
Ramadhan et alimentation, le cas des femmes diabétiques d’Oran,
Upload
univ-lilleCategory
view
3download
0
Les amas coquilliers protobyzantins de Limenas : exploitation des ressources piscicoles, alimentation carnée.Déclin de la cité de Thasos (Grèce)Tarek Oueslati(1)
(1) Halma-IPEL UMR 8164 du CNRS, Domaine universitaire du Pont-de-Bois, BP 60149, F-59653 Villeneuve-d’Ascq <[email protected]>
Résumé. La fouille archéologique de la Domus 5 Protobyzantine de la cité de Limenas a révélé l’existence de deux niveaux de destruction entre lesquels a été découverte une épaisse couche de terre noire constituée d’amas coquilliers datant de la seconde moitié du vie siècle et du début du viie siècle. Ce contexte coïncide avec la destruction de la plupart des habitats et des églises environnants. La problématique archéologique s’ar-ticule ainsi autour du statut et du mode de subsistance des occupants de la maison en ruine. Les résultats obtenus suggèrent une alimentation inhabituelle à l’échelle locale et régionale indiquant une adaptation à la période de crise que connaît la cité.Mots-clés. Domus, protobyzantin, ichtyofaune, ressources cynégétiques, invasions, séismes.
Abstract. The excavation of the protobyzantine house of the city of Limenas has revealed two destruction levels in between which a dark layer of sediment containing shell middens was discovered. This horizon was dated to the second half of the 6th century beginning of the 7th century AD. The particularity of this context which corresponds to the general destruction of the houses and churches of the city has motivated the study of the animal finds in order to shed light on the status of the residents of these ruins and their subsistence strategies. The preliminary results show an unusual composition of the meat diet thus suggesting an adaptation of the subsistence strategy to the crisis through which the city is passing.Keywords. Domus, protobyzantine, ichthyofauna, wild mammal resources, invasions, earth quakes.
* * *
Description du contexte archéologique
Le site de ThANAr fait l’objet d’une fouille archéologique biannuelle alternée avec des campagnes post-fouilles depuis 2004. Cette opération est conduite par
1
ARCHÉologie Du poisson. 30 Ans D’ARCHÉo-iCHtyologie Au CnRsHommage aux travaux de Jean Desse et nathalie Desse-BersetXXViiie rencontres internationales d’archéologie et d’histoire d’AntibesSous la direction de P. Béarez, S. Grouard et B. ClavelÉditions APDCA, Antibes, 2008
2
t. ouesLati
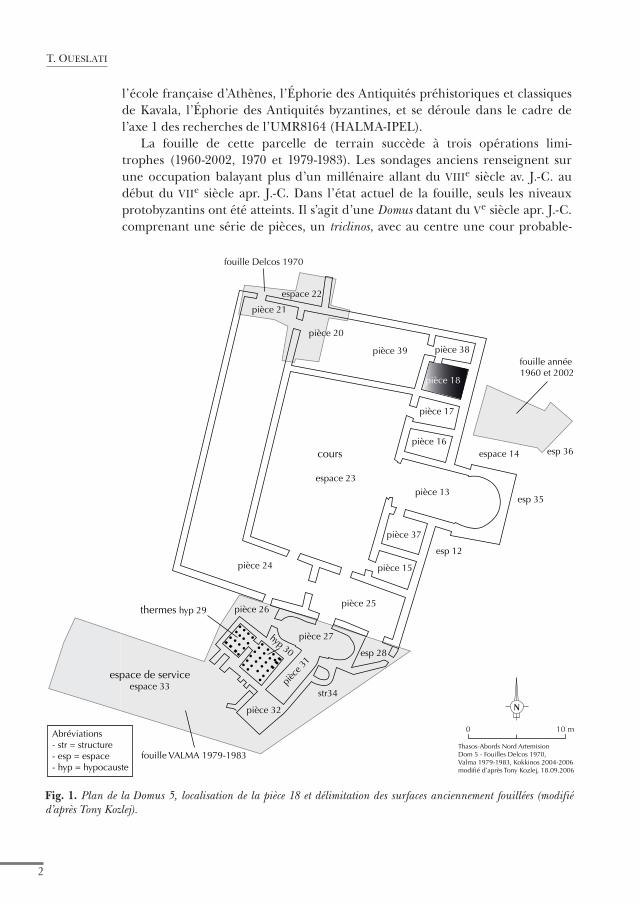
l’école française d’Athènes, l’Éphorie des Antiquités préhistoriques et classiques de Kavala, l’Éphorie des Antiquités byzantines, et se déroule dans le cadre de l’axe 1 des recherches de l’UMR8164 (HALMA-IPEL).
