
Neurotrophin-directed differentiation of human adult marrow stromal cells to dopaminergic-like...
14
Neurotrophin-directed differentiation of human adult marrow stromal cells to dopaminergic-like neurons Valerie M. Tatard a,1 , Gianluca D'Ippolito c,e,1 , Sylma Diabira a,b,c , Alexander Valeyev d,g , John Hackman d,g , Micheline McCarthy d,g,h , Thomas Bouckenooghe a , Philippe Menei a,b , Claudia N. Montero-Menei a,1 , Paul C. Schiller c,d,e,f,1, ⁎ a INSERM Unit 646, Ingénierie de la Vectorisation Particulaire, 10, rue André Bocquel, 49100 Angers, Centre Hospitalier Universitaire d'Angers, 4 rue Larrey, France b Département de Neurochirurgie, Centre Hospitalier Universitaire d'Angers, 4 rue Larrey, France c Geriatric Research, Education, and Clinical Center and Research Service, USA d Research Service, Veterans Affairs Medical Center, USA e Department of Medicine, University of Miami School of Medicine, Miami, FL 33125-1693, USA f Department of Biochemistry and Molecular Biology, University of Miami School of Medicine, Miami, FL 33125-1693, USA g Department of Neurology, University of Miami School of Medicine, Miami, FL 33125-1693, USA h Department of Microbiology and Immunology, University of Miami School of Medicine, Miami, FL 33125-1693, USA Received 27 April 2006; revised 18 August 2006; accepted 5 September 2006 Abstract Marrow-isolated adult multilineage inducible (MIAMI) cells were differentiated in vitro to neuronal cells in a neurotrophin-dependent fashion. After induction, the cells revealed electrophysiological features similar to those observed in mature neurons. Primary early passage human MIAMI cells without any type of co-cultures with other cell types were used. The developmental program involved a multi-step process requiring the concerted action of brain-derived neurotrophic factor, nerve growth factor and depended on neurotrophin-3, after basic fibroblast growth factor withdrawal. MIAMI-derived neuron-like cells sequentially expressed the neuronal markers, developed a complex neurite outgrowth and arborization, and acquired electrophysiological characteristics similar to those observed in mature neurons. The young and old MIAMI-derived neuronal cells developed both inward and outward currents upon depolarization, similar to those observed in normal neurons. These results represent the earliest evidence that neurotrophin-3 can direct the differentiation of non-neural stem cells from human adult bone marrow stroma to neuron-like cells in vitro. Supplementing the aforementioned multi-step process with sonic hedgehog, fibroblast growth factor 8, and retinoic acid increased the expression of molecules involved in dopaminergic differentiation and of tyrosine hydroxylase, the rate limiting enzyme of dopamine synthesis. MIAMI cells from young and old individuals represent autologous human cell populations for the treatment of disorders of the skeletal and nervous systems and for applications in cell therapy and reparative medicine approaches. Published by Elsevier Inc. Keywords: Bone marrow; Differentiation; Neurotrophin-3; Dopamine; Stem cell 1. Introduction The plasticity of bone marrow stem cells has been demonstrated by recent studies showing that subpopulations of marrow stromal cells (MSCs) appear to self-maintain throughout the organism's life; their progeny giving rise to a variety of tissues derived from other embryonic germ layers [1– 4]. Although human MSCs (hMSCs) represent a heterogeneous population of cells, subpopulations of hMSCs exhibiting Bone xx (2006) xxx – xxx + model BON-07574; No. of pages: 14; 4C: www.elsevier.com/locate/bone ⁎ Corresponding author. University of Miami School of Medicine, GRECC Adult Stem Cell Research Program, Veterans Affairs Medical Center-11GRC, 1201 NW 16th Street, Miami, FL 33125-1693, USA. Fax: +1 305 575 3365. E-mail address: [email protected] (P.C. Schiller). 1 These authors contributed equally to this work. 8756-3282/$ - see front matter. Published by Elsevier Inc. doi:10.1016/j.bone.2006.09.013 ARTICLE IN PRESS Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentiation of human adult marrow stromal cells to dopaminergic-like neurons, Bone (2006), doi:10.1016/j.bone.2006.09.013
Transcript of Neurotrophin-directed differentiation of human adult marrow stromal cells to dopaminergic-like...

xxx–xxx
+ model
BON-07574; No. of pages: 14; 4C:
www.elsevier.com/locate/bone
ARTICLE IN PRESS
Bone xx (2006)
Neurotrophin-directed differentiation of human adult marrow stromal cells todopaminergic-like neurons
Valerie M. Tatard a,1, Gianluca D'Ippolito c,e,1, Sylma Diabira a,b,c, Alexander Valeyev d,g,John Hackman d,g, Micheline McCarthy d,g,h, Thomas Bouckenooghe a, Philippe Menei a,b,
Claudia N. Montero-Menei a,1, Paul C. Schiller c,d,e,f,1,⁎
a INSERM Unit 646, Ingénierie de la Vectorisation Particulaire, 10, rue André Bocquel, 49100 Angers,Centre Hospitalier Universitaire d'Angers, 4 rue Larrey, France
b Département de Neurochirurgie, Centre Hospitalier Universitaire d'Angers, 4 rue Larrey, Francec Geriatric Research, Education, and Clinical Center and Research Service, USA
d Research Service, Veterans Affairs Medical Center, USAe Department of Medicine, University of Miami School of Medicine, Miami, FL 33125-1693, USA
f Department of Biochemistry and Molecular Biology, University of Miami School of Medicine, Miami, FL 33125-1693, USAg Department of Neurology, University of Miami School of Medicine, Miami, FL 33125-1693, USA
h Department of Microbiology and Immunology, University of Miami School of Medicine, Miami, FL 33125-1693, USA
Received 27 April 2006; revised 18 August 2006; accepted 5 September 2006
Abstract
Marrow-isolated adult multilineage inducible (MIAMI) cells were differentiated in vitro to neuronal cells in a neurotrophin-dependent fashion.After induction, the cells revealed electrophysiological features similar to those observed in mature neurons. Primary early passage human MIAMIcells without any type of co-cultures with other cell types were used. The developmental program involved a multi-step process requiring theconcerted action of brain-derived neurotrophic factor, nerve growth factor and depended on neurotrophin-3, after basic fibroblast growth factorwithdrawal. MIAMI-derived neuron-like cells sequentially expressed the neuronal markers, developed a complex neurite outgrowth andarborization, and acquired electrophysiological characteristics similar to those observed in mature neurons. The young and old MIAMI-derivedneuronal cells developed both inward and outward currents upon depolarization, similar to those observed in normal neurons. These resultsrepresent the earliest evidence that neurotrophin-3 can direct the differentiation of non-neural stem cells from human adult bone marrow stroma toneuron-like cells in vitro. Supplementing the aforementioned multi-step process with sonic hedgehog, fibroblast growth factor 8, and retinoic acidincreased the expression of molecules involved in dopaminergic differentiation and of tyrosine hydroxylase, the rate limiting enzyme of dopaminesynthesis. MIAMI cells from young and old individuals represent autologous human cell populations for the treatment of disorders of the skeletaland nervous systems and for applications in cell therapy and reparative medicine approaches.Published by Elsevier Inc.
Keywords: Bone marrow; Differentiation; Neurotrophin-3; Dopamine; Stem cell
⁎ Corresponding author. University of Miami School of Medicine, GRECCAdult Stem Cell Research Program, Veterans Affairs Medical Center-11GRC,1201 NW 16th Street, Miami, FL 33125-1693, USA. Fax: +1 305 575 3365.
E-mail address: [email protected] (P.C. Schiller).1 These authors contributed equally to this work.
8756-3282/$ - see front matter. Published by Elsevier Inc.doi:10.1016/j.bone.2006.09.013
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
1. Introduction
The plasticity of bone marrow stem cells has beendemonstrated by recent studies showing that subpopulationsof marrow stromal cells (MSCs) appear to self-maintainthroughout the organism's life; their progeny giving rise to avariety of tissues derived from other embryonic germ layers [1–4]. Although human MSCs (hMSCs) represent a heterogeneouspopulation of cells, subpopulations of hMSCs exhibiting
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone

2 V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
features of primitive adult pluripotent or multipotent stem cellshave been described [5,6]. We have previously reported thecharacterization of human marrow-isolated adult multilineageinducible (MIAMI) cells, a unique subpopulation of non-transformed non-immortalized adult marrow stromal cellscapable of differentiating in vitro into cell lineages derivedfrom all three germ layers [7]. MIAMI cells express uniquemarkers characteristic of developmentally primitive humanembryonic stem cells (hESCs).
Bone is a highly innervated tissue. Recently, it has beenreported that MSCs from rodents could be differentiated intocells with a neural phenotype in vitro [6,8–10]. These cellsunder specific neuron induction conditions expressed markersfound at the different stages of neuronal development. Jiang etal. [10] reported that murine marrow-derived multipotent adultprogenitor cells could be induced to differentiate into neuronal-like phenotype developing electrophysiological propertiessimilar to midbrain neurons. However, this final neuronalmaturation required co-culturing the cells with astrocytes. Inthis regard, a few rodent but also human cells seem to respond tothe environmental signals after transplantation in rodent's brainand give rise to neural cells without eliciting immune response[11–13].
Encouraging data from different groups suggest that MSCscould be strong candidates for cell and gene therapy [14] for avariety of skeletal as well as central nervous system disorders[15–17], particularly for Parkinson' disease (PD) [18,19]. Forcell therapy approaches, it is critical to obtain and use sufficientnumbers of phenotypically homogenous differentiated cellpopulations that have not been exposed to astrocytes,transformed, or co-cultured with other cell types. Clinical trialshave been attempted using fetal dopaminergic transplants in PD,characterized by the progressive loss of dopaminergic neurons.These studies gave some encouraging results, but ethical andpractical concerns like cell availability from fetal tissue limittheir development [20,21]. MIAMI cells could constitute analternative source of cells as they are easily purified and allowautografts to be performed. Recently, two relevant studies havereported the differentiation of human MSC into dopaminergicneuron-like cells. One study showed the differentiation of MSC,transduced with Notch intracellular domain. Approximately40% of the cells had features of mature neuronal cells withelectrophysiological activity, expressing a dopaminergic phe-notype. After their transplantation into an animal model of PD,they improved their behavior [18]. The second study reportedthe induction of approximately 40% of cells into tyrosinehydroxylase (TH)-positive cells using non-transfected humanMSC treated with trophic factors [19]. We are aware of only twopublished reports where non-transformed human MSCs havebeen differentiated in vitro to mature neuron-like cells showingsigns of electrophysiological activity. In one study, there waslittle evidence of ion channel activity in these neuron-like cells.[22]. The second study used retinoic acid to induce thedevelopment of a neuronal phenotype of cells showingspontaneous action potentials without evidence of sodium andpotassium currents. Furthermore, these cells also showedspontaneous postsynaptic currents when voltage clamped
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
[23]. Recently, Wenisch et al. demonstrated that hMSCsdevelop cellular features of neuritogenesis and synaptogenesisin the course of neuronal induction and suggested that the cellshad features similar to those observed in immature neuronswithout electrophysiological activity.
