
Overexpression of a Protein Phosphatase 2C from Beech Seeds in Arabidopsis Shows Phenotypes Related...
11
Overexpression of a Protein Phosphatase 2C from Beech Seeds in Arabidopsis Shows Phenotypes Related to Abscisic Acid Responses and Gibberellin Biosynthesis 1 David Reyes, Dolores Rodrı´guez, Mary Paz Gonza ´lez-Garcı´a, Oscar Lorenzo, Gregorio Nicola ´s, Jose ´ Luis Garcı ´a-Martı ´nez, and Carlos Nicola ´s* Departamento de Fisiologı ´a Vegetal, Centro Hispano-Luso de Investigaciones Agrarias, Facultad de Biologı ´a, Universidad de Salamanca, 37007 Salamanca, Spain (D. Reyes, D. Rodrı ´guez, M.P.G.-G., O.L., G.N., C.N.); and Instituto de Biologı ´a Molecular y Celular de Plantas, Universidad Polite ´cnica de Valencia-Consejo Superior de Investigaciones Cientificas, 46022 Valencia, Spain (J.L.G.-M.) A functional abscisic acid (ABA)-induced protein phosphatase type 2C (PP2C) was previously isolated from beech (Fagus sylvatica) seeds (FsPP2C2). Because transgenic work is not possible in beech, in this study we overexpressed this gene in Arabidopsis (Arabidopsis thaliana) to provide genetic evidence on FsPP2C2 function in seed dormancy and other plant responses. In contrast with other PP2Cs described so far, constitutive expression of FsPP2C2 in Arabidopsis, under the cauliflower mosaic virus 35S promoter, produced enhanced sensitivity to ABA and abiotic stress in seeds and vegetative tissues, dwarf phenotype, and delayed flowering, and all these effects were reversed by gibberellic acid application. The levels of active gibberellins (GAs) were reduced in 35S:FsPP2C2 plants, although transcript levels of AtGA20ox1 and AtGA3ox1 increased, probably as a result of negative feedback regulation, whereas the expression of GASA1 was induced by GAs. Additionally, FsPP2C2-overexpressing plants showed a strong induction of the Responsive to ABA 18 (RAB18) gene. Interestingly, FsPP2C2 contains two nuclear targeting sequences, and transient expression assays revealed that ABA directed this protein to the nucleus. Whereas other plant PP2Cs have been shown to act as negative regulators, our results support the hypothesis that FsPP2C2 is a positive regulator of ABA. Moreover, our results indicate the existence of potential cross-talk between ABA signaling and GA biosynthesis. Abscisic acid (ABA) and GAs regulate several as- pects of plant growth showing antagonistic effects in different developmental processes, including seed dormancy or germination events (for review, see Kermode, 2005; Kucera et al., 2005). Several components implicated in ABA signaling, mainly those regulating seed dormancy, have been identified in ABA-response mutants by isolating the affected genes and their corresponding proteins in- volving a complex network of positive and negative regulators, including kinases, phosphatases, and tran- scriptional regulators (for review, see Finkelstein et al., 2002; Abe et al., 2003). Protein phosphorylation/de- phosphorylation events have been investigated in detail in ABA-mediated processes involving several well-known protein kinases and phosphatases (Leung and Giraudat, 1998; Finkelstein et al., 2002), such as PKABA1, an ABA-induced protein kinase that sup- presses GA-inducible gene expression in barley (Hor- deum vulgare) aleurone layers (Go ´mez-Cadenas et al., 1999, 2001); the guard cell-specific protein kinase AAPK, necessary for stomatal closure mediated by ABA (Li et al., 2000); and the protein kinase OST-1, an essential component mediating stomatal regulation in response to drought (Mustilli et al., 2002). Addition- ally, four Arabidopsis (Arabidopsis thaliana) Ser-Thr pro- tein phosphatase 2Cs (PP2Cs; ABI1, ABI2, AtPP2CA, and HAB1) and one PP2C from beech (Fagus sylvatica; FsPP2C1) have been described as negative regulators of the ABA signal transduction cascade (Gosti et al., 1999; Merlot et al., 2001; Tahtiharju and Palva, 2001; Gonza ´lez-Garcı ´a et al., 2003; Sa ´ez et al., 2004; Kuhn et al., 2006; Yoshida et al., 2006). In addition, analysis of mutants with reduced pro- duction of GAs has demonstrated the role of these hormones in several aspects of plant development, including stem elongation, fruit set, flower induction, or seed germination (for review, see Lange, 1998). For instance, Arabidopsis mutants blocked in GA biosyn- thesis fail to complete germination, this effect being reverted by exogenous addition of GAs (Koornneef and van der Veen, 1980). Three different phases can be considered within the biosynthetic pathway of GAs (Olszewski et al., 2002). The first one is the formation of ent-kaurene, involving geranylgeranyl diphosphate cyclation via copalyl diphosphate as an intermediate. 1 This work was supported by the Ministerio de Ciencia y Tecnologı ´a, Spain (grant nos. BFI2003–01755 and BIO2003–00151), by Junta de Castilla y Leo ´ n (grant no. SA046A05), and by a ‘‘Ramo ´ny Cajal’’ research contract (to O.L.). * Corresponding author; e-mail [email protected]; fax 34–923294682. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantphysiol.org) is: Carlos Nicola ´s ([email protected]). Article, publication date, and citation information can be found at www.plantphysiol.org/cgi/doi/10.1104/pp.106.084681. 1414 Plant Physiology, August 2006, Vol. 141, pp. 1414–1424, www.plantphysiol.org Ó 2006 American Society of Plant Biologists
Transcript of Overexpression of a Protein Phosphatase 2C from Beech Seeds in Arabidopsis Shows Phenotypes Related...

Overexpression of a Protein Phosphatase 2C fromBeech Seeds in Arabidopsis Shows Phenotypes Related toAbscisic Acid Responses and Gibberellin Biosynthesis1
David Reyes, Dolores Rodrıguez, Mary Paz Gonzalez-Garcıa, Oscar Lorenzo, Gregorio Nicolas,Jose Luis Garcıa-Martınez, and Carlos Nicolas*
Departamento de Fisiologıa Vegetal, Centro Hispano-Luso de Investigaciones Agrarias, Facultad de Biologıa,Universidad de Salamanca, 37007 Salamanca, Spain (D. Reyes, D. Rodrıguez, M.P.G.-G., O.L., G.N., C.N.);and Instituto de Biologıa Molecular y Celular de Plantas, Universidad Politecnica de Valencia-ConsejoSuperior de Investigaciones Cientificas, 46022 Valencia, Spain (J.L.G.-M.)
A functional abscisic acid (ABA)-induced protein phosphatase type 2C (PP2C) was previously isolated from beech (Fagussylvatica) seeds (FsPP2C2). Because transgenic work is not possible in beech, in this study we overexpressed this gene inArabidopsis (Arabidopsis thaliana) to provide genetic evidence on FsPP2C2 function in seed dormancy and other plantresponses. In contrast with other PP2Cs described so far, constitutive expression of FsPP2C2 in Arabidopsis, under thecauliflower mosaic virus 35S promoter, produced enhanced sensitivity to ABA and abiotic stress in seeds and vegetativetissues, dwarf phenotype, and delayed flowering, and all these effects were reversed by gibberellic acid application. The levelsof active gibberellins (GAs) were reduced in 35S:FsPP2C2 plants, although transcript levels of AtGA20ox1 and AtGA3ox1increased, probably as a result of negative feedback regulation, whereas the expression of GASA1 was induced by GAs.Additionally, FsPP2C2-overexpressing plants showed a strong induction of the Responsive to ABA 18 (RAB18) gene.Interestingly, FsPP2C2 contains two nuclear targeting sequences, and transient expression assays revealed that ABA directedthis protein to the nucleus. Whereas other plant PP2Cs have been shown to act as negative regulators, our results support thehypothesis that FsPP2C2 is a positive regulator of ABA. Moreover, our results indicate the existence of potential cross-talkbetween ABA signaling and GA biosynthesis.
Abscisic acid (ABA) and GAs regulate several as-pects of plant growth showing antagonistic effects indifferent developmental processes, including seeddormancy or germination events (for review, seeKermode, 2005; Kucera et al., 2005).
