and instead appeared in the cone of inner root sheath asell as the upper parts of hair bulb, the precursor of the inner root
heath and hair shaft. Consequently during the middle and latetages of hair follicle development, the major role of c-Myc maye related to the differentiation initiation of the inner root sheathnd the hair sheath. Furthermore, c-Myc may also participate in
munoreactivity was almost undetectable in the regressed hair follicle, but only inhe growing hair follicle, c-Myc immunoreactivity was also consistently seen in the
the differentiation maintenance of the inner root sheath and thehair sheath, because c-Myc consistently expressed in the inner rootsheath and hair shaft when the hair follicle became more elongatedand mature. So it was proposed that during mouse hair follicle mor-phogenesis, the role of c-Myc may be stage dependent. The stagedependent role of c-Myc was also found in the morphogenesis ofother tissues (Blakely et al., 2005).
Until now, c-Myc expression during hair follicle cycle wasreported only in humans. Here, we firstly defined the hair cycle-dependent expression pattern of c-Myc in mice, which accuratelycoincided with the c-Myc expression pattern in human hair folli-cles as found in previous studies (Rumio et al., 2000; Barajon et al.,2001; Bull et al., 2001), and suggested the potential role of c-Mychair follicle cycle.
For a long time, because there was an argument about the c-Mycdistribution in human hair follicles during the hair follicle cycle, theprecise function of c-Myc in hair follicle was unclear. Rumio et al.(2000) and Barajon et al. (2001) have shown the specific presence ofc-Myc immunoreactivity in the inner root sheath of human anagenhair follicle. Conversely, Bull observed c-Myc immunostaining onlyin the outer root sheath as well as in the hair matrix (Bull et al.,2001). In our present study, the c-Myc immunoreactivity was alsodetected in the inner root sheath and the hair matrix of the mouseanagen hair follicle, but we did not detect c-Myc immunoreactivityin the outer root sheath. The reason for this discrepancy may bedue to the species difference between humans and mice.
In this study, because significant expression of c-Myc was foundin the hair matrix and the inner root sheath of the anagen hairfollicle, we proposed that during the anagen stage of the micehair follicle cycle, c-Myc may be involved in the regulation notonly the hair matrix keratinocyte proliferation, but also the innerroot sheath differentiation. Our in vivo inhibitory experiment fur-ther revealed that the depression of c-Myc function may not onlydepress the dividing hair matrix keratinocytes and lead to the
