
Interlocking Transactions: Obstacles, Precursors or Instruments of Agrarian Capitalism?
Upload
independentCategory
view
0download
0
Ž .Journal of Immunological Methods 210 1997 65–77
Optimization of a peptide-based protocol employing IL-7 for invitro restimulation of human cytotoxic T lymphocyte precursors
Ajit Lalvani, Tao Dong, Graham Ogg, Ansar A. Pathan, Heidi Newell,Adrian V.S. Hill, Andrew J. McMichael, Sarah Rowland-Jones )
Molecular Immunology Group, Institute of Molecular Medicine, UniÕersity of Oxford, Oxford OX3 9DU, UK
Received 7 August 1997; revised 28 August 1997; accepted 21 October 1997
Abstract
Ž . Ž .A variety of different methods for the in vitro restimulation of human cytotoxic T lymphocyte CTL precursors CTLpare in use. Our aim was to enhance the detection of circulating human CTLp in peripheral blood. We have developed astandardized and highly efficient method for restimulating CTLp. Synthetic peptides were used to restimulate cognate CTLp
Ž .from peripheral blood mononuclear cells PBMC , and effector CTL capable of lysing peptide-pulsed and virus infectedtargets were generated. The effects of several parameters on CTL specific for influenza A, EBV and HIV-1 were evaluated,and the optimum peptide concentration for CTL generation was established. Supplementation of initial cultures with IL-7greatly enhanced peptide-specific lytic activity for all peptides tested and the dose–response relationship for IL-7 wasdelineated. A novel technique using peptide-MHC class I molecule tetramers to stain T cells bearing cognate T cell receptorspermitted enumeration of antigen-specific CD8q CTL during in vitro restimulation; IL-7 supplementation selectivelyexpanded the population of peptide-specific CD8q CTL. Importantly, this protocol, whilst enhancing the restimulation andlytic activity of secondary CTL, does not induce primary CTL in vitro. The improved efficiency with which CTL aregenerated in this system substantially enhances the sensitivity of CTL culture and the 51Cr release assay to detect low levelsof CTL activity. q 1997 Elsevier Science B.V.
Keywords: Human cytotoxic; T lymphocyte precursor; IL-7; CTL activity
1. Introduction
Conventional methods of generating CTL cultureshave relied on the use of the pathogen to stimulatethe growth of specific CTL from a memory popula-tion; this requires a careful balance between using asufficient amount of the organism to stimulate CTLgrowth but not so much that the cells are damaged.This is a particular problem for generating CTL
) Corresponding author. Tel: q44-1865-222316; fax: q44-1865-222502; e-mail: [email protected]
specific for HIV, when the use of live HIV tostimulate the CTL, either added exogenously or bystimulating the replication of endogenous HIV with
Ž .mitogens Nixon et al., 1988 , can damage the CD4qT cells which provide help for CTL growth. Somepathogens, for example the liver stage of P. falci-parum, cannot be readily cultured in vitro and syn-thetic peptides may then be the only means of gener-
Ž .ating CTL cultures Hill et al., 1992 . Recombinantvaccinia viruses, when available, are often suitablefor stimulating CTLp in vitro but are inferior to
Ž .peptides in certain systems Aidoo et al., 1997 .
0022-1759r97r$17.00 q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0022-1759 97 00177-4

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–7766
Furthermore, restimulation with synthetic peptidescan circumvent the immunodominance of one epi-tope, which sometimes occurs when using wholevirus, and thereby reveal a latent repertoire of CTL
Žspecific for sub-dominant epitopes Martinon et al.,.1990 .
In conditions where the circulating CTL precursorfrequency may be low, such as in P. falciparum
Ž .malaria Aidoo et al., 1995; Plebanski et al., 1997 ,Ž .tuberculosis Lalvani et al., 1997 individuals heavily
Žexposed to but not infected by HIV Rowland-Jones.et al., 1995 and when assessing CTL induction in
Žhuman vaccine trials Borysiewicz et al., 1996; Ock-.enhouse et al., manuscript in preparation , sensitive
methods of detecting CTL are required. In thesesituations optimal techniques for CTL culture areessential to generate sufficient numbers of effectorCTL to give demonstrable lytic activity in chromiumrelease assays.
We have developed a peptide-based protocol torestimulate CTLp in vitro with maximal efficiency.Restimulation conditions were evaluated for a widerange of peptides, including peptides with low, aswell as those with high affinity for their respectiveMHC class I molecules.
Ž .Interleukin 7 IL-7 is a bone marrow stromalcell-derived cytokine with multiple effects on B cellgrowth. It has been reported to augment proliferationŽ .Carini and Essex, 1994 and antigen-specific cyto-
Žtoxic activity Hickman et al., 1990; Plebanski et al.,.1995 of human CD8q CTL in vitro. The effect of
supplementation of CTL cultures with IL-7 was as-sessed and the dose–response relationship over arange of IL-7 concentrations was investigated. How-ever, IL-7 is a component of protocols for priming
Žhuman antigen-naive CTL in vitro Celis et al., 1994;.Plebanski et al., 1995 . We therefore also investi-
gated whether this protocol induces primary CTL invitro from PBMC from a series of unexposed con-trols. The mechanism by which IL-7 enhances thegeneration of peptide-specific CTL was also anal-ysed; antigen-specific CD8q CTL were enumeratedduring in vitro restimulation and the effects of IL-7on the specific sub-population were assessed.
Procedures for CTL culture vary widely betweendifferent laboratories. The protocol described herehas been developed and critically evaluated to pro-vide a standardized, powerful and robust method forgenerating specific effector CTL from precursors inPBMC without resulting in the priming of antigen-naive CTL precursors in vitro.
2. Materials and methods
2.1. Peptides
Six peptides were used for restimulation and forsensitizing target cells; these are listed in Table 1ŽMorrison et al., 1992; Bowness et al., 1993; Lee et
.al., 1995; Dong et al., 1996 . A panel of HLA-B35-restricted peptide epitopes from HIV-1 were used forrestimulation in control experiments in HIV-1seronegative donors. These were peptides from Nef
Ž .corresponding to residues 75–82 VPLRPMTY ,from Pol corresponding to residues 329–337Ž .HPDIVIYQY , from p24 corresponding to residues
Ž .260–269 PPIPVGDIY and from p17 correspondingŽ . Žto residues 130–138 NSSKVSQNY Rowland-
.Jones et al., 1995 . Peptides were synthesized onsolid phase on a semi-automated peptide synthesizerŽ .Zinsser Analytical, Frankfurt and purity confirmed
Ž .by high pressure liquid chromatography HPLC .
