
Methionine Aminopeptidases from Mycobacterium tuberculosis as Novel Antimycobacterial Targets
Upload
independentCategory
view
0download
0
Novel organization of genes in a phthalatedegradation operon of Mycobacterium vanbaaleniiPYR-1
Robin L. Stingley,1 Barbara Brezna,1,2 Ashraf A. Khan1
and Carl E. Cerniglia1
Correspondence
Ashraf A. Khan
1Division of Microbiology, National Center for Toxicological Research, US Food and DrugAdministration, Jefferson, AR 72079, USA
2Institute of Molecular Biology, Slovak Academy of Sciences, 845 51 Bratislava, Slovakia
Received 20 April 2004
Revised 25 June 2004
Accepted 12 August 2004
Mycobacterium vanbaalenii PYR-1 is capable of degrading polycyclic aromatic hydrocarbons
(PAHs) to ring cleavage metabolites. This study identified and characterized a putative phthalate
degradation operon in the M. vanbaalenii PYR-1 genome. A putative regulatory protein (phtR)
was encoded divergently with five tandem genes: phthalate dioxygenase large subunit (phtAa),
small subunit (phtAb), phthalate dihydrodiol dehydrogenase (phtB), phthalate dioxygenase
ferredoxin subunit (phtAc) and phthalate dioxygenase ferredoxin reductase (phtAd ). A 6?7 kb
EcoRI fragment containing these genes was cloned into Escherichia coli and converted phthalate
to 3,4-dihydroxyphthalate. Homologues to the operon region were detected in a number of
PAH-degrading Mycobacterium spp. isolated from various geographical locations. The operon
differs from those of other Gram-positive bacteria in both the placement and orientation of
the regulatory gene. In addition, the M. vanbaalenii PYR-1 pht operon contains no decarboxylase
gene and none was identified within a 37 kb region containing the operon. This study is the
first report of a phthalate degradation operon in Mycobacterium spp.
INTRODUCTION
Phthalates are a class of widely used industrial compoundsknown technically as dialkyl or alkyl aryl esters of 1,2-benzenedicarboxylic acid. The chemicals are used in themanufacture of many products, including plastics, lubri-cants, solvents and cosmetics (Graham, 1973; Peakall, 1975).As a result of this extensive use, phthalates are commonlyfound at low levels in the environment and humans areexposed to phthalates in consumer products, diet andmedical treatments (Cadogan et al., 1993; Cobellis et al.,2003; Duty et al., 2003; Tickner et al., 2001). Althoughphthalates used in fragrances and cosmetic products do notappear to pose significant health risks (Api, 2001), thosefound in medical devices containing polyvinyl chloride(PVC) may be linked to a number of possible adverse healtheffects in the liver, reproductive tract, kidneys, lungs and
heart (Cobellis et al., 2003; Duty et al., 2003; Tickner et al.,2001).
Because phthalates are widely used, they have undergoneextensive testing to determine their environmental fate.Generally they do not persist in the environment and biode-grade rapidly. Phthalate is an intermediate in the biodegrada-tion pathways of some polycyclic aromatic hydrocarbons(PAHs), including pyrene (Heitkamp et al., 1988b), phenan-threne (Barnsley, 1983; Kiyohara & Nagao, 1978; Moodyet al., 2001), fluorene (Grifoll et al., 1994) and fluoranthene(Kelley et al., 1993; Sepic et al., 1998). The phthalatedegradation pathway of Gram-positive bacteria involvesoxygenation to form 3,4-dihydro-3,4-dihydroxyphthalate,dehydrogenation to 3,4-dihydroxyphthalate and finallydecarboxylation to form protocatechuate (Eaton, 2001;Habe et al., 2003) (Fig. 1). The pathway in Gram-negativebacteria progresses through oxygenation and dehydrogena-tion at carbons 4 and 5 to form 4,5-dihydroxyphthalate,with subsequent decarboxylation to form protocatechuate(Chang & Zylstra, 1998; Nomura et al., 1992).
Mycobacterium vanbaalenii PYR-1 is capable of degrading anumber of aromatic hydrocarbons, such as anthracene,phenanthrene, pyrene, biphenyl, benzo[a]pyrene and 7,12-dimethylbenz[a]anthracene (Heitkamp et al., 1988b; Khanet al., 2002; Moody et al., 2001, 2002, 2003, 2004). Based on
Abbreviations: PAH, polycyclic aromatic hydrocarbon; PFGE, pulsedfield gel electrophoresis; TMS, trimethylchlorosilane.
The GenBank/EMBL/DDBJ accession numbers for the sequencesreported in this paper are AY365117 (M. vanbaalenii PYR-1 pht operonregion), AY372761 and AY372763 (Mycobacterium sp. PAH2.135phtAa and phtB PCR products, respectively), and AY372762 andAY372764 (M. flavescens PYR-GCK phtAa and phtB PCR products,respectively).
0002-7263 G 2004 SGM Printed in Great Britain 3749
Microbiology (2004), 150, 3749–3761 DOI 10.1099/mic.0.27263-0

the degradation of various PAHs by M. vanbaalenii PYR-1,this species is likely to have multiple monooxygenases anddioxygenases to perform the initial steps in degradationpathways, in addition to a number of other enzymes thatperform subsequent aromatic ring cleavage steps in thepathways (Heitkamp et al., 1988b; Moody et al., 2003). Twogenes were characterized previously, nidA and nidB, whichencode large and small subunits, respectively, of a polycyclicaromatic ring dioxygenase in M. vanbaalenii PYR-1 (Khanet al., 2001). The products of these genes share limitedsequence homology with known dioxygenases, includingPhdA and PhdB of Nocardioides sp. KP7, and have
conserved homologues in a number of Mycobacteriumspecies that degrade PAHs (Brezna et al., 2003).
M. vanbaalenii PYR-1 nidA and nidB are clustered on thegenome with an aldehyde dehydrogenase, nidD (Khan et al.,2001). Genes encoding enzymes involved in PAH degrada-tion are clustered in a similar manner in other bacterialspecies, including Arthrobacter keyseri 12B, Terrabacter sp.DBF63 and Nocardioides sp. KP7 (Eaton, 2001; Habe et al.,2003; Nojiri et al., 2002; Saito et al., 1999, 2000). The pre-sent study focuses on molecular cloning, sequencing andcharacterization of additional genes in the nidDBA regionof the M. vanbaalenii PYR-1 genome. Approximately 9 kbupstream of the nidDBA BamHI fragment, we have found aputative operon containing genes that encode enzymesinvolved in the degradation of phthalate. Since phthalatehas been isolated as an intermediate metabolite in thedegradation of pyrene, phenanthrene and fluoranthene inM. vanbaalenii PYR-1 (Heitkamp et al., 1988b; Kelley et al.,1993; Moody et al., 2001, 2002, 2003, 2004), we wanted tocharacterize the genes involved in phthalate degradation.This is the first description of the operon involved in thephthalate degradation pathway from Mycobacterium spp.The operon is conserved in several PAH-degrading Myco-bacterium spp. isolated from various geographical locations(Table 1).
METHODS
Bacterial strains and genomic DNA isolation. Bacterial strainsare listed in Table 1. Mycobacterium vanbaalenii PYR-1 was grownon Middlebrook 7H10 agar at 25 uC for 4 days. Cells were scrapedfrom the plates into 1 ml H2O and pelleted by centrifugation for10 min at 5900 g. Genomic DNA was isolated according to theprotocol for Gram-positive bacteria with the Qiagen DNeasy Kit.Additional strains (Table 1) were cultivated on mineral basal salt(MBS) medium supplemented with sorbitol and phenanthrene asdescribed previously (Brezna et al., 2003).
Cloning, sequencing and sequence analyses. A genomiclibrary was previously prepared in pCC1FOS (Epicentre) and probedfor nidA (Stingley et al., 2004). One clone, pFOS608, contained aputative phthalate degradation operon approximately 9 kb upstreamof nidA. A 6?7 kb EcoRI fragment from pFOS608 containing the phtoperon region, including the putative regulatory protein, was clonedinto pGEM-11z f(+) (pPHT) and introduced into E. coli JM109.
DNA sequencing was performed on an Applied Biosystems model 377DNA sequencer at the University of Arkansas for Medical Sciences.Sequences were compiled, translated and analysed using Lasergenesoftware (DNASTAR) and compared to similar genes and proteinsusing BLAST and Conserved Domain Database searches (Marchler-Bauer et al., 2003). Phylogenetic analyses were performed with puta-tive amino acid sequences by using PHYLIP, version 3.572c. Thepercentage confidence was estimated by a bootstrap analysis with 1000replications.
Screening Mycobacterium spp. and Rhodococcus spp. forpht genes. PCR was performed using genomic DNA as templatewith Taq DNA polymerase and supplied PCR solutions according tothe manufacturer’s instructions (Qiagen). The PCR regime consistedof a 3 min preincubation at 95 uC, 30 cycles of 30 s denaturationat 94 uC, 30 s annealing at 55 uC and 1 min extension at 72 uC,
Fig. 1. Schematic representation of the phthalate degradationpathway in Gram-positive bacteria. Genes identified in M.
vanbaalenii PYR-1 that encode enzymes along the pathway areindicated.
3750 Microbiology 150
R. L. Stingley and others
followed by a final hold at 72 uC for 7 min. All primers used arelisted in Table 2. Primers phtAa-2 and phtAa-3 were used to detectphtAa, primers phtB-1 and phtB-3 to detect phtB, and primersphtAa-1 and phtAd-1 to detect a 3?1 kb region of the pht operon,which includes part of phtAa, spans through phtAb, phtB, phtAc andends inside phtAd.
