
Fate Mapping and Lineage Analyses Demonstrate the Production of a Large Number of Striatal...
26
Fate Mapping and Lineage Analyses Demonstrate the Production of a Large Number of Striatal Neuroblasts after TGFα and Noggin Striatal Infusions into the Dopamine-depleted Striatum Antoine de Chevigny 1 , Oliver Cooper 1 , Angel Vinuela 1 , Casper Reske-Nielsen 1 , Diane C. Lagace 2 , Amelia J. Eisch 2 , and Ole Isacson 1 1Udall Parkinson Disease Research Center of Excellence, Center for Neuroregeneration Research. McLean Hospital/Harvard Medical School, Belmont, MA02478. 2Department of Psychiatry, University of Texas Southwestern Medical Center, Dallas, TX Abstract Infusion of TGFα into the adult dopamine (DA)-depleted striatum generates a local population of nestin + /PCNA + newborn cells [1]. The precise origin and fate of these new striatal cells are unknown, making it difficult to direct them for neural repair in Parkinson’s disease (PD). Experiments in rats using BrdU to label neural progenitor cells (NPCs) showed that during TGFα infusion in the DA- depleted striatum, newborn striatal cells formed a homogenous population of precursors, with the majority coexpressing nestin, Mash1, Olig2 and EGFR, consistent with the phenotype of multipotent C cells. Upon TGFα pump withdrawal, the subventricular zone (SVZ) was repopulated by neuroblasts. Strikingly, during this period, numerous clusters of DCX + /PSANCAM + neuroblasts were also produced in the ipsilateral medial striatum. In parallel, striatal BrdU + /GFAP + astrocytes were generated, but no BrdU + /O4 + /CNPase + oligodendrocytes. Infusion of the neuralizing BMP antagonist noggin after TGFα pump withdrawal increased the neuroblast to astrocyte ratio among new striatal cells by blocking glial differentiation, but did not alter striatal neurogenesis. At no time or no treatment condition were differentiated neurons generated, including DA neurons. Using 6- OHDA lesioned nestin-CreER T2 /R26R-YFP mice that allow genetic fate-mapping of SVZ nestin + cells, we show that TGFα-generated striatal cells originate from SVZ nestin + precursors that confirmed data from the rats on the phenotype and fate of striatal nestin + /PCNA + cells upon TGFα withdrawal. This work demonstrates that a large population of multipotent striatal C-like cells can be generated in the DA-depleted striatum that do not spontaneously differentiate into DA neurons. Keywords Parkinson’s disease; adult neurogenesis; subventricular zone; striatum; TGFα; neuroblasts Corresponding author: Professor Ole Isacson, Neuroregeneration Laboratories, Harvard Medical School, McLean Hospital, MRC I, 115 Mill Street, Belmont, MA 02478, Tel 617-855-3283, Fax 617-855-2522, [email protected]. Author contributions: A. de Chevigny: Design, provision of study material, collection and/or assembly of data, data analysis and interpretation, manuscript writing; O.C.: Design, provision of study material , collection and/or assembly of data, data analysis and interpretation, manuscript writing; A.V.: Design, provision of study material, collection and/or assembly of data, data analysis and interpretation, manuscript writing; C.R.: Collection and/or assembly of data, data analysis and interpretation, manuscript writing; D.L.: Provision of study material, data analysis and interpretation, manuscript writing; A.E.: Provision of study material, data analysis and interpretation, manuscript writing; O.I.: Conception and design, principal investigator and grant support, provision of study material, collection and/or assembly of data, data analysis and interpretation, manuscript writing, final approval of manuscript. NIH Public Access Author Manuscript Stem Cells. Author manuscript; available in PMC 2009 September 1. Published in final edited form as: Stem Cells. 2008 September ; 26(9): 2349–2360. doi:10.1634/stemcells.2008-0080. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Fate Mapping and Lineage Analyses Demonstrate the Production of a Large Number of Striatal...

Fate Mapping and Lineage Analyses Demonstrate the Productionof a Large Number of Striatal Neuroblasts after TGFα and NogginStriatal Infusions into the Dopamine-depleted Striatum
Antoine de Chevigny1, Oliver Cooper1, Angel Vinuela1, Casper Reske-Nielsen1, Diane C.Lagace2, Amelia J. Eisch2, and Ole Isacson1
1Udall Parkinson Disease Research Center of Excellence, Center for Neuroregeneration Research. McLeanHospital/Harvard Medical School, Belmont, MA02478.
2Department of Psychiatry, University of Texas Southwestern Medical Center, Dallas, TX
AbstractInfusion of TGFα into the adult dopamine (DA)-depleted striatum generates a local population ofnestin+/PCNA+ newborn cells [1]. The precise origin and fate of these new striatal cells are unknown,making it difficult to direct them for neural repair in Parkinson’s disease (PD). Experiments in ratsusing BrdU to label neural progenitor cells (NPCs) showed that during TGFα infusion in the DA-depleted striatum, newborn striatal cells formed a homogenous population of precursors, with themajority coexpressing nestin, Mash1, Olig2 and EGFR, consistent with the phenotype of multipotentC cells. Upon TGFα pump withdrawal, the subventricular zone (SVZ) was repopulated byneuroblasts. Strikingly, during this period, numerous clusters of DCX+/PSANCAM+ neuroblastswere also produced in the ipsilateral medial striatum. In parallel, striatal BrdU+/GFAP+ astrocyteswere generated, but no BrdU+/O4+/CNPase+ oligodendrocytes. Infusion of the neuralizing BMPantagonist noggin after TGFα pump withdrawal increased the neuroblast to astrocyte ratio amongnew striatal cells by blocking glial differentiation, but did not alter striatal neurogenesis. At no timeor no treatment condition were differentiated neurons generated, including DA neurons. Using 6-OHDA lesioned nestin-CreERT2/R26R-YFP mice that allow genetic fate-mapping of SVZ nestin+
cells, we show that TGFα-generated striatal cells originate from SVZ nestin+ precursors thatconfirmed data from the rats on the phenotype and fate of striatal nestin+/PCNA+ cells upon TGFαwithdrawal. This work demonstrates that a large population of multipotent striatal C-like cells canbe generated in the DA-depleted striatum that do not spontaneously differentiate into DA neurons.
KeywordsParkinson’s disease; adult neurogenesis; subventricular zone; striatum; TGFα; neuroblasts
Corresponding author: Professor Ole Isacson, Neuroregeneration Laboratories, Harvard Medical School, McLean Hospital, MRC I, 115Mill Street, Belmont, MA 02478, Tel 617-855-3283, Fax 617-855-2522, [email protected] contributions: A. de Chevigny: Design, provision of study material, collection and/or assembly of data, data analysis andinterpretation, manuscript writing; O.C.: Design, provision of study material , collection and/or assembly of data, data analysis andinterpretation, manuscript writing; A.V.: Design, provision of study material, collection and/or assembly of data, data analysis andinterpretation, manuscript writing; C.R.: Collection and/or assembly of data, data analysis and interpretation, manuscript writing; D.L.:Provision of study material, data analysis and interpretation, manuscript writing; A.E.: Provision of study material, data analysis andinterpretation, manuscript writing; O.I.: Conception and design, principal investigator and grant support, provision of study material,collection and/or assembly of data, data analysis and interpretation, manuscript writing, final approval of manuscript.
NIH Public AccessAuthor ManuscriptStem Cells. Author manuscript; available in PMC 2009 September 1.
Published in final edited form as:Stem Cells. 2008 September ; 26(9): 2349–2360. doi:10.1634/stemcells.2008-0080.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