La fouille de cette parcelle de terrain succède à trois opérations limi-trophes (1960-2002, 1970 et 1979-1983). Les sondages anciens renseignent sur une occupation balayant plus d’un millénaire allant du viiie siècle av. J.-C. au début du viie siècle apr. J.-C. Dans l’état actuel de la fouille, seuls les niveaux protobyzantins ont été atteints. Il s’agit d’une Domus datant du ve siècle apr. J.-C. comprenant une série de pièces, un triclinos, avec au centre une cour probable-
10 m0
N
Thasos-Abords Nord ArtemisionDom 5 - Fouilles Delcos 1970, Valma 1979-1983, Kokkinos 2004-2006modifié d’après Tony Kozlej, 18.09.2006
Abréviations- str = structure- esp = espace- hyp = hypocauste
fouille Delcos 1970
pièce 20
espace 22
pièce 21
pièce 18
pièce 38pièce 39fouille année 1960 et 2002
esp 12
esp 35
espace 14 esp 36
pièce 15
pièce 13
pièce 37
espace 23
pièce 16
pièce 17
cours
pièce 25
pièce 24
esp 28
pièce 27
pièce 26thermes hyp 29
hyp 30
pièc
e 31
str34
pièce 32
fouille VALMA 1979-1983
espace de serviceespace 33
Fig. 1. plan de la Domus 5, localisation de la pièce 18 et délimitation des surfaces anciennement fouillées (modifié d’après tony Kozlej).

Les amas coquiLLiers protobyzantins de Limenas
3
ment à péristyle, un complexe thermal et une aire de service (fig. 1). Comparée aux autres édifices civils contemporains de Limenas, cette grande et luxueuse demeure constitue un cas à part. Les opérations de fouille de 2004 ont révélé une première destruction de l’habitat vers 560-570. Une réoccupation précaire des lieux a été à l’origine du dépôt d’une terre noire (US 04.27) contenant des amas coquilliers. Ce niveau est surmonté à son tour par un second niveau de démolition correspondant à la destruction définitive de la demeure peu après 620. Dans l’état actuel de la recherche, la fouille n’a pas livré d’indices sur les facteurs à l’origine de ces événements. Simultanément à la constitution de cette terre noire, on assiste au démontage des pavements en marbre des murs et des sols, et une réoccupation bien moins dense et luxueuse de l’habitat. Le sondage 2 effectué en 2004 couvre partiellement la pièce 18. La terre noire étudiée ici vient combler à la fin du vie siècle et au début du viie siècle cette pièce, ce qui signifie son abandon à cette époque. Dans ce contexte archéologique très parti-culier, les principales problématiques posées s’articulent autour de la nature de l’occupation entre les deux phases de destruction et la définition du statut des occupants du site. Dans cette optique, l’approche archéozoologique exploitera les apports des restes animaux, mollusques et vertébrés. Cette faune a été collectée à vue ainsi que par tamisage à l’eau et par tamisage à sec. Le tamisage à l’eau a concerné la première moitié de la couche tandis que l’autre moitié a été fouillée puis tamisée à sec. Les restes de mammifères ont été étudiés dans le musée de limenas tandis que les restes de poissons ont été déterminés au laboratoire de l’Université de Lille 3 et à l’Institut royal des Sciences naturelles de Belgique.
