
Arabidopsis RNA-binding protein UBA2a relocalizes into nuclear speckles in response to abscisic acid
6
Arabidopsis RNA-binding protein UBA2a relocalizes into nuclear speckles in response to abscisic acid Marta Riera * , Yulia Redko, Jeffrey Leung Institut des Sciences du Ve ´ge ´tal, Centre National de la Recherche Scientifique UPR2355, Av de la Terrasse 91198 Gif-sur-Yvette, France Received 11 April 2006; revised 6 June 2006; accepted 20 June 2006 Available online 30 June 2006 Edited by Ulf-Ingo Flu ¨gge Abstract The Arabidopsis thaliana RNA binding protein UBA2a is the closest homologue of the Vicia faba AKIP1 (56% identity). Like AKIP1, UBA2a is a constitutively-ex- pressed nuclear protein and in response to ABA it is also reorga- nized within the nucleus in ‘‘speckles’’ suggesting a possible role of this protein in the regulation of mRNA metabolism during ABA signaling. AKIP1 interacts with, and is phosphorylated by, the upstream ABA-activated protein kinase AAPK. We have investigated if a pathway similar to that described in Vicia faba also exists in Arabidopsis. Our results showed that despite the resemblance between the corresponding Vicia and Arabidopsis proteins, it appears that the function of UBA2a is independent of OST1 phosphorylation. Ó 2006 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. Keywords: Abscisic acid; RNA binding protein; OST1 kinase; Arabidopsis thaliana 1. Introduction Abscisic acid (ABA) is a plant hormone that regulates essen- tial processes in growth and development, including seed mat- uration or adaptive responses to drought stress [1,2]. With increasing frequency, novel data indicate that RNA metabo- lism underpins many ABA-responsive pathways [3]. In partic- ular, several genes with homology to RNA-binding proteins have been identified in Arabidopsis mutants altered in ABA sensitivity. These include, for example, the mRNA cap-binding protein ABH1 [4], the Sm-like small nuclear ribonucleoprotein SAD1 [5], or HYL1, which binds specifically double-stranded RNA in vitro [6]. Significantly, the RNA-binding protein FCA which controls flowering time has also been described re- cently as an ABA receptor [7], reinforcing the importance of RNA processing to ABA signalling. Characterization of RNA-binding proteins involved in ABA response might pro- vide insights on how they coordinately regulate sets of mRNA during stress adaptation, a mechanism that is conceptually similar to that of operons in bacteria. The physiological context in which ABA signals modify post-transcriptional events has been established in the greatest detail in Vicia faba through the characterization of AAPK interacting protein (AKIP1), an RNA-binding protein with homology to heterogeneous nuclear RNA-binding proteins A/B and D. AKIP is constitutively nuclear, but in response to ABA it is reorganized within the nucleus in ‘‘speckles’’ [8,9], discrete cytological entities that are thought to function as mRNA storage/processing factories. AKIP1 interacts with and is phosphorylated by AAPK (ABA activated protein ki- nase) and once activated it is able to bind a dehydrin mRNA [8]. The protein kinase OPEN STOMATA1 (OST1, also known as SNRK2E and SnRK2.6) is the Arabidopsis ortho- logue of AAPK. Like AAPK, OST1 is activated by ABA and is a key regulator of stomatal closure in response to osmo- tic stress [10–13]. However, it was not known if OST1 could be involved in post-transcriptional regulation. The closest Arabid- opsis homologue of the Vicia faba AKIP1 is UBP1 interacting protein 2a (UBA2a) (56% identity). UBA2a has been described as the interacting partner of UBP1, also an RNA-binding pro- tein [14]. Whereas UBP1 is able to stimulate pre-mRNA splic- ing in transfected protoplasts using reporter constructs, the role of the UBA2a protein in RNA metabolism remains un- known. UBA2a belongs to a family of three related genes, UBA2a, UBA2b, and UBA2c. However, analyses of the UBA2 proteins have not been systematic and none has been studied in the context of physiological functions. Here we tested the possibility that UBA2a might be a downstream tar- get of OST1, and that a pathway similar to that described in Vicia faba might also exist in Arabidopsis. 2. Materials and methods Arabidopsis of the Col-0 accession was used throughout for the prep- aration of protoplasts and transgenic lines. All of the primers used in the present study are described in supplementary data (Table 1). The cDNAs of UBA2a (AGI Accession Number AT3G56860) cloned in pGEX2T, pDEDH and pGBT9 were provided by Z. Lorkovic (Max F Perutz Laboratories, Vienna, Austria). 2.1. Expression of the UBA2 genes The expression of the UBA2 genes was semi-quantified by RT-PCR in leaves from 10-days old plantlets, grown in soil under controlled conditions in growth chambers (200 lE/m 2 /s; 24 °C, 16 h light, 50% humidity) either well-watered or withheld from water for 6 days. To- tal RNA was extracted using QIAGEN purification kits. For cDNA synthesis, 5 lg of total RNA were reverse transcribed using Power- Abbreviations: ABA, abscisic acid; UBA2a, UBP1 interacting protein 2a; OST1, open stomata 1; AAPK, ABA activated kinase; AKIP1, AAPK interacting protein * Corresponding author. Present address: Unite ´ de Mycologie Mole ´- culaire, Institut Pasteur, 25 rue du Dr-Roux, 75724 Paris Cedex 15, France. E-mail address: [email protected] (M. Riera). 0014-5793/$32.00 Ó 2006 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2006.06.064 FEBS Letters 580 (2006) 4160–4165
