
An organotypic culture model to study nigro-striatal degeneration
9
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of An organotypic culture model to study nigro-striatal degeneration

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Journal of Neuroscience Methods 188 (2010) 205–212
Contents lists available at ScienceDirect
Journal of Neuroscience Methods
journa l homepage: www.e lsev ier .com/ locate / jneumeth
An organotypic culture model to study nigro-striatal degeneration
Fabio Cavalierea,∗, Edurne San Vicenteb, Carlos Matutea,1
a Neurotek-UPV/EHU, Department of Neuroscience and CIBERNED, Vizcaya Technology Park, Ed. 205, E-48170 Zamudio, Vizcaya, Spainb Departamento de Bioquímica y Biología Molecular, Facultad de Ciencias, Universidad de Extremadura, Avenida de Elvas s/n, E-06071 Badajoz, Spain
a r t i c l e i n f o
Article history:Received 15 December 2009Received in revised form 3 February 2010Accepted 4 February 2010
Keywords:Dopaminergic neuronSubventricular zoneStriatumSubstantia nigraParkinson’s disease
a b s t r a c t
Functional and reliable in vitro models of Parkinson’s disease (PD) are valuable for studying mechanismsof dopaminergic degeneration before proceeding to animal testing. At present, all in vitro models involvesubstitute cell types and thus their direct relevance to PD is questionable. Here, we describe an organ-otypic culture model which conserves the 3D architecture of the nigro-striatal pathway, together withthe subventricular zone and cerebral cortex, and recapitulates a specific pattern of dopaminergic degen-eration which is the principal hallmark of PD. The organotypic culture is kept in vitro for up to 12 daysand dopaminergic degeneration is induced by the simple cutting of dopaminergic fibers. This organotypicmodel represents a rapid and useful method (30 min/pup for preparation and up to 12 days of cultivation)to investigate in vitro the mechanisms underlying neuronal death and protection, as well as neurogenesisand repair after nigro-striatal neurodegeneration.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Parkinson’s disease (PD) is a chronic and progressive neurode-generative disease involving disturbances in movement control,tremor, rigidity, bradykinesia in all kinds of movements (e.g. walk-ing, sitting, eating, talking) and postural instability. The etiologyof PD is not yet fully understood, but symptoms are clearly asso-ciated with the selective degeneration of dopaminergic neuronsin the substantia nigra (SN). Dopamine deficit in the nigro-striatalpathway which leads from the SN to the striatum, induces furtherloss of striatal neurons, and to a variety of characteristic cytologi-cal changes including the aggregation of alpha-synuclein in Lewybodies (Halliday and McCann, 2008).
Current strategies aimed to restore the dopaminergic systemare either of a pharmacological nature, such as the adminis-tration of levodopa (L-DOPA) or a restorative nature involvingstem cell grafting, as well as endogenous neurogenesis (Winneret al., 2009; Lindvall and Kokaia, 2009). Recently, endogenousstem cells have been identified in neurogenic regions of the adultbrain. In particular, the subventricular zone is believed to be themain neurogenic niche with potential to generate neurons in pri-marily non-neurogenic areas, such as the midbrain and striatum(Arias-Carrión et al., 2006). Modulation of neurogenesis repre-sents an exciting new generation of therapies, since it is the most
∗ Corresponding author. Tel.: +34 94601 8284.E-mail addresses: [email protected] (F. Cavaliere),
[email protected] (E.S. Vicente), [email protected] (C. Matute).1 Tel.: +34 94601 3244.
non-invasive treatment involving autologous neuronal replace-ment.
One well established model of PD is the 6-hydroxy-dopamine(6-OHDA) rat model. 6-OHDA, which is administered stereo-taxically into the brain, induces a selective degeneration ofdopaminergic neurons comparable to that observed in PD patients.It uses the same intracellular transport mechanisms as dopamineand noradrenaline and induces neuronal death by means ofoxidative stress. Intra-carotid injection of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in the monkey or mouse alsorecapitulates features of Parkinson’s disease and representsanother useful model for the study of the neurobiology and ofnew forms of drug therapy for Parkinson’s disease (Jackson-Lewisand Przedborski, 2007). MPTP, which is selectively transportedinto SN neurons, is converted intracellularly into its highly activemetabolite 1-methyl-4-phenylpyridinium (MPP+) which provokesthe block of mitochondrial complex I leading to neuronal death.
