
Olfactory ensheathing cells promote collateral axonal branching in the injured adult rat spinal cord
11
Olfactory ensheathing cells promote collateral axonal branching in the injured adult rat spinal cord M.I. Chuah, * D. Choi-Lundberg, S. Weston, A.J. Vincent, R.S. Chung, J.C. Vickers, and A.K. West NeuroRepair Group, School of Medicine, University of Tasmania, Hobart, Tasmania 7001, Australia Received 13 February 2003; revised 20 August 2003; accepted 16 September 2003 Abstract In recent years, injection of olfactory ensheathing cells (ECs) into the spinal cord has been used as an experimental strategy to promote regeneration of injured axons. In this study, we have compared the effects of transplanting encapsulated ECs with those injected directly into the spinal cord. The dorsal columns of adult rats were cut at T 8–9 and rats in experimental groups received either EC-filled porous polymer capsules or culture medium (CM)-filled capsules with ECs injected at the injury site. Control rats were in three groups: (1) uninjured, (2) lesion with transplantation of CM-filled capsules and (3) lesion with transplantation of CM-filled capsules and injections of CM. Three weeks after injury, Fluororuby was injected into the hindlimb motor and somatosensory cortex to label corticospinal neurons. Observations indicated that there were a few regenerating fibres, up to 10, in the EC-treated groups. In rats that received encapsulated ECs, regenerating fibres were present in close association with the capsule. Rats that received EC injections demonstrated a significant increase in the number of collateral branches from the intact ventral corticospinal tract (vCST) compared with the corresponding control, CM-injected group ( P = 0.003), while a trend for increased collateral branches was observed in rats that received encapsulated ECs ( P = 0.07). D 2003 Elsevier Inc. All rights reserved. Keywords: Olfactory ensheathing cells; Spinal cord; Regeneration; Corticospinal tract; Collateral sprouting Introduction Traumatic injury to the spinal cord triggers a series of cellular reactions that often result in wide ranging functional deficits (e.g., reviews by Fawcett and Asher, 1999; Schwab and Bartholdi, 1996). Within the first few days of injury, there is massive neuronal and glial cell death (Emery et al., 1998). Axons distal to the injury site degenerate while reactive astrocytes divide to form scar tissue, which inhibits subsequent axonal regrowth by a combination of mechan- ical and trophic mechanisms (Lindsay, 1986). The injury site is a complex environment containing reactive astro- cytes, fibroblasts, immune cells and a mix of growth factors, inhibitory factors, cytokines and other agents, some soluble and some associated with the surface of specific cells (Schwab and Bartholdi, 1996). In light of the restrictive nature of the injured central nervous system, various experimental strategies have been developed to alter this environment so that axonal repair can take place. One of these is the use of olfactory ensheathing cells (ECs). Indeed, ECs injected into the injured spinal cord have resulted in a remarkable degree of axonal regeneration and functional recovery (e.g., Imaizumi et al., 1998; Nash et al., 2002; Ramon-Cueto et al., 1998), including remyelina- tion of demyelinated axons (Li et al., 1997; Imaizumi et al., 1998). Perhaps most remarkable is the restoration of climb- ing ability of rats whose spinal cords have undergone complete transection (Ramon-Cueto et al., 2000). However, studies have yet to investigate the mechanisms through which ECs operate in the novel environment of the injured spinal cord. Results of in vitro studies have demonstrated that ECs express cell surface molecules such as N-CAM, N-cadherin and laminin (Chuah and Au, 1994; Chuah et al., 1991; Treloar et al., 1996). They also produce many growth factors including several forms of neuregulins and some neurotrophins (Chuah and West, 2002; Chuah et al., 2000; Woodhall et al., 2001). It is not known whether the 0014-4886/$ - see front matter D 2003 Elsevier Inc. All rights reserved. doi:10.1016/j.expneurol.2003.09.008 * Corresponding author. Discipline of Anatomy and Physiology, School of Medicine, University of Tasmania, Private Bag 24, Medical Sciences Building, College Road, Hobart, Tasmania 7001, Australia. Fax: +61-3-62262679. E-mail address: [email protected] (M.I. Chuah). www.elsevier.com/locate/yexnr Experimental Neurology 185 (2004) 15 – 25
Transcript of Olfactory ensheathing cells promote collateral axonal branching in the injured adult rat spinal cord

www.elsevier.com/locate/yexnr
Experimental Neurology 185 (2004) 15–25
Olfactory ensheathing cells promote collateral axonal branching
in the injured adult rat spinal cord
M.I. Chuah,* D. Choi-Lundberg, S. Weston, A.J. Vincent, R.S. Chung,J.C. Vickers, and A.K. West
NeuroRepair Group, School of Medicine, University of Tasmania, Hobart, Tasmania 7001, Australia
Received 13 February 2003; revised 20 August 2003; accepted 16 September 2003
Abstract
In recent years, injection of olfactory ensheathing cells (ECs) into the spinal cord has been used as an experimental strategy to promote
regeneration of injured axons. In this study, we have compared the effects of transplanting encapsulated ECs with those injected directly into
the spinal cord. The dorsal columns of adult rats were cut at T8–9 and rats in experimental groups received either EC-filled porous polymer
capsules or culture medium (CM)-filled capsules with ECs injected at the injury site. Control rats were in three groups: (1) uninjured, (2)
lesion with transplantation of CM-filled capsules and (3) lesion with transplantation of CM-filled capsules and injections of CM. Three weeks
after injury, Fluororuby was injected into the hindlimb motor and somatosensory cortex to label corticospinal neurons. Observations indicated
that there were a few regenerating fibres, up to 10, in the EC-treated groups. In rats that received encapsulated ECs, regenerating fibres were
present in close association with the capsule. Rats that received EC injections demonstrated a significant increase in the number of collateral
branches from the intact ventral corticospinal tract (vCST) compared with the corresponding control, CM-injected group (P = 0.003), while a
trend for increased collateral branches was observed in rats that received encapsulated ECs (P = 0.07).
D 2003 Elsevier Inc. All rights reserved.
Keywords: Olfactory ensheathing cells; Spinal cord; Regeneration; Corticospinal tract; Collateral sprouting
Introduction
Traumatic injury to the spinal cord triggers a series of
cellular reactions that often result in wide ranging functional
deficits (e.g., reviews by Fawcett and Asher, 1999; Schwab
and Bartholdi, 1996). Within the first few days of injury,
there is massive neuronal and glial cell death (Emery et al.,
1998). Axons distal to the injury site degenerate while
reactive astrocytes divide to form scar tissue, which inhibits
subsequent axonal regrowth by a combination of mechan-
ical and trophic mechanisms (Lindsay, 1986). The injury
site is a complex environment containing reactive astro-
cytes, fibroblasts, immune cells and a mix of growth factors,
inhibitory factors, cytokines and other agents, some soluble
and some associated with the surface of specific cells
(Schwab and Bartholdi, 1996).