INTRODUCTIONIn the context of Parkinson’s disease (PD), subventricular zone (SVZ) neural stem andprecursor cells [herein collectively called neural progenitor cells (NPCs)] do not contribute todopamine (DA) cell replacement [2]. The SVZ is the largest neurogenic zone in the brain andis adjacent to the striatum, the region where most DA neurotransmitter depletion occurs in PD.Given that some SVZ cells continuously differentiate into olfactory bulb (OB) DA neuronsthroughout life, their artificial recruitment into the striatum and differentiation into DA neuronsis an appealing alternative to transplantation therapies [1,3,4].
The cytoarchitecture, cellular composition and lineage relationships between different NPCsubtypes in the mammalian SVZ have been extensively studied [5–8]. There are 3 well-characterized NPC subtypes in the adult SVZ. The primary precursors which are thought to beneural stem cells (B cells) have immunocytochemical and ultrastructural characteristics ofastrocytes [7]. These SVZ astrocytes give rise to transit-amplifying precursor cells (C cells),which have a globular morphology and proliferate rapidly, thus increasing the size of the SVZprecursor pool. The C cells generate neuroblasts (A cells) that express the polysialylatedneuronal cell adhesion molecule (PSANCAM) and doublecortin (DCX) [7]. Those neuroblastsare committed to the neuronal lineage and migrate towards the OB, where they ultimatelydifferentiate into interneurons, which use GABA or both GABA and DA as neurotransmitters.The latter interneurons constitute the A16 population of DA neurons that share functionalcharacteristics with A9 DA neurons [9,10]. Finally, in addition to OB neurons, some SVZ Bcells generate oligodendrocytes, although the precise lineage leading to this fate is unclear[11].
DA secreted by midbrain afferent projections promotes NPC proliferation in the SVZ bybinding directly to D2-like receptors on C cells [2]. Probably as a direct consequence of this,animal models of PD [2,12] and PD patients [2] display reduced SVZ cell proliferation andfewer newborn OB neurons [2]. Surprisingly, SVZ Pax6+ DA neuron precursors and olfactorybulb DA neurons are increased in PD animal models [13–15]. This provides the intriguingpossibility of using chemo-attractants or growth factors to artificially recruit endogenous DAneuron precursors to the DA-depleted striatum.
In the 6-hydroxydopamine (6-OHDA) rodent model of PD, striatal TGFα infusion stimulatesforebrain cell division and induces septal and striatal waves of nestin+/PCNA+ cells orientedtowards the infusion cannula [1,16]. Here we infused the EGFR agonist TGFα into the DA-depleted striatum [1,16], and addressed the following, new questions: (a) what are the originand phenotype(s) of TGFα-activated cells in the striatal wave? (b) What is their fate 2 weeksafter TGFα withdrawal? (c) What is the effect of TGFα and noggin consecutive infusions,given the known neurogenic influence of noggin on SVZ cells [17]?
MATERIALS AND METHODSRats
Adult 250–300 g female Sprague Dawley rats (Taconic Farms, Germantown, NY) administered6-OHDA in the medial forebrain bundle [18] (n=88, 8 per group for all groups except n=4 forTGFα̣ weeks / PBS 4 weeks). The animals were maintained in accordance with current NationalInstitutes of Health guidelines and McLean Hospital/Harvard University Institutional AnimalCare and Use Committee protocols.
TGFα and Noggin infusions in ratsThe infusion procedures for TGFα and noggin were performed as described [1]. Osmoticpumps (Alzet, models 1002 and 2004; Cupertino, CA) delivered either: (a) 0.5 mg/ml TGFα
de Chevigny et al. Page 2
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(R&D Systems, Minneapolis, MN), (b) 6 µg/ml noggin (R&D Systems), (c) 0.5 mg/mlTGFα̣ and 6 µg/ml noggin, or (d) PBS into the right striatum (+1.2 A/P; +2.7 M/L) via a cannula(Brain Infusion Kit II, Alzet) at 0.25µl h−1. Proteins were dissolved in 0.1 M PBS.
For sequential infusions, rats were anesthetized and the osmotic pump was replaced, while thecannula, polyethylene tubing and flow moderator remained in place.
Nestin-CreERT2/R26R-YFP mice experiments5–8 week-old nestin-CreERT2/R26R-YFP mice [19] were anesthetized with ketamine/xylazinesolution and 6-OHDA (5.0 mg/ml in 0.9% NaCl/0.02% ascorbate) was injected by microlitersyringe (Hamilton) at 0.5 µl/min by pump for a total dose of 15.0 mg/3 µl. The right striatumwas targeted at stereotaxic coordinates AP: +0.9 mm; ML: +2.2 mm; DV: −2.5 mm relative tobregma [20]. Two weeks following 6-OHDA injection, mice were administered tamoxifen at180 mg/kg/day for 5 days (intraperitoneally (i.p.); dissolved in 10% EtOH/90% sunflower oil).The first group of mice was sacrificed 2 days after the tamoxifen regimen (Fig. 1C1). For the2 other groups, 1 week following the tamoxifen regimen, osmotic pumps delivered either 0.5mg/ml TGFα or 0.9% saline into the right striatum (AP: +1 mm; ML: +1.3 mm; DV: −3 mmrelative to bregma). Mice were sacrificed after 2 weeks of infusion (Fig. 1C2) or 2 weeks afterpump withdrawal (Fig.1C3).
BrdU administrationRats were administered BrdU (Sigma, St. Louis, MO) i.p. once per day from days 3 to 14 ofthe first TGFα/PBS infusion at a dose of 50 mg/kg body weight (Fig. 1B). We waited 3 daysafter pump implantation before starting BrdU injections in order to avoid BrdU incorporationby reactive astroglia around the cannula site.
Histological proceduresAnimals were terminally anesthetized with sodium pentobarbital (100 mg/kg, i.p.) and perfusedwith intracardial heparin saline (0.1% heparin in 0.9% saline; 100 ml/rat) andparaformaldehyde (4% in PBS; 200 ml/rat). The brains were post-fixed and sectioned (40 µm)as described [18].
ImmunohistochemistryRoutine indirect immunofluorescence was performed on randomly selected series of sectionsthat represented 1/12th of the total brain [18] (see Supplemental Methods for primary antibodydetails). For BrdU immunostaining, the sections were pretreated with 0.2% Triton X-100 for2 h and denatured with 2 N HCl for 40 min at 37 °C. Primary antibodies were kept overnightat room temperature, except for EGFR, Mash1 and Dlx2 antibodies that were kept 48–72 hoursat 4 °C. For double and triple labeling with BrdU, cell markers were detected before BrdUimmunostaining. Primary antibodies were visualized using fluorescent secondary antibodies(AlexaFluor, Molecular Probes). For EGFR, Mash1 and Dlx2 immunostainings, sections weretreated for 2 h at room temperature with a biotinylated secondary antibody (1:300, JacksonLaboratories) and then visualized using the tyramide signal amplification kit (TSA PlusSystem, PerkinElmer Life Science, Waltham, MA).
TUNEL detected apoptosis-related DNA fragmentation using the manufacturer’s protocol(Neurotacs II, R&D Systems).
Cell CountsCells were counted blind to experimental conditions. For striatal counts, a 100–300 µmdiameter circle with its center at the infusion site was excluded in order to avoid contamination
de Chevigny et al. Page 3
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

of analyses by reactive astroglia. Sections were analyzed using either a standard microscope(BX51, Olympus or Axioskop 2+, Carl Zeiss) or a confocal microscope (LSM 510/Meta, CarlZeiss, Thornwood, NY). Cell counts and distances were measured using a stereologyworkstation (Stereoinvestigator, Microbrightfield, Williston, VT) with an integratedepifluorescence microscope (Axioskop 2+, Carl Zeiss). BrdU+ cells colabeled with phenotypicmarkers were identified from orthogonal projections to confirm colocalization. For eachmarker and per animal, we analyzed 130 to 280 BrdU+ cells per section on a total of 4–6 striatalsections spaced by 480 µm (once every 12th section). For nestin expression, we identified highexpression when signal was detectable using the following parameters on the confocalmicroscope: pinhole: 85 µm, Detector gain: 600, Amplifier Offset: −0.02 V. We considerednestin expression to be low when emission fluorescence was not detected using the aboveparameters but was detected using the following parameters: pinhole: 89 µm, Detector gain:814, Amplifier Offset: −0.02 V.
Behavioral testingUnilaterally lesioned rats were tested for rotational behavior in response to amphetamine (4mg/kg i.p.) before infusion and 3 days prior to sacrifice. Animals were randomized and placed intoautomated rotometer bowls and left and right full-body turns were monitored by a computerizedactivity monitor system as described [18].
Statistical AnalysisData are presented as mean ± SEM. One way ANOVA and Student’s unpaired t test wereemployed to assess differences between data groups using the software (SAS Institute, Cary,NC). Differences were considered statistically significant when p<0.05.
RESULTSPhenotypic characterization of newly generated cells in the DA-depleted striatum duringTGFα infusion
To determine the precise phenotype of the striatal nestin+/PCNA+ cells in 6-OHDA lesionedrats [1,16], we investigated NPC marker expression by BrdU+ cells in the ipsilateral striatumafter 2 weeks of TGFα infusion (Fig. 1B1). BrdU was injected throughout the infusion periodto label cells that were in S-phase (Fig. 1B). The majority (> 89 %) of BrdU+ cells in theTGFα-induced ipsilateral striatal and septal waves coexpressed nestin, EGFR and Olig2,indicative of a “C-like” phenotype [13,21] (Suppl. Fig. 1 and Fig. 2A–D, P). We also foundthat 47 ± 8 % of the BrdU+ cells expressed Mash1 (Fig. 2E, P), another marker of an SVZ Ccell subpopulation [22]. Newborn cells in the TGFα-activated contralateral, septal wave [1]and proliferative nodules also displayed this “C-like cell” phenotype (Suppl. Fig. 1). However,BrdU+ “C-like” cells in the waves differed from normal SVZ C cells in that they did not expressDlx2 (data not shown) [21] and had an elongated morphology (Fig. 2C). Almost no BrdU+ cellin the striatum expressed the mature neural cell markers GFAP, Tuj1, NeuN, CNPase or Iba1(Fig. 2I–L and P), confirming that new cells did not differentiate during TGFα infusion. Ofnote, TGFα infusion increased the number of Iba1+ microglial cells near where BrdU+ C-likecells were proliferating (both SVZ and striatal wave) compared to PBS infusion (Fig. 2M–O),although few BrdU+ cells expressed Iba1 (4 ± 3 %) and few Iba1+ cells expressed BrdU (2 ±3 %, Fig. 2K, L and P).
Distribution and phenotypes of SVZ-derived cells after TGFα withdrawalNext, we examined the response of the C-like cells in the ipsilateral striatal wave after TGFαwithdrawal. Unilaterally 6-OHDA lesioned rats received TGFα for 2 weeks followed by a 2week PBS infusion (Fig. 1B2). In these rats, the number of striatal PCNA+ dividing cells had
de Chevigny et al. Page 4
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