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Arabidopsis RNA-binding protein UBA2a relocalizes into nuclear speckles in response to abscisic acid

FEBS Letters 580 (2006) 4160–4165
Arabidopsis RNA-binding protein UBA2a relocalizes into nuclearspeckles in response to abscisic acid
Marta Riera*, Yulia Redko, Jeffrey Leung
Institut des Sciences du Vegetal, Centre National de la Recherche Scientifique UPR2355, Av de la Terrasse 91198 Gif-sur-Yvette, France
Received 11 April 2006; revised 6 June 2006; accepted 20 June 2006
Available online 30 June 2006
Edited by Ulf-Ingo Flugge
Abstract The Arabidopsis thaliana RNA binding proteinUBA2a is the closest homologue of the Vicia faba AKIP1(56% identity). Like AKIP1, UBA2a is a constitutively-ex-pressed nuclear protein and in response to ABA it is also reorga-nized within the nucleus in ‘‘speckles’’ suggesting a possible roleof this protein in the regulation of mRNA metabolism duringABA signaling. AKIP1 interacts with, and is phosphorylatedby, the upstream ABA-activated protein kinase AAPK. We haveinvestigated if a pathway similar to that described in Vicia fabaalso exists in Arabidopsis. Our results showed that despite theresemblance between the corresponding Vicia and Arabidopsisproteins, it appears that the function of UBA2a is independentof OST1 phosphorylation.� 2006 Federation of European Biochemical Societies. Publishedby Elsevier B.V. All rights reserved.
Keywords: Abscisic acid; RNA binding protein; OST1 kinase;Arabidopsis thaliana
1. Introduction
Abscisic acid (ABA) is a plant hormone that regulates essen-
tial processes in growth and development, including seed mat-
uration or adaptive responses to drought stress [1,2]. With
increasing frequency, novel data indicate that RNA metabo-
lism underpins many ABA-responsive pathways [3]. In partic-
ular, several genes with homology to RNA-binding proteins
have been identified in Arabidopsis mutants altered in ABA
sensitivity. These include, for example, the mRNA cap-binding
protein ABH1 [4], the Sm-like small nuclear ribonucleoprotein
SAD1 [5], or HYL1, which binds specifically double-stranded
RNA in vitro [6]. Significantly, the RNA-binding protein
FCA which controls flowering time has also been described re-
cently as an ABA receptor [7], reinforcing the importance of
RNA processing to ABA signalling. Characterization of
RNA-binding proteins involved in ABA response might pro-
vide insights on how they coordinately regulate sets of mRNA
Abbreviations: ABA, abscisic acid; UBA2a, UBP1 interacting protein2a; OST1, open stomata 1; AAPK, ABA activated kinase; AKIP1,AAPK interacting protein
*Corresponding author. Present address: Unite de Mycologie Mole-culaire, Institut Pasteur, 25 rue du Dr-Roux, 75724 Paris Cedex 15,France.E-mail address: [email protected] (M. Riera).
0014-5793/$32.00 � 2006 Federation of European Biochemical Societies. Pu
doi:10.1016/j.febslet.2006.06.064
during stress adaptation, a mechanism that is conceptually
similar to that of operons in bacteria.
The physiological context in which ABA signals modify
post-transcriptional events has been established in the greatest
detail in Vicia faba through the characterization of AAPK
interacting protein (AKIP1), an RNA-binding protein with
homology to heterogeneous nuclear RNA-binding proteins
A/B and D. AKIP is constitutively nuclear, but in response
to ABA it is reorganized within the nucleus in ‘‘speckles’’
[8,9], discrete cytological entities that are thought to function
as mRNA storage/processing factories. AKIP1 interacts with
and is phosphorylated by AAPK (ABA activated protein ki-
nase) and once activated it is able to bind a dehydrin mRNA
[8]. The protein kinase OPEN STOMATA1 (OST1, also
known as SNRK2E and SnRK2.6) is the Arabidopsis ortho-
logue of AAPK. Like AAPK, OST1 is activated by ABA
and is a key regulator of stomatal closure in response to osmo-
tic stress [10–13]. However, it was not known if OST1 could be
involved in post-transcriptional regulation. The closest Arabid-
opsis homologue of the Vicia faba AKIP1 is UBP1 interacting
protein 2a (UBA2a) (56% identity). UBA2a has been described
as the interacting partner of UBP1, also an RNA-binding pro-
tein [14]. Whereas UBP1 is able to stimulate pre-mRNA splic-
ing in transfected protoplasts using reporter constructs, the
role of the UBA2a protein in RNA metabolism remains un-
known. UBA2a belongs to a family of three related genes,
UBA2a, UBA2b, and UBA2c. However, analyses of the
UBA2 proteins have not been systematic and none has been
studied in the context of physiological functions. Here we
tested the possibility that UBA2a might be a downstream tar-
get of OST1, and that a pathway similar to that described in
Vicia faba might also exist in Arabidopsis.