Hybridizations and confirmation of PCR screening. M. van-baalenii PYR-1 fosmid DNA BamHI restriction digests wereseparated on agarose gels stained with ethidium bromide for visuali-zation, then transferred and cross-linked to positively charged nylon
membranes (Roche). The resulting blots were incubated at 65 uCfor at least 30 min in DIG Easy Hyb (Roche) prior to addition
of digoxigenin (DIG)-labelled DNA probes (denatured at 95 uCfor 10 min) specific for nidA. Hybridization was carried out at
65 uC for 16–18 h. DIG-labelled probe was detected with alkaline
phosphate-conjugated anti-DIG antibody (Roche) and the chemi-
luminescent substrate disodium 3-(4-methoxyspiro(1,2-dioxetane-
3,29-(59-chloro)tricyclo[3.3.1]decan)-4-yl)phenyl phosphate (CSPD;
Roche).
PCR products were analysed by ethidium-bromide-stained agarose
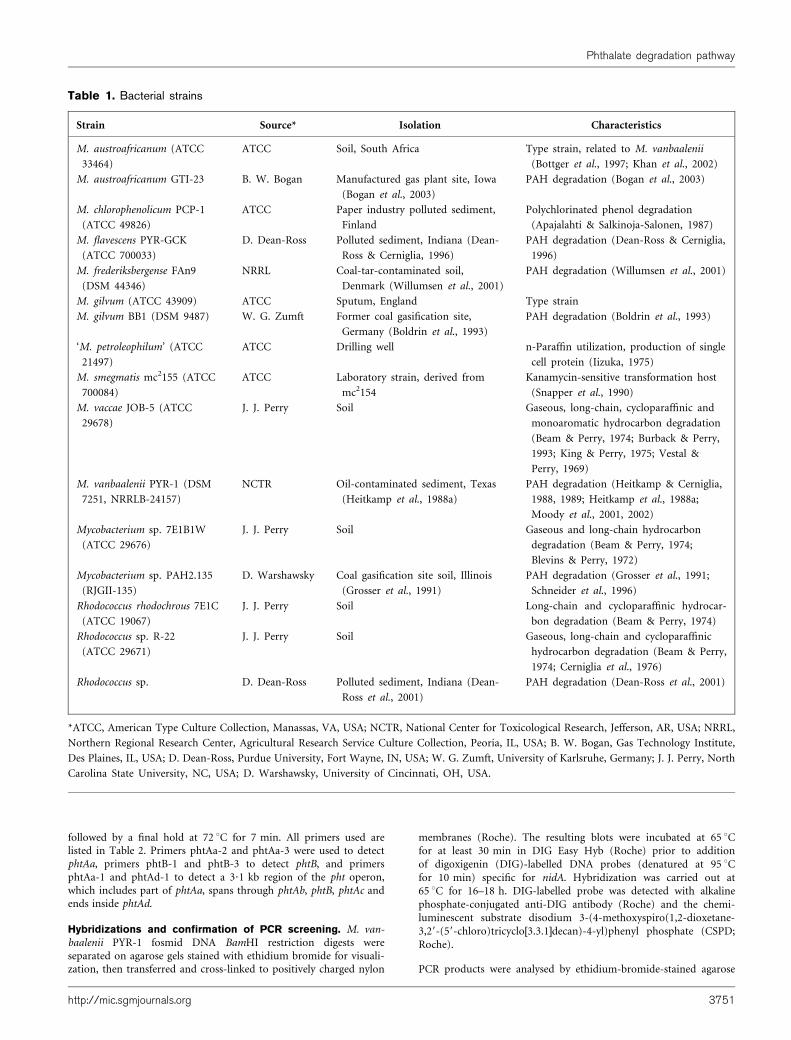
Table 1. Bacterial strains
Strain Source* Isolation Characteristics
M. austroafricanum (ATCC
33464)
ATCC Soil, South Africa Type strain, related to M. vanbaalenii
(Bottger et al., 1997; Khan et al., 2002)
M. austroafricanum GTI-23 B. W. Bogan Manufactured gas plant site, Iowa
(Bogan et al., 2003)
PAH degradation (Bogan et al., 2003)
M. chlorophenolicum PCP-1
(ATCC 49826)
ATCC Paper industry polluted sediment,
Finland
Polychlorinated phenol degradation
(Apajalahti & Salkinoja-Salonen, 1987)
M. flavescens PYR-GCK
(ATCC 700033)
D. Dean-Ross Polluted sediment, Indiana (Dean-
Ross & Cerniglia, 1996)
PAH degradation (Dean-Ross & Cerniglia,
1996)
M. frederiksbergense FAn9
(DSM 44346)
NRRL Coal-tar-contaminated soil,
Denmark (Willumsen et al., 2001)
PAH degradation (Willumsen et al., 2001)
M. gilvum (ATCC 43909) ATCC Sputum, England Type strain
M. gilvum BB1 (DSM 9487) W. G. Zumft Former coal gasification site,
Germany (Boldrin et al., 1993)
PAH degradation (Boldrin et al., 1993)
‘M. petroleophilum’ (ATCC
21497)
ATCC Drilling well n-Paraffin utilization, production of single
cell protein (Iizuka, 1975)
M. smegmatis mc2155 (ATCC
700084)
ATCC Laboratory strain, derived from
mc2154
Kanamycin-sensitive transformation host
(Snapper et al., 1990)
M. vaccae JOB-5 (ATCC
29678)
J. J. Perry Soil Gaseous, long-chain, cycloparaffinic and
monoaromatic hydrocarbon degradation
(Beam & Perry, 1974; Burback & Perry,
1993; King & Perry, 1975; Vestal &
Perry, 1969)
M. vanbaalenii PYR-1 (DSM
7251, NRRLB-24157)
NCTR Oil-contaminated sediment, Texas
(Heitkamp et al., 1988a)
PAH degradation (Heitkamp & Cerniglia,
1988, 1989; Heitkamp et al., 1988a;
Moody et al., 2001, 2002)
Mycobacterium sp. 7E1B1W
(ATCC 29676)
J. J. Perry Soil Gaseous and long-chain hydrocarbon
degradation (Beam & Perry, 1974;
Blevins & Perry, 1972)
Mycobacterium sp. PAH2.135
(RJGII-135)
D. Warshawsky Coal gasification site soil, Illinois
(Grosser et al., 1991)
PAH degradation (Grosser et al., 1991;
Schneider et al., 1996)
Rhodococcus rhodochrous 7E1C
(ATCC 19067)
J. J. Perry Soil Long-chain and cycloparaffinic hydrocar-
bon degradation (Beam & Perry, 1974)
Rhodococcus sp. R-22
(ATCC 29671)
J. J. Perry Soil Gaseous, long-chain and cycloparaffinic
hydrocarbon degradation (Beam & Perry,
1974; Cerniglia et al., 1976)
Rhodococcus sp. D. Dean-Ross Polluted sediment, Indiana (Dean-
Ross et al., 2001)
PAH degradation (Dean-Ross et al., 2001)
*ATCC, American Type Culture Collection, Manassas, VA, USA; NCTR, National Center for Toxicological Research, Jefferson, AR, USA; NRRL,
Northern Regional Research Center, Agricultural Research Service Culture Collection, Peoria, IL, USA; B. W. Bogan, Gas Technology Institute,
Des Plaines, IL, USA; D. Dean-Ross, Purdue University, Fort Wayne, IN, USA; W. G. Zumft, University of Karlsruhe, Germany; J. J. Perry, North
Carolina State University, NC, USA; D. Warshawsky, University of Cincinnati, OH, USA.
http://mic.sgmjournals.org 3751
Phthalate degradation pathway

gels and transferred to positively charged nylon membranes (Roche),
then subjected to Southern hybridization with the following
internal DIG-labelled probes: oligoprobe phtAa-4 (Table 2) for phtAa
and oligoprobe phtB-2 (Table 2) for phtB and phtAaAbBAcAd
PCR products. Oligoprobes were prepared with the DIG oligo-
nucleotide 39-end labelling kit (Roche). Hybridization was at 30 uCand washes with 0?56 SSC, 0?1% SDS were at 48 uC for phtAa and
55 uC for phtB and phtAaAbBAcAd. The phtAa and phtB PCR
products from Mycobacterium sp. strain PAH2.135 were confirmed by
sequencing.
To screen bacterial strains for phtAa, total genomic DNA was
embedded in agarose plugs, digested by XbaI and separated by
pulsed field gel electrophoresis (PFGE) as described previously (Brezna
et al., 2003). Restriction fragments were separated at 14 uC in a 1%
agarose gel in 0?56 TBE buffer using a contour-clamped homogen-
eous electric field (CHEF) apparatus (CHEF-DR II, Bio-Rad). Electro-
phoresis was performed at 6 V cm21 with a 1?2–8?5 s linear ramp
time for 30 h.
DNA was analysed by Southern hybridization with a phtAa-specific
DIG-labelled DNA probe, which was prepared using the PCR DIG
probe synthesis kit (Roche) with primers phtAa-2 and phtAa-3.
Hybridization was performed at 41 uC and washes with 0?56 SSC,
0?1% SDS were performed at 61 uC.