markedly decreased compared to during TGFα infusion (Fig. 3A–C), indicating that striatalmitotic activity was reduced. The number and distribution of BrdU+ cells in the ipsilateralstriatum during, or after TGFα infusion were also determined (Fig. 3D–G). During TGFαinfusion, BrdU+ cells were found in septal and striatal cellular waves (Fig. 3E). After TGFαwithdrawal, fewer BrdU+ cells remained in the striatal wave (Fig. 3F) (40,767 ± 814 BrdU+
cells vs 28,109 ± 2622 BrdU+ cells for TGFα and TGFα/PBS infusion conditions, respectively;34 % reduction, unpaired t test, p<0.05; Fig. 3G). This reduced number of newborn striatalcells was due, at least in part, to apoptotic mechanisms because more TUNEL+ profiles wereobserved in the ipsilateral SVZ and striatum after TGFα withdrawal than in rats infused withPBS (Fig. 3H–K; p<0.05). Hypercellular nodules seen during TGFα infusion were virtuallyabsent 2 weeks after TGFα withdrawal (Fig. 3E, F), and their remnants were undergoingapoptotic cell death (Fig. 3J). Importantly, the number of striatal BrdU+ cells was similar at 2and 4 weeks post-TGFα (Fig. 1B4 and 3G; 26,684 ± 1,534 BrdU+ cells vs 27,003 ± 2,340BrdU+ cells at 2 weeks and 4 weeks post- TGFα withdrawal, respectively), indicating that celldeath was negligible from 2 weeks after TGFα withdrawal.
Next we investigated the phenotype(s) of the remaining ~ 28,000 BrdU+ striatal wave cells perrat (Fig. 4A), 2 weeks after TGFα pump withdrawal. The majority of these BrdU+ cells (> 92%,Fig. 4E) did not express Olig2, Mash1 or EGFR (Fig. 4B and data not shown), indicating thatthe C-like cells had differentiated. However, 79 ± 8 % of BrdU+ striatal cells still expressednestin, although to a lower level than during TGFα infusion (compare Fig. 4B, C to Fig. 2B,C and see Materials and Methods). These data demonstrate that TGFα withdrawal did not resultin full differentiation of many SVZ-derived striatal cells by 2 weeks. Importantly however, wefound that 35 ± 4% of striatal BrdU+ cells were GFAP+ astrocytes (Fig. 4D, E), 70 ± 10 % ofwhich co-expressed low levels of nestin (data not shown). These BrdU+/GFAP+ astrocytes didnot express SSEA-1 or PCNA (data not shown), suggesting that they were post-mitotic cellsand not B-like cells. Importantly, 540 ± 12 BrdU+ cells per DA-depleted striatum co-expressedDCX, PSANCAM and nestin (demonstrated using staining on adjacent sections), whichrepresented ~ 2 % of the striatal BrdU+ population (Fig. 4F, G). In contrast, the number ofectopic striatal DCX+ neuroblasts was approximately 15 times larger (7,606 ± 2,131 cells peripsilateral striatum, Fig. 4H, M). Striatal DCX+/PSANCAM+ cells were proliferating afterTGFα withdrawal because 82 ± 6 % of them expressed the endogenous proliferation markerPCNA (Fig. 4I). Therefore, absence of BrdU labeling in ectopic striatal neuroblasts wasprobably due to BrdU dilution following a large number of divisions during the 2 weeks PBSinfusion period (i.e. “washing period”) post TGFα withdrawal. Seventy-five percent of suchDCX+ cells expressed Tuj1 (Fig. 4J), confirming a neuroblast phenotype. However, none ofthe DCX+ or PSANCAM+ cells in the striatum expressed Pax6, while SVZ neuroblasts werePax6+ in the same tissue section (data not shown). Interestingly, most ectopic striatalneuroblasts formed clusters that were in close contact with GFAP+ processes of astrocytes thathad their end feet on blood vessels (Fig. 4K, L). Furthermore, no BrdU+ cells were CNPase+
or O4+ oligodendrocytes, and only few were Iba1+ microglial cells at this time (1.5 ± 2 %, Fig.4E). Finally, we found similar phenotypes of BrdU+ cells at 4 weeks post-TGFα-withdrawal(data not shown). Strikingly however, the number of PSANCAM+ neuroblasts was increasedat this later time point (Fig. 4M, Suppl. Fig. 2; 7,607 ± 2,031 PSANCAM+ cells vs 13,827 ±1980 PSANCAM+ cells at 2 weeks and 4 weeks post-TGFα withdrawal, respectively), furtherdemonstrating the mitotic activity of striatal neuroblasts.
Distribution and phenotypes of SVZ-derived cells after TGFα followed by noggin infusionsIn an attempt to block astroglial differentiation and to enhance neuronal differentiation ofTGFα-stimulated cells in the DA-depleted striatum, we used the BMP antagonist noggin[23]. Noggin was first infused alone in the rat DA-depleted striatum for 4 weeks, in order toverify previous data indicating that noggin was pro-neuronal for SVZ NPCs in vivo [17].
de Chevigny et al. Page 5
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Interestingly, noggin expanded the PSANCAM+ neuroblast population in the ipsilateral SVZ,when compared to PBS infusion (358 ± 33 cells versus 238 ± 15 cells per SVZ section fornoggin infused and PBS infused rats, respectively; unpaired t test, p<0.01; Fig. 5A, B).Importantly, noggin infusion did not alter the number of nestin+/PCNA+ NPCs in the ipsilateralSVZ or striatum, relative to PBS administration alone (Fig. 5C, D), indicating that proliferation,migration and survival of NPCs were unaltered. These data demonstrate diffusion over longdistances and the induction of a pro-neuronal effect on SVZ cells by noggin.
To test the effect of noggin on TGFα-activated BrdU+ cells recruited in the striatum, rats wereadministered TGFα for 2 weeks, followed by 2-weeks of noggin infusion (Fig. 1B3). The totalnumber of striatal BrdU+ cells was unchanged compared to rats with TGFα/PBS sequentialinfusions (compare Fig. 5E to Fig. 4A; 28,109 ± 2622 BrdU+ cells vs 26,109 ± 2980 BrdU+
cells for TGFα/PBS and TGFα/noggin infusion conditions, respectively), confirming thatnoggin did not alter proliferation, survival or migration. This TGFα/noggin sequential infusionparadigm virtually blocked the generation of BrdU+/GFAP+ astrocytes in the DA-depletedstriatum (Fig. 5F, H–I; 3 ± 1 % versus 35 ± 4 % of astrocytes among new cells, for TGFα/noggin sequential infusion and TGFα/PBS sequential infusion, respectively; unpaired t test,p < 0.01). In contrast, neuronal commitment, as measured by DCX expression, was unchangedcompared to TGFα/PBS sequentially infused rats (Fig. 5H, J; 7,606 ± 2,131 versus 8,345 ±2,512 DCX+ cells for TGFα/PBS and TGFα/noggin infused rats, respectively). The percentageof immature BrdU+ cells (nestin+) tended to increase when compared to TGFα/PBSsequentially infused rats (Fig. 5G, H; p = 0.079), suggesting that BrdU+ cells that did notdifferentiate into astrocytes stayed in a nestin+ immature state rather than differentiated.
We also determined the effect of TGFα and noggin co-infusion for 4 weeks (Fig. 1B5). In thiscondition, most BrdU+ striatal cells were EGFR+/Olig2+/nestin+ “C-like” cells (Suppl. Fig. 2),which was similar to the TGFα only condition. This data shows that the TGFα effect waspredominant and blocked the noggin effect.
Nestin-CreERT2/R26R-YFP mice demonstrate that TGFα-induced striatal cells originate fromSVZ nestin+ precursors
An important basic biological question was to determine whether the striatal wave of C-likecells observed during TGFα infusion originated from nestin+ SVZ precursors. For this purpose,we used inducible nestin-CreERT2/R26R-YFP mice. Upon tamoxifen administration, YFPexpression in SVZ and SGZ precursors [19] allows genetic fate-mapping of adult neurogenesis.We administered tamoxifen to 5–8 week-old transgenic mice that had received a unilateral 6-OHDA injection 2 weeks before (Fig. 1C). Two days after tamoxifen treatment (Fig. 1C1),YFP+ cells were only observed in the SVZ, RMS and dentate gyrus of hippocampus (data notshown). In mice sacrificed during PBS infusion (Fig. 1C2), forebrain YFP+ cells were restrictedto the same regions, except that their distribution had changed, with many YFP+ cells in theOB (Fig. 6A). In contrast, in mice sacrificed during TGFα infusion (Fig. 1C2), YFP+ SVZ cellshad expanded and colonized the ipsilateral medial striatum, septum, corpus callosum and deeplayers of the neocortex (Fig. 6B). The majority of these YFP+ cells expressed nestin and PCNA(Fig. 6C), as well as BrdU (Fig. 6D). These data unambiguously show that striatal nestin+/PCNA+ precursors described by Cooper and Isacson [1] originated from SVZ nestin+
precursors. They also indicate that tamoxifen did not substantially interfere with TGFαsignaling [24]. YFP cells did not express GFAP at this time (Fig. 6E), indicating that we werenot measuring reactive gliosis at this point.
The C-like cell phenotype of ectopic striatal YFP+ SVZ cell progeny was confirmed bycoexpression with EGFR and Olig2 (Fig. 6F, G). The transgenic mouse model also allowed usto determine the fine morphology of striatal C-like cells: YFP+/Olig2+/nestin+/PCNA+ cells
de Chevigny et al. Page 6
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

had a complex morphology, with numerous processes oriented towards the source of TGFα(Fig. 6C, G).
Two weeks after TGFα withdrawal (Fig. 1C3), we observed a decrease in striatal, septal andolfactory bulb YFP+ cells, as well as a dispersion of YFP+ cells in the ipsilateral striatum(compare Fig. 6H with Fig. 6B), which corroborated the rat data. Also, examining the fate ofYFP+ cells in nestin-CreERT2/R26R-YFP mice 2 weeks after TGFα pump withdrawal showedthat some striatal YFP+ cells were GFAP+ astrocytes, while others were PSANCAM+
neuroblasts (Fig. 6I, J), again confirming the rat data. We also observed striatal YFP+ cells thatwere nestin+/GFAP−/PSANCAM− undifferentiated cells, with a morphology reminiscent ofastrocytes (data not shown).
Differential fate of striatal and olfactory bulb C-like cells after TGFα withdrawalIn the olfactory bulb of 6-OHDA lesioned nestin-CreERT2/R26R-YFP mice during PBSinfusion, YFP+ cells were either granular or periglomerular interneurons (data not shown),consistent with the normal fate of SVZ NPCs [25]. In contrast, TGFα infusion generatedolfactory bulb YFP+/NeuN−/Olig2+ cells, reminiscent of the C-like phenotype observed in thestriatum (Fig. 7A–C). The number of YFP+ cells in the olfactory bulb was also greatly enhancedby TGFα infusion (compare Fig. 6A and Fig. 6B). Importantly, upon TGFα withdrawal, mostYFP+ cells were granular or periglomerular interneurons (NeuN expression with typicalneuronal morphology, Fig. 7D, E), and were much more numerous than during PBS infusion(Suppl. Fig. 3). This indicates that olfactory bulb C-like cells, as opposed to their striatalcounterparts, had differentiated into mature neurons in the olfactory bulb upon TGFαwithdrawal, thus highlighting a decisive influence of the microenvironment – striatal versusbulbar – on the fate of the C-like cells.
Absence of new TH+ neurons and behavioral improvementThe ability of TGFα, noggin or TGFα̣ and noggin co- and sequential intrastriatal infusions torestore motor function in rat groups with a unilateral 6-OHDA lesion was examined.Amphetamine-induced rotations were counted before and after the 2–4 weeks infusion period[26]. No significant reductions in rotation scores for any infusion groups were found (Suppl.Fig. 4). Consistent with these findings, not a single TH+ cell that had incorporated BrdU orYFP was seen in the DA-depleted striatum in all rat and mouse groups, respectively (data notshown).
DISCUSSIONIn this study, we first characterized the phenotype of striatal nestin+/PCNA+ cells generatedby TGFα infusion in the rat DA-depleted striatum [1]. These BrdU+ cells coexpressed Olig2,EGFR, nestin and Mash1, consistent with the phenotype of multipotent C cells [21]. Secondly,we determined the fate of these cells upon TGFα withdrawal. We observed large numbers ofnew striatal neuroblasts and astrocytes, confirming the in vivo multipotency of the striatal “C-like” cells while some nestin+ SVZ derived striatal cells remained undifferentiated at this stage.Thirdly, we increased the neuroblast to astrocyte ratio in the DA-depleted striatum byadministering the BMP antagonist noggin after TGFα withdrawal. Interestingly, in our 2 weeksinfusion paradigm noggin treatment blocked glial differentiation but neuroblast differentiationwas not increased. Finally, using nestin-CreERT2/R26R-YFP mice, we showed that striatalneurogenic C-like cells originate from the SVZ and confirmed the phenotype and neuronal/glial fate of such rat cells after TGFα withdrawal. We also determined that, in contrast to theirstriatal counterparts, olfactory bulb C-like cells give rise to mature neurons, emphasizing anenvironmental effect on the fate of C-like cells. The generation of a large number of NPCswith neurogenic potential in the DA-depleted striatum is of therapeutic interest, and our
de Chevigny et al. Page 7
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