2. Materials and methods
Arabidopsis of the Col-0 accession was used throughout for the prep-aration of protoplasts and transgenic lines. All of the primers used inthe present study are described in supplementary data (Table 1). ThecDNAs of UBA2a (AGI Accession Number AT3G56860) cloned inpGEX2T, pDEDH and pGBT9 were provided by Z. Lorkovic (MaxF Perutz Laboratories, Vienna, Austria).
2.1. Expression of the UBA2 genesThe expression of the UBA2 genes was semi-quantified by RT-PCR
in leaves from 10-days old plantlets, grown in soil under controlledconditions in growth chambers (200 lE/m2/s; 24 �C, 16 h light, 50%humidity) either well-watered or withheld from water for 6 days. To-tal RNA was extracted using QIAGEN purification kits. For cDNAsynthesis, 5 lg of total RNA were reverse transcribed using Power-
blished by Elsevier B.V. All rights reserved.

OST1 KIN1 EF1α
UBA2cUBA2bUBA2aC S C S C S
C S C S C S
Fig. 1. Gene regulation of UBA2 and OST1 genes under progressivestress conditions. Total RNA was extracted from A. thaliana ecotypeColumbia plants grown in soil under standard conditions (C) orstressed plants not watered during 6 days (S) and semi-quantitativeRT-PCR was done using specific primers. Constitutive EF1a and stressinduced KIN1 genes were used as controls.
M. Riera et al. / FEBS Letters 580 (2006) 4160–4165 4161
Script RT (Clontech) according the manufacturer’s protocol. As neg-ative controls, each PCR reaction was performed in parallel withoutRT. The following conditions were used for PCR reactions: UBA2a:26 cycles, T annealing at 58 �C; UBA2b and OST1: 26 cycles, Tannealing at 56.4 �C; UBA2c: 32 cycles, T annealing at 51 �C;EF1a 22 cycles, T annealing at 58 �C; KIN1: 32 cycles, T annealingat 48 �C.
2.2. Protoplast transformation, screening of T-DNA mutants, production
and analysis of stable transgenic ArabidopsisThe protoplasts were prepared and transformed with constructs
pDEDH-35S::UBA2aGFP [14] using the PEG-mediated method [15].To produce stable transgenic Arabidopsis plants, the 35S::UBA2afused either to GFP or HA were released from the correspondingpDEDH constructs [14] by digestion with EcoRI and recloned in thebinary vector pGREEN0229 containing the plant selection markerBASTA [16]. The 3 0-UTR of UBA2a was retained as terminator topreserved potential post-transcriptional regulatory signals. These con-structs were introduced into Arabidopsis by Agrobacterium-mediatedtransformation [17]. Soil-grown transgenic plants were selected byspraying with a gardening herbicide (Contour� from a local gardeningcenter) containing the active selection ingredient Glufosinate (used at afinal concentration of 60 mg/L), or in case of in vitro selection, germi-nation on MS-containing agar with glufosinate ammonium (10–15 mg/L; Sigma). The fusion protein UBA2a-GFP was visualized in protop-lasts and in transgenic plants by using a Leica TCS SP confocal laser-scanning microscope (Leica, Heidelberg). For ABA treatment, abscisicacid (mixed isomers, Sigma) was diluted from 10 mM stock solutionprepared in methanol; equivalent volumes of methanol were includedin the ABA free controls. Five-day old seedlings grown in MS mediumwere transferred to similar plates containing 10 lM ABA for 30 min.For Western analysis, protein extraction and blotting were done as de-scribed [18], using the rat monoclonal anti-HA antibody 3F10 (Amer-sham).
Mutants homozygous for the T-DNA insertion into the UBA2a gene(SALK 53281) were selected and confirmed by PCR using specific oli-gonucleotide primers.