Several components implicated in ABA signaling,mainly those regulating seed dormancy, have beenidentified in ABA-response mutants by isolating theaffected genes and their corresponding proteins in-volving a complex network of positive and negativeregulators, including kinases, phosphatases, and tran-scriptional regulators (for review, see Finkelstein et al.,2002; Abe et al., 2003). Protein phosphorylation/de-phosphorylation events have been investigated indetail in ABA-mediated processes involving severalwell-known protein kinases and phosphatases (Leungand Giraudat, 1998; Finkelstein et al., 2002), such as
PKABA1, an ABA-induced protein kinase that sup-presses GA-inducible gene expression in barley (Hor-deum vulgare) aleurone layers (Gomez-Cadenas et al.,1999, 2001); the guard cell-specific protein kinaseAAPK, necessary for stomatal closure mediated byABA (Li et al., 2000); and the protein kinase OST-1, anessential component mediating stomatal regulation inresponse to drought (Mustilli et al., 2002). Addition-ally, fourArabidopsis (Arabidopsis thaliana) Ser-Thr pro-tein phosphatase 2Cs (PP2Cs; ABI1, ABI2, AtPP2CA,and HAB1) and one PP2C from beech (Fagus sylvatica;FsPP2C1) have been described as negative regulatorsof the ABA signal transduction cascade (Gosti et al.,1999; Merlot et al., 2001; Tahtiharju and Palva, 2001;Gonzalez-Garcıa et al., 2003; Saez et al., 2004; Kuhnet al., 2006; Yoshida et al., 2006).
In addition, analysis of mutants with reduced pro-duction of GAs has demonstrated the role of thesehormones in several aspects of plant development,including stem elongation, fruit set, flower induction,or seed germination (for review, see Lange, 1998). Forinstance, Arabidopsis mutants blocked in GA biosyn-thesis fail to complete germination, this effect beingreverted by exogenous addition of GAs (Koornneefand van der Veen, 1980). Three different phases can beconsidered within the biosynthetic pathway of GAs(Olszewski et al., 2002). The first one is the formationof ent-kaurene, involving geranylgeranyl diphosphatecyclation via copalyl diphosphate as an intermediate.
1 This work was supported by the Ministerio de Ciencia yTecnologıa, Spain (grant nos. BFI2003–01755 and BIO2003–00151),by Junta de Castilla y Leon (grant no. SA046A05), and by a ‘‘Ramon yCajal’’ research contract (to O.L.).
* Corresponding author; e-mail [email protected]; fax 34–923294682.The author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Carlos Nicolas ([email protected]).
Article, publication date, and citation information can be found atwww.plantphysiol.org/cgi/doi/10.1104/pp.106.084681.
1414 Plant Physiology, August 2006, Vol. 141, pp. 1414–1424, www.plantphysiol.org � 2006 American Society of Plant Biologists
Ent-kaurene is then converted to GA12 and GA53 (a C-13hydroxylated GA) by oxidative reactions catalyzed byP450-dependent monooxygenases. Finally, during thelast phase, GA 20-oxidases and GA 3-oxidases catalyzethe last steps of the GA biosynthesis pathway. These are
key enzymes controlling active GA biosynthesis fromGA12 andGA53 (Xu et al., 1995;Williams et al., 1998) thatregulate many developmental processes, such as pro-motion of seed germination. Thus, it has been shownthat GAs and the expression of a GA 20-oxidase(FsGA20ox1) play an important role in breaking thedormancy of beech seeds (Calvo et al., 2004). Also,Perez-Flores et al. (2003) have indicated that the level ofseed dormancy presented by different lines of sorghum(Sorghum bicolor) correlates with the level of expressionof two different GA 20-oxidases. Additionally, it hasbeen shown that GA 3-oxidase genes also play an im-portant role in the stimulation of seed germination inArabidopsis (Yamauchi et al., 2004).
In a previous work, we reported the cloning ofFsPP2C2, encoding a functional PP2C from beechnut,whose expression was induced by ABA in seeds andother tissues (Lorenzo et al., 2002). The alignment of thecatalytic cores of several PP2Cs showed that this PP2Cis included in cluster 3, where PP2Cs have not beenstudied in detail (Saez et al., 2004). The commoncharacteristic of all the phosphatases included in this
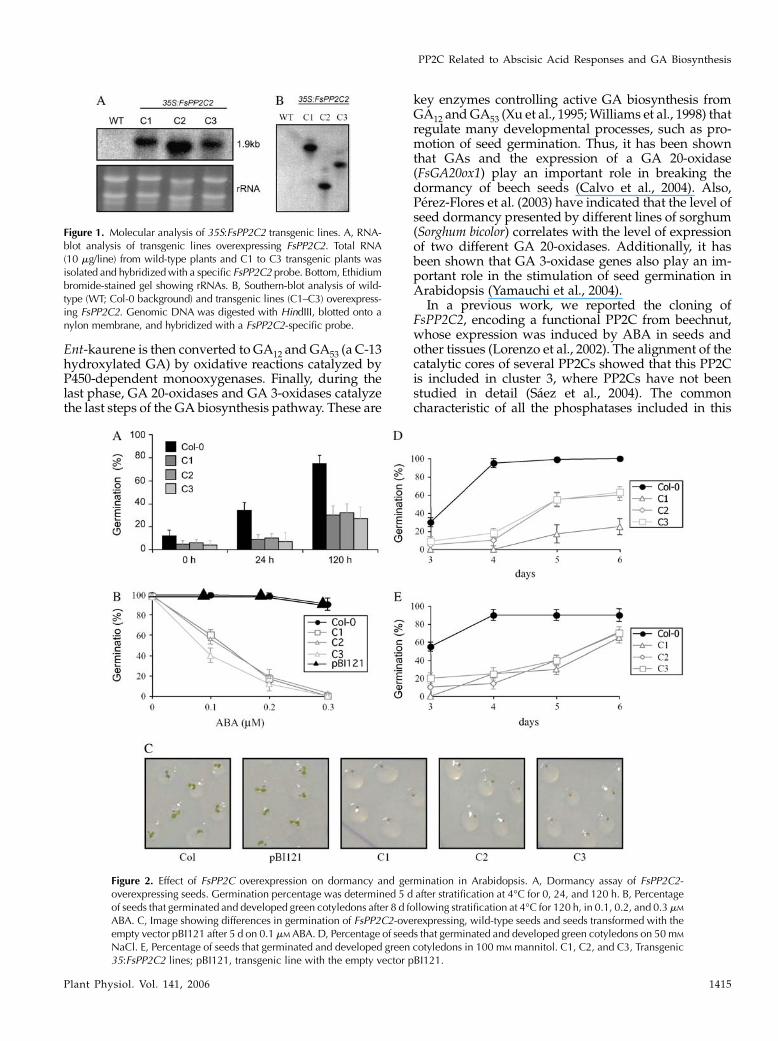
Figure 1. Molecular analysis of 35S:FsPP2C2 transgenic lines. A, RNA-blot analysis of transgenic lines overexpressing FsPP2C2. Total RNA(10 mg/line) from wild-type plants and C1 to C3 transgenic plants wasisolated and hybridizedwith a specific FsPP2C2 probe. Bottom, Ethidiumbromide-stained gel showing rRNAs. B, Southern-blot analysis of wild-type (WT; Col-0 background) and transgenic lines (C1–C3) overexpress-ing FsPP2C2. Genomic DNA was digested with HindIII, blotted onto anylon membrane, and hybridized with a FsPP2C2-specific probe.
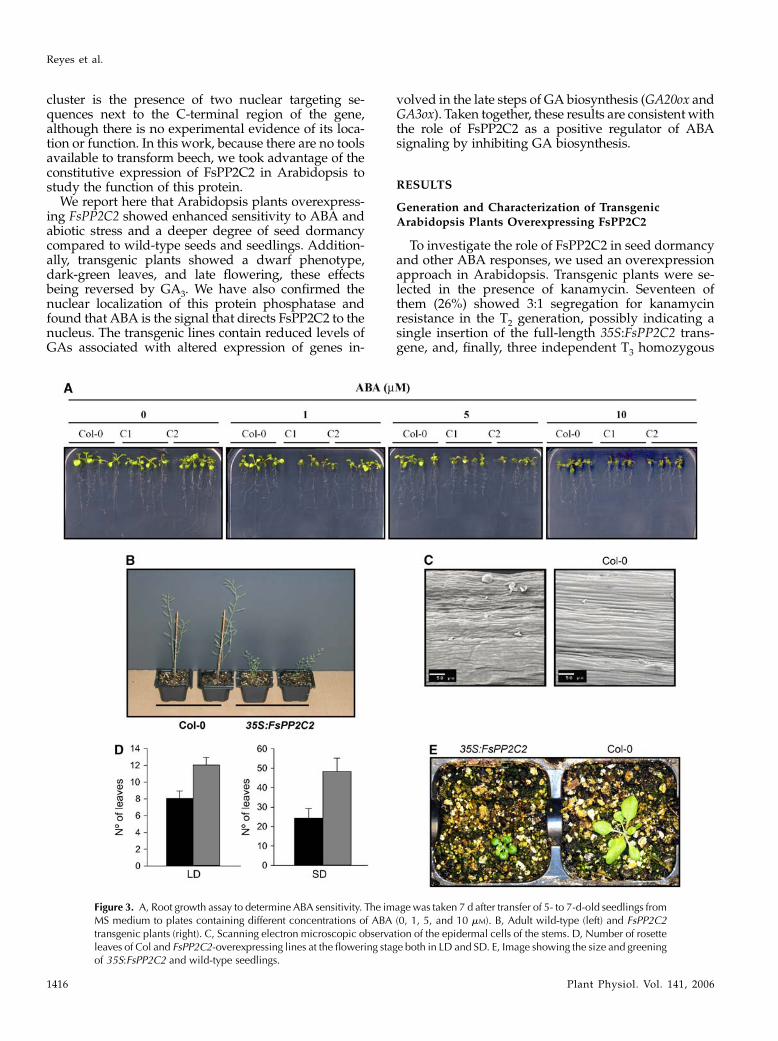
Figure 2. Effect of FsPP2C overexpression on dormancy and germination in Arabidopsis. A, Dormancy assay of FsPP2C2-overexpressing seeds. Germination percentage was determined 5 d after stratification at 4�C for 0, 24, and 120 h. B, Percentageof seeds that germinated and developed green cotyledons after 8 d following stratification at 4�C for 120 h, in 0.1, 0.2, and 0.3mM
ABA. C, Image showing differences in germination of FsPP2C2-overexpressing, wild-type seeds and seeds transformed with theempty vector pBI121 after 5 d on 0.1 mM ABA. D, Percentage of seeds that germinated and developed green cotyledons on 50 mM
NaCl. E, Percentage of seeds that germinated and developed green cotyledons in 100 mM mannitol. C1, C2, and C3, Transgenic35:FsPP2C2 lines; pBI121, transgenic line with the empty vector pBI121.
