
In vivo pharmacology of the dopaminergic stabilizer pridopidine
8
Neuropharmacology and Analgesia In vivo pharmacology of the dopaminergic stabilizer pridopidine Henrik Ponten a, ⁎, Johan Kullingsjö a , Sören Lagerkvist a , Peter Martin b , Fredrik Pettersson a , Clas Sonesson a , Susanna Waters a , Nicholas Waters a a NeuroSearch Sweden AB, Arvid Wallgrens Backe 20, SE-413 46 Gothenburg, Sweden b Vårdcentralen Lerum, Göteborgsvägen 9, SE-443 25, Lerum, Sweden abstract article info Article history: Received 17 February 2010 Received in revised form 18 June 2010 Accepted 11 July 2010 Available online 24 July 2010 Keywords: Pridopidine ACR16 Dopamine Dopaminergic stabilizer Huntington's disease Pharmacology Pridopidine (ACR16) belongs to a new pharmacological class of agents affecting the central nervous system called dopaminergic stabilizers. Dopaminergic stabilizers act primarily at dopamine type 2 (D 2 ) receptors and display state-dependent behavioural effects. This article aims to give an overview of the preclinical neurochemical and behavioural in vivo pharmacological properties of pridopidine. Pridopidine was given s.c. to male Sprague–Dawley rats (locomotor, microdialysis and tissue neurochemistry) and i.p. to Swiss male mice (tail suspension test). Pridopidine dose-dependently increased striatal tissue levels of the dopamine metabolite 3,4-dihydroxyphenylalanin (ED 50 =81 μmol/kg), and prefrontal cortex dialysate levels of dopamine and noradrenaline as measured by high performance liquid chromatography. The agent reduced hyperlocomotion (D-amphetamine: ED 50 =54 μmol/kg; MK-801: ED 50 =40 μmol/kg), but preserved spontaneous locomotor activity, confirming state-dependent behavioural effects. In addition, pridopidine significantly reduced immobility time in the tail suspension test. We conclude that pridopidine state-dependently stabilizes psychomotor activity by the dual actions of functional dopamine D 2 receptor antagonism and strengthening of cortical glutamate functions in various settings of perturbed neurotransmission. The putative restoration of function in cortico–subcortical circuitry by pridopidine is likely to make it useful for ameliorating several neurological and psychiatric disorders, including Huntington's disease. © 2010 Elsevier B.V. All rights reserved. 1. Introduction Pridopidine (ACR16; 4-[3-(methylsulfonyl)phenyl]-1-propylpi- peridine hydrochloride; Huntexil ® ), originally referred to as a modulator of dopamine neurotransmission (Sonesson et al., 2000), belongs to a new pharmacological class of agents acting on the central nervous system. Through their primary action at dopamine type 2 (D 2 ) receptors, these compounds stabilize dysregulated psychomotor functions (i.e. they reverse behavioural states originating from both hypo- and hyper-dopaminergia), while having only subtle effects on normal psychomotor activity (Natesan et al., 2006; Pettersson et al., 2010; Sonesson et al., 2000). Thus, these compounds regulate both hyper- and hypoactive functioning in areas of the brain receiving dopamine input, leading to reduced excitatory and inhibitory stress, resulting in improved behavioural performance. These pharmacody- namic actions of this new class (known as dopaminergic stabilizers) do not appear to fit fully into the current classification of dopamine D 2 receptor-acting compounds (Carlsson, 2000; Rung et al., 2008): antagonists, agonists, partial agonists or inverse agonists (Strange, 2008). Pridopidine has been shown to bind to striatal dopamine D 2 receptors in vivo, in a dose range where the agent is active in models assessing antipsychotic-like efficacy, and, similar to dopamine D 2 receptor antagonists, lacks intrinsic activity at dopamine D 2 receptors in vivo (Natesan et al., 2006) as well as in vitro (Dyhring et al., 2010; Tadori et al., 2007). The functional effects of dopaminergic stabilizers are not confined to aberrations originating from the dopamine system. Pridopidine has also been shown to reverse perturbed behavioural patterns originating from