Degradation of phthalate. E. coli JM109 cells containing either
pPHT, pRS14 or pRS42 were grown in 50 ml LB broth supplemen-
ted with 100 mg ampicillin ml21 at 30 uC with shaking. The cells
were pelleted by centrifugation at 4000 g, washed twice with 50 ml
50 mM phosphate buffer (pH 6?8) and suspended in 20 ml buffer
to an OD600 of 10. IPTG (Invitrogen) and phthalic acid (dipotas-
sium salt; Aldrich) were added to each at a final concentration of
0?1% and the cultures were incubated at 30 uC for 16–18 h. The
cells were pelleted by centrifugation at 4000 g and the resultant
supernatants were extracted three times with an equal volume of
ethyl acetate. The samples were derivatized with 1% trimethylchloro-
silane (TMS; Regis Technologies) for GC-MS analyses. The samples
were dissolved in 250 ml acetonitrile. Dissolved sample (100 ml) was
mixed with 150 ml silylation reagent and allowed to react for 1 h at
60 uC. GC-MS analysis was performed on the Thermo Finnigan TSQ
700 triple quadrupole mass spectrometer operated in EI mode.
Separation was achieved in a J&W DB5-ms capillary column
(30 m60?25 mm i.d.60?25 mm). GC-MS analyses were performed
with a column temperature rate of 10 uC min21 and a total analysis
time of 25 min. The amount of degradation was calculated from the
peak area as compared to control.
RESULTS
Nucleotide sequencing and operon organization
An M. vanbaalenii PYR-1 genomic library was constructedpreviously in a fosmid vector and screened for nidA(Stingley et al., 2004). A single nidA-positive fosmidclone, pFOS608, was chosen for initial sequencing studies.The clone was digested with BamHI and the resultantfragments were subcloned into pGEM-11z and sequenced.Initial sequences were obtained from T7 and Sp6 primerslocated on either end of the insert site in pGEM-11z andsubsequent sequencing used walking primers in bothdirections. Subclone junctions were amplified by high-fidelity PCR using pFOS608 as template and primers thatwere insert-specific. The resulting PCR products weresequenced to confirm the relative organization of theBamHI fragments in the fosmid insert (Fig. 2a).
BLAST searches with the resulting sequence data indicatedthe presence of a putative phthalate-degrading (pht) operon(Fig. 2, Table 3). A putative regulatory protein gene (phtR),transcribed from the opposite strand, is upstream ofthe putative operon. The operon consists of large (phtAa)and small (phtAb) dioxygenase subunits, an unknownORF (phtU) (not shown in Fig. 2), a dihydrodiol dehy-drogenase (phtB), a dioxygenase ferredoxin subunit (phtAc)and a dioxygenase ferredoxin reductase (phtAd). The stopsites of phtAa, phtAb and phtU overlap with the start sitesof phtAb, phtU and phtB, respectively, and the stop site ofphtAc overlaps the start site of phtAd. In each instance,the stop/start overlap consists of four bases, ATGA. Theoperon gene products share 53–78% identity and 66–88% similarity with their counterparts in Terrabacter sp.DBF63 and A. keyseri 12B (Table 3). However, the M.vanbaalenii PYR-1 operon differs from operons in thesespecies in the placement and orientation of the regulatorygene, which is encoded upstream of the operon andtranscribed divergently, rather than encoded in tandemwith the operon (Fig. 2b). In contrast to the pht operonsin Terrabacter sp. DBF63, A. keyseri 12B, Burkholderiacepacia DBO1 and Pseudomonas putida NMH102-2, the
Table 2. PCR primers and oligonucleotide probes
Primer Sequence (5§–3§) Gene Relative position*
phtAa-1 GACGTGCTTTCCCAACATCAG phtAa 2522–2542
phtAa-2 GTACGCACTGGCATGATTC phtAa 1671–1689
phtAa-3 GCCGTTGATTGTTCTCGTTGTAGC phtAa 2928–2905
phtAa-4 TCGTTCATCATCGCTCGTG phtAa 1827–845
phtAd-1 GTGAGGCCGATCTTAAGGTTG phtAd 5680–5660
phtB-1 GGAGCAGGTTCGGGTATCGG phtB 4026–4045
phtB-2 TGTTTCGCATCAACGTCCT phtB 4321–4339
phtB-3 ACTTCGACGCCACATACAG phtB 4470–4452
*Relative position in the pht operon sequence (GenBank accession no. AY365117).
3752 Microbiology 150
R. L. Stingley and others
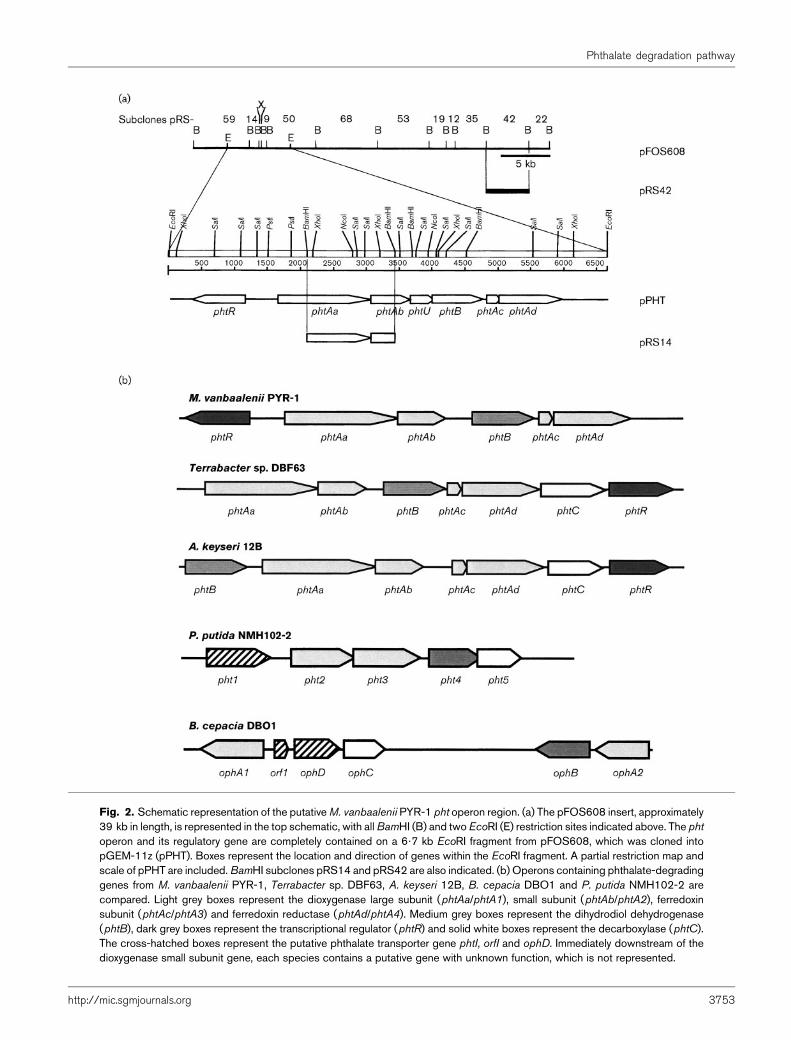
Fig. 2. Schematic representation of the putativeM. vanbaalenii PYR-1 pht operon region. (a) The pFOS608 insert, approximately39 kb in length, is represented in the top schematic, with allBamHI (B) and twoEcoRI (E) restriction sites indicated above. The phtoperon and its regulatory gene are completely contained on a 6?7 kb EcoRI fragment from pFOS608, which was cloned intopGEM-11z (pPHT). Boxes represent the location and direction of genes within the EcoRI fragment. A partial restriction map andscale of pPHT are included.BamHI subclones pRS14 and pRS42 are also indicated. (b) Operons containing phthalate-degradinggenes from M. vanbaalenii PYR-1, Terrabacter sp. DBF63, A. keyseri 12B, B. cepacia DBO1 and P. putida NMH102-2 arecompared. Light grey boxes represent the dioxygenase large subunit (phtAa/phtA1), small subunit (phtAb/phtA2), ferredoxinsubunit (phtAc/phtA3) and ferredoxin reductase (phtAd/phtA4). Medium grey boxes represent the dihydrodiol dehydrogenase(phtB), dark grey boxes represent the transcriptional regulator (phtR) and solid white boxes represent the decarboxylase (phtC).The cross-hatched boxes represent the putative phthalate transporter gene phtI, orfI and ophD. Immediately downstream of thedioxygenase small subunit gene, each species contains a putative gene with unknown function, which is not represented.
http://mic.sgmjournals.org 3753
Phthalate degradation pathway

M. vanbaalenii PYR-1 pht operon did not contain adecarboxylase gene (Fig. 2b) and none was locatedwithin approximately 2?5–3 kb of either end of theoperon region.