0014-4886/$ - see front matter D 2003 Elsevier Inc. All rights reserved.
doi:10.1016/j.expneurol.2003.09.008
* Corresponding author. Discipline of Anatomy and Physiology,
School of Medicine, University of Tasmania, Private Bag 24, Medical
Sciences Building, College Road, Hobart, Tasmania 7001, Australia. Fax:
+61-3-62262679.
E-mail address: [email protected] (M.I. Chuah).
In light of the restrictive nature of the injured central
nervous system, various experimental strategies have been
developed to alter this environment so that axonal repair can
take place. One of these is the use of olfactory ensheathing
cells (ECs). Indeed, ECs injected into the injured spinal cord
have resulted in a remarkable degree of axonal regeneration
and functional recovery (e.g., Imaizumi et al., 1998; Nash et
al., 2002; Ramon-Cueto et al., 1998), including remyelina-
tion of demyelinated axons (Li et al., 1997; Imaizumi et al.,
1998). Perhaps most remarkable is the restoration of climb-
ing ability of rats whose spinal cords have undergone
complete transection (Ramon-Cueto et al., 2000). However,
studies have yet to investigate the mechanisms through
which ECs operate in the novel environment of the injured
spinal cord.
Results of in vitro studies have demonstrated that ECs
express cell surface molecules such as N-CAM, N-cadherin
and laminin (Chuah and Au, 1994; Chuah et al., 1991;
Treloar et al., 1996). They also produce many growth
factors including several forms of neuregulins and some
neurotrophins (Chuah and West, 2002; Chuah et al., 2000;
Woodhall et al., 2001). It is not known whether the

M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–2516
reparative properties of ECs in the spinal cord can be
attributed to these soluble growth factors or whether direct
contact between ECs and host tissue is crucial for spinal
cord repair. In the present study, we have compared the
effect of encapsulated ECs with that of ECs injected directly
into the injured spinal cord. The dorsal corticospinal tract
(dCST) was cut at T8–9 and the extent of repair was
assessed qualitatively and quantitatively by examining seg-
ments of the spinal cord rostral and caudal to the lesion. We
have also investigated the response of the uninjured ventral
corticospinal tract (vCST) to ECs that are encapsulated or
implanted directly into the spinal cord. To our knowledge,
this is the first study to examine the influence of ECs on
collateral sprouting in the spinal cord.
Materials and methods
All procedures involving animals were approved by the
Animal Experimentation Ethics Committee of the Univer-
sity of Tasmania and are consistent with the Australian Code
of Practice for the Care and Use of Animals for Scientific
Purposes.
Ensheathing cell purification and encapsulation
Ensheathing cells were purified and cultured on the basis
of the method of Chuah and Teague (1999) with modifica-
tions (Vincent et al., 2003) to increase the yield of ECs while
maintaining a high level of purity. The olfactory nerve layer
(ONL) and mucosa (OM) were isolated from 3-day-old
hooded Wistar rats euthanised in ice. The ONL was peeled
away from the glomerular and deeper layers of the olfactory
bulb and the OM was scraped from both sides of the nasal
septum after cutting away the surrounding respiratory tissue.
The pooled tissues were digested in MEM HEPES modifi-
cation (MEMH; Sigma, NSW, Australia) containing 0.25%
trypsin (Gibco, NY) and 0.03% collagenase (Sigma) for 15
min at 37jC. This incubation was repeated twice with fresh
solution, then replaced with culture medium (CM) composed
of MEM D-valine modification (Sigma) supplemented with
10% dialysed fetal calf serum (10000 MW cut-off; Sigma)
and penicillin–streptomycin–amphotericin B (Gibco) (Tisay
and Key, 1999). The digested tissue was triturated, filtered
through a sterile nylon gauze (80 Am pore size) and centri-
fuged for 10 min at 500 � g. Resuspended cells were plated
in CM. After 24 h, the culture was treated with 10� 4 M
cytosine-h-D-arabinofuranoside (AraC; Sigma) for 48 h to
minimise the population of fibroblasts. Purified ECs were
enriched in 125 Ag/ml bovine pituitary extract (Sigma) for 2
days before passage.
For encapsulation, ECs were scraped from 25-cm2 flasks
(Sarstedt, NC), centrifuged at 500 � g and resuspended in
CM. By means of a Hamilton syringe, they were introduced
into a polyvinylidene fluoride (PVDF) hollow fibre of 1.0-
mm diameter with a MW cut-off of 300000 Da (Spectrum,
CA). After loading, each end of the fibre was heat sealed to
form a capsule measuring 6–8 mm long and containing
about 105 ECs. Control capsules contained CM alone with
no ECs. All capsules were placed in CM overnight at 37jCin 5% carbon dioxide before transplanting to the spinal cord
the following day.
Hoechst blue-labeling of ensheathing cells
Ensheathing cells that were used for direct injections into
the spinal cord were prelabeled with Hoechst 33342 (Sig-
ma). One day before injection, ECs that were growing in a
25-cm2 flask were incubated in 10 ng/ml Hoechst 33342 in
CM and diluted from a stock of 10 mg/ml in dimethyl
sulfoxide. The cells were incubated for 40 min at 37jC,washed four times in MEMH and returned to fresh CM. A
small sample of cells was scraped off the flask and viewed
under ultraviolet filter to check that the staining had been
successful. Before implantation, the CM was collected to
test whether any unincorporated Hoechst dye had remained
in the solution. A fresh flask of unlabeled cells was
incubated overnight with this medium and checked under
ultraviolet filter. Only labeling of a barely detectable level
was observed. This procedure was done as a precautionary
measure to ascertain that host tissues were not exposed to
excess dye in solution during the process of injecting ECs
into the spinal cord.
Thirty minutes before injection, ECs were scraped from
half the flask, centrifuged at 500 � g and resuspended at a
concentration of about 104 cells/Al CM.
Spinal lesion surgeries
Adult rats weighing 270–380 g were anaesthetised with
3% isoflurane with oxygen, and maintained at 2–2.5%
isoflurane with oxygen at 0.5 l/min throughout the surgery.