The value of in vitro models to effectively recapitulate Parkin-sonian features is less clear. Primary mesencephalic cell lines ordopaminergic-neuron-derived cell lines (e.g. human SH-SY5Y andrat PC12) have been used to study the mechanisms of dopaminergicneuronal degeneration. The model is initiated by chronic appli-cation of drugs affecting mitochondrial complexes with furthergeneration of reactive oxygen species. However, all in vitro modelscurrently in use employ substitute cell types and thus, their rel-evance to Parkinson’s disease is reduced (for a review of in vivoand in vitro models of PD, see Shimohama et al., 2003). In contrast,organotypic cultures from different regions of the CNS have beensuccessfully used to study neurodegeneration associated with dif-ferent brain diseases (Cavaliere et al., 2006; Noraberg et al., 1999;
0165-0270/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.jneumeth.2010.02.008

Author's personal copy
206 F. Cavaliere et al. / Journal of Neuroscience Methods 188 (2010) 205–212
Table 1Schematic view of protocol adopted. DIV (days in vitro); PI (propidium iodide); 6-OHDA (6-hydroxydopamine); IF (immunofluorescence).
Gogolla et al., 2006). This culture model simulates better the invivo situation since it preserves natural cell-to-cell contacts andthe organotypic structure of the area. Moreover, the model permitselectrophysiological, as well as immunofluorescence analyses.
An example of an organotypic culture used to study connectionsbetween the cortex, striatum and substantia nigra is the co-culturemodel described by Plenz and Kitai (1996). These authors platedthree different slices proceeding from different cuts of the samebrain and observed axon elongation from the substantia nigra to thestriatum and from the cortex to the striatum. The principal disad-vantage of this model was the observed remodeling of the originalnigro-striatal pathway, besides of the absence of the subventricularzone.
In the novel protocol which we present here (see Table 1for schematic representation), we employ a modification of thePlenz and Kitai model in which we maintained the 3D structurebetween the substantia nigra–striatum–subventricular zone andcerebral cortex. The presence of the subventricular zone in thesame slice allows neurogenic phenomena, which may occur afternigro-striatal damage, to be studied. Dopaminergic degenerationis induced by mechanical cutting of dopaminergic fibers from thesubstantia nigra to the striatum or by 6-OHDA, with relevance to PD.
2. Experimental methodology
2.1. Organotypic cultures
Organotypic cultures were prepared using a modification of themethod by Plenz and Kitai (1996). Sprague–Dawley rat pups (2–3days old) were decapitated and skulls opened along the commis-sural lines (Fig. 1A). Cultures were prepared also from different agebut slices from 2 to 3 days were selected as more homogeneousand viable (data not shown). Brains were separated in two hemi-spheres and the lateral third of each hemisphere, together withcerebellum, were removed (Fig. 1B). The resulting brain blocks werecut dorso/ventrally (350 �m thickness) with a vibratome (Pelco3000, CA, USA) at 45◦ inclination (Fig. 1C). Finally, slices containingcortex–corpus callosum–subventricular zone–striatum–substantianigra (approximately 6 slices/pup) were selected morphologicallyunder dissection microscope taking as a reference, the subventric-ular zone as depicted in Fig. 1D. Slices were cultivated with themethod of liquid–air interface on Millicell CM culture inserts (oneslice/insert from Millipore, Madrid, Spain) and maintained in Neu-robasal medium supplemented with 0.5% B27 supplement (both
from Gibco, Barcelona, Spain), 2 mM l-glutamine (Sigma, Madrid,Spain), 25% horse serum (Gibco) and 25 mg/ml gentamycin (Sigma,Madrid, Spain) at 37 ◦C, 5%CO2. Organotypic cultures were usedafter 7–12 days in vitro (depending on experimental procedures)in order to avoid apoptotic cell death in the inner layers due to lossof oxygen and nutrients and to allow proper cell maturation. Beforeeach experiment, slices considered to be damaged on the basis ofpropidium iodide (10 �M) incorporation were discarded and notfurther used.
All experiments were conducted under the supervision and withthe approval of our internal animal ethics committee (Neurotek-UPV/EHU), following European Communities Council Directive of24 November 1986 (86/609/EEC). Animals were handled in accor-dance with the European Communities Council Directive. Everypossible effort was made to minimize animal suffering and thenumber of animals used.
2.2. Cell and fiber labeling
The neuronal tracer Vybrant DiD (Molecular probes, Madrid,Spain) is a lipophilic dye that can be used to check cell and tissueviability during the different stages of slice culture (Table 1). DiD is acarbocyanine that can easily diffuse laterally within the membraneand serve as anterograde and retrograde tracer in nervous system(Honig and Hume, 1989). DiD was dissolved at 10 mg/ml in DMSOand 0.5 �l was injected into the substantia nigra with a Hamiltonsyringe. Dye incorporation into the substantia nigra and/or striatumwas visualized 3 days thereafter by using a Cy5 filter.