2.3. Expression of recombinant proteins in E. coli and in vitro
phosphorylation assaysSince the UBA2 genes contain no introns, UBA2b (AT2G41060)
and UBA2c genes (AT3G15010) were amplified from genomicDNA, cloned into the corresponding EcoRI/XhoI sites in pTOPO(Invitrogen) and confirmed by sequencing. For protein expression,UBA2b and UBA2c were cloned in-frame with the glutathione-S-transferase (GST) in pGEX-2T. The fusion proteins were expressedin E. coli and purified according the manufacturer’s protocol (Amer-sham). The purified histidine tagged OST1 kinase produced in E coliwas a gift from C. Belin and S. Thomine (ISV, Gif-surYvette). Forin vitro kinase assay, 100 ng of purified OST1 kinase and 500 ng ofeach of the UBA2 proteins were used. The reaction was carried outin 20 mM Hepes pH 8, 0.5% Triton X-100, 2 mM MnCl2, 10 mMNaF, 5 mM b-glycerophosphate, 10 mM orthovanadate, completeprotease inhibitors cocktail tablets (Roche) and 3 lCi of c-[32P]ATP in a final volume of 25 ll. The reactions were stopped by add-ing the Laemmli buffer after 30 min of incubation at 30 �C and thesamples were separated by SDS–PAGE and visualized by autoradi-ography. Western blot, using anti-GST antibody was done asdescribed [18].
2.4. Directed two-hybrid interactionThe cDNAs encoding proteins to be tested for interaction by the
yeast two-hybrid were cloned in-frame with the respective GAL4 do-mains in both the vectors pGBKT7 and pGADT7. UBA2a was re-leased from pGBT9 [14], and UBA2b from pTOPO and cloned asEcoRI/XhoI fragments. The UBA2c gene was amplified by PCR fromgenomic DNA and cloned as an NdeI/SalI fragment, while the cDNAof OST1 was PCR-amplified from leaves and cloned as EcoRI and SalIfragments. The plasmids were transformed into the yeast strain AH109according to the supplier’s instructions (Clontech) and transformantswere selected for prototrophy on Drop Out medium without leucine,thiamine, (-L-T) and for protein interaction based on prototrophyon the same selective medium without histidine and adenine (-L-T-H-A).
3. Results
3.1. Expression of the UBA2 and OST1 genes under progressive
stress conditions
Since the AAPK-AKIP pathway has been described in Vicia
guard cells and the Arabidopsis AAPK homolog OST1 is in-
volved in regulation of stomatal aperture [8,11], we checked
by semi-quantitative RT-PCR the expression in leaves of the
three AKIP-homologous genes encoding UBA2a, UBA2b
and UBA2c and that for the OST1 kinase (Fig. 1). The tran-
script levels of the UBA2 genes remained constant when plants
were submitted to a progressive drought regime over a 6-day-
period. However, the expression of the OST1 gene is enhanced
under these same long-term stress conditions.
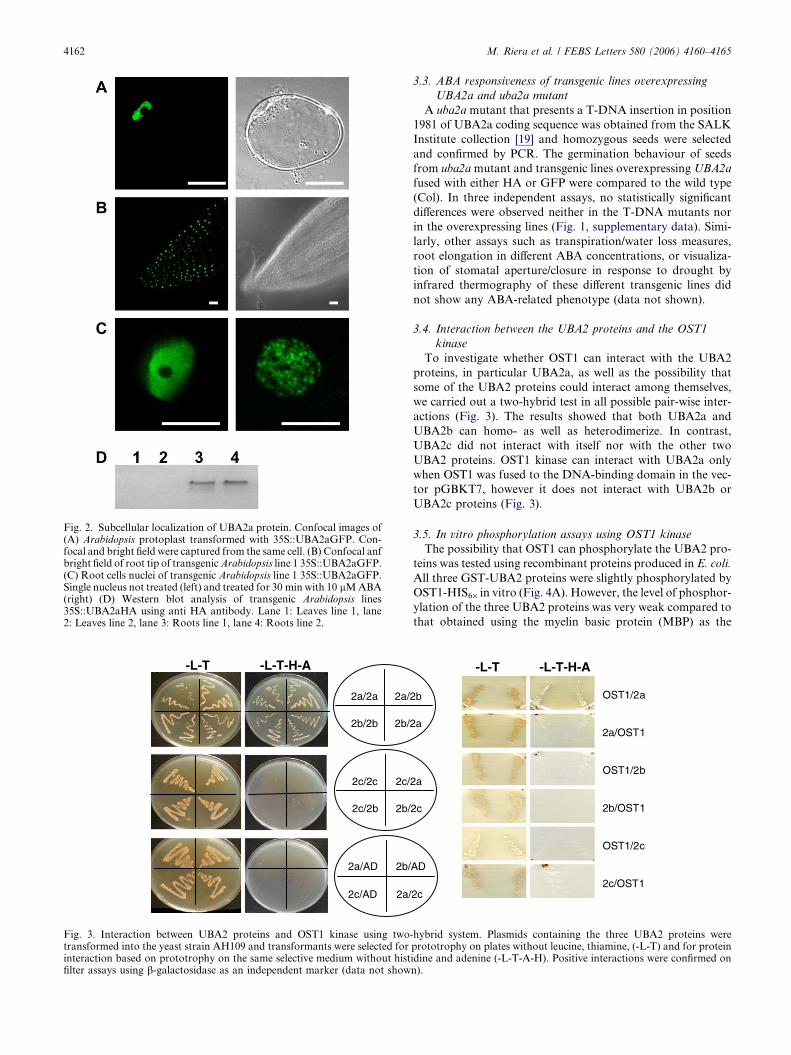
3.2. The UBA2a protein relocalizes into ‘‘nuclear speckles’’ in
response to ABA
To study the regulation of the protein UBA2a in plants,
transient expression of the protein fused to GFP in Arabidopsis
protoplasts was carried out. Our results indicated that the fu-
sion protein is largely or exclusively nuclear and it is noticeable
that the protein is excluded from the nucleolar structures
(Fig. 2A). This subcellular localization is confirmed in stable
transgenic plants (Fig. 2C, left). Observation by confocal
microscopy revealed that UBA2a-GFP expressed under the
35S RNA promoter of CaMV is highly abundant in the nuclei
of root cells (Fig. 2B). Although the overall expression is weak-
er in leaves, UBA2a-GFP seemed nonetheless enriched in
guard cells (data not shown). The much higher accumulation
of the UBA2a protein in the root relative to the arial parts
was confirmed by immunoblotting analyses using an anti-HA