Rats were injected subcutaneously with 0.1 ml/kg Rimadyl
(carprofen 50 mg/ml, a nonsteroidal anti-inflammatory and
analgesic; Pfizer Animal Health, UK), intramuscularly with
0.1 ml/kg Norocillin L.A. (procaine penicillin G 150 mg/ml
and benzathine penicillin G 112.5 mg/ml; Norbrook Labo-
ratories, UK) and intraperitoneally with 40 ml/kg Hart-
mann’s saline or 0.9% saline (Baxter Healthcare, NSW,
Australia). Hair from the back was removed with clippers
and skin swabbed with 70% ethanol and povidone-iodine. A
sterile surgical drape was placed over the rat and the skin
over spinous processes of thoracic vertebrae 6–11 (T6–
T11) was incised and retracted. Attachments of muscles to
T7–T10 spinous processes were cut and the muscles
retracted. The spinous processes of T8 and T9 were re-
moved with rongeurs and a laminectomy performed using a
dental drill and rongeurs to expose the dorsal spinal cord.
The dura was incised with microscissors. The dorsal col-
umns, which include the dCST, were cut by lowering
microscissors attached to a stereotaxic arm to a depth of
1.5 mm below the dorsal surface of the spinal cord and

M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–25 17
cutting twice. The width of the cut was 1.5–2.0 mm, centred
on the midline. Experimental trials were first done to
ascertain that this surgical procedure produced a consistent
total transection of the dCST on both sides.
Two groups of rats received injections of ECs (n = 6) or
CM (n = 5) followed by placement of a CM-containing
PVDF capsule during the same lesion surgery. ECs sus-
pended in CM or CM alone were loaded into a 10-Al syringewith 26-s gauge bevelled needle (Hamilton, Reno, NV)
using the ‘‘fast up’’ function of an sp310i syringe pump
(World Precision Instruments, Victoria, Australia) mounted
on a stereotaxic frame. One microlitre of ECs or CM was
injected with the pump set at an injection rate of 1 Al/min at
each of four sites, 0.3 mm to the right and left of the midline
and 2–3 mm anterior and posterior to the injury site. All
injections were 1.0 mm deep to the dorsal surface of the
spinal cord. These coordinates were selected to place the
injected material just lateral to the dCST. Additionally, 4
Al of ECs or CM was injected at 1 Al/min at the injury site
on the midline. After each of the five injections, the needle
was left in place for 1 min. Following the injections, a
PVDF fibre capsule filled with CM was inserted into the
dorsal median sulcus of the spinal cord, centred anteropos-
teriorly on the injury site. Two additional groups of rats
received a PVDF fibre capsule loaded with ECs (n = 5) or
CM (n = 6), but no injections of ECs or CM, during the
same lesion surgery. The capsule was inserted into the
dorsal median sulcus of the spinal cord, centred anteropos-
teriorly on the injury site.
Following the lesion, injections (in two groups) and
capsule placement (in all four groups), the retracted muscles
were sutured with 5–0 silk and the skin closed with Michel
wound clips. The skin was swabbed with povidone-iodine
and the rat returned to its individual home cage under a heat
lamp for 1 h. Rats were injected daily for 3 days following
surgery subcutaneously with 0.1 ml/kg Rimadyl (carprofen
50 mg/ml).
Cortical injections of Fluororuby
To anterogradely label the corticospinal tract, rats were
injected with Fluororuby (Molecular Probes, Eugene, OR)
into the hindlimb motor and somatosensory cortex, which
contains most corticospinal neurons projecting to the lum-
bosacral enlargement of the spinal cord (Li et al., 1990).
Fluororuby injections were performed 3 weeks after spinal
lesion surgeries and also on a group of uninjured rats
weighing 340–365 g (n = 4). Rats were anaesthetised and
injected with Rimadyl, Norocillin L.A. and saline as for the
spinal surgeries. They were placed in a stereotaxic frame
with gas anaesthesia mask (David Kopf Instruments,
Tujunga, CA). Scalp hair was removed with clippers, the
scalp swabbed with 70% ethanol and povidone-iodine, and
incised. Scalp flaps were retracted and fascia cleared from
the skull with cotton tip applicators. The bregma and
lambda were levelled in the horizontal plane by adjusting
the bite bar. Burr holes were drilled in the skull above the
injection sites at the following coordinates relative to the
bregma: 0.9 mm posterior, 2.0 mm lateral; 1.9 mm posterior,
2.4 mm lateral; and 2.9 mm posterior, 2.6 mm lateral, on
both the right and left (coordinates on the basis location of
hindlimb somatosensory cortex by Paxinos and Watson,
1986). Half a microlitre of Fluororuby solution (10%
rhodamine–dextran, MW 10000 in water with 0.5% Triton
X100) was injected at each of the six sites 1.8 mm below the
dura at a rate of 0.5 Al/min using a 10 Al Hamilton syringe
with 33 gauge needle controlled by an sp310i syringe pump.
The needle was left in place for 1 min after each injection.
Burr holes were filled with bone wax and the scalp closed
with suture clips. The skin was swabbed with povidone-
iodine and the rat returned to its individual home cage under
a heat lamp for 30 min. Rats were injected daily for 2 days
following the surgery subcutaneously with 0.1 ml/kg
Rimadyl (carprofen 50 mg/ml).
Histology
Rats were anaesthetised with 0.4 ml Nembutal (200 mg/
kg; Boehringer Ingelheim, NSW, Australia) 2 weeks after
injection of Fluororuby. They were perfused transcardially
with cold phosphate buffered saline (PBS) followed by 4%
paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). After
perfusion, the vertebral column and attached brain were
dissected and placed in cold 4% buffered paraformaldehyde
overnight. The following day, the spinal cord was removed
en bloc from the rest of the vertebral column and cut into
segments at levels indicated in Fig. 1A. The rostral ends of
spinal cord segments were marked with a solution of 15%
Alcian Blue in 50% glycerol to facilitate orientation of
specimens on the cryostat (Jung Frigocut 2800E, Leica).
All segments were stored in buffered 4% paraformaldehyde
at 4jC until ready for sectioning, at which time they were
embedded in Tissue-TekR O.C.T. compound (ProSciTech,
QLD, Australia). The approximately 2.8-cm length of cord
containing the capsule was sectioned (50 Am thickness) in
the sagittal plane while all other sections were made in the
transverse plane at the cervical, upper thoracic and mid-
thoracic level. Sections were floated on water bath to
remove O.C.T. compound and then mounted on slides
coated with 2% 3-aminopropyltriethoxysilane in acetone
(Sigma) (Henderson, 1989). It should be noted that when
the sections were floated, the capsule often became dis-
lodged from the rest of the spinal cord tissue, leaving an
oval-shaped space in the spinal cord section. Sections were
allowed to dry for 1 h in the dark and coverslipped using
DAKO fluorescent mounting medium. To prevent evapora-
tion and drying of sections, nail polish was used to seal the
edges of coverslips.
