
Hepatic CYP1A, 2B, 2C, 2E and 3A regulation by methoxychlor in male and female rats
11
Hepatic CYP1A, 2B, 2C, 2E and 3A regulation by methoxychlor in male and female rats Luis F. Oropeza-Herna ´ndez, Ricardo Lo ´ pez-Romero, Arnulfo Albores * Seccio ´n de Toxicologı ´a, Cinvestav-IPN, Avenida IPN 2508, Mexico City 07360, Mexico Abstract The effect on liver cytochrome P450 (CYP) by i.p. injections of methoxychlor (MXC) in corn oil at 0, 100, 150, 200 or 250 mg/kg twice daily for 3 days was investigated in adult male and female Wistar rats. The MXC injection (100 mg/kg b.w.) caused a similar increase of total CYP content in males and females as compared with controls who received the vehicle only. In males, this increase continued up to 250 mg/kg. As to the induction of specific CYP activities, the effect of MXC was found to be sex dependent with three different patterns. Males showed the greatest increases of ethoxy- and methoxyresorufin-O -dealkylase (EROD and MROD, respectively), two CYP1A1/1A2-related activities. On the contrary, females were more responsive than males for pentoxyresorufin-O -dealkylase (PROD) and benzyloxyresor- ufin-O -dearylase (BROD), two CYP2B-related activities. Finally, p -nitrophenol hydroxylase (PNPH), a CYP2E1- related activity, showed a similar small, although statistically significant, increase for both sexes. As to CYP apoprotein levels, CYP1A1 and CYP2B1/2B2 showed greater increases in females than in males; whereas, interestingly, CYP2E1 induction was higher in males than in females. These results indicate overall that gender modulates CYP expression after MXC injection both qualitatively and quantitatively, and, therefore, this pesticide is not a pure PB inducer. Moreover, the statistically significant increase of CYP3A2 apoprotein expression observed in females and also, to a lower extent, in males, and the decrease of CYP2C11 apoprotein found in males, two sex-related enzymes, may explain the reported endocrine disrupting effect of MXC. The relevance of the different patterns of rat liver CYP induction observed after MXC treatment, in relationship to the speculated endocrine disrupting potential of MXC in humans potentially exposed to this pesticide, needs further investigation. # 2003 Elsevier Ireland Ltd. All rights reserved. Keywords: Liver CYP; Methoxychlor; Organochlorine pesticides; Sex differences; CYP regulation 1. Introduction Methoxychlor (MXC; 1,1,1-trichloro-2,2-bis-(4- methoxyphenyl)-ethane), a broad spectrum orga- nochlorine (OC) pesticide, gained popularity be- cause of its short half-life in mammals (Kapoor et al., 1970). However, this compound can be depos- ited on the ground, bind to soil particles, thus decreasing its mobility (Derr, 1974). MXC is currently used as a substitute for DDT (1,1,1- trichloro-2,2-bis-(chlorophenyl)ethane) which has been banned or strictly regulated since 2000 in industrially developed and other countries due to * Corresponding author. Tel.: /52-55-5747-3308; fax: /52- 55-5747-7002/111. E-mail address: [email protected]vestav.mx (A. Albores). Toxicology Letters 144 (2003) 93 /103 www.elsevier.com/locate/toxlet 0378-4274/03/$ - see front matter # 2003 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/S0378-4274(03)00230-3
Transcript of Hepatic CYP1A, 2B, 2C, 2E and 3A regulation by methoxychlor in male and female rats

Hepatic CYP1A, 2B, 2C, 2E and 3A regulation bymethoxychlor in male and female rats
Luis F. Oropeza-Hernandez, Ricardo Lopez-Romero, Arnulfo Albores *
Seccion de Toxicologıa, Cinvestav-IPN, Avenida IPN 2508, Mexico City 07360, Mexico
Abstract
The effect on liver cytochrome P450 (CYP) by i.p. injections of methoxychlor (MXC) in corn oil at 0, 100, 150, 200 or
250 mg/kg twice daily for 3 days was investigated in adult male and female Wistar rats. The MXC injection (100 mg/kg
b.w.) caused a similar increase of total CYP content in males and females as compared with controls who received the
vehicle only. In males, this increase continued up to 250 mg/kg. As to the induction of specific CYP activities, the effect
of MXC was found to be sex dependent with three different patterns. Males showed the greatest increases of ethoxy-
and methoxyresorufin-O -dealkylase (EROD and MROD, respectively), two CYP1A1/1A2-related activities. On the
contrary, females were more responsive than males for pentoxyresorufin-O -dealkylase (PROD) and benzyloxyresor-
ufin-O -dearylase (BROD), two CYP2B-related activities. Finally, p -nitrophenol hydroxylase (PNPH), a CYP2E1-
related activity, showed a similar small, although statistically significant, increase for both sexes. As to CYP apoprotein
levels, CYP1A1 and CYP2B1/2B2 showed greater increases in females than in males; whereas, interestingly, CYP2E1
induction was higher in males than in females. These results indicate overall that gender modulates CYP expression
after MXC injection both qualitatively and quantitatively, and, therefore, this pesticide is not a pure PB inducer.
Moreover, the statistically significant increase of CYP3A2 apoprotein expression observed in females and also, to a
lower extent, in males, and the decrease of CYP2C11 apoprotein found in males, two sex-related enzymes, may explain
the reported endocrine disrupting effect of MXC. The relevance of the different patterns of rat liver CYP induction
observed after MXC treatment, in relationship to the speculated endocrine disrupting potential of MXC in humans
potentially exposed to this pesticide, needs further investigation.