specific antibody of two independent transgenic Arabidopsis
lines also expressing UBA2a-HA under the 35S promoter
(Fig. 2D). To check if UBA2a is responsive to ABA, transgenic
plants were treated with ABA (10 lM for 30 min). The
UBA2a-GFP signal was observed to coalesce into structures
resembling ‘speckles’ (Fig. 2C, right) similar to those described
for AKIP1 in Vicia faba guard cells [8,9].
-L-T -L-T-H-A
2a/2a 2a/
2b/2b 2b/
2c/2c 2c/
2c/2b 2b/
2a/
2a/AD
2c/AD
2b/
Fig. 3. Interaction between UBA2 proteins and OST1 kinase using twotransformed into the yeast strain AH109 and transformants were selected forinteraction based on prototrophy on the same selective medium without histfilter assays using b-galactosidase as an independent marker (data not show
Fig. 2. Subcellular localization of UBA2a protein. Confocal images of(A) Arabidopsis protoplast transformed with 35S::UBA2aGFP. Con-focal and bright field were captured from the same cell. (B) Confocal anfbright field of root tip of transgenic Arabidopsis line 1 35S::UBA2aGFP.(C) Root cells nuclei of transgenic Arabidopsis line 1 35S::UBA2aGFP.Single nucleus not treated (left) and treated for 30 min with 10 lM ABA(right) (D) Western blot analysis of transgenic Arabidopsis lines35S::UBA2aHA using anti HA antibody. Lane 1: Leaves line 1, lane2: Leaves line 2, lane 3: Roots line 1, lane 4: Roots line 2.
4162 M. Riera et al. / FEBS Letters 580 (2006) 4160–4165
3.3. ABA responsiveness of transgenic lines overexpressing
UBA2a and uba2a mutant
A uba2a mutant that presents a T-DNA insertion in position
1981 of UBA2a coding sequence was obtained from the SALK
Institute collection [19] and homozygous seeds were selected
and confirmed by PCR. The germination behaviour of seeds
from uba2a mutant and transgenic lines overexpressing UBA2a
fused with either HA or GFP were compared to the wild type
(Col). In three independent assays, no statistically significant
differences were observed neither in the T-DNA mutants nor
in the overexpressing lines (Fig. 1, supplementary data). Simi-
larly, other assays such as transpiration/water loss measures,
root elongation in different ABA concentrations, or visualiza-
tion of stomatal aperture/closure in response to drought by
infrared thermography of these different transgenic lines did
not show any ABA-related phenotype (data not shown).
3.4. Interaction between the UBA2 proteins and the OST1
kinase
To investigate whether OST1 can interact with the UBA2
proteins, in particular UBA2a, as well as the possibility that
some of the UBA2 proteins could interact among themselves,
we carried out a two-hybrid test in all possible pair-wise inter-
actions (Fig. 3). The results showed that both UBA2a and
UBA2b can homo- as well as heterodimerize. In contrast,
UBA2c did not interact with itself nor with the other two
UBA2 proteins. OST1 kinase can interact with UBA2a only
when OST1 was fused to the DNA-binding domain in the vec-
tor pGBKT7, however it does not interact with UBA2b or
UBA2c proteins (Fig. 3).
3.5. In vitro phosphorylation assays using OST1 kinase
The possibility that OST1 can phosphorylate the UBA2 pro-
teins was tested using recombinant proteins produced in E. coli.
All three GST-UBA2 proteins were slightly phosphorylated by
OST1-HIS6· in vitro (Fig. 4A). However, the level of phosphor-
ylation of the three UBA2 proteins was very weak compared to
that obtained using the myelin basic protein (MBP) as the
-L-T -L-T-H-A
2b
2a
2a
2c
2c
AD
OST1/2a
2a/OST1
OST1/2b
2b/OST1
OST1/2c
2c/OST1
-hybrid system. Plasmids containing the three UBA2 proteins wereprototrophy on plates without leucine, thiamine, (-L-T) and for proteinidine and adenine (-L-T-A-H). Positive interactions were confirmed onn).

