
Isolation and Characterization of the Gibberellin Biosynthetic Gene Cluster in Sphaceloma...
15
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2008, p. 5325–5339 Vol. 74, No. 17 0099-2240/08/$08.000 doi:10.1128/AEM.00694-08 Copyright © 2008, American Society for Microbiology. All Rights Reserved. Isolation and Characterization of the Gibberellin Biosynthetic Gene Cluster in Sphaceloma manihoticola † Christiane Bo ¨mke, 1 Maria Cecilia Rojas, 2 Fan Gong, 3 Peter Hedden, 3 and Bettina Tudzynski 1 * Westfa ¨lische Wilhelms-Universita ¨t Mu ¨nster, Institut fu ¨r Botanik, Schlossgarten 3, 48149 Mu ¨nster, Germany 1 ; Laboratorio de Bioorga ´nica, Departamento de Quı ´mica, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile 2 ; and Rothamsted Research, Harpenden, Hertfordshire AL5 2JQ, United Kingdom 3 Received 25 March 2008/Accepted 14 June 2008 Gibberellins (GAs) are tetracyclic diterpenoid phytohormones that were first identified as secondary metabolites of the fungus Fusarium fujikuroi (teleomorph, Gibberella fujikuroi). GAs were also found in the cassava pathogen Sphaceloma manihoticola, but the spectrum of GAs differed from that in F. fujikuroi. In contrast to F. fujikuroi, the GA biosynthetic pathway has not been studied in detail in S. manihoticola, and none of the GA biosynthetic genes have been cloned from the species. Here, we present the identification of the GA biosynthetic gene cluster from S. manihoticola consisting of five genes encoding a bifunctional ent-copalyl/ent-kaurene synthase (CPS/KS), a pathway- specific geranylgeranyl diphosphate synthase (GGS2), and three cytochrome P450 monooxygenases. The functions of all of the genes were analyzed either by a gene replacement approach or by complementing the corresponding F. fujikuroi mutants. The cluster organization and gene functions are similar to those in F. fujikuroi. However, the two border genes in the Fusarium cluster encoding the GA 4 desaturase (DES) and the 13-hydroxylase (P450-3) are absent in the S. manihoticola GA gene cluster, consistent with the spectrum of GAs produced by this fungus. The close similarity between the two GA gene clusters, the identical gene functions, and the conserved intron positions suggest a common evolutionary origin despite the distant relatedness of the two fungi. Although they are ubiquitous phytohormones, gibberellins (GAs) were first identified as secondary metabolites of the fungus Fusarium fujikuroi (teleomorph, Gibberella fujikuroi mating population C [MP-C]). GAs are also present in other fungi, including Sphaceloma manihoticola (teleomorph, Elsi- noe ¨ brasiliensis) (42) and Phaeosphaeria sp. strain L487 (47), and several bacteria, such as Rhizobium phaseoli (5) and some Bacillus species (17). In recent years, studies of GA biosynthe- sis in plants and F. fujikuroi have revealed significant differ- ences at the chemical (pathway), biochemical (enzyme), and genetic levels, indicating that plants and Fusarium have evolved their complex biosynthetic pathways to GAs indepen- dently and that corresponding genes have not been transferred to the fungus by horizontal gene transfer (reviewed in refer- ence 20). Although higher plants (Arabidopsis thaliana) and Fusarium produce structurally identical GAs, there are pro- found differences in the sequences of the biosynthetic steps (reviewed in references 20 and 51). A major difference is the stage at which the hydroxyl groups are introduced. Whereas 3-hydroxylation occurs at an early stage in the fungus, with GA 12 -aldehyde being converted to GA 14 -aldehyde (Fig. 1) (44), in Arabidopsis,3-hydroxylation of GA 9 and GA 20 are the final reactions in the formation of the biologically active end products, GA 4 and GA 1 , respectively. The final biosyn- thetic step in F. fujikuroi is the 13-hydroxylation of GA 7 to form GA 3 (58), whereas in plants, 13-hydroxylation of GA 12 to GA 53 is a relatively early step (20). The members of the two kingdoms also differ in the nature of the enzymes utilized for GA biosynthesis. The formation of plant GAs requires both membrane-bound cytochrome P450 monooxygenases and sol- uble 2-oxoglutarate-dependent dioxygenases (18, 19), whereas in fungal GA biosynthesis, only monooxygenases have been shown to participate in the same reactions. Interestingly, in F. fujikuroi, as well as in the GA-producing fungus Phaeosphaeria sp. strain L487, the two-step cyclization of geranylgeranyl diphosphate (GGDP) to ent-kaurene via ent-copalyl diphos- phate (CDP) is catalyzed by a fungal-type bifunctional cyclase (CPS/KS) (Fig. 1) (54), whereas in plants, two independent diterpene cyclases catalyze these reactions (48, 54, 62). In contrast to Arabidopsis and other plants, where the GA bio- synthetic genes are dispersed throughout the genome (20), in F. fujikuroi the genes are physically linked in a gene cluster (29, 52). The gene cluster in F. fujikuroi includes a GA pathway- specific GGDP synthase-encoding gene (ggs2), whereas a sec- ond gene, ggs1, encodes the enzyme responsible for GGDP synthesis for primary metabolism (35). The ggs2 gene shares a bidirectional promoter with the ent-copalyl diphosphate or ent- kaurene synthase gene, cps/ks. In addition to four cytochrome P450 monooxygenase genes (P450-1 to P450-4), which encode multifunctional enzymes, the GA gene cluster also contains a GA 4 desaturase-encoding gene, des (58). Recently, the GA biosynthetic gene cluster has been identified in a second GA-producing fungus, Phaeosphaeria sp. strain L487 (23). In contrast to F. fujikuroi, this fungus produces GA 1 , rather than GA 3 . Aside from the Phaeosphaeria sp. strain L487 cps/ks gene (Phcps/ks), the GA gene cluster of Phaeosphaeria sp. strain L487 contains four cytochrome P450 monooxygenase genes, three of which are similar to the F. fujikuroi monooxygenase genes P450-1, P450-2, and P450-4. Surprisingly, the Phaeosphaeria * Corresponding author. Mailing address: Westfa ¨lische Wilhelms- Universita ¨t Mu ¨nster, Institut fu ¨r Botanik, Schlossgarten 3, 48149 Mu ¨n- ster, Germany. Phone: (49)251 8324801. Fax: (49)251 8323823. E-mail: [email protected]. † Supplemental material for this article may be found at http://aem .asm.org/. Published ahead of print on 20 June 2008. 5325
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Isolation and Characterization of the Gibberellin Biosynthetic Gene Cluster in Sphaceloma...

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2008, p. 5325–5339 Vol. 74, No. 170099-2240/08/$08.00�0 doi:10.1128/AEM.00694-08Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Isolation and Characterization of the Gibberellin Biosynthetic GeneCluster in Sphaceloma manihoticola�†
Christiane Bomke,1 Maria Cecilia Rojas,2 Fan Gong,3 Peter Hedden,3 and Bettina Tudzynski1*Westfalische Wilhelms-Universitat Munster, Institut fur Botanik, Schlossgarten 3, 48149 Munster, Germany1; Laboratorio de
Bioorganica, Departamento de Quımica, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile2; andRothamsted Research, Harpenden, Hertfordshire AL5 2JQ, United Kingdom3
Received 25 March 2008/Accepted 14 June 2008
Gibberellins (GAs) are tetracyclic diterpenoid phytohormones that were first identified as secondary metabolitesof the fungus Fusarium fujikuroi (teleomorph, Gibberella fujikuroi). GAs were also found in the cassava pathogenSphaceloma manihoticola, but the spectrum of GAs differed from that in F. fujikuroi. In contrast to F. fujikuroi, theGA biosynthetic pathway has not been studied in detail in S. manihoticola, and none of the GA biosynthetic geneshave been cloned from the species. Here, we present the identification of the GA biosynthetic gene cluster from S.manihoticola consisting of five genes encoding a bifunctional ent-copalyl/ent-kaurene synthase (CPS/KS), a pathway-specific geranylgeranyl diphosphate synthase (GGS2), and three cytochrome P450 monooxygenases. The functionsof all of the genes were analyzed either by a gene replacement approach or by complementing the corresponding F.fujikuroi mutants. The cluster organization and gene functions are similar to those in F. fujikuroi. However, the twoborder genes in the Fusarium cluster encoding the GA4 desaturase (DES) and the 13-hydroxylase (P450-3) areabsent in the S. manihoticola GA gene cluster, consistent with the spectrum of GAs produced by this fungus. Theclose similarity between the two GA gene clusters, the identical gene functions, and the conserved intron positionssuggest a common evolutionary origin despite the distant relatedness of the two fungi.
Although they are ubiquitous phytohormones, gibberellins(GAs) were first identified as secondary metabolites of thefungus Fusarium fujikuroi (teleomorph, Gibberella fujikuroimating population C [MP-C]). GAs are also present in otherfungi, including Sphaceloma manihoticola (teleomorph, Elsi-noe brasiliensis) (42) and Phaeosphaeria sp. strain L487 (47),and several bacteria, such as Rhizobium phaseoli (5) and someBacillus species (17). In recent years, studies of GA biosynthe-sis in plants and F. fujikuroi have revealed significant differ-ences at the chemical (pathway), biochemical (enzyme), andgenetic levels, indicating that plants and Fusarium haveevolved their complex biosynthetic pathways to GAs indepen-dently and that corresponding genes have not been transferredto the fungus by horizontal gene transfer (reviewed in refer-ence 20). Although higher plants (Arabidopsis thaliana) andFusarium produce structurally identical GAs, there are pro-found differences in the sequences of the biosynthetic steps(reviewed in references 20 and 51). A major difference is thestage at which the hydroxyl groups are introduced. Whereas3�-hydroxylation occurs at an early stage in the fungus, withGA12-aldehyde being converted to GA14-aldehyde (Fig. 1)(44), in Arabidopsis, 3�-hydroxylation of GA9 and GA20 arethe final reactions in the formation of the biologically activeend products, GA4 and GA1, respectively. The final biosyn-thetic step in F. fujikuroi is the 13-hydroxylation of GA7 toform GA3 (58), whereas in plants, 13-hydroxylation of GA12 to
GA53 is a relatively early step (20). The members of the twokingdoms also differ in the nature of the enzymes utilized forGA biosynthesis. The formation of plant GAs requires bothmembrane-bound cytochrome P450 monooxygenases and sol-uble 2-oxoglutarate-dependent dioxygenases (18, 19), whereasin fungal GA biosynthesis, only monooxygenases have beenshown to participate in the same reactions. Interestingly, in F.fujikuroi, as well as in the GA-producing fungus Phaeosphaeriasp. strain L487, the two-step cyclization of geranylgeranyldiphosphate (GGDP) to ent-kaurene via ent-copalyl diphos-phate (CDP) is catalyzed by a fungal-type bifunctional cyclase(CPS/KS) (Fig. 1) (54), whereas in plants, two independentditerpene cyclases catalyze these reactions (48, 54, 62). Incontrast to Arabidopsis and other plants, where the GA bio-synthetic genes are dispersed throughout the genome (20), inF. fujikuroi the genes are physically linked in a gene cluster (29,52). The gene cluster in F. fujikuroi includes a GA pathway-specific GGDP synthase-encoding gene (ggs2), whereas a sec-ond gene, ggs1, encodes the enzyme responsible for GGDPsynthesis for primary metabolism (35). The ggs2 gene shares abidirectional promoter with the ent-copalyl diphosphate or ent-kaurene synthase gene, cps/ks. In addition to four cytochromeP450 monooxygenase genes (P450-1 to P450-4), which encodemultifunctional enzymes, the GA gene cluster also contains aGA4 desaturase-encoding gene, des (58).