In this study, we demonstrate for the first time thedifferentiation of human marrow-derived cells from youngand old donors, the MIAMI cells, to electrophysiologicallycompetent neuron-like cells, with resting membrane potentials,that are not spontaneously active, and when voltage clampedand depolarized they display ionic currents that are neuron-likein nature. Differentiation is dependent on expansion ofprogenitor cells under low oxygen tension and neuronalinduction requiring continuous treatment with NT-3 and thesequential use of other neurotrophic factors, without co-culturing with any other type of cell. These results advancethe novel concept that NT-3 acting most likely through NTRK-3plays a key role in directing the neuronal differentiation of non-neural early passage human stromal stem cells. After exposureto the neuronal induction protocol, the MIAMI cells undergo aneuronal developmental program resembling that of neural stemcells. A slightly modified protocol yields neuron-like cells witha dopaminergic phenotype. Moreover, these neuron-like cellsexpressed several molecules involved in dopaminergic celldifferentiation at the expected stages. These results stronglysuggest an authentic lineage differentiation rather than abiochemical manipulation. MIAMI cells may therefore con-stitute a population of human cells suitable for the treatment ofdisorders of the skeletal and nervous system in young and oldindividuals.
2. Materials and methods
2.1. Cytokines
Human recombinant basic fibroblast growth factor (bFGF) and all-transretinoic acid were purchased from Sigma Chemical (St. Louis, MO); β-nervegrowth factor (NGF), neurotrophin-3 (NT-3), and brain-derived neurotrophicfactor (BDNF), from Calbiochem (San Diego, CA). Sonic hedgehog (SHH),fibroblast growth factor 8 (FGF-8), and glial cell line derived neurotrophic factor(GDNF) were purchased from R&D Systems Europe (Lille, France).
2.2. Antibodies
The following primary antibodies were used: for immunocytochemistry(dilution from 1:100 to 1:1000) — neuron-specific enolase (NSE), neurofila-ment-L (68 kDa), neurofilament-M (160 kDa), nestin, neuronal nuclear protein(NeuN), glial fibrillary acid protein (GFAP), synaptophysin and neurotrophintyrosine kinase receptor type I (NTRK-1/TrkA/NGF receptor) were fromChemicon (Temecula, CA); neuron-specific class III β-tubulin (TuJ1) was fromCovance (Princeton, NJ), neurotrophin tyrosine kinase receptor type III (NTRK-3/TrkC) was from Santa Cruz Biotechnology (Santa Cruz, CA), and tyrosinehydroxylase (TH) from Jacques Boy (Reims, France). Horseradish peroxidase(HRP), rhodamine, and FITC conjugated secondary antibodies were from SantaCruz Biotechnology (Santa Cruz, CA).
2.3. Bone marrow and cells
Whole bone marrow was obtained from vertebral bodies (T1–L5) of 12cadaveric donors (6 males and 6 females) who died of fatal traumatic injury (agerange; 3–72 years old), following guidelines for informed consent set by the
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone

3V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
University of Miami School of Medicine Committee on the Use of HumanSubjects in Research. Cells from individual donors were cultured separately.
2.4. Selection/expansion of MIAMI cells
As previously described [7], isolated whole bone marrow cells were plated ata constant density of 105 cells/cm2 in DMEM-low glucose media, containing 5%fetal bovine serum (FBS) and 100 U penicillin/1000 U streptomycin infibronectin (10 ng/ml)-coated 10-cm dishes. Whole bone marrow cells,containing adherent and nonadherent cells, were maintained in a humidifiedincubator at 37°C undisturbed in an atmosphere of 3% O2/5% CO2/92% N2;7 days later, half of the culture medium was replaced. Fourteen days after theinitial plating, the nonadherent cells were removed. Single-cell-derived (isolatedusing cloning rings) and pooled colonies of adherent cells were carefully rinsedin medium and subcloned. These cells were selected and expanded at lowdensity (≤30% confluency) by plating single-cell-derived or pooled colonies infibronectin (10 ng/ml)-coated dishes using an expansion medium composed of:98% DMEM-low glucose, 2% FBS (lot selected), or DMEM supplemented with40% MCDB-201 medium (Sigma, M6770) and insulin–transferrin–selenium(Sigma, I3146) and 100 U penicillin/1000 U streptomycin (Gibco InvitrogenCorp, Grand Island, NY) both at 3% O2/5% CO2/92% N2.
2.5. Neuronal induction
For neuronal induction, morphologically homogeneous single-cell- ormulticolony-derived MIAMI cells passage 3 to 9 were seeded at 2500–5000 cells/cm2 in 35 mm dishes (Costar) coated fibronectin (10 ng/ml-coatedcoverslips) in DMEM-high glucose (DMEM-HG) supplemented with 5 or20% FBS, 100 U/ml penicillin, and 1 mg/ml streptomycin and cultured for24 h. Neural specification (step 1) was induced by exposing cells to DMEM-HG/5 or 20% FBS/10 ng/ml bFGF for 24 h. At the end of the neuralspecification treatment, cells were washed twice with PBS, and then neuronalcommitment (step 2) was induced by exposing the cells to DMEM-HG/1 mMβ-mercaptoethanol (βME)/30 ng/ml NT-3 for 2 days. For some experiments,bFGF was maintained during step 2 to increase the number of neuralprecursors and βME concentration was decreased to 100 μM to enhanceviability. Finally, we induced neuronal differentiation (step 3) by first washingtwice with PBS and then exposing the cells to 100–200 μM butylatedhydroxyanisole (BHA, Sigma), 25 mM KCl, 2 mM valproic acid, 4 μMforskolin, 1 μM hydrocortisone, 5 μM insulin, NT-3 (30 ng/ml), NGF (10 ng/ml), and BDNF (50 ng/ml) in DMEM-HG for 3 to 7 days. Importantly, allsolutions were freshly prepared immediately prior to use. Neuronal inductionwas performed at 20% oxygen.
For induction of a dopaminergic phenotype, cells were exposed to SHH(200 ng/ml), FGF-8 (100 ng/ml), and retinoic acid (RA) (0.5 μM) (obtainedfrom Sigma, Saint Quentin Fallavier, France) during neuronal commitment (step2). To improve dopaminergic neuron-like survival, GDNF (10 ng/ml) was addedto the neuronal induction media (step 3). Three different experiments in triplicatewere done, and five fields per slide at 10× objective were counted.
2.6. Human fetal neuroepithelial precursor cells
Human fetal neuroepithelial precursor cells were used as positive controlfor the expression of the intermediate filament protein nestin. Nestin isexpressed by multipotent neuroepithelial cells and by glial or neuronal lineageprecursor cells [24,25] (Fig. 1E). Neuronal lineage committed cells expressclass III β-tubulin (TuJ1), and post-mitotic neurons express the 68 kDaintermediate filament protein neurofilament-L (not shown). Multipotentneuroepithelial stem cells were prepared from dissociated first trimestergestation fetal human CNS as described previously [26]. Fetal CNS tissuespecimens included telencephalon, diencephalon, and rostral brainstem.Procedures for procurement and use of this human fetal CNS tissue wereapproved and monitored by the University of Miami School of Medicine'sMedical Sciences Subcommittee for the Protection of Human Subjects. Cellsderived from distinct fetal specimens were cultured separately; there was noco-culturing of cells from different specimens. Single-cell suspensions werecultured in defined medium supplemented with EGF and bFGF (Neural
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
Progenitor Maintenance Medium [NPMM] Bullet Kit, Clonetics Division ofBioWhittaker, San Diego, CA) or Neurobasal medium plus B27 supplement[27] (Gibco Invitrogen Corp.). These cell suspensions generated proliferatingclones of neuroepithelial precursor cells in floating spheres (“neurospheres”),which were cultured for 7–10 days. To initiate differentiation of precursorcells, the spheres were seeded on an adherent substrate (poly-D, L-ornithineplus fibronectin-coated glass, PO/FBN) in DMEM/F12 medium supplementedwith N2 supplement (Gibco Invitrogen Corp.) and 2.5% (v/v) heat-inactivatedFBS. Proliferating and differentiating neuroepithelial precursor cells migratedout from adherent spheres so that a monolayer culture developed within 1 daypost seeding.
2.7. Immunocytochemistry
Uninduced and induced cells at all steps of neuronal induction were used forimmunocytochemistry. For staining of cytoskeletal proteins at the indicatedtimes, cells were rinsed twice with cold Dulbecco's phosphate-buffered saline(DPBS), pH 7.4, fixed with methanol at −20°C for 2 min and thenpermeabilized with 0.1% Triton X-100 for 10 min. For other intracellularmolecules, cells were fixed with 4% paraformaldehyde at 4°C for 10 min andthen permeabilized with 0.1% Triton X-100 for 10 min. For cell surfacereceptors, cells were fixed with 4% paraformaldehyde at 4°C for 10 min. Slideswere blocked for 30 min. Blocking solution consisted of PBS, 1% bovine serumalbumin (BSA), and 1% serum (Sigma) from the species in which the secondaryantibody was raised. Slides were then incubated sequentially for 30 min eachwith primary specific antibodies or isotypic IgG antibody (controls) followed byincubation with HRP-, FITC- or rhodamine-conjugated secondary antibodies.Between each step, slides were washed with PBS plus 0.3% BSA. Specificimmunostaining was demonstrated in control experiments in which cells wereexposed to primary isotypic antibodies and then incubated with conjugatedantibodies (data shown only for nestin in Fig. 1, but performed in everyexperiment). DAPI (4′,6-diamidino-2-phenylindole dihydrochloride) was pre-sent in the mounting solution. To examine the cells, we used a Nikon microscope(connected to a Retiga 1300 color digital camera and imaging system) or aconfocal fluorescence microscope (Zeiss LSM 510 laser scanning confocalmicroscope). Cells with neural morphological characteristics were expressed asa percentage of total MIAMI cells counted± the standard deviation (percentageof total MIAMI cells±SD).