Histological examinations were performed with a fluo-
rescence attachment mounted on an Olympus BX50 micro-
scope and images captured using an Olympus DP50 digital
camera with the aid of Viewfinder Lite and Studio Lite 1.0

Fig. 1. (A) Dissected brain and spinal cord of rat. Arrows indicate the levels
at which the spinal cord is cut into segments. Transverse sections (50-Amthickness) were made at the cervical (C), upper thoracic (UT), mid-thoracic
(MT) and upper lumbar level (immediately caudal to the bracketed
segment). The bracketed segment contains the lesion and was longitudinally
serially sectioned. Scale bar = 1.5 cm. (B) Transverse section of the spinal
cord at the mid-thoracic level showing the Fluororuby-labeled dorsal
corticospinal tract. Scale bar = 0.8 mm. Inset shows a magnified image of
the fibres of the vCST. Scale bar = 200 Am.
M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–2518
software (Pixera Corporation). To count the number of
dCST fibres in transverse sections, images were enlarged
to A4 size and printed. A plastic transparency was placed
over the fluorescent tract and individual axons were marked
off with a pen as the counting proceeded. Images of vCST at
the lower thoracic level were similarly captured and counted
at a magnification of 450�.
In sagittal sections, an 8-mm length of the spinal cord
caudal to the capsule was used for counting the number of
axonal branches from the vCST fibres. All serial sagittal
sections of each spinal cord segment were examined at
400� magnification, and the number of fibres branching
directly from vCST fibres (as exemplified by branching
shown in Fig. 4H) was counted. Similarly, the number of
terminal branches of axons present in the grey matter was
also counted. Terminal branches appeared characteristically
as an arborisation extending from a single axon and whose
area of innervation was restricted to the grey matter. In the
uninjured animals, counts were conducted at the cor-
responding similar level. All counts were performed by a
person blinded to the treatment group. Statistical evaluation
of counts and comparison among groups were performed
with SigmaStat 2.03 (Jandel Scientific Software Corpora-
tion, San Rafael, CA) and the results presented in Figs. 6A
and B. A one-way ANOVA was applied to detect differ-
ences among groups, followed by multiple pair wise com-
parison (Tukey test). A P value of < 0.05 was considered
statistically significant and data were presented as mean FSEM.
Results
Histology and axonal labeling
Previous studies have shown that corticospinal neurons
are present not only in the motor cortex but also in some
other cortical areas including the supplementary motor area,
prefrontal cortex and somatosensory cortex (Li et al., 1990;
Miller, 1987). On the basis these findings and several
experimental trials, we have found that bilateral injection
of Fluororuby into the hindlimb motor and somatosensory
cortex consistently produced intense labeling of the cortico-
spinal tracts (Fig. 1B). In the uninjured rat, the number of
fibres in the bilateral dCST averaged 339 F 18, 348 F 11
and 284 F 28 at the cervical, upper and middle thoracic
levels, respectively (Fig. 2A). The fibres were present in
compact arrangement as two well-defined tracts. The
smaller component of vCST averaged 55 F 4 fibres bilat-
erally at the mid-thoracic level and fibres were distributed
diffusely over a wide area of the ventral funiculus. As
reported in previous studies (Liang et al., 1991; Weidner
et al., 2001), we also found occasionally a few corticospinal
fibres, usually numbering fewer than six on each side, being
present in the lateral funiculi, just ventral to the dorsal horns.
Axonal counts rostral to the injury site
The number of Fluororuby-labeled dCST axons was
counted in all groups at the cervical, upper and mid-thoracic
levels (Fig. 2A). At the cervical, upper and mid-thoracic
level, there was no statistically significant difference in the
number of dCST fibres in all treatment groups, although a
higher number of fibres in the groups that received ECs
(encapsulated or injected) were present at the mid-thoracic
level. The control group that received CM-containing cap-
sules together with CM injections had the least number of
fibres with 86 F 20 fibres at the mid-thoracic level. This
value translates to 27 F 5% of the number of fibres present
at the cervical level (Fig. 2B). Compared to the other control
group which received only a CM-containing capsule, this

Fig. 2. (A) Graphical presentation of the number of labeled dorsal
corticospinal fibres at the cervical, upper and mid-thoracic level in all
groups. The rats which received EC or CM injections also had culture
medium (CM)-containing capsules placed over the lesion. There was no
statistically significant difference among the treatment groups. (B) The
number of labeled dorsal corticospinal fibres at the upper and mid-thoracic
levels expressed as a percentage of fibres at the cervical level within each
group. Except for the uninjured group, all treatment groups showed a
reduction in the number of dorsal corticospinal fibres from the cervical to
the mid-thoracic level. The data also indicate that many of the corticospinal
neurons still survive 5 weeks post-lesion. Data are presented as mean FSEM.
Fig. 3. Tracing of a typical sagittal section of the spinal cord showing the
cavity which marks the location of the capsule (*) and its spatial orientation
with the dorsal corticospinal tract (dCST). The grey matter constitutes the
middle one-third of the section. The labeled arrows indicate the
approximate locations from which Fig. 4 is obtained. R = rostral, C =
caudal. Scale bar = 2.5 mm.
M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–25 19
group sustained additional acute injury from multiple CM
injections, hence the comparatively fewer fibres that were
labeled. The persistence of Fluororuby-labeled fibres at 5
weeks post-axotomy even in the absence of any cellular
transplants suggests that a considerable proportion of dCST
neurons still survive. This was somewhat surprising given
that it is widely accepted that neurons undergo chromatol-
ysis and eventual cell death following traumatic injury (e.g.,
Schwab and Bartholdi, 1996).
Corticospinal tract fibres near the lesion site
Dissection of the spinal cord at 5 weeks post-surgery
showed that during this time, a layer of fibrous connective
tissue had grown over the capsule, marked by a slight
protrusion on the dorsal surface (Chuah and West, 2002).
Longitudinal sections revealed that the capsule had shifted
ventrally in the spinal cord during the recovery period (Fig.
3) and in some animals had caused some compression of the
vCST. The cut edge of the dCST was visible as a bright red
fluorescent bundle with the axotomised edge directly ap-
posed to the rostral tip of the capsule (Figs. 3 and 4A).
In both control and EC-treated rats, small numbers of
nerve sprouts were observed emanating from the cut stump
and sides of the dCST (Fig. 4B). Small numbers of regener-
ating fibres, up to 10, were present in EC-treated rats. In the
EC-encapsulated rats, the regenerating fibres were observed
in close association with the capsule, along its sides and
caudally as well (Figs. 4C, D and E). These regenerating
fibres were characterised by axonal varicosities and theywere
often closely apposed, running in a parallel direction, typical
of dCST fibres. In rats that received encapsulated ECs, some
nerve fibres were observed to be curving closely around the
caudal edge of the capsule (Fig. 4E), a location and direc-
tional growth that are not associated with the normal trajec-
tory of the dCST. In contrast, in rats that had ECs injected
directly into the spinal cord, fine nerve fibres projected
caudally away from the caudal tip of the CM-containing
capsule (Fig. 4F). The longest of these caudally projecting
fibres extended as far as 10mmdistal to the caudal edge of the
capsule. In summary, the observations indicate that there
were a few regenerating fibres from the sectioned dCST of
rats which had received ECs (encapsulated or injected).