Recently, the GA biosynthetic gene cluster has been identifiedin a second GA-producing fungus, Phaeosphaeria sp. strain L487(23). In contrast to F. fujikuroi, this fungus produces GA1, ratherthan GA3. Aside from the Phaeosphaeria sp. strain L487 cps/ksgene (Phcps/ks), the GA gene cluster of Phaeosphaeria sp. strainL487 contains four cytochrome P450 monooxygenase genes,three of which are similar to the F. fujikuroi monooxygenasegenes P450-1, P450-2, and P450-4. Surprisingly, the Phaeosphaeria
* Corresponding author. Mailing address: Westfalische Wilhelms-Universitat Munster, Institut fur Botanik, Schlossgarten 3, 48149 Mun-ster, Germany. Phone: (49)251 8324801. Fax: (49)251 8323823. E-mail:[email protected].
† Supplemental material for this article may be found at http://aem.asm.org/.
� Published ahead of print on 20 June 2008.
5325

FIG. 1. Biosynthetic pathway for GAs and related diterpenoids in F. fujikuroi. The major pathway is indicated by boldface arrows.
5326

cluster does not contain a pathway-specific GGDP synthase geneequivalent to ggs2 in F. fujikuroi nor genes with homology to the13-hydroxylase-encoding gene P450-3 or the desaturase-encodinggene des (23).
At present, only a few GA-producing fungi are known. Be-sides F. fujikuroi (G. fujikuroi MP-C), Fusarium konzum (G.fujikuroi MP-I) (32), and Phaeosphaeria sp. strain L487, theability to produce GAs has been described for several Sphac-eloma species, mainly for S. manihoticola (reviewed in refer-ences 30, 42, and 43). In contrast to F. fujikuroi, Sphacelomaspp. produce GA4 rather than GA3 as the main end product ofthe GA pathway (8, 38, 42, 43), indicating that the desaturase,for conversion of GA4 to GA7, and the 13-hydroxylase, forconversion of GA7 to GA3, are inactive or missing in thesefungi. However, GA biosynthesis in Sphaceloma has not yetbeen subjected to molecular genetic studies.
In this paper, we focus on the identification of the GAbiosynthetic gene cluster in S. manihoticola, a pathogen of thetropical starchy root crop cassava (Manihot esculenta) thatcauses the hyperelongation of the internodes as a result of GAsecretion. The cluster consists of five genes that share highsimilarity with the corresponding F. fujikuroi homologues.However, the cluster contains neither a desaturase-encodinggene (des) nor a P450-3 homologue, consistent with the inabil-ity to produce GA7, GA3, and GA1. The S. manihoticola clustergenes fully restored GA production in the correspondingFusarium mutants, confirming that the new gene cluster isindeed responsible for GA biosynthesis. Despite some differ-ences in the cluster organization, we suggest that the pathwaygenes in both fungi have a common phylogenetic origin.
MATERIALS AND METHODS
Fungal strains. The S. manihoticola wild-type strain Lu949 (ATCC 44292) wasdonated by W. Rademacher (BASF Agricultural Center, Limburgerhof, Ger-many). IMI58289 (CABI Biosciences, Kew, United Kingdom) is a GA-producingwild-type strain of F. fujikuroi (teleomorph, G. fujikuroi MP-C). Strain Ff-�cps/ks-GD-T22 (54) is an F. fujikuroi IMI58289 disruption mutant that does notproduce GAs or precursors. The GA-defective strain SG139 is an F. fujikuroi UVmutant that lacks the whole GA gene cluster (14, 56). It was kindly provided byE. Cerda-Olmedo and J. Avalos (University of Seville, Seville, Spain). MutantB1-41a, obtained by UV irradiation of F. fujikuroi strain GF-1a (7), was donatedby J. MacMillan (University of Bristol, Bristol, United Kingdom). The strain wasshown to contain a point mutation in the P450-4 locus and is GA deficient (56).
Media and culture conditions. For DNA isolation, Fusarium strains weregrown on cellophane sheets (Alba Gewurze, Bielefeld, Germany) placed on CMagar (39) for 3 days at 28°C. S. manihoticola Lu949 was grown on potato dextroseagar plates for 10 days under the same conditions. The mycelium was harvested,frozen in liquid nitrogen, and lyophilized for 24 h. For RNA isolation, Fusariumstrains were grown in 100%, 20%, or 0% ICI medium (16) containing 8%glucose, 0.5% MgSO4, 0.1% KH2PO4, and 5.0, 1.0, or 0 g/liter NH4NO3, respec-tively. S. manihoticola was grown in optimized production medium (OPM) con-taining 6% sunflower oil, 0.05% (NH4)2SO4, 1.5% corn steep solids (Sigma-Aldrich, Taufkirchen, Germany), and 0.1% KH2PO4. For shift experiments, theSphaceloma wild-type strain Lu949 was grown as indicated below. For GAproduction, Fusarium strains were grown for 7 to 10 days and S. manihoticola forabout 44 days on a rotary shaker (190 rpm) at 28°C in 300-ml Erlenmeyer flaskscontaining 100 ml of either 20% ICI medium or OPM. S. manihoticola was alsocultivated for 30 days on 10 g brown rice that had been autoclaved with 15 mlH2O in 300-ml Erlenmeyer flasks. ICI medium for Sphaceloma cultures wasprepared with 8% maltose instead of glucose. All fungal strains were preculti-vated in Darken medium (11) for 3 (Fusarium) or 7 (Sphaceloma) days.
Bacterial strains and plasmids. Escherichia coli strain Top10F� (Invitrogen,Groningen, The Netherlands) was used for plasmid propagation. The vectorspUC19 (Fermentas, St. Leon-Rot, Germany) and pBluescript II SK(�) (Strat-agene, La Jolla, CA) were used to clone DNA fragments carrying the S. mani-
hoticola Lu949 cluster genes and gene fragments. The plasmid pSm-cps/ks wasused for complementation of the F. fujikuroi �cps/ks-GD-T22 strain. To obtainthe plasmid pSm-cps/ks, a 5,276-kb SacI fragment of � clone 13.1 was cloned intopUC19/SacI. The S. manihoticola cps/ks (Smcps/ks) gene replacement vectorp�Sm-cps/ks was obtained by amplifying two flanking sequences of Smcps/ks byPCR, introducing a KpnI (flank 1) restriction site and HindIII and BamHI (flank2) restriction sites into the oligonucleotides. For the amplified flank 1, a genomicSalI site could be used (see Fig. 3). The flanking sequences (flank1, 1.2 kb; flank2, 1 kb) were cloned into the corresponding sites of the vector pUCH2-8 (1)carrying a hygromycin resistance cassette. The 4.7-kb replacement fragment wasobtained by restriction with KpnI and BamHI. For heterologous expression of S.manihoticola GA genes in the GA-deficient F. fujikuroi mutants SG139 andB1-41a, the three P450 genes were isolated by PCR approaches and cloned intothe vector pCR2.1TOPO, creating the vectors pSm-P450-1, pSm-P450-2, andpSm-P450-4. For expression of the Sphaceloma P450 genes under the control ofthe corresponding F. fujikuroi promoter, the single genes were amplified withintroduced or genomic (SmP450-1) NcoI sites at the translation start codon,cloned into pCR2.1TOPO, excised with EcoRI, and ligated into pUC19. Thecorresponding Fusarium promoters were generated by PCR, introducing NcoIsites at the 5� and 3� ends. They were ligated into the NcoI-digested pUC19containing the appropriate Sphaceloma genes, creating the vectors pFfP1prom::SmP1, pFfP2prom::SmP2, and pFfP4prom::SmP4, respectively. The cosmid pCos1,derived from a cosmid library based on strain m567, contains the entire GA genecluster, including the 5� and 3� regions (an �40-kb insert) (P. Linnemannstonsand B. Tudzynski, unpublished data). In cotransformation experiments, pNR1(nourseothricin resistance) (27, 31) or pAN7-1 (hygromycin resistance) (41) wasused for selection of resistant transformants.
Construction and screening of a genomic library. A � DASH II library (Strat-agene) from genomic DNA of S. manihoticola strain Lu949 was prepared fol-lowing the manufacturer’s instructions. About 35,000 recombinant phage wereplated and transferred to nylon membranes (Whatman GmbH, Dassel, Ger-many). Hybridization was performed at high stringency (65°C) in 5� Denhardt’ssolution containing 5% dextran sulfate. The blots were washed under hybridiza-tion conditions with 0.1� SSC (1� SSC is 0.15 M NaCl, 0.015 M sodium citrate),0.1% sodium dodecyl sulfate (SDS). Positive recombinant phages were used fora second round of plaque purification.
DNA isolation. Genomic DNA was isolated from lyophilized mycelium asdescribed previously by Doyle and Doyle (13). Plasmid DNA was extracted usingthe Qiagen plasmid extraction kit (Qiagen, Hilden, Germany). Lambda DNAfrom positive lambda phages was prepared according to the method of Maniatiset al. (34).
Southern blot and Northern blot analyses. For Southern analysis, genomic,plasmid, or phage DNA was digested to completion with appropriate restrictionenzymes (Fermentas, St. Leon-Rot, Germany), fractionated in 1.0% (wt/vol)agarose gels, and transferred to Hybond N� filters (Amersham Pharmacia,Freiburg, Germany). Hybridization was carried out in 6� SSC, 5� Denhardt’ssolution, 0.1% SDS, and 50 mM phosphate buffer, pH 6.6, at 65°C in thepresence of a random-primed [-32P]dCTP-labeled probe. Membranes werewashed once (2� SSPE [1� SSPE is 0.18 M NaCl, 10 mM NaH2PO4, and 1 mMEDTA {pH 7.7}], 0.1% SDS) before being exposed to autoradiographic film.Total fungal RNA was isolated using the RNAgents total-RNA isolation kit(Promega, Mannheim, Germany). Northern blot hybridizations were done aspreviously described by Church and Gilbert (10).
DNA sequencing and sequence homology searching. DNA sequencing of re-combinant plasmid clones was accomplished by the dideoxy chain terminationmethod (46) with an automatic sequencer, LI-COR 4200 (MWG, Munich, Ger-many). The two strands of overlapping subclones obtained from the genomicDNA clones were sequenced using the universal and the reverse primers. DNAand protein sequence alignments were done using DNA Star (DNA Star Inc.,Madison, WI).
Sequence homology searches were performed using the NCBI database server.Protein homology was based on BlastX searches (2). For further investigations,the programs of DNA Star Inc. (Madison, WI) were used.
PCR and reverse transcription (RT)-PCR. PCR was performed as previouslydescribed (32). Heterologous PCR based on the two known fungal diterpenecyclase sequences of F. fujikuroi (FfCPS/KS) and Phaeosphaeria sp. strain L487(PhCPS/KS) to identify Smcps/ks was performed with the primer combinationcps/ks-Sm-F2 and Sm-cps/ks-5R1 (Table 1). For analysis of the GA gene clusterorganization in S. manihoticola, the following primers (Table 1) were synthesizedon the basis of the determined sequences of lambda clones 12.1, 13.1, and 14.1.They were applied in different combinations to obtain the whole cluster se-quence.
For amplifying the 5� and 3� flanks of the gene replacement vector p�Sm-
VOL. 74, 2008 GIBBERELLIN GENE CLUSTER IN SPHACELOMA MANIHOTICOLA 5327

cps/ks, the following primers were used (Table 1): Sm-GRP-cpsks-KpnI-F1 andSm-GRP-cpsks-LF-R1 (flank 1) and Sm-GRP-cpsks-HindIII-F1 combined withSm-GRP-cpsks-BamHI-R1 (flank 2). The analysis of putative Smcps/ks knockouttransformants was performend with the diagnostic primers Sm-cps/ks-GR-K-F1and PUCH-P for integration in the 5� region of Smcps/ks, and Sm-cps/ks-GR-K-R1 and PUCH-T for integration in the 3� region of Smcps/ks (Table 1). Thedeletion of the wild-type Smcps/ks gene copy was demonstrated by diagnosticPCR using the primers Sm-cps/ks-nF3 and Sm-cps/ks-nR1. The primers used forgeneration of vectors for heterologous gene expression are shown in Table 1.