evidence suggests that such C-like cells require further midbrain or forebrain cues [13,27–29] to differentiate into mature DA neurons suitable to improve function in PD. A longer periodpost-infusion may also be required to completely understand the differentiation potential ofthese newly generated cells.
Phenotype of SVZ-derived cells and striatal response during TGFα infusionIn vivo, EGFR ligands increase C cell proliferation and colonization of the striatum by theircellular progeny [1,21]. However, the progeny phenotype of these striatal C cells has not beendetermined, making it difficult to determine whether such cells can be directed for neural repairin PD. In this study, we found that during TGFα infusion, most SVZ derived cells recruited tothe striatum displayed features of C cells, such as Olig2, nestin, EGFR, and Mash1 expression[1,8,13,16,21,22]. However, such cells were different from SVZ C cells due to an elongatedand complex morphology (while SVZ C cells have a simple, globular morphology), expressednestin more strongly and did not express Dlx2 [7,21].
TGFα-induced striatal “C-like” cells expressed markers of mitotic activity (i.e. BrdU andPCNA), as well as β-catenin, EGFR and Olig2, which are known markers of SVZ C cells andglioma-forming cells [30–33]. Hence, our data emphasize the roles of EGFR signaling, theWnt/β-catenin signaling pathway, and the transcriptional repressor activity of Olig2 in inducingadult NPCs to adopt a high replication state.
Another novel finding in this field is that TGFα̣ infusion elevated the number of microglialcells in both ipsilateral SVZ and striatum. This was not due to a direct mitogenic effect ofTGFα on microglia since they rarely incorporated BrdU (2 ± 3 %). Rather, existing microgliawere probably recruited from other brain regions [34,35]. Such microglial recruitment towardsthe region of cell proliferation was not likely due to a direct action of TGFα, since TGFαoverexpression in transgenic mice does not modify microglia number or behavior [36]. Instead,microglial cells may have been attracted by signals released by cells undergoing apoptosis, asdiscussed below. Recent reports indicate that microglia influence adult neurogenesis [37–40].For example, microglia activated by cytokines associated with T-helper cells may stimulateneurogenesis [41], while inflammation-associated microglia reduces neurogenesis [42,43].Whether the microglia affected neurogenesis in the present paradigm remains to be elucidated.
TGFα withdrawal induces a large population of neuroblasts in the DA-depleted striatumIn the second phase of the investigation, we examined the effect of TGFα withdrawal on thefate of striatal C-like cells. Two weeks after TGFα withdrawal, ~ 34 % fewer BrdU+ cells werepresent. This was at least partly due to apoptotic cell death, because TUNEL+ cells were foundin the striatal cellular wave. The surviving new cells in the striatum were composed ofGFAP+ astrocytes, DCX+/PSANCAM+/nestin+ neuroblasts, and immature cells that expressednestin and/or PCNA only. The BrdU+/GFAP+ astrocytes often expressed low levels of nestinbut did not express SSEA-1 or PCNA. Therefore, these cells likely represent an immatureastrocyte population rather than B-like “neural stem” cells [44]. Among the striatal neuroblastpopulation, ~ 82 % of the cells expressed PCNA while only ~ 7 % expressed BrdU. This, alongwith YFP/PSANCAM coexpression in the striatum of nestin-CreERT2/R26R-YFP mice, isconsistent with the progressive loss of BrdU by dilution over numerous cell divisions [45].Because PCNA was almost restricted to DCX+ cells 2 weeks after TGFα withdrawal, anydisappearance of BrdU by dilution is likely to have preferentially affected this population. Theabsence of YFP+/NeuN+ cells in the DA-depleted striatum of nestin-CreERT2/R26R-YFP miceindicates that mature neurons were not generated by TGFα in this region.
Another surprising finding is that, in contrast to SVZ and RMS neuroblasts, ectopic striatalneuroblasts did not express the transcription factor Pax6. This difference suggests that ectopic
de Chevigny et al. Page 8
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

striatal neuroblasts would differentiate into olfactory bulb GABAergic interneurons, ratherthan A16 DA neurons without further stimulation [13,14].
We also observed the presence of newly generated neuroblast clusters along blood vessels andin contact with astrocytes, similar to their normal SVZ niche [5]. The astrocytes surroundingthe neuroblast clusters likely did not arise from the SVZ given the absence of BrdU, PCNA ornestin coexpression.
Besides the astrocyte and neuroblast populations, the majority of remaining striatal BrdU+ cellswere immature because they expressed nestin, without Olig2, EGFR or Mash1 expression atthis time. The long-term fate of those BrdU+/nestin+ cells is uncertain: they may retain animmature phenotype, or ultimately differentiate or die due to lack of fate specification and/ortrophic supply.
Interestingly, no BrdU+ cells in the striatum were CNPase+, O4+ oligodendrocytes. This resultwas surprising because, in the presence of TGFα, a high proportion of BrdU+ cells expressedOlig2 and Mash1, which are transcription factors involved in oligodendrocyte specification inthe embryo and postnatal SVZ [46,47]. Nevertheless, the absence of NG2 expression by theOlig2+/Mash1+ cells confirmed that they were not typical oligodendrocyte precursors [48]. Arecent study using CNP-hEGFR overexpressing mice indicates that EGFR signaling activationis involved in oligodendrogenesis and remyelination in demyelinating conditions [49].Therefore, it is possible that such demyelinating conditions are required for TGFα-activatedNPCs to undergo oligodendrogenesis [50].
Effects of TGFα and noggin sequential infusions on the striatal neuroblast to astrocyte ratioWe examined whether administration of the BMP antagonist noggin after TGFα withdrawalwould influence the neuroblast to astrocyte ratio by blocking gliogenesis and/or inducingneurogenesis [51]. We found that striatal infusion of noggin without TGFα in 6-OHDAlesioned rats significantly increased SVZ neurogenesis. This is in line with published data[17] and demonstrates that the noggin protein diffuses over long distances (about 2.7 mm) andremains functional when chronically infused in vivo. In contrast to SVZ, the normal striatumis gliogenic for SVZ cells [52], probably due to the absence or too low levels of noggin andother BMP antagonists such as follistatin or chordin [53]. However, virally delivered nogginreverses this effect and induces neurogenesis from SVZ cells recruited to the striatum, eitherafter transplantation or following BDNF-induced recruitment [17,54]. Strikingly, in our invivo DA depletion paradigm noggin did not increase striatal neurogenesis, but only blockedgliogenesis. This difference with previous literature [17,54] may be due to paradigmdifferences. The present data suggest that noggin is primarily anti-gliogenic, while other factorsconfined to SVZ induce neurogenesis when gliogenesis is blocked.
Comparisons with other paradigms inducing adult SVZ neurogenesisAs mentioned, we found ~ 8,000 newly generated striatal neuroblasts 2 weeks after the EGFRligand withdrawal. This cell number is similar to that found by Arvidsson et al. [55] 2 weeksafter striatal stroke induction (6,312 DCX+ neuroblasts), but it is ~ 4 times larger than the 1,600BrdU+/Tuj1+ cells found after adenoviral co-administration of BDNF and noggin [54]. In ourparadigm, striatal neuroblasts formed tight clusters throughout the medial striatum, whereasthey were individualized in the studies from Arvidsson et al. and Chmielnicki et al. [54,55].Furthermore, in our paradigm 82% of neuroblasts continued to divide 2 weeks after growthfactor withdrawal, but this remains unknown in the other paradigms [54,55].
There are both analogies and differences between our in vivo paradigm and published invitro neurosphere culture assays [56]. In neurosphere cultures, SVZ cells grow as clonal spheres
de Chevigny et al. Page 9
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

in the presence of EGF [57]. Notably, neurosphere cells rapidly upregulate Olig2 anddownregulate Pax6 in these proliferative conditions [58], and withdrawing EGF is required fordifferentiation into neurons and glia after plating [21], as shown here in vivo. However, thefact that SVZ cells do not differentiate into oligodendrocytes in vivo in the present study whilesome do in vitro [56] suggests that the striatal microenvironment influences the fate of SVZcells.
CONCLUSIONWe demonstrated that striatal TGFα infusion recruits a large population of SVZ-derivedmultipotent “C-like” cells to the DA-depleted striatum. After TGFα withdrawal, astrocytes,neuroblasts and undifferentiated cells, but no oligodendrocytes or differentiated neurons, aregenerated. Noggin infusion after TGFα blocks astrocyte generation in favor of anundifferentiated phenotype, thus increasing the neuroblast to astrocyte ratio. The findings thatstriatal neural precursors with neurogenic potential are generated in large numbers in the adultDA-depleted striatum provides a unique opportunity to differentiate such cells to functionalDA neurons for PD.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
ACKNOWLEDGMENTSSupported by NIH/NINDS P50 NS39793, the Orchard Foundation, Anti-Aging Foundation, Michael K. SternFoundation, Harold and Ronna Cooper Family (OI); NIDA K02 DA023555, R01 DA16765, NARSAD YoungInvestigator Award (AJE); Canadian Institute of Health Research (DCL).
REFERENCES1. Cooper O, Isacson O. Intrastriatal transforming growth factor alpha delivery to a model of Parkinson's
disease induces proliferation and migration of endogenous adult neural progenitor cells withoutdifferentiation into dopaminergic neurons. J Neurosci 2004;24:8924–8931. [PubMed: 15483111]
2. Hoglinger GU, Rizk P, Muriel MP, et al. Dopamine depletion impairs precursor cell proliferation inParkinson disease. Nat Neurosci 2004;7:726–735. [PubMed: 15195095]
3. Isacson O. The production and use of cells as therapeutic agents in neurodegenerative diseases. LancetNeurol 2003;2:417–424. [PubMed: 12849120]
4. Isacson O, Bjorklund LM, Schumacher JM. Toward full restoration of synaptic and terminal functionof the dopaminergic system in Parkinson's disease by stem cells. Ann Neurol 2003;53:S135–S146.[PubMed: 12666105]discussion S146-138
5. Alvarez-Buylla A, Garcia-Verdugo JM. Neurogenesis in adult subventricular zone. J Neurosci2002;22:629–634. [PubMed: 11826091]
6. Doetsch F. A niche for adult neural stem cells. Curr Opin Genet Dev 2003;13:543–550. [PubMed:14550422]
7. Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A. Cellular composition and three-dimensionalorganization of the subventricular germinal zone in the adult mammalian brain. J Neurosci1997;17:5046–5061. [PubMed: 9185542]
8. Garcia-Verdugo JM, Doetsch F, Wichterle H, et al. Architecture and cell types of the adultsubventricular zone: in search of the stem cells. J Neurobiol 1998;36:234–248. [PubMed: 9712307]
9. Belluzzi O, Benedusi M, Ackman J, et al. Electrophysiological differentiation of new neurons in theolfactory bulb. J Neurosci 2003;23:10411–10418. [PubMed: 14614100]
10. Puopolo M, Bean BP, Raviola E. Spontaneous activity of isolated dopaminergic periglomerular cellsof the main olfactory bulb. Journal of neurophysiology 2005;94:3618–3627. [PubMed: 16033943]
de Chevigny et al. Page 10
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