However, regeneration was probably hampered by large areas
of necrosis, macrophages and tissue scarring in both control
and EC-treated rats. It was observed that some of the
regenerating sprouts terminated when confronted by areas
of brightly fluorescent cells, presumably macrophages (Fig.
4G). The small number of regenerating fibres is consistent
with the finding that there were not statistically significant
differences in the number of dCST fibres rostral to the lesion.
In contrast to the dense, compact arrangement of fibres in
the dCST, the brightly labeled vCST is composed of fibres
that are distributed over a wide area in the ventral half of the
spinal cord. In a few animals that received encapsulated or
injected ECs, branches from the vCST caudal to the lesion
extended to the dorsal columns and grew distally in the
space originally occupied by the dCST (Fig. 4H). These
fibres were new collateral sprouts because they did not
exhibit any axonal branches or terminations as they coursed
through the grey matter. In addition, in some EC-treated
(injected or encapsulated) rats, distally projecting fibres in

Fig. 4. Longitudinal sections of spinal cord segment containing lesion and capsule, 2 weeks after injection of Fluororuby. ‘‘capsule’’ marks the cavity which
previously housed a capsule containing ECs. (A) During post-lesion recovery, the capsule has been displaced ventrally in the spinal cord and appears to be in
the path of the sectioned dorsal corticospinal tract. Nerve sprouts projecting from the sides of the tract (asterisk) are visible at higher magnification. Scale bar =
800 Am. (B) Arrow indicates a fibre sprouting from the dorsal corticospinal tract (dCST) in the area marked by asterisk in A. The fibre ends in an arborisation
of fine sprouts. Scale bar = 300 Am. (C) Arrows indicate two fine nerve fibres present in the tissue overlying the EC-containing capsule. Scale bar = 250 Am.
(D) Arrows indicate a nerve fibre running close to the ventral edge of the capsule (C) containing ECs. Scale bar = 700 Am. (E) Regenerating nerve fibres
(arrows) running close to the EC-containing capsule, curving around its caudal edge and sandwiched between the capsule and clusters of brightly fluorescent
cells, presumably macrophages. Scale bar = 180 Am. (F) Section from spinal cord bearing lesion injected with ECs and transplanted with capsule containing the
culture medium. A nerve fibre (arrows) is observed coursing away from the caudal edge of the capsule. Scale bar = 400 Am. (G) Arborising fine nerve sprouts
(arrow) appear to terminate when confronted with aggregates of fluorescent cells, presumably macrophages. Scale bar = 150 Am. (H) Section from a region of
spinal cord caudal to the lesion and which received encapsulated ECs. A collateral branch (arrows) from the ventral corticospinal tract in the ventral funiculus
extends across the grey matter and then projects dorsally in the region where the original dorsal corticospinal tract was located. Scale bar = 600 Am. (I) Similar
section caudal to the lesion from a different rat which received encapsulated ECs. Fibres of the vCST (arrowheads) are clearly observed in the ventral funiculus.
A caudally projecting fibre (arrows) in the dorsal funiculus originates from the fibre network present in the grey matter. Scale bar = 800 Am.
M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–2520

Fig. 4 (continued).
M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–25 21
the dorsal columns, caudal to the lesion, were observed to
originate from a network in the grey matter (Fig. 4I). This
unusual axonal route as described in Figs. 4H and I was not
observed in animals from both control groups.
Although ECs injected directly into the spinal cord were
prelabeled fluorescently with Hoescht blue (Fig. 5A), there
was difficulty in localising the cells 5 weeks posttransplan-
tation. In some animals, ECs remained near the injection site
(Fig. 5B), while in others, very few ECs were present (Fig.
5C). Viable ECs could be distinguished from the yellowish
crystalloid autofluorescence of macrophages (Bunn et al.,
2002; Fig. 6 in Takami et al., 2002) by their blue fluores-
cence under UV stimulation (Fig. 5B, inset). In small areas
where the two types of fluorescence overlapped, it is possible
that the fluorescence could be a consequence of macro-
phages having phagocytosed dead ECs.
Axonal branching from the ventral corticospinal tract
In addition to the number of labeled vCST fibres, axonal
fibres observed to be clearly branching directly from the
vCST (as shown in Fig. 4H) were also counted. Initial
overview showed that there was variability in the proportion
of labeled fibres in the vCST. This is due in part to the fact that
the total number of fibres in this tract is considerably less than
that of the dCST, about 19% and probably also because the
motor neurons are dispersed over a wider cortical region. To
normalize the data, the number of axonal branches and
terminations was expressed as a ratio of the number of labeled
vCST fibres (Figs. 6A and B). ANOVA analysis of the data
showed that there was a statistically significant difference
among treatment groups (P = 0.01). The two control groups
(lesion with transplantation of CM-filled capsule; lesion with
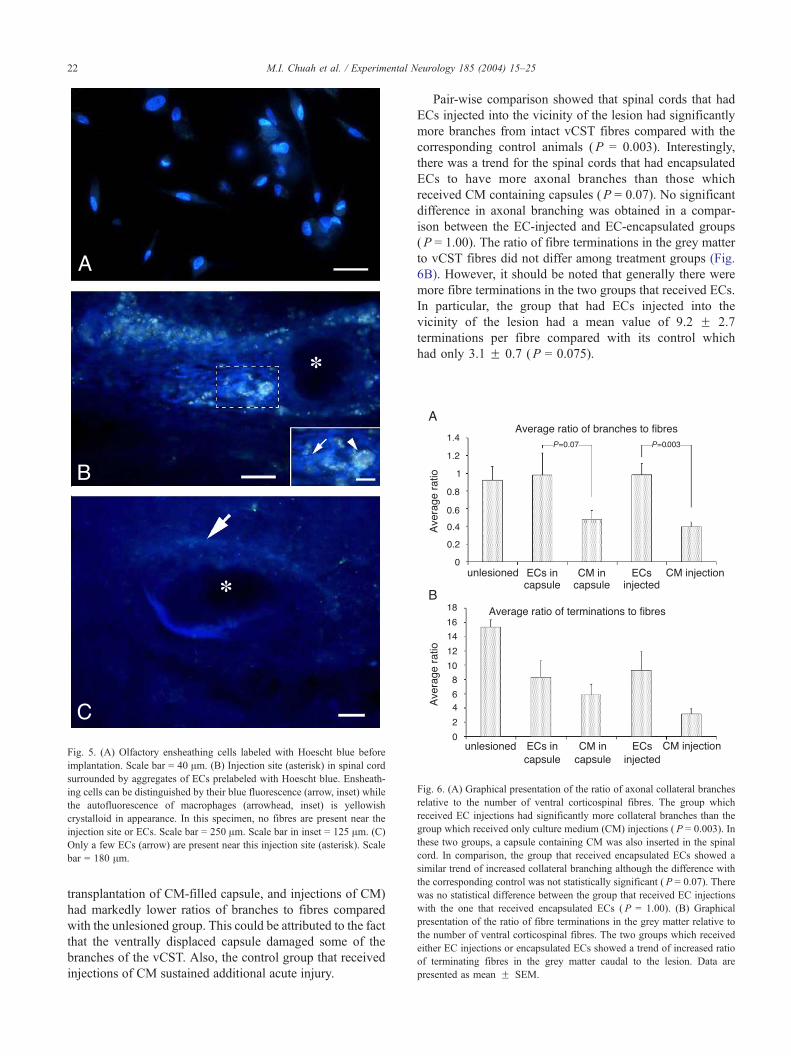
Fig. 5. (A) Olfactory ensheathing cells labeled with Hoescht blue before
implantation. Scale bar = 40 Am. (B) Injection site (asterisk) in spinal cord
surrounded by aggregates of ECs prelabeled with Hoescht blue. Ensheath-
ing cells can be distinguished by their blue fluorescence (arrow, inset) while
the autofluorescence of macrophages (arrowhead, inset) is yellowish
crystalloid in appearance. In this specimen, no fibres are present near the
injection site or ECs. Scale bar = 250 Am. Scale bar in inset = 125 Am. (C)
Only a few ECs (arrow) are present near this injection site (asterisk). Scale
bar = 180 Am.