For RT-PCR, 1 g of total RNA of nitrogen-starved wild-type cells served as
a template, and the nonspecific oligo(dT)18 primer (Fermentas) was used tocreate cDNA by reverse transcription.
Fungal transformations. Preparation of protoplasts of F. fujikuroi was per-formed as previously described (53). For S. manihoticola, the following modifi-cations were used: Lu949 was precultivated for 7 to 10 days on potato dextroseagar and used for inoculation of 100 ml of Darken medium. After 5 days on arotary shaker at 28°C and 190 rpm, 1 ml of this culture was transferred to 100 mlof CM medium for 24 h. Sphaceloma protoplasts were obtained with the samemixture of lysing enzymes as Fusarium, with an additional 250 mg/100 ml muta-nase (InterSpex Products Inc., San Mateo, CA). Protoplasts (108) of each strainwere transformed with up to 13 g of the respective complementation vectors orpCos1 and, if necessary, with 7 g of the nourseothricin resistance-mediatingvector pNR1 or the hygromycin resistance cassette-containing vector pAN7-1,respectively. For deletion of Smcps/ks, 10 g of the KpnI/BamHI fragment of thegene replacement vector p�Sm-cps/ks was used for transformation of S. mani-hoticola strain Lu949. Transformed protoplasts were regenerated at 28°C in acomplete regeneration agar (0.7 M sucrose, 0.05% yeast extract, 0.1% CasaminoAcids) containing 120 g/ml hygromycin B (InvivoGen, San Diego, CA) or 120g/ml nourseothricin (Werner BioAgents, Jena, Germany) for 6 or 7 days.Complementation mutants of the hygromycin-resistant Fusarium strainFf-�cps/ks-GD-T22 were generated by cotransformation with the nourseo-thricin resistance-mediating vector pNR1. For purification of the Smcps/ksdeletion mutants, single protoplasts were regenerated from hygromycin-re-sistant transformants and used for DNA isolation, PCR, and Southern andNorthern blot analyses.
GA analysis. GA3, GA4, and GA7 in culture fluids of all strains were analyzedby high-performance liquid chromatography (HPLC) (6) with a Merck HPLCsystem with a UV detector and a Lichrospher 100 RP18 column (5 m; 250 by4 mm). These GAs were also analyzed by thin-layer chromatography (TLC) onsilica gels eluted with ethyl acetate-chloroform-acetic acid (90:60:7.5). Gaschromatography-mass spectrometry (GC-MS) using a GCQ system (Thermo-Finnigan) as described by Troncoso et al. (50) was used to determine thecomplete GA complements produced by the different strains after extractionfrom the culture fluid and to identify the products from incubations with 14C-labeled substrates after HPLC or TLC separation. Identifications were based oncomparison of mass spectra with published data (15).
Labeled substrates. ent-7-hydroxy[14C4]kaurenoic acid, [14C4]GA12-alde-hyde, ent-6,7-diOH[14C4]kaurenoic acid, and ent-[14C4]kauradienoic acid wereprepared from R-[2-14C]mevalonic acid (Amersham) by incubation with a cell-free system from Cucurbita maxima endosperm in the presence of ATP, MgCl2,and NADPH (59). [14C1]GA14 was prepared from [14C1]GA12 by incubation withcultures of an SG139-P450-1 F. fujikuroi transformant (44). ent-[17-14C]kaure-noic acid, [17-14C]GA12, [17-14C]GA15, and[17-14C]GA24 were obtained from L.Mander (Australian National University, Canberra, Australia).
Incubations with radiolabeled GA precursors. Cultures of SG139-Sm-P450-1or SG139-Sm-P450-2 transformants grown in 40% ICI medium for 3 days at 28°Cwere harvested, and the mycelia were washed with 0% ICI medium (16). Afterresuspension of the mycelia in 0% ICI, 1-ml aliquots were transferred to 25-mlflasks containing 10 ml of the same medium. Radiolabeled substrates were addedas methanol solutions (300,000 dpm to 500,000 dpm per flask), and the cultureswere incubated for 3 days on an orbital shaker at 28°C. After incubation, theculture was filtered and the culture fluid was acidified to pH 3.0 and partitionedagainst ethyl acetate. The organic phase was evaporated, and the residue wasdissolved in 20% MeOH/H2O, pH 3.0. This solution was applied to C18 car-tridges (Bakerbond; Baker), eluted in 2 ml MeOH, and separated by HPLC ona C18 column. A 30-min linear gradient from 60 to 100% MeOH/H2O, pH 3.0,was utilized to separate the products formed from the radiolabeled substrates,except for [14C4]kaurenolides, which were separated from ent-[14C4]kauradienoicacid in a 15-min gradient from 75 to 100% MeOH/H2O, pH 3.0, followed by100% MeOH for 15 min. The flow rate was 1 ml/min. Fractions (1 ml) werecollected, and the radioactivity was measured by scintillation counting.[14C4]fujenoic acids were separated from ent-6,7-diOH[14C4]kaurenoicacid by TLC on silica gel G plates developed with hexane-ethyl acetate (3:7)containing drops of formic acid. The silica was scraped from the plate andeluted with methanol, and the radioactivity was quantified by liquid scintil-lation counting.
Nucleotide sequence accession numbers. The gene sequences of SmP450-1,SmP450-4, SmP450-2, Smggs2, and Smcps/ks of S. manihoticola Lu949 have beendeposited in the GenBank database under accession numbers AM886288,AM886289, AM886290, AM886291, and AM886292, respectively.
TABLE 1. Primers used for GA gene cluster isolation, vectorconstruction, and diagnostic PCR
Primer Sequence (5�33�)a
Sm-P450-1-K-F1 .....................................GGTCATGAGCGTGGCTTTGSm-P450-1-Fus-R2 .................................CGAGGAGCTAAGGCAAGAAGCSm-P450-1-RT-F1...................................GGGTTCGGCGCACCTTGTAASm-P450-1-RT-F2...................................GCAGCTTCTTGCCTTAGCTCCTCGSm-P450-1-RT-PromR3.........................CCTTGGACGAATCTGCGGTGTSm-P450-4-K-F1 .....................................CCATGGTGAGCAACAGGTACACCSm-P450-4-F2..........................................CCGGAGAAGCATGATGAGACCSm-P450-4-K-R1.....................................GGAGGTGGCGGTATTTGGACGSm-P450-2-K-F1 .....................................GGAACTGACCGGCGATGAASm-P450-2-F1..........................................CGTCGTGCTTCCTGAACTTGTSm-P450-2-R1.........................................CATCTACCAATTGTGAACAACSm-P450-2-Prom-K-R1..........................GGAGGTTGAGGGGTTGCATTGGSm-ggs2-RT-F1.......................................GCAACCTTGCTAATCAAGGSm-ggs2-F1..............................................CAGTCTACATCCAATGTCTGGA
GACCSm-ggs2-RT-Prom-R1 ...........................GGGCTCTGACGAGCTGCACASm-ggs2-P450-2-Fus-R1.........................CCATTATTGACGACCGGGTGCSm-cpsks-K-F1........................................CCGCTTTGGTACTCTCCSm-cpsks-nF3 ..........................................GGATGAGTGATCAACTCTCGGGGSm-cpsks-nF4 ..........................................GACCTCATTGCGGAGGCATGTAGSm-cpsks-nR1 .........................................ATTACCGCAAATGACCCAGGSm-cpsks-nR4 .........................................TTCTGCTGTCGAGTCGTACGSm-cpsks-nR6 .........................................GCACGGTAGAGGGGCAGATCGCT
CTCTTGSm-cpsks-RT-R2.....................................CGAGTAGGCGCATGCTACAT
GATCSm-cpsks-K-R1 .......................................CCTGCCAACACGAATGTCCcps/ks-Sm-F2 ...........................................AC(G/C)AGCTGTCAG(A/G)T(C/A)T
A(T/C)GACAC(T/A)GC(C/T)TGGG(T/C)GGCC
Sm-cps/ks-5R1.........................................TTT(T/G)GC(A/C)GTGTC(A/G)TCAACATC
Sm-P450-1-K-F1 .....................................GGTCATGAGCGTGGCTTTGSm-P450-1-Prom-K-R1..........................GCTGGAGGTAATCCGTGCSm-P450-1-RT-Prom-R1 .......................CGAAACAGGTGATTCGATGCSm-P450-4-K-F1 .....................................CCATGGTGAGCAACAGGTACACCSm-P450-4-K-R1.....................................GGAGGTGGCGGTATTTGGACGSm-P450-4-NcoI-F1................................GGATTACCTCCACCATGGTGGAC
GACASm-P450-1-4-Prom-Nco-R1...................TGTCGTCCACCATGGTGGAGGTA
ATCCFf-P450-4-1-Prom-NcoI-F1 ...................TGGACTTACcCATGGTGTTATTGFf-P450-4-1-Prom-NcoI-R1...................GGTTCGCCATGGTCTGACAACSm-P450-2-K-F1 .....................................GGAACTGACCGGCGATGAASm-P450-2-Prom-K-R1..........................GGAGGTTGAGGGGTTGCATTGGSm-P450-2-NcoI-R1 ...............................CCAGAAGAACGcCATGGCACAGG
CAATACCFf-P450-2-Prom-NcoI-F1.......................CCTATGACGcCATGGCTCCTCACA
TAACCFf-P450-2-Prom-NcoI-R1 ......................GATGCcCATGGCGGGTACAATG
TTGGSm-GRP-cpsks-KpnI-F1 ........................CAGAGGTACCATAGAGCGTTGAT
CTACCSm-GRP-cpsks-LF-R1 ...........................GGTTAGGGTGCAACCCTGGSm-GRP-cpsks-HindIII-F1 ...................CTGGAGAAAGCTTTGGCTACTCSm-GRP-cpsks-BamHI-R1 ...................GGCGGATCCGGGCGGAGATG
AGTTSm-cps/ks-GR-K-F1 ...............................CGTCAAATTCATGTAGTCGTCGCGSm-cps/ks-GR-K-R1...............................CGATGAAGACTTCGACCGGGTTCPUCH-P ..................................................CCCTTGGCCTCAGACACCPUCH-T ..................................................TCAACGCATATAGCGCTAGCSm-cps/ks-nF3.........................................GACCTCATTGCGGAGGCATGTAGSm-cps/ks-nR1 ........................................ATTACCGCAAATGACCCAGG
a Underlining indicates introduced restriction sites.
5328 BOMKE ET AL. APPL. ENVIRON. MICROBIOL.

RESULTS
Isolation and organization of the GA gene cluster in S.manihoticola. In order to identify the GA biosynthetic genes inthe Sphaceloma genome, we constructed a lambda DASH IIgenomic library from the S. manihoticola wild-type strainLu949. Degenerate primers were designed from different con-served domains of the two known fungal CPS/KS enzymesfrom F. fujikuroi and Phaeosphaeria sp. strain L487. Only thecombination of primers encoding DVDDTAK (Sm-cps/ks-5R1) and TSCQ(I/V)YDTAW(V/A)A (cps/ks-Sm-F2) yieldeda single DNA fragment of the expected size, approximately 1kb, at an annealing temperature of 47°C. Sequence analysisrevealed significant similarity (51% identity at the amino acidlevel) to both FfCPS/KS and PhCPS/KS. This PCR productwas used as a probe to screen the genomic library. Subcloningof different hybridizing fragments into the vectors pUC19 andpBluescript II SK(�) and subsequent sequencing revealed sev-eral overlapping clones carrying the cps/ks homologous gene, aputative GGDP synthase gene, and parts of a cytochrome P450monooxygenase gene. The sequences possess close similarityto F. fujikuroi GA biosynthetic genes, suggesting that thecloned genes are part of the GA gene cluster in S. manihoti-
cola. Subsequent chromosome walking resulted in identifica-tion of two additional P450 monooxygenase-encoding genesupstream of the first one. Sequencing of cDNA clones of thepredicted GA genes and comparisons with the genomic se-quences revealed their coding regions and intron positions(Table 2).