11. Menn B, Garcia-Verdugo JM, Yaschine C, et al. Origin of oligodendrocytes in the subventricularzone of the adult brain. J Neurosci 2006;26:7907–7918. [PubMed: 16870736]
12. Freundlieb N, Francois C, Tande D, et al. Dopaminergic substantia nigra neurons projecttopographically organized to the subventricular zone and stimulate precursor cell proliferation inaged primates. J Neurosci 2006;26:2321–2325. [PubMed: 16495459]
13. Hack MA, Saghatelyan A, de Chevigny A, et al. Neuronal fate determinants of adult olfactory bulbneurogenesis. Nat Neurosci 2005;8:865–872. [PubMed: 15951811]
14. Kohwi M, Osumi N, Rubenstein JL, et al. Pax6 is required for making specific subpopulations ofgranule and periglomerular neurons in the olfactory bulb. J Neurosci 2005;25:6997–7003. [PubMed:16049175]
15. Winner B, Geyer M, Couillard-Despres S, et al. Striatal deafferentation increases dopaminergicneurogenesis in the adult olfactory bulb. Exp Neurol 2006;197:113–121. [PubMed: 16246330]
16. Fallon J, Reid S, Kinyamu R, et al. In vivo induction of massive proliferation, directed migration,and differentiation of neural cells in the adult mammalian brain. Proc Natl Acad Sci U S A2000;97:14686–14691. [PubMed: 11121069]
17. Lim DA, Tramontin AD, Trevejo JM, et al. Noggin antagonizes BMP signaling to create a niche foradult neurogenesis. Neuron 2000;28:713–726. [PubMed: 11163261]
18. Bjorklund LM, Sanchez-Pernaute R, Chung S, et al. Embryonic stem cells develop into functionaldopaminergic neurons after transplantation in a Parkinson rat model. Proc Natl Acad Sci U S A2002;99:2344–2349. [PubMed: 11782534]
19. Lagace DC, Whitman MC, Noonan MA, et al. Dynamic contribution of nestin-expressing stem cellsto adult neurogenesis. J Neurosci 2007;27:12623–12629. [PubMed: 18003841]
20. Ries V, Henchcliffe C, Kareva T, et al. Oncoprotein Akt/PKB induces trophic effects in murine modelsof Parkinson's disease. Proc Natl Acad Sci U S A 2006;103:18757–18762. [PubMed: 17116866]
21. Doetsch F, Petreanu L, Caille I, et al. EGF converts transit-amplifying neurogenic precursors in theadult brain into multipotent stem cells. Neuron 2002;36:1021–1034. [PubMed: 12495619]
22. Parras CM, Galli R, Britz O, et al. Mash1 specifies neurons and oligodendrocytes in the postnatalbrain. The EMBO journal 2004;23:4495–4505. [PubMed: 15496983]
23. Smith WC, Harland RM. Expression cloning of noggin, a new dorsalizing factor localized to theSpemann organizer in Xenopus embryos. Cell 1992;70:829–840. [PubMed: 1339313]
24. Johnston SR, Dowsett M, Smith IE. Towards a molecular basis for tamoxifen resistance in breastcancer. Ann Oncol 1992;3:503–511. [PubMed: 1498070]
25. Carleton A, Petreanu LT, Lansford R, et al. Becoming a new neuron in the adult olfactory bulb. NatNeurosci 2003;6:507–518. [PubMed: 12704391]
26. Moore AE, Cicchetti F, Hennen J, et al. Parkinsonian motor deficits are reflected by proportional A9/A10 dopamine neuron degeneration in the rat. Exp Neurol 2001;172:363–376. [PubMed: 11716560]
27. Shim JW, Park CH, Bae YC, et al. Generation of functional dopamine neurons from neural precursorcells isolated from the subventricular zone and white matter of the adult rat brain using Nurr1overexpression. Stem Cells 2007;25:1252–1262. [PubMed: 17234994]
28. Papanikolaou T, Lennington JB, Betz A, et al. In vitro generation of dopaminergic neurons from adultsubventricular zone neural progenitor cells. Stem Cells Dev 2008;17:157–172. [PubMed: 18248323]
29. Hermann A, Maisel M, Wegner F, et al. Multipotent neural stem cells from the adult tegmentum withdopaminergic potential develop essential properties of functional neurons. Stem Cells 2006;24:949–964. [PubMed: 16373695]
30. Caricasole A, Bakker A, Copani A, et al. Two sides of the same coin: Wnt signaling inneurodegeneration and neuro-oncology. Biosci Rep 2005;25:309–327. [PubMed: 16307379]
31. Holland EC. Progenitor cells and glioma formation. Curr Opin Neurol 2001;14:683–688. [PubMed:11723374]
32. Ligon KL, Huillard E, Mehta S, et al. Olig2-regulated lineage-restricted pathway controls replicationcompetence in neural stem cells and malignant glioma. Neuron 2007;53:503–517. [PubMed:17296553]
de Chevigny et al. Page 11
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

33. Nicholas MK, Lukas RV, Jafri NF, et al. Epidermal growth factor receptor - mediated signaltransduction in the development and therapy of gliomas. Clin Cancer Res 2006;12:7261–7270.[PubMed: 17189397]
34. Davalos D, Grutzendler J, Yang G, et al. ATP mediates rapid microglial response to local brain injuryin vivo. Nat Neurosci 2005;8:752–758. [PubMed: 15895084]
35. Nimmerjahn A, Kirchhoff F, Helmchen F. Resting microglial cells are highly dynamic surveillantsof brain parenchyma in vivo. Science 2005;308:1314–1318. [PubMed: 15831717]
36. Rabchevsky AG, Weinitz JM, Coulpier M, et al. A role for transforming growth factor alpha as aninducer of astrogliosis. J Neurosci 1998;18:10541–10552. [PubMed: 9852591]
37. Walton NM, Sutter BM, Laywell ED, et al. Microglia instruct subventricular zone neurogenesis. Glia2006;54:815–825. [PubMed: 16977605]
38. Ziv Y, Ron N, Butovsky O, et al. Immune cells contribute to the maintenance of neurogenesis andspatial learning abilities in adulthood. Nat Neurosci 2006;9:268–275. [PubMed: 16415867]
39. Ziv Y, Avidan H, Pluchino S, et al. Synergy between immune cells and adult neural stem/progenitorcells promotes functional recovery from spinal cord injury. Proc Natl Acad Sci U S A2006;103:13174–13179. [PubMed: 16938843]
40. Goings GE, Kozlowski DA, Szele FG. Differential activation of microglia in neurogenic versus non-neurogenic regions of the forebrain. Glia 2006;54:329–342. [PubMed: 16862532]
41. Butovsky O, Ziv Y, Schwartz A, et al. Microglia activated by IL-4 or IFN-gamma differentially induceneurogenesis and oligodendrogenesis from adult stem/progenitor cells. Mol Cell Neurosci2006;31:149–160. [PubMed: 16297637]
42. Ekdahl CT, Claasen JH, Bonde S, et al. Inflammation is detrimental for neurogenesis in adult brain.Proc Natl Acad Sci U S A 2003;100:13632–13637. [PubMed: 14581618]
43. Monje ML, Toda H, Palmer TD. Inflammatory blockade restores adult hippocampal neurogenesis.Science 2003;302:1760–1765. [PubMed: 14615545]
44. Capela A, Temple S. LeX/ssea-1 is expressed by adult mouse CNS stem cells, identifying them asnonependymal. Neuron 2002;35:865–875. [PubMed: 12372282]
45. Dayer AG, Ford AA, Cleaver KM, et al. Short-term and long-term survival of new neurons in the ratdentate gyrus. J Comp Neurol 2003;460:563–572. [PubMed: 12717714]
46. Kondo T, Raff M. Basic helix-loop-helix proteins and the timing of oligodendrocyte differentiation.Development 2000;127:2989–2998. [PubMed: 10862737]
47. Lu QR, Yuk D, Alberta JA, et al. Sonic hedgehog--regulated oligodendrocyte lineage genes encodingbHLH proteins in the mammalian central nervous system. Neuron 2000;25:317–329. [PubMed:10719888]
48. Polito A, Reynolds R. NG2-expressing cells as oligodendrocyte progenitors in the normal anddemyelinated adult central nervous system. J Anat 2005;207:707–716. [PubMed: 16367798]
49. Aguirre A, Dupree JL, Mangin JM, et al. A functional role for EGFR signaling in myelination andremyelination. Nat Neurosci 2007;10:990–1002. [PubMed: 17618276]
50. Chandran S, Compston A. Neural stem cells as a potential source of oligodendrocytes for myelinrepair. Journal of the neurological sciences 2005;233:179–181. [PubMed: 15907942]
51. Sonntag KC, Pruszak J, Yoshizaki T, et al. Enhanced yield of neuroepithelial precursors and midbrain-like dopaminergic neurons from human embryonic stem cells using the bone morphogenic proteinantagonist noggin. Stem Cells 2007;25:411–418. [PubMed: 17038668]
52. Herrera DG, Garcia-Verdugo JM, Alvarez-Buylla A. Adult-derived neural precursors transplantedinto multiple regions in the adult brain. Ann Neurol 1999;46:867–877. [PubMed: 10589539]
53. Balemans W, Patel N, Ebeling M, et al. Identification of a 52 kb deletion downstream of the SOSTgene in patients with van Buchem disease. Journal of medical genetics 2002;39:91–97. [PubMed:11836356]
54. Chmielnicki E, Benraiss A, Economides AN, et al. Adenovirally expressed noggin and brain-derivedneurotrophic factor cooperate to induce new medium spiny neurons from resident progenitor cellsin the adult striatal ventricular zone. J Neurosci 2004;24:2133–2142. [PubMed: 14999064]
55. Arvidsson A, Collin T, Kirik D, et al. Neuronal replacement from endogenous precursors in the adultbrain after stroke. Nat Med 2002;8:963–970. [PubMed: 12161747]
de Chevigny et al. Page 12
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