50
75
100 **
OS
T1
OS
T1+
UB
A2a
UB
A2a
50
75
100
** *
OS
T1
OS
T1+
UB
A2b
OS
T1+
UB
A2c
UB
A2b
UB
A2c
75
100
UB
A2a
UB
A2b
UB
A2c
100
37
50
75
OS
T1
OS
T1+
AB
A
OS
T1
OS
T1+
AB
A
GST MBP
OS
T1
OS
T1+
AB
A
OS
T1
OS
T1+
AB
A
GST MBP
A
B
Fig. 4. In vitro phosphorylation assays using the recombinant proteins UBA2-GST and OST1-His kinase. (A) In vitro phosphorylation of UBA2proteins by OST1 kinase is shown and phosphorylated proteins are detected by autoradiography (right and central panels). Asterisks show positionsof the UBA2 proteins. Highly-phosphorylated band about 50 kDa corresponds to autophosphorylation of the OST1 kinase. The phosphorylatedproducts in these experiments were analyzed in the same gel and for the same duration of exposure. Only the relevant lanes are shown. Left panelshows Western blot of the UBA2 proteins loaded in the assay using anti-GST antibody. The two bands represent the intact proteins and theproteolytic products. (B) Control in vitro phosphorylation assays using MBP or GST as substrates. ABA (50 lM) was added to the in vitro proteinphosphorylation assays (lanes 2, 4) without modifying the OST1 kinase activity. Left panel corresponds to the autoradiography of the gel and rightpanel is the Western blot using anti-GST antibody.
M. Riera et al. / FEBS Letters 580 (2006) 4160–4165 4163
in vitro substrate (Fig. 4B). Furthermore, we did not observe an
improvement in the phosphorylation signal by varying the pH
or the ion (Mg2+ and or Mn2+) concentrations. The catalytic
activities of the Vicia AAPK and the OST1 isolated from plant
cells are stimulated by exogenous ABA [8,11–13]. In our case,
the efficiency of in vitro phosphorylation towards either MBP
or UBA2 proteins was not significantly enhanced by using
OST1-HA recovered from transfected Arabidopsis cells pre-
treated with exogenous ABA (data not shown). Similar nega-
tive results were also obtained by direct ABA treatment of
OST1-His6· produced in E. coli (Fig. 4B). Thus, under our
experimental conditions, OST1 has only weak and most likely
non-physiological kinase activity towards the three UBA2
proteins.
4. Discussion
Based on its close homology to Vicia faba AKIP1, we decided
to investigate the possibility that UBA2 proteins may have a
role in ABA signal transduction. Few experimental data are
available for the Arabidopsis UBA2 family. A systematic classi-
fication of plant RNA binding proteins includes the UBA2
family in the oligouridylate-specific group of RRM proteins
[20]. A common feature of this group of nuclear RRM proteins
(that also includes UBP1, RBP45, RBP47, UBA1 families) is
their in vitro specificity for oligouridylates. Neither UBA1
nor UBA2 proteins have obvious orthologues in metazoan gen-
omes and no physiological role for the UBA2 proteins has been
demonstrated so far. We first tested the expression of the UBA
genes in leaves and we found that under stress conditions, the
expression of the UBA2 genes does not change whereas that
of the OST1 gene is upregulated. It was known previously that
the level of the OST1 transcript is refractory to short treatments
of ABA [11] but here we showed that the transcript accumu-
lated in response to long progressive drought.
To study the physiological role of UBA2a in Arabidopsis, we
constructed transgenic lines expressing the UBA2a protein
fused to either the GFP or the HA epitope tags. We found that
the protein is constitutively nuclear and is mainly detectable in
root cells rather than in leaves of Arabidopsis, even though the
gene was expressed from the constitutive 35S promoter. This
suggests some form of post-transcriptional regulation affecting
the differential translatability of the mRNA and/or the stability
of the protein. When the Arabidopsis transgenic plants express-
ing UBA2a-GFP were treated with ABA UBA2a-GFP re-
sponded by reorganization into multiple foci resembling
nuclear speckles. However, immunoblotting experiments
showed that ABA has no significant influence on the stability
of the protein (1–10 lM ABA for 1–24 h; data not shown).

4164 M. Riera et al. / FEBS Letters 580 (2006) 4160–4165
In other organisms, these interchromatic structures have also
been referred to as stress granules and are thought to be in-
volved in RNA storage and processing [21] thus suggesting a
possible role of the UBA2a protein in the regulation of mRNA
metabolism in response to stress or ABA. Work is in progress
to define more precisely the role of UBA2a in mRNA metab-
olism through the isolation of its mRNA targets to determine
whether some of them encode known elements in stress or
ABA signaling.