# 2003 Elsevier Ireland Ltd. All rights reserved.
Keywords: Liver CYP; Methoxychlor; Organochlorine pesticides; Sex differences; CYP regulation
1. Introduction
Methoxychlor (MXC; 1,1,1-trichloro-2,2-bis-(4-
methoxyphenyl)-ethane), a broad spectrum orga-
nochlorine (OC) pesticide, gained popularity be-
cause of its short half-life in mammals (Kapoor et
al., 1970). However, this compound can be depos-
ited on the ground, bind to soil particles, thus
decreasing its mobility (Derr, 1974). MXC is
currently used as a substitute for DDT (1,1,1-
trichloro-2,2-bis-(chlorophenyl)ethane) which has
been banned or strictly regulated since 2000 in
industrially developed and other countries due to
* Corresponding author. Tel.: �/52-55-5747-3308; fax: �/52-
55-5747-7002/111.
E-mail address: [email protected] (A. Albores).
Toxicology Letters 144 (2003) 93�/103
www.elsevier.com/locate/toxlet
0378-4274/03/$ - see front matter # 2003 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/S0378-4274(03)00230-3

its toxicity and high persistence in the environment
(Matteson and Ramirez, 1999). In Mexico, MXC
is presently used for a large number of crops
(Matteson and Ramirez, 1999). In spite of the
many advantages of MXC use, the widespread
application of this pesticide over several decades is
expected to result in substantial environmental
contamination and, therefore, significant animal
and human exposure to both MXC and its
metabolites (Li and Kupfer, 1998).The presence of the methoxy groups in the
MXC structure probably accounts for the faster
degradation of the compound and its low toxicity
in rats (LD50: 6000�/7000 mg/kg) which is lower
than that of DDT (LD50: 100�/250 mg/kg) (Mur-
phy, 1980). However, MXC presents also some
undesirable side effects. For example, technical
MXC exhibits a higher oestrogenic activity than
that observed for the individual isomers (Bitman
and Cecil, 1970; Nelson, 1974; Tullner, 1961), as
reported both in vivo and in vitro (Bitman and
Cecil, 1970; Bulger, et al., 1978; Nelson, 1974;
Tullner, 1961). In fact, technical MXC induces
oestrogen-like changes in the reproductive tract of
female rats, thus inhibiting fertility (Gray, et al.,
1988; Cummings and Gray, 1989; Cummings and
Laskey, 1993).
OC pesticides, like DDT and its analogues, are
well-known inducers of various hepatic cyto-
chrome P450s (CYP), such as CYP2B1, in rodents
(Hart and Fouts, 1965; Kupfer and Peets, 1966;
Bunyan et al., 1972). This classifies DDT as a
phenobarbital (PB)-type inducer (Lubet et al.,
1992). In comparison with DDT, MXC has
received little attention, probably since early
attempts to provoke a monooxygenase response
to MXC exposure in rats met with little or no
success. For example, a single low DDT dose (5
mg/kg) increased 1.4-fold the total hepatic CYP
content in rats (Sierra-Santoyo et al., 2000),
whereas the administration of 200 mg/kg, twice
daily for 3 days, resulted in just a 1.1-fold increase
(Li et al., 1995). The lack of a significant effect in
rats could be due to the rapid biotransformation of
MXC in this species (Li et al., 1995). However,
multiple dosage treatments with MXC succeeded
in inducing rat hepatic CYP (Li et al., 1995).
The reported endocrine disrupting toxicity ofDDT in rats and other species (Bulger and Kupfer,
1985; Guillette et al., 1994) could be attributed to
its effects on CYPs involved in the metabolism of
sex hormones (Sierra-Santoyo et al., 2000). Prob-
ably, a similar potential ability could also be
attributed to MXC. However, there is no informa-
tion comparing the influence of sex on the CYP
regulation by MXC. Therefore, the aim of thisstudy was to investigate the effect of MXC
administration on the expression of different sex-
related and other CYPs in rat liver in order to
clarify the role of gender, if any, on the response to
this pesticide.
2. Materials and methods
2.1. Reagents
The following reagents were purchased: 7-
ethoxy-, 7-methoxy-, 7-pentoxy, 7-benzyloxy-re-
sorufin, and resorufin from Molecular Probes,
Inc., (Eugene, OR); MXC, NADPH and Tris�/
HCl from Sigma Chemical Co. (St Louis, MO);nitrocellulose paper and other chemicals used for
Western blotting from BIO-RAD (Richmond,
CA). Goat anti-mouse and anti-rabbit IgG con-
jugated to horseradish peroxidase were obtained
from PIERCE (Rockford, IL) and anti-CYP3A2
from Gentest Corp. (Woburn, MA). Anti-
CYP2C11 (Dr. S. Bandiera, University of British
Columbia, Vancouver, BC, Canada), anti-CYP1A1, anti-2B1/2B2 and anti-2E1 (Dr. Colin
Jefcoate, University of Wisconsin-Madison, Ma-
dison, WI) were kindly donated to our group.
2.2. Animal treatment
Male and female Wistar rats (2509/50 g) housed
in polycarbonate cages with sawdust bedding were
fed a standard diet (PMI Feed, Inc., St Louis, MO)and water ad libitum, and were maintained at
21 8C and at a 50% relative humidity in a 12 h
dark/light cycle (starting at 07:00 h). Groups of
five rats were injected with MXC (100, 150, 200, or
250 mg of MXC/kg) as a suspension in 0.4 ml of
corn oil, i.p. twice daily for 3 days. A group of rats
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/10394

was similarly injected with corn oil only. Food waswithdrawn immediately after the last MXC or
corn oil injection and rats were killed 24 h later by
deep anaesthesia with pentobarbital. Livers were
perfused with ice-cold saline (NaCl 0.9%, w/v),
excised and processed to obtain microsomes ac-
cording to Mayer et al. (1990). This study was
conducted in compliance with the Mexican Reg-
ulations of Good Laboratory Practice (NOM,2001).