decrease in the number of the hair matrix cells, but also interferewith the differentiation of the inner root sheath. So, in the normalgrowth of anagen hair follicle, c-Myc may exert its regulation effectby promoting the proliferation of the hair matrix keratinocytes and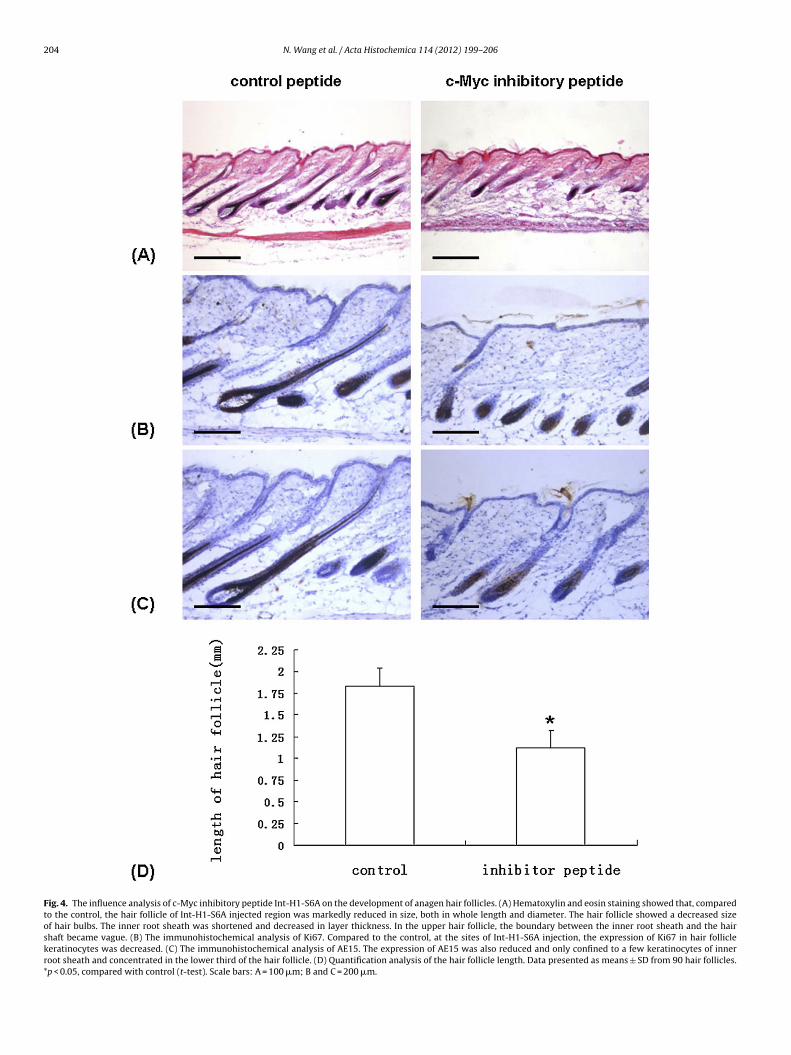
204 N. Wang et al. / Acta Histochemica 114 (2012) 199– 206
Fig. 4. The influence analysis of c-Myc inhibitory peptide Int-H1-S6A on the development of anagen hair follicles. (A) Hematoxylin and eosin staining showed that, comparedto the control, the hair follicle of Int-H1-S6A injected region was markedly reduced in size, both in whole length and diameter. The hair follicle showed a decreased sizeof hair bulbs. The inner root sheath was shortened and decreased in layer thickness. In the upper hair follicle, the boundary between the inner root sheath and the hairshaft became vague. (B) The immunohistochemical analysis of Ki67. Compared to the control, at the sites of Int-H1-S6A injection, the expression of Ki67 in hair folliclekeratinocytes was decreased. (C) The immunohistochemical analysis of AE15. The expression of AE15 was also reduced and only confined to a few keratinocytes of innerroot sheath and concentrated in the lower third of the hair follicle. (D) Quantification analysis of the hair follicle length. Data presented as means ± SD from 90 hair follicles.*p < 0.05, compared with control (t-test). Scale bars: A = 100 �m; B and C = 200 �m.
N. Wang et al. / Acta Histochemica 114 (2012) 199– 206 205
Fig. 5. The influence of plasmid DNA encoding c-Myc on the development of anagen hair follicle. (A) Hematoxylin and eosin staining showed that, compared to the control,in the plasmid DNA encoding c-Myc injection region, the length and size of hair follicle clearly increased. (B–D) The immunohistochemical analysis of c-Myc, Ki67 and AE15.Compared to the control, in the plasmid DNA encoding c-Myc injection region, c-Myc was overexpressed. Elevated Ki67 expression in hair matrix keratinocytes, as well asthe enhanced reactivity of AE15 in the inner root sheath, was also observed. (E) Quantification analysis of the hair follicle length. Data presented as means ± SD from 90 hairfollicles. *p < 0.05, compared with control (t-test). Scale bars: A = 50 �m; B and C = 200 �m; D = 100 �m.

2 ochem
tcTd
lekhrm2oolae
msrdodMf
aMcAtatuge
pmfpOhid
A
Nf
R
A
AB
B
06 N. Wang et al. / Acta Hist
he differentiation of the inner root sheath. This suggestion wasonfirmed by our in vivo injection of plasmid DNA encoding c-Myc.he over expression of c-Myc induced the excessive growth andifferentiation of anagen hair follicles.