Description de l’assemblage faunique
La faune issue du sondage 2 (US 04.27, opération 2004) est constituée de 1 492 restes de vertébrés et 7 156 restes d’huîtres (ostrea edulis). Les espèces de mollusques faiblement représentées ne sont pas prises en compte dans cette approche en raison d’une collecte différentielle des grands mollusques au moment de la fouille. Par ailleurs le tamisage des déblais de la fouille et le tami-sage à l’eau ont livré uniquement de très petits spécimens de mollusques dont la contribution à l’alimentation est à exclure. Les mammifères et les poissons constituent respectivement 56 % et 42 % des vertébrés (fig. 2). En revanche, les restes d’oiseaux sont faiblement représentés.
Les restes de poissons
La diagnose spécifique a permis d’identifier 22 familles dont deux dulça-quicoles (fig. 3). Les sparidés dominent l’assemblage suivis par les scorpénidés, les mugilidés et les cyprinidés. Ces taxons constituent près des trois quarts des poissons déterminés. Au sein des sparidés, la détermination spécifique a permis de souligner la présence du marbré (lithognathus mormyrus NMI = 7), des sars (Diplodus sp. NMI = 4), de la daurade royale (sparus aurata NMI = 2), du denté (Dentex dentex NMI = 1) et d’un reste de pageot (pagellus sp.). Les scorpénidés

4
t. ouesLati
n’ont pas fait l’objet d’une détermination spécifique car les 5 taxons inventoriés dans le bassin méditerranéen occidental ne sont pas réunis dans les collections de référence, et de surcroît la détermination des spécimens frais demeure diffi-cile (Cadenat, 1943). La détermination des mugilidés est également restreinte au rang de la famille en raison de manque de spécimens de référence. Par ailleurs, les cyprinidés ont été attribués pour l’essentiel à la carpe (NR = 27). Toutefois il faut souligner que l’importante diversité spécifique des cyprinidés du nord de la Grèce, la présence de taxons endémiques et la faible variabilité morpho logique au sein de cette famille imposent quelque prudence dans cete détermination. Cette sujétion au manque de spécimens de référence de ces régions a réduit la détermination à un reste de brème (Abramis brama) et un reste de carassin (Carassius sp.). Le cortège de taxons d’eau douce comprend également deux restes de silure (silurus glanis).
La répartition des pièces anatomiques en fonction du nombre de restes pour les taxons les mieux représentés (sparidés, scorpénidés, mugilidés, cypri-nidés, serranidés, mullidés, clupéidés et chondrichthyens) souligne le fort déficit en éléments des ceintures pelvienne et scapulaire (fig. 4). En ce qui concerne les éléments de rachis et de crâne, deux tendances s’observent. Tout d’abord les taxons pourvus d’os de la tête robustes voient ces éléments très bien repré-sentés comme chez les sparidés, les serranidés, les scorpénidés et les cyprinidés. En revanche, pour les mullidés et les clupéidés dont les éléments du crâne sont
Noms vernaculaires Noms latins NR % NR PdR NMI MOW
bœufporcmoutonchèvremouton ou chèvre
Bos primigenius f. taurusSus scrofa f. domesticaOvis orientalis f. ariesCapra aegagrus f. hircus
423071832374
5,439,72,34,148,1
7811524,81572901839
29
8
250380
127,5
Total triade 773 100 4591,8 757,5
chevreuilcerfdaimCervidaesanglierlagomorphe
Capreolus capreolus Cervus elaphus Dama dama Cervus elaphus ou Dama damaSus scrofa
23916321
3,24,814,325,450,81,6
24451432355294
1
1221
Total gibier 63 100 980
coq cygneoieoiseaux indéterminésrat noirchiroptèrehuître
Gallus gallus f. domesticaCygnus cf cygnusAnser cf anser
Rattus rattus
Ostrea edulis
301111117156
6113113628
Abréviations NR : nombre de restes, PdR : poids de restes en gramme, MOW : poids de viande et abats en kg des espèces de la triade.
