
Gibberellin/Abscisic Acid Antagonism In Barley Aleurone Cells: Site of Action of the Protein Kinase...
14
The Plant Cell, Vol. 13, 667–679, March 2001, www.plantcell.org © 2001 American Society of Plant Physiologists Gibberellin/Abscisic Acid Antagonism in Barley Aleurone Cells: Site of Action of the Protein Kinase PKABA1 in Relation to Gibberellin Signaling Molecules Aurelio Gómez-Cadenas, a,1 Rodolfo Zentella, a Mary Kay Walker-Simmons, b and Tuan-Hua David Ho a,2 a Plant Biology Program, Department of Biology, Washington University, St. Louis, Missouri 63130 b United States Department of Agriculture–Agricultural Research Service, Washington State University, Pullman, Washington 99164-6420 The antagonism between gibberellins (GA) and abscisic acid (ABA) is an important factor regulating the developmental transition from embryogenesis to seed germination. In barley aleurone layers, the expression of genes encoding a-amy- lases and proteases is induced by GA but suppressed by ABA. It has been shown that an ABA-induced protein kinase, PKABA1, mediates the ABA suppression of a-amylase expression. Using a barley aleurone transient expression sys- tem, we have now localized the site of action of PKABA1 relative to other signal transduction components governing the expression of a-amylase. The expression of a-amylase can be transactivated by the transcription factor GAMyb, which is itself induced by GA. A truncated GAMyb containing the DNA binding domain but lacking the transactivation domain prevents the GA induction of a-amylase, further supporting the notion that GAMyb mediates the GA induction of a-amylase expression. Although ABA and PKABA1 strongly inhibit the GA induction of a-amylase, they have no ef- fect on GAMyb-transactivated a-amylase expression. Using a GAMyb promoter–b-glucuronidase construct, we also show that both ABA and PKABA1 repress the GA induction of GAMyb. In the slender mutant, GAMyb and a-amylase are highly expressed, even in the absence of GA. However, this constitutive expression can still be inhibited by ABA, PKABA1, or an inhibitor of cGMP synthesis. On the basis of these observations, we suggest that PKABA1 acts up- stream from the formation of functional GAMyb but downstream from the site of action of the Slender gene product. Because PKABA1 inhibits the GA induction of the GAMyb promoter–b-glucuronidase construct, it appears that at least part of the action of PKABA1 is to downregulate GAMyb at the transcriptional level. INTRODUCTION The interaction between phytohormones, particularly that between gibberellin (GA) and abscisic acid (ABA), is an im- portant factor controlling the transition from embryogenesis to germination in seed. During germination of cereal grains, the embryo secretes GA to the aleurone layer, where it pro- motes the expression of several genes encoding hydrolytic enzymes (Ritchie and Gilroy, 1998b; Lovegrove and Hooley, 2000, and references therein). Expression of these genes is blocked by ABA during seed development, in dormant seeds, and in seedlings under unfavorable germination conditions. Cereal aleurone layers, therefore, are an excellent system in which to explore the molecular mechanisms involved in hor- monally regulated gene expression, particularly the antago- nism between GA and ABA (Bethke et al., 1997; Lovegrove and Hooley, 2000). The promoter sequences important for the GA induction and ABA suppression of a-amylase genes have been stud- ied extensively (Skriver et al., 1991; Gubler and Jacobsen, 1992; Lanahan et al., 1992; Rogers and Rogers, 1992). Three regions—box1 (amylase box), GARE (for gibberellin response element), and pyrimidine box—are found in the promoters of all GA-inducible a-amylase genes. An addi- tional region important for GA inducibility (O2S binding box) is found in low pI a-amylase gene promoters. The promoter of a GA-induced cysteine proteinase has elements similar to those found in a-amylase promoters, such as the GARE and the pyrimidine box, and a newly described upstream ele- ment necessary for GA responsiveness (Cercós et al., 1999). The characterization of these promoter elements has pro- vided the basis for the discovery of trans-acting factors in- volved in the transduction of GA signals in barley aleurone layers. A DNA binding protein complex capable of interact- ing with the GARE box and surrounding regions has been shown to be present only in GA-treated aleurone layers This paper is dedicated to Maarten Chrispeels and Joe Varner, who first reported the GA/ABA antagonism in the mid sixties. 1 Current address: Department of Experimental Sciences, Universitat Jaume I, 12071 Castellón, Spain. 2 To whom correspondence should be addressed. E-mail ho@ biology.wustl.edu; fax 314-935-4432.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Gibberellin/Abscisic Acid Antagonism In Barley Aleurone Cells: Site of Action of the Protein Kinase...

The Plant Cell, Vol. 13, 667–679, March 2001, www.plantcell.org © 2001 American Society of Plant Physiologists
Gibberellin/Abscisic Acid Antagonism in Barley Aleurone Cells: Site of Action of the Protein Kinase PKABA1 in Relation to Gibberellin Signaling Molecules
Aurelio Gómez-Cadenas,
a,1
Rodolfo Zentella,
a
Mary Kay Walker-Simmons,
b
and Tuan-Hua David Ho
a,2
a
Plant Biology Program, Department of Biology, Washington University, St. Louis, Missouri 63130
b
United States Department of Agriculture–Agricultural Research Service, Washington State University, Pullman, Washington 99164-6420
The antagonism between gibberellins (GA) and abscisic acid (ABA) is an important factor regulating the developmental
transition from embryogenesis to seed germination. In barley aleurone layers, the expression of genes encoding
a
-amy-lases and proteases is induced by GA but suppressed by ABA. It has been shown that an ABA-induced protein kinase,PKABA1, mediates the ABA suppression of
a
-amylase
expression. Using a barley aleurone transient expression sys-tem, we have now localized the site of action of PKABA1 relative to other signal transduction components governingthe expression of
a
-amylase
. The expression of
a
-amylase
can be transactivated by the transcription factor GAMyb,which is itself induced by GA. A truncated GAMyb containing the DNA binding domain but lacking the transactivationdomain prevents the GA induction of
a
-amylase
, further supporting the notion that GAMyb mediates the GA inductionof
a
-amylase
expression. Although ABA and PKABA1 strongly inhibit the GA induction of
a
-amylase
, they have no ef-fect on GAMyb-transactivated
a
-amylase
expression. Using a
GAMyb
promoter–
b
-glucuronidase
construct, we alsoshow that both ABA and PKABA1 repress the GA induction of
GAMyb
. In the
slender
mutant,
GAMyb
and
a
-amylase
arehighly expressed, even in the absence of GA. However, this constitutive expression can still be inhibited by ABA,PKABA1, or an inhibitor of cGMP synthesis. On the basis of these observations, we suggest that PKABA1 acts up-stream from the formation of functional GAMyb but downstream from the site of action of the
Slender
gene product.Because PKABA1 inhibits the GA induction of the
GAMyb
promoter–
b
-glucuronidase
construct, it appears that at leastpart of the action of PKABA1 is to downregulate
GAMyb
at the transcriptional level.
INTRODUCTION
The interaction between phytohormones, particularly thatbetween gibberellin (GA) and abscisic acid (ABA), is an im-portant factor controlling the transition from embryogenesisto germination in seed. During germination of cereal grains,the embryo secretes GA to the aleurone layer, where it pro-motes the expression of several genes encoding hydrolyticenzymes (Ritchie and Gilroy, 1998b; Lovegrove and Hooley,2000, and references therein). Expression of these genes isblocked by ABA during seed development, in dormant seeds,and in seedlings under unfavorable germination conditions.Cereal aleurone layers, therefore, are an excellent system inwhich to explore the molecular mechanisms involved in hor-monally regulated gene expression, particularly the antago-
nism between GA and ABA (Bethke et al., 1997; Lovegroveand Hooley, 2000).
The promoter sequences important for the GA inductionand ABA suppression of
a
-amylase
genes have been stud-ied extensively (Skriver et al., 1991; Gubler and Jacobsen,1992; Lanahan et al., 1992; Rogers and Rogers, 1992).Three regions—box1 (amylase box), GARE (for gibberellinresponse element), and pyrimidine box—are found in thepromoters of all GA-inducible
a
-amylase
genes. An addi-tional region important for GA inducibility (O2S binding box)is found in low pI
a
-amylase
gene promoters. The promoterof a GA-induced cysteine proteinase has elements similar tothose found in
a
-amylase
promoters, such as the GARE andthe pyrimidine box, and a newly described upstream ele-ment necessary for GA responsiveness (Cercós et al., 1999).The characterization of these promoter elements has pro-vided the basis for the discovery of
trans
-acting factors in-volved in the transduction of GA signals in barley aleuronelayers. A DNA binding protein complex capable of interact-ing with the GARE box and surrounding regions has beenshown to be present only in GA-treated aleurone layers
This paper is dedicated to Maarten Chrispeels and Joe Varner, whofirst reported the GA/ABA antagonism in the mid sixties.
1
Current address: Department of Experimental Sciences, UniversitatJaume I, 12071 Castellón, Spain.
2
To whom correspondence should be addressed. E-mail [email protected]; fax 314-935-4432.
668 The Plant Cell
(Sutliff et al., 1993). Gubler et al. (1995) cloned a GA-induced Myb-like protein (GAMyb) that binds specifically tothe GARE box of an
a
-amylase
promoter. GAMyb is able totransactivate the expression of
a
-amylase
and other GA-regulated genes (Gubler et al., 1995, 1999; Cercós et al.,1999). Several negative regulators of the GA induction ofgene expression have been identified in barley. A zinc fin-gerlike protein has been shown to bind the GARE box andrepress the expression of
a
-amylase
and other genes(Raventos et al., 1998). A barley homolog of the Arabidop-sis
SPY
gene (Jacobsen et al., 1996), which encodes a pu-tative O-linked
N
-acetylglucosamine transferase, stronglydecreases GA induction of the
a
-amylase
gene (Robertsonet al., 1998).