We also observed the c-Myc expression in the mouse hair fol-icle during the catagen phase of hair follicle cycle. The centrallement of the catagen phase is the apoptosis of the hair follicleeratinocyte. So the characteristic of c-Myc expression in catagenair follicle may indicate its possible role in the apoptosis of inneroot sheath. Although the apoptosis driven by c-Myc was shown inany cell types (You et al., 2002; Morrish et al., 2003; Shiio et al.,
006), to date there has been no direct proof about the c-Myc effectn hair follicle apoptosis. On the contrary, It was found that thever-expression of c-Myc did not enhance the apoptosis of hair fol-icles (Bull et al., 2005). Thus the significance of c-Myc expressions well as its role in the catagen hair follicle needs to be furtherlucidated.
In this study, we found that accompanying the hair follicleorphogenesis, the sebaceous gland developed with c-Myc expres-
ion detected in developing sebaceous gland of P4 and P8. Theseesults were similar to a previous report (Allen et al., 2003) andemonstrated the possible role of c-Myc in sebaceous gland devel-pment. c-Myc may also have effect on the sebaceous glandsuring the hair follicle cycle, because here we detected the c-yc expression in sebaceous gland of the anagen and telogen hair
ollicle.It was reported that c-Myc could regulate the proliferation
nd the differentiation of sebaceous gland. The data from the c-yc-ERTM transgenic mice indicated that the overexpression of
-Myc resulted in enlarged sebaceous glands (Bull et al., 2005).fter being transduced with a retrovirus encoding c-Myc-ERTM,
he lipid droplets of sebaceous gland cells increased in numbernd size (Lo Celso et al., 2008). So the possible role of c-Myc inhe sebaceous gland described above may be involved in its reg-lation in the proliferation and the differentiation of sebaceousland cells, but this needs to be further confirmed by more directxperiments.
In conclusion, we demonstrated for the first time the expressionattern of c-Myc in the mouse skin during the hair follicle develop-ent. Our data suggested that at different stages of the mouse hair
ollicle morphogenesis, c-Myc may play an important role in theroliferation or the differentiation of the hair follicle keratinocytes.n the basis of defining the expression pattern of c-Myc duringair cycle, we also directly verified the promotion role of c-Myc
n the proliferation of the hair matrix keratinocytes as well as theifferentiation of inner root sheath.
cknowledgements
This study was supported by grants 30972645 from the Nationalature Science Foundation of China and grants CSTC 2007BB5010
rom the National Nature Science Foundation of Chongqing, China.
eferences
llen M, Grachtchouk M, Sheng H, Grachtchouk V, Wang A, WeiL, et al. Hedgehog signaling regulates sebaceous gland develop-ment. Am J Pathol 2003;163:2173–8.
lonso L, Fuchs E. The hair cycle. J Cell Sci 2006;119:391–3.arajon I, Rumio C, Donetti E, Imberti A, Brivio M, Castano P. Pattern
of expression of c-Myc, Max and Bin1 in human anagen hairfollicles. Br J Dermatol 2001;144:1193–203.
arinov EF, Sulaeva. On histophysiology of hair follicles: currentconcepts. Usp Fiziol Nauk 2004;35:65–77.
ica 114 (2012) 199– 206
Blakely CM, Sintasath L, D’Cruz CM, Hahn KT, Dugan KD, Belka GK,et al. Developmental stage determines the effects of MYC in themammary epithelium. Development 2005;132:1147–60.
Botchkarev VA, Paus R. Molecular biology of hair morphogen-esis: development and cycling. J Exp Zool B Mol Dev Evol2003;298:164–80.
Bull JJ, Müller-Röver S, Patel SV, Chronnell CM, McKay IA, PhilpottMP. Contrasting localization of c-Myc with other Myc superfam-ily transcription factors in the human hair follicle and during thehair growth cycle. J Invest Dermatol 2001;116:617–22.