Fig. 2. effectifs des mammifères, oiseaux et huîtres mis au jour dans l’us 04-27.

Les amas coquiLLiers protobyzantins de Limenas
5
fragiles seuls des éléments de vertèbres sont attestés. Il en ressort que la conser-vation différentielle est probablement le principal facteur de modification des fréquences des parties du squelette. Cela permet de supposer que la préparation et la consommation se faisaient à proximité de la zone de rejet.
L’estimation de la taille moyenne des poissons dont est issue l’ichtyofaune repose sur la comparaison directe des spécimens archéologiques avec des séries de tailles de spécimens actuels. Elle renseigne sur la présence de spécimens dont la longueur standard dépasse 30 cm (fig. 5). Il s’agit des mugilidés (SL = 33 cm,
Taxon NR NMI
Squatina cf. squatina 23 1
Raja clavata Rajidae indéterminés
63
Total Rajidae 9 1
Cyprinus carpio Abramis brama Carassius sp. cyprinidés indéterminés
3111
22
311
Total Cyprinidae 55 5
Silurus glanis 2 1
Lithognathus mormyrus Diplodus sp.Sparus aurataDentex dentexPagellus sp.Sparidae indéterminés
86211
188
74211
Total Sparidae 206 15
ScorpaenidaeMugilidaeSerranus sp.ClupeidaeMullidaeCarangidaePleuronectiformeCentracanthidaeScomber scombrus SciaenidaeMoronidaeConger congerGobiidaeLabridaeSoleidaeTrachinidaeZeus faber
1146927232116138875444111
133611111111211111
Total poissons 621 60
Fig. 3. effectifs en nombre de restes (nR) des familles de poissons identifiées.

6
t. ouesLati
n = 5), de la carpe (SL = 32 cm, n = 16) et des moronidés (SL = 31 cm, n = 2). Les spécimens de plus petite taille sont représentés par le genre serranus avec une longueur standard moyenne de 25,6 cm (n = 5), les sparidés (SL = 20,7 cm, n = 27), les scorpénidés (SL = 16,6 cm, n = 21) et les labridés (SL = 16 cm, n = 2). mesures étant les tailles standards moyennes pour ces familles.
La reconstitution du poids des poissons a reposé sur la comparaison avec des spécimens actuels. Elle permet d’estimer approximativement le poids total de poissons ostéichtyens consommés à 43 kg. La difficulté que posent l’estima-tion du nombre minimum d’individus et la reconstitution des tailles de squati-nidés et de rajidés ne permet pas d’avoir une estimation précise de l’apport de ces poissons à l’alimentation carnée. Néanmoins, la taille du centre vertébral des individus du genre squatina correspond fort probablement à des individus d’au moins 120 cm, soit un poids de plusieurs dizaines de kilogrammes.
L’amoncellement de débris ostréaires
L’amoncellement de tests d’huître caractérise ce niveau archéologique. En effet, les valves de cet ostréidé constituent, à la fouille, des amas coquilliers parti-culièrement denses. Au total 7 156 restes ont été décomptés. La latéralisation des valves permet d’estimer le nombre minimum d’individus à 3 628 spécimens. Afin d’évaluer l’apport de ce mollusque dans l’alimentation carnée, nous avons eu recours à la détermination du calibre moyen des spécimens étudiés d’après
Chondrichtyens
Clupeidae
Cyprinidae
Mugillidae
Mullidae
Scorpaeanidae
Serranidae
Sparidae
Rachis Ceinture pelvienne Ceinture thoracique Tête
0 20 40 60 80 100
Fig. 4. Répartition des parties du squelette regroupées en tête, rachis et ceintures scapulaire et pelvienne.