hypoglutamatergic states induced by the N-methyl-D-aspartate (NMDA) receptor antagonist MK-801 (Nilsson et al., 2004; Pettersson et al., 2002; Rung et al., 2005; Waters et al., 2002). In line with this, increased expression of the gene encoding the activity-regulated cytoskeletal protein (Arc) has been observed in rat cortex after acute systemic administration of pridopidine, which is probably the result of synaptic NMDA receptor activation (Waters et al., 2009a). Hence, at the system level, pridopidine is hypothesized to have a dual mechanism of action consisting of both functional modulation of dopamine transmission and activation of cortical glutamate transmission through synaptic NMDA receptors. European Journal of Pharmacology 644 (2010) 88–95 ⁎ Corresponding author. Tel.: + 46 31 7727 743; fax: + 46 31 7727 701. E-mail addresses: [email protected] (H. Ponten), [email protected] (J. Kullingsjö), [email protected] (S. Lagerkvist), [email protected] (P. Martin), [email protected] (F. Pettersson), [email protected] (C. Sonesson), [email protected] (S. Waters), [email protected] (N. Waters). 0014-2999/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2010.07.023 Contents lists available at ScienceDirect European Journal of Pharmacology journal homepage: www.elsevier.com/locate/ejphar
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of In vivo pharmacology of the dopaminergic stabilizer pridopidine

European Journal of Pharmacology 644 (2010) 88–95
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar
Neuropharmacology and Analgesia
In vivo pharmacology of the dopaminergic stabilizer pridopidine
Henrik Ponten a,⁎, Johan Kullingsjö a, Sören Lagerkvist a, Peter Martin b, Fredrik Pettersson a, Clas Sonesson a,Susanna Waters a, Nicholas Waters a
a NeuroSearch Sweden AB, Arvid Wallgrens Backe 20, SE-413 46 Gothenburg, Swedenb Vårdcentralen Lerum, Göteborgsvägen 9, SE-443 25, Lerum, Sweden
⁎ Corresponding author. Tel.: +46 31 7727 743; fax:E-mail addresses: [email protected] (H
[email protected] (J. Kullingsjö), soren.la(S. Lagerkvist), [email protected] (P. Martin), fredrik.pet(F. Pettersson), [email protected] (C. [email protected] (S. Waters), nicholas.w(N. Waters).
0014-2999/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.ejphar.2010.07.023
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 February 2010Received in revised form 18 June 2010Accepted 11 July 2010Available online 24 July 2010
Keywords:PridopidineACR16DopamineDopaminergic stabilizerHuntington's diseasePharmacology
Pridopidine (ACR16) belongs to a new pharmacological class of agents affecting the central nervoussystem called dopaminergic stabilizers. Dopaminergic stabilizers act primarily at dopamine type 2 (D2)receptors and display state-dependent behavioural effects. This article aims to give an overview of thepreclinical neurochemical and behavioural in vivo pharmacological properties of pridopidine. Pridopidinewas given s.c. to male Sprague–Dawley rats (locomotor, microdialysis and tissue neurochemistry) and i.p.to Swiss male mice (tail suspension test). Pridopidine dose-dependently increased striatal tissue levels ofthe dopamine metabolite 3,4-dihydroxyphenylalanin (ED50=81 μmol/kg), and prefrontal cortex dialysatelevels of dopamine and noradrenaline as measured by high performance liquid chromatography. The agentreduced hyperlocomotion (D-amphetamine: ED50=54 μmol/kg; MK-801: ED50=40 μmol/kg), butpreserved spontaneous locomotor activity, confirming state-dependent behavioural effects. In addition,pridopidine significantly reduced immobility time in the tail suspension test. We conclude thatpridopidine state-dependently stabilizes psychomotor activity by the dual actions of functional dopamineD2 receptor antagonism and strengthening of cortical glutamate functions in various settings of perturbedneurotransmission. The putative restoration of function in cortico–subcortical circuitry by pridopidine islikely to make it useful for ameliorating several neurological and psychiatric disorders, includingHuntington's disease.