PP2C Related to Abscisic Acid Responses and GA Biosynthesis
Plant Physiol. Vol. 141, 2006 1415
cluster is the presence of two nuclear targeting se-quences next to the C-terminal region of the gene,although there is no experimental evidence of its loca-tion or function. In this work, because there are no toolsavailable to transform beech, we took advantage of theconstitutive expression of FsPP2C2 in Arabidopsis tostudy the function of this protein.
We report here that Arabidopsis plants overexpress-ing FsPP2C2 showed enhanced sensitivity to ABA andabiotic stress and a deeper degree of seed dormancycompared to wild-type seeds and seedlings. Addition-ally, transgenic plants showed a dwarf phenotype,dark-green leaves, and late flowering, these effectsbeing reversed by GA3. We have also confirmed thenuclear localization of this protein phosphatase andfound that ABA is the signal that directs FsPP2C2 to thenucleus. The transgenic lines contain reduced levels ofGAs associated with altered expression of genes in-
volved in the late steps of GA biosynthesis (GA20ox andGA3ox). Taken together, these results are consistent withthe role of FsPP2C2 as a positive regulator of ABAsignaling by inhibiting GA biosynthesis.
RESULTS
Generation and Characterization of Transgenic
Arabidopsis Plants Overexpressing FsPP2C2
To investigate the role of FsPP2C2 in seed dormancyand other ABA responses, we used an overexpressionapproach in Arabidopsis. Transgenic plants were se-lected in the presence of kanamycin. Seventeen ofthem (26%) showed 3:1 segregation for kanamycinresistance in the T2 generation, possibly indicating asingle insertion of the full-length 35S:FsPP2C2 trans-gene, and, finally, three independent T3 homozygous
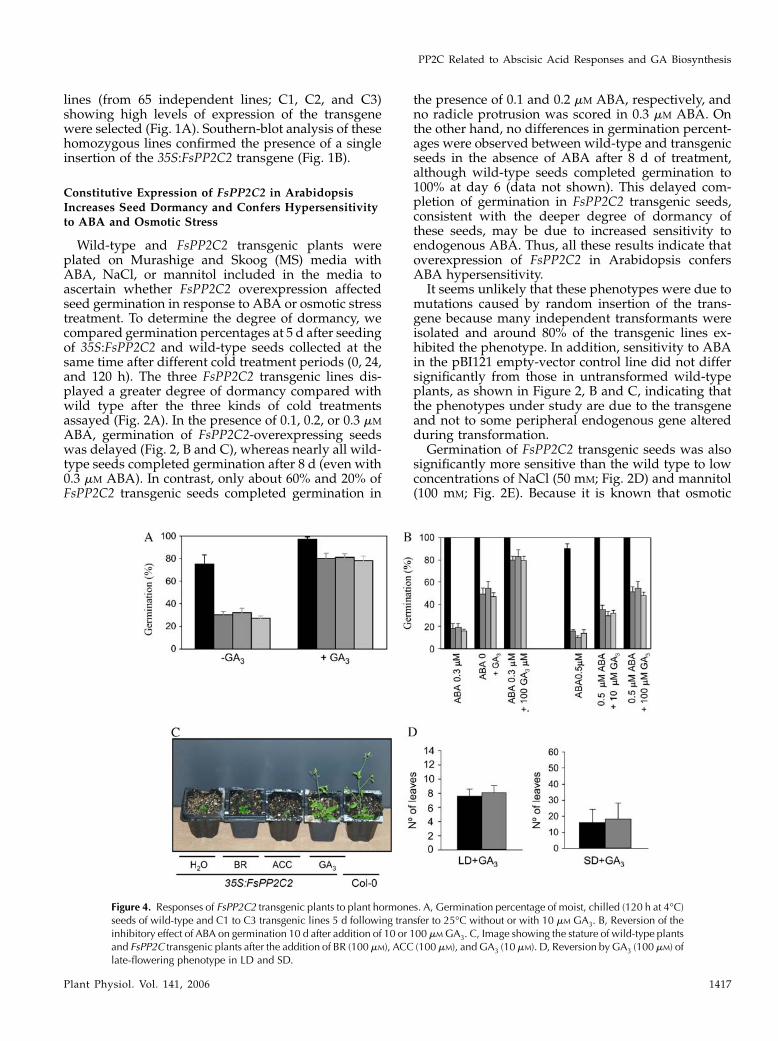
Figure 3. A, Root growth assay to determine ABA sensitivity. The image was taken 7 d after transfer of 5- to 7-d-old seedlings fromMS medium to plates containing different concentrations of ABA (0, 1, 5, and 10 mM). B, Adult wild-type (left) and FsPP2C2transgenic plants (right). C, Scanning electron microscopic observation of the epidermal cells of the stems. D, Number of rosetteleaves of Col and FsPP2C2-overexpressing lines at the flowering stage both in LD and SD. E, Image showing the size and greeningof 35S:FsPP2C2 and wild-type seedlings.
Reyes et al.
1416 Plant Physiol. Vol. 141, 2006
lines (from 65 independent lines; C1, C2, and C3)showing high levels of expression of the transgenewere selected (Fig. 1A). Southern-blot analysis of thesehomozygous lines confirmed the presence of a singleinsertion of the 35S:FsPP2C2 transgene (Fig. 1B).
Constitutive Expression of FsPP2C2 in Arabidopsis
Increases Seed Dormancy and Confers Hypersensitivityto ABA and Osmotic Stress
Wild-type and FsPP2C2 transgenic plants wereplated on Murashige and Skoog (MS) media withABA, NaCl, or mannitol included in the media toascertain whether FsPP2C2 overexpression affectedseed germination in response to ABA or osmotic stresstreatment. To determine the degree of dormancy, wecompared germination percentages at 5 d after seedingof 35S:FsPP2C2 and wild-type seeds collected at thesame time after different cold treatment periods (0, 24,and 120 h). The three FsPP2C2 transgenic lines dis-played a greater degree of dormancy compared withwild type after the three kinds of cold treatmentsassayed (Fig. 2A). In the presence of 0.1, 0.2, or 0.3 mM
ABA, germination of FsPP2C2-overexpressing seedswas delayed (Fig. 2, B and C), whereas nearly all wild-type seeds completed germination after 8 d (even with0.3 mM ABA). In contrast, only about 60% and 20% ofFsPP2C2 transgenic seeds completed germination in
the presence of 0.1 and 0.2 mM ABA, respectively, andno radicle protrusion was scored in 0.3 mM ABA. Onthe other hand, no differences in germination percent-ages were observed between wild-type and transgenicseeds in the absence of ABA after 8 d of treatment,although wild-type seeds completed germination to100% at day 6 (data not shown). This delayed com-pletion of germination in FsPP2C2 transgenic seeds,consistent with the deeper degree of dormancy ofthese seeds, may be due to increased sensitivity toendogenous ABA. Thus, all these results indicate thatoverexpression of FsPP2C2 in Arabidopsis confersABA hypersensitivity.
It seems unlikely that these phenotypes were due tomutations caused by random insertion of the trans-gene because many independent transformants wereisolated and around 80% of the transgenic lines ex-hibited the phenotype. In addition, sensitivity to ABAin the pBI121 empty-vector control line did not differsignificantly from those in untransformed wild-typeplants, as shown in Figure 2, B and C, indicating thatthe phenotypes under study are due to the transgeneand not to some peripheral endogenous gene alteredduring transformation.