Neither the transgenic lines overexpressing UBA2a nor the
uba2a T-DNA mutant showed any obvious ABA-related phe-
notype. In the uba2a mutant, this absence of phenotype could
be due to a compensatory effect provided by UBA2b (highly
homologue to UBA2a).
Interactions between the different UBA2 proteins were
tested by using the two-hybrid system. Many RNA-binding
proteins form homo- and heterodimers. The different subunit
compositions in the complex may contribute to the binding
specificity of the proteins. It was previously described that
UBA2a is able to form homodimers [14]; however, nothing
was known about UBA2b or UBA2c. Results obtained here
demonstrated that only UBA2a and UBA2b but not UBA2c
could form homo and heterodimers. Thus UBA2b, like its
close homolog UBA2a, could also be found in a complex with
UBP1 [14]. In the case of UBA2c, the absence of the acidic N-
terminal domain present in UBA2a and 2b could be a reason
for this lack of interaction with the others. In Vicia faba
AKIP1 protein interacts with AAPK kinase in two-hybrid sys-
tem [8]. In this same assay we did not detect interaction be-
tween OST1 and UBA2b or UBA2c proteins, but we were
able to demonstrate a weak interaction between OST1 and
UBA2a protein only when OST1 is cloned in-frame with the
yeast GAL4 DNA-binding domain. One possibility is that
the affinity of OST1 for UBA2a is relatively weak and can
be disrupted by subtle changes in the conformations of either
one or both of the partner proteins caused by the different fu-
sion tags. This interpretation is consistent with the observation
that OST1 has only a very modest kinase activity towards
UBA2a as an in vitro substrate (see below). OST1 can also
interact with itself in this two-hybrid analysis, which might
suggest its capacity to trans autophosphorylate in vitro. More-
over, phosphorylation assays showed that at least under the
in vitro conditions tested, the other UBA2 proteins are also
barely phosphorylated by the OST1 kinase. There are 11 ser-
ines and nine threonines theoretically phosphorylable in the
UBA2a, but none is associated with the core consensus motif
(R-XX-S/T) found in the putative substrates of rice and Ara-
bidopsis SnRK2 kinases [22,23]. One possibility is that OST1
docks onto UBA2a via intermediates that were not reproduced
in our experimental systems (yeast two-hybrid, and in vitro
using OST1 produced in protoplasts or bacteria). An alterna-
tive explanation is that despite the functional similarities in
the individual components, the physiological relationship be-
tween the kinases and their respective RNA-binding protein(s)
in ABA signalling has not been conserved between Vicia faba
and Arabidopsis thaliana.
Acknowledgements: We thank Z. Lorkovic and W. Filipowicz for pro-viding plasmids, C. Belin for the purified OST1 kinase and S. Thominefor thoughtful suggestions. M.R. was supported by a Marie Curie FP5Individual Post-doctoral Fellowship (MCFI-2002-01624) and Y.R. bya CNRS post-doctoral fellowship. The project is supported in part bythe Genoplante program Phase III GEN026 (J.L.).
Appendix A. Supplementary data
Supplementary data associated with this article can be found,
in the online version, at doi:10.1016/j.febslet.2006.06.064.
References
[1] Finkelstein, R.R., Gampala, S.S. and Rock, C.D. (2002) Abscisicacid signaling in seeds and seedlings. Plant Cell 14 (Suppl), S15–S45.
[2] Riera, M., Valon, C., Fenzi, F., Giraudat, J. and Leung, J. (2005)The genetics of adaptive responses to drought stress: abscisic acid-dependent and abscisic acid-independent signalling components.Physiol. Plant. 123, 111–119.
[3] Kuhn, J.M. and Schroeder, J.I. (2003) Impacts of altered RNAmetabolism on abscisic acid signaling. Curr. Opin. Plant. Biol. 6,463–469.
[4] Hugouvieux, V., Kwak, J.M. and Schroeder, J.I. (2001) AnmRNA cap binding protein, ABH1, modulates early abscisic acidsignal transduction in Arabidopsis. Cell 106, 477–487.
[5] Xiong, L., Gong, Z., Rock, C.D., Subramanian, S., Guo, Y., Xu,W., Galbraith, D. and Zhu, J.K. (2001) Modulation of abscisicacid signal transduction and biosynthesis by an Sm-like protein inArabidopsis. Dev. Cell 1, 771–781.
[6] Lu, C. and Fedoroff, N. (2000) A mutation in the ArabidopsisHYL1 gene encoding a dsRNA binding protein affects responsesto abscisic acid, auxin, and cytokinin. Plant Cell 12, 2351–2366.
[7] Razem, F.A., El-Kereamy, A., Abrams, S.R. and Hill, R.D.(2006) The RNA-binding protein FCA is an abscisic acidreceptor. Nature 439, 290–294.