Neither an FfP450-3 homologue nor a desaturase-encodinggene was detected at the right and left borders, respectively,indicating that the putative GA biosynthesis gene cluster in S.manihoticola consists of only five genes (Fig. 2 and Table 2).The lack of these enzymes is consistent with the accumulationof GA4 and GA9, rather than GA3 and GA1, as final products(43) (Fig. 1). Furthermore, there are no homologues of genesdefined immediately outside of the F. fujikuroi GA gene clusterpresent at the borders in S. manihoticola. The putative genecluster is flanked by two open reading frames, a chitinase-encoding gene (orf1) upstream and a methyltransferase-encod-ing gene (orf2; pfam05148 family) downstream, which probablydo not belong to the GA gene cluster (Fig. 2).
The structure of the gene cluster in S. manihoticola Lu949 isvery similar to that in F. fujikuroi. The most striking similarityis that both clusters contain a pathway-specific GGDP synthase
FIG. 2. Comparison of the GA gene clusters in S. manihoticola and F. fujikuroi. Homologous genes are shown in the same colors. Fusariumgenes without homologues in the Sphaceloma gene cluster are shown in black. The crosses indicate gene inversions. The arrows indicate theorientations of transcription. Genes not belonging to the cluster are shown in gray (without indication of intron positions). � clones used forisolation and sequencing of the cluster are displayed above the S. manihoticola cluster.
TABLE 2. Gibberellin biosynthetic gene cluster in S. manihoticola
Gene (accession no.) Length (bp) Intronno. Function (family) No. of
amino acids
Identity/similarity on aminoacid level to F. fujikuroi
orthologue (%)a
SmP450-1(AM886288)
1,704 3 GA14 synthase (CYP68A) 513 56/75
SmP450-4(AM886289)
1,679 2 ent-kaurene oxidase (CYP503) 525 51/69
SmP450-2(AM886290)
1,768 4 C-20 oxidase (CYP68B) 520 53/72
Smggs2 (AM886291) 1,203 5 GGDP synthase 309 47/66Smcps/ks (AM886292) 3,046 3 CDP/ent-kaurene synthase 962 44/59
a BlastP results.
VOL. 74, 2008 GIBBERELLIN GENE CLUSTER IN SPHACELOMA MANIHOTICOLA 5329

gene that is physically linked to the cps/ks homologous genesharing the same promoter region (Fig. 2). As in F. fujikuroi,two of the cytochrome P450 monooxygenase genes with thehighest levels of sequence similarity to FfP450-1 (GA14 syn-thase) (44) and FfP450-4 (ent-kaurene oxidase) (56) are alsophysically linked and form a transcriptional unit by sharing abidirectional promoter. Compared with the situation in F. fu-jikuroi, the latter complex is inverted in the Sphaceloma ge-nome. The third P450 monooxygenase gene, SmP450-2, whichrevealed the highest degree of sequence similarity to the GA
20 oxidase gene FfP450-2 (57), is also inverted with respect tothe cluster in F. fujikuroi (Fig. 2).
Knockout of Smcps/ks. To confirm that the identified genesare indeed responsible for GA biosynthesis in S. manihoticola,we performed a knockout of the putative ent-copalyl diphos-phate/ent-kaurene synthase-encoding gene (Smcps/ks). In F.fujikuroi, Ffcps/ks encodes a bifunctional enzyme that catalyzestwo specific steps of the GA biosynthetic pathway, the cycliza-tion of GGDP via CDP to ent-kaurene. Knockout mutantswould be characterized by the total lack of GAs and their
FIG. 3. Targeted replacement of the Smcps/ks gene. (A) Replacement strategy showing physical maps of the gene replacement fragmentSm-�cps/ks and the Smcps/ks wild type (WT) and the gene locus from an Sm-�cps/ks knockout mutant showing the components of the hygromycinresistance cassette (hyg; black boxes), restriction sites used for generating the replacement fragment, and digestion of the genomic DNA forSouthern blot analysis (panel B) (see the text). The small arrows indicate the positions of primers used for cloning the 5� and 3� flanks andscrutinizing the correct integration of the replacement cassette (see Materials and Methods). The sizes of hybridizing fragments are also indicated.Note that the drawing is not to scale. (B) Southern blot analysis of genomic DNA of the WT and some homokaryotic single protoplast isolatesof Sm-�cps/ks deletion mutants T1 (T1-1), T2 (T2-1, T2-2, and T2-3), and T12 (T12-3, T12-5, and T12-7). DNA was digested with HindIII,electrophoresed, blotted, and hybridized with two different probes (panel A) (see the text). The hybridizing patterns demonstrate the successfuldeletion of Smcps/ks in all transformants, although some additional ectopic integrations appear to indicate the WT fragment. (C) Northern blotanalysis of the S. manihoticola �cps/ks deletion mutants T1-1, T2-1, and T12-3. Total RNA was hybridized to the probes as indicated. Thehybridization patterns confirm the deletion of Smcps/ks in all transformants, although rRNA appears in the background with approximately thesame size as Smcps/ks and Smggs2 transcripts. All mutants showed downregulation of the expression of the other GA cluster genes, probably dueto the integration of the larger replacement fragment and not because of feedback regulation, since single genes are expressed at a high level inF. fujikuroi SG139, a mutant lacking the whole GA gene cluster (Fig. 6).
5330 BOMKE ET AL. APPL. ENVIRON. MICROBIOL.

diterpenoid precursors. For this experiment, we needed toestablish a transformation system for S. manihoticola, whichwas achieved by modifying the transformation procedure usedfor F. fujikuroi. Transformation of the wild-type strain, Lu949,with the 4.8-kb KpnI/BamHI fragment of the Smcps/ks replace-ment vector (Fig. 3A) resulted in about 300 hygromycin-resis-tant transformants, 13 of which had integrated the replacementfragment at the cps/ks locus, as shown by diagnostic PCR usingthe primer pairs pUCH-P and Sm-cps/ks-GR-K-F1, andpUCH-T and Sm-cps/ks-GR-K-R1, respectively (Fig. 3A). Theheterokaryotic transformants were purified to homokaryons byprotoplast isolation. Three of the transformants, T1, T2, andT12, did not contain an intact wild-type Smcps/ks gene, asdemonstrated by diagnostic PCR using primers Sm-cps/ks-nF3and Sm-cps/ks-nR1 and by Southern blot hybridization of Hin-dIII-digested genomic DNAs of the transformants and thewild-type Lu949 using flank 1 as a probe (probe 1) (Fig. 3B;shown for T1 and T12). Due to the homologous integration ofthe replacement cassette, the hybridizing 3.9-kb wild-typefragment was replaced by a larger (5.3-kb) band in the trans-formants. Transformants T2 and T12 showed one or two ad-ditional hybridizing fragments resulting from ectopic integra-tions of the cassette or parts of it, whereas T1 contained onlyone copy of the replacement fragment. This was also demon-strated by probing with an internal cps/ks fragment (probe 2),which was replaced by the resistance marker in the mutants(Fig. 3B). The successful deletion of the Smcps/ks gene copy inthese transformants was confirmed by Northern blot analysis.
Only the wild-type strain Lu949 produced the correct 2.9-kbSmcps/ks transcript (Fig. 3C). Furthermore, the purified dele-tion mutants expressed none of the other cluster genes or theflanking genes orf1 and orf2 (data not shown). Possible reasonsfor this are discussed below.
To show if the deletion of the Smcps/ks gene affects GAbiosynthesis, two purified clones of transformants T1 and T12,as well as the wild-type strain, Lu949, were cultivated underGA production conditions for 44 days in the synthetic ICImedium containing maltose instead of glucose and on auto-claved brown rice kernels. The culture extracts were analyzedby GC-MS. No GAs or any intermediates were found, consis-tent with SmCPS/KS acting as a bifunctional ent-kaurene syn-thase (Fig. 4).
Restoration of GA production in F. fujikuroi by complemen-tation with Smcps/ks. To confirm the function of Smcps/ks, wecomplemented an F. fujikuroi cps/ks disruption strain with thehomologous gene from S. manihoticola. We transformed theplasmid pSm-cps/ks, carrying the entire S. manihoticola cps/ksgene copy with its endogenous promoter, into Ff-�cps/ks-GD-T22, an F. fujikuroi cps/ks disruption mutant, which was shownto be unable to produce GAs or precursors (54). Cotransfor-mation of pSm-cps/ks with the vector pNR1 carrying thenourseothricin resistance cassette yielded 40 resistant transfor-mants, 26 of which were shown to contain the Smcps/ks gene inthe F. fujikuroi genome by PCR analysis using the primer pairSm-cpsks-K-F1 and Sm-cpsks-K-R1. Heterologous expressionof the transgene was confirmed by Northern blot analysis witha 1.1-kb SalI fragment of Smcps/ks as a probe (Fig. 5A). Toconfirm the restoration of GA biosynthesis in the comple-mented mutants, we cultivated three transformants (aT2, aT3,and aT4), the F. fujikuroi wild-type strain IMI58289, and theF. fujikuroi �cps/ks mutant under GA production conditionsfor 7 days in 20% ICI medium and analyzed the culture mediaby GC-MS (Fig. 5B). Production of GA3 and some precursorswas restored in the complemented strains aT2 and aT3, but notin aT4. These results are consistent with the observation thatonly transformants aT2 and aT3 express Smcps/ks at a detect-able level (Fig. 5A).
Functional analysis of the three P450 monooxygenase genes.The deduced amino acid sequences of the proteins encoded bythe three cytochrome P450 genes in the Sphaceloma clusterwere found to be similar to those of the F. fujikuroi enzymesP450-1, P450-2, and P450-4 (Table 2). To demonstrate thatthey possessed the same activities as the F. fujikuroi homo-logues, we transformed the Sphaceloma P450 genes into dif-ferent F. fujikuroi mutant strains (see below). In order to en-sure high expression levels of the Sphaceloma genes in the F.fujikuroi background, transformations were also carried outwith the Sphaceloma P450 genes driven by the correspondingFusarium promoters (pFfP1prom::SmP1, pFfP2prom::SmP2, andpFfP4prom::SmP4) (Fig. 6).
To find out which of the two physically linked Sphacelomagenes SmP450-4 and SmP450-1 encodes ent-kaurene oxidase,each of the genes was transformed into the F. fujikuroi B1-41amutant, which lacks ent-kaurene oxidase activity due to a mu-tation in FfP450-4 (7, 56). Ethyl acetate extracts of the culturefluids from the transformants were analyzed by TLC andGC-MS to determine GA production (Fig. 7). The extracts ofB1-41a transformants complemented with the vectors pSm-
FIG. 4. GC-MS analysis of culture filtrates from wild-type S. mani-hoticola and of the cps/ks disruption mutant Sm-�cps/ks. Total ioncurrent chromatograms are shown for methyl esters trimethylsilylethers. Compounds identified in the wild type by comparison of themass spectra with published spectra (15) are as follows: 1, GA14; 2,GA4; 3, fujenoic diacid; 4, GA13; 5, GA36; 6, 7�,18-dihydroxykaureno-lide; and 7, 7�,13-dihydroxykarenolide.
VOL. 74, 2008 GIBBERELLIN GENE CLUSTER IN SPHACELOMA MANIHOTICOLA 5331

P450-4 (T3) and pFfP4 prom::SmP4 (T4) contained high levelsof GA3 and its precursors GA4 and GA7 in contrast to therecipient strain, B1-41a, which contained only traces of GA3
(Fig. 7). Transformation of B1-41a with SmP450-1 did notrestore GA production (Fig. 7), although the gene was ex-pressed in the F. fujikuroi background (data not shown). Theseresults confirm that SmP450-4 indeed encodes ent-kaureneoxidase, which catalyzes the three-step oxidation of ent-kau-rene to ent-kaurenoic acid.