+46 31 7727 701.. Ponten),[email protected]@neurosearch.seson),[email protected]
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Pridopidine (ACR16; 4-[3-(methylsulfonyl)phenyl]-1-propylpi-peridine hydrochloride; Huntexil®), originally referred to as amodulator of dopamine neurotransmission (Sonesson et al., 2000),belongs to a new pharmacological class of agents acting on the centralnervous system. Through their primary action at dopamine type 2(D2) receptors, these compounds stabilize dysregulated psychomotorfunctions (i.e. they reverse behavioural states originating from bothhypo- and hyper-dopaminergia), while having only subtle effects onnormal psychomotor activity (Natesan et al., 2006; Pettersson et al.,2010; Sonesson et al., 2000). Thus, these compounds regulate bothhyper- and hypoactive functioning in areas of the brain receivingdopamine input, leading to reduced excitatory and inhibitory stress,resulting in improved behavioural performance. These pharmacody-namic actions of this new class (known as dopaminergic stabilizers)
do not appear to fit fully into the current classification of dopamine D2
receptor-acting compounds (Carlsson, 2000; Rung et al., 2008):antagonists, agonists, partial agonists or inverse agonists (Strange,2008).
Pridopidine has been shown to bind to striatal dopamine D2
receptors in vivo, in a dose range where the agent is active in modelsassessing antipsychotic-like efficacy, and, similar to dopamine D2
receptor antagonists, lacks intrinsic activity at dopamine D2 receptorsin vivo (Natesan et al., 2006) as well as in vitro (Dyhring et al., 2010;Tadori et al., 2007). The functional effects of dopaminergic stabilizersare not confined to aberrations originating from the dopaminesystem. Pridopidine has also been shown to reverse perturbedbehavioural patterns originating from hypoglutamatergic statesinduced by the N-methyl-D-aspartate (NMDA) receptor antagonistMK-801 (Nilsson et al., 2004; Pettersson et al., 2002; Rung et al., 2005;Waters et al., 2002). In line with this, increased expression of the geneencoding the activity-regulated cytoskeletal protein (Arc) has beenobserved in rat cortex after acute systemic administration ofpridopidine, which is probably the result of synaptic NMDA receptoractivation (Waters et al., 2009a). Hence, at the system level,pridopidine is hypothesized to have a dual mechanism of actionconsisting of both functional modulation of dopamine transmissionand activation of cortical glutamate transmission through synapticNMDA receptors.

89H. Ponten et al. / European Journal of Pharmacology 644 (2010) 88–95
Current theories implicate aberrant dopamine transmission indisorders such as schizophrenia, Huntington's disease (HD) andParkinson's disease (PD). However, there are suggestions thatglutamate transmission, for example at the NMDA receptor subunitNR2B, might also be involved in these disorders, for review see Loftisand Janowsky (2003). Furthermore, pharmacological therapiestargeting the glutamate system that aim to modulate transmissionat NMDA receptors are hypothesized to be useful in schizophrenia(Kim et al., 2009), PD (Johnson et al., 2009) and HD (Frank andJankovic, 2010). Based on hypotheses regarding the mode of action ofpridopidine and putative treatment opportunities, clinical trials havebeen initiated and show encouraging results in patients with PD(Tedroff et al., 2004), schizophrenia (Carlsson and Carlsson, 2006) andHD (Tedroff et al., 2009). Pridopidine is currently in clinicaldevelopment for the treatment of HD.
The aim of this article is to give an overview of the preclinical invivo pharmacological studies performed during the discovery ofpridopidine. The data have previously been published, in part, inarticle, patent, poster and abstract form (Pettersson et al., 2010;Pettersson et al., 2002; Sonesson et al., 2000; Waters et al., 2002;Waters et al., 2009a).