Table 1Peptide epitopes used to restimulate and test CTL for optimization of the protocol
Virus Protein Position Sequence HLA restriction
Influenza A matrix M1 58–66 GILGFVFTL HLA-A201Influenza A matrix M1 128–135 ASCMGLIY HLA-B35Influenza A nucleoprotein NP 380–388 ELRSRYWAI HLA-B8Influenza A nucleoprotein NP 383–391 SRYWAIRTR HLA-B2705EBV EBNA 3A EBNA 458–466 YPLHEQHGM HLA-B35HIV Gag GAG 77–85 SLYNTVATL HLA-A201

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–77 67
2.2. Donors and HLA typing
Subjects were healthy laboratory personnel andwere HLA typed serologically with complement me-diated lymphocytotoxicity using fresh peripheral
Ž .blood mononuclear cells PBMC and Terasaki tis-sue typing plates. Donors SD and AS were molecu-larly typed by ARMS-PCR with sequence-specific
Ž .primers Krausa et al., 1993 . For the studies demon-strating that the protocol without IL-7 does not gen-erate in vitro primary HIV-specific CTL, subjectswere healthy Gambian blood donors who tested neg-
Žative for HIV antibodies by ELISA Murex Diagnos-.tics, Dartford, Kent .
2.3. Virus-stimulated specific CTL
Influenza-specific CTL were generated fromŽhealthy volunteers as previously described Gotch et
.al., 1987 . Briefly, PBMC were incubated with aŽ .low-dose of 6 haemagglutination units HAU In-
fluenza A virus, strain X31, for 1 h in the absence ofŽ .serum, then cultured in 10% foetal calf serum FCS
Ž .supplemented medium R10 for 7–10 d. CTL lineswere generated by mixing equal numbers of bulk-
Ž .culture cells and irradiated 2000 rad , washedŽ .EBV-transformed B-lymphoblastoid cells B-LCL
preincubated with 50 mM peptide, and grown in R10with 10 Urml recombinant human interleukin 2Ž .rhIL-2, a kind gift from Cetus, Emeryville, CAadded on day 3 and every 4 d thereafter.
HIV-specific CTL were generated from HIVŽseropositive donors as previously described Nixon
.et al., 1988 . Briefly, the PBMC were divided intotwo portions with approximately 7r8 in one flask ata density of 1–1.5 million cellsrml. The remainingcells were stimulated with PHA at 5 mgrml in a 2ml well of a 24 well tissue culture plate; after 24 hthese cells were washed once and added back to theflask. After 7 d 10 Urml rhIL-2 was added to theculture.
2.4. Peptide-stimulated specific CTL
Peptide-stimulated cultures were set up fromhealthy volunteers and HIV seropositive donors.PBMC at a high cell density in 100 ml R10 werepulsed for 1 h at 378C in 5% CO at a peptide2
concentration of 100 mM and then diluted up to
8–12 ml in R10 and plated out into 2 ml wells of24-well tissue culture plates. PBMC were seeded at 2millionrml, a cell density that we have previouslyfound to be optimal for CTL generation. Wheredifferent peptide concentrations were evaluated, thepeptide concentration used is stated in the figurelegends. For certain cultures, recombinant human
ŽIL-7 rhIL-7, R and D Systems Europe, Abingdon or.supplied as a kind gift by Sanofi-Recherche, Labege
was added at a concentration of 25 ngrml at thisŽ .time point. 10 Urml rhIL-2 Cetus, Emeryville, CA
were added on day 3, and every 4 d thereafter, to allcultures. All experiments were performed with bulkculture CTL that were restimulated only once withpeptide on day 0; further restimulation with peptide-pulsed B-LCL was not employed. CTL were testedin 51Cr release CTL assays after 8–14 d of restimula-tion.
2.5. 51Cr release CTL assays
Target cell lines were HLA-matched B-LCL.Standard chromium release assays were performed
Ž .as previously described Hill et al., 1992 . Briefly,51 ŽB-LCLs were labeled with 100 mCi Cr Amersham
.International, Amersham for 1 h at 378C, washedtwice in R10 then pulsed with peptide at a concentra-tion of 10 mM in 100–200 ml R10 for 1 h at 378C.Cells were then washed twice more in R10 andplated out at 5000 cells per well of a U-bottomed 96well plate. CTL, R10 or 5% Triton X-100 wereadded; final volumes were 150 ml per well. Plateswere incubated for 4 h at 378C, 5% CO and 20 ml2
supernatant harvested and spotted onto filtermatsfrom which 51Cr release was measured in a LKB
Ž .1205 beta-plate scintillation counter Wallac .Virus-infected targets were prepared as follows.
For influenza, B-LCL were incubated for 1 h with100 ml influenza A Xr31 of titre no less than 2000HAUrml in serum free medium. Infection was haltedby adding 2 ml R10 and cells were incubated for afurther 3 h to allow expression of viral proteins. ForHIV, 1 million B-LCL were infected with recombi-
Ž .nant vaccinia viruses kind gifts of B. Moss at aŽ .multiplicity of infection MOI of 3–6 pfurcell in
R10 for 1 h at 378C, washed and then incubatedovernight at 378C in 1 ml of R10; alternativelyB-LCL were infected at a MOI of 10 pfurcell for 1h at 378C, washed and incubated for 2–3 h at 378C.

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–7768
Specific lysis was calculated as follows:
% lysiss test well cpmyspontaneous cpmŽ .
r maximum cpmyspontaneous cpm =100Ž .
% specific lysis was calculated by subtracting the %lysis on the control targets from the % lysis on thepeptide pulsed or virus-infected targets. Spontaneousrelease refers to targets incubated with R10 aloneand maximal release refers to targets incubated with5% Triton X-100; cpmscounts per minute. Maxi-mum spontaneous release never exceeded 30%.