2.3. Other treatments
Positive CYP1A1, CYP2B1/2B2 or CYP2E1
control microsomes were obtained from male
rats treated i.p. with b-naphthoflavone (BNF, 80
mg/kg�/2 days), PB (80 mg/kg�/3 days) orpyridine (Pyr, 100 mg/kg�/3 days), respectively;
and killed under deep anaesthesia with pentobar-
bital.
2.4. CYP content and alkoxyresorufin-O-
dealkylation assays
Total hepatic microsomal CYP content wasmeasured according to Omura and Sato (1964).
Microsomal O -dealkylation of 7-ethoxy- (EROD),
7-methoxy- (MROD), 7-pentoxy- (PROD), and 7-
benzyloxy-resorufin (BROD) were assayed fluor-
imetrically at 37 8C using 530 and 585 nm as the
excitation and emission wavelengths, respectively
(Burke et al., 1985; Lubet et al., 1985; Nerurkar et
al., 1993). The hydroxylation of p -nitrophenol(PNPH) was assayed by the formation of p -
nitrocatechol detected at 510 nm in a Lambda 2S
spectrophotometer (Perkin�/Elmer, Co., Uberlin-
gen, Germany). The PNPH activity was expressed
as nmol of p-nitrocatechol formed per min per mg
of microsomal protein (Reinke and Moyer, 1985).
Proteins were measured according to Lowry et al.
(1951) using bovine serum albumin (BSA) as thestandard.
2.5. Western immunoblots
Microsomal proteins were electrophoretically
separated in SDS-10% polyacrylamide gels (SDS-
PAGE), according to the method of Laemmli
(1970). The proteins were then electrotransferredto a nitrocellulose membrane and the plate
blocked with 100 mM glycine, 1% BSA and 5%
non-fat milk powder overnight at 4 8C. The
membrane was incubated with anti-rat CYP1A1,
2B1/2B2, 2C11, 2E1, or 3A2 antibodies for 1 h,
followed by another 1 h incubation with a horse-
radish peroxidase-conjugated secondary antibody
(goat anti-rabbit IgG). The protein bands werevisualized by chemiluminescence (Amersham, Ar-
lington Heights, IL) followed by a brief exposure
to Kodak XAR Scientific Imaging film. A Foto/
Eclipse videocamera (Fotodyne, Inc.) and Collage
ver. 3.0 software were used to scan the blotted
negatives. Band intensities were compared with the
groups of different treatment.
2.6. Statistical analysis
The analysis of variance (ANOVA) and the
Dunnett’s t-test were used to assess differences
between the experimental groups. A positive
association between any enzyme activities and
protein content was established by linear regres-sion analysis. The level of statistically significant
difference between the mean values was set at P 5/
0.05. All calculations were performed using a
Sigma Stat, ver. 1.0 software (Jandel Corporation,
San Rafael, CA).
3. Results
3.1. MXC acute toxicity
Any of the MXC doses assayed in this investiga-
tion caused significant effects on body weight,
food or water consumption. The highest dose used
in this experiment represents 4.1�/3.6% of theMXC LD50 values mentioned above. However,
MXC administration significantly increased the
relative liver weight of both male and female rats.
Males were affected at all doses tested, while
females were effected at the highest doses only
(data not shown).
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/103 95

3.2. MXC effects on total hepatic CYP content
MXC administration resulted in a dose-depen-
dent increase of the total hepatic CYP content.
Similar increases were observed up to the dose of
100 mg/kg for both sexes, whereas CYP content
increased differently at the following doses in
males and females (Table 1). In males, the effect
of MXC started at 100 mg/kg, was dose-dependentand reached a maximum (2.1-fold) at the highest
dose (250 mg/kg), whereas, in females, total
hepatic CYP content increased 1.6-fold at 100
mg/kg and remained constant at the highest doses.
The comparison of the slopes of the MXC effect
on total hepatic CYP content showed a greater
statistically significant response in males (b�/
0.002) than in females (b�/4.13�/10�5) indicatingthat the response depends on gender.
3.3. Effect of MXC on CYP-related activities
MXC administration resulted in a dose-depen-
dent increase of all the CYP-dependent activities
studied in this investigation, regardless of sex. At
the highest MXC dose (250 mg/kg), for example,
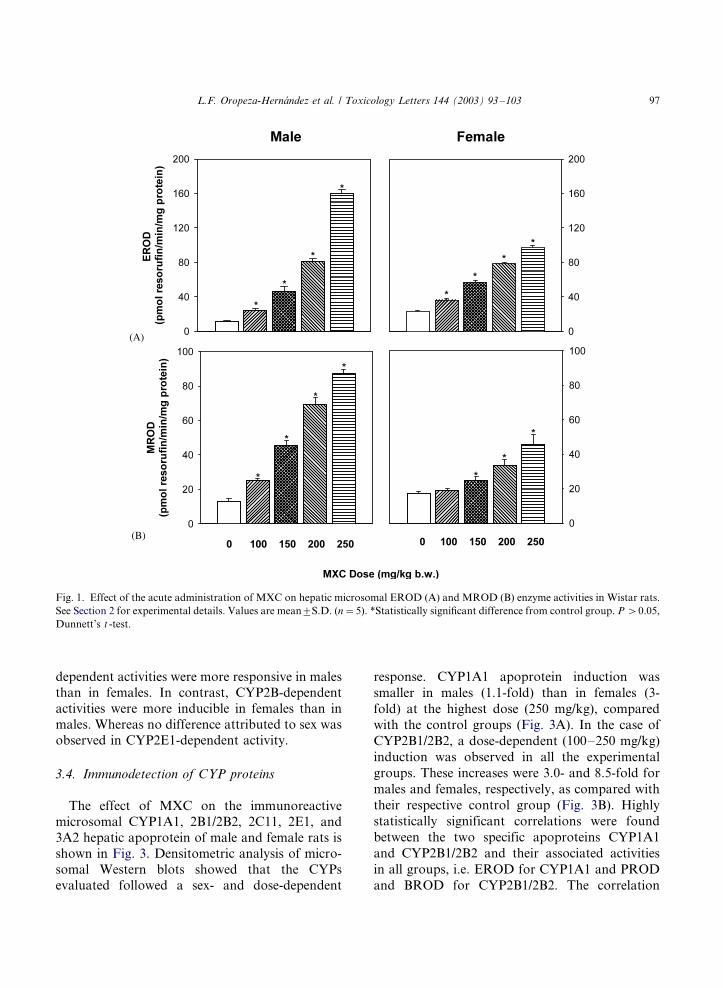
EROD, a relatively specific CYP1A1 activity,increased 13.6-fold in males; whereas, a rather
small response was observed in females (4.2-fold)
(Fig. 1A). This observation was in agreement with
the higher calculated slope of the EROD activity
measured after MXC administration in males(b�/35.4), a value 1.8-fold greater than that
observed in females (b�/19.0), the difference
between slopes being statistically significant. On
the other hand, a smaller increase (males, 5.3-fold;
females, 2.3-fold) of MROD activity, a relatively
specific CYP1A2 activity, was observed after
MXC administration. Male rats showed a slope
of b�/19.4 that was 2.6-fold greater than thatobserved in females (b�/7.2) (Fig. 1B).