M. vanbaalenii PYR-1 phthalate dioxygenase(PhtAaAbAcAd)
Sequence alignments with proteins from BLASTX searchesrevealed that the large subunit of the phthalate dioxygenase(PhtAa) was closely related to aromatic ring hydroxylationdioxygenase E (gene E) of Rhodococcus sp. RHA1 (78%identical and 88% similar residues), PhtAa of A. keyseri 12B(75% identical and 87% similar residues) and PhtA1 ofTerrabacter sp. DBF63 (71% identical and 82% similar
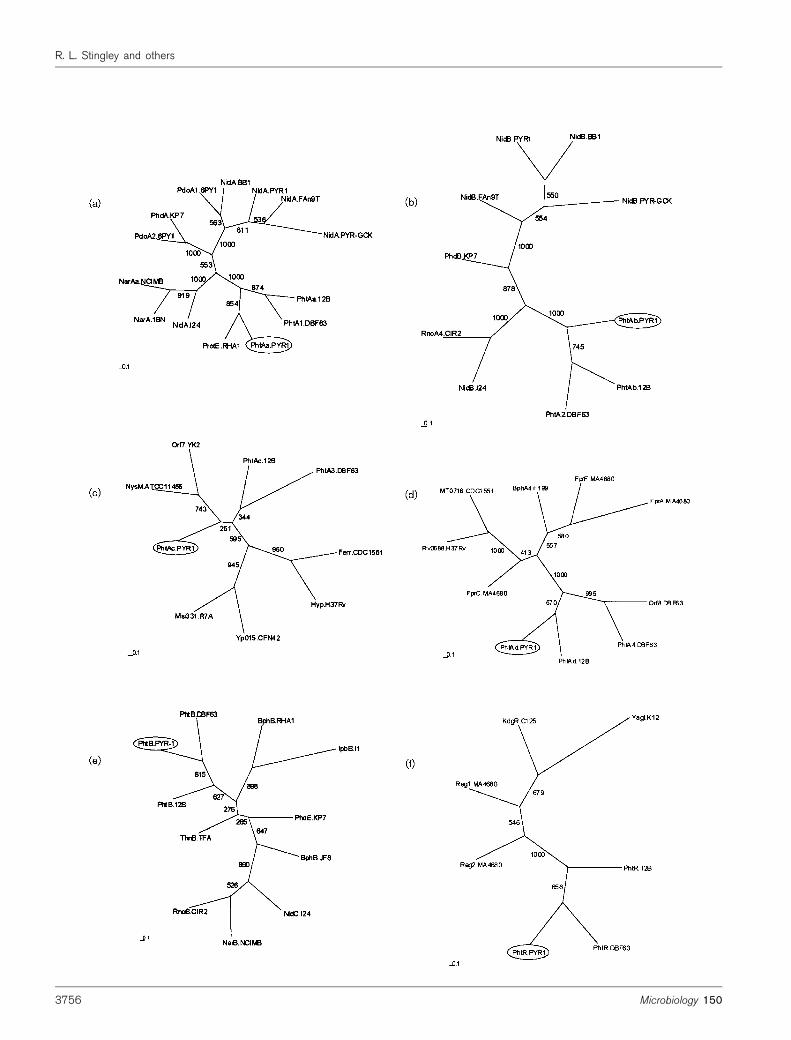
residues) (Table 3). The putative protein sequence con-tained a number of conserved domains related to those oflarge subunit ring-hydroxylating dioxygenases (Table 4).M. vanbaalenii PYR-1 PhtAa and the product of Rhodo-coccus sp. RHA1 gene E branch together in an unrootedphylogenetic tree (Fig. 3a). These proteins share member-ship in a clade with PhtAa and PhtA1 of A. keyseri 12Band Terrabacter sp. DBF63, respectively. PhtAa was moredistantly related to the previously characterized largesubunit dioxygenase, NidA, of M. vanbaalenii PYR-1,which is involved in pyrene degradation (Khan et al.,2001) (Fig. 3a). These two enzymes share only 41% identityand 56% similarity over 437 residues. Alignment of thephthalate dioxygenase large subunit from M. vanbaaleniiPYR-1 and 13 of the most closely related Nocardioform spp.
Table 3. Putative pht operon gene products
Putative
gene
Relative
position*
Deduced Mr
(aa residues)
Gene product Related proteins, GenBank accession no.,
percentage identity/percentage similarityD
phtR 3886–4690 28 971 (268) pht operon regulator Regulatory protein, from Terrabacter sp. DBF63, AB55883, 55/69%
over 245 residues; putative pht operon regulator, A. keyseri 12B,
AF331043, 53/71% over 225 residues; putative regulator from
E. coli K-12, NP_414806, 32/53% over 245 residues
phtAa 5153–6616 54 484 (487) Phthalate dioxygenase,
large subunit
Aromatic ring hydroxylation dioxygenase E from Rhodococcus sp.
RHA1, BAB62289, 78/88% over 356 residues; oxygenase large
subunit of phthalate dioxygenase from Terrabacter sp. DBF63,
BAC54156, 71/82% over 477 residues; phthalate dioxygenase large
subunit from A. keyseri 12B, AF331043, 75/87% over 429 residues
phtAb 6613–7212 22 563 (199) Phthalate dioxygenase,
small subunit
Phthalate dioxygenase small subunit from A. keyseri 12B, AF331043,
68/80% over 199 residues; oxygenase small subunit of phthalate
dioxygenase from Terrabacter sp. DBF63, BAC54157, 67/79%
over 191 residues; Fe–S protein from Rhodococcus sp. CIR2,
BAA76339, 44/61% over 158 residues
orf9-1 7209–7514 10 414 (101) Unknown Unknown protein of Terrabacter sp. DBF63, BAC54158, 57/71%
over 91 residues; hypothetical protein from Mycobacterium sp.
6PY1, CAD38645, 44/63% over 61 residues; Orf72 from
Nocardioides sp. KP7, BAA94710, 53/73% over 44 residues
phtB 7511–8314 28 296 (267) Dihydrodiol
dehydrogenase
3,4-Dihydroxy-3,4-dihydrophthalate dehydrogenase from Terrabacter
sp. DBF63, BAC54159, 65/76% over 263 residues; cis-naphthalene
dihydrodiol dehydrogenase from Rhodococcus sp. NCIMB12038,
AAD30203, 49/66% over 258 residues; dihydrodiol dehydrogenase
from Nocardioides sp. KP7, BAA94705, 45/58% over 256 residues
phtAc 8342–8542 7 200 (66) Phthalate dioxygenase,
ferredoxin subunit
Phthalate dioxygenase ferredoxin from A. keyseri 12B, AF331043,
69/77% over 61 residues; ferredoxin of phthalate dioxygenase
from Terrabacter sp. DBF63, BAC54160, 55/66% over 55
residues; ferredoxin-related protein from M. tuberculosis,
NP_335215, 38/58% over 62 residues
phtAd 8539–9780 44 562 (413) Phthalate dioxygenase,
ferredoxin reductase
Phthalate dioxygenase reductase subunit from A. keyseri 12B,
AF331043, 58/73% over 403 residues; ferredoxin reductase of
phthalate dioxygenase from Terrabacter sp. DBF63, BAC54161,
57/71% over 398 residues; ferredoxin reductase from Terrabacter
sp. DBF63, BAB55881, 57/71% over 398 residues
*Relative position in bases on pht operon sequence, accession no. AY365117.
DPercentages based on BLASTX alignments.
3754 Microbiology 150
R. L. Stingley and others
proteins from a BLASTX search indicates significant homol-ogy in the Rieske domain and a consensus Fe–S-bindingmotif, especially with A. keyseri 12B PhtAa, Terrabacter sp.DBF63 PhtA1 and the Rhodococcus spp. large subunitdioxygenases. Alignment of two Gram-negative phthalatedioxygenases from B. cepacia DBO1 and P. putidaNMH102-2 showed less than 24% similarity with PYR-1phthalate dioxygenase.
The small subunit phthalate dioxygenase (PhtAb) was mostclosely related by sequence alignment to PhtAb of A. keyseri12B (68% identical and 80% similar residues) and PhtA2 ofTerrabacter sp. DBF63 (67% identical and 79% similarresidues) (Table 3). The putative protein sequence con-tained domains related to small subunit dioxygenases(Table 4). In addition, M. vanbaalenii PYR-1 PhtAb is amember of the same clade of an unrooted phylogenetic treeas PhtAb and PhtA2 of A. keyseri 12B and Terrabacter sp.DBF63, respectively (Fig. 3b). PhtAb shares only 45%identity and 61% similarity with NidB, a previouslydescribed dioxygenase small subunit involved in pyrenemetabolism in M. vanbaalenii PYR-1 (Khan et al., 2001).
The putative protein sequence of the phthalate dioxygenase
ferredoxin subunit (PhtAc) contains a ferredoxin domain(Table 4). Upon sequence alignment, PhtAc shared thegreatest homology with PhtAc of A. keyseri 12B (69%identical and 77% similar residues) and PhtA3 of Terra-bacter sp. DBF63 (55% identical and 66% similar residues)(Table 3). M. vanbaalenii PYR-1 PhtAc shares membershipin a clade with these proteins; however, it appears to bemorephylogenetically related to NysM of Streptomyces nourseiATCC 11455 and the product of orf7 of Rhodococcus sp. YK2(Fig. 3c). NysM is a ferredoxin that participates in electrontransfer in P450 monooxygenase systems (Brautaset et al.,2000) and the product of orf7 is a putative ferredoxin.
Sequence alignment of the phthalate dioxygenase ferredoxinreductase (PhtAd) indicated that it was most similar toPhtAd of A. keyseri 12B (58% identical and 73% similarresidues) and PhtA4 of Terrabacter sp. DBF63 (57%identical and 71% similar residues) (Table 3). The puta-tive protein sequence contained several domains related toferredoxin reductases (Table 4). Upon phylogenetic analy-sis, these proteins cluster together, along with the productof orf8 of Terrabacter sp. DBF63 (a ferredoxin reductase) toform a clade, with M. vanbaalenii PYR-1 PhtAd branchingmost closely to PhtAd of A. keyseri 12B (Fig. 3d).