Fig. 6. (A) Graphical presentation of the ratio of axonal collateral branches
relative to the number of ventral corticospinal fibres. The group which
received EC injections had significantly more collateral branches than the
group which received only culture medium (CM) injections ( P = 0.003). In
these two groups, a capsule containing CM was also inserted in the spinal
cord. In comparison, the group that received encapsulated ECs showed a
similar trend of increased collateral branching although the difference with
the corresponding control was not statistically significant ( P = 0.07). There
was no statistical difference between the group that received EC injections
with the one that received encapsulated ECs ( P = 1.00). (B) Graphical
presentation of the ratio of fibre terminations in the grey matter relative to
the number of ventral corticospinal fibres. The two groups which received
either EC injections or encapsulated ECs showed a trend of increased ratio
of terminating fibres in the grey matter caudal to the lesion. Data are
presented as mean F SEM.
M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–2522
transplantation of CM-filled capsule, and injections of CM)
had markedly lower ratios of branches to fibres compared
with the unlesioned group. This could be attributed to the fact
that the ventrally displaced capsule damaged some of the
branches of the vCST. Also, the control group that received
injections of CM sustained additional acute injury.
Pair-wise comparison showed that spinal cords that had
ECs injected into the vicinity of the lesion had significantly
more branches from intact vCST fibres compared with the
corresponding control animals (P = 0.003). Interestingly,
there was a trend for the spinal cords that had encapsulated
ECs to have more axonal branches than those which
received CM containing capsules (P = 0.07). No significant
difference in axonal branching was obtained in a compar-
ison between the EC-injected and EC-encapsulated groups
(P = 1.00). The ratio of fibre terminations in the grey matter
to vCST fibres did not differ among treatment groups (Fig.
6B). However, it should be noted that generally there were
more fibre terminations in the two groups that received ECs.
In particular, the group that had ECs injected into the
vicinity of the lesion had a mean value of 9.2 F 2.7
terminations per fibre compared with its control which
had only 3.1 F 0.7 (P = 0.075).

M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–25 23
Discussion
We have compared the effect of encapsulated versus
injected ECs on collateral axonal sprouting and regeneration
in the injured spinal cord. We have shown for the first time
that collateral axonal branching from the intact vCST is
significantly increased in the presence of ECs. A small
number of regenerating fibres were present in the two
groups of rats which received ECs (encapsulated and
injected), consistent with results that have been reported
previously by other research groups (Ramon-Cueto et al.,
2000; Takami et al., 2002).
Response of vCST to injury in the dorsal columns
In this study, ECs transplanted into the spinal cord
following injury to dCST resulted in increased collateral
branches from the vCST. These new branches either termi-
nated in the grey matter or extended dorsally and caudally
along the original path of the dCST. The response of vCST
to injury in the dCST has generally been overlooked in EC-
transplantation studies, but this observation of collateral
sprouting is particularly important in the accurate interpre-
tation of results. Unless nerve fibres are traced to their
origin, their mere presence in the dorsal column caudal to a
dCST lesion may not be solely attributed to regenerating
fibres. It is possible that previous studies which reported the
presence of regenerating dCST fibres following EC trans-
plantation could have included axons from vCST in their
observations (Li et al., 1998; Nash et al., 2002; Takami et
al., 2002). Li et al. (1998) labeled CST axons by injecting
biotin dextran either into the medullary pyramid or the entire
contralateral sensorimotor cortex. Such an extensive injec-
tion would most likely have labeled the vCST. In a recent
study in which the thoracic spinal cord was contused, a
quantitative analysis was done on the number of fibres in the
dorsal column rostral to and near the lesion (Takami et al.,
2002). Although the authors inferred that the presence of the
fibres near the lesion is indicative of regeneration, they also
conceded that some could be originating from the spared
vCST. Unless a total transection of the spinal cord is
performed, the neuroanatomical contribution of collateral
sprouting to fibres caudal to the lesion cannot be entirely
discounted.
Similarly, the contribution of collateral sprouting from
descending motor tracts to functional recovery should not be
underestimated (Murray and Goldberger, 1974). An early
report has suggested limited sprouting from mid-brain
derived tracts following thoracic spinal cord sectioning
(Prendergast and Misantone, 1980). More recently, the
sparing of a few fibres in the ventral funiculus of the rat
spinal cord was sufficient to significantly improve stepping
abilities in locomotion (Schucht et al., 2002). A study on
adult rats reported that spontaneous sprouting from the
vCST following lesion of the dCST correlated with the rats’
ability to retrieve food pellets. If the vCSTwas subsequently
injured, functional recovery was abolished (Weidner et al.,
2001).