Les amas coquiLLiers protobyzantins de Limenas
7
les mesures moyennes de la longueur et de la largeur des valves. Ces données métriques ont permis de déterminer le poids total de l’animal (Robert, Pichot et al., 1990) qui associe à notre assemblage un calibre moyen compris entre 2 et 3 (Le Gall, 1948). En tenant compte du rendement pondéral et calorique de la chair d’huître plate de ce calibre (Le Gall, 1948), il devient possible d’estimer l’apport des 3 628 individus (NMI) en termes de chair d’huître à 43 536 g soit 36 004 calories réparties en 57 % de protéines, 35 % de glucides et 8 % de lipides. Cela renseigne sur le rôle important joué par ce mollusque dans l’alimentation carné.
Les mammifères et les oiseaux
La triade domestique bœuf, porc et caprinés représente près de 90 % des mammifères terrestres. Les caprinés constituent près de la moitié des restes de cette triade et ils sont suivis de près par le porc représenté à la hauteur de 40 % au sein de ce groupe. L’apport de la triade domestique en viande et abats a été estimé en tenant compte de la stature des caprinés et des suinés ainsi que des âges auxquels ces animaux ont été abattus (Vigne, 1991). Pour le bœuf, l’absence d’os longs complets permettant d’estimer la hauteur au garrot nous a contraints à estimer cette valeur par la comparaison des mesures des phalanges avec d’autres corpus antiques (Oueslati, 2006). Ainsi l’apport en viande et abats de la triade est évalué à près de 750 kg avec en première place le porc (380 kg) puis le bœuf (250 kg) et enfin les caprinés (127 kg). Cette recons titution de l’apport carné de chaque taxon illustre la place privilégiée du porc et du bœuf.
Par ailleurs, le gibier à poils rassemble 8 % des restes de mammifères et comprend le daim (NR = 9), le cerf (NR = 3), le chevreuil (NR = 2), le sanglier
Ouslati 05
Carpe
Moronidés
Mugillidés
Scorpénidés
Serranidés
Sparidés
Labridés
0 5 10 15 20 25 30 35
Fig. 5. taille moyenne des différents taxons d’après la longueur standard (sl).

8
t. ouesLati
(NR = 32) et un reste de lagomorphe. Par ailleurs 16 restes de cervidés n’ont pas pu être déterminés au rang de l’espèce (cerf ou daim). Les animaux chassés comprennent également des restes d’oiseaux, dont le cygne attesté par un reste portant des traces de découpe. Par ailleurs, les oiseaux de basse-cour sont représentés par le coq (NR = 30). Enfin un reste de chiroptère et un autre de rat ont été identifiés. La présence des rongeurs est également illustrée par les nombreuses traces de rongement relevées sur les os.
Nature des milieux exploités
Les ressources dulçaquicoles
La présence de la carpe, de la brème et du silure, espèces fréquentant les eaux calmes à fond vaseux, témoigne de la pêche en eau douce. Le paysage actuel de l’île est caractérisé par l’absence de cours d’eau permanents, ce qui ne permet pas d’envisager, en un premier temps, une exploitation locale. Dans ce cas on pourrait suggérer un commerce de ces espèces à partir du continent où le fleuve Nestos, la lagune naturelle de Vistonida et les zones marécageuses aux environs de Kiramouti constituent autant de milieux propices à ce type de pêche. Toutefois, les aménagements récents sur l’île de Thasos avec le captage systématique des sources et l’assainissement des zones marécageuses laissent présager un réseau hydrographique antique différent. Dans cette optique, les études géomorphologiques menées sur l’île nous éclairent sur l’existence, jusqu’à l’époque moderne, d’étangs littoraux étendus entre Skala sotiros et Prinos par exemple (Lespez, sous presse), ainsi que d’écosystèmes favorables au niveau des parties en aval des cours d’eau les plus importants telles que les rivières de Mariès, Liménaria, Kastri (Lespez, comm. pers.). Ainsi, la restitution des carac-téristiques physiques des bassins versants renseigne sur l’existence d’un environ-nement compatible avec le développement de petites populations