2.6. Enumeration of peptide-specific CD8q T cellswith tetrameric peptide-MHC complexes
Tetramers of the HLA-A2 molecule and the HIV-1GAG 77–85 peptide were constructed in order toproduce a ligand cognate for the T cell receptorŽ .TCR on HLA-A2-restricted CD8q T cells specificfor the GAG 77–85 epitope; the synthetic ligand wasthen used to stain specifically these cells. A 15amino acid substrate peptide for BirA-dependent bi-otinylation was added to the COOH-terminus of the
Ž .HLA-A2 heavy chain Schatz, 1993 . After foldingthe heavy chain fusion protein in vitro in the pres-ence of beta-2-microglobulin and the specific peptide
Ž .ligand GAG 77–85 Garboczi et al., 1992 , thepurified MHC-peptide complex was biotinylated ona single lysine within the BirA substrate peptideŽ .Altman et al., 1996 . Tetramers were produced bymixing the biotinylated peptide-MHC complex withphycoerythrin-labeled deglycosylated avidin at a mo-lar ratio of 4:1. The avidity of these tetramericcomplexes for their cognate TCR permitted efficientstaining of CD8q T cells; FACS analysis thenquantified the percentage of specific T cells in cul-ture.
2.7. Freezing and thawing PBMC
PBMC were cryopreserved at a density of 5 mil-lionrml in 10% DMSO and 90% FCS in cryotubesand stored at y708C. The cells were thawed after 1week, 1 month or 2 months, immediately washedtwice in cold R10 and plated out for restimulationwith peptide as described above.
3. Results
3.1. Peptide stimulation generates MHC class I-re-stricted CTL more efficiently than whole Õirus
Influenza-specific HLA-B35-restricted CTL wereŽgenerated from a healthy donor PTE class I HLA
.haplotype: A24, A29; B35, B4802 . CTL were gen-erated by restimulation with either influenza virus AXr31 or with the HLA-B35-restricted influenza ma-trix peptide M1 128–135. The resultant bulk culture
Ž .CTL were HLA-B35-restricted data not shown .CTL were tested for peptide-specific cytotoxicityagainst autologous B-LCL pulsed with the peptideM1 128–135 in 51Cr release assays. Fig. 1A showsthat the peptide-stimulated CTL show enhanced spe-
Ž .cific lysis at each effector:target E:T ratio com-pared to virus-stimulated cultures, notwithstandingmodestly increased non-specific lysis for the peptidestimulated CTL at the highest E:T ratio.
3.2. Peptide stimulated CTL recognize naturally pro-cessed antigen
Influenza-specific CTL stimulated with peptiderecognized influenza virus-infected targets in anHLA-restricted manner. Fig. 1B shows a representa-tive experiment with lysis of heterologous HLA-B35matched influenza-infected targets by CTL fromdonor PTE restimulated with the HLA-B35-restrictedinfluenza epitope M1 128–135. CTL generated witha variety of other influenza and HIV peptides havealso been shown to recognize endogenously pro-
Ž .cessed antigen data not shown .
3.3. Effects of peptide concentration on efficiency ofrestimulation
The concentration of peptide used for restimula-tion was varied. Increasing the peptide concentrationin the initial 1 h prepulse of freshly isolated PBMCaugmented the resultant lytic activity of CTL bulkcultures, as shown for the HLA-A2.01-restricted in-
Ž .fluenza matrix epitope M1 58–66 Fig. 2A and theHLA-B8-restricted influenza nucleoprotein epitope
Ž .NP 380–88 Fig. 2B . The initial high concentrationprepulse appears to be important; when the prepulsewas dispensed with and PBMC were cultured at a

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–77 69
Ž . 51Fig. 1. A Comparison of percentage lysis in Cr release assayby CTL restimulated with peptide and CTL restimulated with
Žvirus. PBMC from HLA-B35 positive donor PTE class I HLA.haplotype: A24, A29; B35, B48.02 were stimulated with in-
fluenza A Xr31 or the HLA-B35 restricted influenza A matrixpeptide ASCMGLIY; targets were autologous B-LCL. Assays
Ž .were performed after 14 d culture. B CTL from donor PTErestimulated with the HLA-B35-restricted peptide ASCMGLIYefficiently recognize influenza A virus-infected HLA-B35 matchedheterologous B-LCL in the 51Cr release assay. The target B-LCLis an HLA-A201, -B35 homozygous typing B-LCL. The assaywas performed at day 14.
single continuous peptide concentration from the be-ginning of restimulation, peptide concentrations ofup to 25 mM in the culture medium did not generatedetectable CTL, as shown for the HLA-B35-re-
Žstricted influenza matrix epitope M1 128–135 Fig..2C .
3.4. Effects of IL-7 on peptide-specific CTL actiÕity
IL-7, added to the culture medium once at thestart of restimulation, reproducibly enhanced the pep-tide-specific responses of peptide-stimulated CTL
Ž .from all donors tested ns10 with no increase in
non-specific lysis. Representative 51Cr release assaysfor CTL cultured with and without IL-7 are illus-trated for a range of HLA class I-restricted CTLepitopes in Fig. 3, which shows IL-7-induced aug-mentation of responses to the HLA-A201-restricted
Ž .influenza peptide M1 58–66 Fig. 3A , the HLA-ŽB35-restricted influenza peptide M1 128–135 Fig.
.3B , the HLA-B2705-restricted influenza peptide NPŽ .383–391 Fig. 3C and the HLA-B35-restricted EBV
Ž .EBNA 3A peptide 458–466 Fig. 3D . The enhance-ment of specific lysis with IL-7 was observed to asimilar extent whether CTL were assayed after 7Ž . Ž . Ž .Fig. 3B and C , 10 Fig. 3A or 14 d Fig. 3D in
Žculture. In some instances as, for example, in Fig..3D , responses from donors known to give a weak
response were below the threshold level of 10%peptide-specific lysis but became detectable if CTLwere cultured in the presence of IL-7.