2.3. Immunofluorescence
Slices were fixed for 40 min in 4% paraformaldehyde andthen incubated for 30 min, at room temperature in 1% BSA inPBS containing 0.5% Tween (all from Sigma). Subsequently, sliceswere incubated overnight at 4 ◦C with antibodies to tyrosinehydroxylase (1:1000, Chemicon, Madrid, Spain), NeuN (a neuronalnucleus marker; 1:500, Chemicon),or �-synuclein (1:300, Novo-castra, Newcastle, UK), all in PBS containing 1% BSA. After furtherwashing, the cultures were incubated for one hour in a solution con-taining a mixture of the secondary antisera and nuclei were stainedwith Hoechst.
After final washing, the plates were coverslipped with anti-fading gel/mount (Biomeda, CA, USA). Immunofluorescence wasvisualized by a scanning Apotome system microscope (Zeiss).

Author's personal copy
F. Cavaliere et al. / Journal of Neuroscience Methods 188 (2010) 205–212 207
Fig. 1. Schematic view of culture preparation. (A) Lines indicate the location and direction of cut to open the skull and remove the brain for further dissection. b in the circleindicates the Bregma point. (B) The outer third of the lateral cortex and the entire cerebellum were removed (arrows). (C) Slices (350 �m) are cut at 45◦ in order to includethe substantia nigra (SN), striatum–caudate putamen (cp), subventricular zone of the lateral ventricle (lv) and cerebral cortex (cx). Diagonal lines indicate the cutting plane;the vertical line indicates the glued surface for vibratome-cut. (D and E) Microphotographs of slices cultivated for 10 days in vitro by the interface method, before (D) andafter (E) denervation of the nigro-striatal pathway (dashed line).
Fig. 2. Neuronal connections are maintained during slice cultivation. Slices at 1 DIV were labeled in the region of the substantia nigra with 0.5 �l of Vybrant DiD. At 7 daysin vitro, dopaminergic connections between the substantia nigra and the striatum were transected and DiD labeling was analyzed in the striatum of uncut (CTRL) slices (A)and lesioned slices (B) by using a Cy5 filter. Reduced fluorescence following transection is apparent.
Author's personal copy
208 F. Cavaliere et al. / Journal of Neuroscience Methods 188 (2010) 205–212
2.4. Cell proliferation
Slices were incubated for two hours with 20 �M bromod-eoxyuridine (BrdU, Sigma) which is incorporated into proliferatingcells. Immunofluorescent detection of BrdU was performed withan anti-BrdU antibody (Sigma) used at 1:200 concentration after30 min of denaturation with 3 N HCl at 37 ◦C for elimination of non-specific background (see Section 2.3 for more detailed information).
2.5. Statistical analysis
Values were normalized as specified in each figure legend.Statistical differences were evaluated by one-way analysis ofvariance (ANOVA), followed by a post hoc test (HSD Tukey).Fisher’s F was also calculated. All values reported, where nis the number of experiments, are significant, with at leastp < 0.05.