2. Materials and methods
2.1. Animals
Male Sprague–Dawley rats from B&K Scanbur were used forlocomotor, tissue neurochemistry and microdialysis studies. Ratsweighed 160–180 g at the time of arrival. Rats weighed 220–270 g atthe time of the locomotor and tissue neurochemistry studies and 240–355 g at the time of microdialysis studies. They were housed fiveanimals per cage with lights on between 06:00 h and 18:00 h. Swissmale mice from Depré, France (weighing 19–24 g) were used for thetail suspension tests. All experiments were carried out in accordancewith the European Community guidelines for animal studies and withthe approval of local animal ethics committees.
2.2. Locomotor activity
Locomotor activity was recorded for 60 min in motility metersgenerating a time series of x, y (horizontal activity) and z (verticalactivity) coordinates sampled at 25 Hz. This time series wassubsequently converted into locomotor variables studied at sevenfrequencies from 25 Hz to 0.25 Hz (Ponten et al., 2005). For thestatistical analysis applied in the present article, only the variabledistance, derived from the highest frequency (25 Hz), was used.Pridopidine was administered using the NeuroSearch Sweden ABstandard screening protocol for investigation of novel compounds,consisting of subcutaneous (s.c.) injection 4 min before initiation oflocomotor recordings. D-amphetamine (1.5 mg/kg) and MK-801(0.7 mg/kg) were injected intraperitoneally (i.p.) 6 and 86 min,respectively, before s.c. administration of pridopidine. Our laboratoryhas found that these doses and timings of D-amphetamine and MK-801 administration produce locomotor behavioural patterns with ahigh degree of inter-trial reproducibility. In the habituated-stateexperiments, the animals were placed in the motility meter arena for30 min to allow habituation before s.c. injection of pridopidine andstart of locomotor recordings.
2.3. Ex vivo tissue preparation and analysis
Animals were decapitated 64 min after drug administration andbrains dissected into striatum, cortex and limbic regions (containingthe nucleus accumbens — both the core and shell — most parts of theolfactory tubercle and ventral pallidum). The tissue samples werefrozen immediately and stored at –80 °C before preparation, see
Ponten et al. (2005) for further details. Alpha-methyl-dopa (0.25 μM)was used as internal standard in high performance liquid chroma-tography analysis with electrochemical detection (HPLC/EC) of tissueeluates. The separation of the monoamine transmitter substances(noradrenaline, dopamine, 5-hydroxytryptamine [5-HT]) as well asthe amine metabolite, 3-methoxytyramine (3-MT) and acid metabo-lites (3,4-dihydroxyphenylalanine [DOPAC], 5-hydrocyindoleaceticacid [5-HIAA], homovanillic acid [HVA]) was performed on a reversephase column (Nucleosil ODS, 5 μm, 150×4.6 mm internal diameter)with an aqueous mobile phase (1 ml/min) containing 40 mM citricacid, 12 mMK2HPO4, 1.7 mMH3PO4, 0.35 mM sodium octylsulfate, 6%(volume/volume) of methanol and 0.05 mM EDTA.