Influence of ECs on the response of the vCST
Although a statistically significant increase in collateral
sprouting was only observed in rats that had ECs injected
directly into the injury site, those that received encapsulated
ECs demonstrated a trend for increased branching (P =
0.07). When the group that received EC injections was
compared with the one that received encapsulated ECs,
there was no clear statistically significant difference between
them (P = 1.00). The promotion of collateral sprouting by
diffusible growth factors is supported by evidence from
numerous studies on different types of neurons (Cohen-Cory
and Fraser, 1995; Gallo and Letourneau, 1998; He et al.,
1992; Schnell et al., 1994). The peculiar course of a few
regenerating fibres closely enveloping the caudal end of a
capsule containing ECs, as shown in Fig. 4E, is suggestive
of an affinity between the nerve fibres and capsule. Whether
diffusible factors are involved in inducing this behaviour
will need to be confirmed in future experiments.
Other controlling mechanisms could account for our
observations. The fact that only the group that received
EC-injections demonstrated a statistically significant in-
crease in the ratio of branches when compared with their
corresponding control group suggests that injected ECs
could have altered the inflammatory response at the
injury site. One possibility is by suppressing the expres-
sion of inhibitory molecules such as chondroitin sulphate
proteoglycan.
Developmental studies have shown that an increase in
the polysialic acid level of the neural cell adhesion molecule
(NCAM) on CST axons coincides with the initiation of
collateral branching (Daston et al., 1996; Joosten et al.,
1996). The cell adhesion molecule NCAM is expressed by
ECs both in vivo and in vitro (Chuah and Au, 1993;
Miragall et al., 1988). It is possible that in our study,
NCAM expressed by encapsulated ECs was isolated within
the capsule, and hence lesioned fibres were subjected to
diminished stimulation, thus accounting for the less exuber-
ant collateral branching. Preliminary scanning electron mi-
croscopy of encapsulated ECs retrieved from the spinal cord
shows that ECs appear to be embedded within a mucoid
material, suggesting that secreted extracellular matrix may
be trapped within the capsule and cannot be accessed by
regenerating fibres.
Survival of ECs in the spinal cord
Although the exact extent of EC survival in the injured
spinal cord has yet to be quantified, several studies (Nash et
al., 2002; Ramon-Cueto et al., 1998, 2000) have shown that
ECs are able to survive in the spinal cord over a long period,
up to 8 months after implantation (Ramon-Cueto et al.,
2000). In this study, it is expected that some ECs will have

M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–2524
died in the spinal cord by 5 weeks post-implantation.
However, it is unlikely that most of them had been phago-
cytosed by macrophages. In our study, autofluorescence of
macrophages could be clearly distinguished from Hoechst-
labeled ECs, and for most instances these two types of
fluorescence did not overlap. Most recently, DeLucia et al.
(2003) showed that a cell line of Hoescht-labeled ECs
remained viable and was not phagocytosed by macrophages
or microglia 2 weeks following transplantation.
In recent experiments on encapsulated ECs in the spinal
cord, we have found that the ECs survive in the capsules
and continue to express growth-promoting molecules such
as neuregulins (Woodhall et al., 2003). Real time quantita-
tive RT-PCR performed on encapsulated ECs retrieved from
the spinal cord showed that the expression profile of the
various isoforms of neuregulins changed in response to the
novel CNS environment.
One possible reason why it was difficult to localize ECs
injected into the spinal cord may be that ECs continued to
proliferate, and hence the fluorescent label had been pro-
gressively diluted. Preliminary experiments in our laborato-
ry show that ECs undergo increased proliferation when
exposed to spinal cord tissue. Another reason is that the
conditions influencing migration of ECs in the spinal cord
are not clearly understood. It has been reported that ECs
tended to migrate preferentially to the vicinity of the lesion,
suggesting that they may be responding to signals from
injured neurons (Boruch et al., 2002). However, in a study
in which the medial longitudinal fasciculus was transected,
ECs migrated preferentially away from the lesion (Gudino-
Cabrera et al., 1999). The resolution of the fate and
migratory behaviour of ECs in the spinal cord is particularly
important if they are to be developed into a clinically
relevant therapeutic agent.
Acknowledgments
This work was funded by the Motor Accident and
Insurance Board (MAIB) of Tasmania, International In-
stitute for Research in Paraplegia and the National Health
and Medical Research Council. We thank Dr Jim Stanko-
vich for advice on statistical analysis.
References
Boruch, A.V., Conners, J.J., Pipitone, M., Deadwyler, G., Storer, P.D.,
Devries, G.H., Jones, K.J., 2002. Neurotrophic and migratory properties
of an olfactory ensheathing cell line. Glia 33, 225–229.
Bunn, H.J., Woltmann, G., Grigg, J., 2002. Applicability of laser scanning
cytometry to study paediatric alveolar macrophages. Eur. Respir. J. 20,
1437–1443.
Chuah, M.I., Au, C., 1993. Cultures of ensheathing cells from neonatal rat
olfactory bulbs. Brain Res. 601, 213–220.
Chuah, M.I., Au, C., 1994. Olfactory cell cultures on ensheathing cell
monolayers. Chem. Senses 19, 25–34.
Chuah, M.I., Teague, R., 1999. Basic fibroblast growth factor in the pri-
mary olfactory pathway: mitogenic effect on ensheathing cells. Neuro-
science 88, 1043–1050.
Chuah, M.I., West, A.K., 2002. Cellular and molecular biology of en-
sheathing cells. Microsc. Res. Tech. 58, 216–227.
Chuah, M.I., David, S., Blaschuk, O., 1991. Differentiation and survival of
rat olfactory epithelial neurons in dissociated cell culture. Dev. Brain
Res. 60, 123–132.
Chuah, M.I., Cossins, J.-M., Woodhall, E., Tennant, R., Nash, G., West,
A.K., 2000. Glial Growth Factor 2 induces proliferation and structural
changes in ensheathing cells. Brain Res. 857, 265–274.
Cohen-Cory, S., Fraser, S.E., 1995. Effects of brain-derived neurotrophic
factor on optic axon branching and remodelling in vivo. Nature 378,
192–196.
Daston, M.M., Bastmeyer, M., Rutishauser, U., O’Leary, D.D.M., 1996.
Spatially restricted increase in polysialic acid enhances corticospinal
axon branching related to target recognition and innervation. J. Neuro-
sci. 16, 5497–5499.
DeLucia, T.A., Conners, J.J., Brown, T.J., Cronin, C.M., Khan, T., Jones,
K.J., 2003. Use of a cell line to investigate olfactory ensheathing cell-
enhanced axonal regeneration. Anat. Rec. 271B, 61–70.
Emery, E., Aldana, P., Bunge, M.B., Puckett, W., Srinivasan, A., Keane,
R.W., Bethea, J., Levi, A.D., 1998. Apoptosis after traumatic human
spinal cord injury. J. Neurosurg. 89, 911–920.
Fawcett, J.W., Asher, R.A., 1999. The glial scar and central nervous system
repair. Brain Res. Bull. 49, 377–391.