MXC was an effective inducer of the CYP2B
subfamily dependent enzyme activities PROD and
BROD, since these activities showed even greater
responses than CYP1A-dependent activities. The
dose�/response effect after exposure to MXC
started at 100 mg/kg and reached the maximum
effect at 250 mg/kg both in males and females. Inmales, PROD and BROD activities increased 14.8-
and 3.2-fold, respectively. Notably, the response
was more marked in females than in males, since at
the dose of 250 mg/kg, PROD and BROD
activities were increased 16.4- and 16-fold, respec-
tively, as compared with their respective control
group (Fig. 2A and B). Indeed, slope calculations
for PROD and BROD activities for the doses of100�/250 mg/kg showed different values in male vs.
female animals. The slope for PROD response in
females was steeper (b�/27.0) than that of males
(b�/21.6). A similar pattern was observed for
BROD in females (b�/76.4) and males (b�/46.7).
With both PROD and BROD, sex differences in
the slopes were statistically significant and showed
that females were more responsive than males.As for PNPH, a CYP2E1 dependent activity, a
small but statistically significant increase was
observed. This effect was similar for both sexes
(Fig. 2C). In male rats significant increases were
observed starting from 150 mg/kg, whereas in
females from 200 mg/kg; the maximum effect
(2.0- and 1.8-fold, for males and females, respec-
tively) occurred at the highest dose tested. Theeffect of MXC on the slopes of PNPH activity was
similar in males and females (b�/0.42 and 0.39,
respectively). Thus, sex does not seem to influence
the effect of MXC on CYP2E1 activity.
The MXC induction pattern was different for
males and females in terms of induction potency
and CYP isoform response. In general, CYP1A1-
Table 1
Total hepatic CYP in male and female Wistar rats after MXC
treatment
Dose (mg/kg) Total CYP content (nmol/mg of microsomal
protein)
Male Female
0 0.4509/0.01 0.4109/0.06
100 0.5599/0.02a 0.6529/0.01a
150 0.9269/0.02a 0.5919/0.01a
200 0.9399/0.01a 0.4649/0.02
250 0.9459/0.01a 0.4669/0.02
Values are given as mean9/S.D.; n�/5.a P 5/0.05, male or female vs. the respective control at dose
0. Dunnett?s t -test. See Section 2 for further experimental
details.
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/10396
dependent activities were more responsive in males
than in females. In contrast, CYP2B-dependent
activities were more inducible in females than in
males. Whereas no difference attributed to sex wasobserved in CYP2E1-dependent activity.
3.4. Immunodetection of CYP proteins
The effect of MXC on the immunoreactive
microsomal CYP1A1, 2B1/2B2, 2C11, 2E1, and
3A2 hepatic apoprotein of male and female rats is
shown in Fig. 3. Densitometric analysis of micro-
somal Western blots showed that the CYPs
evaluated followed a sex- and dose-dependent
response. CYP1A1 apoprotein induction was
smaller in males (1.1-fold) than in females (3-
fold) at the highest dose (250 mg/kg), compared
with the control groups (Fig. 3A). In the case of
CYP2B1/2B2, a dose-dependent (100�/250 mg/kg)
induction was observed in all the experimental
groups. These increases were 3.0- and 8.5-fold for
males and females, respectively, as compared with
their respective control group (Fig. 3B). Highly
statistically significant correlations were found
between the two specific apoproteins CYP1A1
and CYP2B1/2B2 and their associated activities
in all groups, i.e. EROD for CYP1A1 and PROD
and BROD for CYP2B1/2B2. The correlation
Fig. 1. Effect of the acute administration of MXC on hepatic microsomal EROD (A) and MROD (B) enzyme activities in Wistar rats.
See Section 2 for experimental details. Values are mean9/S.D. (n�/5). *Statistically significant difference from control group. P �/0.05,
Dunnett’s t -test.