Table 4. Conserved domains in putative pht operon proteins
Gene Amino acid
residues
Conserved domains*
phtAa 487 Residues 18–355 aligned with COG4638, HcaE, ring-hydroxylating dioxygenase large
terminal subunit
Residues 165–447 aligned with pfam00848, ring hydroxylating alpha subunit
Residues 43–123 aligned with pfam00355, Rieske [2Fe–2S] domain
Residues 43–122 aligned with COG2146, ferredoxin subunits of ring-hydroxylating dioxygenases
phtAb 199 Residues 49–193 aligned with pfam00866, ring-hydroxylating beta subunit
Residues 45–199 aligned with COG5517, small subunit of phenylpropionate dioxygenase
phtAc 66 Residues 9–65 aligned with COG1141, ferredoxin
phtAd 413 Residues 17–282 aligned with pfam00070, pyridine nucleotide-disulphide oxidoreductase
Residues 17–390 aligned with COG0446, HcaD, uncharacterized NAD(FAD)-dependent
dehydrogenases
Residues 17–323 aligned with COG1252, Ndh, NADH dehydrogenase, FAD-containing subunit
phtB 267 Residues 6–258 aligned with pfam00106, adh_short, short-chain dehydrogenase
Residues 5–259 aligned with COG1028, FabG, dehydrogenases with different specificities
(related to short-chain alcohol dehydrogenase)
Residues 5–192 aligned with COG4221, short-chain alcohol dehydrogenase of unknown
specificity
Residues 5–189 aligned with COG0300, DltE, short-chain dehydrogenases of various substrate
specificities
Residues 5–186 aligned with COG3967, DltE, short-chain dehydrogenase involved in D-alanine
esterification of lepoteichoic acid and wall teichoic acid (D-alanine transfer protein)
Residues 2–260 aligned with COG0623, FabI, enoyl-(acyl carrier protein)
phtR 268 Residues 24–268 aligned with COG1414, IclR, transcriptional regulator
Residues 79–265 aligned with pfam01614, IclR, bacterial transcriptional regulator
Residues 22–111 aligned with smart00346, HTH-ICLR, helix–turn–helix isocitrate lyase
regulation
*Conserved domains based on BLASTP conserved domain search (Marchler-Bauer et al., 2003).
http://mic.sgmjournals.org 3755
Phthalate degradation pathway
3756 Microbiology 150
R. L. Stingley and others

Other proteins from the M. vanbaalenii PYR-1pht operon region
Sequence alignments indicate that the phthalate dihydrodioldehydrogenase (PhtB) is most closely related to PhtB ofTerrabacter sp. DBF63 (65% identical and 76% similarresidues) and NarB of Rhodococcus sp. NCIMB12038(49% identical and 66% similar residues) (Table 3). Thealignments suggest that M. vanbaalenii PYR-1 PhtB is notsignificantly similar to that of A. keyseri 12B. Phylogeneticanalysis, however, indicates thatM. vanbaalenii PYR-1 PhtBis more closely related to that of A. keyseri 12B than toRhodococcus sp. NCIMB12038 NarB (Fig. 3e). In addition,the putative protein sequence of PhtB contained a numberof dehydrogenase domains (Table 4).
In sequence alignments the M. vanbaalenii PYR-1 phtregulatory protein (PhtR) was most similar to PhtR ofTerrabacter sp. DBF63 (55% identical and 69% similarresidues) and PhtR of A. keyseri 12B (53% identical and71% similar residues) (Table 3). Phylogenetic analysissupports the relatedness of these regulatory proteins, asthey are co-members of a clade in an unrooted tree (Fig. 3f).TheM. vanbaalenii PYR-1 PhtR contains a helix–turn–helixmotif near the N terminus and a conserved region near theC terminus that is typical of the isocitrate lyase regulation(IclR) family (Table 4) (Donald et al., 2001). Members of
this family have been implicated in both repression(Yamamoto & Ishihama, 2003) and activation (Torreset al., 2003; Trautwein & Gerischer, 2001) of bacterialtranscription. A number of aromaticmetabolic pathways areregulated by members of the IclR family (Contzen & Stolz,2000; Eulberg & Schlomann, 1998; Martin & Mohn, 2000;Torres et al., 2003; Trautwein & Gerischer, 2001).
A putative protein of unknown function is encoded by phtUand contains no conserved domains. This protein is similarto hypothetical proteins of unknown function in the Terra-bacter sp. DBF63 pht operon (57% identical and 71%similar residues) and in a region of the Mycobacterium sp.6PY1 genome that contains PAH-degradation enzymes(44% identical and 63% similar residues) (Krivobok et al.,2003) (Table 3). In addition, the A. keyseri 12B pht operoncontains a putative ORF that encodes a similar protein and isimmediately downstream of the phtAb ORF (analysis ofGenBank accession no. AF331043).
Degradation of phthalate
To determine whether enzymes from the M. vanbaaleniiPYR-1 pht operon were capable of phthalate degradation,the pht region, which is completely contained on a 6?7 kbEcoRI fragment from pFOS608 (Fig. 2a), was cloned intothe pGEM-11z vector (pPHT).M. vanbaalenii PYR-1, clone
Fig. 3. Phylogenetic analyses of phthalate dioxygenase proteins. Protein sequences with the greatest homology to M.
vanbaalenii PYR-1 phthalate enzymes, based on BLASTX results, were used in the analyses. The multiple-alignment analysiswas performed using the PHYLIP software package and phylogenetic unrooted trees were drawn using TREEVIEW. Thenumbers on some branches refer to the percentage confidence, estimated by a bootstrap analysis with 1000 replications. Thescale bars indicate percentage divergence for each tree. Large subunit dioxygenases (a), small subunit dioxygenases(b), dioxygenase ferredoxin subunits (c), dioxygenase ferredoxin reductases (d), dihydrodiol dehydrogenases (e) andtranscriptional regulators (f) were analysed. GenBank accession numbers are as follows. (a) PhtAa of M. vanbaalenii PYR-1,AY365117; Protein E of Rhodococcus sp. RHA1, BAB62289; PhtA1 of Terrabacter sp. DBF63, BAC54156; PhtAa of A.keyseri 12B, AAK16534; NidA of Rhodococcus sp. I24, AAD25395; NarAa of Rhodococcus sp. NCIMB12038, AAD28100;NarA of Rhodococcus sp. 1BN, CAC14063; PhdA of Nocardioides sp. KP7, BAA84712; NidA of M. vanbaalenii PYR-1,AAF75991; NidA of M. gilvum BB1, AAN78316; NidA of M. flavescens PYR-GCK, AAN78312; PdoA1 of Mycobacterium sp.6PY1, CAD38647; NidA of M. frederiksbergense FAn9, AAN78314; PdoA2 of Mycobacterium sp. 6PY1, CAD38643.(b) PhtAb of M. vanbaalenii PYR-1, AY365117; PhtAb of A. keyseri 12B, AAK16535; PhtA2 of Terrabacter sp. DBF63,BAC54157; RnoA4 of Rhodococcus sp. CIR2, BAA76339; NidB of M. frederiksbergense FAn9, AAN78315; NidB ofRhodococcus sp. I24, AAD25396; PhdB of Nocardioides sp. KP7, BAA84713; NidB of M. vanbaalenii PYR-1, AAF75992;NidB of M. gilvum BB1, AAN78317; NidB of M. flavescens PYR-GCK, AAN78313. (c) PhtAc of M. vanbaalenii PYR-1,AY365117; PhtAc of A. keyseri 12B, AAK16536; PhtA3 of Terrabacter sp. DBF63, BAC54160; ferredoxin-related protein ofM. tuberculosis CDC1551, NP_335215; hypothetical protein of M. tuberculosis H37Rv, NP_215277; Orf7 protein ofRhodococcus sp. YK2, BAC00808; Msi331 of Mesorhizobium loti R7A, CAD31363; Yp015 of Rhizobium etli CFN42,NP_659824; NysM of Streptomyces noursei ATCC 11455, AAF71770. (d) PhtAd of M. vanbaalenii PYR-1, AY365117;PhtAd of A. keyseri 12B, AAK16537; PhtA4 of Terrabacter sp. DBF63, BAC54161; Orf8 protein of Terrabacter sp. DBF63,BAB55881; FprC of Streptomyces avermitilis MA-4680, NP_828132; Rv0688 of M. tuberculosis H37Rv, NP_215202;MT0716 of M. tuberculosis CDC1551, NP_335128; BphA4 of Novosphingobium aromaticivorans F199, NP_049182; FprAof S. avermitilis MA-4680, NP_821758; FprF of S. avermitilis MA-4680, NP_828132. (e) PhtB of M. vanbaalenii PYR-1,AY365117; PhtB of Terrabacter sp. DBF63, BAC54159; NarB of Rhodococcus sp. NCIMB12038, AAD30203; BphB ofBacillus sp. JF8, BAC79228; NidC of Rhodococcus sp. I24, AAD25397; PhdE of Nocardioides sp. KP7, BAA94705; ThnBof Sphingopyxis macrogoltabida TFA, AAN26445; IpbB of Rhodococcus sp. I1, CAA06877; RnoB of Rhodococcus sp.CIR2, BAA76340; BphB of Rhodococcus sp. RHA1, BAA06873; PhtB of A. keyseri 12B, AAK16533. (f) PhtR of M.
vanbaalenii PYR-1, AY365117; PhtR of Terrabacter sp. DBF63, BAB55883; PhtR of A. keyseri 12B, AAK16539; YagI ofE. coli K-12, NP_414806; regulatory protein 1 of S. avermitilis MA-4680, NP_828130; regulatory protein 2 of S. avermitilis
MA-4680, NP_826918; KdgR of Bacillus halodurans C-125, NP_244592.
http://mic.sgmjournals.org 3757
Phthalate degradation pathway
pFOS608, and subclones pRS42 and pRS14 (negativecontrol) were incubated with 0?1% phthalate in phos-phate buffer (pH 6?8) at 30 uC for 16–18 h and the resultingsupernatant was extracted with ethyl acetate for GC-MSanalysis. M. vanbaalenii PYR-1 and clone pFOS608 pro-duced 3,4-dihydroxyphthalate which eluted at 16?2 min andgave major ions at m/z (percentage intensity, proposedcomposition) 486 (8, M+), 471 (57, [M-CH3]
+), 349 (30),309 (20, [M-CH3-OTMS-TMS]+), 259 (38), 220 (45), 206(100), 192 (49), 103 (40) and 91 (21).M. vanbaalenii PYR-1was able to degrade 95% of phthalate and clone pFOS608degraded 75% of phthalate under similar conditions. Sub-clones pRS14 (containing the 39 end of phtAa and the 59end of phtAb) and pRS42 (containing unrelated putativegenes for membrane proteins) (Fig. 2a) were used in con-trol experiments. 3,4-Dihydroxyphthalate was not formedby control strains.