[8] Li, J., Kinoshita, T., Pandey, S., Ng, C.K., Gygi, S.P., Shimazaki,K. and Assmann, S.M. (2002) Modulation of an RNA-bindingprotein by abscisic-acid-activated protein kinase. Nature 418,793–797.
[9] Ng, C.K., Kinoshita, T., Pandey, S., Shimazaki, K. and Assmann,S.M. (2004) Abscisic acid induces rapid subnuclear reorganizationin guard cells. Plant Physiol. 134, 1327–1331.
[10] Li, J. and Assmann, S.M. (1996) An abscisic acid-activated andcalcium-independent protein kinase from guard cells of fava bean.Plant Cell 8, 2359–2368.
[11] Mustilli, A.C., Merlot, S., Vavasseur, A., Fenzi, F. and Giraudat,J. (2002) Arabidopsis OST1 protein kinase mediates the regula-tion of stomatal aperture by abscisic acid and acts upstream ofreactive oxygen species production. Plant Cell 14, 3089–3099.
[12] Yoshida, R., Hobo, T., Ichimura, K., Mizoguchi, T., Takahashi,F., Aronso, J., Ecker, J.R. and Shinozaki, K. (2002) ABA-activated SnRK2 protein kinase is required for dehydration stresssignaling in Arabidopsis. Plant Cell Physiol. 43, 1473–1483.
[13] Boudsocq, M., Barbier-Brygoo, H. and Lauriere, C. (2004)Identification of nine SNF1-related protein kinases 2 activatedby hyperosmotic and saline stresses in Arabidopsis thaliana. J.Biol. Chem. 279, 41758–41766.
[14] Lambermon, M.H., Fu, Y., Wieczorek Kirk, D.A., Dupasquier,M., Filipowicz, W. and Lorkovic, Z.J. (2002) UBA1 andUBA2, two proteins that interact with UBP1, a multifunctionaleffector of pre-mRNA maturation in plants. Mol. Cell. Biol. 22,4346–4357.
[15] Mathur, J. and Koncz, C. (1998) PEG-mediated protoplasttransformation with naked DNA methods. Mol. Biol. 82, 267–276.
[16] Hellens, R.P., Edwards, E.A., Leyland, N.R., Bean, S. andMullineaux, P.M. (2000) pGREEN: a versatile and flexible binaryTi vector for Agrobacterium-mediated plant transformation.Plant Mol. Biol. 42, 819–832.
[17] Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified methodfor Agrobacterium-mediated transformation of Arabidopsis tha-liana. Plant J. 16, 735–743.
[18] Bensmihen, S., To, A., Lambert, G., Kroj, T., Giraudat, J. andParcy, F. (2004) Analysis of an activated ABI5 allele using a newselection method for transgenic Arabidopsis seeds. FEBS Lett.561, 127–131.

M. Riera et al. / FEBS Letters 580 (2006) 4160–4165 4165
[19] Alonso, J.M., Stepanova, A.N., Leisse, T.J., Kim, C.J., Chen, H.,Shinn, P., Stevenson, D.K., Zimmerman, J., Barajas, P., Cheuk,R., Gadrinab, C., Heller, C., Jeske, A., Koesema, E., Meyers,C.C., Parker, H., Prednis, L., Ansari, Y., Choy, N., Deen, H.,Geralt, M., Hazari, N., Hom, E., Karnes, M., Mulholland, C.,Ndubaku, R., Schmidt, I., Guzman, P., Aguilar-Henonin, L.,Schmid, M., Weigel, D., Carter, D.E., Marchand, T., Risseeuw,E., Brogden, D., Zeko, A., Crosby, W.L., Berry, C.C. and Ecker,J.R. (2003) Genome-wide insertional mutagenesis of Arabidopsisthaliana Science 301, 653–657.
[20] Lorkovic, Z.J. and Barta, A. (2002) Genome analysis: RNArecognition motif (RRM) and K homology (KH) domain RNA-binding proteins from the flowering plant Arabidopsis thaliana.Nucleic Acids Res. 30, 623–635.
[21] Lorkovic, Z.J. and Barta, A. (2004) Compartmentalization of thesplicing machinery in plant cell nuclei. Trends Plant Sci. 9, 565–568.
[22] Kobayashi, Y., Murata, M., Minami, H., Yamamoto, S., Kagaya,Y., Hobo, T., Yamamoto, A. and Hattori, T. (2005) Abscisic acid-activated SNRK2 protein kinases function in the gene-regulationpathway of ABA signal transduction by phosphorylating ABAresponse element-binding factors. Plant J. 44, 939–949.
[23] Furihata, T., Maruyama, K., Fujita, Y., Umezawa, T., Yoshida,R., Shinozaki, K. and Yamaguchi-Shinozaki, K. (2006) Abscisicacid-dependent multisite phosphorylation regulates the activity ofa transcription activator AREB1. Proc. Natl. Acad. Sci. USA 103,1988–1993.




