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/103 97
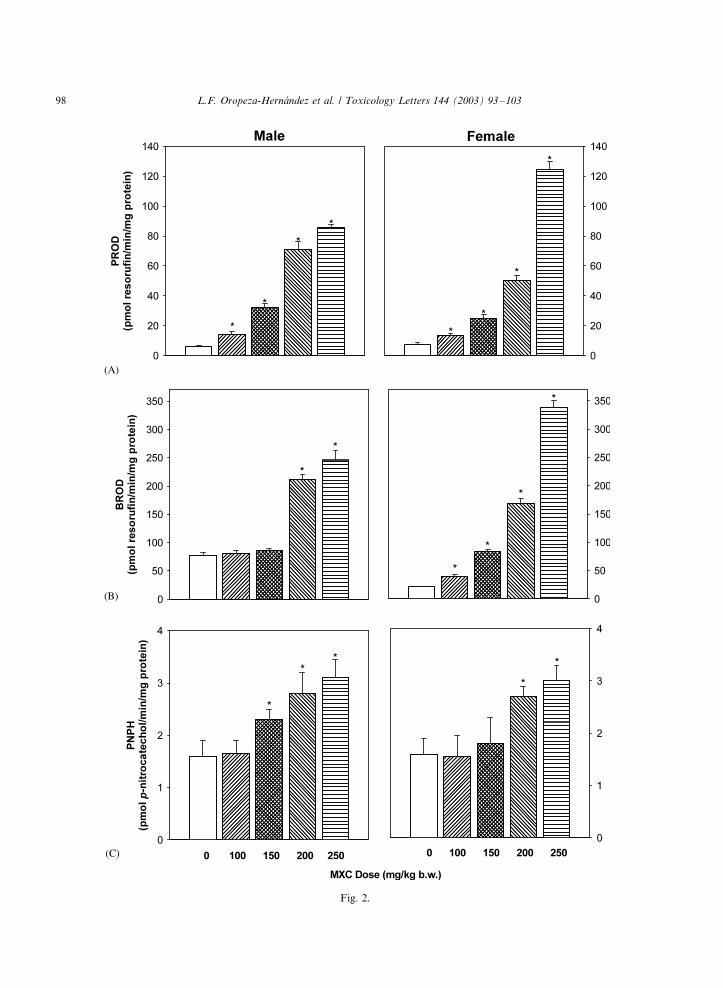
Fig. 2.
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/10398

coefficients between CYP1A1 apoprotein level and
EROD activity in males and females were r�/
0.9485 (P B/0.01) and r�/0.9576 (P B/0.01), re-
spectively.
In males, CYP2B1/2B2 activities and the apo-
protein level showed a slightly better correlation
for PROD (r�/0.9454; P B/0.01) than for BROD
(r�/0.8967; P B/0.01), whereas in females, the
correlations were similar, i.e. r�/0.9933 (P B/
0.01) and r�/0.9817 (P B/0.01), for BROD and
PROD, respectively.
The MXC-dependent increase of the CYP2E1
immunoreactive apoprotein at the highest dose
(250 mg/kg) as compared with the untreated
control was 3.0-fold in males and 1.5-fold in
females (Fig. 3D). These differences in apoprotein
content were not reflected by a corresponding
increase of PNPH activity.
Notably, exposure to MXC decreased the ex-
pression of the male-specific CYP2C11 in a dose-
dependent manner (Fig. 3C), being 0.62-fold that
of the untreated control at the highest MXC dose
(250 mg/kg). Instead, in females, CYP2C11 was
not detected in any group (data not shown).
In addition, MXC also increased the apoprotein
level of the male-specific CYP3A2 in females, the
highest dose used (250 mg/kg) resulting in a 18-fold induction. In contrast, a smaller effect (three-
fold) was observed in males (Fig. 3E). Table 2
summarizes the relative level of induction of the
hepatic CYP apoproteins investigated after MXC
exposure.
4. Discussion
MXC is known to affect the hepatic CYP-
dependent drug metabolizing enzymes (Li et al.,
1995). However, the mechanism of this effect, and
particularly the role of gender, have not been fully
clarified. The results of this study show that MXC
induces several CYP-dependent monooxygenasesin a sex-specific manner. As a result, the endocrine
disruptive effect of MXC may be related with the
observed different regulation of CYPs expression
in the two sexes. Moreover, the data presented
here show that the magnitude of the response in
each gender depends on sex-specific steroidal
Fig. 3. Western blotting analysis of hepatic CYP isoforms obtained from MXC treated rats. Microsomes were subjected to SDS-
PAGE electrophoresis and probed for different CYPs using anti-rat monoclonal antibodies. Ten micrograms of microsomal protein
were loaded on each lane. Lane 1: microsomes from male rats treated with typical inducers, i.e. BNF (CYP1A1) and PB (CYP2B1/2B2
and Pyr (2E1)), or non-treated (CYP2C11, 2E1 and 3A2). Lane 2: microsomes from non-treated animals. Lanes 3�/6: microsomes from
animals treated with 100, 150, 200 and 250 mg of MXC/kg, respectively.
Fig. 2. Effect of the acute administration of MXC on hepatic microsomal PROD (A), BROD (B) and PNPH (C) enzyme activities in
Wistar rats. See Section 2 for experimental details. Values are mean9/S.D. (n�/5). *Statistically significant difference from control
group. P �/0.05, Dunnett’s t -test.
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/103 99
hormones which may be co-involved in the mod-
ulation exerted on the drug metabolism enzymes
by MXC or/and its metabolites. It is possible that
MXC acts as a steroidogenesis regulator or as a
xenoestrogen itself through the activation of the
constitutive androstane receptor system (Blizard et
al., 2001). The modulation of CYPs expression by
MXC may alter the individual capacity to meta-
bolize endogenous and exogenous compounds
such as prostaglandins, drugs, carcinogens and
the steroid hormones themselves (Soucek and Gut,
1992).Interestingly, MXC significantly induced
CYP3A2 expression in a dose-dependent manner
in females (18-fold) as compared with males (3-
fold) and markedly inhibited CYP2C11 in males
(�/38%), as compared with controls. The observa-
tion of the MXC inductive effect on CYP3A2 of
adult female rats is remarkable because CYP3A2
is a male-specific isoform (Waxman et al., 1985).