Another putative negative regulator of GA signaling is theprotein encoded by the
Slender1
(
SLN1
) gene. A recessivemutation in this gene leads to pleiotropic phenotypes, suchas rapid elongation of stems and leaves, sterility, and high
a
-amylase production, even in the presence of GA biosyn-thesis inhibitors (Foster, 1977; Chandler, 1988; Lanahan andHo, 1988). The
slender
(
sln1
) mutant phenotype can bemimicked in wild-type plants by exogenous applications ofa high dose of GA. However, the endogenous levels of ac-tive GA in the
slender
mutant are known to be lower thanthose in the wild type (Croker et al., 1990).
Several biochemical studies indicate that GA is perceivedat the plasma membrane (Hooley et al., 1991; Gilroy andJones, 1994) and that increased levels of cytosolic calcium(Gilroy, 1996) and calmodulin (Schuurink et al., 1996) areearly events in signal transduction. G proteins and proteinphosphatases also may be involved in GA signaling (Kuo etal., 1996; Jones et al., 1998). In addition, Penson et al.(1996) determined that cGMP is an important component inthe transduction of the GA signal.
The ABA signal transduction pathway in aleurone layers islargely unknown (for recent reviews, see Leung and Giraudat,1998; Lovegrove and Hooley, 2000). Both plasma mem-brane and internal receptors have been postulated (Gilroyand Jones, 1994; Gilroy, 1996). Phospholipase D has beenproposed as an intermediate in the propagation of ABA sig-naling (Ritchie and Gilroy, 1998a). The levels of cytosolic cal-cium decrease in response to ABA treatment (Gilroy, 1996),suggesting a possible role of calcium as a second messen-ger. The importance of protein phosphorylation in transduc-ing the ABA signal also has been shown in several reports(Sheen, 1996; Bethke et al., 1997; Grill and Himmelbach,1998; Li et al., 2000). Specifically, it has been demonstratedthat ABA’s induction of genes such as
HVA1
and suppres-sion of
a
-amylase
genes diverge in two different signaltransduction pathways, with the induction branch beingsensitive to a protein phosphatase 2C (Shen, 1996) and thesuppressive pathway being modulated by an ABA-respon-sive serine/threonine protein kinase, PKABA1 (Anderbergand Walker-Simmons, 1992; Gómez-Cadenas et al., 1999).
PKABA1
transcript levels increase in response to ABA inscutellar, root, and shoot tissues (Holappa and Walker-
Simmons, 1995). As ABA levels increase in drying seeds,
PKABA1
transcript levels increase, reaching their peak atseed maturity.
PKABA1
transcript levels remain high inABA-treated aleurone layers but decrease below detectablelevels in GA-treated aleurone.
Although it is known that PKABA1 represses the GA in-duction of genes coding for hydrolytic enzymes, the molecu-lar mechanism underlying the GA/ABA antagonism has notbeen fully explored. In this work, using particle bombard-ment coupled with the transient expression of transgenes inbarley aleurone layers, we studied how the ABA and GA sig-
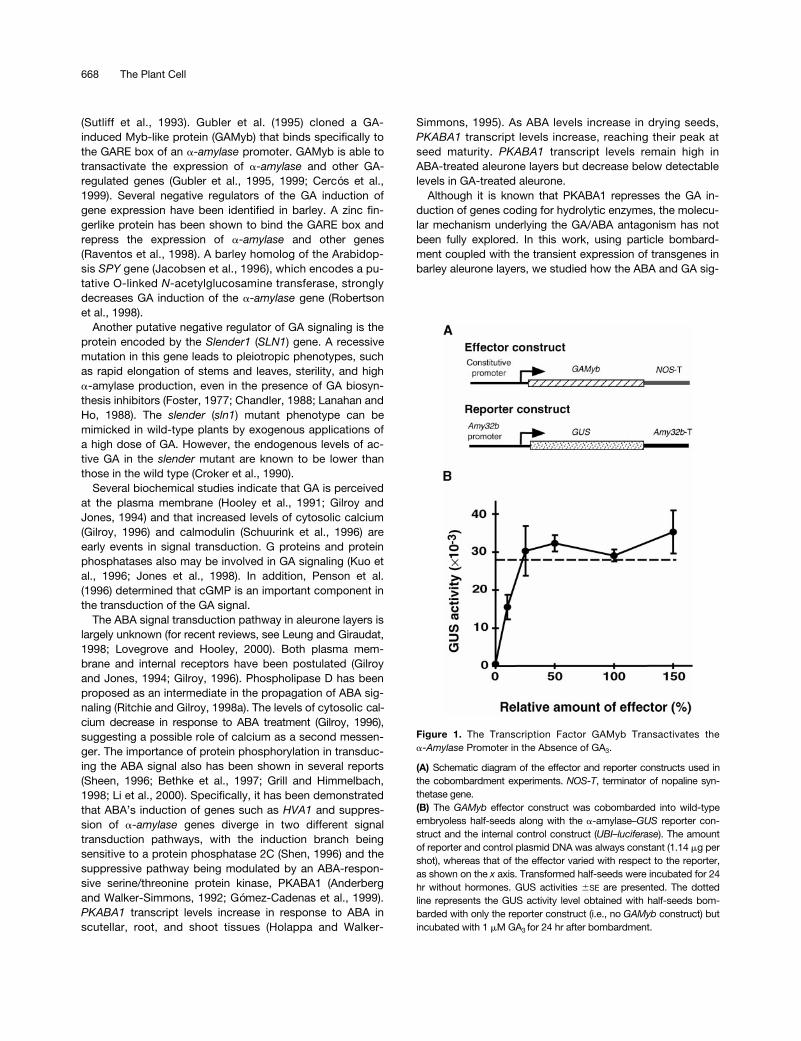
Figure 1. The Transcription Factor GAMyb Transactivates thea-Amylase Promoter in the Absence of GA3.
(A) Schematic diagram of the effector and reporter constructs used inthe cobombardment experiments. NOS-T, terminator of nopaline syn-thetase gene.(B) The GAMyb effector construct was cobombarded into wild-typeembryoless half-seeds along with the a-amylase–GUS reporter con-struct and the internal control construct (UBI–luciferase). The amountof reporter and control plasmid DNA was always constant (1.14 mg pershot), whereas that of the effector varied with respect to the reporter,as shown on the x axis. Transformed half-seeds were incubated for 24hr without hormones. GUS activities 6SE are presented. The dottedline represents the GUS activity level obtained with half-seeds bom-barded with only the reporter construct (i.e., no GAMyb construct) butincubated with 1 mM GA3 for 24 hr after bombardment.
GA/ABA Antagonism in Barley Aleurone 669
nal transduction cascades interact with each other. We showthat PKABA1 acts as an intermediary of the ABA signal trans-duction pathway, repressing the GA induction of
GAMyb
and therefore of hydrolytic enzymes. The involvement of the
slender
mutant is also addressed, and our observations al-low us to locate SLN1 in the GA signal transduction path-way acting upstream of GAMyb and PKABA1.
RESULTS
The Transcription Factor GAMyb Transactivates
a
-Amylase
Promoters
The GA induction of
a
-amylase
gene expression has beenstudied extensively (Ritchie and Gilroy, 1998b, and refer-ences therein). The role of the transcription factor GAMyb asa regulator of
a
-amylase
expression has been shown(Gubler et al., 1995). To test this effect in our experimentalsystem, we cotransformed barley embryoless half-seed (i.e.,without an endogenous source of GA) with the
GAMyb
cDNA driven by the constitutive
Ubi-1
promoter along with alow pI
a
-amylase
–
b
-
glucuronidase
(
GUS
) reporter constructand subsequently incubated the transformed tissue for 24 hrin the absence of hormones (Figure 1). When the
a
-amylase
reporter construct was bombarded alone (Figure 1B, 0 onthe
x
axis), low basal GUS activity levels were detected.Cotransformation of the reporter construct with the
GAMyb
effector construct (Figure 1A) strongly increased the GUSactivity level in the absence of gibberellic acid (GA
3
) treat-ment (Figure 1B). This transactivation of
a
-amylase
expres-sion (as measured by GUS activity) followed a dosage-dependent pattern and eventually reached a level compara-ble to that of GA
3
-treated half-seed bombarded only withthe reporter construct (Figure 1B, dotted line). Similar resultswere obtained with a high pI
a
-amylase
reporter construct(data not shown). The maize C1 protein, a different Myb-likeprotein (Paz-Ares et al., 1987) driven by the same constitu-tive promoter, failed to transactivate the
a
-amylase
promot-ers (data not shown; Gubler et al., 1995).