Screening Mycobacterium and Rhodococcusstrains for pht genes
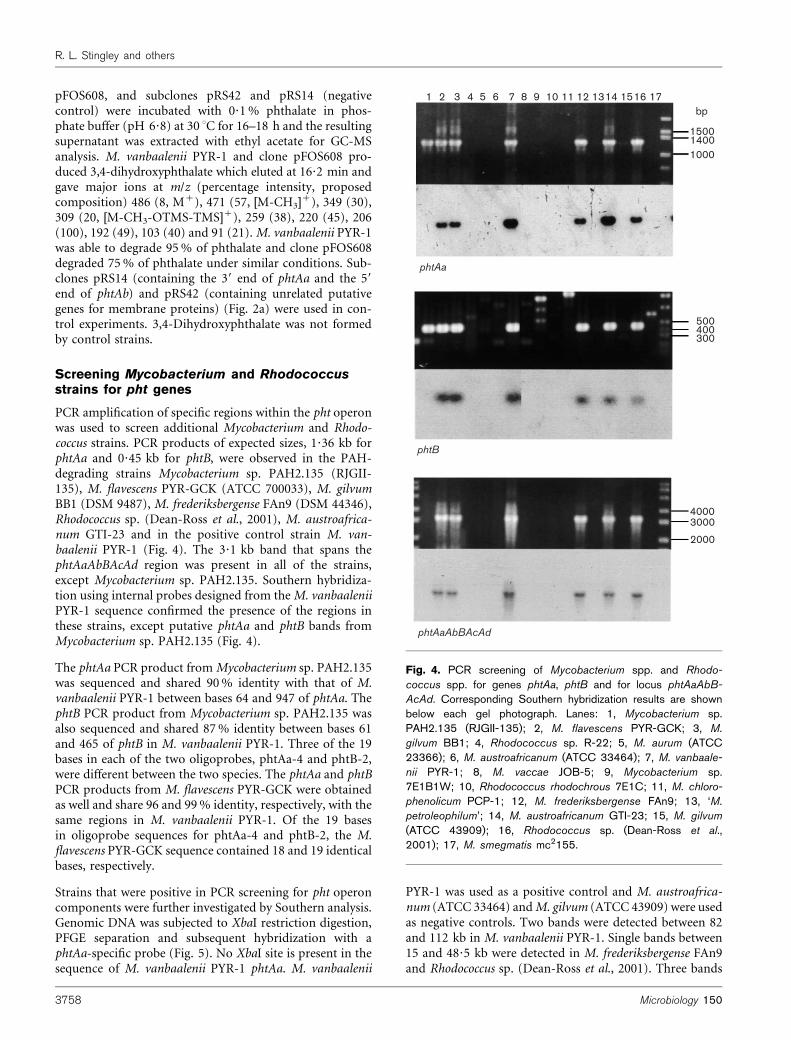
PCR amplification of specific regions within the pht operonwas used to screen additional Mycobacterium and Rhodo-coccus strains. PCR products of expected sizes, 1?36 kb forphtAa and 0?45 kb for phtB, were observed in the PAH-degrading strains Mycobacterium sp. PAH2.135 (RJGII-135), M. flavescens PYR-GCK (ATCC 700033), M. gilvumBB1 (DSM 9487), M. frederiksbergense FAn9 (DSM 44346),Rhodococcus sp. (Dean-Ross et al., 2001), M. austroafrica-num GTI-23 and in the positive control strain M. van-baalenii PYR-1 (Fig. 4). The 3?1 kb band that spans thephtAaAbBAcAd region was present in all of the strains,except Mycobacterium sp. PAH2.135. Southern hybridiza-tion using internal probes designed from theM. vanbaaleniiPYR-1 sequence confirmed the presence of the regions inthese strains, except putative phtAa and phtB bands fromMycobacterium sp. PAH2.135 (Fig. 4).
The phtAa PCR product fromMycobacterium sp. PAH2.135was sequenced and shared 90% identity with that of M.vanbaalenii PYR-1 between bases 64 and 947 of phtAa. ThephtB PCR product from Mycobacterium sp. PAH2.135 wasalso sequenced and shared 87% identity between bases 61and 465 of phtB in M. vanbaalenii PYR-1. Three of the 19bases in each of the two oligoprobes, phtAa-4 and phtB-2,were different between the two species. The phtAa and phtBPCR products from M. flavescens PYR-GCK were obtainedas well and share 96 and 99% identity, respectively, with thesame regions in M. vanbaalenii PYR-1. Of the 19 basesin oligoprobe sequences for phtAa-4 and phtB-2, the M.flavescens PYR-GCK sequence contained 18 and 19 identicalbases, respectively.
Strains that were positive in PCR screening for pht operoncomponents were further investigated by Southern analysis.Genomic DNA was subjected to XbaI restriction digestion,PFGE separation and subsequent hybridization with aphtAa-specific probe (Fig. 5). No XbaI site is present in thesequence of M. vanbaalenii PYR-1 phtAa. M. vanbaalenii
PYR-1 was used as a positive control and M. austroafrica-num (ATCC 33464) andM. gilvum (ATCC 43909) were usedas negative controls. Two bands were detected between 82and 112 kb in M. vanbaalenii PYR-1. Single bands between15 and 48?5 kb were detected in M. frederiksbergense FAn9and Rhodococcus sp. (Dean-Ross et al., 2001). Three bands
1 2 3 4 5 6 7 8 9 10 11 12 1314 1516 17bp
150014001000
500400300
40003000
phtAa
phtB
phtAaAbBAcAd
2000
Fig. 4. PCR screening of Mycobacterium spp. and Rhodo-
coccus spp. for genes phtAa, phtB and for locus phtAaAbB-
AcAd. Corresponding Southern hybridization results are shownbelow each gel photograph. Lanes: 1, Mycobacterium sp.PAH2.135 (RJGII-135); 2, M. flavescens PYR-GCK; 3, M.
gilvum BB1; 4, Rhodococcus sp. R-22; 5, M. aurum (ATCC23366); 6, M. austroafricanum (ATCC 33464); 7, M. vanbaale-
nii PYR-1; 8, M. vaccae JOB-5; 9, Mycobacterium sp.7E1B1W; 10, Rhodococcus rhodochrous 7E1C; 11, M. chloro-
phenolicum PCP-1; 12, M. frederiksbergense FAn9; 13, ‘M.
petroleophilum’; 14, M. austroafricanum GTI-23; 15, M. gilvum
(ATCC 43909); 16, Rhodococcus sp. (Dean-Ross et al.,2001); 17, M. smegmatis mc2155.
3758 Microbiology 150
R. L. Stingley and others

between 82 and 112 kb were detected inM. austroafricanumGTI-23. Three bands in M. flavescens PYR-GCK and twobands inM. gilvum BB1 were detected, all within a 33–97 kbrange. No hybridization signal was detected in Myco-bacterium sp. PAH2.135.
DISCUSSION
In an effort to identify additional M. vanbaalenii PYR-1genes involved in aromatic hydrocarbon degradation, agenomic library was constructed and used to examine aregion of the genome near previously characterizeddioxygenase genes (Khan et al., 2001; Stingley et al.,2004). Sequence from one fosmid clone of the region,pFOS608, indicated that multiple PAH-degrading genes areclustered together on the genome.
Based upon sequencing data and metabolite studies, aphthalate-degrading operon is positioned approximately
12–19 kb upstream of the dioxygenase large subunit gene,nidA. Putative products from genes in the operon sharehomology with those characterized in other species,especially Terrabacter sp. DBF63 (Habe et al., 2003) andA. keyseri 12B (Eaton, 2001). Although BLAST searchesindicate little homology at the DNA level, searches of theprotein database suggest substantial homology to proteinsfrom the pht operons in these species. Phthalate dioxygenasefrom two Gram-negative strains, P. putida NMH102-2 andB. cepacia DBO1, showed different genetic organization ascompared to nocardiforms and showed less than 25%sequence homology. The M. vanbaalenii PYR-1 phthalateoperon genes were therefore named in the manner of thoseof A. keyseri 12B (Eaton, 2001).
Phylogenetic analyses of the dioxygenase proteins (PhtAa,PhtAb, PhtAc and PhtAd) from the M. vanbaalenii PYR-1pht operon confirm a significant relationship with counter-part genes from the Terrabacter sp. DBF63 (Habe et al.,2003) and A. keyseri 12B (Eaton, 2001) pht operons.Sequence alignments indicated that PhtB showed consider-able divergence from A. keyseri 12B PhtB (Eaton, 2001),yet was related to Terrabacter sp. DBF63 PhtB (Habe et al.,2003) and other dihydrodiol dehydrogenases (Larkin et al.,1999; Masai et al., 1995; Saito et al., 2000; Treadwayet al., 1999). Phylogenetic analysis of PhtB suggests,however, that the protein is closely related to PhtB ofboth Terrabacter sp. DBF63 and A. keyseri 12B. The M.vanbaalenii PYR-1 regulatory protein, PhtR, and a putativeprotein of unknown function are also related to similarproteins encoded by Terrabacter sp. DBF63 and A. keyseri12B genes.
