
Striatal Sensitivity During Reward Processing in Attention-Deficit/Hyperactivity Disorder

94:4168-4177, 2005. First published Sep 7, 2005; doi:10.1152/jn.00698.2005 JN
G. Gonzalez-Burgos, S. Kroener, J. K. Seamans, D. A. Lewis and G. BarrionuevoDorsolateral Prefrontal Cortex Plasticity in Fast-Spiking Interneurons of Primate Dopaminergic Modulation of Short-Term Synaptic
You might find this additional information useful...
59 articles, 37 of which you can access free at: This article cites http://jn.physiology.org/cgi/content/full/94/6/4168#BIBL
including high-resolution figures, can be found at: Updated information and services http://jn.physiology.org/cgi/content/full/94/6/4168
can be found at: Journal of Neurophysiologyabout Additional material and information http://www.the-aps.org/publications/jn
This information is current as of November 17, 2005 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the
publishes original articles on the function of the nervous system. It is published 12 times a yearJournal of Neurophysiology
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

Dopaminergic Modulation of Short-Term Synaptic Plasticity in Fast-SpikingInterneurons of Primate Dorsolateral Prefrontal Cortex
G. Gonzalez-Burgos,1 S. Kroener,2 J. K. Seamans,3 D. A. Lewis,1,2 and G. Barrionuevo2
1Departments of Psychiatry and 2Neuroscience, University of Pittsburgh, Pittsburgh, Pennsylvania; and3Department of Physiology and Neuroscience, Medical University of South Carolina, Charleston, South Carolina
Submitted 1 July 2005; accepted in final form 1 September 2005
Gonzalez-Burgos, G., S. Kroener, J. K. Seamans, D. A. Lewis, andG. Barrionuevo. Dopaminergic modulation of short-term synapticplasticity in fast-spiking interneurons of primate dorsolateral prefron-tal cortex. J Neurophysiol 94: 4168–4177, 2005. First publishedSeptember 7, 2005; doi:10.1152/jn.00698.2005. Dopaminergic regu-lation of primate dorsolateral prefrontal cortex (PFC) activity isessential for cognitive functions such as working memory. However,the cellular mechanisms of dopamine neuromodulation in PFC are notwell understood. We have studied the effects of dopamine receptoractivation during persistent stimulation of excitatory inputs ontofast-spiking GABAergic interneurons in monkey PFC. Stimulation at20 Hz induced short-term excitatory postsynaptic potential (EPSP)depression. The D1 receptor agonist SKF81297 (5 �M) significantlyreduced the amplitude of the first EPSP but not of subsequentresponses in EPSP trains, which still displayed significant depression.Dopamine (DA; 10 �M) effects were similar to those of SKF81297and were abolished by the D1 antagonist SCH23390 (5 �M), indi-cating a D1 receptor-mediated effect. DA did not alter miniatureexcitatory postsynaptic currents, suggesting that its effects were ac-tivity dependent and presynaptic action potential dependent. In con-trast to previous findings in pyramidal neurons, in fast-spiking cells,contribution of N-methyl-D-aspartate receptors to EPSPs at subthresh-old potentials was not significant and fast-spiking cell depolarizationdecreased EPSP duration. In addition, DA had no significant effectson temporal summation. The selective decrease in the amplitude of thefirst EPSP in trains delivered every 10 s suggests that in fast-spikingneurons, DA reduces the amplitude of EPSPs evoked at low frequencybut not of EPSPs evoked by repetitive stimulation. DA may thereforeimprove detection of EPSP bursts above background synaptic activity.EPSP bursts displaying short-term depression may transmit spike-timing-dependent temporal codes contained in presynaptic spiketrains. Thus DA neuromodulation may increase the signal-to-noiseratio at fast-spiking cell inputs.
I N T R O D U C T I O N
Dopaminergic regulation of prefrontal cortex (PFC) activityis essential for cognitive functions such as working memory(Goldman-Rakic et al. 2004). Dopamine (DA) neurons inner-vate the primate PFC (Lewis et al. 1987; Williams and Gold-man-Rakic 1998) and release DA during working-memorytasks (Watanabe et al. 1997). Activation of D1-like receptors isnecessary for persistent firing in PFC (Sawaguchi 2001; Wil-liams and Goldman-Rakic 1995), which is thought to be thecellular basis of information storage in working memory (Fus-ter 1997). Although DA appears to be essential for workingmemory, the cellular mechanisms of DA action in PFC are not
well understood. In vitro, DA enhances the response of mon-key PFC pyramidal neurons to injection of excitatory current.However, DA does not elicit pyramidal neuron firing in theabsence of excitatory stimuli, and firing induced by depolariz-ing current pulses is not sustained after the stimulus ceases, inthe presence or absence of DA (Gonzalez-Burgos et al. 2002;Henze et al. 2000). Thus DA modulation of pyramidal cellexcitability is not sufficient to produce persistent activity.Indeed computational models have suggested that the effects ofDA on persistent activity may involve modulation of synapticstrength (Brunel and Wang 2001; Durstewitz et al. 2000;Fellous and Sejnowski 2003; Wang 2001).
In PFC neurons, DA modulates the amplitude of excitatorypostsynaptic potentials (EPSPs) evoked by single stimuli (Gaoand Goldman-Rakic 2003; Gao et al. 2001; Gonzalez-Islas andHablitz 2003; Law-Tho et al. 1994; Seamans et al. 2001; Urban etal. 2002). However, persistent stimulation causes short-term syn-aptic plasticity, and thus EPSPs evoked by temporally isolatedstimuli provide limited information on synaptic interactions dur-ing sustained activity. In PFC pyramidal neurons, stimulus trainselicit short-term EPSP depression (Gonzalez-Burgos et al. 2004;Hempel et al. 2000; Seamans et al. 2001). Synapses with short-term depression are not optimized to support persistent postsyn-aptic firing during persistent synaptic input because their efficacydecreases markedly shortly after the onset of a presynaptic spiketrain (Pouille and Scanziani 2004). However, recent data suggestthat in PFC pyramidal neurons DA modulation counteracts EPSPdepression by decreasing the initial EPSPs and potentiating thelate postsynaptic responses in EPSP trains elicited by sustainedpresynaptic stimulation (Seamans et al. 2001). This effect of DAmay contribute to the recurrent excitation thought to underliesustained firing in local circuits of PFC (Durstewitz et al. 2000).Because recurrent excitation requires inhibitory control, the ef-fects of DA in PFC circuits may involve GABAergic neurons, inaddition to pyramidal cells (Brunel and Wang 2001; Durstewitz etal. 2000).
Inhibitory GABA neurons in monkey PFC include two elec-trophysiological subgroups: fast-spiking (FS) and non-FS cells(Gonzalez-Burgos et al. 2004, 2005). The electrophysiologicalproperties of FS neurons in monkey PFC (Gonzalez-Burgos et al.2004, 2005; Zaitsev et al. 2005) are similar to those described forFS cells in rat neocortex. In the frontal cortex of either rats ormacaque monkeys, FS interneurons selectively express the cal-cium-binding protein parvalbumin (Kawaguchi and Kubota 1997;Zaitsev et al. 2005). Compared with other interneuron classes,
Address for reprint requests and other correspondence: G. Gonzalez-Burgos,Translational Neuroscience Program, Dept. of Psychiatry, University of Pitts-burgh School of Medicine, Rm. W1651 Biomedical Science Tower, 3811O’Hara St. Pittsburgh, PA 15213 (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
J Neurophysiol 94: 4168–4177, 2005.First published September 7, 2005; doi:10.1152/jn.00698.2005.
4168 0022-3077/05 $8.00 Copyright © 2005 The American Physiological Society www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

parvalbumin-containing FS cells express a higher density of DAreceptors (Le Moine and Gaspar 1998; Muly et al. 1998; Paspalasand Goldman-Rakic 2005), are preferentially innervated by DAfibers (Sesack et al. 1995, 1998), and selectively respond to DAreceptor activation (Gorelova et al. 2002; Porter et al. 1999). ThusFS cells may be the main interneuron target of DA in the PFC.