CYP3A is a gene subfamily composed of multiple
isoforms that have already been characterized by
immunochemistry, catalytic activities, and cDNA
cloning and expression. Hepatic CYP3As, are
responsible for the enzyme oxidation of numerous
clinically used drugs, and this subfamily is induced
by a variety of compounds, including naturally
occurring and synthetic glucocorticoids (e.g., dex-
amethasone), pregnane compounds (e.g., pregne-
nolone 16-carbonitrile), and macrolide antibiotics
(e.g., rifampicin) (Quattrochi and Guzelian, 2001),
among others. Although CYPs 3A23 and 3A2
share a high degree of structural homology, they
are regulated independently (Telhada et al., 1992).
For instance, dexamethasone is a more potent
inducer of CYP3A23 than PB. In contrast,
CYP3A2 is induced only by PB (Ghosal et al.,
1996; Gonzalez et al., 1986). The increase of
CYP3A2 observed here confirms the findings of
Li et al. (1995) who reported increases in the
hydroxylation of testosterone at the 6b, 2b and
15b positions, suggesting that MXC enhances
CYP3A apoprotein level and its related activities.
MXC diminished the CYP2C11 apoprotein level
in male rats, an observation in agreement with the
mechanism proposed by Akingbemi et al. (2000) of
the participation of 2,2-bis-(p-hydroxyphenyl)-
1,1,1-trichloroethen, an MXC metabolite, in the
reduction of testosterone biosynthesis in rat Ley-
dig cells by suppressing the mRNA steady-state
and the enzyme cleavage of the cholesterol side-
chain, thus suggesting that MXC may act as an
anti-androgen. Furthermore, a marked decrease of
2a- and 16a-testosterone hydroxylation was ob-
served when liver microsomes from PB male
treated rats were co-incubated with MXC and
testosterone, again indicating a strong inhibition
of MXC on CYP2C11-related enzyme activity (Li
et al., 1993).
Notably, MXC treatment-induced CYP1A1/
1A2, as measured by their associated enzyme
activities EROD and MROD and by their apo-
protein content, being males more responsive than
females. The differences between the relative
increase of the apoprotein content and the asso-
ciated enzyme activities observed in this study for
the CYP1A subfamily in male and female rats are
consistent with previous reports for the analogues
Table 2
Relative induction of hepatic CYP isoforms after an MXC treatment in male and female Wistar rats
Dose (mg/kg) 1A1 2B1/2B2 2C11 2E1 3A2
M F M F M M F M F
0 1.0 1.0 1.0 1.0 1.0 1.0 1.0 1.0 1.0
100 0.4 0.6 0.9 1.3 0.80 1.2 0.5 1.3 2.2
150 0.5 0.9 1.2 2.5 0.72 1.5 0.8 1.8 4.5
200 0.6 1.5 1.8 4.5 0.66 2.0 1.0 2.3 7.0
250 1.1 3.0 3.0 8.5 0.62 3.0 1.5 3.0 18.0
Note: Relative induction�/exposed group/control group. Immunoreactive protein was determined in liver microsome samples
pooled for each group. See Section 2 for details.
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/103100

of MXC, DDT or PCBs (Sierra-Santoyo et al.,2000; Flodstrom et al., 1990). The MXC-depen-
dent induction of the CYP1A subfamily apopro-
teins and related activities contrasts with the
observations by Li and Kupfer (1998) who found
no increase of the mRNA levels of CYP1A1.
However, no attempt was made by these authors
to measure the apoprotein content and the related
enzyme activity. Taking these observations to-gether it is possible to suggest that MXC regula-
tion of CYP1A is posttransductional rather than
posttranscriptional. Other chemicals, like the
MXC analogue DDT, are also able to increase
EROD activity without affecting the correspon-
dent apoprotein content (Sierra-Santoyo et al.,
2000), thus supporting the fact that CYP1A may
be regulated at different levels.In contrast, the levels of CYP2B1/2B2 apopro-
tein and its associated enzyme activities (PROD
and BROD) showed that female rats to be more
responsive to MXC than males. In addition, both
females and males displayed a greater response of
PROD than of BROD activities. These results are
in agreement with those reported by Li et al.
(1995) showing that CYP2B1/2B2 apoproteinswere induced in mature female rats. In addition,
these authors suggested that MXC increased the
16a- and 6b-hydroxylation of testosterone and
androstenedione formation, indicating an MXC-
mediated induction of CYP2B enzyme activities.
Although we could not find any studies on MXC
reproductive toxicity in humans, several studies in
animals, both males and females, suggest thatMXC is able to affect the reproductive system
(Bulger and Kupfer, 1985). These effects are
thought to be due to the oestrogenic activity of
both O -demethylated metabolites of MXC and
some of the O -demethylated contaminants of
technical grade MXC (Bulger and Kupfer, 1985),
CYP2B1 being the isoform responsible for the O -
demethylation and ring-hydroxylation of MXC(Dehal and Kupfer, 1994).
The inductive pattern of MXC on CYP reported
here mimics that observed for DDT (Sierra-
Santoyo et al., 2000). Although MXC showed to
be an effective inducer in both sexes, a larger dose
and a repetitive exposure scheme were needed with
MXC to reach a similar level of induction to that
caused by a small single dose of DDT (Sierra-Santoyo et al., 2000).
In summary, our results showed that MXC is a
relatively weak, but still effective, inducer of CYP
and related activities. The magnitude of the
response to this pesticide depends on gender and
involves sex-specific isoforms. The preferential
induction observed for CYPs involved in steroid
metabolism (2B1, 2B2 and 3A2) implies that MXCmay be able to change the pattern of steroid
turnover. In addition, the MXC-mediated induc-
tion of the CYP1A subfamily suggests that this
pesticide is not a pure PB inducer. Taken together
these results indicate that the induction by MXC
of several CYP isoforms in rat liver is capable of
altering the hormonal homeostasis in this species.