Germination of FsPP2C2 transgenic seeds was alsosignificantly more sensitive than the wild type to lowconcentrations of NaCl (50 mM; Fig. 2D) and mannitol(100 mM; Fig. 2E). Because it is known that osmotic
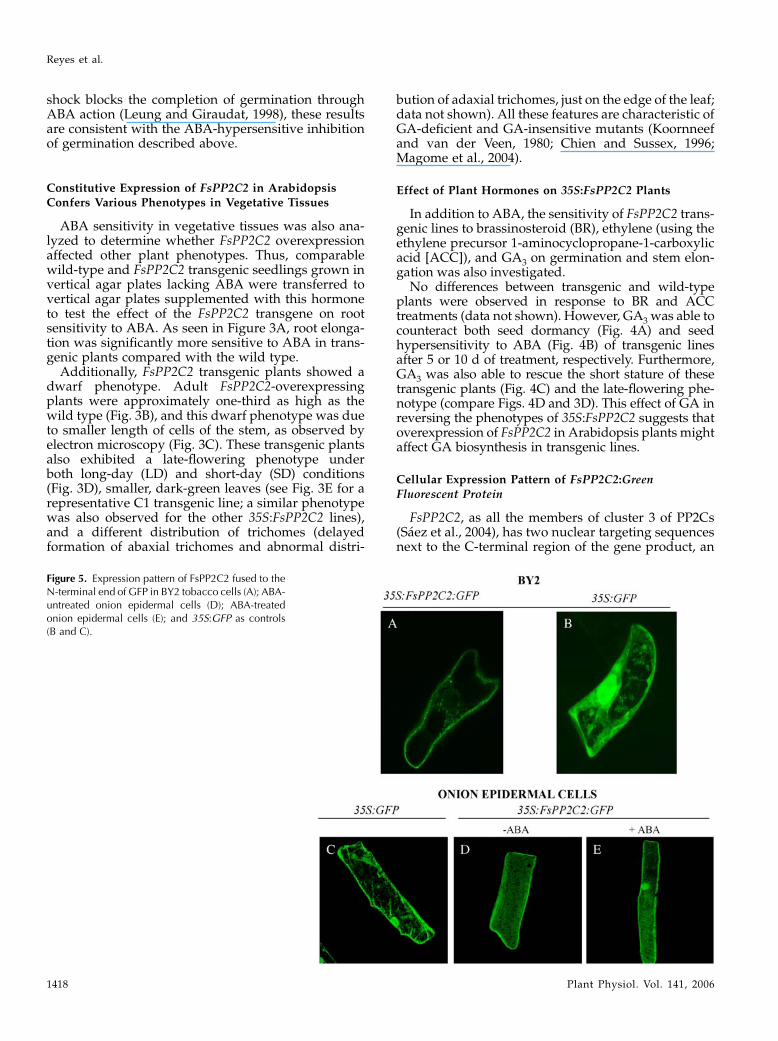
Figure 4. Responses of FsPP2C2 transgenic plants to plant hormones. A, Germination percentage of moist, chilled (120 h at 4�C)seeds of wild-type and C1 to C3 transgenic lines 5 d following transfer to 25�C without or with 10 mM GA3. B, Reversion of theinhibitory effect of ABA on germination 10 d after addition of 10 or 100 mMGA3. C, Image showing the stature of wild-type plantsand FsPP2C transgenic plants after the addition of BR (100mM), ACC (100mM), and GA3 (10mM). D, Reversion byGA3 (100mM) oflate-flowering phenotype in LD and SD.
PP2C Related to Abscisic Acid Responses and GA Biosynthesis
Plant Physiol. Vol. 141, 2006 1417
shock blocks the completion of germination throughABA action (Leung and Giraudat, 1998), these resultsare consistent with the ABA-hypersensitive inhibitionof germination described above.
Constitutive Expression of FsPP2C2 in ArabidopsisConfers Various Phenotypes in Vegetative Tissues
ABA sensitivity in vegetative tissues was also ana-lyzed to determine whether FsPP2C2 overexpressionaffected other plant phenotypes. Thus, comparablewild-type and FsPP2C2 transgenic seedlings grown invertical agar plates lacking ABA were transferred tovertical agar plates supplemented with this hormoneto test the effect of the FsPP2C2 transgene on rootsensitivity to ABA. As seen in Figure 3A, root elonga-tion was significantly more sensitive to ABA in trans-genic plants compared with the wild type.
Additionally, FsPP2C2 transgenic plants showed adwarf phenotype. Adult FsPP2C2-overexpressingplants were approximately one-third as high as thewild type (Fig. 3B), and this dwarf phenotype was dueto smaller length of cells of the stem, as observed byelectron microscopy (Fig. 3C). These transgenic plantsalso exhibited a late-flowering phenotype underboth long-day (LD) and short-day (SD) conditions(Fig. 3D), smaller, dark-green leaves (see Fig. 3E for arepresentative C1 transgenic line; a similar phenotypewas also observed for the other 35S:FsPP2C2 lines),and a different distribution of trichomes (delayedformation of abaxial trichomes and abnormal distri-
bution of adaxial trichomes, just on the edge of the leaf;data not shown). All these features are characteristic ofGA-deficient and GA-insensitive mutants (Koornneefand van der Veen, 1980; Chien and Sussex, 1996;Magome et al., 2004).
Effect of Plant Hormones on 35S:FsPP2C2 Plants
In addition to ABA, the sensitivity of FsPP2C2 trans-genic lines to brassinosteroid (BR), ethylene (using theethylene precursor 1-aminocyclopropane-1-carboxylicacid [ACC]), and GA3 on germination and stem elon-gation was also investigated.
No differences between transgenic and wild-typeplants were observed in response to BR and ACCtreatments (data not shown). However, GA3 was able tocounteract both seed dormancy (Fig. 4A) and seedhypersensitivity to ABA (Fig. 4B) of transgenic linesafter 5 or 10 d of treatment, respectively. Furthermore,GA3 was also able to rescue the short stature of thesetransgenic plants (Fig. 4C) and the late-flowering phe-notype (compare Figs. 4D and 3D). This effect of GA inreversing the phenotypes of 35S:FsPP2C2 suggests thatoverexpression of FsPP2C2 in Arabidopsis plants mightaffect GA biosynthesis in transgenic lines.
Cellular Expression Pattern of FsPP2C2:GreenFluorescent Protein
FsPP2C2, as all the members of cluster 3 of PP2Cs(Saez et al., 2004), has two nuclear targeting sequencesnext to the C-terminal region of the gene product, an
Figure 5. Expression pattern of FsPP2C2 fused to theN-terminal end of GFP in BY2 tobacco cells (A); ABA-untreated onion epidermal cells (D); ABA-treatedonion epidermal cells (E); and 35S:GFP as controls(B and C).
Reyes et al.
1418 Plant Physiol. Vol. 141, 2006

unusual feature among PP2Cs (Lorenzo et al., 2002).However, no experimental evidence on the subcellularlocalization of this group of PP2Cs has been described.Thus, to verify the possible nuclear localization of thisphosphatase, the FsPP2C2 coding region was fused tothe C- or N-terminal end of the green fluorescentprotein (GFP) under the control of the cauliflowermosaic virus (CaMV) 35S promoter and transientexpression assays were developed in BY2 tobacco(Nicotiana tabacum) cells and in onion (Allium cepa)epidermal cells. The results obtained with the twokinds of constructs were similar, so only picturescorresponding to the construct of the coding regionof FsPP2C2 fused to the N-terminal end of the GFP arepresented (Fig. 5). The results show that, in spite of thetwo nuclear targeting sequences present in FsPP2C2,this protein did not migrate to the nucleus in BY2tobacco (Fig. 5A). This was confirmed in onion epi-dermal cells transformed with FsPP2C2:GFP, where, asin the case of tobacco cells, cytoplasmic distribution ofthe protein was observed in more than 90% of un-treated cells (Fig. 5D). However, ABA applicationinduced migration of FsPP2C2 to the nucleus inmore than 85% of ABA-treated onion cells (Fig. 5E).On the other hand, when cells were treated with GA3,no nuclear localization was observed in any of the cells(data not shown). This indicates the involvement ofFsPP2C2 in ABA signaling because of the absence orpresence of this hormone is a signal that modulates thelocalization of this protein in the nucleus.
Quantification of GAs
Because many of the effects observed in 35S:FsPP2C2 plants were reversed by GAs, we quantifiedthe levels of endogenous GAs in 4-week-old trans-genic and wild-type plants to determine whether GAmetabolism was altered in FsPP2C2-overexpressinglines. As shown in Table I, the levels of the interme-
diate GA19 and the end products GA20 (in the earlyC-13 hydroxylation pathway) and GA9 (in the non-C-13 hydroxylation pathway) of GA 20-oxidase activ-ity were not significantly altered in our transgeniclines with respect to wild-type plants. However, thelevels of GA4 (the main active GA in Arabidopsis) andof GA34 (the inactive metabolite of GA4) were reducedto about 60% in the transgenic lines. Interestingly, thelevels of GA1 and GA8 (the active GA and its inactivemetabolite in the early C-13 hydroxylation pathway)were also reduced in the transgenic lines, as well asGA29 (the inactive metabolite of GA20). These resultsshow that the decrease of bioactive GAs in FsPP2C2transgenic plants was probably due to lower GA3-oxidase activity.