SmP450-1 and SmP450-2 were transformed into the F.fujikuroi SG139 mutant, which lacks the entire GA genecluster and thus allows investigation of the catalytic func-tions of GA biosynthetic enzymes in isolation, as describedpreviously for the F. fujikuroi enzymes (44, 57, 58). The
FIG. 5. Heterologous expression of Smcps/ks in F. fujikuroi.(A) Northern blot analysis of the heterologous expression of Smcps/ksin the F. fujikuroi disruption mutant Ff-�cps/ks-GD-T22. Total RNAof S. manihoticola and F. fujikuroi wild-type (WT) strains, Ff-�cps/ks-GD-T22, and the complemented mutants Ff-�cps/ks-Sm-C-aT2, -aT3,-aT4, -T1, -3, -4, -6, -14, -16, -19, -20, and -22 was hybridized withFfcps/ks and Smcps/ks (indicated by cps/ks) as probes. (B) GC-MSanalysis of culture filtrates of wild-type F. fujikuroi (strain IMI58389),the F. fujikuroi disruption mutant Ff-�cps/ks-GD-T22 (�cps/ks), andthe complemented strains Ff-�cps/ks-Sm-C-aT2, -aT3, and -aT4. Totalion current chromatograms are shown for methyl esters trimethylsilylethers. Compounds identified in the wild type and strains Ff-�cps/ks-Sm-C-aT2 and -aT3 by comparison of the mass spectra with publishedspectra (15) are as follows: 1, GA7; 2, fujenal diacid; 3, fujenoic triacid;4, GA13; 5, isoGA3; 6, 7�,18-dihydroxykaurenolide; and 7, GA3.
FIG. 6. Northern blot analysis of heterologously expressed Sphac-eloma GA genes in Fusarium SG139 (a mutant lacking the entire GAgene cluster) or B1-41a (P450-4 mutant) background driven by theirown and corresponding Fusarium promoters (as described in the text)hybridized to probes as indicated. (A) Expression of SmP450-1 in F.fujikuroi SG139. (B) Expression of SmP450-2 in F. fujikuroi SG139.(C) Expression of SmP450-4 in F. fujikuroi B1-41a.
5332 BOMKE ET AL. APPL. ENVIRON. MICROBIOL.

reactions catalyzed by SmP450-1 and SmP450-2 were dem-onstrated by adding 14C-labeled GA precursors to culturesof SG139, complemented with the gene SmP450-1 orSmP450-2 driven by its own promoter or the promoter of thecorresponding F. fujikuroi gene. The reaction products wereseparated by HPLC and identified by GC-MS (see Tables S1and S2 in the supplemental material). SG139-Sm-P450-1(T5) and SG139-FfP1prom::SmP1 (T1) efficiently convertedent-[14C1]kaurenoic acid, ent-7-hydroxy [14C4]kaurenoicacid, or [14C4]GA12-aldehyde into one main labeled productidentified as the 3�-hydroxylated GA [14C]GA14 (see TableS1 in the supplemental material). SmP450-1 also catalyzed3�-hydroxylation of [14C1]GA12, which was completely con-verted into [14C1]GA14 (see Table S1 in the supplementalmaterial). Thus, SmP450-1 is indeed the functional homologueof the F. fujikuroi GA14 synthase FfP450-1 (44), catalyzing thefour oxidation steps from ent-kaurenoic acid to GA14 via ent-7-hydroxykaurenoic acid and GA12-aldehyde. Two additional prod-ucts were formed from ent-[14C1]kaurenoic acid, identified as7�,18-dihydroxy[14C1]kaurenolide and 3�,7�-dihydroxy[14C1]-kaurenolide (see Table S1 in the supplemental material; Fig. 8).These, as well as 7�-hydroxy[14C4]kaurenolide, were formed froment-[14C4]kauradienoic acid, which indicates that, besides cata-lyzing GA14 synthesis, SmP450-1 catalyzes kaurenolide produc-tion through the same reaction sequence described for F. fujikuroi(45). Finally, the reactions of the branch pathway to fujenoic acidswere demonstrated in SG139 complemented with SmP450-1 byincubation with ent-6,7-diOH-[14C4]kaurenoic acid. This pre-cursor was completely converted into [14C4]fujenal diacid and[14C4]fujenoic triacid (see Table S1 in the supplemental material;Fig. 8). Therefore, SmP450-1 catalyzes oxidation at several car-bon centers of ent-kaurenoic acid (C-7, C-6, C-3, and C-18),resulting in the formation of GA14, as well as kaurenolides andfujenoic acids, as previously shown for the F. fujikuroi homologue(44, 45).
Functional analysis of SmP450-2 was performed with cul-tures of SG139 transformed with SmP450-2 driven by the F.fujikuroi P450-2 promoter (pFfP2prom::SmP2). TransformantsT2 and T4 oxidized the C-20 methyl precursors [14C1]GA12
and [14C1]GA14 to give several C-20 oxidation products thatwere identified by GC-MS (see Table S2 in the supplementalmaterial). The non-3�-hydroxylated precursor [14C1]GA12
gave mainly the C-20 aldehyde product [14C1]GA24 (Fig. 8)plus a smaller amount of the 19,10-�-lactone [14C1]GA9, whilethe 3�-hydroxylated substrate [14C1]GA14 was completelyconverted into the 19,10-�-lactone [14C1]GA4. The C-20 alco-hol and C-20 carboxylic acid products, [14C1]GA15 and[14C1]GA25, were also detected in incubations with [14C1]-GA12, the latter as a minor product (see Table S2 in thesupplemental material). C-20 alcohol and C-20 aldehyde GAswould be intermediates in the C-20 oxidation sequence cata-lyzed by SmP450-2. Added [14C1]GA15 (C-20 alcohol) was notutilized and was recovered unconverted in the culture fluid,while [14C1]GA24 showed only a trace conversion into the C-20carboxylic acid product [14C1]GA25 (see Table S2 in the sup-plemental material). Thus, GA C-20 oxidation to form thebiologically active C19-GA GA4 is catalyzed by a P450 mono-oxygenase in S. manihoticola, as in F. fujikuroi (57).
Regulation of GA gene expression. In F. fujikuroi, GA bio-synthesis is mainly controlled by the general transcription reg-
FIG. 7. Analysis of F. fujikuroi culture filtrates. (A) TLC analysis ofthe wild type (WT), the P450-4 mutant B1-41a and B1-41a comple-mented with SmP450-1, and SmP450-4 driven by its own or the F.fujikuroi P450-4 promoter. (B) Total ion current chromatograms fromGC-MS analysis of B1-41a, B1-41a complemented with SmP450-4driven by its own promoter (SmP450-4), or the F. fujikuroi P450-4promoter (FfP4 prom::SmP4) and SmP450-1. The positions of elution ofgibberellenic acid (1), GA13 (2), isoGA3 (3), GA1 (4), and GA3 (5) aremarked.
VOL. 74, 2008 GIBBERELLIN GENE CLUSTER IN SPHACELOMA MANIHOTICOLA 5333

ulator AreA, so that expression of six of the seven genes (withFfP450-3 as the exception) is strictly repressed by largeamounts of nitrogen (e.g., ammonium, glutamine, glutamate,and nitrate) in the culture medium (36, 55). AreA directlybinds to GATA motifs in the promoter regions of the FusariumGA biosynthetic genes, triggering transcription under nitrogenstarvation conditions (36). To determine whether GA biosyn-thesis was regulated similarly in Sphaceloma, the effect of anitrogen source on the expression of the GA biosyntheticgenes was examined by Northern blot analysis. After growingthe S. manihoticola wild-type strain Lu949 in liquid OPM for 7days, equal amounts of the washed mycelium were transferredto nitrogen-free ICI medium (0% ICI) or to ICI medium with100 mM NH4NO3 (100% ICI) for 20 h. Hybridization exper-iments showed that the expression of the GA genes in S.manihoticola also depends on the level of nitrogen (Fig. 9), butto a lesser extent than in F. fujikuroi. This is consistent with thelower number of putative AreA binding sites in the Sphac-eloma promoter regions than in the Fusarium GA gene pro-moters (only 3 to 6 GATA sequence elements compared to 6to 12 in Fusarium). As in F. fujikuroi, the expression of the GAgenes was not affected by the level of the carbon source or
phosphate or the pH value (data not shown). However, therewas a detectable difference in the expression levels of the S.manihoticola genes driven by their original or the correspond-ing Fusarium promoters. In the Fusarium background, expres-sion was always better if the gene was under the control of theF. fujikuroi promoter (Fig. 6). TLC and GC-MS analyses alsorevealed higher GA production in these transformants (Fig. 7).
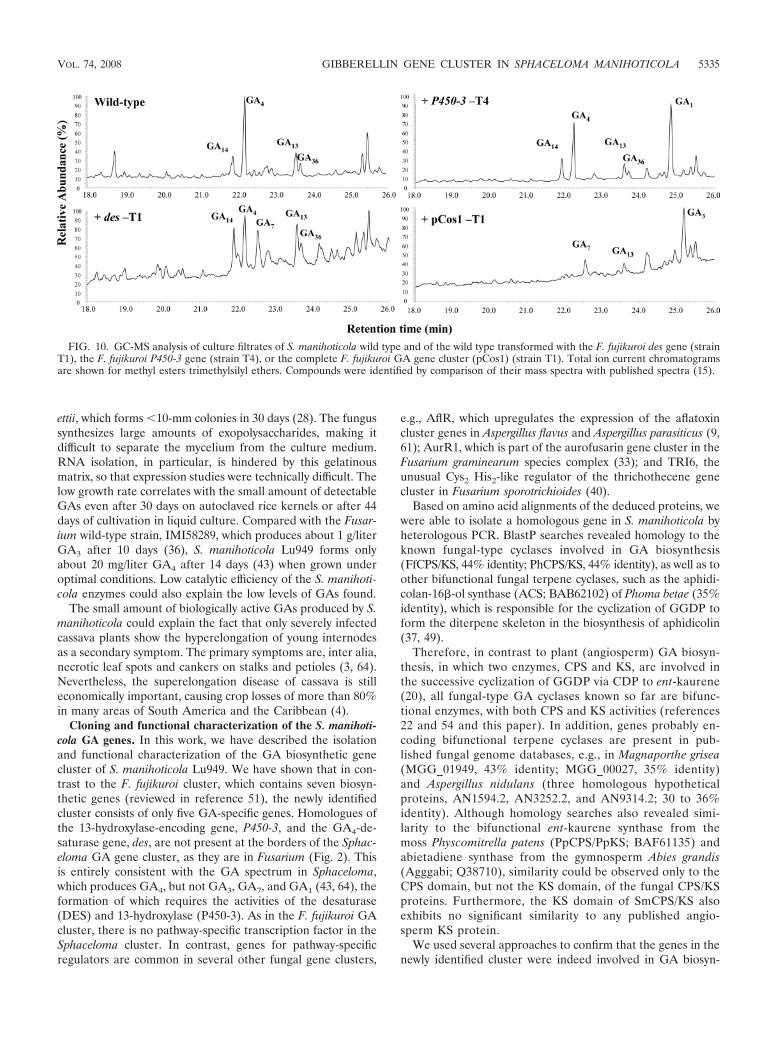
Complementation of S. manihoticola Lu949 with the genesP450-3 and des from F. fujikuroi. In order to determinewhether GA3 production could be restored in Sphaceloma, wetransformed the fungus with the cosmid pCos1 containing theentire GA gene cluster of F. fujikuroi (Linnemannstons andTudzynski, unpublished), or with vectors carrying the two sin-gle Fusarium GA genes des (porf3-Sal) and P450-3 (p450-3-GC) (58), which are absent in the Sphaceloma gene cluster. Ifthe fungus was able to express the heterologous genes from theF. fujikuroi gene cluster, pCos1 transformants of Sphacelomawould be able to produce GA7 and GA1 in addition to GA3,while Sphaceloma transformants carrying the F. fujikuroiP450-3 gene should produce GA1, and des transformantsshould form GA7 (Fig. 1). The Fusarium genes were shown byNorthern analyses and RT-PCR to be expressed in Sphaceloma(data not shown). Furthermore, GC-MS analyses of the culturefluids of the transformants confirmed GA3 synthesis in pCos1transformants while des transformants formed GA7 and P450-3transformants produced GA1, as postulated (Fig. 10).