In monkey PFC, excitatory inputs onto FS cells and inhibitoryconnections made by FS neurons onto nearby pyramidal cellsdisplay short-term depression (Gonzalez-Burgos et al. 2004,2005). Short-term depression decreases the efficiency of synaptictransmission during sustained activation and thus to encode thepresynaptic firing rate per se. However, synaptic depression en-hances the ability to encode changes in presynaptic firing rate(Abbott et al. 1997; Tsodyks and Markram 1997). Thus thefunctional properties of their synaptic inputs and outputs suggestthat FS cells in monkey PFC may constitute an inhibitory neuronsystem specialized to transiently signal changes in firing fre-quency (Beierlein et al. 2003; Pouille and Scanziani 2004). How-ever, DA may counteract short-term depression of excitatoryinputs onto FS neurons as it does in pyramidal cells (Seamans etal. 2001). If so, DA would enable FS neurons to produce sustainedinhibition during persistent network activity. Here, we determinedthe effects of DA on short-term depression of excitatory synapticinputs onto FS interneurons in monkey PFC.
M E T H O D S
Slice preparation and electrophysiological recording
All procedures were carried out in accordance with the NationalInstitutes of Health Guide for the Care and Use of Laboratory Animalsand were approved by the University of Pittsburgh Institutional AnimalCare and Use Committee. Slices were prepared from tissue blockscontaining portions of the dorsolateral PFC (areas 9 and 46) of youngadult macaque monkeys as described in detail elsewhere (Gonzalez-Burgos et al. 2004). Experiments were conducted in 43 slices obtainedfrom 11 animals. Slices obtained from the dorsolateral PFC of these sameanimals were used to collect data for two other studies (Gonzalez-Burgoset al. 2004, 2005). For recording, slices were placed in a submersionchamber and superfused at 32–33°C with an oxygenated solution of thefollowing composition (in mM): 126 NaCl, 2.5 KCl, 1.2 Na2HPO4, 25NaHCO3, 2.0 CaCl2, 1.0 MgCl2, and 10 glucose. Voltage recordingswere obtained with patch pipettes from neurons identified visually inlayers 2/3 of either the medial or lateral bank of the principal sulcus (area46), using infrared differential interference contrast video microscopy.Recordings were obtained using Axoclamp 2B (Axon Instruments, UnionCity, CA) or BVC700A (Dagan, Minneapolis, MN) amplifiers operatingin current-clamp mode, using bridge balance and pipette capacitancecompensation. Pipettes had a resistance of 4–6 M� when filled with asolution of the following composition (in mM): 120 KMethylsulphate, 10HEPES, 0.2 EGTA, 4.5 ATP, 0.3 GTP, 14 phosphocreatine, and 0.5%biocytin, pH adjusted to 7.2–7.3. Focal stimulation was applied usingtheta glass pipettes pulled to a tip diameter or 3–5 �m and filled withextracellular solution. A chlorided silver wire was inserted into eachcompartment of the theta glass stimulation pipette and connected to astimulus isolation unit. External trigger pulses were generated by acomputer-controlled data acquisition board and were used to commandthe stimulus isolation unit. Focal stimulation was used to elicit smallEPSPs (mean amplitude smaller than 5 mV) in the presence of 10 �Mbicuculline (Gonzalez-Burgos et al. 2004). The duration of the stimuluspulses was 100 �s, and the stimulation current intensity varied from �20to �150 �A. Stimulation parameters were adjusted in each experiment toelicit single EPSPs with characteristics of monosynaptic responses, i.e., ashort latency (�3 ms) with small trial-to-trial variability and absence ofpolysynaptic components overlapping with the EPSP decay phase
(Gonzalez-Burgos et al. 2000). Once a satisfactory response was ob-tained, the stimulation parameters were kept constant throughout theexperiment. EPSPs were elicited first at a frequency of 0.1 Hz; next,stimulus trains of 20-Hz frequency were delivered every 10 s, using thesame stimulus parameters. To determine if in addition to short-termdepression, stimulus trains had long-term effects on EPSP amplitude, in12 experiments EPSPs were initially elicited by single stimulus shocksdelivered every 10 s (0.1 Hz) during a �5-min baseline recording period.After this period, 20-Hz stimulus trains were delivered every 10 s for�10 min.
In current-clamp recordings, injection of steady-state current wasemployed to compensate for any significant changes in the FS cellmembrane potential induced by application of DA or D1 agonists,relative to control resting membrane potential. In voltage-clamp experi-ments, the membrane potential was held at �70 mV, and excitatorypostsynaptic currents (EPSCs) were pharmacologically isolated by add-ing 20 �M bicuculline to the bath. Spontaneous EPSCs (sEPSCs) andminiature EPSCs (mEPSCs) were recorded under the same conditionsexcept that mEPSCs were recorded in the presence of 1 �M of thesodium channel blocker tetrodotoxin (Sigma, St. Louis, MO).
Fresh stock solutions of all dopaminergic pharmacological com-pounds were prepared on the day of each experiment and wereprotected from light exposure until bath application. DA stock solu-tions were prepared by dissolving the compound shortly beforeapplication to each slice. The stocks and bath solutions contained theanti-oxidant sodium metabisulfite at a final concentration of 75 �M.All pharmacological compounds used in this study were obtainedfrom Research Biochemicals, unless otherwise indicated.
Data analysis
To determine if stimulus trains had long-term effects on EPSPamplitude, we compared the amplitude of single EPSPs with theamplitude of the first EPSP (EPSP1) in 20-Hz trains elicited in thesame cells. Single EPSPs were elicited by stimulus shocks deliveredevery 10 s (0.1 Hz). EPSP trains were elicited by trains of 10 stimuluspulses at 20 Hz (unless otherwise indicated) and delivered at aninter-train frequency of 0.1 Hz (Fig. 1F). The amplitude of singleEPSPs elicited at 0.1 Hz was estimated by averaging the EPSPsrecorded during a 5-min baseline period of stimulation with singleshocks. The amplitude of EPSP1 in trains was estimated by averagingthe first EPSPs of trains recorded in the same neurons, during a10-min period of stimulation with 20-Hz trains.
Analysis of the effect of DA agonists on EPSP trains was based onmonitoring the amplitude of the first EPSP in the trains before, duringa 5-min drug application, and after washout. In most recorded neurons(SKF81297, 9 of 12 cells; DA 5 of 7 cells), shortly after applicationof DA agonists, a significant reduction of the first EPSP amplitudewas observed in the FS neuron (Fig. 2). In the remaining cases (n �5 FS neurons), in contrast, DA agonists produced no detectabledecrease or apparently an increase in EPSP amplitude compared withthe baseline EPSP amplitude before drug application. The effect ofDA receptor activation was somewhat weaker in the entire population(EPSP1 reduced to 87 � 7% of control) than in the 14 cells showinga decrease of EPSP1 amplitude (EPSP1 reduced to 70 � 5% ofcontrol). Nevertheless, DA receptor activation had a statisticallysignificant effect on the first EPSP in trains (P � 0.05) in the entirepopulation (n � 19) of FS cells in which EPSP trains were tested withDA agonists. Previous studies suggest the presence of heterogeneoussubpopulations of FS neurons in PFC, with regard to the effects of DAon excitatory synaptic inputs (Gao and Goldman-Rakic 2003; C .D.Paspalas, personal communication) (see DISCUSSION). In light of theseprevious studies suggesting heterogeneity, the results presented hereare based on the analysis of short-term plasticity in the group of FSneurons in which D1 receptor activation produced a decrease in theamplitude of the first EPSP in the trains, the decrease being �10% ofthe baseline EPSP amplitude during control period.
4169DOPAMINE AND SHORT-TERM PLASTICITY IN FAST-SPIKING CELLS
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from
The effect of DA receptor activation on the EPSP trains wasdetermined on traces obtained by averaging the last 10–20 consecu-tive EPSP trains recorded in the presence of the drugs and before theonset of washout. These average traces were compared with controlaverage traces obtained just prior to drug application. In eitherexperimental and control average traces, EPSP amplitude was nor-malized relative to that of the first EPSP in the trains recorded incontrol conditions. The effects of DA receptor activation were typi-cally reversed by drug washout (Fig. 2B). However, in some slices theeffects were long-lasting as described previously for certain D1receptor-mediated effects (Seamans and Yang 2004). Potential differ-ences between FS neuron EPSP trains exhibiting short- versus long-lasting D1 receptor effects were not investigated further and resultswere pooled. To determine if DA receptor activation changed the rateof synaptic depression after the first EPSP in the trains, EPSPs werealso normalized to the second control EPSP (Fig. 2D).