Expression of GA20ox, GA3ox, and a GASA1 Gene
Expression of two genes involved in the last stepsof GA biosynthesis, GA5 (AtGA20ox1, encoding a GA20-oxidase; Xu et al., 1995) and GA4 (AtGA3ox1, en-coding a GA 3-oxidase; Williams et al., 1998), keyenzymes in controlling active GA biosynthesis, werealso analyzed. Transcripts of AtGA20ox1 were notdetected in the wild type but were very abundant intransgenic seedlings (Fig. 6A). Slightly higher levels ofAtGA3ox1 transcripts were also observed in transgenicseedlings, as compared with the wild type (Fig. 6A).Expression of many GA20ox and GA3ox genes indifferent species is subjected to negative feedbackregulation (for review, see Olszewski et al., 2002).Thus, the increase of AtGA20ox1 and AtGA3ox1 tran-scripts found in our transgenic lines is probably aconsequence of the reduction in the level of active GAsdescribed before. To check whether the GA signaltransduction pathway was affected in FsPP2C2-over-expressing lines, we also analyzed the expressionof GASA1, a GA-induced gene (Herzog et al., 1995),in both GA-treated and untreated transgenic and
Figure 6. A, Expression of AtGA20ox1 (GA5) andAtGA3ox1 (GA4) genes. B, GASA1 gene expressionin FsPP2C2-overexpressing plants (C1 and C2) com-pared to wild type. mRNA levels of the indicatedgenes were determined by semiquantitative RT-PCR(A) or northern-blot analysis (B) using total RNAs(10 mg/line) isolated from 7-d-old seedlings. Bottom,Ubiquitin (A) and ethidium bromide-stained gelshowing rRNAs (B) as controls.
Table I. GA amount (ng g21 fresh weight) in Arabidopsis plants from wild-type and FsPP2C2 transgenic lines (C1 and C2)
Levels of active GAs (GA1 and GA4) are in bold. Values are means of three biological replicates6SE, except in the case of GA8 (only two replicates).
GA19 GA20 GA29 GA1 GA8 GA9 GA4 GA34
Wild type 0.47 6 0.13 0.23 6 0.03 0.20 6 0.07 1.06 6 0.26 1.00 0.22 6 0.07 1.25 6 0.18 0.68 6 0.14C1 0.51 6 0.11 0.17 6 0.03 0.09 6 0.07 0.83 6 0.04 0.08 0.29 6 0.10 0.71 6 0.10 0.43 6 0.10C2 0.40 6 0.12 0.13 6 0.02 0.04 6 0.02 0.45 6 0.14 0.10 0.15 6 0.07 0.73 6 0.13 0.42 6 0.06
PP2C Related to Abscisic Acid Responses and GA Biosynthesis
Plant Physiol. Vol. 141, 2006 1419

wild-type plants. As seen in Figure 6B, expression ofthis gene is up-regulated in our transgenic lines afterGA treatment, although such up-regulation was not soclear in wild-type plants, as previously described(Herzog et al., 1995; Aubert et al., 1998). This resultindicates that the mechanism of GA action is notaffected in transgenic lines overexpressing FsPP2C2.
Expression of an ABA-Responsive Gene
We compared the expression of the RAB18 gene, aknown ABA-responsive gene (Lang and Palva, 1992)in 35S:FsPP2C2 plants, with that of Columbia (Col-0)to examine whether the enhanced ABA sensitivity intransgenic plants was accompanied by altered expres-sion of ABA-responsive genes. The hyperinduction ofthis ABA-responsive gene further confirms that thePP2C from beech is involved in ABA signal transduc-tion. RAB18 transcripts were not detected in wild-typeand untreated transgenic lines (Fig. 7, lanes 1–3), butwere induced by ABA treatment, much more so intransgenic that in wild-type lines (Fig. 7, lanes 4–6).
DISCUSSION
Only six PP2Cs are found in the yeast (Saccharomycescerevisiae) genome (Stark, 1996) and no more than 15 inthe human genome (Cheng et al., 2000). In contrast, thehigh number of PP2C genes (at least 69) present inthe Arabidopsis genome denotes the importance ofthe PP2C class of Ser-Thr phosphatases in plants (Kerket al., 2002) and suggests that plant PP2Cs have anarrower substrate specificity than other organisms(Merlot et al., 2001).
One plant PP2C family (cluster 5; Saez et al., 2004) hasbeen identified as constituted by components of ABAsignaling. For instance, ABI1, ABI2, AtPP2CA, HAB1,and FsPP2C1 act as negative regulators of the ABAsignal transduction cascade (Gosti et al., 1999; Merlotet al., 2001; Tahtiharju and Palva, 2001; Gonzalez-Garcıaet al., 2003; Saez et al., 2004; Kuhn et al., 2006; Yoshidaet al., 2006). Other members of PP2C, such as MP2C(cluster 6; Saez et al., 2004) from ice plant (Mesembry-anthemum crystallinum), act as negative regulators of themitogen-activated protein kinase pathway activated bysalt stress or wounding (Meskiene et al., 1998, 2003).
Interestingly, all these PP2Cs act as negative regulatorsin different signal transduction cascades. In fact, exceptfor the recent controversy on the role of ABI1 from thework of Wu et al. (2003), all the PP2Cs described andcharacterized so far would act as negative regulators indifferent signal transduction cascades.
We have previously isolated FsPP2C2, a PP2C frombeech that is up-regulated by ABA in seeds andvegetative tissues (Lorenzo et al., 2002), making it areasonable candidate as a regulator of ABA signaling.However, genetic evidence of the role of this phos-phatase was lacking. Studies of orthologous genes andfunctional tests in heterologous systems have shownthat the ABA signal transduction cascade is essentiallyconserved among evolutionarily distant plant species(for review, see Finkelstein et al., 2002). Therefore, inthis study, we used an overexpression approach inArabidopsis to investigate FsPP2C2 function. It wasexpected that if FsPP2C2 is a negative regulator ofABA responses, as is another PP2C described in beech(FsPP2C1; Lorenzo et al., 2001; Gonzalez-Garcıa et al.,2003), the overexpression of FsPP2C2 in Arabidopsiswould lead to ABA insensitivity in seeds and, conse-quently, to reduced dormancy. Surprisingly, 35S:FsPP2C2plants exhibited an enhanced response to ABA andosmotic stress (Figs. 2 and 3) together with otherphenotypes, such as dwarfism, late flowering, smallerand dark-green leaves (Fig. 3), and abnormal trichomedistribution (data not shown), all these typical
Figure 8. Proposed model for the role of FsPP2C2 in ABA signalingthrough a reduction of GA biosynthesis.
Figure 7. Expression of the RAB18 ABA-regulated gene in FsPP2C2-overexpressing plants (C1 and C2) compared to wild type. mRNA levelsof the indicated genes were determined by northern-blot analysis usingtotal RNAs (10 mg/line) isolated from mock-treated (2) or ABA-treated(150 mM ABA for 24 h) plants. Bottom, Ethidium bromide-stained gelshowing rRNAs.
Reyes et al.