3.5. The peptide based restimulation protocol, withor without IL-7, does not induce primary CTL re-sponses in Õitro
The peptide restimulation protocol without IL-7,using the HLA-B35 restricted HIV-1 epitopes from
Ž .Nef, Pol, p24 and p17 listed in Section 2.1 wereused to culture PBMC from a series of 21 healthyHIV seronegative donors with HLA-B35. Cultureswere assayed at days 7, 14 and 21 against targetspulsed with the respective peptides; CTL activity
Žwas not detected in any of the 21 donors data not.shown .
The peptide restimulation protocol incorporatingIL-7 was used to culture PBMC from a series ofhealthy donors who were all laboratory personnelwith no history of HIV exposure; a total of 22distinct HIV specific responses were assessed through7 different HLA class I alleles. Where epitopes ininfluenza A virus or EBV are known for each ofthese alleles, CTL responses were assessed. Table 2shows that under the conditions of our protocol, CTLto the influenza and EBV epitopes were readilydetected in several individuals but in none of thedonors could HIV-specific CTL be detected.
3.6. Dose–response studies with IL-7 suggest anoptimal IL-7 concentration for CTL generation
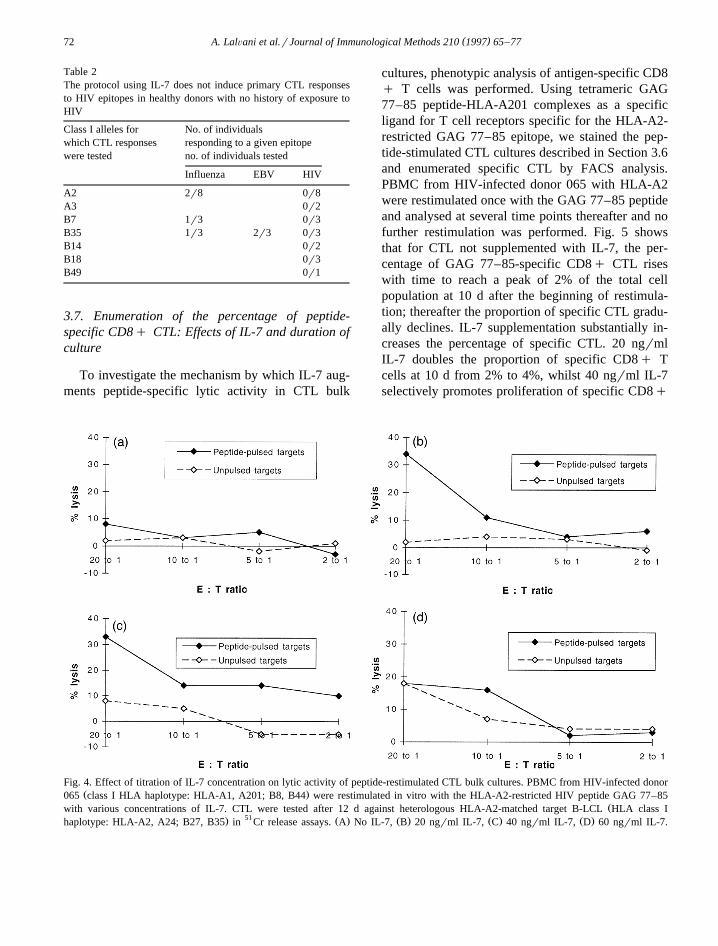
The effect of a broad range of initial IL-7 concen-trations on lytic activity in CTL bulk cultures was

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–7770
assessed. Fig. 4 shows the dose–response relation-ship for CTL specific for the HLA-A2-restrictedHIV epitope GAG 77–85. CTL were generated from
Ždonor 065 HLA class I haplotype: HLA-A1, A2;
.B8, B44 with the peptide GAG 77–85 and testedafter 12 d restimulation against peptide-pulsed andunpulsed heterologous HLA-A2-matched target B-LCL in 51Cr release assays. IL-7 concentrations

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–77 71
Ž .Fig. 3. IL-7 augments lytic activity of peptide-restimulated CTL for a range of HLA class I-restricted epitopes. A CTL from donor ASwith HLA-A2.01 generated with the HLA-A2.01-restricted influenza matrix peptide M1 58–66 tested after 10 d culture across a range of
Ž . Ž .E:T ratios against HLA-A2.01-matched peptide-pulsed heterologous B-LCL JN . B CTL from donor PTE, generated on the HLA-B35Ž .restricted influenza matrix peptide M1 128–135, assayed on peptide-pulsed autologous B-LCL after 7 d culture at an E:T ratio of 5:1. C
Ž .CTL from donor SD class I HLA haplotype: A201, A32; B2705, B49 , generated on the HLA-B2705 restricted influenza nucleoproteinŽ .peptide NP 383–391, assayed after 7 d culture against heterologous HLA-B2705 positive, peptide-pulsed B-LCL at an E:T ratio of 20:1. D
illustrates how IL-7 augmented an otherwise undetectable response. CTL from HLA-B35 positive donor WR were restimulated with theHLA-B35 restricted EBV EBNA3 peptide 458–466 and assayed after 14 d on heterologous peptide-pulsed HLA-B35 B-LCL at an E:T ratioof 5:1.
ranging from 0–60 ngrml were studied; lytic activ-ity was highest for concentrations of 20–40 ngrml.However, non-specific lysis of unpulsed targets wassignificantly increased at higher effector:target ratios
for CTL cultured at 40 ngrml IL-7, whilst no in-crease in non-specific lysis was observed at 20 ngrmlIL-7. Net peptide-specific lysis was thus maximal atthe 20 ngrml IL-7 concentration.