DISCUSSION
Sphaceloma manihoticola was the second fungus after F.fujikuroi shown to produce GAs, mainly GA4 (42). The fungusis only distantly related to F. fujikuroi, making it especiallyinteresting to compare its GA biosynthetic genes, their orga-nization in the genome, their regulation, and the functions ofthe encoded enzymes with those in F. fujikuroi. However, noth-ing was known about the genetic background of GA biosyn-thesis in S. manihoticola. In order to undertake such studies, itwas first necessary to establish standard molecular methods,such as transformation and nucleic acid preparation, for thefungus.
S. manihoticola grows extremely slowly, as already describedby Zeigler et al. in 1980 (64). The strain used in our study,Lu949, takes about 5 days to build colonies 0.5 cm in diameterand is not able to overgrow the agar plate if only a piece ofmycelium is transferred as the inoculum. This has also beendescribed for isolates of the related species Sphaceloma fawc-
FIG. 8. Structures of products formed from labeled precursors. (A) Kaurenolides. (B) Fujenoic acids. (C) C-20 oxidation intermediates.
FIG. 9. Effects of various nitrogen concentrations on the expres-sion levels of the S. manihoticola GA genes cps/ks, P450-2, and ggs2.
5334 BOMKE ET AL. APPL. ENVIRON. MICROBIOL.
ettii, which forms �10-mm colonies in 30 days (28). The fungussynthesizes large amounts of exopolysaccharides, making itdifficult to separate the mycelium from the culture medium.RNA isolation, in particular, is hindered by this gelatinousmatrix, so that expression studies were technically difficult. Thelow growth rate correlates with the small amount of detectableGAs even after 30 days on autoclaved rice kernels or after 44days of cultivation in liquid culture. Compared with the Fusar-ium wild-type strain, IMI58289, which produces about 1 g/literGA3 after 10 days (36), S. manihoticola Lu949 forms onlyabout 20 mg/liter GA4 after 14 days (43) when grown underoptimal conditions. Low catalytic efficiency of the S. manihoti-cola enzymes could also explain the low levels of GAs found.
The small amount of biologically active GAs produced by S.manihoticola could explain the fact that only severely infectedcassava plants show the hyperelongation of young internodesas a secondary symptom. The primary symptoms are, inter alia,necrotic leaf spots and cankers on stalks and petioles (3, 64).Nevertheless, the superelongation disease of cassava is stilleconomically important, causing crop losses of more than 80%in many areas of South America and the Caribbean (4).
Cloning and functional characterization of the S. manihoti-cola GA genes. In this work, we have described the isolationand functional characterization of the GA biosynthetic genecluster of S. manihoticola Lu949. We have shown that in con-trast to the F. fujikuroi cluster, which contains seven biosyn-thetic genes (reviewed in reference 51), the newly identifiedcluster consists of only five GA-specific genes. Homologues ofthe 13-hydroxylase-encoding gene, P450-3, and the GA4-de-saturase gene, des, are not present at the borders of the Sphac-eloma GA gene cluster, as they are in Fusarium (Fig. 2). Thisis entirely consistent with the GA spectrum in Sphaceloma,which produces GA4, but not GA3, GA7, and GA1 (43, 64), theformation of which requires the activities of the desaturase(DES) and 13-hydroxylase (P450-3). As in the F. fujikuroi GAcluster, there is no pathway-specific transcription factor in theSphaceloma cluster. In contrast, genes for pathway-specificregulators are common in several other fungal gene clusters,
e.g., AflR, which upregulates the expression of the aflatoxincluster genes in Aspergillus flavus and Aspergillus parasiticus (9,61); AurR1, which is part of the aurofusarin gene cluster in theFusarium graminearum species complex (33); and TRI6, theunusual Cys2 His2-like regulator of the thrichothecene genecluster in Fusarium sporotrichioides (40).
Based on amino acid alignments of the deduced proteins, wewere able to isolate a homologous gene in S. manihoticola byheterologous PCR. BlastP searches revealed homology to theknown fungal-type cyclases involved in GA biosynthesis(FfCPS/KS, 44% identity; PhCPS/KS, 44% identity), as well as toother bifunctional fungal terpene cyclases, such as the aphidi-colan-16�-ol synthase (ACS; BAB62102) of Phoma betae (35%identity), which is responsible for the cyclization of GGDP toform the diterpene skeleton in the biosynthesis of aphidicolin(37, 49).
Therefore, in contrast to plant (angiosperm) GA biosyn-thesis, in which two enzymes, CPS and KS, are involved inthe successive cyclization of GGDP via CDP to ent-kaurene(20), all fungal-type GA cyclases known so far are bifunc-tional enzymes, with both CPS and KS activities (references22 and 54 and this paper). In addition, genes probably en-coding bifunctional terpene cyclases are present in pub-lished fungal genome databases, e.g., in Magnaporthe grisea(MGG_01949, 43% identity; MGG_00027, 35% identity)and Aspergillus nidulans (three homologous hypotheticalproteins, AN1594.2, AN3252.2, and AN9314.2; 30 to 36%identity). Although homology searches also revealed simi-larity to the bifunctional ent-kaurene synthase from themoss Physcomitrella patens (PpCPS/PpKS; BAF61135) andabietadiene synthase from the gymnosperm Abies grandis(Agggabi; Q38710), similarity could be observed only to theCPS domain, but not the KS domain, of the fungal CPS/KSproteins. Furthermore, the KS domain of SmCPS/KS alsoexhibits no significant similarity to any published angio-sperm KS protein.
We used several approaches to confirm that the genes in thenewly identified cluster were indeed involved in GA biosyn-
FIG. 10. GC-MS analysis of culture filtrates of S. manihoticola wild type and of the wild type transformed with the F. fujikuroi des gene (strainT1), the F. fujikuroi P450-3 gene (strain T4), or the complete F. fujikuroi GA gene cluster (pCos1) (strain T1). Total ion current chromatogramsare shown for methyl esters trimethylsilyl ethers. Compounds were identified by comparison of their mass spectra with published spectra (15).
VOL. 74, 2008 GIBBERELLIN GENE CLUSTER IN SPHACELOMA MANIHOTICOLA 5335

thesis: gene replacement of the putative Smcps/ks, functionalcomplementation of F. fujikuroi mutants, and expression ofsingle Sphaceloma genes in the well-established background ofthe F. fujikuroi mutant SG139, which lacks the entire GA genecluster (56).
The function of SmCPS/KS as the GA-specific diterpene cy-clase was confirmed by targeted disruption of the Smcps/ks geneand by expression of this gene in an F. fujikuroi cps/ks deletionmutant. Both approaches demonstrated that Smcps/ks is the S.manihoticola orthologue of Ffcps/ks: the deletion led to the totalloss of GAs and precursors in the Sphaceloma �cps/ks mutant,and the complementation of F. fujikuroi �cps/ks with Smcps/ksrestored GA biosynthesis in the mutant.
Most steps of GA biosynthesis in S. manihoticola are cata-lyzed by P450 monooxygenases, as described for F. fujikuroi(20, 51), and in contrast to plant systems, which utilize solubledioxygenases, as well as P450 enzymes (20). Three P450 mono-oxygenase-encoding genes were identified in the S. manihoti-cola GA gene cluster: SmP450-1, SmP450-2, and SmP450-4;their annotation was based on their close sequence similarity tothe F. fujikuroi GA genes, placing them in the same CYPgroup. It was, however, important to confirm their catalyticfunctions, particularly since there was an inversion of the P450-1-P450-4 unit in comparison with the F. fujikuroi cluster (Fig.2). After complementation of the F. fujikuroi mutant B1-41a,which has a point mutation in the P450-4 gene (56), withSmP450-1 or SmP450-4, only SmP450-4 restored GA biosyn-thesis in the mutant, and it is therefore the orthologue of theF. fujikuroi gene FfP450-4 (Fig. 7). Thus, SmP450-4 encodesent-kaurene oxidase, which oxidizes the C-19 methyl of ent-kaurene to a carboxylic acid to give ent-kaurenoic acid.
By heterologous expression of SmP450-1 in the clusterlessmutant F. fujikuroi SG139, we could demonstrate thatSmP450-1 is a multifunctional enzyme involved in GA14 syn-thesis from ent-kaurenoic acid. As for FfP450-1, the interme-diacy of ent-7-hydroxykaurenoic acid and GA12-aldehyde wasindicated by their efficient conversion into GA14. GA12 wasalso converted by SG139-SmP450-1 into GA14, although it isnot a main intermediate in GA14 synthesis, since mycelialmicrosomal fractions convert [14C]GA12-aldehyde, but not[14C]-GA12, into [14C]GA14 in the presence of NADPH (un-published results). This indicates that GA biosynthesis in S.manihoticola involves 3�-hydroxylation of GA12 aldehyde, as itdoes in F. fujikuroi (44). This contrasts with the situation in theGA1-producing fungus Phaeosphaeria sp., in which 3�-hy-droxylation occurs late in the pathway after formation of C19
GAs (22). It was suggested that PhP450-1 (ent-kaurenoic acidoxidase) lacked 3�-hydroxylase activity (23), although this hasnot been demonstrated experimentally, and the enzyme re-sponsible for 3�-hydroxylation in Phaeosphaeria has not yetbeen characterized.
SmP450-2 was shown to function as a 20-oxidase convertingGA14 to GA4 without accumulation of intermediates. The non-hydroxylated substrate GA12 was oxidized with lower effi-ciency, as shown by accumulation of the C-20 aldehyde productGA24, together with the C-20 alcohol GA15. This differs fromthe Fusarium enzyme FfP450-2, which oxidizes both GA14 andGA12 efficiently to give GA4 and GA9, respectively, withoutaccumulation of intermediates (57).
Regulation of gene expression. Northern blot analysis re-vealed that the Sphaceloma GA genes are already highly ex-pressed after 7 days of incubation in liquid media. However,GA accumulation could be demonstrated only after 44 days inliquid culture, perhaps due to difficulty in extracting GAs fromthe slimy culture fluid at an earlier stage.
Expression of the Sphaceloma GA genes is regulated bynitrogen, although to a lesser extent than has been demon-strated for the Fusarium genes (36, 55). The promoters of theSphaceloma GA genes contain far fewer potential AreA bind-ing sites (double GATA motifs) than the Fusarium genes. Forexample, there are only three GATA motifs in the bidirec-tional promoter shared by SmP450-1 and SmP450-4, and noneof them as a double GATA motif, which have been demon-strated to be the most important AreA binding sites in thecorresponding promoter region of F. fujikuroi (36). Neverthe-less, shift experiments revealed that large amounts of ammo-nium nitrate (100 mM) in the culture medium led to reducedexpression levels of the GA genes, compared to nitrogen-freemedium, also in S. manihoticola (Fig. 9). Since the largeamounts of exopolysaccharides also complicate shift experi-ments, impeding accurate purification of the mycelium fromthe precultivation broth, the cultivation conditions have to beoptimized in order to investigate regulation mechanisms inmore detail.
In general, the main regulatory elements appear to be con-served between Fusarium and Sphaceloma, as Sphaceloma GAgenes are expressed and the encoded enzymes are functional inthe Fusarium background and vice versa. Moreover, weshowed that the Fusarium GA genes P450-3 and des, whosehomologues are missing in Sphaceloma, can be expressed in S.manihoticola and can generate the complete biosynthetic path-way to the main F. fujikuroi end product, GA3. It is particularlynoteworthy that the Fusarium gene P450-3 is expressed and theencoded 13-hydroxylase is functional in the genetic back-ground of this distantly related fungus, but it was not expressedat detectable levels when transformed into the Fusarium mu-tant strain SG139, lacking the entire GA gene cluster, and no13-hydroxylase activity has been obtained in these transfor-mants (36, 58). Furthermore, although fusion to the strong F.fujikuroi glnA promoter enabled expression of P450-3 inSG139, enzyme functionality could not be demonstrated (C.Bomke and B. Tudzynski, unpublished data). Another pecu-liarity of P450-3 is its distinct regulation. It is the only FusariumGA cluster gene whose expression is not nitrogen repressedand therefore seems to be controlled by at least one additionaltranscription factor, other than AreA. This difference mightpoint to an origin of this gene different from those of the otherGA cluster genes and could explain the lack of a homologousgene in Sphaceloma. It is also possible that the correspondinggene was lost in Sphaceloma ancestors during evolution, al-though the possibility that a homologue is located at anotherlocus in the Sphaceloma genome, though probably in an inac-tive form, cannot be excluded.