1420 Plant Physiol. Vol. 141, 2006

characteristics of GA-deficient and/or insensitive mu-tants (Koornneef and van der Veen, 1980; Magomeet al., 2004). Furthermore, these phenotypes, includ-ing enhanced response to ABA, were reversed by GA3application (Fig. 4).In addition to the genes ABI1, ABI2, PP2CA, HAB1,
and FsPP2C1 (Gosti et al., 1999; Merlot et al., 2001;Tahtiharju and Palva, 2001; Gonzalez-Garcıa et al.,2003; Saez et al., 2004; Kuhn et al., 2006; Yoshida et al.,2006) encoding PP2C, other genes have been identifiedas negative regulators of the ABA-signaling pathway,such as the farnesyl transferase b-subunit (ERA1;Cutler et al., 1996), the mRNA cap-binding protein(ABH1; Hugouvieux et al., 2001), the homeodomainprotein 6 (ATHB6; Himmelbach et al., 2002), and thecalcium sensor CBL-9 (Pandey et al., 2004). Moreover,ABI3, ABI4, and ABI5 encode different types of tran-scription factors (Giraudat et al., 1992; Finkelstein et al.,1998; Finkelstein and Lynch, 2000) known to act aspositive regulators of ABA-mediated regulation ofseed development, germination, and early seedlinggrowth (Finkelstein et al., 2002; Arroyo et al., 2003).However, FsPP2C2 is described as a positive regulatorof the ABA signal transduction pathway in seeds andother vegetative tissues. Evidence of the role of thisPP2C on ABA action was obtained from the observa-tion that plants overexpressing FsPP2C2 had hyper-sensitivity to ABA and osmotic stress and a deeperdegree of dormancy compared with wild-type plants(Figs. 2 and 3). Expression of the ABA-responsive geneRAB18 was also increased in 35S:FsPP2C2 plants (Fig.7). Moreover, the fact that FsPP2C2 is localized in thenucleus only in the presence of ABA (Fig. 5) stronglysupports its involvement in ABA signaling.Evidence of the mode of action of FsPP2C2 in ABA
responses was initially obtained from the observationthat GA3 was able to rescue the poor germination,dwarfism, and delay in flowering of transgenic plantsoverexpressing FsPP2C2 (Fig. 4). Because GA actiondid not seem to be affected in these plants (Fig. 6B),this indicated that FsPP2C2 might exert its action bymodulating GA metabolism. Quantification of endog-enous GAs showed that the levels of active GAs (GA4and GA1) were significantly reduced in transgeniclines, whereas the contents of their immediate meta-bolic precursors (GA9 and GA20, respectively) were notaffected (Table I). The low reduction of GA4 content(about 50%) in transgenic lines displaying phenotypeagrees with the low increase of active GA4 (2- to 3-fold)in transgenic Arabidopsis overexpressing GA20ox.This is in contrast with the 10-fold increases in appliedGA3 needed to get partial reversal of germinationphenotypes in the presence of ABA (Fig. 4B). However,this is similar to what happens in pea (Pisum sativum)ovary, where small differences in active GA content(within 1 order of magnitude) have very significantphysiological effects, whereas higher doses of GA1(orders of magnitude) have to be applied exogenouslyto produce a similar effect probably due to transportand/or metabolism problems (Rodrigo et al., 1997).
Our results suggest that overexpression of FsPP2C2decreased GA 3-oxidase activity. The lower content ofinactive metabolites of GA4 and GA1 (GA34 and GA8,respectively) also supports this conclusion, whileshowing that the decrease of active GA content wasnot the result of higher inactivating metabolism. In-terestingly, transcript levels of AtGA3ox1 (gene GA4)were not reduced, but rather increased in transgeniclines (Fig. 6A). An even more dramatic effect wasobserved on AtGA20ox1 (gene GA5) transcript levels(Fig. 6A). It is known that expression of many GA20oxand GA3ox genes is controlled by active GAs throughnegative feedback regulation (Olszewski et al., 2002).Therefore, the increase of GA5 and GA4 transcripts isprobably a result of this kind of regulation and we canpostulate that the regulation of GA 3-oxidase activityby FsPP2C2 is probably carried out at the posttransla-tional level.
In conclusion, the results presented in this work areconsistent with the role of the ABA-induced FsPP2C2as a positive regulator of ABA signaling through areduction of GA biosynthesis, probably affecting GA3-oxidase activity (Fig. 8). In this proposed model, wehypothesize that a phosphorylated protein in thenucleus may activate an unknown element responsiblefor the activation of GA 3-oxidase, but ABA actionthrough FsPP2C2 would dephosphorylate this pro-tein, leading to inactivation of this key enzyme in GAbiosynthesis. Thus, a potential interaction betweenABA action and GA biosynthesis is described, show-ing another junction in the complex mechanism ofhormone signaling. The data presents new insightsinto future research about the antagonistic effect ofABA and GAs in plant development.
MATERIALS AND METHODS
Plant Materials
Arabidopsis (Arabidopsis thaliana) plants, ecotype Col-0, were used in this
research. They were normally grown in a growth chamber with 40% humidity,
at 22�C, under SD (8 h light/16 h dark) or LD (16 h light/8 h dark; light
intensity of 80–100 mE m22 s21) in pots containing a 1:3 vermiculite:soil
mixture, as described previously (Gonzalez-Garcıa et al., 2003). For in vitro
culture, seeds were surface sterilized in 70% (v/v) ethanol solution containing
1% (v/v) Triton X-100 and washed four times in sterile distilled water.
Stratification of seeds was conducted during 3 d at 4�C, unless otherwise
indicated. Afterward, seeds were sowed on MS plates (Murashige and Skoog,
1962) containing solid medium composed of MS basal salts and 1% (w/v) Suc,
pH 5.7, solidified with 1% (w/v) agar. Plates were sealed and incubated in a
controlled-environment growth chamber.
Vector Construction and Plant Transformation
To produce transgenic plants, the coding region of the FsPP2C2 cDNAwas
cloned into the pBIN121 vector, which contains the modified CaMV 35S
promoter. 5#-GGATCCATGTTTTCGGA-3# (sense) and 5#- GAGCTCTTAGG-
TGCTGCCAAC-3# (antisense), containing the BamHI and SacI cloning sites,
were used as primers to amplify FsPP2C2 cDNA and subcloned in the
corresponding sites of the pBIN121 vector. The pBIN121-FsPP2C2 construct,
after confirmation of the nucleotide sequence, was introduced into Agro-
bacterium tumefaciens C58C1 (pGV2260; Deblaere et al., 1985) by heat shock.
Arabidopsis plants (Col-0 ecotype) were transformed by the floral-dip method
(Clough and Bent, 1998) and transgenic seedlings were selected on kanamycin
PP2C Related to Abscisic Acid Responses and GA Biosynthesis
Plant Physiol. Vol. 141, 2006 1421

medium (50 mg mL21). T2 plants that produced 100% kanamycin-resistant
plants in the T3 generation were considered homozygous for the selection
marker and used for further studies. Nontransformed plants and transgenic
plants transformed with the empty vector pBI121 were used as controls.
From 65 kanamycin-resistant independent lines rescued, around 80%
showed an ABA-hypersensitive phenotype in seed germination compared
with controls that did not display any phenotypes related to ABA responses.
Germination Assays
Seeds were plated on solid medium composed of MS basal salts, 1% (w/v)
Suc, and different concentrations of ABA (Sigma-Aldrich; 0.1, 0.2, 0.3 mM) to
determine ABA sensitivity. For the dormancy assay, seed lots to be compared
were harvested on the same day from individual plants grown in identical
conditions and were stratified during 0, 1, and 5 d at 4�C.To determine sensitivity to inhibition of germination by osmotic stress, the
medium was complemented with low concentrations of mannitol (100 mM) or
NaCl (50 mM).
Reversion of the inhibitory effect of ABA in seed germination was carried
out using media containing 10 and 100 mM GA3 (Sigma-Aldrich).
Seed germination was scored by determining daily the percentage of seeds
that had germinated and developed green cotyledons for 5 to10 d. At least
three replicates of each germination assay were performed.
Root Growth and Flowering Time
Root growth assay for scoring ABA sensitivity was carried out by mea-
suring root growth 7 d after transferring 5- to 7-d-old seedlings onto vertical
MS plates containing different concentrations of ABA (0, 1, 5, and 10 mM). The
leaf number used to determine flowering phenotype was performed as
described by Wilson et al. (1992).
Hormone Dose Response Experiments
Ten-day-old seedlings were transferred to pots containing a 1:3 vermiculite:
soil mixture. The seedlings were sprayed every 2 d with GA3 (100 mM), BR
(Epibrassinolide; 10 mM), or ACC (100 mM; Sigma-Aldrich).
Northern-Blot Analysis
Total RNA was extracted using the RNA wiz kit (Ambion) following the
manufacturer’s protocol, separated on formaldehyde-agarose gels, and blot-
ted onto a nylon membrane. Blots were hybridized with [32P]-labeled specific
probes. All RNA gel-blot experiments were repeated at least three times, and
results from one representative experiment are shown in the figures. RAB18
and GASA1 probes were prepared by reverse transcription (RT)-PCR with the
primers described elsewhere (Herzog et al., 1995; Gonzalez-Guzman et al.,
2002). Blots were exposed for 24 h in a phosphor imager screen (Fujitsu).
Semiquantitative RT-PCR
cDNA was synthesized from total RNA using the first-strand cDNA
synthesis kit for RT-PCR of Avian myeloblastosis virus (Roche Diagnostics) with
oligo-p(dT) as primer, following the manufacturer’s instruction. Two micro-
liters were used for each PCR consisting of 1-min cycles at 94�C, 50�C, and72�C. AtGA20ox1, AtGA3ox1, and UBQ14 primers were those used by Oka
et al. (2001), Williams et al. (1998), and Vriezen et al. (2004), respectively. Each
PCR was performed for 15, 20, 25, and 30 cycles following the procedure
described by Vriezen et al. (2004). After Southern blotting and hybridizing
with the purified PCR products, the signals were quantified by phosphor
imager analysis (Fujitsu). The amplification reactions were exponential be-
tween 15 and 25 cycles, and the products of the reactions after 20 cycles were
used for quantification. The experiment was performed twice and produced
reproducible patterns.