Analysis of m- and sEPSC data (event frequency, amplitude, andkinetics) was performed using MiniAnalysis (Synaptosoft, Decatur,GA). Events were detected when they crossed a threshold set at threetimes the SD of the baseline noise. The detected events were con-firmed as synaptic events by visual inspection. For s- and mEPSCs,the total number of events from 8 min of continuous recordings eachbefore and �2 min after DA application was compared.
Results are shown as means � SE unless otherwise indicated. Thestatistical significance of differences between group means was tested
using Student’s t-test or ANOVA followed by Dunnett’s test. Differ-ences between group means were considered significant if P � 0.05.
The morphological properties of neurons filled with biocytin wereassessed after staining and reconstruction. After recording, slices werefixed in 4% paraformaldehyde in 0.1 M phosphate-buffered saline andresectioned at 50 �m on a microtome. The biotin label was visualizedby standard 3,3�-diaminobenzidine histochemistry, using the Vec-tastain Elite ABC kit (Vector Laboratories, Burlingame, CA). Indi-vidual cells were reconstructed using Neurolucida software (Micro-brightfield, Williston, VT). As reported previously (Gonzalez-Burgoset al. 2005), FS cells were classified into the chandelier cell groupbased on the presence of vertical cartridges of axonal boutons (Fig.1B, right), which are distinctive of this cell type. FS neurons lackingcartridges had axon varicosities more evenly distributed throughoutthe axonal tree and were classified as basket cells (Fig. 1B, left).
R E S U L T S
D1 receptor activation reduces the initial EPSP but not laterresponses in EPSP trains
To focus the present study on FS interneurons, we includedin the data analysis only layers 2/3 interneurons having spikeduration �0.5 ms at half-amplitude, no significant spike fre-quency adaptation, and spike afterhyperpolarizations with
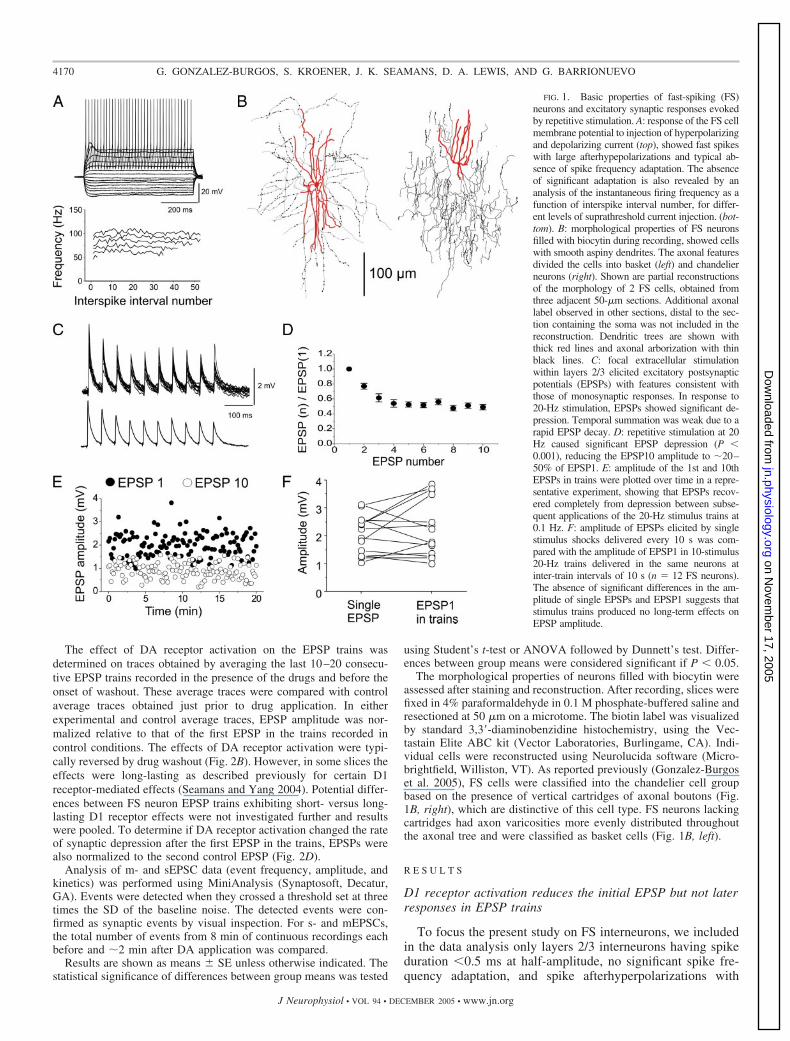
FIG. 1. Basic properties of fast-spiking (FS)neurons and excitatory synaptic responses evokedby repetitive stimulation. A: response of the FS cellmembrane potential to injection of hyperpolarizingand depolarizing current (top), showed fast spikeswith large afterhypepolarizations and typical ab-sence of spike frequency adaptation. The absenceof significant adaptation is also revealed by ananalysis of the instantaneous firing frequency as afunction of interspike interval number, for differ-ent levels of suprathreshold current injection. (bot-tom). B: morphological properties of FS neuronsfilled with biocytin during recording, showed cellswith smooth aspiny dendrites. The axonal featuresdivided the cells into basket (left) and chandelierneurons (right). Shown are partial reconstructionsof the morphology of 2 FS cells, obtained fromthree adjacent 50-�m sections. Additional axonallabel observed in other sections, distal to the sec-tion containing the soma was not included in thereconstruction. Dendritic trees are shown withthick red lines and axonal arborization with thinblack lines. C: focal extracellular stimulationwithin layers 2/3 elicited excitatory postsynapticpotentials (EPSPs) with features consistent withthose of monosynaptic responses. In response to20-Hz stimulation, EPSPs showed significant de-pression. Temporal summation was weak due to arapid EPSP decay. D: repetitive stimulation at 20Hz caused significant EPSP depression (P �0.001), reducing the EPSP10 amplitude to �20–50% of EPSP1. E: amplitude of the 1st and 10thEPSPs in trains were plotted over time in a repre-sentative experiment, showing that EPSPs recov-ered completely from depression between subse-quent applications of the 20-Hz stimulus trains at0.1 Hz. F: amplitude of EPSPs elicited by singlestimulus shocks delivered every 10 s was com-pared with the amplitude of EPSP1 in 10-stimulus20-Hz trains delivered in the same neurons atinter-train intervals of 10 s (n � 12 FS neurons).The absence of significant differences in the am-plitude of single EPSPs and EPSP1 suggests thatstimulus trains produced no long-term effects onEPSP amplitude.