Bull JJ, Pelengaris S, Hendrix S, Chronnell CM, Khan M, Philpott MP.Ectopic expression of c-Myc in the skin affects the hair growthcycle and causes an enlargement of the sebaceous gland. Br JDermatol 2005;152:1125–33.
Giorello L, Clerico L, Pescarolo MP, Vikhanskaya F, Salmona M,Colella G, et al. Inhibition of cancer cell growth and c-Myc tran-scriptional activity by a c-Myc helix 1-type peptide fused to aninternalization sequence. Cancer Res 1998;58:3654–9.
Gorpinich IV, Nozdrin VI. Morpho-functional changes in hair duringtheir renewal. Morfologiia 2007;132:7–17.
Gruppuso PA, Sanders JA. Coordinated regulation of c-Myc and Maxin rat liver development. Am J Physiol Gastrointest Liver Physiol2006;290:G145–55.
Krause K, Foitzik K. Biology of the hair follicle: the basics. SeminCutan Med Surg 2006;25:2–10.
Lo Celso C, Berta MA, Braun KM, Frye M, Lyle S, Zouboulis CC, et al.Characterization of bipotential epidermal progenitors derivedfrom human sebaceous gland: contrasting roles of c-Myc andbeta-catenin. Stem Cells 2008;26:1241–52.
Meuli M, Liu Y, Liggitt D, Kashani-Sabet M, Knauer S, Meuli-SimmenC, et al. Efficient gene expression in skin wound sites followinglocal plasmid injection. J Invest Dermatol 2001;116:131–5.
Morrish F, Giedt C, Hockenbery D. c-MYC apoptotic functionis mediated by NRF-1 target genes. Genes Dev 2003;17:240–55.
Müller-Röver S, Handjiski B, van der Veen C, Eichmüller S, FoitzikK, McKay IA, et al. A comprehensive guide for the accurate clas-sification of murine hair follicles in distinct hair cycle stages. JInvest Dermatol 2001;117:3–15.
Nakhai H, Siveke JT, Mendoza-Torres L, Schmid RM. Conditionalinactivation of Myc impairs development of the exocrine pan-creas. Development 2008;135:3191–6.
Paus R, Handjiski B, Czarnetzki BM, EichmuÈ ller S. A murine modelfor inducing and manipulating hair follicle regression (catagen):effects of dexamethasone and cyclosporin A. J Invest Dermatol1994;103:143–7.
Paus R, Müller-Röver S, Van Der Veen C, Maurer M, Eichmüller S,Ling G, et al. A comprehensive guide for the recognition andclassification of distinct stages of hair follicle morphogenesis. JInvest Dermatol 1999;113:523–32.
Rogers GE. Hair follicle differentiation and regulation. Int J Dev Biol2004;48:163–70.
Rumio C, Donetti E, Imberti A, Barajon I, Prosperi E, Brivio MF, et al.c-Myc expression in human anagen hair follicles. Br J Dermatol2000;142:1092–9.
Schneider MR, Schmidt-Ullrich R, Paus R. The hair follicle as adynamic miniorgan. Curr Biol 2009;19:R132–42.
Shiio Y, Suh KS, Lee H, Yuspa SH, Eisenman RN, Aebersold R. Quan-titative proteomic analysis of myc-induced apoptosis: a directrole for Myc induction of the mitochondrial chloride ion channel,mtCLIC/CLIC4. J Biol Chem 2006;281:2750–6.
Stenn KS, Paus R. Controls of hair follicle cycling. Phys Rev
2001;81:449–94.You Z, Saims D, Chen S, Zhang Z, Guttridge DC, Guan KL, et al. Wntsignaling promotes oncogenic transformation by inhibiting c-Myc-induced apoptosis. J Cell Biol 2002;57:429–40.




