Ž . Ž . ŽFig. 2. A–C The effect of peptide concentration on the efficiency of restimulation of CTL. A CTL line from donor AS class I HLA.haplotype: HLA-A1, -A2; -B8, -B35 restimulated with the HLA-A2.01-restricted influenza matrix peptide M1 58–66 at differing peptide
prepulse concentrations. The line was tested after 10 d culture in a 51Cr release assay against a HLA-A2.01-matched heterologous B-LCLŽ . Ž . ŽJN; class I HLA haplotype: HLA-A201, -1101: B-51, -52 . B CTL line from donor AVSH class I HLA haplotype: HLA-A1, -24; B-8,
.-39 restimulated with the HLA-B8-restricted influenza nucleoprotein peptide NP 380–388 at differing peptide prepulse concentrations. The51 Žline was tested after 8 d culture in a Cr release assay against a HLA-B8-matched heterologous B-LCL PC; class I HLA haplotype: A-1,
. Ž .-3; B-7, -8 . C Using the HLA-B35-restricted matrix peptide M1 128–135, PBMC from donor PTE were restimulated with differingconcentrations of free peptide, up to 25 mM, and with a prepulse at a high concentration of 200 mM; only CTL generated with the highconcentration prepulse had demonstrable cytolytic activity when tested on autologous peptide-pulsed B-LCL.
( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–7772
Table 2The protocol using IL-7 does not induce primary CTL responsesto HIV epitopes in healthy donors with no history of exposure toHIV
Class I alleles for No. of individualswhich CTL responses responding to a given epitopewere tested no. of individuals tested
Influenza EBV HIV
A2 2r8 0r8A3 0r2B7 1r3 0r3B35 1r3 2r3 0r3B14 0r2B18 0r3B49 0r1
3.7. Enumeration of the percentage of peptide-specific CD8q CTL: Effects of IL-7 and duration ofculture
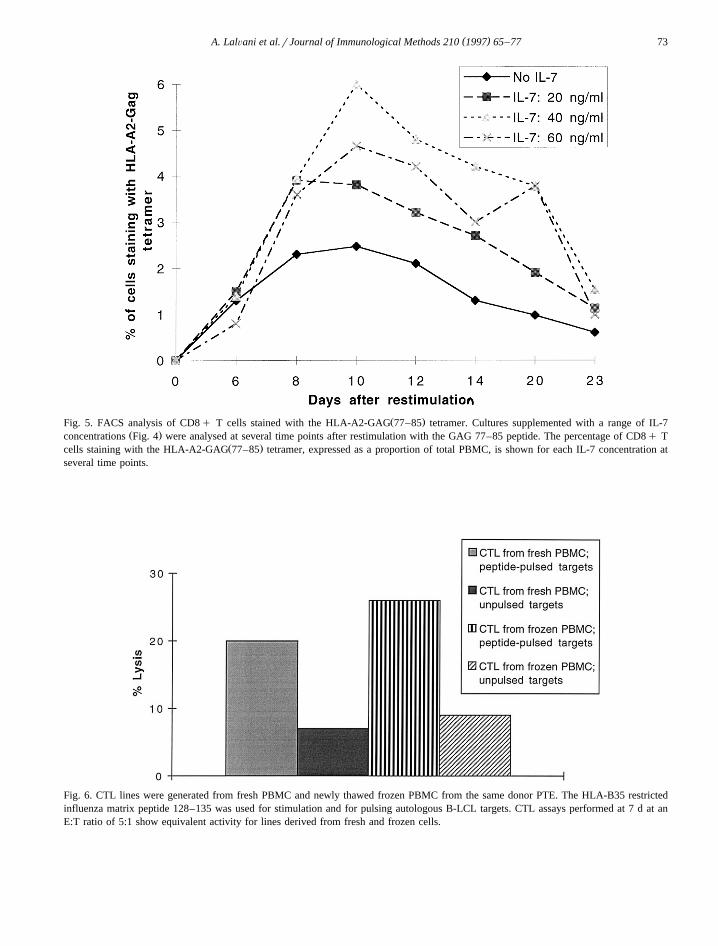
To investigate the mechanism by which IL-7 aug-ments peptide-specific lytic activity in CTL bulk
cultures, phenotypic analysis of antigen-specific CD8q T cells was performed. Using tetrameric GAG77–85 peptide-HLA-A201 complexes as a specificligand for T cell receptors specific for the HLA-A2-restricted GAG 77–85 epitope, we stained the pep-tide-stimulated CTL cultures described in Section 3.6and enumerated specific CTL by FACS analysis.PBMC from HIV-infected donor 065 with HLA-A2were restimulated once with the GAG 77–85 peptideand analysed at several time points thereafter and nofurther restimulation was performed. Fig. 5 showsthat for CTL not supplemented with IL-7, the per-centage of GAG 77–85-specific CD8q CTL riseswith time to reach a peak of 2% of the total cellpopulation at 10 d after the beginning of restimula-tion; thereafter the proportion of specific CTL gradu-ally declines. IL-7 supplementation substantially in-creases the percentage of specific CTL. 20 ngrmlIL-7 doubles the proportion of specific CD8q Tcells at 10 d from 2% to 4%, whilst 40 ngrml IL-7selectively promotes proliferation of specific CD8q
Fig. 4. Effect of titration of IL-7 concentration on lytic activity of peptide-restimulated CTL bulk cultures. PBMC from HIV-infected donorŽ .065 class I HLA haplotype: HLA-A1, A201; B8, B44 were restimulated in vitro with the HLA-A2-restricted HIV peptide GAG 77–85
Žwith various concentrations of IL-7. CTL were tested after 12 d against heterologous HLA-A2-matched target B-LCL HLA class I. 51 Ž . Ž . Ž . Ž .haplotype: HLA-A2, A24; B27, B35 in Cr release assays. A No IL-7, B 20 ngrml IL-7, C 40 ngrml IL-7, D 60 ngrml IL-7.
( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–77 73
Ž .Fig. 5. FACS analysis of CD8q T cells stained with the HLA-A2-GAG 77–85 tetramer. Cultures supplemented with a range of IL-7Ž .concentrations Fig. 4 were analysed at several time points after restimulation with the GAG 77–85 peptide. The percentage of CD8q T
Ž .cells staining with the HLA-A2-GAG 77–85 tetramer, expressed as a proportion of total PBMC, is shown for each IL-7 concentration atseveral time points.
Fig. 6. CTL lines were generated from fresh PBMC and newly thawed frozen PBMC from the same donor PTE. The HLA-B35 restrictedinfluenza matrix peptide 128–135 was used for stimulation and for pulsing autologous B-LCL targets. CTL assays performed at 7 d at anE:T ratio of 5:1 show equivalent activity for lines derived from fresh and frozen cells.