56. Doetsch F, Caille I, Lim DA, et al. Subventricular zone astrocytes are neural stem cells in the adultmammalian brain. Cell 1999;97:703–716. [PubMed: 10380923]
57. Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A. Regeneration of a germinal layer in the adultmammalian brain. Proc Natl Acad Sci U S A 1999;96:11619–11624. [PubMed: 10500226]
58. Hack MA, Sugimori M, Lundberg C, et al. Regionalization and fate specification in neurospheres:the role of Olig2 and Pax6. Mol Cell Neurosci 2004;25:664–678. [PubMed: 15080895]
de Chevigny et al. Page 13
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
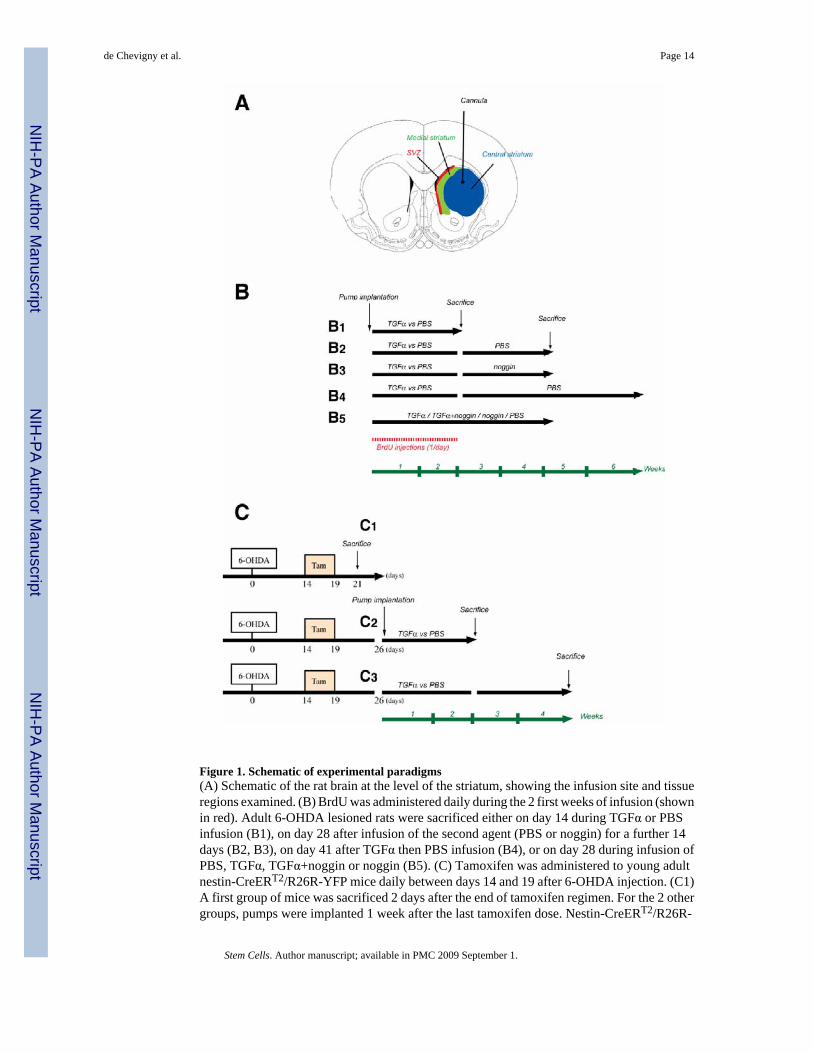
Figure 1. Schematic of experimental paradigms(A) Schematic of the rat brain at the level of the striatum, showing the infusion site and tissueregions examined. (B) BrdU was administered daily during the 2 first weeks of infusion (shownin red). Adult 6-OHDA lesioned rats were sacrificed either on day 14 during TGFα or PBSinfusion (B1), on day 28 after infusion of the second agent (PBS or noggin) for a further 14days (B2, B3), on day 41 after TGFα then PBS infusion (B4), or on day 28 during infusion ofPBS, TGFα, TGFα+noggin or noggin (B5). (C) Tamoxifen was administered to young adultnestin-CreERT2/R26R-YFP mice daily between days 14 and 19 after 6-OHDA injection. (C1)A first group of mice was sacrificed 2 days after the end of tamoxifen regimen. For the 2 othergroups, pumps were implanted 1 week after the last tamoxifen dose. Nestin-CreERT2/R26R-
de Chevigny et al. Page 14
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

YFP mice were sacrificed either on day 14 during TGFα or PBS infusion (C2), or on day 28after infusion of PBS for a further 14 days period (C3).
de Chevigny et al. Page 15
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
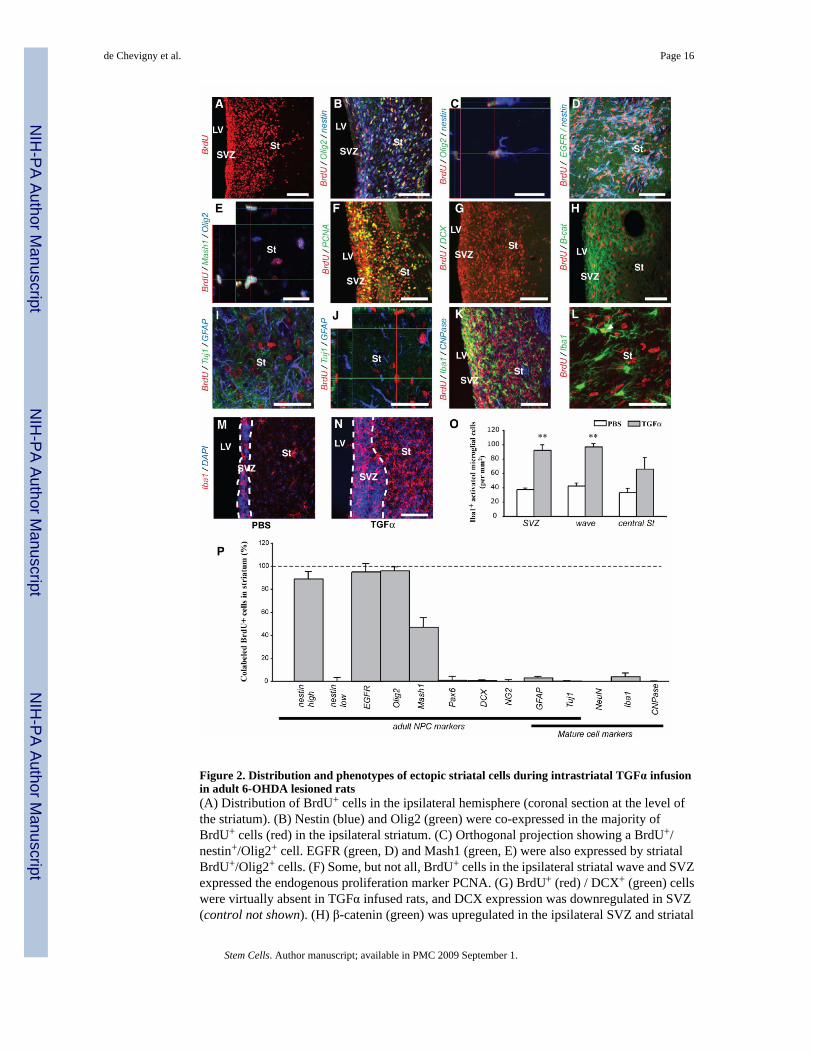
Figure 2. Distribution and phenotypes of ectopic striatal cells during intrastriatal TGFα infusionin adult 6-OHDA lesioned rats(A) Distribution of BrdU+ cells in the ipsilateral hemisphere (coronal section at the level ofthe striatum). (B) Nestin (blue) and Olig2 (green) were co-expressed in the majority ofBrdU+ cells (red) in the ipsilateral striatum. (C) Orthogonal projection showing a BrdU+/nestin+/Olig2+ cell. EGFR (green, D) and Mash1 (green, E) were also expressed by striatalBrdU+/Olig2+ cells. (F) Some, but not all, BrdU+ cells in the ipsilateral striatal wave and SVZexpressed the endogenous proliferation marker PCNA. (G) BrdU+ (red) / DCX+ (green) cellswere virtually absent in TGFα infused rats, and DCX expression was downregulated in SVZ(control not shown). (H) β-catenin (green) was upregulated in the ipsilateral SVZ and striatal
de Chevigny et al. Page 16
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

wave. (I, J) No BrdU+ cells (red) expressed either Tuj1 (green) or GFAP (blue) in the striatalwave (orthogonal projection in J). (K, L) Few striatal BrdU+ cells (red) were Iba1+ microglialcells (green) and none were CNPase+ oligodendrocytes (blue). Only one BrdU+ cell expressesIba1 in (L). (M, N) Representative images of Iba1+ microglia in the SVZ and ipsilateral striatumduring PBS or TGFα infusion. (O) Upon quantification, a higher Iba1+ microglial density wasfound in the ipsilateral SVZ and striatum during TGFα administration (grey) than during PBSinfusion (white). (P) Histogram showing the percentage of marker-positive cells among striatalBrdU+ cells. Scale bars: A, B, F, G, K, M, N, 100 µm; C, E, 20 µm; D, H, 50 µm; I, J, L, 40µm.
de Chevigny et al. Page 17
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
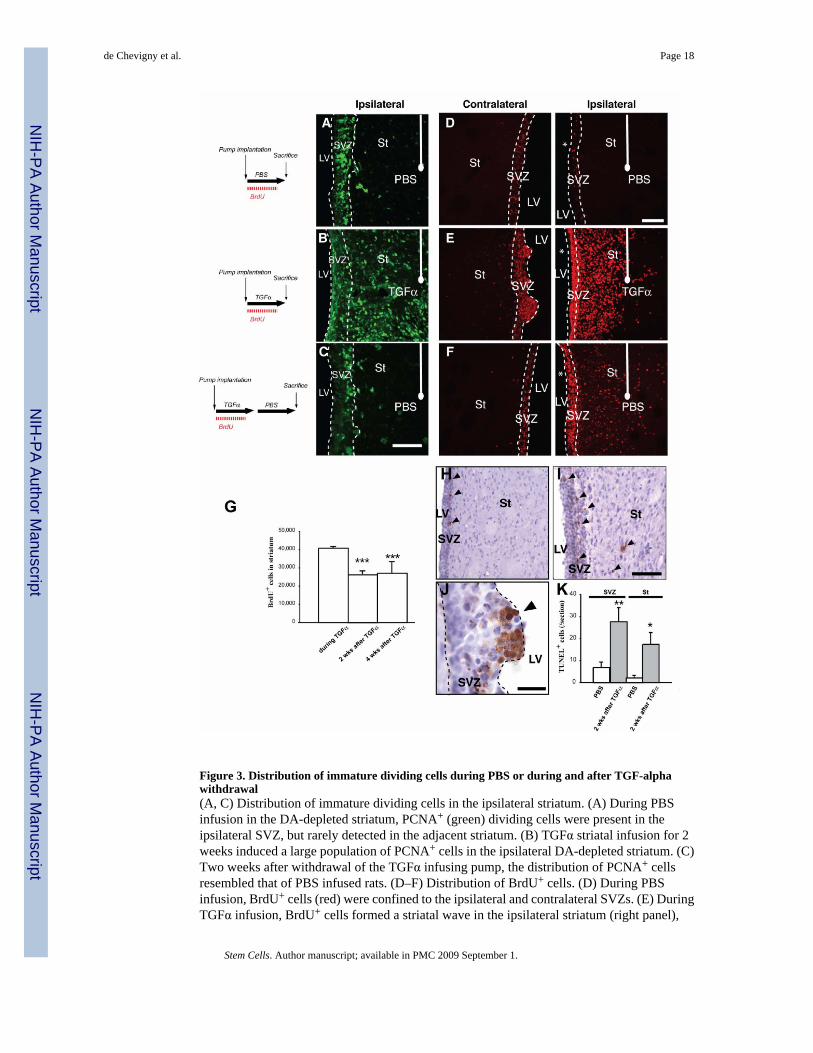
Figure 3. Distribution of immature dividing cells during PBS or during and after TGF-alphawithdrawal(A, C) Distribution of immature dividing cells in the ipsilateral striatum. (A) During PBSinfusion in the DA-depleted striatum, PCNA+ (green) dividing cells were present in theipsilateral SVZ, but rarely detected in the adjacent striatum. (B) TGFα striatal infusion for 2weeks induced a large population of PCNA+ cells in the ipsilateral DA-depleted striatum. (C)Two weeks after withdrawal of the TGFα infusing pump, the distribution of PCNA+ cellsresembled that of PBS infused rats. (D–F) Distribution of BrdU+ cells. (D) During PBSinfusion, BrdU+ cells (red) were confined to the ipsilateral and contralateral SVZs. (E) DuringTGFα infusion, BrdU+ cells formed a striatal wave in the ipsilateral striatum (right panel),
de Chevigny et al. Page 18
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