2.8. RNA isolation and analysis
Before induction and at each step of the dopaminergic differentiationprotocol, MIAMI cells were lysed in a 1% β-mercaptoethanol containingbuffer obtained from an RNA extraction kit (Macherey Nagel, Hoerdt,France). RNA was extracted as described by the manufacturer and cDNAsynthesized using random hexamers (Amersham Biosciences, Orsay,France). Semi-quantitative noncompetitive RT-PCR was performed withBioTaq (AbCys, Paris, France). Amplification was achieved with human-specific primer pairs of the target sequence, as described in Table 1. PCRwas performed with 3 μg of cDNA from each sample in the presence of200 μM deoxynucleotide triphosphate, 1.5 mM MgCl2, 5 U Taq DNApolymerase, and 25 pM of each primer. PCR was carried out in athermal cycler (MyCycler, Biorad, Marnes-la-Coquette, France) with eachcycle consisting of denaturation at 94°C for 60 s, annealing at 60°C for60 s, and polymerization at 72°C for 60 s. The last PCR step was afinal extension at 72°C for 10 min. The PCR products wereelectrophoresed in 2% agarose gel, and bands were scanned with anintegration camera CDD (Fisher Bioblock Scientific, Illkirch, France).Positive controls were performed with the appropriate human fetalneuroepithelial precursor cells.
2.9. ELISA analysis
CNTF, TGF-β1, and TGF-β3 ELISA kits were purchased from R&DSystems, and the analysis was performed according to the manufacturer'sinstructions for uninduced cells and throughout the dopaminergic neuronalinduction protocol.
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone
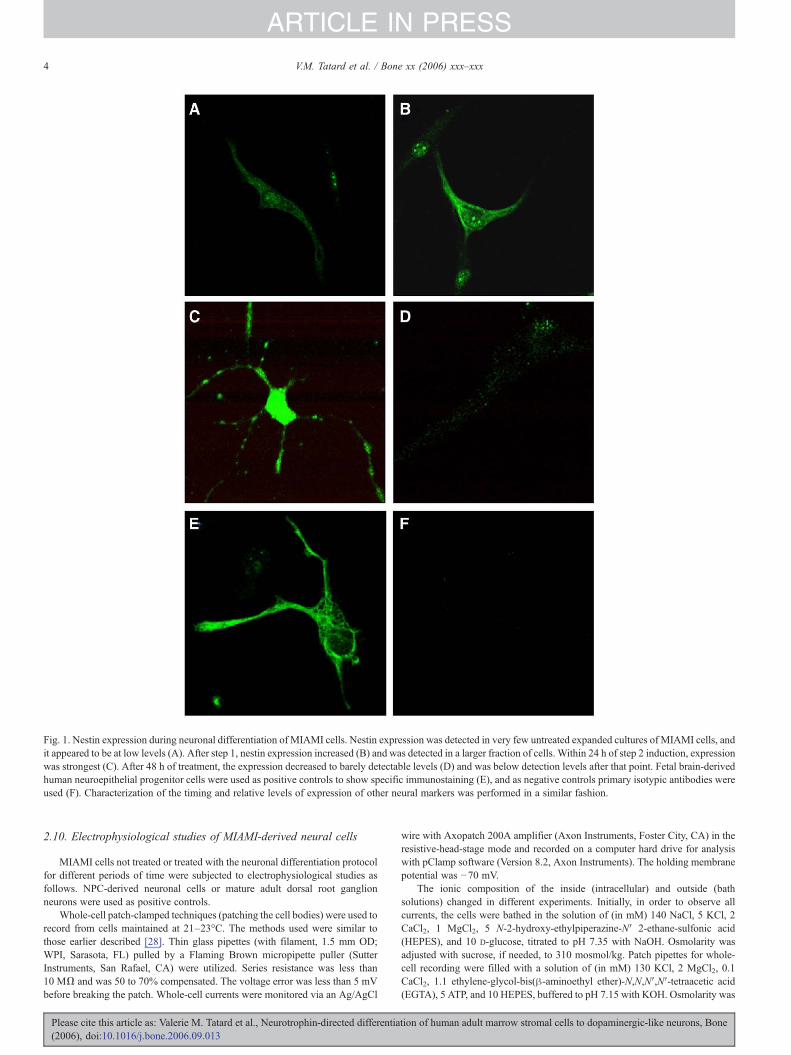
Fig. 1. Nestin expression during neuronal differentiation of MIAMI cells. Nestin expression was detected in very few untreated expanded cultures of MIAMI cells, andit appeared to be at low levels (A). After step 1, nestin expression increased (B) and was detected in a larger fraction of cells. Within 24 h of step 2 induction, expressionwas strongest (C). After 48 h of treatment, the expression decreased to barely detectable levels (D) and was below detection levels after that point. Fetal brain-derivedhuman neuroepithelial progenitor cells were used as positive controls to show specific immunostaining (E), and as negative controls primary isotypic antibodies wereused (F). Characterization of the timing and relative levels of expression of other neural markers was performed in a similar fashion.
4 V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
2.10. Electrophysiological studies of MIAMI-derived neural cells
MIAMI cells not treated or treated with the neuronal differentiation protocolfor different periods of time were subjected to electrophysiological studies asfollows. NPC-derived neuronal cells or mature adult dorsal root ganglionneurons were used as positive controls.
Whole-cell patch-clamped techniques (patching the cell bodies) were used torecord from cells maintained at 21–23°C. The methods used were similar tothose earlier described [28]. Thin glass pipettes (with filament, 1.5 mm OD;WPI, Sarasota, FL) pulled by a Flaming Brown micropipette puller (SutterInstruments, San Rafael, CA) were utilized. Series resistance was less than10 MΩ and was 50 to 70% compensated. The voltage error was less than 5 mVbefore breaking the patch. Whole-cell currents were monitored via an Ag/AgCl
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
wire with Axopatch 200A amplifier (Axon Instruments, Foster City, CA) in theresistive-head-stage mode and recorded on a computer hard drive for analysiswith pClamp software (Version 8.2, Axon Instruments). The holding membranepotential was −70 mV.
The ionic composition of the inside (intracellular) and outside (bathsolutions) changed in different experiments. Initially, in order to observe allcurrents, the cells were bathed in the solution of (in mM) 140 NaCl, 5 KCl, 2CaCl2, 1 MgCl2, 5 N-2-hydroxy-ethylpiperazine-N′ 2-ethane-sulfonic acid(HEPES), and 10 D-glucose, titrated to pH 7.35 with NaOH. Osmolarity wasadjusted with sucrose, if needed, to 310 mosmol/kg. Patch pipettes for whole-cell recording were filled with a solution of (in mM) 130 KCl, 2 MgCl2, 0.1CaCl2, 1.1 ethylene-glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid(EGTA), 5 ATP, and 10 HEPES, buffered to pH 7.15 with KOH. Osmolarity was
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone
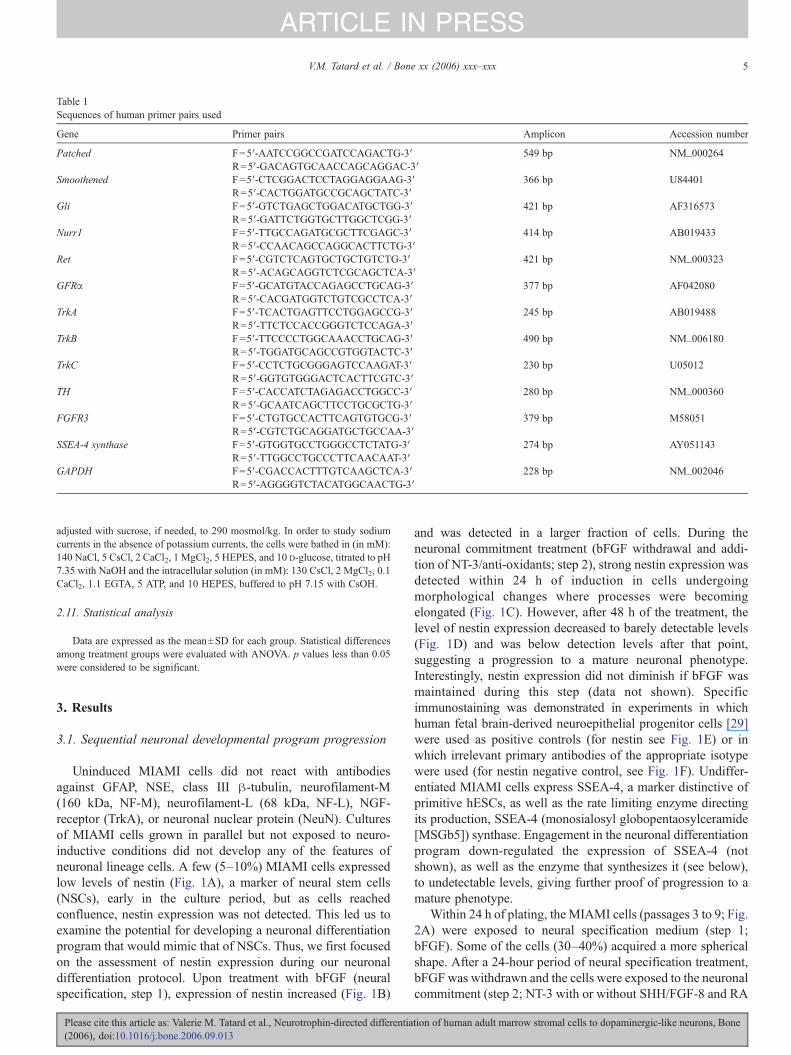
Table 1Sequences of human primer pairs used
Gene Primer pairs Amplicon Accession number
Patched F=5′-AATCCGGCCGATCCAGACTG-3′ 549 bp NM_000264R=5′-GACAGTGCAACCAGCAGGAC-3′
Smoothened F=5′-CTCGGACTCCTAGGAGGAAG-3′ 366 bp U84401R=5′-CACTGGATGCCGCAGCTATC-3′
Gli F=5′-GTCTGAGCTGGACATGCTGG-3′ 421 bp AF316573R=5′-GATTCTGGTGCTTGGCTCGG-3′
Nurr1 F=5′-TTGCCAGATGCGCTTCGAGC-3′ 414 bp AB019433R=5′-CCAACAGCCAGGCACTTCTG-3′
Ret F=5′-CGTCTCAGTGCTGCTGTCTG-3′ 421 bp NM_000323R=5′-ACAGCAGGTCTCGCAGCTCA-3′
GFRα F=5′-GCATGTACCAGAGCCTGCAG-3′ 377 bp AF042080R=5′-CACGATGGTCTGTCGCCTCA-3′
TrkA F=5′-TCACTGAGTTCCTGGAGCCG-3′ 245 bp AB019488R=5′-TTCTCCACCGGGTCTCCAGA-3′
TrkB F=5′-TTCCCCTGGCAAACCTGCAG-3′ 490 bp NM_006180R=5′-TGGATGCAGCCGTGGTACTC-3′
TrkC F=5′-CCTCTGCGGGAGTCCAAGAT-3′ 230 bp U05012R=5′-GGTGTGGGACTCACTTCGTC-3′
TH F=5′-CACCATCTAGAGACCTGGCC-3′ 280 bp NM_000360R=5′-GCAATCAGCTTCCTGCGCTG-3′
FGFR3 F=5′-CTGTGCCACTTCAGTGTGCG-3′ 379 bp M58051R=5′-CGTCTGCAGGATGCTGCCAA-3′
SSEA-4 synthase F=5′-GTGGTGCCTGGGCCTCTATG-3′ 274 bp AY051143R=5′-TTGGCCTGCCCTTCAACAAT-3′
GAPDH F=5′-CGACCACTTTGTCAAGCTCA-3′ 228 bp NM_002046R=5′-AGGGGTCTACATGGCAACTG-3′
5V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
adjusted with sucrose, if needed, to 290 mosmol/kg. In order to study sodiumcurrents in the absence of potassium currents, the cells were bathed in (in mM):140 NaCl, 5 CsCl, 2 CaCl2, 1 MgCl2, 5 HEPES, and 10 D-glucose, titrated to pH7.35 with NaOH and the intracellular solution (in mM): 130 CsCl, 2 MgCl2, 0.1CaCl2, 1.1 EGTA, 5 ATP, and 10 HEPES, buffered to pH 7.15 with CsOH.