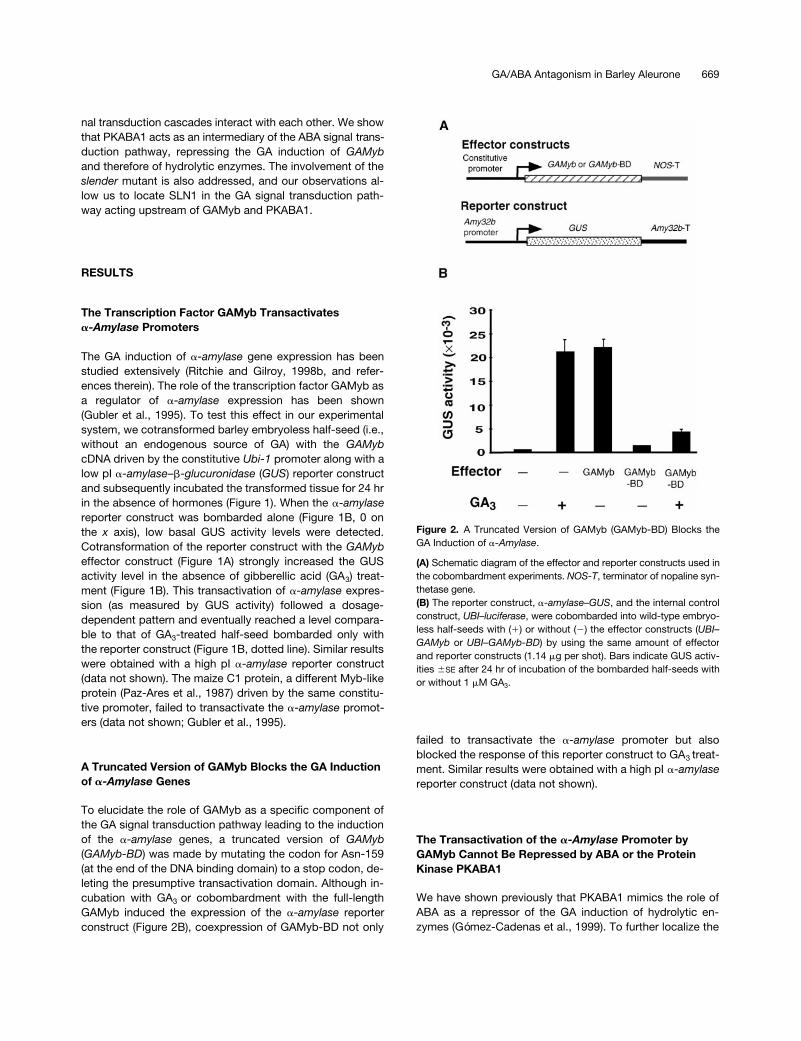
A Truncated Version of GAMyb Blocks the GA Induction of
a
-Amylase
Genes
To elucidate the role of GAMyb as a specific component ofthe GA signal transduction pathway leading to the inductionof the
a
-amylase
genes, a truncated version of
GAMyb
(
GAMyb-BD
) was made by mutating the codon for Asn-159(at the end of the DNA binding domain) to a stop codon, de-leting the presumptive transactivation domain. Although in-cubation with GA
3
or cobombardment with the full-lengthGAMyb induced the expression of the
a
-amylase
reporterconstruct (Figure 2B), coexpression of GAMyb-BD not only
failed to transactivate the
a
-amylase
promoter but alsoblocked the response of this reporter construct to GA
3
treat-ment. Similar results were obtained with a high pI
a
-amylase
reporter construct (data not shown).
The Transactivation of the
a
-Amylase
Promoter by GAMyb Cannot Be Repressed by ABA or the Protein Kinase PKABA1
We have shown previously that PKABA1 mimics the role ofABA as a repressor of the GA induction of hydrolytic en-zymes (Gómez-Cadenas et al., 1999). To further localize the
Figure 2. A Truncated Version of GAMyb (GAMyb-BD) Blocks theGA Induction of a-Amylase.
(A) Schematic diagram of the effector and reporter constructs used inthe cobombardment experiments. NOS-T, terminator of nopaline syn-thetase gene.(B) The reporter construct, a-amylase–GUS, and the internal controlconstruct, UBI–luciferase, were cobombarded into wild-type embryo-less half-seeds with (1) or without (2) the effector constructs (UBI–GAMyb or UBI–GAMyb-BD) by using the same amount of effectorand reporter constructs (1.14 mg per shot). Bars indicate GUS activ-ities 6SE after 24 hr of incubation of the bombarded half-seeds withor without 1 mM GA3.
670 The Plant Cell
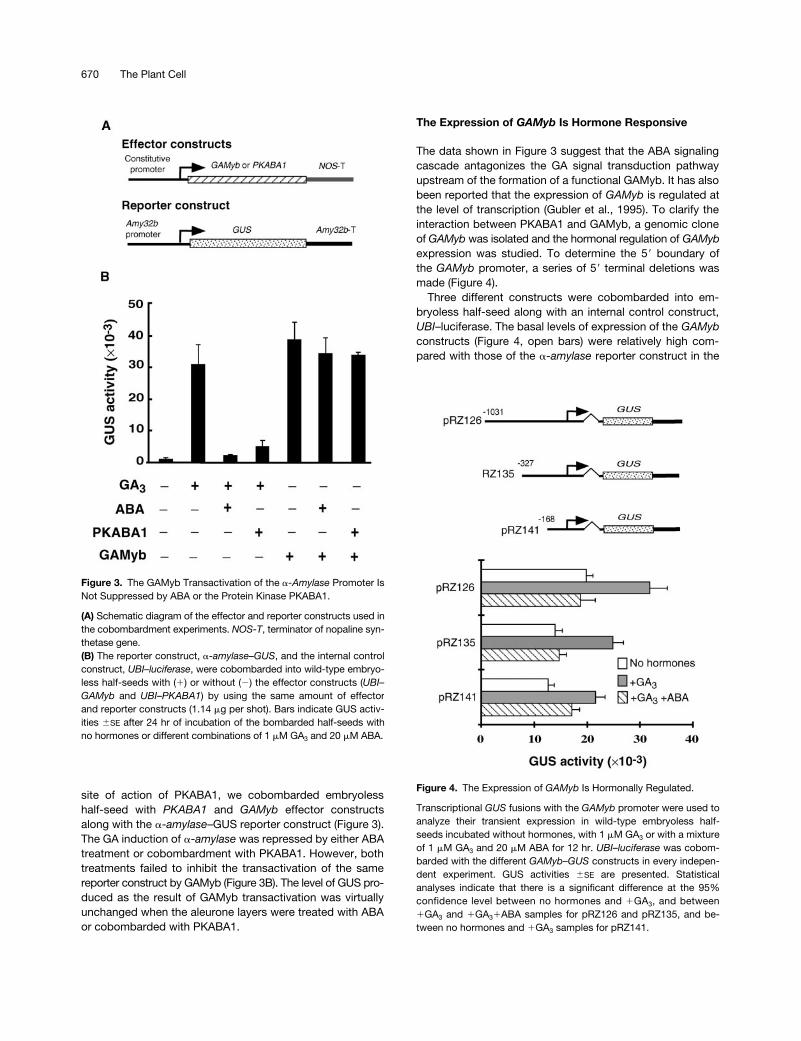
site of action of PKABA1, we cobombarded embryolesshalf-seed with PKABA1 and GAMyb effector constructsalong with the a-amylase–GUS reporter construct (Figure 3).The GA induction of a-amylase was repressed by either ABAtreatment or cobombardment with PKABA1. However, bothtreatments failed to inhibit the transactivation of the samereporter construct by GAMyb (Figure 3B). The level of GUS pro-duced as the result of GAMyb transactivation was virtuallyunchanged when the aleurone layers were treated with ABAor cobombarded with PKABA1.
The Expression of GAMyb Is Hormone Responsive
The data shown in Figure 3 suggest that the ABA signalingcascade antagonizes the GA signal transduction pathwayupstream of the formation of a functional GAMyb. It has alsobeen reported that the expression of GAMyb is regulated atthe level of transcription (Gubler et al., 1995). To clarify theinteraction between PKABA1 and GAMyb, a genomic cloneof GAMyb was isolated and the hormonal regulation of GAMybexpression was studied. To determine the 59 boundary ofthe GAMyb promoter, a series of 59 terminal deletions wasmade (Figure 4).
Three different constructs were cobombarded into em-bryoless half-seed along with an internal control construct,UBI–luciferase. The basal levels of expression of the GAMybconstructs (Figure 4, open bars) were relatively high com-pared with those of the a-amylase reporter construct in the
Figure 3. The GAMyb Transactivation of the a-Amylase Promoter IsNot Suppressed by ABA or the Protein Kinase PKABA1.
(A) Schematic diagram of the effector and reporter constructs used inthe cobombardment experiments. NOS-T, terminator of nopaline syn-thetase gene.(B) The reporter construct, a-amylase–GUS, and the internal controlconstruct, UBI–luciferase, were cobombarded into wild-type embryo-less half-seeds with (1) or without (2) the effector constructs (UBI–GAMyb and UBI–PKABA1) by using the same amount of effectorand reporter constructs (1.14 mg per shot). Bars indicate GUS activ-ities 6SE after 24 hr of incubation of the bombarded half-seeds withno hormones or different combinations of 1 mM GA3 and 20 mM ABA.
Figure 4. The Expression of GAMyb Is Hormonally Regulated.
Transcriptional GUS fusions with the GAMyb promoter were used toanalyze their transient expression in wild-type embryoless half-seeds incubated without hormones, with 1 mM GA3 or with a mixtureof 1 mM GA3 and 20 mM ABA for 12 hr. UBI–luciferase was cobom-barded with the different GAMyb–GUS constructs in every indepen-dent experiment. GUS activities 6SE are presented. Statisticalanalyses indicate that there is a significant difference at the 95%confidence level between no hormones and 1GA3, and between1GA3 and 1GA31ABA samples for pRZ126 and pRZ135, and be-tween no hormones and 1GA3 samples for pRZ141.
GA/ABA Antagonism in Barley Aleurone 671
absence of any effector construct (Figures 1 and 2). Incuba-tion with GA3 for 12 hr induced the expression of all threeconstructs (Figure 4, gray versus open bars). The constructwith 327 bp of 59 flanking sequence (pRZ135) displayed thehighest GA induction. On the other hand, the GA inductionof these constructs was still repressed by the presence ofABA, although deleting to 2168 (pRZ141) resulted in thepartial loss of ABA suppression, that is, no significant differ-ence between 1GA3 and 1GA31ABA samples anymore.However, the levels of basal expression were still high (Fig-ure 4). In light of these observations, the pRZ135 constructwas chosen for subsequent experiments.