while in the contralateral hemisphere they formed hyper-proliferative nodules protruding fromthe SVZ towards the source of TGFα (left panel). (F) Two weeks after TGFα withdrawal,contralateral ventricular BrdU+ hyper-proliferative nodules were not found (left panel).BrdU+ cells were observed in the ipsilateral striatum (right), although their distribution pattern(C, right) and number had changed. Note the ~ 35% reduction in total BrdU+ cell number afterTGFα withdrawal (G). (H–K) Programmed cell death is induced by TGFα treatment. (H, I)TUNEL+ cells in ipsilateral SVZ and striatum of rats infused with PBS (H) or TGFα (I), 2weeks after its withdrawal. Arrowheads show TUNEL+ cells. The number of TUNEL+ cellswas significantly higher in rats sacrificed after TGFα infusion than in their control counterparts(K). (J) In one rat that died 5 days after TGFα withdrawal, ventricular nodules were found andcontained a high proportion of TUNEL+ cells. Scale bars: A–B, 50 µm; D–F, 100 µm; H, I,200 µm; J, 50 µm.
de Chevigny et al. Page 19
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
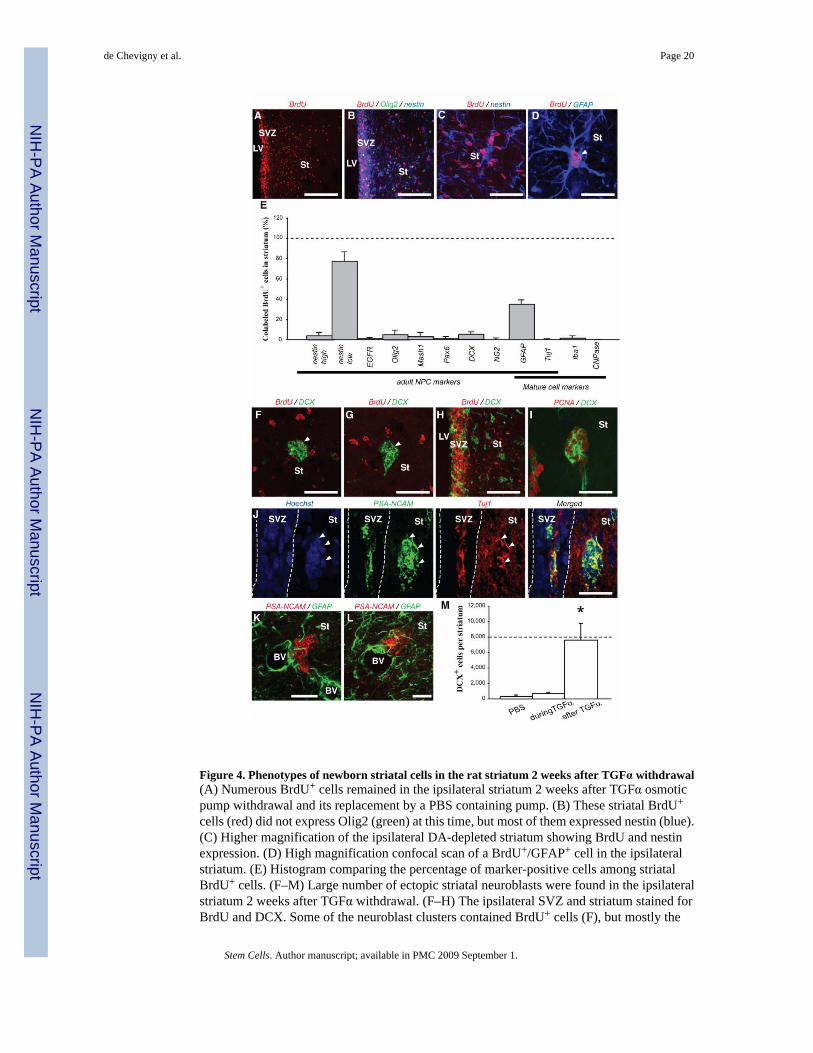
Figure 4. Phenotypes of newborn striatal cells in the rat striatum 2 weeks after TGFα withdrawal(A) Numerous BrdU+ cells remained in the ipsilateral striatum 2 weeks after TGFα osmoticpump withdrawal and its replacement by a PBS containing pump. (B) These striatal BrdU+
cells (red) did not express Olig2 (green) at this time, but most of them expressed nestin (blue).(C) Higher magnification of the ipsilateral DA-depleted striatum showing BrdU and nestinexpression. (D) High magnification confocal scan of a BrdU+/GFAP+ cell in the ipsilateralstriatum. (E) Histogram comparing the percentage of marker-positive cells among striatalBrdU+ cells. (F–M) Large number of ectopic striatal neuroblasts were found in the ipsilateralstriatum 2 weeks after TGFα withdrawal. (F–H) The ipsilateral SVZ and striatum stained forBrdU and DCX. Some of the neuroblast clusters contained BrdU+ cells (F), but mostly the
de Chevigny et al. Page 20
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

neuroblast clusters were BrdU− (G). (H) Clusters of DCX+ neuroblasts were numerous anddistributed throughout the striatum. (I) Most DCX+ cells inside striatal neuroblast clustersexpressed PCNA, demonstrating a mitotic state. (J) Neuroblasts in striatal clusters coexpressedPSANCAM (green) and the neuronal marker Tuj1 (red). Arrowheads show striatalPSANCAM+/Tuj1+ cells. (K, L) Most PSANCAM+ neuroblast clusters (red) were directlyapposed to blood vessels and astrocyte processes (green). (M) Histogram showing an increasednumber of striatal neuroblasts 2 weeks after TGFα withdrawal as compared to control, PBSinfused rats. Scale bars: A, 100 µm; B, 50 µm; C, 25 µm; D, 20 µm, F, G, I–K, 25 µm; H, 50µm.
de Chevigny et al. Page 21
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
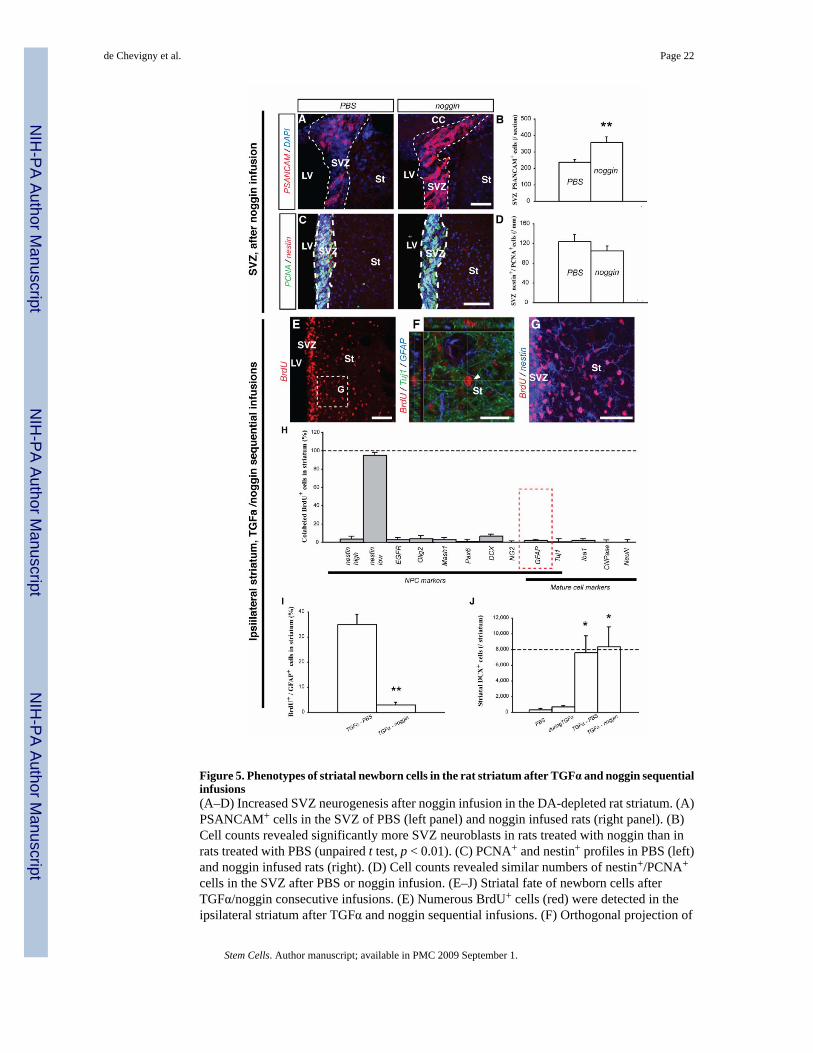
Figure 5. Phenotypes of striatal newborn cells in the rat striatum after TGFα and noggin sequentialinfusions(A–D) Increased SVZ neurogenesis after noggin infusion in the DA-depleted rat striatum. (A)PSANCAM+ cells in the SVZ of PBS (left panel) and noggin infused rats (right panel). (B)Cell counts revealed significantly more SVZ neuroblasts in rats treated with noggin than inrats treated with PBS (unpaired t test, p < 0.01). (C) PCNA+ and nestin+ profiles in PBS (left)and noggin infused rats (right). (D) Cell counts revealed similar numbers of nestin+/PCNA+
cells in the SVZ after PBS or noggin infusion. (E–J) Striatal fate of newborn cells afterTGFα/noggin consecutive infusions. (E) Numerous BrdU+ cells (red) were detected in theipsilateral striatum after TGFα and noggin sequential infusions. (F) Orthogonal projection of
de Chevigny et al. Page 22
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