FsPP2C2:GFP Constructs
The FsPP2C2 coding region was fused to the N-terminal end of GFP in the
vector pMON30063 (Pang et al., 1996), under the control of the CaMV 35S
promoter. A fragment corresponding to the FsPP2C2 coding region was ampli-
fied by PCR using synthetic anchored primers that contained aNcoI-compatible
restriction site at their 3# and 5# ends. In addition, the coding region of FsPP2C2
was also fused to the C-terminal end of GFP in the same vector, although in this
case the subsequent cloning required a different strategy. The pMON vector was
digested with EcoRI restriction enzyme, whereas FsPP2C2 was digested with
NdeI and XhoI restriction enzymes because its coding region contained an EcoRI
restriction site. Both the linearized vector and the DNA fragment containing
FsPP2C2 coding regions were treated with the Klenow fragment of DNA
polymerase in order to obtain blunt ends before ligation. Following sequence
verification of the inserts, GFP fusion and control constructs were transiently
expressed by particle bombardment.
Transient Expression Assays
Transformation was carried out following the procedure described by
Varagona et al. (1992). DNA absorption to gold particles and bombardment
using a helium-driven particle accelerator (PDS-1000/He; Bio-Rad) was
performed according to the manufacturer’s recommendations. One micro-
gram of plasmid was used for transformation, and the target material (BY2
tobacco [Nicotiana tabacum] and onion [Allium cepa] epidermal cells) were
bombarded three times. After bombardment, onion epidermal cells were
transferred to MS medium supplemented or not with 100 mM ABA and kept at
28�C in the dark for 18 h before analysis by fluorescence microscopy. Cells
were examined using a Zeiss Axiovert 200 microscope equipped with
epifluorescence optics. GFP-specific fluorescence was detected using Zeiss
filters 515 to 530 nM. Confocal images were obtained on a Zeiss LSM 510
confocal laser-scanning microscope with Zeiss LSM version 2.8 software.
Scanning Electron Microscopy
Stems were harvested from mature plants and observed directly by a
scanning microscope (Zeiss DSM 940) equipped with a cooling stage.
Quantification of GAs
Endogenous GAs were quantified by gas chromatography-mass spectrom-
etry as described elsewhere (Garcıa-Martınez et al., 1997). The material (about
10 g fresh weight of rosette leaves from 4-week-old plants grown at 22�Cunder LD conditions, at the vegetative state, and collected 3 h after starting
the light period) was spiked at the time of extraction with [3H]GA20 (1.41
TBq mmol21) and [3H]GA9 (1.41 TBq mmol21) to check recoveries during the
purification procedure, and with [17-2H2]GA1, [17-2H2]GA4, [17-2H2]GA8,
[17-2H2]GA9, [17-2H2]GA19, [17-2H2]GA20, [17-2H2]GA29, and [17-2H2]GA34
(purchased from Prof. L. Mander, Research School of Chemistry, Australian
National University, Canberra, Australia) as internal standards for quantifi-
cation. Fractions of HPLC containing the GAs of interest were methylated and
trimethylsilylated before analysis by gas chromatography-mass spectrometry
using a Hewlett-Packard 5890 gas chromatograph coupled to a 5971A mass
selective detector. The peak-area ratios of the following ion pairs, in the
appropriate HPLC fractions and having a retention time similar to that of the
corresponding GAs, were determined to calculate the concentrations of
endogenous GAs by reference to calibration curves: 506/508 (GA1), 284/286
(GA4), 594/596 (GA8), 298/300 (GA9), 434/436 (GA19), 418/420 (GA20), 506/
508 (GA29), and 506/508 (GA34).
We only measured GA amounts in the transgenic C1 and C2 lines because
the size and production of seeds of the C3 line were not sufficient to perform
this set of experiments.
Sequence data from this article can be found in the GenBank/EMBL data
libraries under accession number AJ277744.
ACKNOWLEDGMENTS
We thank Dr. P.L. Rodriguez (Instituto de Biologıa Molecular y Celular de
Plantas-Consejo Superior de Investigaciones Cientificas, Valencia), for
providing us with Arabidopsis seeds transformed with the empty vector
pBI121, used as a control in this study.
Received June 8, 2006; revised June 8, 2006; accepted June 21, 2006; published
June 30, 2006.
Reyes et al.
1422 Plant Physiol. Vol. 141, 2006

LITERATURE CITED
Abe H, Urao T, Seki M, Shinozaki K, Yamaguchi-Shinozaki K (2003)
Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as tran-
scriptional activators in abscisic acid signaling. Plant Cell 15: 63–78
Arroyo A, Bossi F, Finkelstein RR, Leon P (2003) Three genes that affect
sugar sensing (abscisic acid insensitive 4, abscisic acid insensitive 5, and
constitutive triple response 1) are differentially regulated by glucose in
Arabidopsis. Plant Physiol 133: 231–242
Aubert D, Chevillard M, Dorne AM, Arlaud G, Herzog M (1998) Expres-
sion patterns of GASA genes in Arabidopsis thaliana: The GASA4 gene is
up-regulated by gibberellins in meristematic regions. Plant Mol Biol 36:
871–883
Calvo AP, Nicolas C, Nicolas G, Rodrıguez D (2004) Evidence of a cross-
talk regulation of a GA 20-oxidase (FsGA20ox1) by gibberellins and
ethylene during the breaking of dormancy in Fagus sylvatica seeds.
Physiol Plant 120: 623–630
Cheng A, Kaldis P, Solomon MJ (2000) Dephosphorylation of human
cyclin-dependent kinases by protein phosphatase type 2C alpha and
beta isoforms. J Biol Chem 275: 34744–34749
Chien JC, Sussex IM (1996) Differential regulation of trichome formation
on the adaxial and abaxial leaf surfaces by gibberellins and photoperiod
in Arabidopsis thaliana (L.) Heynh. Plant Physiol 111: 1321–1328
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacte-
rium-mediated transformation of Arabidopsis thaliana. Plant J 16:
735–743
Cutler S, Ghassemian M, Bonetta D, Cooney S, McCourt P (1996) A
protein farnesyl transferase involved in abscisic acid signal transduc-
tion in Arabidopsis. Science 273: 1239–1241
Deblaere R, Bytebier B, De Greve H, Deboeck F, Schell J, Van Montagu
M, Leemans J (1985) Efficient octopine Ti plasmid-derived vectors for
Agrobacterium-mediated gene transfer to plants. Nucleic Acids Res 13:
4777–4788
Finkelstein RR, Gampala SS, Rock CD (2002) Abscisic acid in seeds and
seedlings. Plant Cell 14: S15–S45
Finkelstein RR, Lynch TJ (2000) The abscisic acid response gene ABI5
encodes a basic leucine zipper transcription factor. Plant Cell 12: 599–610
Finkelstein RR, Wang ML, Lynch TJ, Rao S, Goodman HM (1998) The
Arabidopsis abscisic acid response locus ABI4 encodes an APETALA2
domain protein. Plant Cell 10: 1043–1054
Garcıa-Martınez JL, Santes C, Croker SJ, Hedden P (1997) Identification,
quantification and distribution of gibberellins in fruits of Pisum sativum
L. cv Alaska during pod development. Planta 184: 53–60
Giraudat J, Hauge BM, Valon C, Smalle J, Parcy F, Goodman HM (1992)
Isolation of the Arabidopsis ABI3 gene by positional cloning. Plant Cell
4: 1251–1261
Gomez-Cadenas A, Verhey SD, Holappa LD, Shen Q, Ho THD, Walker-
Simmons MK (1999) An abscisic acid induced protein kinase, PKABA1,
mediates abscisic acid-suppressed gene expression in barley aleurone.
Proc Natl Acad Sci USA 96: 1767–1772
Gomez-Cadenas A, Zentella R, Walker-Simmons MK, Ho THD (2001)
Gibberellin/abscisic acid antagonism in barley aleurone cells: site of
action of the protein kinase PKABA1 in relation to gibberellin signaling
molecules. Plant Cell 13: 667–679
Gonzalez-Garcıa MP, Rodrıguez D, Nicolas C, Rodrıguez PL, Nicolas G,
Lorenzo O (2003) Negative regulation of abscisic acid signaling by the
Fagus sylvatica FsPP2C1 plays a role in seed dormancy regulation and
promotion of seed germination. Plant Physiol 133: 135–144
Gonzalez-Guzman M, Apostolova N, Belles JM, Barrero JM, Piqueras P,
Ponce MR, Micol JL, Serrano R, Rodrıguez PL (2002) The short-chain
alcohol dehydrogenase ABA2 catalyzes the conversion of xanthoxin to
abscisic aldehyde. Plant Cell 14: 1833–1846
Gosti F, Beaudoin N, Serizet C, Webb AA, Vartanian N, Giraudat J (1999)
ABI1 protein phosphatase 2C is a negative regulator of abscisic acid.