To analyze the time course of the GA induction and ABAsuppression of GAMyb, the construct pRZ135 was bom-barded into embryoless half-seed, which were then incu-
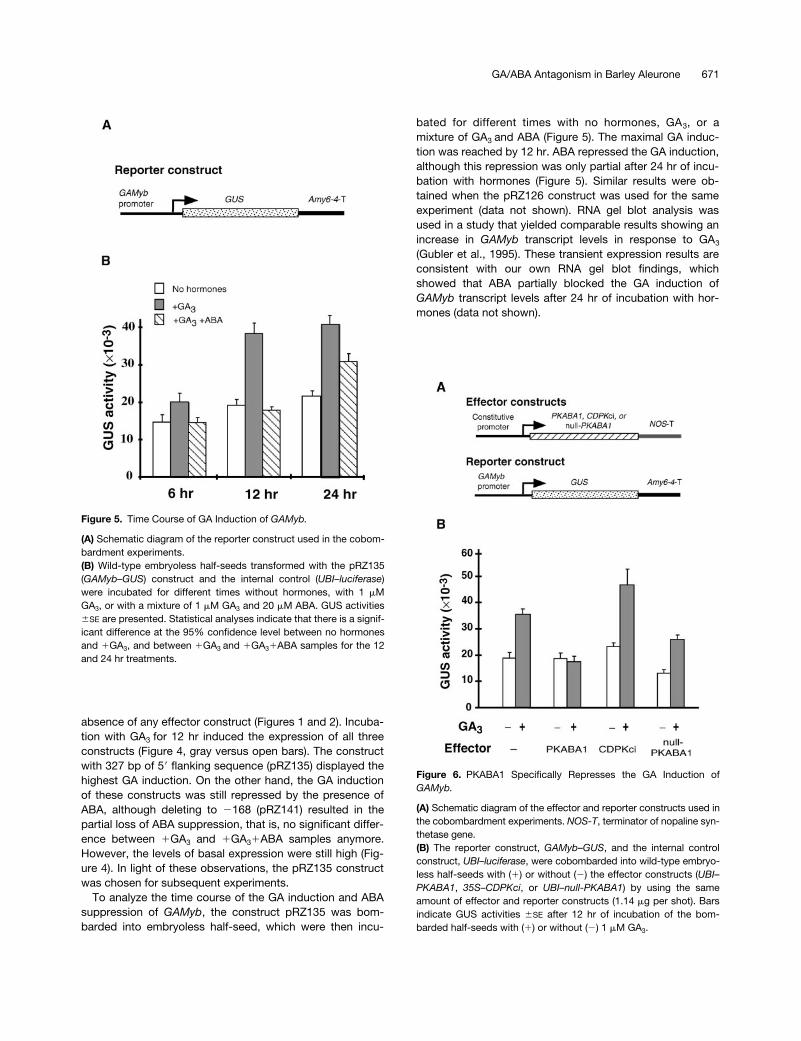
bated for different times with no hormones, GA3, or amixture of GA3 and ABA (Figure 5). The maximal GA induc-tion was reached by 12 hr. ABA repressed the GA induction,although this repression was only partial after 24 hr of incu-bation with hormones (Figure 5). Similar results were ob-tained when the pRZ126 construct was used for the sameexperiment (data not shown). RNA gel blot analysis wasused in a study that yielded comparable results showing anincrease in GAMyb transcript levels in response to GA3
(Gubler et al., 1995). These transient expression results areconsistent with our own RNA gel blot findings, whichshowed that ABA partially blocked the GA induction ofGAMyb transcript levels after 24 hr of incubation with hor-mones (data not shown).
Figure 5. Time Course of GA Induction of GAMyb.
(A) Schematic diagram of the reporter construct used in the cobom-bardment experiments.(B) Wild-type embryoless half-seeds transformed with the pRZ135(GAMyb–GUS) construct and the internal control (UBI–luciferase)were incubated for different times without hormones, with 1 mMGA3, or with a mixture of 1 mM GA3 and 20 mM ABA. GUS activities6SE are presented. Statistical analyses indicate that there is a signif-icant difference at the 95% confidence level between no hormonesand 1GA3, and between 1GA3 and 1GA31ABA samples for the 12and 24 hr treatments.
Figure 6. PKABA1 Specifically Represses the GA Induction ofGAMyb.
(A) Schematic diagram of the effector and reporter constructs used inthe cobombardment experiments. NOS-T, terminator of nopaline syn-thetase gene.(B) The reporter construct, GAMyb–GUS, and the internal controlconstruct, UBI–luciferase, were cobombarded into wild-type embryo-less half-seeds with (1) or without (2) the effector constructs (UBI–PKABA1, 35S–CDPKci, or UBI–null-PKABA1) by using the sameamount of effector and reporter constructs (1.14 mg per shot). Barsindicate GUS activities 6SE after 12 hr of incubation of the bom-barded half-seeds with (1) or without (2) 1 mM GA3.

672 The Plant Cell
PKABA1 Specifically Represses the GA Inductionof GAMyb
To examine the possible role of PKABA1 in mediating ABAsuppression of the GAMyb promoter, we cobombarded em-bryoless barley half-seed with the effector construct UBI–PKABA1 along with the GAMyb reporter construct (Figure 6).In the absence of the effector construct, the expression ofGAMyb was induced after 12 hr of incubation with GA3. How-ever, coexpression of PKABA1 along with the GAMyb reporterconstruct blocked the response to GA3. Similar results wereobtained using PKABA1 cDNA driven by the cauliflower mo-saic virus 35S promoter (35S) (data not shown). As controls,cobombardment of a different protein kinase, CDPKci (Harperet al., 1994), or a null version of PKABA1 (without the glycine-rich loop that is thought to be necessary for nucleotide bind-ing and protein kinase activity) (Gómez-Cadenas et al., 1999)driven by the 35S or Ubi-1 promoter, failed to repress GA in-duction of the reporter construct (Figure 6).
PKABA1 Blocks the Constitutive Expression ofa-Amylase and GAMyb in the slender Mutant
The slender mutation in barley is a single-locus recessivemutation that causes a plant to appear as if it had beengrown in saturating concentrations of GA, even in the pres-ence of GA biosynthesis inhibitors (Foster, 1977; Chandler,1988; Lanahan and Ho, 1988). This phenotype is observeddespite the fact that endogenous levels of active GA arelower in the slender mutant than in the wild type (Croker etal., 1990). Thus, SLN1 is probably involved in the repressionof the GA signal transduction pathway. We studied the roleof SLN1 in GA-mediated gene expression in aleurone layersusing transient expression analysis. As shown in Table 1, inthe wild-type aleurone layers, expression of a-amylase andGAMyb reporter constructs was induced by GA3 treatment.However, in the slender aleurone layers, both reporter con-structs showed increased levels of expression in the ab-sence of GA3 treatment. GA3 treatment of slender aleuronelayers did not enhance the expression of a-amylase orGAMyb reporter constructs.
To understand the relationship among PKABA1, SLN1,
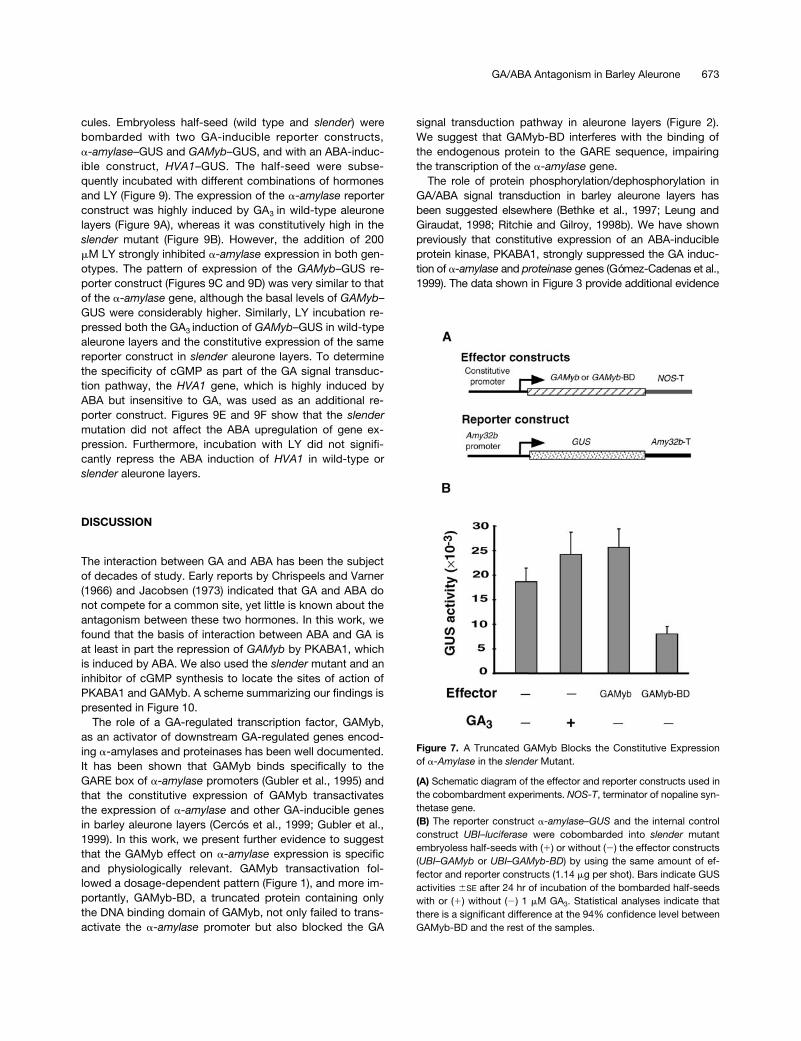
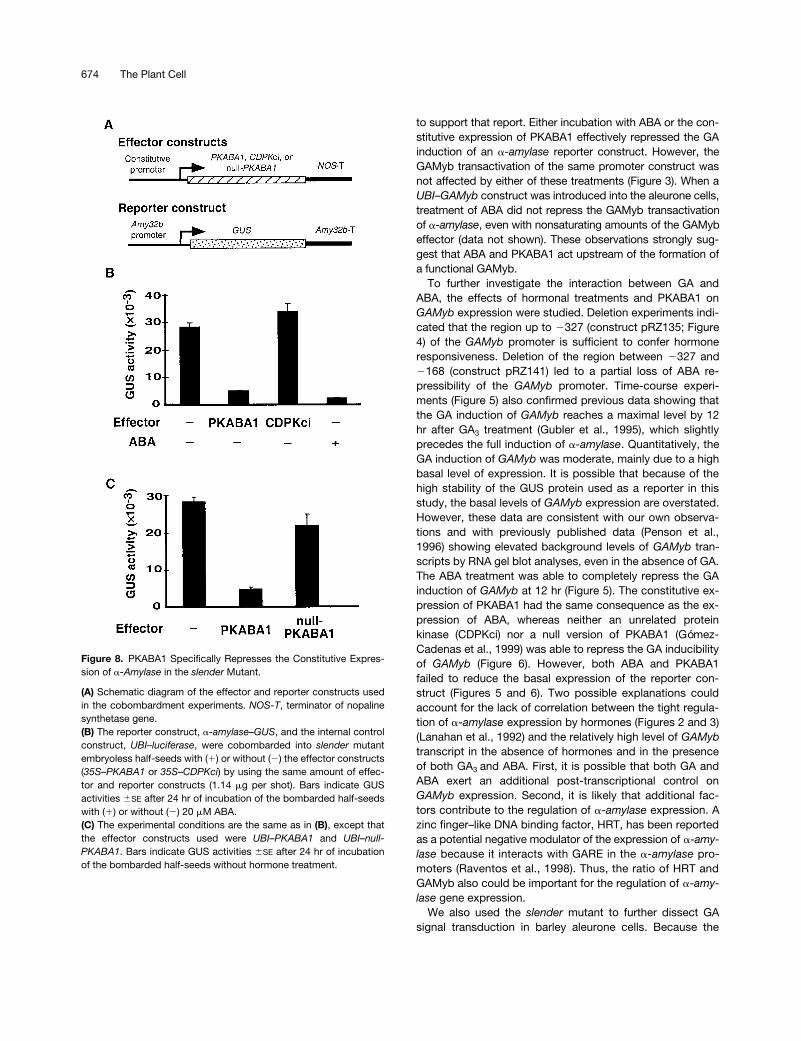