4170 G. GONZALEZ-BURGOS, S. KROENER, J. K. SEAMANS, D. A. LEWIS, AND G. BARRIONUEVO
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

large amplitude (Fig. 1A). The results of this study were basedon recordings from a total of 44 FS neurons from the superfi-cial layers of the principal sulcus region (area 46) of macaque
monkey PFC. The morphology of the recorded FS cells (Fig.1B) was consistent with either chandelier or basket types asdescribed previously (Gonzalez-Burgos et al. 2004, 2005; Zait-
FIG. 2. Effects of DA receptor activation on EPSP trains recorded from FS neurons. A: representative example showing the effects of the D1 agonistSKF81297 (5 �M) on EPSP trains recorded from FS neurons. Note the significant reduction in the amplitude of the 1st EPSP without significant change in theEPSPs evoked by repetitive stimulation. Traces shown are the average of 10 consecutive sweeps right before SKF81297 application (control) and right beforebeginning of washout (SKF81297). B: plot of the amplitude of the 1st and 10th EPSPs in trains versus time in 2 representative experiments. Note the significantreduction in EPSP1 amplitude shortly after beginning of SKF81297 application (indicated by the bar), and the absence of significant change in the EPSP10amplitude. In most neurons, the effect of SKF81297 was reversed after a short period of washout (top); however, in some cells, the effect of a brief applicationof SKF81297 was long-lasting (bottom). C: summary graph showing the effects of SKF81297 on EPSP depression in FS neurons (n � 9). The depolarizationelicited by each EPSP in the trains was divided by the amplitude of the 1st EPSP recorded in control conditions and plotted vs. stimulus number. ANOVAfollowed by contrasts indicated a significant reduction of the EPSP1 amplitude by SKF81297 (*P � 0.05) but no significant change of subsequent EPSPs in thetrains (P � 0.1). In either control recording conditions or after SKF81297 application, repetitive stimulation caused significant depression of the EPSP amplitude(P � 0.001), and EPSPs1-3 were significantly different from EPSP10 (P � 0.01). These data show that D1 receptor stimulation reduced EPSP1 but did notsignificantly alter the rate of depression of EPSPs elicited by repetitive presynaptic spikes (i.e., EPSPs2-10). D: ratio between EPSP10 and EPSP1 within a trainwas increased significantly during D1 receptor activation (P � 0.05). E: relative to EPSP2 recorded in control conditions, the amplitude of EPSP2 and subsequentEPSPs in trains was not altered by D1 receptor stimulation. F–H: effects of DA (10 �M) on the 1st and subsequent EPSPs in FS cell EPSP trains (n � 5) weresimilar to those of SKF81297. DA produced a significant reduction of EPSP1 amplitude (*P � 0.05), but no significant change of subsequent EPSPs in the trains(P � 0.1). Repetitive stimulation caused significant depression of the EPSP amplitude (P � 0.001) and EPSPs1-3 were significantly different from EPSP10 (P �0.01). These data show that D1 receptor stimulation reduced EPSP1 but did not significantly alter the rate of depression of EPSPs elicited by repetitive presynapticspikes (i.e., EPSPs2-10).
4171DOPAMINE AND SHORT-TERM PLASTICITY IN FAST-SPIKING CELLS
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

sev et al. 2005). Stimulus trains of 20-Hz frequency, whichcorresponds to typical mean frequencies of persistent firing inmonkey PFC during working-memory tasks (Sawaguchi 2001;Wang 2001; Williams and Goldman-Rakic 1995), were deliv-ered every 10 s. These 20-Hz input trains produced significantEPSP depression (P � 0.001) with weak temporal summationdue to a fast EPSP decay (Fig. 1C). During synaptic depressioninduced by 20-Hz stimulation, the amplitude of the 10thresponse (EPSP10) was reduced to 20–50% of the first re-sponse (EPSP1) amplitude (Fig. 1D). Between subsequentstimulus trains, the EPSP1 amplitude completely recoveredfrom depression (Fig. 1E). Furthermore, the amplitude ofEPSP1 in trains delivered every 10 s did not differ significantlyfrom the amplitude of single EPSPs evoked every 10 s in thesame neurons, prior to the onset of stimulation with 20-Hztrains (Fig. 1F). The results displayed in Fig. 1, E and F,suggest that stimulus trains did not have long-term effects onthe EPSP amplitude and that EPSP1 in trains delivered at a lowfrequency (0.1 Hz) equals single EPSPs elicited at the samelow frequency.
In monkey dorsolateral PFC, D1-like receptors are signifi-cantly more abundant than receptors of the D2 family (Gold-man-Rakic et al. 1990). Thus to determine the effects of DAreceptor activation on the 20-Hz EPSP trains recorded from FScells (n � 9), we first applied the selective D1 receptor agonistSKF81297 (5 �M). SKF81297 application significantly re-duced the EPSP1 amplitude (to �80-50% of EPSP1 control),whereas the depolarization produced by later EPSPs appearedto be unaffected (Fig. 2A). Figure 2B shows the time course ofthe D1 agonist effect on EPSP1 and EPSP10 amplitudes. Asdisplayed in Fig. 2B, top, in some cells, the effect elicited byDA agonists was short-lasting, whereas in other neurons theDA agonist effect was persistent and did not reverse onwashout (Fig. 2B, bottom). As summarized in Fig. 2C, D1receptor stimulation significantly reduced the EPSP1 ampli-tude to �70% of the EPSP1 recorded in control conditions(Fig. 2C; P � 0.05). In contrast, compared with controlconditions, D1 receptor activation did not change significantlythe depolarization elicited by EPSP2 or the late, stationaryEPSPs (Fig. 2C; P � 0.1). Despite reduction of the first EPSP,short-term EPSP depression induced by input trains in thepresence of SKF81297 was statistically significant (Fig. 2C,P � 0.001). However, the reduction in EPSP1 amplitudewithout significant change in stationary EPSPs suggests thatrepetitive stimulation elicits less depression during D1 receptoractivation than in control conditions. Indeed, SKF81297 sig-nificantly increased the EPSP10/EPSP1 ratio within a train(Fig. 2D). To further examine the effect of D1 receptor stim-ulation on the time course of depression, we fit a single-exponential decay function to the changes in EPSP amplitudeduring trains. The time constant of decay increased followingD1 receptor stimulation, although the difference was not sig-nificant (control: 79 � 19 ms; SKF81297: 142 � 58 ms, n �8, P � 0.3, paired Student’s t-test). Because D1 receptoractivation significantly decreased EPSP1 but not EPSP2 orlater responses (Fig. 2C), we next examined whether the rate ofEPSP depression after EPSP2 was changed by 5 �MSKF81297. Normalization of EPSP amplitude relative toEPSP2 showed that synaptic depression after the second EPSPin the trains was not significantly altered by D1 receptoractivation compared with control conditions (Fig. 2E). More-
over, the time constant of exponential decay after EPSP2 wasnot altered by D1 receptor activation (control: 85 � 28 ms;SKF81297: 108 � 37 ms, n � 8, P � 0.3, paired Student’st-test). Thus the increase in EPSP10/EPSP1 ratio depicted inFig. 2D results only from the reduction of EPSP1 by D1receptor activation without additional changes in the temporalfeatures of synaptic depression. Together, these data show thatthe main effect of D1 receptor stimulation on EPSP trainsrecorded from FS neurons is a reduction of the first EPSP,without changes in the absolute depolarization elicited by thefollowing EPSPs in the trains, once stimulation switches to arepetitive regime. In agreement with these results, D1 receptoractivation selectively reduced, in a frequency-dependent man-ner, the first response during repetitive stimulation of depress-ing excitatory inputs in nucleus accumbens neurons (Hjelmstad2004).
Application of the endogenous agonist DA (10 �M), alsoreduced EPSP1 without affecting the repetitive EPSPs in trainsrecorded from FS neurons (n � 5), thus having effects similarto those of the D1 agonist SKF81297 (Fig. 2, F–H). In thepresence of the D1 receptor antagonist SCH23390 (5 �M), DA(10 �M) failed to decrease the amplitude of the first EPSP(EPSP1 was 90 � 13% of control; n � 3, P � 0.48) orsubsequent EPSPs in the trains (data not shown). Moreover, inthe presence of SCH23390, DA failed to increase the EPSP10/EPSP1 ratio within a train (control ratio, 0.31 � 0.09;DASCH23390 ratio, 0.32 � 0.04; n � 3, P � 0.91). Takentogether, these results suggest that the effects of DA on EPSPtrains in FS interneurons are mediated by activation of D1receptors.