A clone containing the M. vanbaalenii PYR-1 pht operonin E. coli exhibited the ability to degrade phthalate to theexpected dihydroxyphthalate. The fragmentation pattern ofthe TMS derivative of the metabolite obtained was similarto those previously reported for 3,4-dihydroxyphthalate(Chang & Zylstra, 1998; Eaton, 2001; Habe et al., 2003).Based on the sequence homologies among M. vanbaaleniiPYR-1 pht operon proteins and those of Terrabacter sp.DBF63 and A. keyseri 12B, the product of phthalatedegradation by enzymes from this operon is most likely3,4-dihydroxyphthalate, rather than 4,5-dihydroxyphtha-late, which is more commonly produced by Gram-negativespecies (Chang & Zylstra, 1998; Eaton, 2001; Habe et al.,2003; Nomura et al., 1992).
The pht operon appears to be conserved within PAH-degrading Mycobacterium spp. Based on PCR and Southernhybridization analyses, all of the PAH-degrading strainsexamined in this study, except strain PAH2.135, appear tohave homologues to phtAa, phtB and the phtAaAbBAcAdregion. Although phtAa and phtB probes did not bind toPCR products or genomic digests of strain PAH2.135, thesequences of the PCR products share significant homologywith the related portions of strain PYR-1 genes. Among thestrains analysed in this study, a correlation between the PAH
1 2 3 4 5 6 7 8 9
kb
145.5130.5
112.0
97.0
82.0
63.5
48.5
33.5
15.0
Fig. 5. Screening Mycobacterium spp. and Rhodococcus sp.for phtAa. Bacterial genomic DNA was digested with XbaI,separated by PFGE and hybridized with a phtAa-specific probe.Lanes: 1, M. vanbaalenii PYR-1; 2, Mycobacterium sp.PAH2.135 (RJGII-135); 3, M. gilvum BB1; 4, M. flavescens
PYR-GCK; 5, M. frederiksbergense FAn9; 6, M. austroafrica-
num GTI-23; 7, Rhodococcus sp. (Dean-Ross et al., 2001);8, M. austroafricanum (ATCC 33464); 9, M. gilvum (ATCC43909).
http://mic.sgmjournals.org 3759
Phthalate degradation pathway

degradative phenotype and the presence of nidA, nidB and phtgenes was observed (Brezna et al., 2003; Khan et al., 2001).
This report is the first to identify and characterize afunctional operon involved in biodegradation of phthalateinMycobacterium spp. Although putative proteins from thisoperon share significant homology with those from similaroperons in Terrabacter sp. DBF63 (Habe et al., 2003) and A.keyseri 12B (Eaton, 2001), the M. vanbaalenii PYR-1 phtoperon is distinct both in its overall organization and in itsnucleotide sequence. The placement and orientation of thegene encoding the putative regulatory protein are distinctivein M. vanbaalenii PYR-1 (Fig. 2b). In addition, in contrastto the composition of pht operons in Terrabacter sp. DBF63(Habe et al., 2003) and A. keyseri 12B (Eaton, 2001), nodecarboxylase gene is present in the M. vanbaalenii PYR-1pht operon and none has been identified within that regionof the genome. Therefore, the M. vanbaalenii PYR-1 phtoperon is distinctly different from related operons in otherbacterial genera.
ACKNOWLEDGEMENTS
This work was supported in part by an appointment to the Post-graduate Research Program at the National Center for Toxico-logical Research administered by the Oak Ridge Institute for Scienceand Education through an interagency agreement between theUS Department of Energy and the US Food and Drug Adminis-tration. The authors would like to thankMr Allen Gies at the Universityof Arkansas for Medical Sciences for sequencing, Dr James P. Freemanfor GC-MS analyses, Ms Joanna Moody and Ms Lisa Mullis fortechnical support, and Dr Mark Hart and Dr Chris Elkins for criticalreview of the manuscript.
REFERENCES
Apajalahti, J. H. & Salkinoja-Salonen, M. S. (1987). Dechlorinationand para-hydroxylation of polychlorinated phenols by Rhodococcuschlorophenolicus. J Bacteriol 169, 675–681.
Api, A. M. (2001). Toxicological profile of diethyl phthalate: a vehiclefor fragrance and cosmetic ingredients. Food Chem Toxicol 39,97–108.
Barnsley, E. A. (1983). Phthalate pathway of phenanthrenemetabolism: formation of 2-carboxybenzalpyruvate. J Bacteriol 154,113–117.
Beam, H. W. & Perry, J. J. (1974). Microbial degradation ofcycloparaffinic hydrocarbons via co-metabolism and commensalism.J Gen Microbiol 82, 163–169.
Blevins, W. T. & Perry, J. J. (1972). Metabolism of propane, n-propylamine, and propionate by hydrocarbon-utilizing bacteria.J Bacteriol 112, 513–518.
Bogan, B. W., Lahner, L. M., Sullivan, W. R. & Paterek, J. R. (2003).Degradation of straight-chain aliphatic and high-molecular-weightpolycyclic aromatic hydrocarbons by a strain of Mycobacteriumaustroafricanum. J Appl Microbiol 94, 230–239.
Boldrin, B., Tiehm, A. & Fritzsche, C. (1993). Degradation ofphenanthrene, fluorene, fluoranthene, and pyrene by a Mycobac-terium sp. Appl Environ Microbiol 59, 1927–1930.
Bottger, E. C., Kirschner, P., Springer, B. & Zumft, W. (1997).
Mycobacteria degrading polycyclic aromatic hydrocarbons. Int J Syst
Bacteriol 47, 247.
Brautaset, T., Sekurova, O. N., Sletta, H., Ellingsen, T. E., StrLm,
A. R., Valla, S. & Zotchev, S. B. (2000). Biosynthesis of the polyene
antifungal antibiotic nystatin in Streptomyces noursei ATCC 11455:
analysis of the gene cluster and deduction of the biosynthetic
pathway. Chem Biol 7, 395–403.
Brezna, B., Khan, A. A. & Cerniglia, C. E. (2003). Molecular charac-
terization of dioxygenases from polycyclic aromatic hydrocarbon-
degrading Mycobacterium spp. FEMS Microbiol Lett 223, 177–183.
Burback, B. L. & Perry, J. J. (1993). Biodegradation and biotrans-
formation of groundwater pollutant mixtures by Mycobacterium
vaccae. Appl Environ Microbiol 59, 1025–1029.
Cadogan, D. F., Papez, M., Poppe, A. C., Pugh, D. M. & Scheubel, J.(1993). An assessment of the release, occurrence and possible effects
of plasticizers in the environment. Prog Rubber Plastics Technol 10, 1–19.
Cerniglia, C. E., Blevins, W. T. & Perry, J. J. (1976). Microbial oxidation
and assimilation of propylene. Appl Environ Microbiol 32, 764–768.
Chang, H.-K. & Zylstra, G. J. (1998). Novel organization of the
genes for phthalate degradation from Burkholderia cepacia DBO1.
J Bacteriol 180, 6529–6537.
Cobellis, L., Latini, G., Felice, C. D., Razzi, S., Paris, I., Ruggieri, F.,Mazzeo, P. & Petraglia, F. (2003). High plasma concentrations of
di(2-ethylhexyl)-phthalate in women with endometriosis. Hum
Reprod 18, 1512–1515.
Contzen, M. & Stolz, A. (2000). Characterization of the genes for
two protocatechuate 3,4-dioxygenases from the 4-sulfocatechol-
degrading bacterium Agrobacterium radiobacter strain S2. J Bacteriol
182, 6123–6129.
Dean-Ross, D. & Cerniglia, C. E. (1996). Degradation of pyrene by
Mycobacterium flavescens. Appl Microbiol Biotechnol 46, 307–312.
Dean-Ross, D., Moody, J. D., Freeman, J. P., Doerge, D. R. &Cerniglia, C. E. (2001). Metabolism of anthracene by a Rhodococcus
species. FEMS Microbiol Lett 204, 205–211.
Donald, L. J., Hosfield, D. J., Cuvelier, S. L., Ens, W., Standing, K. G.
& Duckworth, H. W. (2001). Mass spectrometric study of the
Escherichia coli repressor proteins, Ic1R and Gc1R, and their
complexes with DNA. Protein Sci 10, 1370–1380.
Duty, S. M., Singh, N. P., Silva, M. J., Barr, D. B., Brock, J. W., Ryan, L.,Herrick, R., Christiani, D. C. & Hauser, R. (2003). The relationship
between environmental exposures to phthalates and DNA damage in
human sperm using the neutral comet assay. Environ Health Perspect
111, 1164–1169.
Eaton, R. W. (2001). Plasmid-encoded phthalate catabolic pathway in
Arthrobacter keyseri 12B. J Bacteriol 183, 3689–3703.
Eulberg, D. & Schlomann, M. (1998). The putative regulator of
catechol catabolism in Rhodococcus opacus 1CP – an IclR-type, not a
LysR-type transcriptional regulator.Antonie van Leeuwenhoek 74, 71–82.
Graham, P. R. (1973). Phthalate ester plasticizers – why and how
they are used. Environ Health Perspect 3, 3–12.