and GAMyb, two experiments were performed in the slendermutant background. First, to further investigate the role ofGAMyb as a key element in the activation of a-amylasegenes, the effect of GAMyb-BD was tested on the slenderaleurone layers. As expected, the expression of the a-amy-lase reporter construct was high in the absence of hor-mones (Figure 7). Incubation with GA3 or cobombardmentwith GAMyb slightly increased the already high levels of ex-pression of a-amylase. However, cobombardment withGAMyb-BD reduced the constitutive expression of a-amy-lase by 50%. Second, to test the role of PKABA1 as a puta-tive repressor of the constitutive expression of a-amylasegenes in the slender mutant, several effector constructswere cobombarded into slender embryoless half-seed alongwith the a-amylase reporter construct (Figure 8). Coexpres-sion of either 35S–PKABA1 (Figure 8B) or UBI–PKABA1(Figure 8C) strongly repressed the constitutively high levelsof expression of the a-amylase construct. However, coex-pression of another protein kinase (CDPKci) or a null versionof PKABA1 did not significantly alter the high levels of ex-pression of a-amylase in slender aleurone layers. Figure 8Balso shows that incubation with ABA effectively blocked theexpression of a-amylase in the mutant tissue.
When GAMyb–GUS was used as a reporter construct(data not shown), ABA treatment or cobombardment withPKABA1 significantly repressed the constitutive expressionof the GAMyb construct by 40% in the aleurone layers ofslender mutant barley.
LY83583, a Guanylyl Cyclase Inhibitor, Blocks GA Signal Transduction Upstream of GAMyb but Downstreamof SLN1
The role of cGMP in hormone-regulated gene expressionwas established by Penson et al. (1996), who showed thatcGMP levels increased in response to GA treatment in bar-ley aleurone layers. Furthermore, the presence of a guanylylcyclase inhibitor, LY83583 (LY), blocked the GA-inducedaccumulation of a-amylase and GAMyb transcripts. How-ever, cGMP alone was not sufficient to increase either ofthese transcripts (Penson et al., 1996). We also have testedthe site of action of cGMP relative to other regulatory mole-
Table 1. Levels of Expression of a-Amylase and GAMyb in the Wild Type and the slender Mutant Barleya
Expression of a-Amylase–GUS Expression of GAMyb–GUS
Hormone Treatment Wild Type slender Mutant Wild Type slender Mutant
No hormone 0.2 6 0.0 8.7 6 1.2 14.4 6 2.0 20.5 6 2.11GA3 16.6 6 2.7 10.3 6 1.2 22.9 6 1.3 22.2 6 3.8
a Either the a-amylase or the GAMyb reporter construct was bombarded into slender mutant embryoless half-seed along with the internal controlconstruct UBI–luciferase, using the same amount of effector and reporter constructs (1.14 mg per shot). Transformed half-seeds were incubatedfor 12 hr with or without 1 mM GA3. GUS activities (31023) 6SE are presented.
GA/ABA Antagonism in Barley Aleurone 673
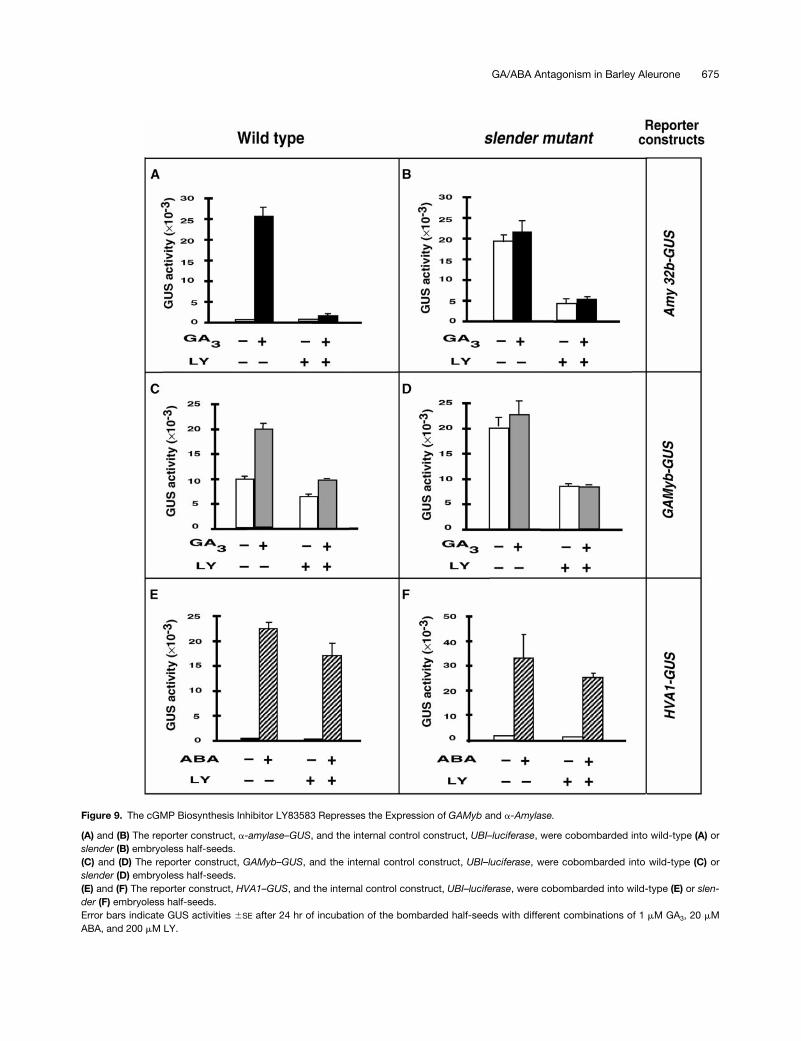
cules. Embryoless half-seed (wild type and slender) werebombarded with two GA-inducible reporter constructs,a-amylase–GUS and GAMyb–GUS, and with an ABA-induc-ible construct, HVA1–GUS. The half-seed were subse-quently incubated with different combinations of hormonesand LY (Figure 9). The expression of the a-amylase reporterconstruct was highly induced by GA3 in wild-type aleuronelayers (Figure 9A), whereas it was constitutively high in theslender mutant (Figure 9B). However, the addition of 200mM LY strongly inhibited a-amylase expression in both gen-otypes. The pattern of expression of the GAMyb–GUS re-porter construct (Figures 9C and 9D) was very similar to thatof the a-amylase gene, although the basal levels of GAMyb–GUS were considerably higher. Similarly, LY incubation re-pressed both the GA3 induction of GAMyb–GUS in wild-typealeurone layers and the constitutive expression of the samereporter construct in slender aleurone layers. To determinethe specificity of cGMP as part of the GA signal transduc-tion pathway, the HVA1 gene, which is highly induced byABA but insensitive to GA, was used as an additional re-porter construct. Figures 9E and 9F show that the slendermutation did not affect the ABA upregulation of gene ex-pression. Furthermore, incubation with LY did not signifi-cantly repress the ABA induction of HVA1 in wild-type orslender aleurone layers.
DISCUSSION
The interaction between GA and ABA has been the subjectof decades of study. Early reports by Chrispeels and Varner(1966) and Jacobsen (1973) indicated that GA and ABA donot compete for a common site, yet little is known about theantagonism between these two hormones. In this work, wefound that the basis of interaction between ABA and GA isat least in part the repression of GAMyb by PKABA1, whichis induced by ABA. We also used the slender mutant and aninhibitor of cGMP synthesis to locate the sites of action ofPKABA1 and GAMyb. A scheme summarizing our findings ispresented in Figure 10.