However, the mechanisms responsible for thegeneral and gender-specific modulation by MXC
of the expression of different CYP isoforms in rats
and the relevance of these events for humans
remain to be established.
Acknowledgements
The European Community, EC INCO-DC con-
tract ERB IC18-CT980341, supported the present
research. L.F.O.H. was a recipient of a doctoral
scholarship from CONACyT, Mexico.
References
Akingbemi, B.T., Ge, R.S., Klinefelter, G.R., Gunsalus, G.L.,
Hardy, M.P., 2000. A metabolite of methoxychlor, 2,2-
bis(p -hydroxyphenyl)-1,1, 1-trichloroethane, reduces testos-
terone biosynthesis in rat leydig cells through suppression of
steady-state messenger ribonucleic acid levels of the choles-
terol side-chain cleavage enzyme. Biol. Reprod. 62, 571�/
578.
Bitman, J., Cecil, H.C., 1970. Estrogenic activity of DDT
analogs and polychlorinated biphenyls. J. Agric. Food
Chem. 18, 1108�/1112.
Blizard, D., Sueyoshi, T., Negishi, M., Dehal, S.S., Kupfer, D.,
2001. Mechanism of induction of cytochrome P450 enzymes
by the proestrogenic endocrine disruptor pesticide-methox-
ychlor: interactions of methoxychlor metabolites with the
constitutive androstane receptor system. Drug Metab.
Dispos. 29, 781�/785.
Bulger, W.H., Kupfer, D., 1985. Estrogenic activity of pesti-
cides and other xenobiotics on the uterus and male
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/103 101

reproductive tract. In: Thomas, J.A., Korach, K.S., McLa-
chlan, J.A. (Eds.), Endocrine Toxicology. Raven, New
York, pp. 1�/3.
Bulger, W.H., Muccitelli, R.M., Kupfer, D., 1978. Studies on
the in vivo and in vitro estrogenic activities of methoxychlor
and its metabolites: Role of hepatic monooxigenase in
methoxychlor activation. Biochem. Pharmacol. 27, 2417�/
2423.
Bunyan, P.J., Townsend, M.G., Taylor, A., 1972. Pesticide-
induced changes in hepatic microsomal enzyme systems.
Some effects of 1,1-DI(p -chlorophenyl)-2,2,2-trichlor-
oethane (DDT) and 1,1-DI(p -chlorophenyl)-2,2-dichlor-
oethylene (DDE) in the rat and Japanese quail. Chem.
Biol. Interact. 5, 13�/26.
Burke, M.D., Thompson, S., Elcombe, C.E., Halpert, J.,
Haaparanta, T., Mayer, R.T., 1985. Ethoxy-, pentoxy-
and benzyloxyphenoxazone and homologues: a series of
substrates to distinguish between different induced cyto-
chromes P-450. Biochem. Pharmacol. 34, 3337�/3345.
Cummings, A.M., Gray, L.E., Jr., 1989. Antifertility effect of
methoxychlor in female rats: dose- and time-dependent
blockade of pregnancy. Toxicol. Appl. Pharmacol. 97,
454�/462.
Cummings, A.M., Laskey, J., 1993. Effect of methoxychlor on
ovarian steroidogenesis: role in early pregnancy loss.
Reprod. Toxicol. 7, 17�/23.
Dehal, S.S., Kupfer, D., 1994. Metabolism of the proestrogenic
pesticide methoxychlor by hepatic P450 monooxygenases in
rats and humans. Dual pathways involving novel ortho
ring-hydroxylation by CYP2B. Drug Metab. Dispos. 22,
937�/946.
Derr, S.K., 1974. Bioactive compounds in the aquatic environ-
ment. Loss of methoxychlor from autumn-shed leaves into
the aquatic environment. Bull. Environ. Contam. Toxicol.
11, 500�/502.
Flodstrom, S., Hemming, H., Warngard, L., Ahlborg, U.G.,
1990. Promotion of altered hepatic foci development in rat
liver, cytochrome P450 enzyme induction and inhibition of
cell-cell communication by DDT and some structurally
related organohalogen pesticides. Carcinogenesis 11, 1413�/
1417.
Ghosal, A., Satoh, H., Thomas, P.E., Bush, E., Moore, D.,
1996. Inhibition and kinetics of cytochrome P4503A activity
in microsomes from rat, human, and cDNA-expressed
human cytochrome P450. Drug Metab. Dispos. 24, 940�/
947.
Gonzalez, F.J., Song, B.J., Hardwick, J.P., 1986. Pregnenolone
16 alpha-carbonitrile-inducible P-450 gene family: gene
conversion and differential regulation. Mol. Cell. Biol. 6,
2969�/2976.
Gray, L.E., Jr., Ostby, J., Sigmon, R., Ferrell, J., Rehnberg, G.,
Linder, R., Cooper, R., Goldman, J., Laskey, J., 1988. The
development of a protocol to assess reproductive effects of
toxicants in the rat. Reprod. Toxicol. 2, 281�/287.
Guillette, L.J., Jr., Gross, T.S., Masson, G.R., Matter, J.M.,
Percival, H.F., Woodward, A.R., 1994. Developmental
abnormalities of the gonad and abnormal sex hormone
concentrations in juvenile alligators from contaminated and
control lakes in Florida. Environ. Health Perspect. 102,
680�/688.
Hart, L.G., Fouts, J.R., 1965. Further studies on the stimula-
tion of hepatic microsomal drug metabolizing enzymes by
DDT and its analogs. Arch. Exp. Pathol. Pharmacol. 249,
486�/500.
Kapoor, I.P., Metcalf, R.L., Nystrom, R.F., Sangha, G.K.,
1970. Comparative metabolism of methoxychlor, methio-
chlor, and DDT in mouse, insects, and in a model
ecosystem. J. Agric. Food Chem. 18, 1145�/1152.