Plant Cell 11: 1897–1910
Herzog M, Dorne AM, Grellet F (1995) GASA, a gibberellin regulated gene
family from Arabidopsis thaliana related to the tomato GAST1 gene.
Plant Mol Biol 27: 743–752
Himmelbach A, Hoffmann T, Leube M, Hohener B, Grill E (2002)
Homeodomain protein ATHB6 is a target of the protein phosphatase
ABI1 and regulates hormone responses in Arabidopsis. EMBO J 21:
3029–3038
Hugouvieux V, Kwak JM, Schroeder JI (2001) An mRNA cap binding
protein, ABH1, modulates early abscisic acid signal transduction in
Arabidopsis. Cell 106: 477–487
Kerk D, Bulgrien J, Smith DW, Barsam B, Veretnik S, Gribskov M (2002)
The complement of protein phosphatase catalytic subunits encoded in
the genome of Arabidopsis. Plant Physiol 129: 908–925
Kermode AR (2005) Role of abscisic acid in seed dormancy. J Plant Growth
Regul 24: 319–344
Koornneef M, van der Veen JH (1980) Induction and analysis of gibberellin-
sensitive mutants in Arabidopsis thaliana L. Theor Appl Genet 58: 257–263
Kucera B, Cohn MA, Leubner-Metzger G (2005) Plant hormone interac-
tions during seed dormancy release and germination. Seed Sci Res 15:
281–307
Kuhn JM, Boisson-Dernier A, Dizon MB, Maktabi MH, Schroeder JI
(2006) The protein phosphatase AtPP2CA negatively regulates abscisic
acid signal transduction in Arabidopsis, and effects of abh1 on AtPP2CA
mRNA. Plant Physiol 140: 127–139
Lang V, Palva ET (1992) The expression of a rab-related gene, rab18, is
induced by abscisic acid during the cold acclimation process of
Arabidopsis thaliana (L.) Heynh. Plant Mol Biol 20: 951–962
Lange T (1998) Molecular biology of gibberellin synthesis. Planta 204:
404–419
Leung J, Giraudat J (1998) Abscisic acid signal transduction. Annu Rev
Plant Physiol Plant Mol Biol 19: 199–222
Li J, Wang XQ, Watson MB, Assmann SM (2000) Regulation of abscisic
acid-induced stomatal closure and anion channels by guard cells AAPK
kinase. Science 287: 300–303
Lorenzo O, Nicolas C, Nicolas G, Rodrıguez D (2002) Molecular cloning of
a functional protein phosphatase 2C (FsPP2C2) with unusual features
and synergistically up-regulated by ABA and calcium in dormant seeds
of Fagus sylvatica. Physiol Plant 114: 482–490
Lorenzo O, Rodrıguez D, Nicolas G, Rodrıguez PL, Nicolas C (2001) A
new protein phosphatase 2C (FsPP2C1) induced by abscisic acid is
specifically expressed in dormant beechnut seeds. Plant Physiol 125:
1949–1956
Magome H, Yamaguchi S, Hanada A, Kamiya Y, Oda K (2004) dwarf and
delayed flowering 1, a novel Arabidopsis mutant deficient in gibberellin
biosynthesis because of overexpression of a putative AP2 transcription
factor. Plant J 37: 720–729
Merlot S, Gosti F, Guerrier D, Vavasseur A, Giraudat J (2001) The ABI1
and ABI2 protein phosphatases 2C act in a negative feedback regulatory
loop of the abscisic acid pathway. Plant J 25: 295–303
Meskiene I, Beaudoin E, Schweighofer A, Liwosz A, Jonak C, Rodrıguez
PL, Jelinek H, Hirt H (2003) The stress-induced protein phosphatase 2C
is a negative regulator of a mitogen-activated protein kinase. J Biol
Chem 278: 18945–18952
Meskiene I, Bogre L, Glaser W, Balog J, Brandstotter M, Zwerger K,
Ammerer G, Hirt H (1998) MP2C, a plant protein phosphatase 2C,
functions as a negative regulator of mitogen-activated protein kinase
pathways in yeast and plants. Proc Natl Acad Sci USA 95: 1938–1943
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco. Physiol Plant 15: 473–497
Mustilli AC, Merlot S, Vavasseur A, Fenzi F, Giraudat J (2002) Arabi-
dopsis OST1 protein kinase mediates the regulation of stomatal aperture
by abscisic acid and acts upstream of reactive oxygen species produc-
tion. Plant Cell 14: 3089–3099
Oka M, Tasaka Y, Iwabuchi M, Masanobu M (2001) Elevated sensitivity to
gibberellin by vernalization in the vegetative rosette plants of Eustoma
grandiflorum and Arabidopsis thaliana. Plant Sci 160: 1237–1245
Olszewski N, Sun TP, Gubler F (2002) Gibberellin: biosynthesis, catabo-
lism and response pathways. Plant Cell 14: S61–S80
Pandey GK, Cheong YH, Kim N, Grant JJ, Li L, Hung W, D’Angelo C,
Weinl S, Kudla J, Luan S (2004) The calcium sensor calcineurin B-like 9
modulates abscisic acid sensitivity and biosynthesis in Arabidopsis.
Plant Cell 16: 1912–1924
Pang SZ, DeBoer D, Wan Y, Ye C, Layton JC, Neher MK, Armstrong C, Fry
JE (1996) An improved green fluorescent protein gene as a vital marker
in plants. Plant Physiol 112: 893–900
Perez-Flores L, Carrari F, Osuna-Fernandez R, Rodrıguez MV, Enciso S,
Stanelloni R, Sanchez RA, Bottini R, Iusem ND, Benech-Arnold RL
(2003) Expression analysis of a GA 20-oxidase in embryos from
two sorghum lines with contrasting dormancy: possible participation
of this gene in the hormonal control of germination. J Exp Bot 54:
2071–2079
PP2C Related to Abscisic Acid Responses and GA Biosynthesis
Plant Physiol. Vol. 141, 2006 1423

Rodrigo MJ, Garcıa-Martınez JL, Santes CM, Gaskin P, Hedden P (1997)
The role of gibberellins A1 and A3 in fruit growth of Pisum sativum L.,
and the identification of gibberellins A4 and A7 in young seeds. Planta
201: 446–455
Saez A, Apostolova N, Gonzalez-Guzman M, Gonzalez-Garcıa MP,
Nicolas C, Lorenzo O, Rodrıguez PL (2004) Gain of function and loss
of function phenotypes of the protein phosphatase 2C HAB1 reveal its
role as a negative regulator of abscisic acid. Plant J 37: 354–369
Stark MJ (1996) Yeast protein serine/threonine phosphatases: multiple
roles and diverse regulation. Yeast 12: 1647–1675
Tahtiharju S, Palva T (2001) Antisense inhibition of protein phosphatase
2C accelerates cold acclimation in Arabidopsis thaliana. Plant J 26:
461–470
Varagona MJ, Schmidt RJ, Raikhel NV (1992) Nuclear localization sig-
nal(s) required for nuclear targeting of the maize regulatory protein
Opaque-2. Plant Cell 4: 1213–1227
Vriezen WH, Achard P, Harberd NP, van der Straeten D (2004) Ethylene-
mediated enhancement of apical hook formation in etiolated Arabidopsis
thaliana seedlings is gibberellin dependent. Plant J 37: 505–516
Williams J, Phillips AL, Gaskin P, Hedden P (1998) Function and substrate
specificity of the gibberellin 3b-hydroxylase encoded by the Arabidop-
sis GA4 gene. Plant Physiol 117: 559–563
Wilson RN, Heckman JW, Somerville CR (1992) Gibberellin is required for
flowering inArabidopsis thaliana under short days. Plant Physiol 100: 403–408
Wu Y, Sanchez JP, Lopez-Molina L, Himmelbach A, Grill E, Chua NH
(2003) The abi1-1 mutation blocks ABA downstream cADPR action.
Plant J 34: 307–315
Xu YL, Li L, Wu K, Peeters AJ, Gage DA, Zeevaart JAD (1995) The GA5
locus of Arabidopsis thaliana encodes a multifunctional gibberellin
20-oxidase: molecular cloning and functional expression. Proc Natl
Acad Sci USA 92: 6640–6644
Yamauchi Y, Ogawa M, Kuwahara A, Hanada A, Kamiya Y, Yamaguchi S
(2004) Activation of gibberellin biosynthesis and response pathways by
low temperature during imbibition of Arabidopsis thaliana seeds. Plant
Cell 16: 367–378
Yoshida T, Nishimura N, Kitahata N, Kuromori T, Ito T, Asami T,
Shinozaki K, Hirayama T (2006) ABA-Hypersensitive Germination3 en-
codes a protein phosphatase 2C (AtPP2CA) that strongly regulates
abscisic acid signaling during germination among Arabidopsis protein
phosphatase 2Cs. Plant Physiol 140: 115–126
Reyes et al.
1424 Plant Physiol. Vol. 141, 2006




