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–7774
T cells to 6%. Still higher concentrations of IL-7 are,however, less effective. Interestingly, the expansionof the antigen-specific population of CD8q CTLinduced by different concentrations of IL-7 broadlycorrelates with the lytic activity of the same CTL
Ž .cultures on peptide-pulsed targets Fig. 4 , althoughfor higher E:T ratios the increase in IL-7 concentra-tion from 20 to 40 ngrml gave no further incrementin peptide-specific lytic activity. The temporal pat-tern of the expansion, and subsequent contraction, ofthe specific CTL during in vitro restimulation is notaltered by IL-7 and the selective increase of thepeptide-specific population of CD8q T cells is sus-tained for up to three weeks.
3.8. The restimulation protocol is effectiÕe for frozenPBMC
Fresh PBMC and newly thawed cryopreservedPBMC from the same donor were set up with pep-tide in identical fashion at the same time point. CTLassays were performed after 7 and 14 d culture; therewas no difference between CTL derived from freshor frozen PBMC. Fig. 6 shows a representativeexperiment out of several that were performed. Linescould be generated with peptide pulsed B-LCL res-timulation from frozen cells as efficiently as fromfresh cells.
4. Discussion
Peptides have become widely used for the in vitrogeneration of specific effector CTL from the pool ofmemory CTL in PBMC. They are more convenientto use than the natural pathogen, especially whenculturing the organism damages PBMC as is the casewith HIV and Mycobacterium tuberculosis; in otherinstances, such as the liver stage of P. falciparum, invitro culture is not practical. The techniques used forpeptide restimulation differ considerably. Althougheffector CTL activity generated in vitro from CTLprecursors does not necessarily reflect effector CTLactivity in vivo, there is a need for a sensitive andreliable method for detecting CTL precursors, partic-ularly one which could be used to measure CTLinduction after vaccination. We have described herea standardized, peptide-based protocol for generatingeffector CTL; several parameters have been opti-
mized to maximize the efficiency of in vitro restimu-lation.
We have demonstrated that peptide restimulation,as measured by the magnitude of specific lysis in51Cr release CTL assays, is more effective than virusrestimulation with influenza. This is in line with ourprevious observation that in HIV-exposed but unin-fected Gambian women, in vitro stimulation withpeptide successfully generated secondary effector
ŽCTL but stimulation with HIV failed to do so Row-.land-Jones et al., 1995 . Parallel experiments have
shown that peptides from P. falciparum liver stageantigens are more proficient at restimulation thanvaccinia viruses recombinant for genes encoding
Ž .these antigens Aidoo et al., 1997 . CTL generatedwith peptide have similar fine antigen specificities tothose generated by conventional virus stimulationand can recognize naturally processed antigen invirus-infected targets.
The concentration of peptide used for restimula-tion is important; for 3 different peptides we found
Žthat a high concentration of peptide typically 100.mM in the initial 1 h prepulse at the start of
Žrestimulation promoted stronger CTL responses Fig..2 A–C . This was true of epitopes with a high
affinity for their respective HLA class I moleculeŽŽthe HLA-A2.01-restricted influenza peptide M1
Ž .58–66 Morrison et al., 1992 and the HLA-B8-re-Ž ..stricted peptide NP 380–388 Sutton et al., 1993 as
well as the HLA-B35-restricted peptide M1 128–135Žwhich has a low affinity for HLA-B35 Dong et al.,
.1996 . Thus, when mapping unknown putative epi-topes it is necessary to use a high concentration ofthe candidate peptides for the prepulse; we routinelyuse a prepulse concentration of 100 mM.
Supplementation of initial PBMC cultures withIL-7 at a concentration of 25 ngrml was shown toaugment peptide-specific lytic activity of bulk cul-
Ž .ture CTL for 5 different peptides Figs. 3 and 4 .Low-level and borderline responses became readilydetectable with IL-7; indeed this protocol has oftenmade it possible to detect weak responses which
Žwould otherwise have been missed unpublished ob-.servations .
We studied the dose–response effect of IL-7 bytitrating the concentration of IL-7 over a range from0 to 60 ngrml. Lysis of peptide-pulsed targets at low
Ž .E:T ratios 5:1 or less was augmented by increasing

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–77 75
IL-7 concentrations up to 40 ngrml with no increasein non-specific lysis of unpulsed targets. At the
Ž .higher E:T ratio 20:1 , increasing the IL-7 concen-tration from 20 to 40 ngrml did not further enhancelysis of peptide-pulsed targets yet resulted in in-creased levels of non-specific lysis of unpulsed tar-gets. Increasing the IL-7 concentration to 60 ngrmlfurther promoted non-specific lysis and actually di-minished lysis of peptide-pulsed targets. The in-creased non-specific lysis observed at higher concen-trations may reflect the fact that IL-7 can increasethe cytotoxic activity of the CD56 bright subset of
Ž .natural killer cells Dadmarz et al., 1994 . Our re-sults indicate that the optimum concentration of IL-7for maximizing peptide-specific lysis, whilst keepingnon-specific lysis to a minimum, lies between 20 and40 ngrml. This is consistent with previous dose–re-
Žsponse studies Hickman et al., 1990; Carini and.Essex, 1994 which have shown that the increased
cytolytic activity of CTL reaches a plateau at rhIL-7concentrations of 1000–1,200 Urml, equivalent tothe 25 ngrml concentration used in our protocol.
How does IL-7 enhance peptide-specific lytic ac-tivity? The effect of IL-7 on the generation of sec-ondary antigen-specific responses appears to be adirect effect on CD8q T cells and is independent of
ŽIL-2 Kos and Muellbacher, 1993; Carini and Essex,.1994 . IL-7 augments proliferation of CD8q T cells
Ž .Carini and Essex, 1994 , promotes in vitro differen-Žtiation of memory CTLp to effector CTL Kos and
.Muellbacher, 1993 and increases specific cytolyticŽactivity of CTL Hickman et al., 1990; Plebanski et
.al., 1995 . We performed a quantitative analysis ofthe effect of IL-7 on peptide-specific CD8q CTL inculture. A novel technique using peptide-MHC classI molecule tetramers enabled us to enumerate specif-ically those CD8q T cells cognate for the peptideepitope used for restimulation in cultures supple-mented with a variety of different IL-7 concentra-tions and at several different time points after initia-tion of restimulation. In this way both the optimalconcentration of IL-7 for specific CTL generationand the kinetics of CTL generation in vitro could
Ž .clearly be seen Fig. 5 . This powerful technique,Žwhich correlates with the results of CTL assays Fig.