D1 receptor activation had differential effects on the firstresponse in EPSP trains delivered every 10 s. These resultssuggest that DA modulation of excitatory synaptic inputs ontoFS neurons is activity dependent, selectively decreasing theamplitude of EPSPs evoked at low frequency (0.1 Hz). Wetherefore tested the effects of DA on mEPSCs, which resultfrom low-frequency spontaneous release of glutamate at singlesynapses. As shown in Fig. 3, DA did not have significanteffects on the frequency, amplitude, or kinetics of mEPSCs inFS neurons. mEPSCs are recorded in the presence of thesodium channel blocker tetrodotoxin and result from actionpotential-independent (and possibly extracellular calcium-in-dependent) spontaneous release mechanisms (Simkus andStricker 2002). In a previous study of excitatory synapticinputs onto pyramidal cells during normal aging of the monkeyPFC, the frequency of mEPSCs was apparently lower than thatof sEPSCs recorded in the absence of tetrodotoxin (Luebke etal. 2004). These results suggest that low-frequency spontane-ous action potential firing occurred in cells providing excita-tory inputs onto pyramidal neurons. Thus we also tested theeffects of DA on sEPSCs recorded from FS neurons in theabsence of tetrodotoxin. As shown in Table 1, DA applicationdid not alter the frequency, amplitude, or kinetics of sEPSCsrecorded from FS cells. In addition, we found no significantdifferences in frequency, amplitude, or kinetics of sEPSCsrecorded from individual FS neurons before and after tetrodo-toxin application, respectively (Table 1). These results suggestthat in our experimental conditions there is no significantspontaneous presynaptic action potential firing in excitatoryinputs onto FS neurons and that therefore sEPSCs essentiallyrepresent mEPSCs, as reported previously for layers 2/3 neu-
4172 G. GONZALEZ-BURGOS, S. KROENER, J. K. SEAMANS, D. A. LEWIS, AND G. BARRIONUEVO
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

rons in rat neocortex (Simkus and Stricker 2002). In addition,the m- and sEPSC data indicate that DA does not change thepostsynaptic response to synaptic release of glutamate and thatthe effects of DA are exclusively associated with action po-tential-evoked glutamate release.
Effects of DA receptor activation do not involve temporalEPSP summation or activation of N-methyl-D-aspartatereceptor channels
In rat PFC pyramidal neurons, the depolarization elicited bylate EPSPs in trains is enhanced by DA via a D1 receptor-mediated increase in temporal summation (Seamans et al.2001). Here we found that in PFC FS neurons, D1 receptoractivation did not enhance the depolarization elicited by lateEPSPs. Thus we tested whether this difference is due to a lackof DA effect on temporal summation in FS cell EPSP trains. Toestimate the effect of temporal summation, we calculated thefraction of the EPSP-induced depolarization, relative to theresting potential, that was due to summation of each EPSP withthe decay phase of the prior response in the train (Fig. 4A). Onaverage, EPSP summation accounted for only 15–30% of thedepolarization elicited by the EPSPs and did not change sig-nificantly throughout the trains (Fig. 4B). In addition, a com-parison between EPSP trains recorded before and after DAreceptor activation, showed that while reducing EPSP1 (Fig.2), activation of D1 receptors produced no significant differ-ences in temporal summation in the same cells (Fig. 4B). Thusin contrast to the effect observed in PFC pyramidal cells(Seamans et al. 2001), D1 receptor activation did not increasesummation in EPSP trains recorded from FS neurons.
The D1 receptor-mediated increase in EPSP summation thatenhances late responses in pyramidal cell trains, is N-methyl-D-aspartate (NMDA) receptor dependent (Seamans et al. 2001).Previous studies of rat neocortex and hippocampus suggest thatexcitatory synapses in FS interneurons have a low NMDAreceptor content (Angulo et al. 1999; Geiger et al. 1997;Goldberg et al. 2003a,b; Nyiri et al. 2003). Therefore the lackof effect of D1 receptor activation on temporal summation inPFC FS neurons may be explained by a small NMDA EPSPcomponent. To determine the contribution of NMDA receptorsto EPSP trains, we tested the effects of the NMDA receptorantagonist D,L-2-amino-5-phosphonovaleric acid (D,L-APV).Figure 5A shows that application of APV (100 �M) did notalter significantly the EPSP trains, suggesting a small or absentNMDA EPSP component in FS interneurons from monkeyPFC.
FIG. 3. Dopamine receptor activation does not alter spontaneous glutamaterelease at synapses onto FS neurons. A: representative recordings of mEPSCsfrom a PFC FS neuron. Application of DA (10 �M) did not significantlychange mEPSC frequency or amplitude compared with baseline conditions. B:analysis of the distribution and cumulative distribution of mEPSC amplitudeindicated that DA had no significant effects. C, left: averages of all mEPSCsrecorded from a representative FS neuron in control vs. DA condition show nosignificant effects of DA application on mEPSCs amplitude and time course.Right: bar graphs showing the absence of effects of DA receptor stimulation onthe mean mEPSC frequency, amplitude, 10–90% rise time, and monoexpo-nential decay time constant. Results were obtained after recording from n � 6FS neurons.
TABLE 1. Effects of dopamine receptor activation and tetrodotoxin on spontaneous EPSCs recorded from FS neurons in monkey PFC
Experiment 1 Experiment 2
Control Dopamine Control Tetrodotoxin
Frequency, Hz 3.18 � 0.93 3.10 � 0.78 2.34 � 0.42 2.06 � 0.33Amplitude, pA 13.4 � 2.1 13.3 � 1.9 11.4 � 0.6 10.4 � 0.97Rise time, msec 0.85 � 0.04 0.85 � 0.05 0.89 � 0.05 0.93 � 0.04Decay time, msec 1.84 � 0.16 1.85 � 0.18 1.89 � 0.20 1.85 � 0.21
Two groups of experiments were performed. In experiment 1, (sEPSCs) were recorded from fast-spiking (FS) neurons (n � 5) before and after a 5-minapplication of dopamine (DA) (10 �M). In experiment 2, sEPSCs were recorded from a different group of FS neurons (n � 4) before and after addition oftetrodotoxin (1 �M). Paired t-test was used to compare sEPSC parameters in control vs. DA or tetrodotoxin conditions. No significant differences were observedbetween group means (P � 0.05). Data are expressed as means � SE.
4173DOPAMINE AND SHORT-TERM PLASTICITY IN FAST-SPIKING CELLS
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

A smaller NMDA component in FS neurons than in pyra-midal cells may result from a more significant voltage-depen-dent Mg2 block of NMDA channels. In monkey dorsolateralPFC in vitro, the resting potential is significantly depolarized inFS cells (�67.7 � 1.2 mV) compared with pyramidal neurons(�74.4 � 1.1 mV) (Gonzalez-Burgos et al. 2004). In thepresent study, FS cells showing D1 effect and FS cells testedwith APV had resting potentials of �66.7 � 5.2 and �67.2 �2.9 mV, respectively. However, the resting potential of FS cellsin monkey PFC is actually hyperpolarized compared with therange of potentials (�65 to �58 mV) at which D1 receptormodulation of EPSP trains in pyramidal cells is NMDA de-pendent (Seamans et al. 2001). Therefore subthreshold mem-brane depolarization may reveal an NMDA-mediated increasein EPSP duration that could enhance EPSP summation. Asshown in Fig. 5B, depolarization of FS neurons, however,produced effects opposite to those of NMDA channel activa-tion because EPSP duration was significantly shortened atpotentials near spike threshold (Fig. 5B). These results confirmprevious findings indicating that in FS neurons voltage-depen-dent conductances curtail the EPSP duration and decreasetemporal summation at depolarized subthreshold potentials(Galarreta and Hestrin 2001; Gonzalez-Burgos et al. 2004). Incontrast, in cortical pyramidal cells from both rat and monkey,voltage-gated channels increase EPSP duration, via an effect
that is independent of, but potentially synergistic with, theeffect of NMDA receptors (Gonzalez-Burgos and Barrionuevo2001; Gonzalez-Burgos et al. 2004; Stuart and Sakmann 1995).Therefore the actions of voltage-gated and synaptic conduc-tances give rise to significant differences in temporal EPSPsummation in FS neurons and pyramidal cells. Together, thisand previous studies suggest that, in FS neurons at the sub-threshold membrane potential range, NMDA receptors have asmall impact on somatic EPSP duration and thus on temporalsummation.
D I S C U S S I O N
In this study, we found that at excitatory inputs onto FSneurons of monkey PFC, DA modulated EPSP amplitude in anactivity-dependent manner. Specifically, when excitatory in-puts were stimulated with trains at inter-train intervals of 10 s,D1 receptor activation decreased the amplitude of the firstEPSP, but not the amplitude of later EPSPs in the train. Therepetitive EPSPs (EPSPs2-10 in the trains), showed similardegrees of short-term depression in control or DA conditions.This suggests that the basic mechanisms of temporal codingand gain control are retained during DA neuromodulation ofFS cell inputs. In contrast with previous findings in PFCpyramidal cells, we found that in FS neurons D1 receptoractivation did not change temporal summation of EPSPs. Inaddition, NMDA receptors did not contribute significantly toEPSPs in the subthreshold potential range. This suggests thatNMDA receptors are an unlikely substrate for DA modulationof EPSP integration in FS neurons. Thus D1-NMDA receptorinteractions in PFC microcircuits appear to be specific topyramidal cell inputs.