2.11. Statistical analysis
Data are expressed as the mean±SD for each group. Statistical differencesamong treatment groups were evaluated with ANOVA. p values less than 0.05were considered to be significant.
3. Results
3.1. Sequential neuronal developmental program progression
Uninduced MIAMI cells did not react with antibodiesagainst GFAP, NSE, class III β-tubulin, neurofilament-M(160 kDa, NF-M), neurofilament-L (68 kDa, NF-L), NGF-receptor (TrkA), or neuronal nuclear protein (NeuN). Culturesof MIAMI cells grown in parallel but not exposed to neuro-inductive conditions did not develop any of the features ofneuronal lineage cells. A few (5–10%) MIAMI cells expressedlow levels of nestin (Fig. 1A), a marker of neural stem cells(NSCs), early in the culture period, but as cells reachedconfluence, nestin expression was not detected. This led us toexamine the potential for developing a neuronal differentiationprogram that would mimic that of NSCs. Thus, we first focusedon the assessment of nestin expression during our neuronaldifferentiation protocol. Upon treatment with bFGF (neuralspecification, step 1), expression of nestin increased (Fig. 1B)
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
and was detected in a larger fraction of cells. During theneuronal commitment treatment (bFGF withdrawal and addi-tion of NT-3/anti-oxidants; step 2), strong nestin expression wasdetected within 24 h of induction in cells undergoingmorphological changes where processes were becomingelongated (Fig. 1C). However, after 48 h of the treatment, thelevel of nestin expression decreased to barely detectable levels(Fig. 1D) and was below detection levels after that point,suggesting a progression to a mature neuronal phenotype.Interestingly, nestin expression did not diminish if bFGF wasmaintained during this step (data not shown). Specificimmunostaining was demonstrated in experiments in whichhuman fetal brain-derived neuroepithelial progenitor cells [29]were used as positive controls (for nestin see Fig. 1E) or inwhich irrelevant primary antibodies of the appropriate isotypewere used (for nestin negative control, see Fig. 1F). Undiffer-entiated MIAMI cells express SSEA-4, a marker distinctive ofprimitive hESCs, as well as the rate limiting enzyme directingits production, SSEA-4 (monosialosyl globopentaosylceramide[MSGb5]) synthase. Engagement in the neuronal differentiationprogram down-regulated the expression of SSEA-4 (notshown), as well as the enzyme that synthesizes it (see below),to undetectable levels, giving further proof of progression to amature phenotype.
Within 24 h of plating, the MIAMI cells (passages 3 to 9; Fig.2A) were exposed to neural specification medium (step 1;bFGF). Some of the cells (30–40%) acquired a more sphericalshape. After a 24-hour period of neural specification treatment,bFGF was withdrawn and the cells were exposed to the neuronalcommitment (step 2; NT-3 with or without SHH/FGF-8 and RA
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone
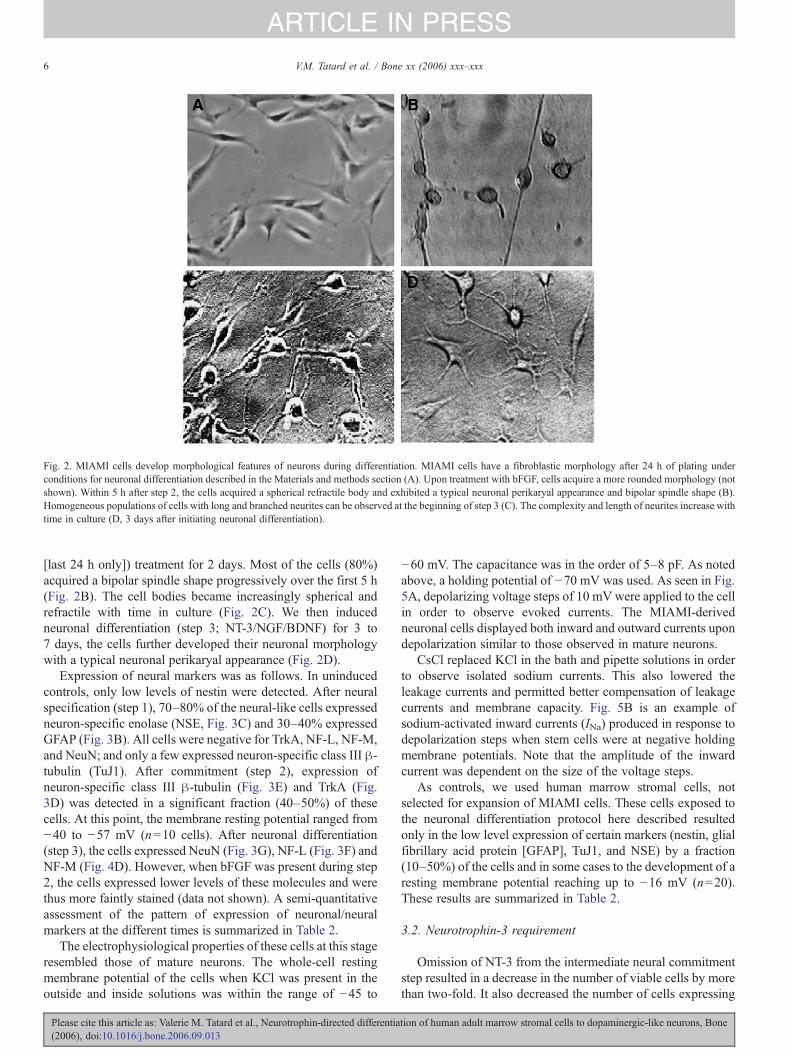
Fig. 2. MIAMI cells develop morphological features of neurons during differentiation. MIAMI cells have a fibroblastic morphology after 24 h of plating underconditions for neuronal differentiation described in the Materials and methods section (A). Upon treatment with bFGF, cells acquire a more rounded morphology (notshown). Within 5 h after step 2, the cells acquired a spherical refractile body and exhibited a typical neuronal perikaryal appearance and bipolar spindle shape (B).Homogeneous populations of cells with long and branched neurites can be observed at the beginning of step 3 (C). The complexity and length of neurites increase withtime in culture (D, 3 days after initiating neuronal differentiation).
6 V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
[last 24 h only]) treatment for 2 days. Most of the cells (80%)acquired a bipolar spindle shape progressively over the first 5 h(Fig. 2B). The cell bodies became increasingly spherical andrefractile with time in culture (Fig. 2C). We then inducedneuronal differentiation (step 3; NT-3/NGF/BDNF) for 3 to7 days, the cells further developed their neuronal morphologywith a typical neuronal perikaryal appearance (Fig. 2D).
Expression of neural markers was as follows. In uninducedcontrols, only low levels of nestin were detected. After neuralspecification (step 1), 70–80% of the neural-like cells expressedneuron-specific enolase (NSE, Fig. 3C) and 30–40% expressedGFAP (Fig. 3B). All cells were negative for TrkA, NF-L, NF-M,and NeuN; and only a few expressed neuron-specific class III β-tubulin (TuJ1). After commitment (step 2), expression ofneuron-specific class III β-tubulin (Fig. 3E) and TrkA (Fig.3D) was detected in a significant fraction (40–50%) of thesecells. At this point, the membrane resting potential ranged from−40 to −57 mV (n=10 cells). After neuronal differentiation(step 3), the cells expressed NeuN (Fig. 3G), NF-L (Fig. 3F) andNF-M (Fig. 4D). However, when bFGF was present during step2, the cells expressed lower levels of these molecules and werethus more faintly stained (data not shown). A semi-quantitativeassessment of the pattern of expression of neuronal/neuralmarkers at the different times is summarized in Table 2.
The electrophysiological properties of these cells at this stageresembled those of mature neurons. The whole-cell restingmembrane potential of the cells when KCl was present in theoutside and inside solutions was within the range of −45 to
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
−60 mV. The capacitance was in the order of 5–8 pF. As notedabove, a holding potential of −70 mV was used. As seen in Fig.5A, depolarizing voltage steps of 10 mV were applied to the cellin order to observe evoked currents. The MIAMI-derivedneuronal cells displayed both inward and outward currents upondepolarization similar to those observed in mature neurons.
CsCl replaced KCl in the bath and pipette solutions in orderto observe isolated sodium currents. This also lowered theleakage currents and permitted better compensation of leakagecurrents and membrane capacity. Fig. 5B is an example ofsodium-activated inward currents (INa) produced in response todepolarization steps when stem cells were at negative holdingmembrane potentials. Note that the amplitude of the inwardcurrent was dependent on the size of the voltage steps.