.4 , will be valuable for assessing the efficiency ofother methods of CTL generation in the future.
Fig. 5 also demonstrates that for both IL-7-supp-
lemented and -unsupplemented cultures, the size ofthe antigen-specific CTL population peaks at around10 d, suggesting that this is the best time point atwhich to test CTL bulk cultures in 51Cr release CTLassays. Although IL-7 did not prolong the survival ofthe expanded peptide-specific CTL population in thebulk cultures illustrated in Fig. 5, we have occasion-ally observed that regular supplementaion of ex-tended cultures with IL-7 enabled specific CTL to begrown in culture for up to 7 weeks without restimu-lation with peptide-pulsed B-LCL and with no lossof antigen-specifity.
FACS analysis of the CTL bulk cultures in Figs. 4and 5 also demonstrated that as the IL-7 concentra-tion was titrated upwards, the number of large cellsand blasts increased significantly, suggesting an in-
Žcrease in the proportion of activated T cells data not.shown . This is consistent with a previous cytofluo-
rometric analysis of CTL generated in a recombinantvacciniarHIV-1 stimulation strategy from PBMCfrom HIV-infected patients; IL-7 preferentially ex-
Ž .panded CD8 memory cells CD45ROq and CD8qlymphocytes expressing the activation molecules
Ž .HLA-DR, CD25 and CD38 Ferrari et al., 1995 .The fact that IL-7 is an important component of
protocols for generating primary CTL responses fromŽantigen-naive precursors in vitro Celis et al., 1994;
.Plebanski et al., 1995 raises the possibility that inour method antigen naive cells may be primed,expanded and recruited to make a significant contri-bution to the effector response. However, protocolsfor generating primary CTL in vitro have all requiredrepeated restimulation with peptide-pulsed fresh au-
Ž .tologous PBMC or adherent fractions thereof , or, inaddition, the use of powerful adjuvant moleculessuch as KLH. Presumably the fresh autologousPBMC are an abundant source of antigen presentingcells bearing accessory molecules necessary forpriming. It would therefore appear unlikely that ourprotocol could induce primary CTL. Nonetheless wetested our protocol, both with and without IL-7,using a panel of HIV epitopes, on PBMC from alarge series of healthy donors with no history of HIVexposure; CTL activity to the HIV peptides was notdetected in any of these subjects. We are thereforeconfident that this protocol, whilst enhancing low-level secondary CTL responses, does not lead to theinduction of primary CTL in vitro.

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–7776
Several features of this protocol make it readilyreproducible and versatile: peptides are simpler touse and transport than natural pathogens, and PBMCcould be frozen after separation with no loss in theefficacy of peptide restimulations as compared with
Ž .fresh PBMC Fig. 6 .In conclusion, a standardized and highly efficient
protocol for generating effector CTL from CTLp inPBMC has been developed. The use of IL-7 selec-tively expands the population of peptide-specific CTLthereby enhancing the sensitivity with which CTLcan be detected, yet it does not elicit primary re-sponses. The protocol is simple, versatile and effec-tive at detecting low level CTL responses whereprecursor frequencies may be low. These advantagesmake it suitable for use in the field when studyinginfectious diseases and for evaluating the immuno-genicity of new CTL-inducing vaccines in clinicaltrials.
Acknowledgements
AL and TD have contributed equally to the workpresented here. AL and GO are MRC Clinical Train-ing Fellows, AVSH a Wellcome Trust PrincipalResearch Fellow and SR-J an MRC Senior ClinicalFellow. TD and AJM are supported by the MRC andAAP by the Wellcome Trust. We are very grateful to
Ž .Dr. Adrian Minty of Sanofi-Recherche Labege forthe kind gift of rhIL-7 and to Dr. Magdalena Pleban-ski for advice and helpful discussions.
References
Aidoo, M., Lalvani, A., Allsopp, C.E.M., Plebanski, M., Meisner,S.J., Browning, M., Morris-Jones, S., Gotch, F., Fidock, D.A.,Takiguchi, M., Robson, K.J.H., Greenwood, B.M., Druilhe, P.,Whittle, H.C., Hill, A.V.S., 1995. Identification of conservedantigenic components for a cytotoxic T lymphocyte-inducingvaccine against malaria. Lancet 345, 1003.
Aidoo, M., Lalvani, A., Whittle, H.C., Hill, A.V.S., Robson,K.J.H., 1997. Recombinant vaccinia viruses for the characteri-zation of Plasmodium falciparum-specific cytotoxic T lympho-cytes: Recognition of processed antigen despite limited restim-ulation efficacy. Int. Immunol. 9, 731.
Altman, J.D., Moss, P.A.H., Goulder, P.J.R., Barouch, D.H.,McHeyzer-Williams, M.G., Bell, J.I., McMichael, A.J., Davis,M.M., 1996. Phenotypic analysis of antigen-specific T lym-phocytes. Science 274, 94.
Borysiewicz, L.K., Fiander, A., Nimako, M., Man, S., Wilkinson,G.W.G., Westmoreland, D., Evans, A.S., Adams, M., Stacey,S.N., Boursnell, M.E.G., Rutherford, E., Hickling, J.K., Inglis,S.C., 1996. A recombinant vaccinia virus encoding humanpapilloma virus types16 and 18, E6 and E7 proteins as im-munotherapy for cervical cancer. Lancet 347, 1523.
Bowness, P., Moss, P.A.H., Rowland-Jones, S.L., Bell, J.I.,McMichael, A.J., 1993. Conservation of T cell receptor usageby HLA-B27-restricted influenza-specific CTL suggests a gen-eral pattern for antigen-specific MHC class I-restricted re-sponses. Eur. J. Immunol. 23, 1417.