An interesting observation is the downregulation of theother GA cluster genes, as well as the flanking genes orf1 andorf2, as a consequence of the knockout of the Smcps/ks gene.This effect is more likely due to the integration of the largerreplacement fragment than to a positive feedback regulation ofthe GA genes, because the single Sphaceloma (and Fusarium)
5336 BOMKE ET AL. APPL. ENVIRON. MICROBIOL.

P450 monooxygenase genes are highly expressed in the geneticbackground of F. fujikuroi SG139, which lacks the entire GAgene cluster (Fig. 6).
Evolution of fungal diterpene gene clusters. The differentcluster organization in Sphaceloma (inversion of the P450-1-P450-4 transcriptional unit and P450-2), as well as the lack ofthe border genes of the Fusarium GA cluster (Ffdes andFfP450-3), raises questions about the evolution of the GA geneclusters in fungi and GA biosynthesis in general. Previouscomparisons of GA biosynthesis in plants and F. fujikuroi dis-proved the hypothesis of horizontal gene transfer betweenhigher plants and fungi and suggested independent evolutionof GA biosynthesis pathways (20).
The organizations of different fungal diterpene gene clustersare very similar, suggesting a common origin. It is particularlynoticeable that a large number of diterpene gene clusters containa pathway-specific GGDP synthase-encoding gene, for example,Ffggs2 and Smggs2 in the GA biosynthetic gene clusters, PbGGSin the aphidicolin gene cluster (49), and paxG, ltmG, and atmG inthe indole-diterpene gene clusters for paxilline (63), lolitrem(63a), and aflatrem (65) biosynthesis in Penicillium paxilli, Neoty-phodium lolii, and A. flavus, respectively. The identification of thepathway-specific ggs gene in the S. manihoticola GA gene clustersupports the hypothesis that the presence of two copies of GGDPsynthase genes may be a molecular signature for diterpene bio-synthesis (65). Remarkably, the unit of a diterpene cyclase(MGG_00027) and a GGDP synthase gene (MGG_14382) isalso conserved in a putative rudimentary diterpene cluster inM. grisea. An exception is the GA gene cluster in Phaeosphaeriasp. strain L487, which does not contain a GGDP synthase gene(23). Interestingly, a couple of diterpenoid gene clusters havebeen identified in Streptomyces species that also contain aGGDP synthase-encoding gene next to cyclase genes (21, 24).However, phylogenetic studies with bacterial, plant, fungal,and animal GGDP synthases and diterpene cyclases clearlyshow that the bacterial genes are very distant from fungalgenes and share a much higher similarity with the plant en-zymes (Bomke and Tudzynski, unpublished).
In addition to the similar cluster organizations of fungalditerpene gene clusters, the conserved intron positions, espe-cially in fungal cps/ks genes (Ffcps/ks and Smcps/ks), as well asthe P. betae homologue PbACS and both Magnaporthe cps/kshomologous genes, MGG_00027 and MGG_01949, is a furtherindication of a common origin. The conservation of some in-tron positions can also be observed in the P450 monooxygen-ase genes and ggs2, although fungal P450 genes in generalexhibit low conservation of intron-exon structure (12). Sinceonly the mRNA sequence of Phcps/ks (AB003395) and that ofonly one monooxygenase (GA 20-oxidase)-encoding gene(AB106677) of Phaeosphaeria sp. strain L487 are available, it isnot possible to include these genes in this comparison.
How gene clusters are formed during evolution and theirevolutionary advantage are unknown, but there are severalhypotheses concerning cluster assembly and the putative ben-efits of clustering genes in genomes. The “selfish-gene” hy-pothesis was established to be the driving force of comobiliza-tion of these genes within species through mating or into otherspecies by horizontal gene transfer (60). The occurrence ofhorizontal gene transfer in fungi was recently demonstrated forthe fungal ACEI gene cluster (25), and it represents a convinc-
ing explanation for the existence of shared gene clusters indistantly related fungi, such as the GA gene clusters in F.fujikuroi and S. manihoticola. However, other genome events,such as gene duplications and differential gene losses, are al-ternative explanations for the discontinuous distribution offungal gene clusters (26). Thus, it remains unclear if the GAgene cluster of Sphaceloma ancestors has lost flanking geneshomologous to des and P450-3 or if an ancient Fusarium spe-cies acquired a basic GA gene cluster via horizontal genetransfer and it was coincidentally inserted next to a P450 mo-nooxygenase-encoding gene, P450-3, that could contribute toGA biosynthesis. The increasing number of sequenced ge-nomes might help to advance the theory of the evolution ofgene clusters, especially those for diterpene biosynthesis, suchas the GA gene cluster, in fungi.
Conclusions. In this paper, we describe the identificationand functional characterization of all genes involved in GAbiosynthesis in the cassava pathogen S. manihoticola. There areseveral features, such as gene structure, their organization in agene cluster, the presence of a pathway-specific GGDP syn-thase-encoding gene transcribed from the same promoter asthe cps/ks gene, and the order of enzymatic steps, that aresimilar to those in F. fujikuroi, suggesting a common origin ofGA clusters in F. fujikuroi and S. manihoticola. The Sphac-eloma genes fully complement the corresponding F. fujikuroimutants, demonstrating that general regulatory features, in-cluding nitrogen regulation, are similar in these fungi. On theother hand, there are only five genes in this gene cluster,because the flanking genes of the F. fujikuroi gene cluster areabsent in Sphaceloma. As a consequence, the biosyntheticpathway ends with GA4 instead of being further converted toGA7, GA1, and GA3, as occurs in F. fujikuroi (Fig. 11).
ACKNOWLEDGMENTS
We thank W. Rademacher (BASF Agricultural Center, Limburgerhof,Germany) for providing the strain S. manihoticola Lu949, J. Avalos (Uni-versity of Seville, Seville, Spain) for providing F. fujikuroi strains IMI58289and SG139, and J. MacMillan (University of Bristol, Bristol, United King-dom) for providing the F. fujikuroi mutant strain B1-41a.
The work was supported by the Deutsche Forschungsgemeinschaft(DFG) (Tu101/9-5, SPP 1152 “Evolution of Metabolic Diversity”) andFondo Nacional de Desarrollo Cientifico y Tecnologico (grant1061127). Rothamsted Research receives grant-aided support from the
FIG. 11. End products of the GA biosynthetic pathways in F. fu-jikuroi and S. manihoticola.
VOL. 74, 2008 GIBBERELLIN GENE CLUSTER IN SPHACELOMA MANIHOTICOLA 5337

Biotechnology and Biological Sciences Research Council of the UnitedKingdom. Furthermore, we thank the DAAD (Deutscher Akademis-cher Austausch Dienst) and CONICYT/DFG (project 105-2007) forgranting short-term scholarships (D/06/47017).
REFERENCES
1. Alexander, N. J., T. M. Hohn, and S. P. McCormick. 1998. The TRI11 geneof Fusarium sporotrichioides encodes a cytochrome P-450 monooxygenaserequired for C-15 hydroxylation in trichothecene biosynthesis. Appl. Envi-ron. Microbiol. 64:221–225.
2. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990.Basic local alignment search tool. J. Mol. Biol. 215:403–410.
3. Alvarez, E., and M. L. Molina. 2000. Characterizing the Sphaceloma fungus,causal agent of superelongation disease in cassava. Plant Dis. 84:423–428.
4. Alvarez, E., G. Llano, J. B. Loke, and J. F. Mejıa. 2005. Applying biotech-nology tools to control plant diseases. Fitopatol. Colombiana 28:93–97.
5. Atzorn, R., A. Crozier, C. T. Wheeler, and G. Sandberg. 1988. Production ofgibberellins and indole-3-acetic acid by Rhizobium phaseoli in relation tonodulation of Phaseolus vulgaris roots. Planta 175:532–538.
6. Barendse, G. W. M., P. H. van de Werken, and N. Takahashi. 1980. High-performance liquid chromatography of gibberellins. J. Chromatogr. 198:449–455.
7. Bearder, J. R., J. MacMillan, M. Wels, M. B. Chaffey, and B. O. Phinney.1974. Position of the metabolic block for gibberellin biosynthesis in mutantB1-41a of Gibberella fujikuroi. Phytochemistry 13:911–917.
8. Bearder, J. R. 1983. In vivo diterpenoid biosynthesis in Gibberella fujikuroi:the pathway after ent-Kaurene, p. 251–387. In A. Crozier (ed.), The bio-chemistry and physiology of gibberellins, vol. 1. Praeger, New York, NY.
9. Chang, P. K., J. W. Cary, J. Yu, D. Bhatnagar, and T. E. Cleveland. 1995.The Aspergillus parasiticus polyketide synthase gene pksA, a homolog ofAspergillus nidulans wA, is required for aflatoxin B1 biosynthesis. Mol. Gen.Genet. 248:270–277.
10. Church, G. M., and W. Gilbert. 1984. Genomic sequencing. Proc. Natl. Acad.Sci. USA 81:1991–1995.
11. Darken, M. A., A. A. L. Jensen, and P. Shu. 1959. Production of gibberellicacid by fermentation. Appl. Microbiol. 7:301–303.
12. Deng, J., I. Carbone, and R. A. Dean. 2007. The evolutionary history ofcytochrome P450 genes in four filamentous ascomycetes. BMC Evol. Biol.7:30.
13. Doyle, J. J., and J. L. Doyle. 1990. Isolation of plant DNA from fresh tissue.Focus 12:13–15.
14. Fernandez-Martın, R., F. Reyes, C. E. Domenech, E. Cabrera, P. M. Bramley,A. F. Barrero, J. Avalos, and E. Cerda-Olmedo. 1995. Gibberellin biosynthesis ingib mutants of Gibberella fujikuroi. J. Biol. Chem. 270:14970–14974.
15. Gaskin, P., and J. MacMillan. 1992. GC-MS of the gibberellins and relatedcompounds: methodology and a library of spectra. Cantock’s Enterprises,Bristol, United Kingdom.
16. Geissman, T. A., A. J. Verbiscar, B. O. Phinney, and G. Cragg. 1966. Studieson the biosynthesis of gibberellins from (�)-kaurenoic acid in cultures ofGibberella fujikuroi. Phytochemistry 5:933–947.
17. Gutierrez-Manero, F., B. Ramos-Solano, A. Probanza, J. Mehouachi, F. R.Tadeo, and M. Talon. 2001. The plant-growth-promoting rhizobacteriaBacillus pumilus and Bacillus licheniformis produce high amounts of physio-logically active gibberellins. Physiol. Plant. 111:206–211.
18. Hedden, P., and Y. Kamiya. 1997. Gibberellin biosynthesis: enzymes, genesand their regulation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48:431–460.
19. Hedden, P., and A. L. Phillips. 2000. Gibberellin metabolism: new insightsrevealed by the genes. Trends Plant Sci. 5:523–530.
20. Hedden, P., A. L. Phillips, M. C. Rojas, E. Carrera, and B. Tudzynski. 2002.Gibberellin biosynthesis in plants and fungi: a case of convergent evolution?J. Plant Growth Reg. 20:317–331.
21. Ikeda, C., Y. Hayashi, N. Itoh, H. Seto, and T. Dairi. 2007. Functional analysisof eubacterial ent-copalyl diphosphate synthase and pimara-9(11),15-diene syn-thase with unique primary sequences. J. Biochem. 141:37–45.