FIG. 5. Contribution of NMDA receptors to excitatory transmission in FSneurons. A: application of the NMDA receptor blocker 2-amino-5-phospho-novaleric acid (100 �M) did not affect the EPSPs trains recorded at the FS cellresting membrane potential. The traces show EPSP trains recorded from a FSneuron that exhibited one of the greatest degrees of temporal summationduring the trains. B: depolarization of FS neurons in the subthreshold mem-brane potential range had effects opposite to those expected for NMDAchannel activation because it reduced the EPSP duration (n � 3); *, truncatedaction potential.
FIG. 4. Temporal summation during FS cell EPSP trains was not influencedby dopamine receptor activation. A: contribution of temporal summation to thedepolarization produced by EPSPs in trains was estimated by calculating thefraction of the total depolarization (2) that was accounted by summation withthe decay phase of the previous EPSP in the train (1). B: temporal summationduring FS cell EPSP trains was calculated as described in A for trains recordedbefore and after DA receptor stimulation. Data were pooled for experiments inwhich either SKF81297 or DA was applied to the slices and producedsignificant changes in EPSP1 amplitude (see Fig. 2). Note the absence ofsignificant effects of DA receptor activation on the degree of temporalsummation.
4174 G. GONZALEZ-BURGOS, S. KROENER, J. K. SEAMANS, D. A. LEWIS, AND G. BARRIONUEVO
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

DA modulation of short-term plasticity in FS neurons
At excitatory connections onto neocortical neurons, depres-sion predominates if the synapses display high probability ofglutamate release (Pr) in response to single stimuli (Atzori etal. 2001; Rozov et al. 2001). Manipulations that reduce Prdecreased the amplitude of EPSP1 but not of subsequentEPSPs in trains elicited at depressing synapses (Markram andTsodyks 1996; Tsodyks and Markram 1997). Similarly, in thisstudy, D1 receptor activation selectively reduced EPSP1, sug-gesting that DA may produce a reduction in Pr via presynapticeffects. Because the amplitude of EPSP2 and subsequent re-sponses in the trains were not altered by DA relative to controlconditions, short-term synaptic depression of repetitive EPSPsseemed to be independent of the DA-induced decrease inEPSP1. To examine the complex mechanisms underlying thiseffect was beyond the scope of the present study. A mechanismthat could yield depressed EPSPs independent of prior re-sponses in the trains is release-independent depression (RID).During RID, EPSPs depress independent of whether precedingstimuli fail to evoke release (Thomson and Bannister 1999).The mechanisms of RID imply significant differences betweenrelease evoked by single stimuli or repetitive stimulation(Fuhrmann et al. 2004). Thus during RID glutamate releasemay be refractory to DA effects that, however, decrease releaseevoked by temporally isolated spikes, i.e., the first spike in atrain. The contribution of RID to repetitive EPSPs in FSneurons remains to be determined.
If D1 receptor activation modulates glutamate release at FScell inputs, then DA effects should be at least in part presyn-aptic. In the deep layers of monkey PFC, D1 receptors arelocalized in excitatory boutons synapsing with pyramidal cells,but are distinctively absent from excitatory boutons targetingparvalbumin-positive interneurons (Paspalas and Goldman-Rakic 2005). These data argue against presynaptic effects ofDA at glutamate synapses onto FS neurons. However, in thesuperficial layers of monkey PFC area 46, certain excitatoryboutons synapsing with parvalbumin-containing dendritesovertly express D1 receptors and contain numerous D1 recep-tor immunoparticles (C. D. Paspalas, personal communica-tion). Ultrastructural studies thus imply both target- and layer-specific heterogeneity in the expression of D1 receptors inexcitatory axons synapsing onto PFC neurons. Likewise, ourphysiological experiments also suggest heterogeneity becausein a minority of the recorded FS cells, DA caused no reductionof EPSP1 amplitude (see METHODS). This subgroup includedboth basket and chandelier cells, suggesting no correlationbetween heterogeneity in DA effects and morphological sub-populations of FS neurons. In contrast with our findings, inlayer 5 of ferret PFC, DA did not affect EPSPs in most FSneurons but strongly reduced EPSP amplitude in a minority ofthese cells (Gao and Goldman-Rakic 2003). Besides differ-ences between species, the discrepancy between our results andthose of Gao and Goldman-Rakic (2003) may be explained bya greater incidence of D1 receptor expression in terminalscontacting FS neurons in the superficial compared with thedeep cortical layers of PFC (C. D. Paspalas, personal commu-nication).
Our results suggesting a small NMDA receptor contributionto EPSP trains in PFC FS neurons are consistent with previousstudies indicating that a small NMDA EPSC component and a
low NMDA content are found in excitatory synapses onto FSneurons from rat hippocampus and neocortex (Angulo et al.1999; Geiger et al. 1997; Nyiri et al. 2003). In FS neurons,synaptically evoked dendritic Ca2 transients are not sensitiveto NMDA antagonists or are significantly less sensitive than inother interneuron subtypes (Goldberg et al. 2003a,b). In someFS cell synapses, NMDA antagonists partially reduced thedendritic Ca2 transients, without simultaneously altering theamplitude or time course of somatic EPSPs (Goldberg et al.2003a,b). Thus NMDA receptors may mediate Ca2 influx atcertain FS cell synapses without having a significant impact onsomatic EPSP time course. Voltage-clamp experiments suggestthat some NMDA receptor activation occurs at very depolar-ized potentials in FS neurons from rat neocortex (Angulo et al.1999). However, our data suggest that DA regulation of short-term depression of subthreshold EPSPs in FS cells is unlikelyto involve D1 modulation of synaptically activated NMDAreceptors in contrast to PFC pyramidal cells (Seamans et al.2001).
Functional implications
In computational and in vitro experimental models, persis-tent firing in PFC is strongly influenced or dominated byrecurrent inhibition (Fellous and Sejnowski 2003; McCormicket al. 2003; Seamans et al. 2003; Wang et al. 2004). Modelingstudies indeed show that DA-mediated modulation of GABA-mediated inhibition is critical for the stability of persistentactivity (Brunel and Wang 2001; Durstewitz and Seamans2002; Durstewitz et al. 2000). The prevalent synaptic depres-sion in their inputs and outputs suggests that FS neuronsgenerate transient inhibition but are not a significant source ofsustained recurrent inhibition (Beierlein et al. 2003; Pouilleand Scanziani 2004). In pyramidal cells, DA counteracts EPSPdepression by increasing the amplitude of late, stationaryEPSPs, potentially favoring sustained postsynaptic firing bysustained presynaptic stimulation. In FS cells, in contrast, DAdid not potentiate the late EPSPs in trains, suggesting that DAmodulation does not favor sustained recruitment of FS neuronsby input trains. Recurrent inhibition during persistent firingmay thus be mediated by non-FS interneurons, which typicallyexhibit short-term facilitation instead of depression, in eitherinputs or outputs (Beierlein et al. 2003; Gonzalez-Burgos et al.2004).