As controls, we used human marrow stromal cells, notselected for expansion of MIAMI cells. These cells exposed tothe neuronal differentiation protocol here described resultedonly in the low level expression of certain markers (nestin, glialfibrillary acid protein [GFAP], TuJ1, and NSE) by a fraction(10–50%) of the cells and in some cases to the development of aresting membrane potential reaching up to −16 mV (n=20).These results are summarized in Table 2.
3.2. Neurotrophin-3 requirement
Omission of NT-3 from the intermediate neural commitmentstep resulted in a decrease in the number of viable cells by morethan two-fold. It also decreased the number of cells expressing
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone
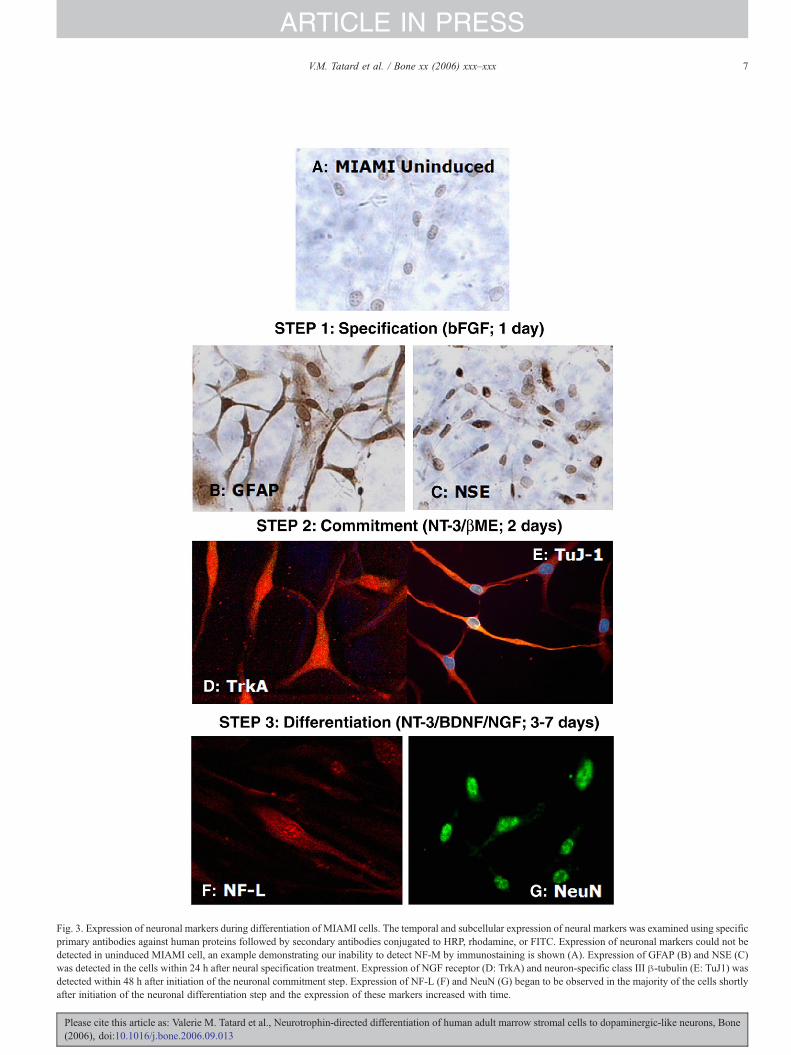
Fig. 3. Expression of neuronal markers during differentiation of MIAMI cells. The temporal and subcellular expression of neural markers was examined using specificprimary antibodies against human proteins followed by secondary antibodies conjugated to HRP, rhodamine, or FITC. Expression of neuronal markers could not bedetected in uninduced MIAMI cell, an example demonstrating our inability to detect NF-M by immunostaining is shown (A). Expression of GFAP (B) and NSE (C)was detected in the cells within 24 h after neural specification treatment. Expression of NGF receptor (D: TrkA) and neuron-specific class III β-tubulin (E: TuJ1) wasdetected within 48 h after initiation of the neuronal commitment step. Expression of NF-L (F) and NeuN (G) began to be observed in the majority of the cells shortlyafter initiation of the neuronal differentiation step and the expression of these markers increased with time.
7V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentiation of human adult marrow stromal cells to dopaminergic-like neurons, Bone(2006), doi:10.1016/j.bone.2006.09.013
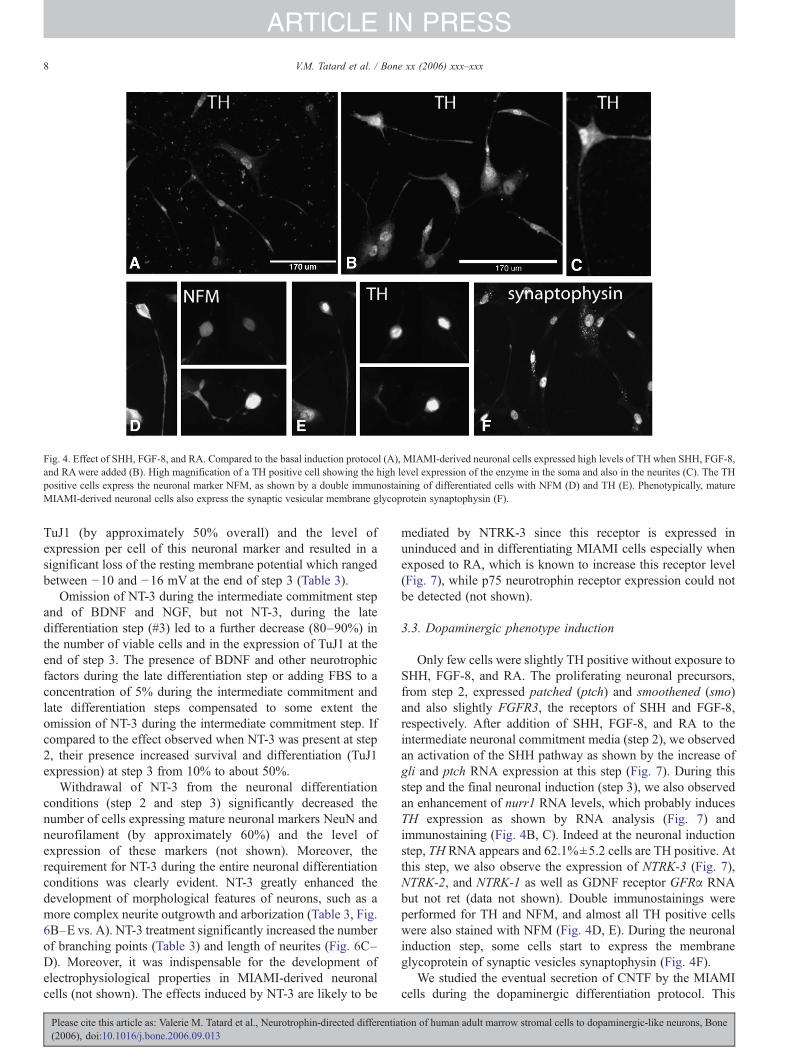
Fig. 4. Effect of SHH, FGF-8, and RA. Compared to the basal induction protocol (A), MIAMI-derived neuronal cells expressed high levels of TH when SHH, FGF-8,and RAwere added (B). High magnification of a TH positive cell showing the high level expression of the enzyme in the soma and also in the neurites (C). The THpositive cells express the neuronal marker NFM, as shown by a double immunostaining of differentiated cells with NFM (D) and TH (E). Phenotypically, matureMIAMI-derived neuronal cells also express the synaptic vesicular membrane glycoprotein synaptophysin (F).
8 V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
TuJ1 (by approximately 50% overall) and the level ofexpression per cell of this neuronal marker and resulted in asignificant loss of the resting membrane potential which rangedbetween −10 and −16 mV at the end of step 3 (Table 3).
Omission of NT-3 during the intermediate commitment stepand of BDNF and NGF, but not NT-3, during the latedifferentiation step (#3) led to a further decrease (80–90%) inthe number of viable cells and in the expression of TuJ1 at theend of step 3. The presence of BDNF and other neurotrophicfactors during the late differentiation step or adding FBS to aconcentration of 5% during the intermediate commitment andlate differentiation steps compensated to some extent theomission of NT-3 during the intermediate commitment step. Ifcompared to the effect observed when NT-3 was present at step2, their presence increased survival and differentiation (TuJ1expression) at step 3 from 10% to about 50%.
Withdrawal of NT-3 from the neuronal differentiationconditions (step 2 and step 3) significantly decreased thenumber of cells expressing mature neuronal markers NeuN andneurofilament (by approximately 60%) and the level ofexpression of these markers (not shown). Moreover, therequirement for NT-3 during the entire neuronal differentiationconditions was clearly evident. NT-3 greatly enhanced thedevelopment of morphological features of neurons, such as amore complex neurite outgrowth and arborization (Table 3, Fig.6B–E vs. A). NT-3 treatment significantly increased the numberof branching points (Table 3) and length of neurites (Fig. 6C–D). Moreover, it was indispensable for the development ofelectrophysiological properties in MIAMI-derived neuronalcells (not shown). The effects induced by NT-3 are likely to be
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
mediated by NTRK-3 since this receptor is expressed inuninduced and in differentiating MIAMI cells especially whenexposed to RA, which is known to increase this receptor level(Fig. 7), while p75 neurotrophin receptor expression could notbe detected (not shown).
3.3. Dopaminergic phenotype induction
Only few cells were slightly TH positive without exposure toSHH, FGF-8, and RA. The proliferating neuronal precursors,from step 2, expressed patched (ptch) and smoothened (smo)and also slightly FGFR3, the receptors of SHH and FGF-8,respectively. After addition of SHH, FGF-8, and RA to theintermediate neuronal commitment media (step 2), we observedan activation of the SHH pathway as shown by the increase ofgli and ptch RNA expression at this step (Fig. 7). During thisstep and the final neuronal induction (step 3), we also observedan enhancement of nurr1 RNA levels, which probably inducesTH expression as shown by RNA analysis (Fig. 7) andimmunostaining (Fig. 4B, C). Indeed at the neuronal inductionstep, TH RNA appears and 62.1%±5.2 cells are TH positive. Atthis step, we also observe the expression of NTRK-3 (Fig. 7),NTRK-2, and NTRK-1 as well as GDNF receptor GFRα RNAbut not ret (data not shown). Double immunostainings wereperformed for TH and NFM, and almost all TH positive cellswere also stained with NFM (Fig. 4D, E). During the neuronalinduction step, some cells start to express the membraneglycoprotein of synaptic vesicles synaptophysin (Fig. 4F).