The role of a GA-regulated transcription factor, GAMyb,as an activator of downstream GA-regulated genes encod-ing a-amylases and proteinases has been well documented.It has been shown that GAMyb binds specifically to theGARE box of a-amylase promoters (Gubler et al., 1995) andthat the constitutive expression of GAMyb transactivatesthe expression of a-amylase and other GA-inducible genesin barley aleurone layers (Cercós et al., 1999; Gubler et al.,1999). In this work, we present further evidence to suggestthat the GAMyb effect on a-amylase expression is specificand physiologically relevant. GAMyb transactivation fol-lowed a dosage-dependent pattern (Figure 1), and more im-portantly, GAMyb-BD, a truncated protein containing onlythe DNA binding domain of GAMyb, not only failed to trans-activate the a-amylase promoter but also blocked the GA
signal transduction pathway in aleurone layers (Figure 2).We suggest that GAMyb-BD interferes with the binding ofthe endogenous protein to the GARE sequence, impairingthe transcription of the a-amylase gene.
The role of protein phosphorylation/dephosphorylation inGA/ABA signal transduction in barley aleurone layers hasbeen suggested elsewhere (Bethke et al., 1997; Leung andGiraudat, 1998; Ritchie and Gilroy, 1998b). We have shownpreviously that constitutive expression of an ABA-inducibleprotein kinase, PKABA1, strongly suppressed the GA induc-tion of a-amylase and proteinase genes (Gómez-Cadenas et al.,1999). The data shown in Figure 3 provide additional evidence
Figure 7. A Truncated GAMyb Blocks the Constitutive Expressionof a-Amylase in the slender Mutant.
(A) Schematic diagram of the effector and reporter constructs used inthe cobombardment experiments. NOS-T, terminator of nopaline syn-thetase gene.(B) The reporter construct a-amylase–GUS and the internal controlconstruct UBI–luciferase were cobombarded into slender mutantembryoless half-seeds with (1) or without (2) the effector constructs(UBI–GAMyb or UBI–GAMyb-BD) by using the same amount of ef-fector and reporter constructs (1.14 mg per shot). Bars indicate GUSactivities 6SE after 24 hr of incubation of the bombarded half-seedswith or (1) without (2) 1 mM GA3. Statistical analyses indicate thatthere is a significant difference at the 94% confidence level betweenGAMyb-BD and the rest of the samples.
674 The Plant Cell
to support that report. Either incubation with ABA or the con-stitutive expression of PKABA1 effectively repressed the GAinduction of an a-amylase reporter construct. However, theGAMyb transactivation of the same promoter construct wasnot affected by either of these treatments (Figure 3). When aUBI–GAMyb construct was introduced into the aleurone cells,treatment of ABA did not repress the GAMyb transactivationof a-amylase, even with nonsaturating amounts of the GAMybeffector (data not shown). These observations strongly sug-gest that ABA and PKABA1 act upstream of the formation ofa functional GAMyb.
To further investigate the interaction between GA andABA, the effects of hormonal treatments and PKABA1 onGAMyb expression were studied. Deletion experiments indi-cated that the region up to 2327 (construct pRZ135; Figure4) of the GAMyb promoter is sufficient to confer hormoneresponsiveness. Deletion of the region between 2327 and2168 (construct pRZ141) led to a partial loss of ABA re-pressibility of the GAMyb promoter. Time-course experi-ments (Figure 5) also confirmed previous data showing thatthe GA induction of GAMyb reaches a maximal level by 12hr after GA3 treatment (Gubler et al., 1995), which slightlyprecedes the full induction of a-amylase. Quantitatively, theGA induction of GAMyb was moderate, mainly due to a highbasal level of expression. It is possible that because of thehigh stability of the GUS protein used as a reporter in thisstudy, the basal levels of GAMyb expression are overstated.However, these data are consistent with our own observa-tions and with previously published data (Penson et al.,1996) showing elevated background levels of GAMyb tran-scripts by RNA gel blot analyses, even in the absence of GA.The ABA treatment was able to completely repress the GAinduction of GAMyb at 12 hr (Figure 5). The constitutive ex-pression of PKABA1 had the same consequence as the ex-pression of ABA, whereas neither an unrelated proteinkinase (CDPKci) nor a null version of PKABA1 (Gómez-Cadenas et al., 1999) was able to repress the GA inducibilityof GAMyb (Figure 6). However, both ABA and PKABA1failed to reduce the basal expression of the reporter con-struct (Figures 5 and 6). Two possible explanations couldaccount for the lack of correlation between the tight regula-tion of a-amylase expression by hormones (Figures 2 and 3)(Lanahan et al., 1992) and the relatively high level of GAMybtranscript in the absence of hormones and in the presenceof both GA3 and ABA. First, it is possible that both GA andABA exert an additional post-transcriptional control onGAMyb expression. Second, it is likely that additional fac-tors contribute to the regulation of a-amylase expression. Azinc finger–like DNA binding factor, HRT, has been reportedas a potential negative modulator of the expression of a-amy-lase because it interacts with GARE in the a-amylase pro-moters (Raventos et al., 1998). Thus, the ratio of HRT andGAMyb also could be important for the regulation of a-amy-lase gene expression.
We also used the slender mutant to further dissect GAsignal transduction in barley aleurone cells. Because the
Figure 8. PKABA1 Specifically Represses the Constitutive Expres-sion of a-Amylase in the slender Mutant.
(A) Schematic diagram of the effector and reporter constructs usedin the cobombardment experiments. NOS-T, terminator of nopalinesynthetase gene.(B) The reporter construct, a-amylase–GUS, and the internal controlconstruct, UBI–luciferase, were cobombarded into slender mutantembryoless half-seeds with (1) or without (2) the effector constructs(35S–PKABA1 or 35S–CDPKci) by using the same amount of effec-tor and reporter constructs (1.14 mg per shot). Bars indicate GUSactivities 6SE after 24 hr of incubation of the bombarded half-seedswith (1) or without (2) 20 mM ABA.(C) The experimental conditions are the same as in (B), except thatthe effector constructs used were UBI–PKABA1 and UBI–null-PKABA1. Bars indicate GUS activities 6SE after 24 hr of incubationof the bombarded half-seeds without hormone treatment.
GA/ABA Antagonism in Barley Aleurone 675
Figure 9. The cGMP Biosynthesis Inhibitor LY83583 Represses the Expression of GAMyb and a-Amylase.
(A) and (B) The reporter construct, a-amylase–GUS, and the internal control construct, UBI–luciferase, were cobombarded into wild-type (A) orslender (B) embryoless half-seeds.(C) and (D) The reporter construct, GAMyb–GUS, and the internal control construct, UBI–luciferase, were cobombarded into wild-type (C) orslender (D) embryoless half-seeds.(E) and (F) The reporter construct, HVA1–GUS, and the internal control construct, UBI–luciferase, were cobombarded into wild-type (E) or slen-der (F) embryoless half-seeds.Error bars indicate GUS activities 6SE after 24 hr of incubation of the bombarded half-seeds with different combinations of 1 mM GA3, 20 mMABA, and 200 mM LY.

676 The Plant Cell
slender mutation is recessive, the wild-type Slender gene(SLN1) is likely to encode a negative regulator of the GA sig-nal cascade (Foster, 1977; Chandler, 1988; Lanahan andHo, 1988). More importantly, the effect of the slender muta-tion on a-amylase expression can be blocked completely bytreatment with 20 mM ABA (Lanahan and Ho, 1988), whichsuggests that the regulatory function of this gene is exertedearly in the GA signal transduction cascade. The expressionof both the GAMyb and a-amylase reporter constructs wasincreased in the slender aleurone layers, which is compara-ble to the expression in GA3-treated wild-type aleurone lay-ers (Figures 7 to 9). Moreover, GA3 treatment of slenderaleurone layers enhanced a-amylase and GAMyb expres-sion only slightly beyond the already high levels. Our datareinforce the importance of GAMyb as a central regulator ofGA signal transduction, even in the slender mutant back-ground, because GAMyb-BD still blocked the expression ofthe a-amylase reporter construct in the slender mutant (Fig-ure 7). Furthermore, the constitutive expression of a-amylaseand GAMyb can be repressed by ABA treatment or cotrans-formation with PKABA1 (Figure 8).
The specificity of PKABA1 mimicking the ABA effect is sug-gested by the fact that a different protein kinase or a null ver-sion of PKABA1 did not have a significant effect on theexpression of GAMyb or a-amylase. Moreover, the ABA sig-nal transduction pathway does not appear to be affected bythe slender mutation (Chandler, 1988; Lanahan and Ho,1988). In fact, Figures 9E and 9F show that the ABA upregula-tion of the HVA1 gene was not affected by the slender muta-tion. Collectively, these data suggest that the point ofinteraction between the ABA (mediated by PKABA1) and GAsignal transduction pathways occurs downstream of the ac-tion of SLN1 but upstream of the action of GAMyb. The mo-lecular nature of SLN1 has been the subject of intenseinvestigation. By means of restriction length polymorphismanalysis, it has been shown that the SLN1 and HvSPY genesare two different loci (Robertson et al., 1998). Because theslender mutation is recessive, it is likely that SLN1 encodes arepressor and that the role of GA is to inhibit the repressiveaction of SLN1, thus allowing the downstream signaling to
proceed, similar to what has been proposed for the action ofGAI and RGA in Arabidopsis (Peng et al., 1999; Sun, 2000).