Gallo, G., Letourneau, P.C., 1998. Localised sources of neurotrophins ini-
tiate axon collateral sprouting. J. Neurosci. 18, 5403–5414.
Gudino-Cabrera, G., Pastor, A.M., de la Cruz, R.R., Delgado-Garcia, J.M.,
Nieto-Sampedro, M., 1999. Limits to the capacity of transplants of
olfactory glia to promote axonal regrowth in the CNS. NeuroReport
11, 467–471.
He, Y., Yao, Z., Gu, Y., Kuang, G., Chen, Y., 1992. Nerve growth factor
promotes collateral sprouting of cholinergic fibres in the septohippo-
campal cholinergic system of aged rats with fimbria transection. Brain
Res. 586, 27–35.
Henderson, C., 1989. Aminoalkylsilane: an inexpensive simple preparation
for slide adhesion. J. Histotechnol. 12, 123–124.
Imaizumi, T., Lankford, K.L., Waxman, S.G., Greer, C.A., Kocsis, J.D.,
1998. Transplanted olfactory ensheathing cells remyelinate and enhance
axonal conduction in the demyelinated dorsal columns of the rat spinal
cord. J. Neurosci. 18, 6176–6185.
Joosten, E.A.J., Reshilov, L.N., Gispen, W.H., Bar, P.R., 1996. Embryonic
form of N-CAM and development of the rat corticospinal tract: immu-
no-electron microscopical localization during spinal white matter in-
growth. Dev. Brain Res. 94, 99–105.
Li, X.G., Florence, S.L., Kaas, J.H., 1990. Areal distributions of cortical
neurons projecting to different levels of the caudal brain stem and spinal
cord in rats. Somatosens. Motor Res. 7, 315–335.
Li, Y., Field, P.M., Raisman, G., 1997. Repair of adult rat corticospinal tract
by transplants of olfactory ensheathing cells. Science 277, 2000–2002.
Li, Y.L., Field, P.M., Raisman, G., 1998. Regeneration of adult rat cortico-
spinal axons induced by transplanted olfactory ensheathing cells. J.
Neurosci. 18, 10514–10524.
Liang, L., Moret, V., Wiesendanger, M., Rouiller, E., 1991. Corticomoto-
neuronal connections in the rat: evidence from double-labeling of mo-
toneurons and corticospinal axon arborizations. J. Comp. Neurol. 311,
356–366.
Lindsay, R.M., 1986. Reactive gliosis. In: Fedoroff, S., Vernadakis, A.
(Eds.), Astrocytes: Cell Biology and Pathology of Astrocytes. Academ-
ic Press, Orlando, pp. 231–262.
Miller, M.W., 1987. The origin of corticospinal projection neurons in rat.
Exp. Brain Res. 67, 339–351.
Miragall, F., Kadmon, G., Husmann, M., Schachner, M., 1988. Expres-
sion of cell adhesion molecules in the olfactory system of the adult
mouse: presence of the embryonic form of N-CAM. Dev. Biol. 129,
516–531.
Murray, M., Goldberger, M.E., 1974. Restitution of function and collateral

M.I. Chuah et al. / Experimental Neurology 185 (2004) 15–25 25
sprouting in the cat spinal cord: the partially hemisected animal. J.
Comp. Neurol. 158, 19–36.
Nash, H.H., Borke, R.C., Anders, J.J., 2002. Ensheathing cells and methyl-
prednisolone promote axonal regeneration and functional recovery in
the lesioned adult rat spinal cord. J. Neurosci. 22, 7111–7120.
Paxinos, G., Watson, C., 1986. The Rat Brain in Stereotaxic Coordinates,
second ed. Academic Press, Sydney.
Prendergast, J., Misantone, L.J., 1980. Sprouting by tracts descending from
the midbrain to the spinal cord: the result of thoracic funiculotomy in
the newborn, 21-day-old, and adult rat. Exp. Neurol. 69, 458–480.
Ramon-Cueto, A., Plant, G.W., Avila, J., Bunge, M.B., 1998. Long-distance
axonal regeneration in the transected adult rat spinal cord is promoted by
olfactory ensheathing glia transplants. J. Neurosci. 18, 3803–3815.
Ramon-Cueto, A., Cordero, M.I., Santos-Benito, F.F., Avila, J., 2000.
Functional recovery of paraplegic rats and motor axon regeneration in
their spinal cords by olfactory ensheathing glia. Neuron 25, 425–435.
Schnell, L., Schneider, R., Kolbeck, R., Barde, Y., Schwab, M.E., 1994.
Neurotrophin-3 enhances sprouting of corticospinal tract during devel-
opment and after adult spinal cord lesion. Nature 367, 170–173.
Schucht, P., Raineteau, O., Schwab, M.E., Fouad, K., 2002. Anatomical
correlates of locomotor recovery following dorsal and ventral lesions of
the rat spinal cord. Exp. Neurol. 176, 143–153.
Schwab, M.E., Bartholdi, D., 1996. Degeneration and regeneration of ax-
ons in the lesioned spinal cord. Physiol. Rev. 76, 319–370.
Takami, T., Oudega, M., Bates, M.L., Wood, P.M., Kleitman, N.,
Bunge, M.B., 2002. Schwann cell but not olfactory ensheathing glia
transplants improve hindlimb locomotor performance in the moder-
ately contused adult rat thoracic spinal cord. J. Neurosci. 22,
6670–6681.
Tisay, K.T., Key, B., 1999. The extracellular matrix modulates olfactory
neurite growth on ensheathing cells. J. Neurosci. 19, 9890–9899.
Treloar, H.B., Nurcombe, V., Key, B., 1996. Expression of extracellular
matrix molecules in the embryonic rat olfactory pathway. J. Neurobiol.
31, 41–55.
Vincent, A.J., West, A.K., Chuah, M.I., 2003. Morphological plasticity of
ensheathing cells is regulated by cAMP and endothelin-1. Glia 41,
393–403.
Weidner, N., Ner, A., Salimi, N., Tuszynski, M.H., 2001. Spontane-
ous corticospinal axonal plasticity and functional recovery after
adult central nervous system injury. Proc. Natl. Acad. Sci. 98,
3513–3518.
Woodhall, E., West, A.K., Chuah, M.I., 2001. Cultured olfactory ensheath-
ing cells express Nerve Growth Factor, Brain-derived Neurotrophic
Factor, Glia Cell Line-derived Neurotrophic Factor, and their receptors.
Mol. Brain Res. 88, 203–213.
Woodhall, E., West, A.K., Vickers, J.C., Chuah, M.I., 2003. Olfactory
ensheathing cell phenotype following implantation in the lesioned spi-
nal cord. Cell. Mol. Life Sci. 60, 2241–2253.




