Carini, C., Essex, M., 1994. Il-2 independent IL-7 activity en-hances cytotoxic immune response of HIV-1 infectedc individ-uals. Aids Res. Hum. Retr. 10, 121.
Celis, E., Tsai, V., Crimi, C., deMars, R., Wentworth, P.A.,Chesnut, R.W., Grey, H.M., sette, A., Serra, H.M., 1994.Induction of anti-tumor CTL in normal humans using primarycultures and synthetic peptide epitopes. Proc. Natl. Acad. Sci.USA 91, 2105.
Dadmarz, R., Bockstoce, D.C., Golub, S.H., 1994. IL-7 selec-tively enhances natural kill cytotoxicity mediated by theCD56-bright natural killer population. Lymphokine CytokineRes. 13, 349.
Dong, T., Boyd, D., Rosenberg, W., Alp, N., Takiguchi, M.,McMichael, A.J., Rowland-Jones, S., 1996. Increased affinityfor HLA-B35 of an influenza A epitope modified at an anchorresidue results in an antagonistic peptide. Eur. J. Immunol. 26,335.
Garboczi, D.N., Hung, D.T., Wiley, D.C., 1992. HLA-A2-peptidecomplexes: Refolding and crystallization of molecules ex-pressed in Escherichia coli and complexed with single anti-genic peptides. Proc. Natl. Acad. Sci. USA 89, 3429.
Hickman, C.J., Crim, J.A., Mostowski, H.S., Siegel, J.P., 1990.Regulation of human cytotoxic T lymphocyte development byIL-7. J. Immunol. 145, 2415.
Hill, A.V.S., Elvin, J., Willis, A., Aidoo, M., Allsopp, C.E.M.,Gotch, F.M., Gao, X.M., Takiguchi, M., Greenwood, B.M.,Townsend, A.R.M., McMichael, A.J., Whittle, H.C., 1992.Molecular analysis of the association of HLA-B53 and resis-tance to severe malaria. Nature 360, 434.
Kos, F.J., Muellbacher, A., 1993. IL-2 independent activity ofIL-7 in the generation of secondary antigen-specific cytotoxicT cell responses in vitro. J. Immunol. 150, 387.
Ferrari, G., King, K., Rathbun, K., Place, C.A., Packard, M.V.,Bartlett, J.A., Bolognesi, D.P., Weinhold, K.J., 1995. IL-7enhancement of antigen-driven activationrexpansion of HIV-
Ž .1-specific cytotoxic T lymphocyte precursors CTLp . Clin.Exp. Immunol. 101, 239.
Gotch, F.M., Rothbard, J., Howland, K., Townsend, A.,McMichael, A.J., 1987. Cytotoxic T lymphocytes recognise afragment of influenza virus matrix protein in association withHLA-A2. Nature 326, 881.
Krausa, P., Bodmer, J.G., Browning, M.J., 1993. Defining thecommon subtypes of HLA A9, A10, A28 and A19 by use ofARMSrPCR. Tissue Antigens 42, 91–99.
Lalvani, A., Brookes, R., Wilkinson, R., Malin, A.S., Pathan,A.A., Dockrell, H., Andersen, P., Pasvol, G., Hill, A.V.S.,

( )A. LalÕani et al.rJournal of Immunological Methods 210 1997 65–77 77
1997. Human cytolytic and interferon-gamma secreting CD8qT cells specific for Mycobacterium tuberculosis. J. Immunol.Methods, submitted.
Lee, S.P., Morgan, S., Skinner, J., Thomas, W.A., Rowland-Jones,S.L., Khanna, R., Whittle, H.C., Rickinson, A.B., 1995. EBVisolates with the major HLA B35.01-restricted CTL epitopeare prevalent in a highly B35.01-positive African population.Eur. J. Immunol. 25, 102.
Martinon, F., Gomard, E., Hannoun, C., Levy, J.-P., 1990. In vitrohuman cytotoxic T cell responses against influenza A viruscan be induced and selected by synthetic peptides. Eur. J.Immunol. 20, 2171.
Morrison, J., Elvin, J., Latron, F., Gotch, F., Moots, R., Stro-minger, J.L., McMichael, A.J., 1992. Identification of thenonamer peptide from influenza-A matrix protein and the roleof pockets of HLA-A2 in its recognition by cytotoxic Tlymphocytes. Eur. J. Immunol. 22, 903–907.
Nixon, D., Townsend, A.R.M., Elvin, J., Rizza, C., Gallway, J.,McMichael, A.J., 1988. HIV-1 gag-specific cytotoxic T lym-phocytes defined with recombinant vaccinia virus and syn-thetic peptides. Nature 336, 484.
Plebanski, M., Allsopp, C.E.M., Aidoo, M., Reyburn, H., Hill,
A.V.S., 1995. Induction of peptide-specific primary CTL re-sponses from human peripheral blood. Eur. J. Immunol. 25,1783.
Plebanski, M., Aidoo, M., Whittle, H.C., Hill, A.V.S., 1997.Precursor frequency analysis of cytotoxic T lymphocytes topre-erythrocytic antigens of Plasmodium falciparum in WestAfrica. J. Immunol. 158, 2849.
Rowland-Jones, S., Sutton, J., Ariyoshi, K., Dong, T., Gotch, F.,McAdam, S., Whitby, D., Sabally, S., Galimore, A., Corrah,T., Takiguchi, M., Schultz, T., McMichael, A.J., Whittle, H.,1995. HIV-specific CTL in HIV-exposed but uninfected Gam-bian women. Nat. Med. 1, 59.
Schatz, P.J., 1993. Use of peptide libraries to map the substratespecificity of a peptide-modifying enzyme: A 13 residue con-sensus peptide specifies biotinylation in Escherichia coli.Biotechnology 11, 1138.
Sutton, J., Rowland-Jones, S., Rosenberg, W., Nixon, D., Gotch,F., Gao, X.M., Murray, N., Spoonas, A., Driscoll, P., 1993. Asequence pattern for peptides presented to cytotoxic T lympho-cytes by HLA-B8 revealed by analysis of epitopes and elutedpeptides. Eur. J. Immunol. 23, 447.
Copyright © 2022 FDOKUMEN




