dulçaquicoles. Il faut rajouter à cela toutes les dépressions gorgées d’eau qui auraient pu être colonisées par des larves de poissons transportées, notamment par les oiseaux comme c’est le cas aujourd’hui sur l’île (Manuella et Tony Kozlej, comm. pers.). À l’échelle de l’Égée septentrionale, les études des ichtyofaunes datées des périodes préhistoriques et protohistoriques attestent une exploitation impor-tante de poissons d’eau douce concentrée sur les sites charnières et continentaux (Theodoropoulou, 2007). Dans ces assemblages, les cyprinidés représentent plus de 30 % des restes de poisson. Au sein de cette famille, c’est à proximité des côtes que la carpe est la mieux représentée. Si cette pêche en eau douce aux époques préhistoriques se limitait aux occupations continentales et à faible distance des côtes, la donne semble modifiée aux périodes plus récentes dans la mesure où l’occupation hellénistique de Krania (Égée septentrionale) et la villa romaine de Mytilène (contexte insulaire, Égée septentrionale) attestent l’exploitation de pois-sons d’eau douce (Mylona, 2007). Ainsi l’assemblage de ThANAr, bien que plus tardif que les deux sites côtiers précédents, conforte la place des poissons d’eau douce dans l’alimentation carnée des sites côtiers. Par ailleurs la prédominance

Les amas coquiLLiers protobyzantins de Limenas
9
de la carpe sur ces sites côtiers suit la tendance observée aux époques plus anciennes.
Les ressources marines
Les taxons dominants sont les sparidés, les scorpénidés et les mugilidés. Les eaux côtières sableuses ou rocheuses ont probablement constitué l’essen-tiel des zones de pêche des petits sparidés dont la longueur standard moyenne est de 20 cm. Ces jeunes individus sont souvent regroupés en bancs et occupent des eaux plus superficielles que les grands individus solitaires. Une capture au filet maillant est fort probable. Par ailleurs, les zones rocheuses, caractéris-tiques de Thasos, ont probablement constitué des zones favorables à la capture des scorpénidés au filet ou à la ligne et hameçon. Enfin les mugilidés, euryha-lins, occupent essentiellement les eaux côtières et peuvent pénétrer dans les zones saumâtres et dans les eaux douces. Les mulets vivent en banc et occupent de faibles tranches d’eau. Leur capture se fait aisément au filet maillant ou à l’épervier. L’utilisation de nasses est possible pour l’ensemble de ces taxons. Cette ichtyofaune marine dominée par les espèces littorales de tailles petite et moyenne contraste avec les assemblages du Néolithique moyen de l'île qui ont livré de grands spécimens de Scombridae, de Sparidae et de Sciaenidae (Theodoropoulo, 2007). A l'échelle régionale, les sites des périodes histo riques ont livré de grands poissons notamment des mérous (Mylona, 2007) qui ne sont pas attestés dans nos ensembles. Ces différences de taille des poissons et l'absence des pélagiques de grande taille confèrent aux techniques de pêche à l'origine de notre assemblage un caractère peu organisé et probablement limi-tées à des captures côtières voir une pêche du bord.
En ce qui concerne les mollusques, la contribution importante des huîtres à l’alimentation carnée illustre l’exploitation de l’étage infralittoral. Avant l’intensification des prélèvements des bancs à l’époque moderne, l’huître plate formait un cordon ininterrompu sur toute la côte occidentale d’Europe ainsi qu’en Méditerranée. Cette ressource a été exploitée par les occupants du site par dragage ou par collecte sur le fond. Ces accumulations d’amas ostréaires sont considérées par les archéologues comme étant typiques de cette occupa-tion protobyzantine puisqu’aucun antécédent n’a été enregistré auparavant. De même, d’après Theodoropoulou (2007), la consommation de l’huître aux époques antérieures (âge du Bronze et Néolithique) occupait une place margi-nale sur l’île ainsi que sur la plupart des sites côtiers de l’Égée septentrional.