Grifoll, M., Selifonov, S. A. & Chapman, P. J. (1994). Evidence for a
novel pathway in the degradation of fluorene by Pseudomonas sp.
strain F274. Appl Environ Microbiol 60, 2438–2449.
Grosser, R. J., Warshawsky, D. & Vestal, J. R. (1991). Indigenousand enhanced mineralization of pyrene, benzo[a]pyrene, and
carbazole in soils. Appl Environ Microbiol 57, 3462–3469.
Habe, H., Miyakoshi, M., Chung, J., Kasuga, K., Yoshida, T., Nojiri, H.& Omori, T. (2003). Phthalate catabolic gene cluster is linked to the
angular dioxygenase gene in Terrabacter sp. strain DBF63. Appl
Microbiol Biotechnol 61, 44–54.
3760 Microbiology 150
R. L. Stingley and others

Heitkamp, M. A. & Cerniglia, C. E. (1988). Mineralization of
polycyclic aromatic hydrocarbons by a bacterium isolated
from sediment below an oil field. Appl Environ Microbiol 54,
1612–1614.
Heitkamp, M. A. & Cerniglia, C. E. (1989). Polycyclic aromatic
hydrocarbon degradation by a Mycobacterium sp. in microcosms
containing sediment and water from a pristine ecosystem. ApplEnviron Microbiol 55, 1968–1973.
Heitkamp, M. A., Franklin, W. & Cerniglia, C. E. (1988a). Microbial
metabolism of polycyclic aromatic hydrocarbons: isolation and
characterization of a pyrene-degrading bacterium. Appl Environ
Microbiol 54, 2549–2555.
Heitkamp, M. A., Freeman, J. P., Miller, D. W. & Cerniglia, C. E.(1988b). Pyrene degradation by a Mycobacterium sp. Identification of
ring oxidation and ring fission products. Appl Environ Microbiol 54,
2556–2565.
Iizuka, H. E. A. (1975). Method of recovering microbial cells containing
protein. US Patent No. 3888736.
Kelley, I., Freeman, J. P., Evans, F. E. & Cerniglia, C. E. (1993).Identification of metabolites from the degradation of fluorantheneby Mycobacterium sp. strain PYR-1. Appl Environ Microbiol 59,
800–806.
Khan, A. A., Wang, R.-F., Cao, W.-W., Doerge, D. R., Wennerstrom, D.& Cerniglia, C. E. (2001). Molecular cloning, nucleotide sequence,
and expression of genes encoding a polycyclic aromatic ring
dioxygenase from Mycobacterium sp. strain PYR-1. Appl Environ
Microbiol 67, 3577–3585.
Khan, A. A., Kim, S.-J., Paine, D. D. & Cerniglia, C. E. (2002).Classification of a polycyclic aromatic hydrocarbon-metabolizing
bacterium, Mycobacterium sp. strain PYR-1 as Mycobacterium
vanbaalenii sp. nov. Int J Syst Evol Microbiol 52, 1997–2002.
King, D. H. & Perry, J. J. (1975). The origin of fatty acids in the
hydrocarbon-utilizing microorganism Mycobacterium vaccae. CanJ Microbiol 21, 85–89.
Kiyohara, H. & Nagao, K. (1978). The catabolism of phenanthrene
and anthracene by bacteria. J Gen Microbiol 105, 69–75.
Krivobok, S., Kuony, S., Meyer, C., Louwagie, M., Willison, J. C. &Jouanneau, Y. (2003). Identification of pyrene-induced proteins in
Mycobacterium sp. strain 6PY1: evidence for two ring-hydroxylating
dioxygenases. J Bacteriol 185, 3828–3841.
Larkin, M. J., Allen, C. C., Kulakov, L. A. & Lipscomb, D. A. (1999).Purification and characterization of a novel naphthalene dioxygenase
from Rhodococcus sp. strain NCIMB12038. J Bacteriol 181, 6200–6204.
Marchler-Bauer, A., Anderson, J. B., DeWeese-Scott, C. & 24 otherauthors (2003). CDD: a curated Entrez database of conserved
domain alignments. Nucleic Acids Res 31, 383–387.
Martin, V. J. & Mohn, W. W. (2000). Genetic investigation of the
catabolic pathway for degradation of abietane diterpenoids by
Pseudomonas abietaniphila BKME-9. J Bacteriol 182, 3784–3793.
Masai, E., Yamada, A., Healy, J. M., Hatta, T., Kimbara, K., Fukuda, M.& Yano, K. (1995). Characterization of biphenyl catabolic genes of
gram-positive polychlorinated biphenyl degrader Rhodococcus sp.strain RHA1. Appl Environ Microbiol 61, 2079–2085.
Moody, J. D., Freeman, J. P., Doerge, D. R. & Cerniglia, C. E. (2001).Degradation of phenanthrene and anthracene by cell suspensions
of Mycobacterium sp. strain PYR-1. Appl Environ Microbiol 67,
1476–1483.
Moody, J. D., Doerge, D. R., Freeman, J. P. & Cerniglia, C. E. (2002).Degradation of biphenyl by Mycobacterium sp. strain PYR-1. Appl
Environ Microbiol 58, 364–369.
Moody, J. D., Fu, P. P., Freeman, J. P. & Cerniglia, C. E. (2003). Regio-and stereoselective metabolism of 7,12-dimethylbenz[a]anthraceneby Mycobacterium vanbaalenii PYR-1. Appl Environ Microbiol 69,3924–3931.
Moody, J. D., Freeman, J. P., Fu, P. P. & Cerniglia, C. E. (2004).Degradation of benzo[a]pyrene by Mycobacterium vanbaalenii PYR-1.Appl Environ Microbiol 70, 340–345.
Nojiri, H., Kamakura, M., Urata, M., Tanaka, T., Chung, J. S.,Takemura, T., Yoshida, T., Habe, H. & Omori, T. (2002). Dioxincatabolic genes are dispersed on the Terrabacter sp. DBF63 genome.Biochem Biophys Res Commun 296, 233–240.
Nomura, Y., Nakagawa, M., Ogawa, N., Harashima, S. & Oshima, Y.(1992).Genes in PHT plasmid encoding the initial degradation pathwayof phthalate in Pseudomonas putida. J Ferment Bioeng 74, 333–344.
Peakall, D. B. (1975). Phthalate esters: occurrence and biologicaleffects. Residue Rev 54, 1–41.
Saito, A., Iwabuchi, T. & Harayama, S. (1999). Characterization ofgenes for enzymes involved in the phenanthrene degradation inNocardioides sp. KP7. Chemosphere 38, 1331–1337.
Saito, A., Iwabuchi, T. & Harayama, S. (2000). A novel phenanthrenedioxygenase from Nocardioides sp. strain KP7: expression inEscherichia coli. J Bacteriol 182, 2134–2141.
Schneider, J., Grosser, R., Jayasimhulu, K., Xue, W. & Warshawsky, D.(1996). Degradation of pyrene, benz[a]anthracene, and benzo[a]pyreneby Mycobacterium sp. strain RJGII-135, isolated from a former coalgasification site. Appl Environ Microbiol 62, 13–19.
Sepic, E., Bricelj, M. & Leskovsek, H. (1998). Degradation offluoranthene by Pasteurella sp. IFA and Mycobacterium sp. PYR-1:isolation and identification of metabolites. J Appl Microbiol 85, 746–754.
Snapper, S. B., Melton, R. E., Mustafa, S., Kieser, T. & Jacobs,W. R., Jr (1990). Isolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatis. Mol Microbiol4, 1911–1919.
Stingley, R. L., Khan, A. A. & Cerniglia, C. E. (2004). Molecular charac-terization of a phenanthrene degradation pathway in Mycobacteriumvanbaalenii PYR-1. Biochem Biophys Res Commun 322, 133–146.
Tickner, J. A., Schettler, T., Guidotti, T., McCally, M. & Rossi, M.(2001). Health risks posed by use of di-2-ethylhexyl phthalate(DEHP) in PVC medical devices: a critical review. Am J Ind Med 39,100–111.
Torres, B., Porras, G., Garcia, J. L. & Diaz, E. (2003). Regulation ofthe mhp cluster responsible for 3-(3-hydroxyphenyl)propionic aciddegradation in Escherichia coli. J Biol Chem 278, 27575–27585.
Trautwein, G. & Gerischer, U. (2001). Effects exerted by transcrip-tional regulator PcaU from Acinetobacter sp. strain ADP1. J Bacteriol183, 873–881.
Treadway, S. L., Yanagimachi, K. S., Lankenau, E., Lessard, P. A.,Stephanopoulos, G. & Sinskey, A. J. (1999). Isolation andcharacterization of indene bioconversion genes from Rhodococcusstrain I24. Appl Microbiol Biotechnol 51, 786–793.
Vestal, J. R. & Perry, J. J. (1969). Divergent metabolic pathways forpropane and propionate utilization by a soil isolate. J Bacteriol 99,216–221.
Willumsen, P., Karlson, U., Stackebrandt, E. & Kroppenstedt, R. M.(2001). Mycobacterium frederiksbergense sp. nov., a novel polycyclicaromatic hydrocarbon-degrading Mycobacterium species. Int J SystBacteriol 51, 1715–1722.
Yamamoto, K. & Ishihama, A. (2003). Two different modes oftranscription repression of the Escherichia coli acetate operon by IclR.Mol Microbiol 47, 183–194.
http://mic.sgmjournals.org 3761
Phthalate degradation pathway
Copyright © 2022 FDOKUMEN




