We studied the eventual secretion of CNTF by the MIAMIcells during the dopaminergic differentiation protocol. This
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone

Table 2Fraction of cells expressing particular neuronal/neural features at specific times in comparison with control populations
Neuronal/Neural feature Uninduced controls Step 1 Step 2 Step 3 Treated hMSCs a
Nestin 5–10%b 90% 80–90%c ND LowGFAP NDd 30–40% ND ND LowNSE ND 70–80% ND ND 5–40%Class III β-tubulin ND Lowe 40–50% 50–60% 5–15%TrkA ND ND 40–50% 50–60% NDNF-M ND ND 5–10% 50–60% NDNF-L ND ND ND 50–60% NDNeuN ND ND Low 60–80% NDSpherical shape ND 30–40% − − 3–5%Bipolar spindle shape ND ND 80% 80–90% 5–30%Resting membrane potential (mV) ND ND −40 to −57 −45 to −60 up to −16Inward and outward currents ND ND ND Yes NDa Highest levels observed for marrow stromal cells isolated using standard procedures, not the MIAMI cell conditions, at any point during treatment steps 1 to 3.b The number represents the range of the percentage of cells strongly expressing the feature in control or treated MIAMI cell populations isolated from four to eight
different individuals.c The expression of nestin was very high at the onset of step 2, but dramatically decreased toward the end of step 2.d ND: not detected.e Low: indicates that a small fraction of the cells (1–5%) weakly expressed the protein marker.
9V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
cytokine is implicated in the differentiation of astrocyteswhich are involved in the dopaminergic differentiation ofneurons. No CNTF secretion was detected. We have alsostudied TGF-β secretion as it was described that TGF-β mayact with SHH and FGF-8 to induce dopaminergic differentiationof midbrain precursors. Uninduced MIAMI cells secrete TGF-β1 (408±179 pg/ml), they stop producing it during neuronalcommitment and secrete (155±23 pg/ml) again during theneuronal induction step 3. No TGF-β3 secretion was detected.
3.4. Age-dependent responses
MIAMI cells isolated from donors of various ages (#519 [3-year-old male], #869 [55-year-old male], #812 [72-year-oldmale], and #889 [59-year-old female]) were used for theneuronal differentiation experiments described above, and allresponded in a similar fashion. Single-cell-derived coloniesfrom MIAMI cells isolated from sample #519 were expandedunder conditions described and tested for their ability to
Fig. 5. Voltage-activated ion currents in MIAMI-derived neuronal cells. (A) Inwardsample #519 (3-year-old male). Ion currents were recorded without using compensatfrom cultured #519 MIAMI-derived neuronal cells. Leak currents have been subtracA and B show the depolarizing voltage steps. Cells were whole-cell patch-clampeactivated by 10 mV depolarizing steps. (C) Inward and outward currents recorded
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
differentiate toward the ectodermal-derived neuronal pheno-type. These cells responded to neuronal differentiation in amanner similar to that described. Development of themorphological characteristic, sequential expression of neuronalproteins, and acquisition of resting membrane potential within arange of −45 to −60 mV was consistently observed (70–90%).However, acquisition of inward and outward currents wasobserved only in about 20% of the inductions from the samedonors within 7–10 days of initiating the neuronal induction ofMIAMI cells isolated from donors of all ages. At this point intime, the reason for this is unclear.
4. Discussion
We report here that a subpopulation of human bone marrow-derived stromal cells, the MIAMI cells, can be induced todifferentiate into neuron-like cells with characteristics similar tothose of central nervous system mature neurons. The sequentialexpression of markers of MIAMI cells, isolated from either
and outward currents were recorded in the MIAMI-derived neuronal cells fromion. (B) Potential-activated inward currents recorded in whole-cell configurationted from signals obtained during the voltage steps commands. Arrows in panelsd techniques at the holding membrane potential −70 mV, and currents werein MIAMI-derived neuronal cells from sample #869 (55-year-old male).
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone
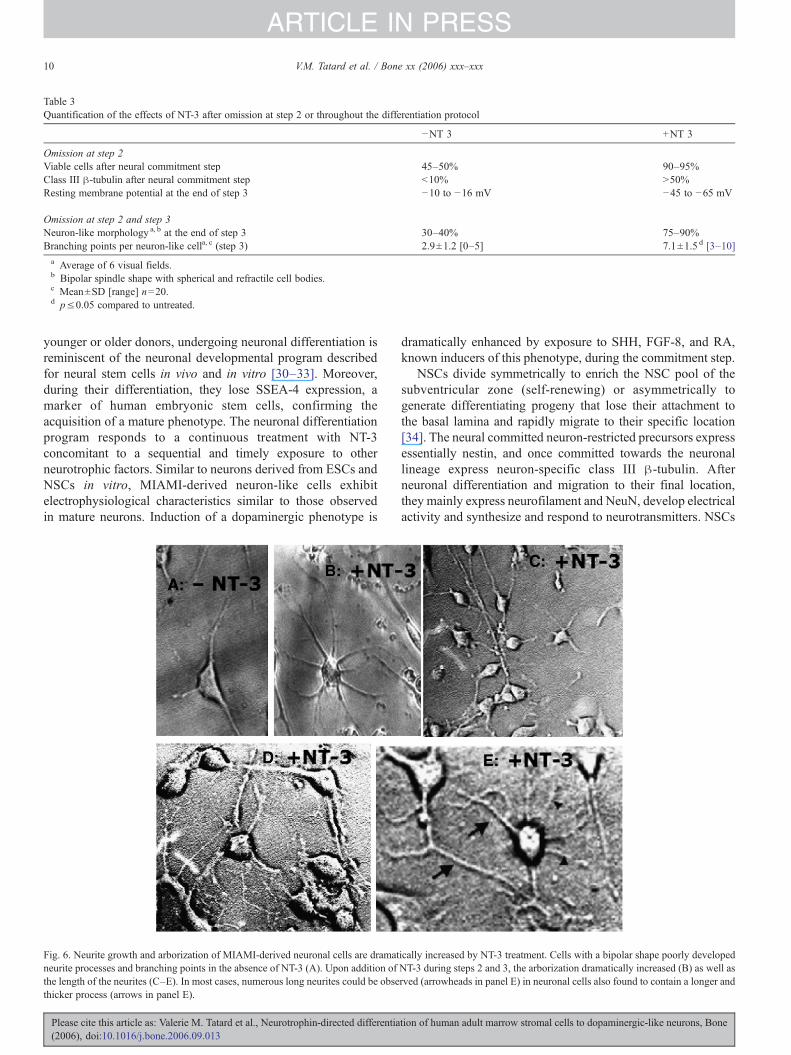
Table 3Quantification of the effects of NT-3 after omission at step 2 or throughout the differentiation protocol
−NT 3 +NT 3
Omission at step 2Viable cells after neural commitment step 45–50% 90–95%Class III β-tubulin after neural commitment step <10% >50%Resting membrane potential at the end of step 3 −10 to −16 mV −45 to −65 mV
Omission at step 2 and step 3Neuron-like morphology a, b at the end of step 3 30–40% 75–90%Branching points per neuron-like cella, c (step 3) 2.9±1.2 [0–5] 7.1±1.5 d [3–10]a Average of 6 visual fields.b Bipolar spindle shape with spherical and refractile cell bodies.c Mean±SD [range] n=20.d p≤0.05 compared to untreated.
10 V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
younger or older donors, undergoing neuronal differentiation isreminiscent of the neuronal developmental program describedfor neural stem cells in vivo and in vitro [30–33]. Moreover,during their differentiation, they lose SSEA-4 expression, amarker of human embryonic stem cells, confirming theacquisition of a mature phenotype. The neuronal differentiationprogram responds to a continuous treatment with NT-3concomitant to a sequential and timely exposure to otherneurotrophic factors. Similar to neurons derived from ESCs andNSCs in vitro, MIAMI-derived neuron-like cells exhibitelectrophysiological characteristics similar to those observedin mature neurons. Induction of a dopaminergic phenotype is
Fig. 6. Neurite growth and arborization of MIAMI-derived neuronal cells are dramatneurite processes and branching points in the absence of NT-3 (A). Upon addition ofthe length of the neurites (C–E). In most cases, numerous long neurites could be obsethicker process (arrows in panel E).
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
dramatically enhanced by exposure to SHH, FGF-8, and RA,known inducers of this phenotype, during the commitment step.
NSCs divide symmetrically to enrich the NSC pool of thesubventricular zone (self-renewing) or asymmetrically togenerate differentiating progeny that lose their attachment tothe basal lamina and rapidly migrate to their specific location[34]. The neural committed neuron-restricted precursors expressessentially nestin, and once committed towards the neuronallineage express neuron-specific class III β-tubulin. Afterneuronal differentiation and migration to their final location,they mainly express neurofilament and NeuN, develop electricalactivity and synthesize and respond to neurotransmitters. NSCs
ically increased by NT-3 treatment. Cells with a bipolar shape poorly developedNT-3 during steps 2 and 3, the arborization dramatically increased (B) as well asrved (arrowheads in panel E) in neuronal cells also found to contain a longer and
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone

Fig. 7. Modulation of gene expression during neuronal differentiation of MIAMIcells. Gene expression during neuronal differentiation of MIAMI cells assessedby semi-quantitative RT-PCR as described. SSEA-4 synthase is expressed by theMIAMI cells and then when the cells begin to differentiate it disappears.Patched is also expressed before treatment and at step 1 and is still present afterstep 2 when RA, SHH, and FGF-8 are added. Gli as well as Nurr1 mRNAappear at step 2, and Nurr1 is still present and is more strongly expressed afterthe long term differentiation. At this step, TH mRNA is also expressed.Expression of NTRK-3 is observed during all the steps of the protocol. SSEA-4:stage-specific embryonic antigen 4, NTRK-3: neurotrophic tyrosine kinasereceptor 3, TH: tyrosine hydroxylase.