The DA effects found here are consistent with a reduction ofEPSPs elicited by temporally isolated presynaptic spikes(EPSP1) and no reduction of EPSPs elicited by repetitivespikes (EPSPs2-10), i.e., spikes elicited shortly after the pre-ceding one. Repetitive spikes elicited short-term depression ofEPSPs2-10 and because the absolute amplitude of EPSPs2-10was not affected by D1 receptor activation, this depression wasnot altered by DA. Temporal coding by depressing synapsesdepends strongly on the effect of the initial EPSPs afterpresynaptic firing frequency switches from low to high, be-cause the initial EPSPs are not significantly depressed com-pared with the stationary responses (Abbott et al. 1997; Tso-dyks and Markram 1997). In either control conditions or afterD1 receptor stimulation, EPSPs1-3 were significantly differentfrom stationary EPSPs (P � 0.05, ANOVA followed bycontrasts) despite the reduction of EPSP1 amplitude by DAreceptor activation. In addition, the absence of DA modulation
4175DOPAMINE AND SHORT-TERM PLASTICITY IN FAST-SPIKING CELLS
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

of stationary EPSPs during 20-Hz stimulation suggests that DAdoes not enhance the ability of synapses onto FS neurons tocode presynaptic firing rate per se (Abbott et al. 1997; Tsodyksand Markram 1997). As suggested previously (Abbott et al.1997), these results indicate that the basic temporal coding andgain control mechanisms at depressing synapses onto FS neu-rons are retained during DA neuromodulation. Spike trainsrecorded from the monkey PFC during working-memory tasksdisplay increased inter-spike interval variability, approximat-ing a Poisson process, and suppression of specific frequenciesin the power spectra (Compte et al. 2003). Thus spike trains inPFC may contain spike timing-dependent information relevantfor task-related events, although the precise nature of thisinformation remains to be determined.
If spike timing-dependent information is transmitted viapresynaptic action potential bursts at depressing synapses, thenEPSP bursts in FS neurons may contain an activity-dependentsignal related to transitions in the level of network activity.Because the effects of DA described here may favor detectionof bursts relative to low-frequency background synaptic activ-ity, then DA may actually enhance the signal-to-noise ratio atFS cell inputs. The results reported here as well as those of aprevious electrophysiological and ultrastructural studies (Gaoand Goldman-Rakic 2003; Paspalas and Goldman-Rakic 2005;C. Paspalas, personal communication) suggest heterogeneity inthe effects of DA on FS cells. Therefore it is possible that thepotential enhancenment of the signal-to-noise ratio is observedonly at particular subtypes of depressing synaptic inputs ontoFS cells. Similar mechanisms may operate at depressing syn-apses onto pyramidal neurons so long as transmission is me-diated mostly by non-NMDA receptors, as in hyperpolarizedstates. When PFC pyramidal cells are depolarized, activation ofNMDA receptors, together with the subthreshold effects ofNa channels counteract synaptic depression, actually convert-ing it into facilitation (Gonzalez-Burgos and Barrionuevo2001; Williams and Stuart 1999). This NMDA-dependentfacilitation may effectively increase the strength of connectionsabove a critical threshold necessary for the stability of persis-tent firing (Brunel and Wang 2001; Wang 2001). In contrast, inthe absence of significant NMDA receptor contribution, in FScells, depolarization decreases EPSP duration and thus en-hances the effects of synaptic depression. Our results thereforesuggest that the contribution of D1-NMDA receptor synergy totemporal EPSP integration is pyramidal cell-specific and thatDA has different physiological effects at different componentsof the local PFC circuits.
A C K N O W L E D G M E N T S
We thank Drs. Daniel Durstewitz, Edda Thiels, and Etienne Sibille foruseful comments on a previous version of this manuscript.
Present address of S. Kroener: Dept. of Physiology and Neuroscience,Medical University of South Carolina.
G R A N T S
This work was supported by National Institute of Mental Health GrantsMH-45156 and MH-51234 and by a National Alliance for Research onSchizophrenia and Depression Young Investigator Award to G. Gonzalez-Burgos.
R E F E R E N C E S
Abbott LF, Varela JA, Sen K, and Nelson SB. Synaptic depression andcortical gain control. Science 275: 220–224, 1997.
Angulo MC, Rossier J, and Audinat E. Postsynaptic glutamate receptors andintegrative properties of fast-spiking interneurons in the rat neocortex.J Neurophysiol 82: 1295–1302, 1999.
Atzori M, Lei S, Evans DI, Kanold PO, Phillips-Tansey E, McIntyre O,and McBain CJ. Differential synaptic processing separates stationary fromtransient inputs to the auditory cortex. Nat Neurosci 4: 1230–1237, 2001.
Beierlein M, Gibson JR, and Connors BW. Two dynamically distinctinhibitory networks in layer 4 of the neocortex. J Neurophysiol 90: 2987–3000, 2003.
Brunel N and Wang X-J. Effects of neuromodulation in a cortical networkmodel of object working memory dominated by recurrent inhibition. J Com-put Neurosci 11: 63–85, 2001.
Compte A, Constantinidis C, Tegner J, Raghavachari S, Chafee MV,Goldman-Rakic PS, and Wang XJ. Temporally irregular mnemonic per-sistent activity in prefrontal neurons of monkeys during a delayed responsetask. J Neurophysiol 90: 3441–3454, 2003.
Durstewitz D and Seamans JK. The computational role of dopamine D1receptors in working memory. Neural Networks 15: 561–572, 2002.
Durstewitz D, Seamans JK, and Sejnowski TJ. Dopamine-mediated stabi-lization of delay-period activity in a network model of prefrontal cortex.J Neurophysiol 83: 1733–1750, 2000.
Fellous JM and Sejnowski TJ. Regulation of persistent activity by back-ground inhibition in an in vitro model of a cortical microcircuit. CerebCortex 13: 1232–1241, 2003.
Fuhrmann G, Cowan A, Segev I, Tsodyks M, and Stricker C. Multiplemechanisms govern the dynamics of depression at neocortical synapses ofyoung rats. J Physiol 557: 415–438, 2004.
Fuster JM. The Prefrontal Cortex: Anatomy, Physiology and Neuropsychol-ogy of the Frontal Lobe. New York: Raven, 1997.
Galarreta M and Hestrin S. Spike transmission and synchrony detection innetworks of GABAergic interneurons. Science 292: 2295–2299, 2001.
Gao WJ and Goldman-Rakic PS. Selective modulation of excitatory andinhibitory microcircuits by dopamine. Proc Natl Acad Sci USA 100: 2836–2841, 2003.
Gao WJ, Krimer LS, and Goldman-Rakic PS. Presynaptic regulation ofrecurrent excitation by D1 receptors in prefrontal circuits. Proc Natl AcadSci USA 98: 295–300, 2001.
Geiger JR, Lubke J, Roth A, Frotscher M, and Jonas P. SubmillisecondAMPA receptor-mediated signaling at a principal neuron-interneuron syn-apse. Neuron 18: 1009–1023, 1997.
Goldberg JH, Tamas G, Aronov D, and Yuste R. Calcium microdomains inaspiny dendrites. Neuron 40: 807–821, 2003a.
Goldberg JH, Yuste R, and Tamas G. Ca2 imaging of mouse neocorticalinterneurone dendrites: contribution of Ca2-permeable AMPA and NMDAreceptors to subthreshold Ca2dynamics. J Physiol 551: 67–78, 2003b.
Goldman-Rakic PS, Castner SA, Svensson TH, Siever LJ, and WilliamsGV. Targeting the dopamine D1 receptor in schizophrenia: insights forcognitive dysfunction. Psychopharmacology 174: 3–16, 2004.
Goldman-Rakic PS, Lidow MS, and Gallager DW. Overlap of dopaminer-gic, adrenergic, and serotoninergic receptors and complementarity of theirsubtypes in primate prefrontal cortex. J Neurosci 10: 2125–2138, 1990.
Gonzalez-Burgos G, and Barrionuevo G. Voltage-gated sodium channelsshape subthreshold EPSPs in layer 5 pyramidal neurons from rat prefrontalcortex. J Neurophysiol 86: 1671–1684, 2001.
Gonzalez-Burgos G, Barrionuevo G, and Lewis DA. Horizontal synapticconnections in monkey prefrontal cortex: an in vitro electrophysiologicalstudy. Cereb Cortex 10: 82–92, 2000.
Gonzalez-Burgos G, Krimer LS, Povysheva NV, Barrionuevo G, andLewis DA. Functional properties of fast spiking interneurons and theirsynaptic connections with pyramidal cells in primate dorsolateral prefrontalcortex. J Neurophysiol 93: 942–953, 2005.
Gonzalez-Burgos G, Krimer LS, Urban NN, Barrionuevo G, and LewisDA. Synaptic efficacy during repetitive activation of excitatory inputs inprimate dorsolateral prefrontal cortex. Cereb Cortex 14: 530–542, 2004.