Les ressources terrestres
Avec plus de 92 % des restes de mammifères, les animaux domestiques occupent une place dominante dans l’alimentation carnée. La prédominance des restes de caprinés et de suinés illustre très probablement les contraintes imposées par la topographie de l’île sur l’élevage, comme c’est le cas aujourd’hui avec un élevage spécialisé de moutons. Par ailleurs l’exploitation du milieu forestier est

10
t. ouesLati
illustrée par une proportion relativement importante de cervidés et de sangliers si l’on tient en compte la datation haute de l’occupation étudiée. Enfin la présence d’un reste de cygne, probablement sauvage, illustre une capture hiver-nale, saison à laquelle le cygne fréquente les zones littorales et marécageuses de ces régions.
Discussion
L’alimentation carnée au sein du dépotoir étudié révèle que la triade domes-tique constitue l’apport principal en viande et abats. Toutefois un complément non négligeable est apporté sous forme de poissons et d’huîtres dont les parts respectives sont sensiblement égales. Par ailleurs, l’étude de l’ichtyofaune révèle un territoire de pêche côtier à caractère peu sélectif et opportuniste exploi-tant les ressources disponibles à proximité du rivage. Il ne faut pas perdre de vue que ces restes de poissons consommés peuvent constituer une partie sélec-tionnée du produit de la pêche original. En effet des facteurs socio-économiques peuvent régir l’accès à des spécimens d’espèces et de tailles adaptées au statut du consommateur. De même, l’évaluation des stratégies de pêche doit prendre en considération la productivité des fonds marins limitrophes de l’île de Thasos. Cette précaution est d’autant plus de rigueur que la mer de ces contrées est peu productive selon les études actuelles d’évaluation des prises et des stocks de pois-sons. Toutefois il a été déjà montré que l’exploitation des données des pêcheries modernes est source d’erreurs et qu’on doit leur substituer par les données histo-riques, biogéographiques et paléoclimatique (Jacobsen 2005). De même, la forte diversité des fonds marins helléniques rend prévisible une forte variabilité dans la production. Si l’on focalise sur l’Égée septentrional, la synthèse bibliographique de Mylona (2007) révèle que cette mer est particulièrement riche, voire eutro-phique, ce qui confère un potentiel de pêche considérable. En ce qui concerne le statut socio-économique des habitants de cette Domus, au moment de l’accumu-lation des rejets de consommation, on assiste à un passage d’un statut exception-nellement privilégié à l’échelle de la cité de Limenas à un statut indéterminé, associé à une époque de crise et de destruction de l’habitat avec une réoccupa-tion partielle de la Domus. Simultanément, la cité se dépeuple après la première phase de destruction, ce qui laisse présager un changement dans les mécanismes de l’approvisionnement de la population. Ainsi l’accès sélectif à une partie du produit de la pêche est à écarter dans la mesure où les logiques associées à l’offre et à la demande sur le marché ne sont plus de rigueur. Par ailleurs, l’exploitation intensive et sans précédent des bancs d’huîtres ainsi que la pression cynégétique sur les ressources forestières nous éclairent sur un comportement atypique des occupants des ruines de cette Domus, suggérant des difficultés d’approvisionne-ment et un recours à la chasse et à la collecte qui constituent des compléments non négligeables à l’alimentation. Si l’on considère les hypothèses associées à la destruction de la cité avec d’un côté les séismes et de l’autre les invasions slaves, il est possible de voir dans les stratégies de subsistance une adaptation à une

Les amas coquiLLiers protobyzantins de Limenas
11
situation de crise. Dans cette optique, les premiers résultats obtenus à partir de l’étude des contextes plus anciens de notre site témoignent de la consommation de taxons de grandes tailles comme l’espadon et le maigre, ce qui renseigne sur les modifications de la composition de l’ichtyofaune après la destruction de la domus.