Finally, the data in Figure 9 show that LY, a guanylyl cy-clase inhibitor, is able to repress the GA induction of GAMyband a-amylase reporter constructs in both wild-type andslender aleurone layers. However, the same inhibitor doesnot affect the ABA induction of HVA1 in both genetic back-grounds. These data, together with the observation that GAinduces a rapid accumulation of cGMP in barley aleuronelayers (Penson et al., 1996), reinforce the model suggestingthat cGMP acts as a second messenger in the GA signaltransduction pathway (Penson et al., 1996). cGMP alonecannot substitute for the GA regulation of a-amylase syn-thesis, which suggests cooperation among cGMP and otherelements (probably Ca21 and/or calmodulin) in the transduc-tion of the GA signal (Penson et al., 1996). Our results sug-gest that SLN1 acts upstream from or at the same level asguanylyl cyclase(s) in GA signal transduction. The relation-ship between guanylyl cyclase(s) and PKABA1 is not yetclear, and more investigation is needed. However, ABAtreatment does not modify the steady state levels of cGMPin aleurone layers (Penson et al., 1996). Therefore, we spec-ulate that ABA (and PKABA1) blocks the GA signaling cas-cade downstream of the point of action of cGMP. Guanylylcyclases appear to be a central component in other plantsignal transduction networks, including light detection(Brown et al., 1989) and phytochrome A responses (Bowlerand Chua, 1994). However, the molecular mechanism ofcGMP action in plants is not yet well understood. In mam-malian systems, cGMP can alter calcium levels that in turncontrol protein kinase activities and gene transcription (Buttet al., 1993; Wang and Robinson, 1997).
In summary, the data presented here provide new insightsinto the molecular interaction between the ABA and GAsignal transduction pathways. We propose that proteinphosphorylation, and particularly PKABA1, are important el-ements connecting the ABA cascade of events to the sup-pression of a-amylase gene expression. PKABA1 blocks theGA upregulation of GAMyb and therefore the expression ofa-amylase in both wild-type and slender mutant aleuronelayers. Our data also reinforce the role of cGMP as an im-portant intermediary in GA signal transduction, most likelyacting downstream of SLN1. Although additional regulatorymolecules or different levels of regulation of PKABA1 and/orGAMyb cannot be excluded, we have demonstrated that atleast part of the action of PKABA1 is to downregulateGAMyb at the transcriptional level.
METHODS
Plant Material
Barley (Hordeum vulgare cv Himalaya) seed were used for all of theexperiments reported in this work. For the experiments involving
Figure 10. Diagram Summarizing the Molecular Events Involved inthe Interaction between ABA and GA Signal Transduction Pathways.

GA/ABA Antagonism in Barley Aleurone 677
slender seed, due to the sterility of the homozygous mutant, theprogeny had to be maintained as a heterozygous population. Half-seed–containing embryos were planted to determine their pheno-types, and only the slender homozygous embryoless half-seed wereused for transient expression.
Preparation of the DNA Constructs
Reporter constructs were prepared as follows. (1) a-Amylase–b-glucuronidase (GUS) was constructed by linking the promoter(up to 2331), the entire 59 untranslated sequence, and the first in-tron of the low pI a-amylase gene, Amy 32b, to the GUS codingsequence and the 39 untranslated region of the same a-amylasegene (Lanahan et al., 1992). (2) HVA1–GUS was made by ligatingthe 68-bp ABRC3 (abscisic acid [ABA] response complex 3,derived from the HVA1 promoter sequence) to the GUS coding se-quence (Shen et al., 1996). (3) GAMyb–GUS constructs are de-scribed below.
Effector constructs were prepared as follows. (1) UBI–GAMyb con-sisted of the maize Ubi-1 promoter linked to the coding region ofGAMyb and to the 39 untranslated region of the nopaline synthetasegene, NOS-T (Gubler et al., 1995). (2) GAMyb-BD was made by mu-tating the Asn at position 159 to a stop codon in the UBI–GAMybconstruct by site-directed mutagenesis (Cercós et al., 1999). (3) UBI–PKABA1 was made by cloning a reconstituted PKABA1 cDNA(Anderberg and Walker-Simmons, 1992; Gómez-Cadenas et al.,1999) into the plant expression vector pAHC17 (Christensen andQuail, 1996). (4) UBI–null-PKABA1 is identical to UBI–PKABA1 butlacks the glycine-rich loop (GSGNFG, amino acids 11 to 16), which ispart of the nucleotide binding site (Gómez-Cadenas et al., 1999). (5)35S–CDPKci was made by linking the cauliflower mosaic virus 35Spromoter to the coding region of a calcium-independent mutantCDPK gene of Arabidopsis thaliana (Harper et al., 1994; J.F. Harper,unpublished observations) and the 39 untranslated region of the NOSgene. (6) 35S–PKABA1 was made by substituting the Ubi-1 promoterin UBI–PKABA1 for the 35S promoter used in the 35S–CDPK con-struct.
Isolation of a GAMyb Genomic Clone
A genomic library of barley (cv Igri) in the vector Lambda FIX II wasobtained from Stratagene (La Jolla, CA). A KpnI fragment of 1.6 kbcontaining the entire coding region of GAMyb (Gubler et al., 1995),which was kindly provided by Drs. Frank Gubler and John Jacobsen(Commonwealth Scientific and Industrial Research Organization[CSIRO], Australia), was used as a probe to screen 106 colony-forming units. One clone of z16.5 kb containing the entire codingregion and at least 4 kb upstream and downstream of the openreading frame was isolated. A 3.5-kb BamHI fragment containingthe region 21031 to 12177 of the GAMyb gene was subcloned intothe BamHI site of pBluescript KS1 (Stratagene), generating theplasmid pRZ118. The GAMyb genomic sequence was submitted toGenBank with accession number AY008692. The clone pRZ118 wassubsequently cut with SapI, blunt ended with the Klenow fragmentof the DNA polymerase I, and then recut with KpnI. The fragmentKpnI/SapI was subcloned into pJR268b (kindly provided by Dr.John C. Rogers, Washington State University, Pullman) previouslycut with BamHI, blunt ended with the Klenow fragment, and thenrecut with KpnI. This new plasmid (pRZ126) contains 1031 bp of the
promoter, the first intron, and the sequence encoding the firstseven amino acids of the GAMyb protein fused in frame to the cod-ing region of GUS and the 39 untranslated region of the Amy6-4gene from barley.
The 2327 deletion construct (pRZ135) was generated by cuttingpRZ126 with XhoI and BglII, filling in with the Klenow fragment, andreligating the plasmid. The 2168 deletion construct (pRZ141) wasobtained by cutting pRZ126 with XhoI and partially with MfeI, treatingwith the Klenow fragment, and religating the plasmid. All constructswere created and maintained in the Escherichia coli strain XL1-BlueMRF9 (Stratagene).
Particle Bombardment and Transient Expression Assays
Detailed descriptions of the transient expression procedure with thebarley aleurone system and the particle bombardment techniquehave been published (Lanahan et al., 1992; Shen et al., 1993). Briefly,the mixture (in equal molar ratio) of a reporter construct and a maizeUBI–luciferase internal control construct, pAHC18 (Christensen andQuail, 1996), was bombarded into barley embryoless half-seed (fourreplicates per test construct).
To test the effect of regulatory molecules such as PKABA1 andGAMyb, different amounts of effector constructs were included, asindicated in the figures. After incubation in the presence or absenceof hormones for different times, the bombarded half-seed in sets offour were homogenized with the aleurone still attached to thestarchy endosperm in 800 mL of grinding buffer (Shen et al., 1993).After centrifugation at 12,000g for 10 min at 68C, 100 mL of the su-pernatant was assayed for luciferase activity using a luminometer(Moonlight 2010; Analytic Luminescence Laboratory, San Diego,CA). For GUS assays, 50 mL of the supernatant was diluted with200 mL of GUS assay buffer (Shen et al., 1993) and incubated at378C for 20 hr. Fifty microliters of the reaction mixture was diluted in2 mL of 0.2 M Na2CO3, and the resulting fluorescence was mea-sured in a Sequoia-Turner model 450 fluorometer (Unipath, Moun-tain View, CA) that was adjusted to read 1000 units for 1 mM4-methylumbelliferone. The GUS activities of all samples were nor-malized against those of the luciferase internal control. The normal-ized GUS activity represents the total number of fluorescence unitsfrom an aliquot of extract that contained 2 3 106 relative light unitsof luciferase activity. The data reported in each figure represent theresults of four independent transformations in a typical experiment.Each experiment was repeated at least two times with similar re-sults. Statistical analysis was performed with the program Stat-graphics Plus, v.2.1.
ACKNOWLEDGMENTS
We thank Drs. Frank Gubler and John Jacobsen (CSIRO, CanberraCity, Australia) for the barley GAMyb cDNA clone, Dr. Karen Cone(University of Missouri, Columbia) for the maize C1 Myb cDNA, Dr.John Rogers (Washington State University, Pullman) for the a-amy-lase clones, Dr. Peter Chandler (CSIRO) for the slender barley seed,Dr. Jeff Harper (Scripps Institute, La Jolla, CA) for the CDPK cDNAs,Dr. Lynn Hollapa (U.S. Department of Agriculture/Washington StateUniversity) for the PKABA1 null mutant, and Dr. Daisuke Yamauchi(Washington University, St. Louis, MO) for help in the initial screeningfor barley GAMyb genomic clones. This work was supported by

678 The Plant Cell
grants from the U.S. Department of Agriculture (No. NRICGP97-35100-4228) and the National Science Foundation (No. IBN-9983126).
Received September 14, 2000; accepted January 8, 2001.
REFERENCES
Anderberg, R.J., and Walker-Simmons, M.K. (1992). Isolation of awheat cDNA clone for an abscisic acid-inducible transcript withhomology to protein kinase. Proc. Natl. Acad. Sci. USA 89,10183–10187.
Bethke, P.C., Schuurink, R., and Jones, R.L. (1997). Hormonal sig-nalling in cereal aleurone. J. Exp. Bot. 48, 1337–1356.
Bowler, C., and Chua, N.H. (1994). Emerging themes of plant signaltransduction. Plant Cell 6, 1529–1541.
Brown, E.G., Newton, R.P., Evans, D.E., Walton, T.J., Younis,L.M., and Vaughan, J.M. (1989). Influence of light on cyclicnucleotide metabolism in plants: Effects of dibutyl cyclic nucle-otides on chloroplast components. Phytochemistry 28, 2559–2563.