a striatal BrdU+ cell (red) that neither expressed Tuj1 (green), nor GFAP (blue). (G) Highermagnification image of the region in (E) showed that most striatal BrdU+ cells expressed nestin(blue). (H) Histogram comparing the percentage of marker-positive cells among striatalBrdU+ cells. (I) Histogram comparing the proportion of BrdU+ cells colabeled with GFAP, inrats sequentially infused with TGFα and then PBS, or TGFα and then noggin. (J) Mean striatalnumber of DCX+ neuroblasts per ipsilateral striatum in the 4 conditions. Scale bars: A, G, 50µm; C, E, 100 µm; F, 25 µm.
de Chevigny et al. Page 23
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
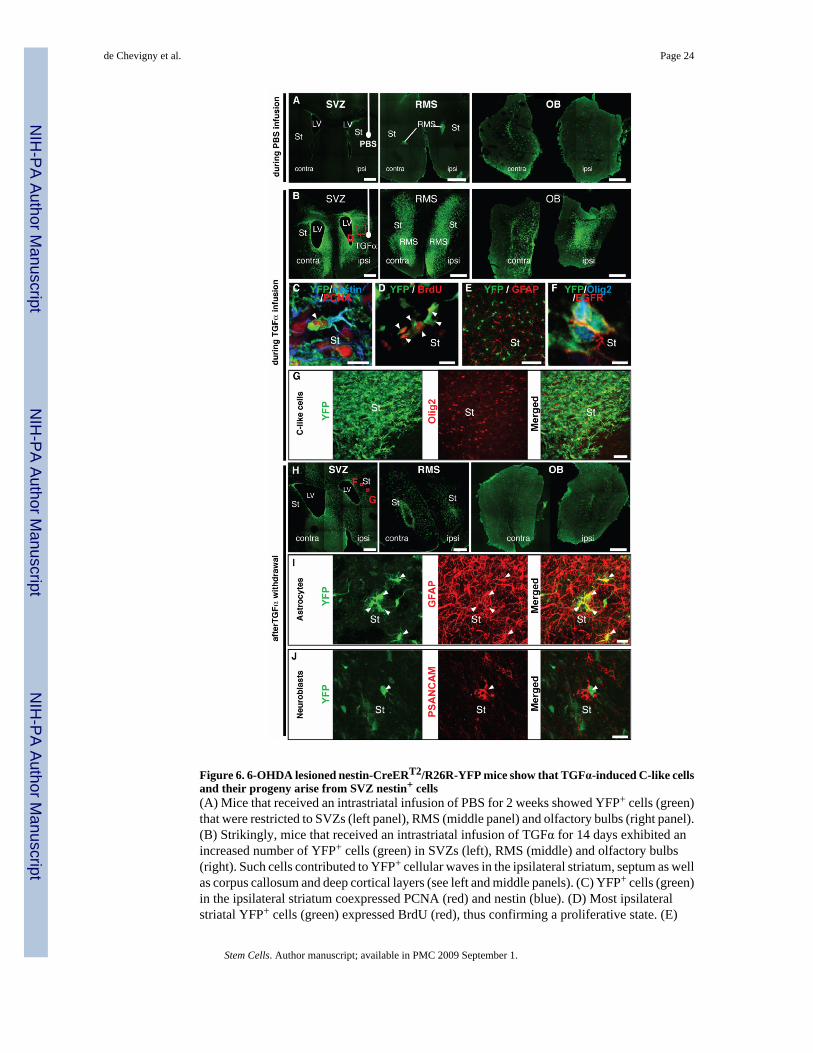
Figure 6. 6-OHDA lesioned nestin-CreERT2/R26R-YFP mice show that TGFα-induced C-like cellsand their progeny arise from SVZ nestin+ cells(A) Mice that received an intrastriatal infusion of PBS for 2 weeks showed YFP+ cells (green)that were restricted to SVZs (left panel), RMS (middle panel) and olfactory bulbs (right panel).(B) Strikingly, mice that received an intrastriatal infusion of TGFα for 14 days exhibited anincreased number of YFP+ cells (green) in SVZs (left), RMS (middle) and olfactory bulbs(right). Such cells contributed to YFP+ cellular waves in the ipsilateral striatum, septum as wellas corpus callosum and deep cortical layers (see left and middle panels). (C) YFP+ cells (green)in the ipsilateral striatum coexpressed PCNA (red) and nestin (blue). (D) Most ipsilateralstriatal YFP+ cells (green) expressed BrdU (red), thus confirming a proliferative state. (E)
de Chevigny et al. Page 24
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Single plan confocal scan showing that no YFP+ cells (green) were GFAP+ astrocytes (red) inmice sacrificed during TGFα infusion. (F) YFP+ cells (green) coexpressing EGFR (red) andOlig2 (blue) in the DA-depleted striatum. (G) At a lower magnification, most ipsilateral striatalYFP+ cells (green) coexpressed Olig2 (red). (G–I) Distribution and phenotypes of SVZ-derivedNPCs 2 weeks after TGFα withdrawal. (G) Numerous YFP+ cells (green) were observed inthe SVZs, ipsilateral striatum, septum (left), RMS (middle) and olfactory bulbs (right). (H)Numerous ipsilateral striatal YFP+ cells (green, left) coexpressed GFAP (red, middle). Rightpanel depicts a merged image. (I) Other striatal YFP+ cells (green, left) coexpressedPSANCAM (red, middle), indicating a neuroblast phenotype (see right panel for a mergedimage). Scale bars: A, B, H, 500 µm; C, D, 10 µm, E, 400 µm; F, 5 µm; G, 50 µm, I, J, 20 µm.
de Chevigny et al. Page 25
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
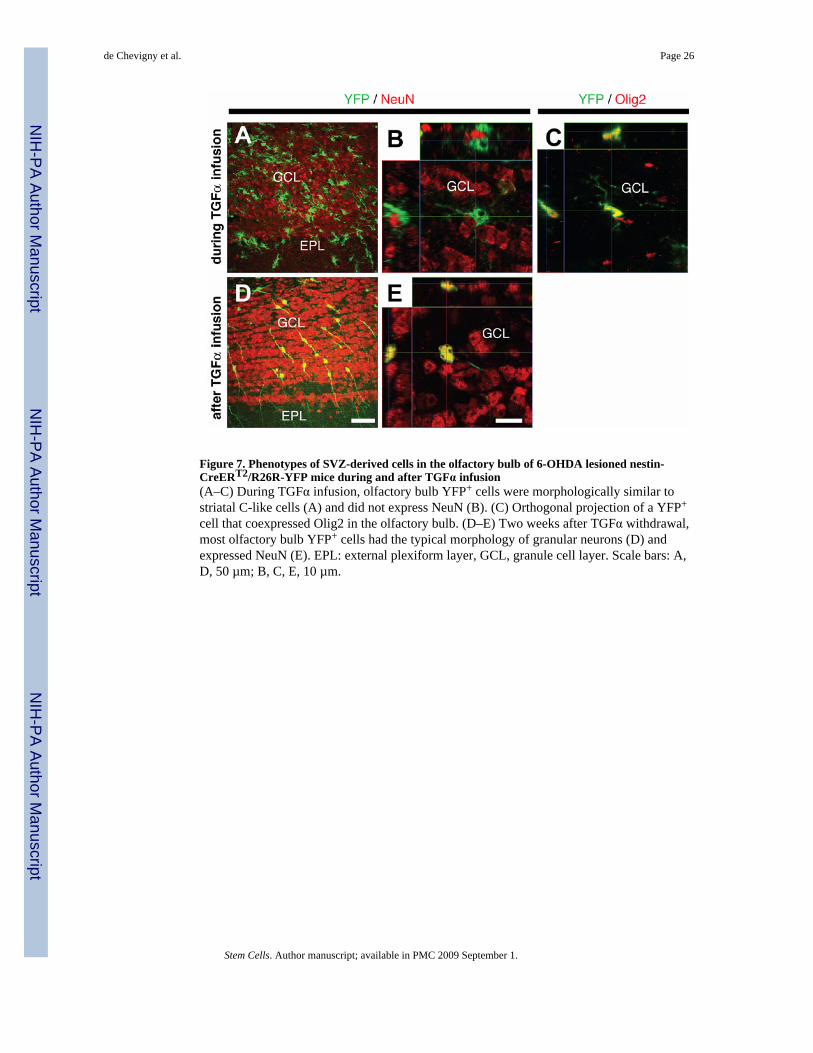
Figure 7. Phenotypes of SVZ-derived cells in the olfactory bulb of 6-OHDA lesioned nestin-CreERT2/R26R-YFP mice during and after TGFα infusion(A–C) During TGFα infusion, olfactory bulb YFP+ cells were morphologically similar tostriatal C-like cells (A) and did not express NeuN (B). (C) Orthogonal projection of a YFP+
cell that coexpressed Olig2 in the olfactory bulb. (D–E) Two weeks after TGFα withdrawal,most olfactory bulb YFP+ cells had the typical morphology of granular neurons (D) andexpressed NeuN (E). EPL: external plexiform layer, GCL, granule cell layer. Scale bars: A,D, 50 µm; B, C, E, 10 µm.
de Chevigny et al. Page 26
Stem Cells. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript














![Compensation for cranial spill-in into the cerebellum improves quantitation of striatal dopamine D2/3 receptors in rats with prolonged [18F]-DMFP infusions](https://static.fdokumen.com/doc/165x107/633a5d216cd679033b0e56dd/compensation-for-cranial-spill-in-into-the-cerebellum-improves-quantitation-of-striatal.jpg)