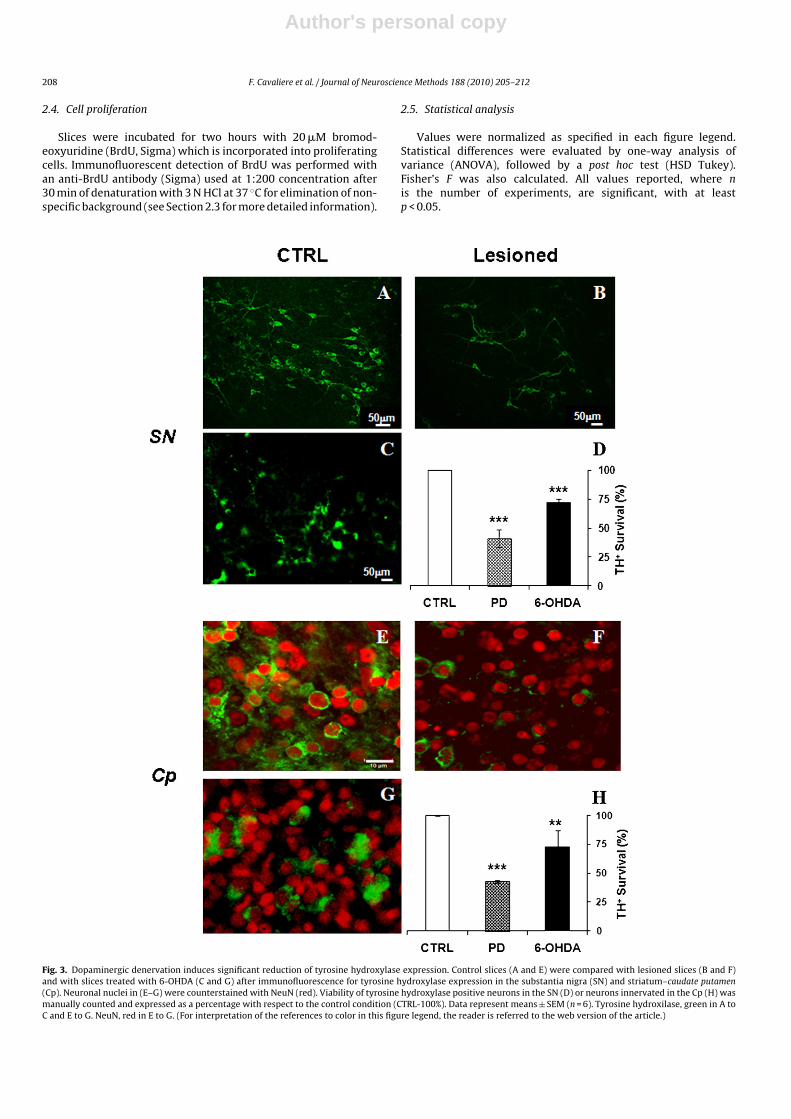
Fig. 3. Dopaminergic denervation induces significant reduction of tyrosine hydroxylase expression. Control slices (A and E) were compared with lesioned slices (B and F)and with slices treated with 6-OHDA (C and G) after immunofluorescence for tyrosine hydroxylase expression in the substantia nigra (SN) and striatum–caudate putamen(Cp). Neuronal nuclei in (E–G) were counterstained with NeuN (red). Viability of tyrosine hydroxylase positive neurons in the SN (D) or neurons innervated in the Cp (H) wasmanually counted and expressed as a percentage with respect to the control condition (CTRL-100%). Data represent means ± SEM (n = 6). Tyrosine hydroxilase, green in A toC and E to G. NeuN, red in E to G. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)

Author's personal copy
F. Cavaliere et al. / Journal of Neuroscience Methods 188 (2010) 205–212 209
3. Results
Organotypic slices were cultivated in such a way as to main-tain the dopaminergic (nigro-striatal pathway) and glutamatergicpathways (cortical–striatal) in the same slice. Thus, rat brains weresectioned on a vibratome to obtain 350 �m slice series with organ-otypic connections involving the substantia nigra, caudate striatum(caudate putamen), and part of the lateral ventricle, with thesubventricular zone connected to the cortex via the corpus callo-sum (Fig. 1C and D). Dopaminergic degeneration was successivelyinduced by mechanical cut of fibers from the substantia nigra to thestriatum (Fig. 1E and Table 1 for protocol description).
Dopaminergic connection between the substantia nigra andcaudate putamen in the striatum was demonstrated by labelingthe substantia nigra with 0.5 �l of the cellular tracer DiD (Table 1).Three days after labeling we transected the slices and three dayslater we analyzed slices under a fluorescence microscope. In thecontrol slices, we observed labeled neuronal terminals in the stria-tum contacting with striatal cell bodies (Fig. 2A). Anterogradelabeling in the striatum was reduced after transaction of dopamin-ergic fibers from the substantia nigra to the striatum (Fig. 2B).
In order to confirm that reduction of labeled neuronal ter-minals in the caudate putamen was due to degeneration ofthe nigro-striatal pathway affecting dopaminergic innervations,we examined by immunofluorescence the expression of tyrosinehydroxylase in the substantia nigra (Fig. 3A and B) and striatum(Fig. 3E and F). Slices at 7 DIV were damaged at the nigro-striatallevel by mechanical lesion and maintained in culture 3–5 daysmore, before immunofluorescence (Table 1). In parallel, we alsocompared the effect of axon transection with the more specificdamage induced by 6-OHDA (Fig. 3C and G) degeneration (see e.g.Kress and Reynolds, 2005). Cell viability in both models was mon-itored by propidium iodide incorporation after induction of celldamage (data not shown).