Butt, E., Geiger, J., Jarchau, T., Lohmann, S.M., and Walter, U.(1993). The cGMP-dependent protein kinase: Gene, protein, andfunction. Neurol. Res. 18, 27–42.
Cercós, M., Gómez-Cadenas, A., and Ho, T.-H.D. (1999). Hor-monal regulation of a cysteine proteinase gene, EPB1, in barleyaleurone layers: Cis- and trans-acting elements involved in thecoordinated gene expression regulated by gibberellins and absci-sic acid. Plant J. 19, 107–118.
Chandler, P.M. (1988). Hormonal regulation of gene expression inthe “slender” mutant of barley (Hordeum vulgare L.). Planta 175,115–120.
Chrispeels, M.J., and Varner, J.E. (1966). Inhibition of gibberellicacid induced formation of a-amylase by abscisin II. Nature 212,1066–1067.
Christensen, A.H., and Quail, P.H. (1996). Ubiquitin promoter-based vectors for high-level expression of selectable and/orscreenable marker genes in monocotyledonous plants. Trans-genic Res. 5, 213–218.
Croker, S.J., Hedden, P., Lenton, J.R., and Stoddart, J.L. (1990).Comparison of gibberellins in normal and slender seedlings. PlantPhysiol. 94, 194–200.
Foster, C.A. (1977). Slender: An accelerated extension growthmutant of barley. Barley Genet. Newslett. 7, 24–27.
Gilroy, S. (1996). Signal transduction in barley aleurone protoplastsis calcium dependent and independent. Plant Cell 8, 2193–2209.
Gilroy, S., and Jones, R.L. (1994). Perception of gibberellin andabscisic acid at the external face of the plasma membrane of bar-ley (Hordeum vulgare L.) aleurone protoplasts. Plant Physiol. 104,1185–1192.
Gómez-Cadenas, A., Verhey, S.D., Holappa, L.D., Shen, Q., Ho,T.-H.D., and Walker-Simmons, M.K. (1999). An abscisic acid-induced protein kinase, PKABA1, mediates abscisic acid-sup-pressed gene expression in barley aleurone layers. Proc. Natl.Acad. Sci. USA 96, 1767–1772.
Grill, E., and Himmelbach, A. (1998). ABA signal transduction. Curr.Opin. Plant Biol. 1, 412–418.
Gubler, F., and Jacobsen, J.V. (1992). Gibberellin-responsive ele-ments in the promoter of a barley high-pI a-amylase gene. PlantCell 4, 1435–1441.
Gubler, F., Kalla, R., Roberts, J.K., and Jacobsen, J.V. (1995).Gibberellin-regulated expression of a myb gene in barley aleuronecells: Evidence for Myb transactivation of a high-pI a-amylasegene promoter. Plant Cell 7, 1879–1891.
Gubler, F., Raventos, D., Keys, M., Watts, R., Mundy, J., andJacobsen, J.V. (1999). Target genes and regulatory domains ofthe GAMYB transcriptional activator in cereal aleurone. Plant J.17, 1–9.
Harper, J.F., Huang, J.-F., and Lloyd, S.J. (1994). Genetic identifi-cation of an autoinhibitor in CDPK, a protein kinase with a cal-modulin-like domain. Biochemistry 33, 7267–7277.
Holappa, L.D., and Walker-Simmons, M.K. (1995). The wheatabscisic acid-responsive protein kinase mRNA, PKABA1, is up-regulated by dehydration, cold temperature, and osmotic stress.Plant Physiol. 108, 1203–1210.
Hooley, R., Beale, M.H., and Smith, S.J. (1991). Gibberellin per-ception at the plasma membrane of Avena fatua aleurone proto-plasts. Planta 183, 274–280.
Jacobsen, J.V. (1973). Interactions between gibberellic acid, ethyl-ene, and abscisic acid in control of amylase synthesis in barley.Plant Physiol. 51, 198–202.
Jacobsen, S.E., Binkowski, K.A., and Olszewski, N.E. (1996).SPINDLY, a tetratricopeptide repeat protein involved in gibberellinsignal transduction in Arabidopsis. Proc. Natl. Acad. Sci. USA 93,9292–9296.
Jones, H.D., Smith, S.J., Desikan, R., Plakidou-Dymock, S.,Lovegrove, A., and Hooley, R. (1998). Heterotrimeric G proteinsare implicated in gibberellin induction of a-amylase gene expres-sion in wild oat aleurone. Plant Cell 10, 245–253.
Kuo, A., Cappelluti, S., Cervantes-Cervantes, M., Rodriguez, M.,and Bush, D.S. (1996). Okadaic acid, a protein phosphataseinhibitor, blocks calcium changes, gene expression, and celldeath induced by gibberellin in wheat aleurone cells. Plant Cell 8,259–269.
Lanahan, M.B., and Ho, T.-H.D. (1988). Slender barley: A constitu-tive gibberellin-response mutant. Planta 175, 107–114.
Lanahan, M.B., Ho, T.-H.D., Rogers, S.W., and Rogers, J.C.(1992). A gibberellin response complex in cereal a-amylase genepromoters. Plant Cell 4, 203–211.
Leung, J., and Giraudat, J. (1998). Abscisic acid signal transduc-tion. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 199–222.
Li, J., Wang, X.Q., Watson, M.B., and Assmann, S.M. (2000). Reg-ulation of abscisic acid–induced stomatal closure and anion chan-nels by guard cell AAPK kinase. Science 287, 300–303.
Lovegrove, A., and Hooley, R. (2000). Gibberellin and abscisic acidsignalling in aleurone. Trends Plant Sci. 5, 102–110.
Paz-Ares, J., Ghosal, D., Wienand, U., Peterson, P., and Saedler,H. (1987). The regulatory locus c1 of Zea mays encodes a proteinwith homology to myb proto-oncogene products and with struc-

GA/ABA Antagonism in Barley Aleurone 679
tural similarities to transcriptional activators. EMBO J. 6, 3553–3558.
Peng, J., et al. (1999). ‘Green revolution’ genes encode mutant gib-berellin response modulators. Nature 400, 256–261.
Penson, S.P., Schuurink, R.C., Fath, A., Gubler, F., Jacobsen,J.V., and Jones, R.L. (1996). cGMP is required for gibberellicacid-induced gene expression in barley aleurone. Plant Cell 8,2325–2333.
Raventos, D., Skriver, K., Schelein, M., Karnahl, K., Rogers,S.W., Rogers, J.C., and Mundy, J. (1998). HRT, a novel zinc fin-ger transcriptional repressor from barley. J. Biol. Chem. 273,23313–23320.
Ritchie, S., and Gilroy, S. (1998a). Abscisic acid signal transductionin the barley aleurone is mediated by phospholipase D activity.Proc. Natl. Acad. Sci. USA 95, 2697–2702.
Ritchie, S., and Gilroy, S. (1998b). Gibberellins: Regulating genesand germination. New Phytol. 140, 363–383.
Robertson, M., Swain, S.M., Chandler, P.M., and Olszewski, N.E.(1998). Identification of a negative regulator of gibberellin action,HvSPY, in barley. Plant Cell 10, 995–1007.
Rogers, J.C., and Rogers, S.W. (1992). Definition and functionalimplications of gibberellin and abscisic acid cis-acting hormoneresponse complexes. Plant Cell 4, 1443–1451.
Schuurink, R.C., Chan, P.V., and Jones, R.L. (1996). Modulation ofcalmodulin mRNA and protein levels in barley aleurone. PlantPhysiol. 111, 371–380.
Sheen, J. (1996). Ca21-dependent protein kinases and stress signaltransduction in plants. Science 274, 1900–1902.
Shen, Q. (1996). Involvement of PP2C in the signal transduction ofABA action in barley aleurone cells. In Phosphorylation-Dephos-phorylation of Plant Proteins: Current Topics in Plant Biochemis-try, Physiology and Molecular Biology, D.D. Randall, ed (Columbia,MO: University of Missouri), pp. 42–43.
Shen, Q., Uknes, S.J., and Ho, T.-H.D. (1993). Hormone responsecomplex of a novel abscisic acid and cycloheximide induciblebarley gene. J. Biol. Chem. 268, 23652–23660.
Shen, Q., Zhang, P., and Ho, T.-H.D. (1996). Modular nature ofabscisic acid (ABA) response complexes: Composite promoterunits that are necessary and sufficient for ABA induction of geneexpression in barley. Plant Cell 8, 1107–1119.
Skriver, K., Olsen, F.L., Rogers, J.C., and Mundy, J. (1991). Cis-acting DNA elements responsive to gibberellin and its antagonistabscisic acid. Proc. Natl. Acad. Sci. USA 88, 7266–7270.
Sun, T.-P. (2000). Gibberellin signal transduction. Curr. Opin. PlantBiol. 3, 374–380.
Sutliff, T.D., Lanahan, M.B., and Ho, T.-H.D. (1993). Gibberellintreatment stimulates nuclear factor binding to the gibberellinresponse complex in a barley a-amylase promoter. Plant Cell 5,1681–1692.
Wang, X., and Robinson, P.J. (1997). Cyclic GMP-dependent pro-tein kinase and cellular signalling in the nervous system. J. Neuro-chem. 68, 443–456.

DOI 10.1105/tpc.13.3.667 2001;13;667-679Plant Cell
Aurelio Gómez-Cadenas, Rodolfo Zentella, Mary Kay Walker-Simmons and Tuan-Hua David HoKinase PKABA1 in Relation to Gibberellin Signaling Molecules
Gibberellin/Abscisic Acid Antagonism in Barley Aleurone Cells: Site of Action of the Protein
This information is current as of January 8, 2016
References http://www.plantcell.org/content/13/3/667.full.html#ref-list-1
This article cites 44 articles, 26 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists




