11V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
can be isolated based on their dependence on bFGF for growth[35], and bFGF appears to be a prerequisite for neuronaldifferentiation [36,37]. Similarly, neural specification (step 1) ofcompetent MIAMI cells requires bFGF which leads to theupregulation of nestin expression. MIAMI cells express NTRK-3, and it has been reported that treatment of NSCs with NT-3promotes neuronal maturation [38]. Progression of inducedMIAMI cells toward a neuronal phenotype required subsequentexposure to NT-3 and low levels of anti-oxidants (step 2). Thisneuronal commitment step induced the expression of neuron-specific class III β-tubulin and TrkAwith the paralleled decreasein nestin expression, a phenotype consistent with that of aneuronal precursor cell [33]. Final neuronal differentiation (step3) also required treatment with the neurotrophic factor NT-3;however, addition of NGF and BDNF enhanced the NT-3 effect.While these factors are known to support differentiation andsurvival of neurons [39,40], this is the first report showing thatNT-3 is required for the neuronal differentiation of human adultnon-neural stem cells. Neuronal differentiation is demonstratedby the expression of NeuN, neurofilament, synaptophysin,neuronal morphology, extensive neurite growth and arboriza-tion, and acquisition of electrophysiological properties indis-tinguishable from bona fide neurons.
Of tantamount importance is the fact that MIAMI-derivedneuron-like cells from young and old donors developed neuron-like electrophysiological properties without the need for co-culture with astrocytes as published for rat bone marrow-
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
derived cells [10] and other brain-derived stem cells [41]. Whilethis demonstrates that differentiation of human MIAMI cells toneuron-like cells can be achieved without any type of co-culture, it cannot be excluded that co-culturing with primaryastrocytes may further enhance the efficiency of neuronaldifferentiation of MIAMI cells.
The unique ability of normal early passage human MIAMIcells to differentiate to neuron-like cells, under defined cultureconditions using characterized growth factor and without theneed of co-culturing with other cell types, is of criticalimportance in the context of cellular transplantation. Our resultsdemonstrate that marrow stem cells can differentiate to aneuron-like phenotype without the need for fusion withprecursor cells of the neuroectodermal lineage. While it cannotbe excluded that NSCs may be present among the MIAMI cellpopulation, this possibility is unlikely since the response ofMIAMI cells to neuronal induction was rapid and homoge-neous, and it did not require a long period of time that mightinvolve selective proliferation and differentiation of a smallpopulation of NSCs.
For the same reasons, it is unlikely that neuronal differentia-tion of MIAMI cells is a cell culture artifact produced after long-term expansion or a biochemical manipulation of a subpopula-tion of cells [42,43]. Although there is the possibility forheterogeneity of the initial cell population, starting MIAMI cellsare morphologically homogeneous and consistently express thesame markers including the expression of certain neural markerssuch as NT-3 receptor NTRK-3.
MIAMI cells are capable of differentiating into cells thatexpress neuronal proteins, maintain a neuron-like morphology,and respond to neurotrophic factors. However, acquisition ofelectrophysiological characteristics indicates that MIAMI-derived neuronal cells exhibit unique neuronal-like properties.The cells maintained a resting membrane potential anddepolarizing voltage steps activated ion currents. In cells withthe potassium currents blocked, the inward current displayedthe features of fast neuronal sodium currents, however, furtherdetailed characterization of these currents is needed. Further-more, the expression of the synaptic vesicular membraneglycoprotein, synaptophysin, after the neuronal inductionprotocol suggests the capacity for synaptic activity.
Acquisition of electrophysiological properties in cellsderived from MIAMI cells was dependent on NT-3 treatment.MIAMI cells are derived from MSCs after selection/expansionrequiring a low oxygen tension environment. Since the moreprimitive marrow stem cells are thought to be maintained at lowoxygen tension in vivo [44,45], low oxygen tension mayrepresent a key factor in the selection and expansion of earlyprogenitor cells capable of maintaining their potential to developinto cells of the neuroectodermal lineage. It has been shown thatdecreased oxygen levels promoted the survival and proliferationof a population of mammalian neural stem cells [46].Furthermore, reduced oxygen levels promote the survival,proliferation, and catecholaminergic differentiation of CNSstem cells [47] and stimulate sympathoadrenal differentiation byisolated neural crest stem cells [48], suggesting that neural stemcells may exhibit a conserved response to reduced oxygen levels.
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone

12 V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
Although, MIAMI cells were differentiated into neuron-likecells at 20% oxygen (air), these results open the possibility that alower oxygen tension may further enhance their neuronaldifferentiation.
The addition of NT-3 was required in order to obtain maturefunctional neuron-like cells in our studies even when two otherneurotrophic factors, BDNF and NGF, were present. Thissuggests that specific signals mediated by TrkC/NTRK-3 arerequired to induce the development of a more complexarborization pattern and the acquisition of the electrophysiolo-gical properties described. These results bring forward the novelconcept that NT-3 acting most likely through NTRK-3 candirect the neuron-like differentiation of non-neural earlypassage human stem cells. It has recently been reported thatanti-NT-3 neutralizing antibodies decreased the survival anddifferentiation of cortical progenitors [39].
From a clinical perspective, it is important to show thatMIAMI cells from young and old individuals can serve as astarting material to generate neuron-like cells because this canbe accomplished in vitro under defined conditions and stronglysuggests that autologous cells could be used in a transplantationsetting in young and older persons. In this regard and for celltherapy applications, it is also important to be able to direct theirdifferentiation to neuron-like cells expressing a specificneurotransmitter. The proliferating neuronal precursors at step2 expressed Ptc and Smo and also slightly FGFR3, the receptorsof SHH and FGF-8, which are required for inducingdopaminergic neurons [49,50]. By adding SHH, FGF-8, andRA at this step, we have activated the SHH pathway as shownby an increase of gli and ptch expression and finally obtainedapproximately 60% of TH expressing MIAMI-derived matureneurons. Without these factors, only a very slight TH expressionis observed. These neuron-like cells also express NF-M andNTRK-1, 2, and 3, probably due in part to RA treatment, whichincreased the expression of these receptors rendering the cellsmore responsive to the neurotrophins subsequently added. It iswell known that BDNF and NT-3 may increase the survival ofdopaminergic neurons [51,52]. The treatment activated thedopaminergic induction pathway as a strong expression ofNurr1, a key element in this pathway, was also observed 24 hafter addition of these molecules [53]. TH expression followedat step 3 confirming their dopaminergic phenotype.
As suggested by the study of Jiang [10], astrocytes seem toenhance MSC differentiation into mature dopaminergic cells.Moreover, it was described that MSC are able to differentiateinto astrocytes [8,10,15]. In this way, we have performedimmunostaining of the astrocyte marker GFAP and quantifiedthe release of CNTF, a cytokine that may be secreted byastrocytes and is involved in their differentiation [54,55]. Onlyfew GFAP positives cells were observed and no CNTF wasdetected. These data suggest that the induction protocol usingSHH, FGF-8, and RA by itself is sufficient to obtaindopaminergic cells. Nevertheless, the presence of astrocytesobtained from MIAMI cells would be useful to increase thepercentage of dopaminergic cells, without the disadvantage ofco-culture conditions and may allow a better integration in thebrain after implantation. Interestingly, the secretion of TGF-β,
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
involved in dopaminergic differentiation, was regulated duringthe neuronal induction protocol. No TGF-βwas detected duringneuronal commitment, but it was secreted a few days later,during step 3 of the induction protocol. It was shown that TGF-β's act as differentiation inducers of rat fetal ventralmesencephalon cells into TH positive cells as well as survivalfactors [56]. We postulate that an autocrine stimulation of theMIAMI cells on their own differentiation into dopaminergiccells may occur. Nevertheless, it would be interesting to analyzehow an addition of TGF-β can promote MIAMI cellsdifferentiation into TH positive cells during the 3 steps inducingprotocol, especially in combination with SHH.
In conclusion, we report here for the first time that normalnon-infected human BM-derived MIAMI cells, from young andold, can be induced to differentiate to cells with morphological,biochemical, and more importantly with electrophysiologicalcharacteristics similar to those observed in mature neurons, inresponse to defined cytokines, dependent on NT-3 (likelymediated by the activation of NTRK-3) and without the need ofco-culturing with any other cell types. Furthermore, thedevelopment of dopaminergic neuronal cells can be dramaticallyenhanced by the action of known inducers of this phenotype.The sequential neuronal differentiation program of these cellsclosely mimics that of NSCs at the molecular level. Therefore,MIAMI cells may represent an autologous source of cells for therepair of damaged, diseased, or aged neuronal tissue.
Acknowledgments
This work was in part supported by the Department ofVeterans Affairs and the ‘Fondation de la Recherché Medicale.’The authors are grateful to Drs. Guy Howard and Carlos Perez-Stable for rewarding discussions and critical review of themanuscript; Drs. Camillo Ricordi and Antonello Pileggi andMs. Julie Allickson from the Diabetes Research Institute(University of Miami School of Medicine) for providing thewhole bone marrow; David Vazquez, Irvin Vidaurre and BlancaRodriguez for expert technical assistance in cell culture and cellassays; and Virginia Roos for help in preparing the manuscript.
References
[1] Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, et al. Pluripotency of mesenchymal stem cells derived fromadult marrow. Nature 2002;418:41–9.
[2] Colter DC, Sekiya I, Prockop DJ. Identification of a subpopulation ofrapidly self-renewing and multipotential adult stem cells in colonies ofhuman marrow stromal cells. Proc Natl Acad Sci U S A 2001;98:7841–5.
[3] D'Ippolito G, Schiller PC, Ricordi C, Roos BA, Howard GA. Age-relatedosteogenic potential of mesenchymal stromal stem cells from humanvertebral bone marrow. J Bone Miner Res 1999;14:1115–22.
[4] Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietictissues. Science 1997;276:71–4.
[5] Caplan AI. Mesenchymal stem cells. J Orthop Res 1991;9:641–50.[6] Reyes M, Verfaillie CM. Characterization of multipotent adult progenitor
cells, a subpopulation of mesenchymal stem cells. Ann N Y Acad Sci2001;938:231–3 (discussion 233–5).
[7] D'Ippolito G, Diabira S, Howard GA, Menei P, Roos BA, Schiller PC.Marrow-isolated adult multilineage inducible (MIAMI) cells, a unique
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone

13V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
population of postnatal young and old human cells with extensiveexpansion and differentiation potential. J Cell Sci 2004;117:2971–81.
[8] Sanchez-Ramos J, Song S, Cardozo-Pelaez F, Hazzi C, Stedeford T,Willing A, et al. Adult bone marrow stromal cells differentiate into neuralcells in vitro. Exp Neurol 2000;164:247–56.
[9] Woodbury D, Schwarz EJ, Prockop DJ, Black IB. Adult rat and humanbone marrow stromal cells differentiate into neurons. J Neurosci Res2000;61:364–70.