Kupfer, D., Peets, L., 1966. The effect of o ,p ?DDD on cortisol
and hexobarbital metabolism. Biochem. Pharmacol. 15,
573�/581.
Laemmli, U.K., 1970. Cleavage of structural proteins during
the assembly of head of bacteriophage T4. Nature 227, 680�/
685.
Li, H.C., Kupfer, D., 1998. Mechanism of induction of rat
hepatic CYP2B and 3A by the pesticide methoxychlor. J.
Biochem. Mol. Toxicol. 12, 315�/323.
Li, H.C., Mani, C., Kupfer, D., 1993. Reversible and time-
dependent inhibition of the hepatic cytochrome P450
steroidal hydroxylases by the proestrogenic pesticide meth-
oxychlor in rat and human. J. Biochem. Toxicol. 8, 195�/
206.
Li, H.C., Dehal, S.S., Kupfer, D., 1995. Induction of the
hepatic CYP2B and CYP3A enzymes by the proestrogenic
pesticide methoxychlor and by DDT in the rat. Effects on
methoxychlor metabolism. J. Biochem. Toxicol. 10, 51�/
61.
Lowry, O.H., Rosenbrough, N.F., Farr, A.L., Randall, R.J.,
1951. Protein measurement with the Folin phenol reagent. J.
Biol. Chem. 193, 265�/275.
Lubet, R.A., Mayer, R.T., Cameron, J.W., Nims, R.W., Burke,
M.D., Wolff, T., Guenguerich, P.F., 1985. Dealkylation of
pentoxyresorufin: a rapid and sensitive assay for measuring
induction of cytochrome(s) P-450 by phenobarbital and
other xenobiotics in the rat. Arch. Biochem. Biophys. 238,
43�/48.
Lubet, R.A., Nims, R.W., Dragnev, K.H., Jones, C.R., Diwan,
B.A., Devor, D.E., Ward, J.M., Miller, M.S., Rice, J.M.,
1992. A markedly diminished pleiotropic response to
phenobarbital and structurally-related xenobiotics in
Zucker rats in comparison with F344/NCr or DA rats.
Biochem. Pharmacol. 43, 1079�/1087.
Matteson, P., Ramirez, J., 1999. Phasing out the use of DDT
for malaria control in Mexico. In: Matteson, P.C. (Ed.),
Disease Vector Management for Public Health and Con-
servation. World Wildlife Fund, Washington, pp. 8.1�/
8.38.
Mayer, R.T., Netter, K.J., Heubel, F., Hahnemann, B.,
Buccheister, A., Mayer, G.K., Burke, M.D., 1990. 7-
Alkoxyquinolines: new fluorescent substrates for cyto-
chrome P450 monooxygenases. Biochem. Pharmacol. 40,
1645�/1655.
Murphy, S.D., 1980. Pesticides: Casarett and Doull’s toxicol-
ogy. In: Doull, J., Klaassen, C.D., Amdur, M.O. (Eds.), The
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/103102

Basic Science of Poisons. Macmillan, New York, pp. 357�/
408.
Nelson, J.A., 1974. Effects of dichlorodiphenyltrichloroethane
DDT analogs and polychlorinated byphenyl (PCB) mixtures
on 17b-(3H)estradiol binding to rat uterine receptor. Bio-
chem. Pharmacol. 23, 447�/451.
Nerurkar, P.V., Park, S.S., Thomas, P.E., Bims, R.W., Lubet,
R.A., 1993. Methoxyresorufin and benzyloxy: substrates
preferentially metabolised by cytochromes P4501A2 and 2B,
respectively, in the rat and mouse. Biochem. Pharmacol. 46,
933�/943.
NOM, 2001. Especificaciones tecnicas para la produccion,
cuidado y uso de los animales de laboratorio. Norma Oficial
Mexicana NOM-062-ZOO-1999. Diario Oficial de la Fed-
eracion 2a, Seccion 575, 20�/72.
Omura, T., Sato, R., 1964. The carbon monoxide-binding
pigment of liver microsomes. J. Biol. Chem. 239, 2370�/
2378.
Quattrochi, L.C., Guzelian, P.S., 2001. Cyp3A regulation: from
pharmacology to nuclear receptors. Drug Metab. Dispos.
29, 615�/622.
Sierra-Santoyo, A., Hernandez, M., Albores, A., Cebrian,
M.E., 2000. Sex-dependent regulation of hepatic cyto-
chrome P-450 by DDT. Toxicol. Sci. 54, 81�/87.
Soucek, P., Gut, I., 1992. Cytochromes P-450 in rats: structures,
functions, properties and relevant human forms. Xenobio-
tica 22, 83�/103.
Reinke, L.A., Moyer, M.J., 1985. p -Nitrophenol hydroxyla-
tion. A microsomal oxidation which is highly inducible by
ethanol. Drug Metab. Dispos. 13, 548�/552.
Telhada, M.B., Pereira, T.M., Lechner, M.C., 1992. Effect of
dexamethasone and phenobarbital on run-on transcription
rate and CYP3A mRNA concentration in rat liver: changes
during development. Arch. Biochem. Biophys. 298, 715�/
725.
Tullner, W.W., 1961. Uterotropic action of the insecticide
methoxychlor. Science 133, 647�/648.
Waxman, D.J., Dannan, G.A., Guengerich, F.P., 1985. Reg-
ulation of rat hepatic cytochrome P-450: age-dependent
expression, hormonal imprinting, and xenobiotic inducibil-
ity of sex-specific isoenzymes. Biochemistry 24, 4409�/4417.
L.F. Oropeza-Hernandez et al. / Toxicology Letters 144 (2003) 93�/103 103




