Gonzalez-Burgos G, Kroner S, Krimer LS, Seamans JK, Urban NN,Henze DA, Lewis DA, and Barrionuevo G. Dopamine modulation ofneuronal function in the monkey prefrontal cortex. Physiol Behav 77:537–543, 2002.
Gonzalez-Islas C and Hablitz JJ. Dopamine enhances EPSCs in layer II-IIIpyramidal neurons in rat prefrontal cortex. J Neurosci 23: 867–875, 2003.
Gorelova N, Seamans JK, and Yang CR. Mechanisms of dopamine activa-tion of fast-spiking interneurons that exert inhibition in rat prefrontal cortex.J Neurophysiol 88: 3150–3166, 2002.
4176 G. GONZALEZ-BURGOS, S. KROENER, J. K. SEAMANS, D. A. LEWIS, AND G. BARRIONUEVO
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from

Hempel CM, Hartman KH, Wang XJ, Turrigiano GG, and Nelson SB.Multiple forms of short-term plasticity at excitatory synapses in rat medialprefrontal cortex. J Neurophysiol 83: 3031–3041, 2000.
Henze DA, Gonzalez-Burgos GR, Urban NN, Lewis DA, and BarrionuevoG. Dopamine increases excitability of pyramidal neurons in primate pre-frontal cortex. J Neurophysiol 84: 2799–2809, 2000.
Hjelmstad GO. Dopamine excites nucleus accumbens neurons through thedifferential modulation of glutamate and GABA release. J Neurosci 24:8621–8628, 2004.
Kawaguchi Y and Kubota Y. GABAergic cell subtypes and their synapticconnections in rat frontal cortex. Cereb Cortex 7: 476–486, 1997.
Law-Tho D, Hirsch JC, and Crepel F. Dopamine modulation of synaptictransmission in rat prefrontal cortex: an in vitro electrophysiological study.Neurosci Res 21: 151–160, 1994.
Le Moine C and Gaspar P. Subpopulations of cortical GABAergic interneu-rons differ by their expression of D1 and D2 dopamine receptor subtypes.Brain Res Mol Brain Res 58: 231–236, 1998.
Lewis DA, Campbell MJ, Foote SL, Goldstein M, Morrison, and JH. Thedistribution of tyrosine hydroxylase-immunoreactive fibers in primate neo-cortex is widespread but regionally specific. J Neurosci 7: 279–290, 1987.
Luebke JI, Chang YM, Moore TL, and Rosene DL. Normal aging results indecreased synaptic excitation and increased synaptic inhibition of layer 2/3pyramidal cells in the monkey prefrontal cortex. Neuroscience 125: 277–288, 2004.
Markram H and Tsydyks M. Redistribution of synaptic efficacy betweenneocortical pyramidal neurons. Nature 382: 807–810, 1996.
McCormick DA, Shu Y, Hasenstaub A, Sanchez-Vives M, Badoual M, andBal T. Persistent cortical activity: mechanisms of generation and effects onneuronal excitability. Cereb Cortex 13: 1219–1231, 2003.
Muly EC, Szigeti K, and Goldman-Rakic PS. D1 receptor in interneurons ofmacaque prefrontal cortex: distribution and subcellular localization. J Neu-rosci 18: 10553–10565, 1998.
Nyiri G, Stephenson BM, Freund TF, and Somogyi P. Large variability insynaptic N-methyl-D-aspartate receptor density on interneurons and a com-parison with pyramidal-cell spines in the rat hippocampus. Neuroscience119: 347–363, 2003.
Paspalas CD and Goldman-Rakic PS. Presynaptic D1 dopamine receptors inprimate prefrontal cortex: target-specific expression in the glutamatergicsynapse. J Neurosci 25: 1260–1267, 2005.
Porter LL, Rizzo E, and Hornung JP. Dopamine affects parvalbuminexpression during cortical development in vitro. J Neurosci 19: 8990–9003,1999.
Pouille F and Scanziani M. Routing of spike series by dynamic circuits in thehippocampus. Nature 429: 717–723, 2004.
Rozov A, Burnashev N, Sakmann B, and Neher E. Transmitter releasemodulation by intracellular Ca2 buffers in facilitating and depressing nerveterminals of pyramidal cells in layer 2/3 of the rat neocortex indicates atarget cell-specific difference in presynaptic calcium dynamics. J Physiol531: 807–826, 2001.
Sawaguchi T. The effects of dopamine and its antagonists on directionaldelay-period activity of prefrontal neurons in monkeys during an oculomo-tor delayed-response task. Neurosci Res 41: 115–128, 2001.
Seamans JK, Durstewitz D, Christie B, Stevens CF, and Sejnowski TJ.Dopamine D1/D5 receptor modulation of excitatory synaptic inputs to layerV prefrontal cortex neurons. Proc Natl Acad Sci USAmerica 98: 301–306,2001.
Seamans JK, Nogueira L, and Lavin A. Synaptic basis of persistent activityin prefrontal cortex in vivo and in organotypic cultures. Cereb Cortex 13:1242–1250, 2003.
Seamans JK and Yang CR. The principal features and mechanisms ofdopamine modulation in the prefrontal cortex. Prog Neurobiol 74: 1–58,2004.
Sesack SR, Bressler CN, and Lewis DA. Ultrastructural associations betweendopamine terminals and local circuit neurons in the monkey prefrontalcortex: a study of calretinin-immunoreactive cells. Neurosci Lett 200: 9–12,1995.
Sesack SR, Hawrylak VA, Melchitzky DS, and Lewis DA. Dopamineinnervation of a subclass of local circuit neurons in monkey prefrontalcortex: ultrastructural analysis of tyrosine hydroxylase and parvalbuminimmunoreactive structures. Cereb Cortex 8: 614–622, 1998.
Simkus CR and Stricker C. Properties of mEPSCs recorded in layer IIneurones of rat barrel cortex. J Physiol 545: 509–520, 2002.
Stuart G and Sakmann B. Amplification of EPSPs by axosomatic sodiumchannels in neocortical pyramidal neurons. Neuron 15: 1065–1076, 1995.
Thomson AM and Bannister AP. Release-independent depression at pyra-midal inputs onto specific cell targets: dual recordings in slices of rat cortex.J Physiol 519: 57–70, 1999.
Tsodyks MV and Markram H. The neural code between neocortical pyra-midal neurons depends on neurotransmitter release probability. Proc NatlAcad Sci USA 94: 719–723, 1997.
Urban NN, Gonzalez-Burgos G, Henze DA, Lewis DA, and BarrionuevoG. Selective reduction by dopamine of excitatory synaptic inputs to pyra-midal neurons in primate prefrontal cortex. J Physiol 539: 707–712, 2002.
Wang X-J. Synaptic reverberation underlying mnemonic persistent activity.Trends Neurosci 24: 455–463, 2001.
Wang XJ, Tegner J, Constantinidis C, and Goldman-Rakic PS. Division oflabor among distinct subtypes of inhibitory neurons in a cortical microcircuitof working memory. Proc Natl Acad Sci USA 101: 1368–1373, 2004.
Watanabe M, Kodama T, and Hikosaka K. Increase of extracellular dopa-mine in primate prefrontal cortex during a working memory task. J Neuro-physiol 78: 2795–2798, 1997.
Williams GV and Goldman-Rakic PS. Modulation of memory fields bydopamine D1 receptors in prefrontal cortex. Nature 376: 572–575, 1995.
Williams SM and Goldman-Rakic PS. Widespread origin of the primatemesofrontal dopamine system. Cereb Cortex 8: 321–345, 1998.
Williams SR and Stuart GJ. Mechanisms and consequences of actionpotential burst firing in rat neocortical pyramidal neurons. J Physiol 521:467–482, 1999.
Zaitsev AV, Gonzalez-Burgos G, Povysheva NV, Kroner S, Lewis DA, andKrimer LS. Localization of calcium-binding proteins in physiologically andmorphologically characterized interneurons of monkey dorsolateral prefron-tal cortex. Cereb Cortex 15: 1178–1186, 2005.
4177DOPAMINE AND SHORT-TERM PLASTICITY IN FAST-SPIKING CELLS
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on Novem
ber 17, 2005 jn.physiology.org
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