[10] Jiang Y, Henderson D, Blackstad M, Chen A, Miller RF, Verfaillie CM.Neuroectodermal differentiation from mouse multipotent adult progenitorcells. Proc Natl Acad Sci U S A 2003;100(Suppl 1):11854–60.
[11] Azizi SA, Stokes D, Augelli BJ, DiGirolamo C, Prockop DJ. Engraftmentand migration of human bone marrow stromal cells implanted in the brainsof albino rats—similarities to astrocyte grafts. Proc Natl Acad Sci U S A1998;95:3908–13.
[12] Brazelton TR, Rossi FM, Keshet GI, Blau HM. From marrow to brain:expression of neuronal phenotypes in adult mice. Science 2000;290:1775–9.
[13] Mezey E, Chandross KJ, Harta G, Maki RA, McKercher SR. Turningblood into brain: cells bearing neuronal antigens generated in vivo frombone marrow. Science 2000;290:1779–82.
[14] Prockop DJ. Marrow stromal cells as stem cells for continual renewal ofnonhematopoietic tissues and as potential vectors for gene therapy. J CellBiochem 1998;30–31:284–5 (Suppl.).
[15] Zhao LR, Duan WM, Reyes M, Keene CD, Verfaillie CM, Low WC.Human bone marrow stem cells exhibit neural phenotypes and ameliorateneurological deficits after grafting into the ischemic brain of rats. ExpNeurol 2002;174:11–20.
[16] ChoppM, Zhang XH, Li Y, Wang L, Chen J, Lu D, et al. Spinal cord injuryin rat: treatment with bone marrow stromal cell transplantation.NeuroReport 2000;11:3001–5.
[17] Schwarz EJ, Alexander GM, Prockop DJ, Azizi SA. Multipotentialmarrow stromal cells transduced to produce L-DOPA: engraftment in a ratmodel of Parkinson disease. Hum Gene Ther 1999;10:2539–49.
[18] Dezawa M, Kanno H, Hoshino M, Cho H, Matsumoto N, Itokazu Y, et al.Specific induction of neuronal cells from bone marrow stromal cells andapplication for autologous transplantation. J Clin Invest 2004;113:1701–10.
[19] Tao H, Rao R, Ma DD. Cytokine-induced stable neuronal differentiation ofhuman bone marrow mesenchymal stem cells in a serum/feeder cell-freecondition. Dev Growth Differ 2005;47:423–33.
[20] Freed CR, Breeze RE, Rosenberg NL, Schneck SA, Kriek E, Qi JX, et al.Survival of implanted fetal dopamine cells and neurologic improvement 12to 46 months after transplantation for Parkinson's disease. N Engl J Med1992;327:1549–55.
[21] Schierle GS, Hansson O, Leist M, Nicotera P, Widner H, Brundin P.Caspase inhibition reduces apoptosis and increases survival of nigraltransplants. Nat Med 1999;5:97–100.
[22] Hung SC, Cheng H, Pan CY, Tsai MJ, Kao LS, Ma HL. In vitrodifferentiation of size-sieved stem cells into electrically active neural cells.Stem Cells 2002;20:522–9.
[23] Cho KJ, Trzaska KA, Greco SJ, McArdle J, Wang FS, Ye JH, et al.Neurons derived from human mesenchymal stem cells show synaptictransmission and can be induced to produce the neurotransmitter substanceP by interleukin-1 alpha. Stem Cells 2005;23:383–91.
[24] Lendahl U, Zimmerman LB, McKay RD. CNS stem cells express a newclass of intermediate filament protein. Cell 1990;60:585–95.
[25] Cattaneo E, McKay R. Proliferation and differentiation of neuronal stemcells regulated by nerve growth factor. Nature 1990;347:762–5.
[26] Quinn SM, Walters WM, Vescovi AL, Whittemore SR. Lineage restrictionof neuroepithelial precursor cells from fetal human spinal cord. J NeurosciRes 1999;57:590–602.
[27] Brewer GJ, Torricelli JR, Evege EK, Price PJ. Optimized survival ofhippocampal neurons in B27-supplemented Neurobasal, a new serum-freemedium combination. J Neurosci Res 1993;35:567–76.
[28] Valeyev AY, Hackman JC, Holohean AM, Wood PM, Katz JL, DavidoffRA. GABA-induced Cl− current in cultured embryonic human dorsal rootganglion neurons. J Neurophysiol 1999;82:1–9.
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
[29] McCarthy M, Auger D, Whittemore SR. Human cytomegalovirus causesproductive infection and neuronal injury in differentiating fetal humancentral nervous system neuroepithelial precursor cells. J Hum Virol2000;3:215–28.
[30] Taupin P, Gage FH. Adult neurogenesis and neural stem cells of the centralnervous system in mammals. J Neurosci Res 2002;69:745–9.
[31] McKay R. Stem cells in the central nervous system. Science 1997;276:66–71.
[32] Zimmerman L, Parr B, Lendahl U, Cunningham M, McKay R, Gavin B, etal. Independent regulatory elements in the nestin gene direct transgeneexpression to neural stem cells or muscle precursors. Neuron1994;12:11–24.
[33] Messam CA, Hou J, Major EO. Coexpression of nestin in neural and glialcells in the developing human CNS defined by a human-specific anti-nestin antibody. Exp Neurol 2000;161:585–96.
[34] Cai L, Hayes NL, Nowakowski RS. Synchrony of clonal cell proliferationand contiguity of clonally related cells: production of mosaicism in theventricular zone of developing mouse neocortex. J Neurosci 1997;17:2088–100.
[35] Shihabuddin LS, Ray J, Gage FH. FGF-2 is sufficient to isolate progenitorsfound in the adult mammalian spinal cord. Exp Neurol 1997;148:577–86.
[36] Kennea NL, Mehmet H. Neural stem cells. J Pathol 2002;197:536–50.[37] Craig CG, Tropepe V, Morshead CM, Reynolds BA, Weiss S, van der
Kooy D. In vivo growth factor expansion of endogenous subependymalneural precursor cell populations in the adult mouse brain. J Neurosci1996;16:2649–58.
[38] Takahashi J, Palmer TD, Gage FH. Retinoic acid and neurotrophinscollaborate to regulate neurogenesis in adult-derived neural stem cellcultures. J Neurobiol 1999;38:65–81.
[39] Barnabe-Heider F, Miller FD. Endogenously produced neurotrophinsregulate survival and differentiation of cortical progenitors via distinctsignaling pathways. J Neurosci 2003;23:5149–60.
[40] Park JK, Williams BP, Alberta JA, Stiles CD. Bipotent cortical progenitorcells process conflicting cues for neurons and glia in a hierarchical manner.J Neurosci 1999;19:10383–9.
[41] Song H, Stevens CF, Gage FH. Astroglia induce neurogenesis from adultneural stem cells. Nature 2002;417:39–44.
[42] Meletis K, Frisen J. Blood on the tracks: a simple twist of fate? TrendsNeurosci 2003;26:292–6.
[43] Bertani N, Malatesta P, Volpi G, Sonego P, Perris R. Neurogenic potentialof human mesenchymal stem cells revisited: analysis by immunostaining,time-lapse video and microarray. J Cell Sci 2005;118:3925–36.
[44] Chow DC, Wenning LA, Miller WM, Papoutsakis ET. Modeling pO(2)distributions in the bone marrow hematopoietic compartment. II. ModifiedKroghian models. Biophys J 2001;81:685–96.
[45] Danet GH, Pan Y, Luongo JL, Bonnet DA, Simon MC. Expansion ofhuman SCID-repopulating cells under hypoxic conditions. J Clin Invest2003;112:126–35.
[46] Morrison SJ, White PM, Zock C, Anderson DJ. Prospective identification,isolation by flow cytometry, and in vivo self-renewal of multipotentmammalian neural crest stem cells. Cell 1999;96:737–49.
[47] Studer L, Csete M, Lee SH, Kabbani N, Walikonis J, Wold B, et al.Enhanced proliferation, survival, and dopaminergic differentiation of CNSprecursors in lowered oxygen. J Neurosci 2000;20:7377–83.
[48] Morrison SJ, Csete M, Groves AK, Melega W, Wold B, Anderson DJ.Culture in reduced levels of oxygen promotes clonogenic sympathoadrenaldifferentiation by isolated neural crest stem cells. J Neurosci 2000;20:7370–6.
[49] Ye W, Shimamura K, Rubenstein JL, Hynes MA, Rosenthal A. FGF andShh signals control dopaminergic and serotonergic cell fate in the anteriorneural plate. Cell 1998;93:755–66.
[50] Simon HH, Bhatt L, Gherbassi D, Sgado P, Alberi L. Midbraindopaminergic neurons: determination of their developmental fate bytranscription factors. Ann N YAcad Sci 2003;991:36–47.
[51] Espejo M, Cutillas B, Arenas TE, Ambrosio S. Increased survival ofdopaminergic neurons in striatal grafts of fetal ventral mesencephalic cellsexposed to neurotrophin-3 or glial cell line-derived neurotrophic factor.Cell Transplant 2000;9:45–53.
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone

14 V.M. Tatard et al. / Bone xx (2006) xxx–xxx
ARTICLE IN PRESS
[52] Baquet ZC, Bickford PC, Jones KR. Brain-derived neurotrophic factor isrequired for the establishment of the proper number of dopaminergicneurons in the substantia nigra pars compacta. J Neurosci 2005;25:6251–9.
[53] Arenas E. Engineering a dopaminergic phenotype in stem/precursor cells:role of Nurr1, glia-derived signals, and Wnts. Ann N Y Acad Sci2005;1049:51–66.
[54] Kamiguchi H, Yoshida K, Sagoh M, Sasaki H, Inaba M, Wakamoto H, et
Please cite this article as: Valerie M. Tatard et al., Neurotrophin-directed differentia(2006), doi:10.1016/j.bone.2006.09.013
al. Release of ciliary neurotrophic factor from cultured astrocytes and itsmodulation by cytokines. Neurochem Res 1995;20:1187–93.
[55] Johe KK, Hazel TG, Muller T, Dugich-Djordjevic MM, McKay RD.Single factors direct the differentiation of stem cells from the fetal andadult central nervous system. Genes Dev 1996;10:3129–40.
[56] Roussa E, Farkas LM, Krieglstein K. TGF-beta promotes survival onmesencephalic dopaminergic neurons in cooperation with Shh and FGF-8.Neurobiol Dis 2004;16:300–10.
tion of human adult marrow stromal cells to dopaminergic-like neurons, Bone




