3.1. Substantia nigra
Cells in slices maintained for up to 12 days in vitro, withoutdopaminergic degeneration (Fig. 3A), express high levels of tyrosinehydroxylase. Tyrosine hydroxylase is specifically expressed in cellswith the typical morphology of dopaminergic neurons. Three tofive days after dopaminergic denervation, the number of dopamin-ergic neurons in the substantia nigra decreased by approximately
60% (F = 171.2, p = 0.005) as can be seen in Fig. 3B and D, whereastreatment of slices with 100 �M 6-OHDA (Fig. 3C and D) for 60 min(Kress and Reynolds, 2005) reduced the number of dopaminergicneurons by 30–35% (F = 240.1, p = 0.004).
3.2. Striatum (caudate putamen)
Denervation also affected tyrosine hydroxylase expression inthe caudate putamen. In control slices with no denervation (Fig. 3E),a moderate density of dopaminergic terminals surrounded striatalcells; this dramatically decreased after denervation of the nigro-striatal pathway (Fig. 3F and H), practically to the same extent asin the substantia nigra (57%; F = 1392, p = 0.0007). As in the sub-stantia nigra, degeneration caused by 6-OHDA treatment reduceddopaminergic innervation of the caudate putamen by only about30% (F = 10.33, p = 0.084), as described in Fig. 3G and H, calculatedas the number of striatal neurons which lose tyrosine hydroxylase-associated staining.
It is important to note that at this time point (i.e. three days afterdopaminergic denervation), neuronal viability of the remainingneurons, as indicated by NeuN (neuronal nuclei) immunofluores-cence (Fig. 3E–G), appears to be unaffected despite the reduction indopaminergic terminals. Therefore, this time window seems appro-priate to study some of the cellular and molecular consequences ofdopaminergic denervation.
Three days after dopaminergic denervation, the caudate puta-men presented symptoms of neuronal damage, similar to thoseobserved in Parkinson’s disease. Thus, as illustrated in Fig. 4, weobserved an increase of alpha-synuclein aggregation in the caudateputamen, in addition to a reduction in the number of dopaminergicneurons and degeneration of dopaminergic fibers (consistent withFig. 3A–H).
3.3. Subventricular zone
Due to the presence of the subventricular zone, the modelpresented here also represents a useful tool for the study of regen-eration induced by dopaminergic denervation. This region, afterBrdU staining, appears as a highly proliferative area (Fig. 5A).Degeneration of nigro-striatal fibers induced 3 days after section-ing increased the proliferative state in the subventricular zone byabout 80% (Fig. 5B); in contrast, in 6-OHDA treated slices, only a4–5% increase was apparent (Fig. 5C). Thus, the subventricular zone
Fig. 4. Denervation produces alpha synuclein aggregation in the striatum. Three days after lesion, the caudate putamen was analyzed by immunofluorescence for alpha-synuclein expression/aggregation. Nuclei were counterstained with Hoechst (blue). An increase in alpha-synuclein aggregation is apparent in this nucleus. (For interpretationof the references to color in this figure legend, the reader is referred to the web version of the article.)
Author's personal copy
210 F. Cavaliere et al. / Journal of Neuroscience Methods 188 (2010) 205–212
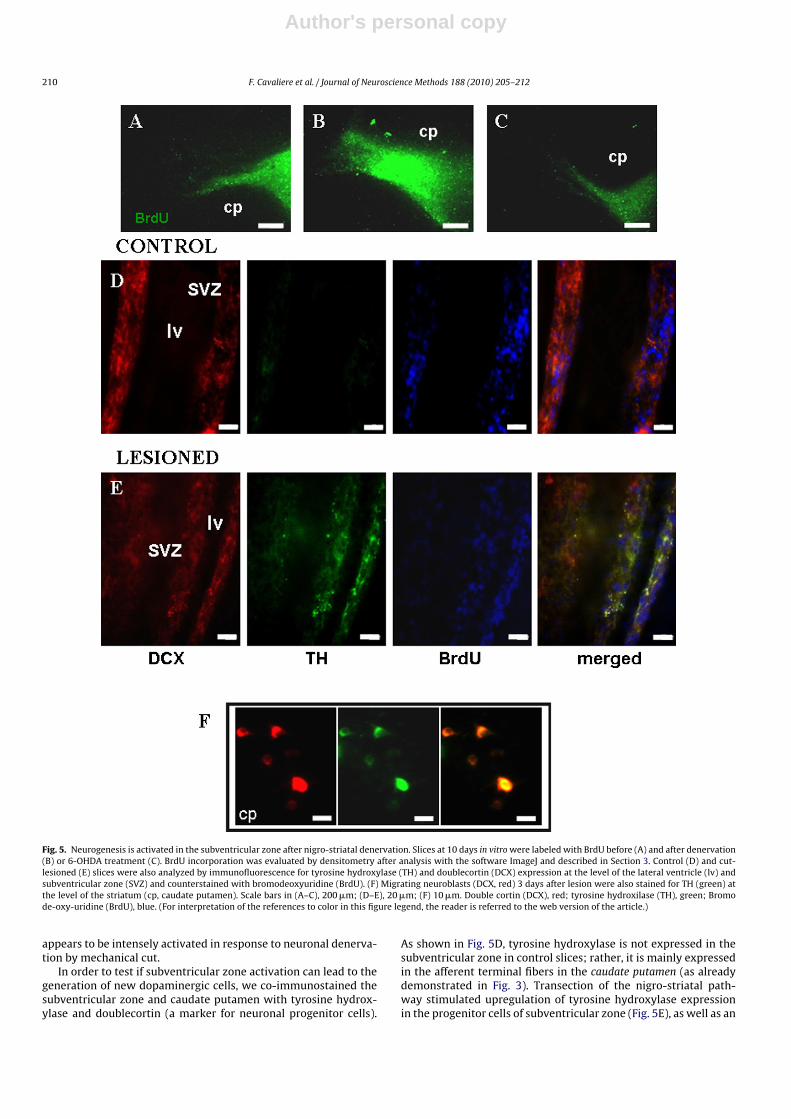