22. Kawaide, H., R. Imai, T. Sassa, and Y. Kamiya. 1997. cDNA isolation,characterization and bacterial expression of a bifunctional diterpene cyclasein fungal gibberellin biosynthesis. J. Biol. Chem. 272:21706–21712.
23. Kawaide, H. 2006. Biochemical and molecular analysis of gibberellin biosyn-thesis in fungi. Biosci. Biotechnol. Biochem. 70:583–590.
24. Kawasaki, T., T. Kuzuyama, Y. Kuwamori, N. Matsuura, N. Itoh, K. Furihata,H. Seto, and T. Dairi. 2004. Presence of copalyl diphosphate synthase gene in anactinomycete possessing the mevalonate pathway. J. Antibiot. 57:739–747.
25. Khaldi, N., J. Collemare, M.-H. Lebrun, and K. H. Wolfe. 2008. Evidence forhorizontal transfer of a secondary metabolite gene cluster between fungi.Genome Biol. 9:R18.
26. Kroken, S., N. L. Glass, J. W. Taylor, O. C. Yoder, and B. G. Turgeon. 2003.Phylogenomic analysis of type I polyketide synthase genes in pathogenic andsaprobic ascomycetes. Proc. Natl. Acad. Sci. USA 100:15670–15675.
27. Krugel, H., G. Fiedler, C. Smithand, and S. Baumberg. 1993. Sequence andtranscriptional analysis of the nourseothricin acetyltransferase-encodinggene nat1 from Streptomyces noursei. Gene 127:127–131.
28. Liao, H.-L., and K.-R. Chung. 2008. Cellular toxicity of elsinochrome phy-totoxins produced by the pathogenic fungus, Elsinoe fawcettii causing citrusscab. New Phytol. 177:239–250.
29. Linnemanstons, P., T. Voß, P. Hedden, P. Gaskin, and B. Tudzynski. 1999.Deletions in the gibberellin biosynthesis gene cluster of Gibberella fujikuroiby restriction enzyme-mediated integration and conventional transforma-tion-mediated mutagenesis. Appl. Environ. Microbiol. 65:2558–2564.
30. MacMillan, J. 2002. Occurrence of gibberellins in vascular plants, fungi, andbacteria. J. Plant Growth Reg. 20:387–442.
31. Malonek, S., M. C. Rojas, P. Hedden, P. Gaskin, and B. Tudzynski. 2004.The NADPH: cytochrome P450 reductase gene from Gibberella fujikuroi isessential for gibberellin biosynthesis. J. Biol. Chem. 279:25075–25084.
32. Malonek, S., C. Bomke, E. Bornberg-Bauer, M. C. Rojas, P. Hedden, and B.Tudzynski. 2005. Distribution of gibberellin biosynthetic genes and gibberel-lin production in the Gibberella fujikuroi species complex. Phytochemistry66:1296–1311.
33. Malz, S., M. N. Grell, C. Thrane, F. J. Maier, P. Rosager, A. Felk, K. S.Albertsen, S. Salomon, L. Bohn, W. Schafer, and H. Giese. 2005. Identifi-cation of a gene cluster responsible for the biosynthesis of aurofusarin in theFusarium graminearum species complex. Fungal Genet. Biol. 42:420–433.
34. Maniatis, T., J. Sambrook, and E. F. Fritsch. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
35. Mende, K., V. Homann, and B. Tudzynski. 1997. The geranylgeranyl diphos-phate synthase gene of Gibberella fujikuroi: isolation and expression. Mol.Gen. Genet. 255:96–105.
36. Mihlan, M., V. Homann, T.-W. D. Liu, and B. Tudzynski. 2003. AREAdirectly mediates nitrogen regulation of gibberellin biosynthesis in Gibberellafujikuroi, but its activity is not affected by NMR. Mol. Microbiol. 47:975–991.
37. Oikawa, H., T. Toyomasu, H. Toshima, S. Ohashi, H. Kawaide, Y. Kamiya,M. Ohtsuka, S. Shinoda, W. Mitsuhashi, and T. Sassa. 2001. Cloning andfunctional expression of cDNA encoding aphidicolan-16�-ol synthase: a keyenzyme responsible for formation of an unusual diterpene skeleton in bio-synthesis of DNA polymerase inhibitor aphidicolin. J. Am. Chem. Soc.123:5154–5155.
38. Phinney, B. O. 1979. Gibberellin biosynthesis in the fungus Gibberella fu-jikuroi and in higher plants, p. 57–58. In N. B. Mandava (ed.), Plant growthsubstances. ACS Symposium Series 111. American Chemical Society, Wash-ington, DC.
39. Pontecorvo, G. V., J. A. Poper, L. M. Hemmonns, K. D. MacDonald, andA. W. J. Buften. 1953. The genetics of Aspergillus nidulans. Adv. Genet.141:141–238.
40. Proctor, R. H., T. M. Hohn, S. P. McCormick, and A. E. Desjardins. 1995.Tri6 encodes an unusual zinc finger protein involved in regulation of tricho-thecene biosynthesis in Fusarium sporotrichioides. Appl. Environ. Microbiol.61:1923–1930.
41. Punt, P. J., R. P. Oliver, M. A. Dingenanse, P. H. Puwels, and C. A. M. J. J.van den Hondel. 1987. Transformation of Aspergillus nidulans based on thehygromycin B resistance marker from Escherichia coli. Gene 56:117–124.
42. Rademacher, W., and J. E. Graebe. 1979. Gibberellin A4 produced by Sphac-eloma manihoticola, the cause of the superelongation disease of cassava(Manihot esculenta). Biochem. Biosphys. Res. Commun. 91:35–40.
43. Rademacher, W. 1992. Occurrence of gibberellins in different species of thefungal genera Sphaceloma and Elsinoe. Phytochemistry 31:4155–4157.
44. Rojas, M. C., P. Hedden, P. Gaskin, and B. Tudzynski. 2001. The P450-1gene of Gibberella fujikuroi encodes a multifunctional enzyme in gibberellinbiosynthesis. Proc. Natl. Acad. Sci. USA 98:5838–5843.
45. Rojas, M. C., O. Urrutia, C. Cruz, P. Gaskin, B. Tudzynski, and P. Hedden.2004. Kaurenolides and fujenoic acids are side products of the P450-1 mono-oxygenase in Gibberella fujikuroi. Phytochemistry 65:821–830.
46. Sanger, F., S. Nicklen, and A. R. Coulson. DNA sequencing with chain-terminating inhibitors. 1977. Proc. Natl. Acad. Sci. USA 74:5463–5467.
47. Sassa, T., K. Suzuki, and E. Haruki. 1989. Isolation and identification ofgibberellins A4 and A9 from a fungus Phaeosphaeria sp. Agric. Biol. Chem.53:303–304.
48. Sun, T.-P., and Y. Kamiya. 1994. The Arabidopsis GA1 locus encodes thecyclase ent-kaurene synthetase A of gibberellin biosynthesis. Plant Cell6:1509–1518.
49. Toyomasu, T., K. Nakaminami, H. Toshima, T. Mie, K. Watanabe, H. Ito, H.Matsui, W. Mitsuhashi, T. Sassa, and H. Oikawa. 2004. Cloning of a genecluster responsible for the biosynthesis of diterpene aphidicolin, a specificinhibitor of DNA polymerase alpha. Biosci. Biotechnol. Biochem. 68:146–152.
50. Troncoso, C., J. Carcamo, P. Hedden, B. Tudzynski, and M. C. Rojas. 2008.Influence of electron transport proteins on the reactions catalyzed by Fusar-ium fujikuroi gibberellin monooxygenases. Phytochemistry 69:672–683.
51. Tudzynski, B. 2005. Gibberellin biosynthesis in fungi: genes, enzymes, evolution,and impact on biotechnology. Appl. Microbiol. Biotechnol. 66:597–611.
52. Tudzynski, B., and K. Holter. 1998. The gibberellin biosynthetic pathway inGibberella fujikuroi: evidence or a gene cluster. Fungal Genet. Biol. 25:157–170.
53. Tudzynski, B., K. Mende, K.-M. Weltring, S. E. Unkles, and J. R. Kinghorn.
5338 BOMKE ET AL. APPL. ENVIRON. MICROBIOL.

1996. The Gibberella fujikuroi niaD gene encoding nitrate reductase: isola-tion, sequence, homologous transformation and electrophoretic karyotypelocation. Microbiology 142:533–539.
54. Tudzynski, B., H. Kawaide, and Y. Kamiya. 1998. Gibberellin biosynthesis inGibberella fujikuroi: cloning and characterization of the copalyl diphosphatesynthase gene. Curr. Genet. 34:234–240.
55. Tudzynski, B., V. Homann, B. Feng, and G. A. Marzluf. 1999. Isolation,characterization and disruption of the areA nitrogen regulatory gene ofGibberella fujikuroi. Mol. Gen. Genet. 261:106–114.
56. Tudzynski, B., P. Hedden, E. Carrera, and P. Gaskin. 2001. The P450-4 geneof Gibberella fujikuroi encodes ent-kaurene oxidase in the gibberellin biosyn-thetic pathway. Appl. Environ. Microbiol. 67:3514–3522.
57. Tudzynski, B., M. C. Rojas, P. Gaskin, and P. Hedden. 2002. The Gibberellafujikuroi gibberellin 20-oxidase is a multifunctional monooxygenase. J. Biol.Chem. 277:21246–21253.
58. Tudzynski, B., M. Mihlan, M. C. Rojas, P. Linnemannstons, P. Gaskin, andP. Hedden. 2003. Characterization of the final two genes of the gibberellinbiosynthesis gene cluster of Gibberella fujikuroi: des and P450-3 encode GA4desaturase and the 13-hydroxylase, respectively. J. Biol. Chem. 278:28635–28643.
59. Urrutia, O., P. Hedden, and M. C. Rojas. 2001. Monooxygenases involved inGA12 and GA14 synthesis in Gibberella fujikuroi. Phytochemistry 56:505–511.
60. Walton, J. D. 2000. Horizontal gene transfer and the evolution of secondarymetabolite gene clusters in fungi: an hypothesis. Fungal Genet. Biol. 30:167–171.
61. Woloshuk, C. P., K. R. Foutz, J. F. Brewer, D. Bhatnagar, T. E. Cleveland,and G. A. Payne. 1994. Molecular characterization of aflR, a regulatory locusfor aflatoxin biosynthesis. Appl. Environ. Microbiol. 60:2408–2414.
62. Yamaguchi, S., T.-P. Sun, H. Kawaide, and Y. Kamiya. 1998. The GA2 locusof Arabidopsis thaliana encodes ent-kaurene synthase of gibberellin biosyn-thesis. Plant Physiol. 116:1271–1278.
63. Young, C. A., L. McMillan, E. Telfer, and B. Scott. 2001. Molecular cloningand genetic analysis of an indole-diterpene gene cluster from Penicilliumpaxilli. Mol. Microbiol. 39:754–764.
63a.Young, C. A., S. Felitti, K. Shields, G. Spangenberg, R. D. Johnson, G. T.Bryan, S. Saikia, and B. Scott. 2006. A complex gene cluster for indole-diterpene biosynthesis in the grass endophyte Neotyphodium lolii. FungalGenet. Biol. 43:679–693.
64. Zeigler, R. S., L. E. Powell, and H. D. Thurston. 1980. Gibberellin A4production by Sphaceloma manihoticola, causal agent of cassava superelon-gation disease. Phytopathology 70:589–593.
65. Zhang, S., B. J. Monahan, J. S. Tkacz, and B. Scott. 2004. Indole-diterpenegene cluster from Aspergillus flavus. Appl. Environ. Microbiol. 70:6875–6883.
VOL. 74, 2008 GIBBERELLIN GENE CLUSTER IN SPHACELOMA MANIHOTICOLA 5339




