Conclusions
La faune de la seconde moitié du vie siècle apr. J.-C. et du début du viie siècle du site de ThANAr revêt un caractère inhabituel en comparaison avec les autres occupations antiques de Grèce (Mylona, 1999 ; 2007). La réoccupation sommaire de la Domus 5 après sa destruction et l’accumulation sans précédent à Thasos d’amas coquilliers nous éclairent sur un mode de subsistance original. Par ailleurs, une pêche opportuniste et non organisée est suggérée par la domi-nance des poissons littoraux et de taille moyenne. Enfin, la part inhabituelle-ment élevée de la chasse et l’exploitation des poissons d’eau douce suggèrent un mode de subsistance en rapport avec la période de crise que traverse la cité. Si ces hypothèses découlent en partie des comparaisons avec des contextes contem-porains ou plus anciens de l’Égée septentrional et méridional, la fouille à venir des niveaux antérieurs à la première destruction et l’abandon du site devraient nous éclairer avec plus de précision sur les circonstances et sur le statut des occu-pants de la Domus, ainsi que sur leur mode de subsistance. Pour conclure, souli-gnons que les données historiques et archéologiques témoignent de la fréquence de la réoccupation des sites d’habitat dans des contextes de catastrophes natu-relles, d’insécurité et de repli sur les sites fortifiés, de déclin démographique, etc. Dans cette optique, les hypothèses avancées par Patterson (2006) sur l’abandon de sites et leur réoccupation par des catégories socio-économiques différentes se matérialisent à l’échelle des fouilles archéologiques par un changement net de la qualité du vaisselier ou par l’affectation de l’habitat à des fonctions économiques (pressoirs, étables, etc.). Ainsi une approche interdisciplinaire est en cours pour affiner la nature des événements associés à la constitution du dépotoir étudié notamment par la caractérisation de la céramique qui y a été mise au jour.
Bibliographie
cadenat J., 1943.– Les scorpaenidae de l’Atlantique et de la Méditerranée, première note, le genre scorpaena, Revue des travaux de l’institut des pêches maritimes, 13, 1-4, p. 525-563.
Jacobsen A. L. L., 2005.– The reliability of Fishing Statistics as a source for Catches and Fish Stocks in Antiquity, in : T. Bekker-Nielsen (dir.), Ancient Fishing and Fish processing in the Black sea Region, proceedings of the interdisciplinary workshop on marine resources and trade in fish products in the Black Sa region in antiquity, avril 2003, Esbherg, Aarhus University Press, p. 97-104.
Le GaLL J. V., 1948.– Valeur nutritive et valeur thérapeutique de l’huître. office scientifique et technique des pêches maritimes, notes et rapports (nouvelle série), 2, 80 p.

12
t. ouesLati
Lespez L., sous presse.– Environnement thasien depuis le Néolithique, l’apport des recherches géoarchéologiques récentes, actes du symposium international à la mémoire de M. Sgourou, thasos, Métropole et colonies, septembre 2006.
myLona D., 2003.– Fishing in Late Antiquity : The case of Itanos, Crete, in : C. Gambe, P. Halstead, Y. Hamilakis, E. Kotjabopoulou (dir.), Zooarchaeology in greece : Recent advances, British School at Athens Studies, 9, p. 95-101.
myLona D., 2007.– Fish-eating in greece from the fifth century BC to the seventh century AD. A story of impoverished fishermen or luxurious fish banquets ?, thèse de doctorat de l’Université de Southampton, 287 p.
patterson J. R., 2006.– landscapes & Cities, Rural settlement and Civic transformation in early imperial italy, Oxford University Press, 348 p.
robert r., pichot y., comps m., 1990.– Essais de culture de l’huître plate ostrea edulis dans le bassin d’Arcachon, Résultats préliminaires, Rapports internes de la Direction des ressources vivantes de l’iFReMeR, 15 p.
theodoropouLou T., 2007.– L’exploitation des ressources aquatiques en Égée septen-trionale aux périodes pré- et protohistoriques, thèse de doctorat de l’Université Paris 1, 628 p. + 498 p. annexes.
viGne J.-D., 1991.– The meat and offal weight (MOW) method and the relative propor-tion of ovicaprines in some ancient meat diets of the north-western Mediterranean, Rivista di studi liguri, 57, 1-4, p. 21-47.
Copyright © 2022 FDOKUMEN




