Fig. 5. Neurogenesis is activated in the subventricular zone after nigro-striatal denervation. Slices at 10 days in vitro were labeled with BrdU before (A) and after denervation(B) or 6-OHDA treatment (C). BrdU incorporation was evaluated by densitometry after analysis with the software ImageJ and described in Section 3. Control (D) and cut-lesioned (E) slices were also analyzed by immunofluorescence for tyrosine hydroxylase (TH) and doublecortin (DCX) expression at the level of the lateral ventricle (lv) andsubventricular zone (SVZ) and counterstained with bromodeoxyuridine (BrdU). (F) Migrating neuroblasts (DCX, red) 3 days after lesion were also stained for TH (green) atthe level of the striatum (cp, caudate putamen). Scale bars in (A–C), 200 �m; (D–E), 20 �m; (F) 10 �m. Double cortin (DCX), red; tyrosine hydroxilase (TH), green; Bromode-oxy-uridine (BrdU), blue. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
appears to be intensely activated in response to neuronal denerva-tion by mechanical cut.
In order to test if subventricular zone activation can lead to thegeneration of new dopaminergic cells, we co-immunostained thesubventricular zone and caudate putamen with tyrosine hydrox-ylase and doublecortin (a marker for neuronal progenitor cells).
As shown in Fig. 5D, tyrosine hydroxylase is not expressed in thesubventricular zone in control slices; rather, it is mainly expressedin the afferent terminal fibers in the caudate putamen (as alreadydemonstrated in Fig. 3). Transection of the nigro-striatal path-way stimulated upregulation of tyrosine hydroxylase expressionin the progenitor cells of subventricular zone (Fig. 5E), as well as an

Author's personal copy
F. Cavaliere et al. / Journal of Neuroscience Methods 188 (2010) 205–212 211
increase in cell proliferation as shown in Fig. 5B. In fact, we foundthat many DCX+ cells also expressed tyrosine hydroxylase, indi-cating that dopaminergic denervation stimulates precursor cellsin the subventricular zone to pre-differentiate into dopaminergicneuronal precursors. Finally, we suggest that degeneration of nigro-striatal fibers indeed stimulates neurogenesis as a few neuroblasts(DCX+) in the caudate putamen were found to express tyrosinehydroxylase (Fig. 5F).
4. Discussion
Here, we describe a novel organotypic culture system com-prising the cortex–striatum–substantia nigra–corpus callosum–subventricular zone in one single slice. For the first time, we report a3D model of cortical-basal structures in which many of the originalconnections between the cortex, striatum and substantia nigra arerepresented. In contrast to previously reported organotypic models(e.g. that of Plenz and Kitai, 1996), the glutamatergic pathway fromthe cortex to the striatum, the GABAergic projection from the stria-tum to the substantia nigra and the dopaminergic projection fromthe substantia nigra to the striatum are all maintained intact. More-over, the presence of the subventricular zone in the vicinity of thestriatum permits a study of neurogenesis. Thus, the model hereinreported is a versatile tool to study mechanisms of neurodegen-erative diseases which involve cortico-striatal-basal degeneration,as well as neuroregeneration. In particular, we adapted the modelto study Parkinson’s disease, since it is possible to reproduce thecharacteristics of PD due to the anatomy and original connectionsof the slice. More generally, our model can be also be used to modelother degenerative diseases caused by cortico-striatal-basal degen-eration, such as cortico-basal degeneration (Riley et al., 1990), Lewybody dementia (Hanson and Lippa, 2009), and even schizophrenia(Arakawa et al., 2009).
This organotypic model involves dopaminergic degenerationdue to mechanical cutting of tissue between the striatum and sub-stantia nigra which leads to retrograde and anterograde neuronaldegeneration in the substantia nigra and in the striatum respec-tively (as demonstrated by reduction in the number of tyrosinehydroxylase positive neurons, Fig. 3). Nevertheless, the model isquite versatile and can emulate PD quite well with respect to themore canonical methods, such as 6-OHDA treatment. In this paper,we compared the two methods and found that damage to dopamin-ergic neurons due to 6-OHDA is less intense than that due todissection (50–60% less). This difference is likely due to the massivedamage induced after transection which also affects glutamatergicand GABAergic systems. In any event, transection leads to degen-eration, but the organotypic slices retain viability, as demonstratedby NeuN immunofluorescence and Hoechst staining (Figs. 3 and 4).
Both researchers and clinicians agree that multiple, alter-native, therapeutic strategies are necessary to treat complexdiseases which have different etiologies and underlying mecha-nisms (Björklund and Kirik, 2009). The tissue model presentedherein may well facilitate the development of novel therapeutictools, since it is highly versatile and recapitulates well the featuresof dopaminergic degeneration. In fact, organotypic slices can beused to model dopaminergic degeneration after mechanical lesionor even by drug administration (6-OHDA). Nevertheless, we do notto exclude that, differently from mechanical lesion, drugs used toinduce neuronal damage can be buffered or even cleared by glialcells, including oligodendrocytes and astrocytes.
As stated previously, the subventricular zone is also representedin these organotypic slices, thus making the model relevant for thestudy of neurogenesis in neurodegenerative disease Arias-Carriónet al. (2007). In particular, the generation of new dopaminergic neu-rons under conditions of striatal degeneration, by stimulation of
endogenous neurogenesis or by cell grafts, still represents a com-pelling alternative to PD therapy which at the moment only usespalliative treatments. Our results suggest that striatal deafferenta-tion in vivo induces dopaminergic neurogenesis in the striatum,as well as in the subventricular zone, in corroboration with otherreports. Thus, it has been demonstrated that degeneration ofdopaminergic fibers is associated with the increased generation ofdopaminergic neurons in the olfactory bulb (Winner et al., 2006),which is the natural site of neurogenesis activated in the subven-tricular zone. Moreover, Arias-Carrión et al. (2004, 2006) showedthat 6-OHDA lesion in vivo generated functional tyrosine hydrox-ylase positive neuroblasts in the subventricular zone of lesionedanimals. In these studies, dopaminergic neurogenesis was effectiveonly if animals were treated in parallel with transcranial magneticfield stimulation or chromaffin cell transplantation.
Our results corroborate the general hypothesis that neurode-generative insult (such as deafferentation, 6-OHDA treatment, aswell as ischemia or trauma) triggers neurogenesis in the ependy-mal cell layer of the subventricular zone where cells are normallyquiescent. Cell division can be activated by a combination of injuryand growth factor stimulation (Gleason et al., 2008). Alternatively,division may be inhibited by a negative environment, such as thatassociated with ischemia, (Vergni et al., 2009). On the basis of theresults obtained with the novel organotypic model which we havedescribed here, it seems reasonable to think that dopamine deple-tion predisposes neural stem cells of the subventricular zone todifferentiate into dopaminergic cells (Fig. 5). However, neuronalregeneration is not sustained because of the absence of appropri-ate growth factor stimulation, and/or the presence of unfavorableconditions which inhibit regeneration.
In this paper, besides of technical advances in modeling nigro-striatal degeneration in tissue cultures, we want to highlight theaptitude of the subventricular zone to activate specific neuro-genesis following severe dopamine depletion with generation ofmultipotent tyrosine hydroxilase positive cells. Pharmacologicalmodulation of tyrosine hydroxilase positive neurons generationmay represent a promised perspective for studies of striatal neu-roregeneration.
Acknowledgments
This work was supported by CIBERNED (Ministerio de Sanidady Consumo) and University of Basque Country.
References
Arakawa R, Ichimiya T, Ito H, Takano A, Okumura M, Takahashi H, et al.Increase in thalamic binding of [(11)C]PE2I in patients with schizophrenia: Apositron emission tomography study of dopamine transporter. J Psychiatr Res2009;43(15):1219–23.
Arias-Carrión O, Verdugo-Díaz L, Feria-Velasco A, Millán-Aldaco D, Gutiérrez AA,Hernández-Cruz A, et al. Neurogenesis in the subventricular zone followingtranscranial magnetic field stimulation and nigrostriatal lesions. J Neurosci Res2004;78:16–28.
Arias-Carrión O, Hernández-López S, Ibanez-Sandoval O, Bargas J, Hernández-CruzA, Drucker-Colín R. Neuronal precursors within the adult rat subventricularzone differentiate into dopaminergic neurons after substantia nigra lesion andchromaffin cell transplant. J Neurosci Res 2006;84:1425–37.
Arias-Carrión O, Freundlieb N, Oertel WH, Höglinger GU. Adult neurogenesis andParkinson’s disease CNS. Neurol Disord Drug Targets 2007;6:326–35.
Björklund T, Kirik D. Scientific rationale for the development of gene therapy strate-gies for Parkinson’s disease. Biochim Biophys Acta 2009;1792:703–13.
Cavaliere F, Dinkel K, Reymann K. The subventricular zone releases factors whichcan be protective in oxygen/glucose deprivation-induced cortical damage: anorganotypic study. Exp Neurol 2006;201:66–74.
Gleason D, Fallon JH, Guerra M, Liu JC, Bryant PJ. Ependymal stem cells divide asym-metrically and transfer progeny into the subventricular zone when activated byinjury. Neuroscience 2008;156:81–8.
Gogolla N, Galimberti I, DePaola V, Caroni P. Preparation of organotypic hippocampalslice cultures for long-term live imaging. Nat Protocols 2006;1:1165–71.
Halliday GM, McCann H. Human-based studies on alpha-synuclein deposition andrelationship to Parkinson’s disease symptoms. Exp Neurol 2008;209:12–21.

Author's personal copy
212 F. Cavaliere et al. / Journal of Neuroscience Methods 188 (2010) 205–212
Hanson JC, Lippa CF. Lewy body dementia. Int Rev Neurobiol 2009;84:215–28.Honig MG, Hume RI. Dil and diO: versatile fluorescent dyes for neuronal labelling
and pathway tracing. Trends Neurosci 1989;12:333–5.Jackson-Lewis V, Przedborski S. Protocol for the MPTP mouse model of Parkinson’s
disease. Nat Protocols 2007;2:141–51.Kress GJ, Reynolds IJ. Dopaminergic neurotoxins require excitotoxic stimulation in
organotypic cultures 2005;20:639–45.Lindvall O, Kokaia Z. Prospects of stem cell therapy for replacing dopamine neurons
in Parkinson’s disease. Trends Pharmacol Sci 2009;30:260–7.Noraberg J, Kristensen BW, Zimmer J. Markers for neuronal degeneration in organ-
otypic slice cultures. Brain Res Brain Res Protoc 1999;3:278–90.Plenz D, Kitai ST. Organotypic cortex–striatum–mesencephalon cultures: the nigros-
triatal pathway. Neurosci Lett 1996;209:177–80.
Riley DE, Lang AE, Lewis A, Resch L, Ashby P, Hornykiewicz O, et al. Cortical-basalganglionic degeneration. Neurology 1990;40:1203–12.
Shimohama S, Sawada H, Kitamura Y, Taniguchi T. Disease model: Parkinson’s dis-ease. Trends Mol Med 2003;9:360–5.
Vergni D, Castiglione F, Briani M, Middei S, Alberdi E, Reymann KG, et al. A model ofischemia-induced neuroblast activation in the adult subventricular zone. PLoSOne 2009;4:e5278.
Winner B, Desplats P, Hagl C, Klucken J, Aigner R, Ploetz S, et al. Dopamine receptoractivation promotes adult neurogenesis in an acute Parkinson model. Exp Neurol2009;219:543–52.
Winner B, Geyer M, Couillard-Despres S, Aigner R, Bogdahn U, Aigner L, et al. Stri-atal deafferentation increases dopaminergic neurogenesis in the adult olfactorybulb. Exp Neurol 2006;197:113–21.




