2.4. Microdialysis
Experiments were conducted in accordance with Waters et al.(1993), with some modifications as described below. For experi-ments performed in the dorsal striatum, prefrontal cortex andnucleus accumbens, probes with an exposed length of 3, 2.5 and2 mm of the dialysis membrane, respectively, were used. Coordi-nates were calculated relative to bregma: prefrontal cortex(antero-posterior, +3.2 mm from bregma; medio-lateral, 1.2 mmfrom midline; and dorsal–ventral, –4.0 mm from the surface ofskull), nucleus accumbens (antero-posterior, +1.85 mm; medio-lateral, 1.6 mm; dorsal–ventral, –7.8 mm) or dorsal striatum(antero-posterior +1.0 mm; medio-lateral, 2.6 mm; dorsal–ven-tral, –6.2 mm) according to Paxinos and Watson (1986). The probedirected at the prefrontal cortex was angled 8° from the mid-sagittal plane. Before insertion into the brain, the dialysis probeswere perfused with 0.6 ml ethanol 70% and 1 ml of Ringer solutionconsecutively. The rats were housed individually in spherical cagesfor 36 to 48 h before the dialysis experiments. The brains wereremoved rapidly and fixed in Neo-fix solution (Kebo-lab, Sweden).Determination of dialysate content of dopamine, noradrenaline, 5-HT, DOPAC, HVA, 3-MT and 5-HIAA was performed in accordancewith the HPLC/EC system described in Ponten et al. (2005).
2.5. Tail suspension test
Four groups of 10mice eachwere used for tail suspension tests. Thefirst group received saline vehicle, the second desipramine 10 mg/kg(as method control substance), and groups three and four receivedpridopidine (10 μmol/kg and 40 μmol/kg, respectively). Given that theinitial pharmacokinetic and pharmacological exploration of pridopi-dine in mice was performed using i.p. administration, each agent wasalso administered i.p. for the tail suspension test. The mice wereobserved at about 27 min after dosing to confirm that treatment hadnomarked effect on normal behaviour and at 30 min after dosing weresuspended by their tail in the cage. The sum of immobility periods wasrecorded over a period of 6 min. Experiments were performed byCentre de Recherches Biologiques (CERB), France.
2.6. Data analysis
Data are expressed as mean±S.E.M. percentage of vehicle-treatedgroup in thenormal state and thehabituated state, and asmean±S.E.M.percentage of the D-amphetamine or MK-801 pretreated controls inthe D-amphetamine and MK-801 state experiments, respectively. TheED50 values for DOPAC in striatum and locomotor activity werecalculated by curve fitting and fitted by least square minimization tothe function: End–(End–Control)/(1+(dose/ED50)Slope), where Controlis control level and End is end level. The DOPAC levels were normalizedto control (within eachexperiment), ED50 and slopewerefittedwith therestrictions: ED50N0, 0.5bSlopeb3, End=350% of control. For ED50
value calculation of pridopidine-induced reduction of spontaneous ordrug-induced hyperlocomotion by D-amphetamine or MK-801,

Table 1Ex vivo brain monoaminergic biochemistry in rats given various doses of pridopidine.
Analyte Dose ofpridopidine(μmol/kg)
Limbic region Striatum Cortex
% ofvehicle
S.E.M. % ofvehicle
S.E.M. % ofvehicle
S.E.M.
Noradrenaline 3.7 96.2 4.0 108.2 3.8 91.0b 1.211 96.4 2.2 104.3 11.7 96.6 4.433 88.6 5.0 96.3 4.8 90.4b 2.0
100 93.8 2.8 90.8 4.5 80.8c 1.5300 74.9c 5.1 96.7 7.0 61.8c 3.5
Dopamine 3.7 97.1 6.3 88.4 4.0 95.4 25.311 98.6 4.8 87.0a 3.1 154.1 39.433 102.0 5.2 93.5 2.1 121.4 11.3
100 88.7 3.2 83.7b 2.3 86.7 5.7300 75.5c 1.1 70.9c 1.9 64.8b 4.3
DOPAC 3.7 97.7 4.9 95.2 2.1 89.7 17.5
90 H. Ponten et al. / European Journal of Pharmacology 644 (2010) 88–95
distances were normalized to vehicle, D-amphetamine or MK-801controls, respectively, and fitted with the restrictions: ED50N0,0.5bSlopeb3, End=0% of control. In addition, statistical significancewas assessed versus appropriate control groups using the Student's t-test. ED50 and t-tests were calculated using Excel (Microsoft).
Microdialysis data are expressed as the mean±S.E.M. percentageof baseline. Baseline was defined as the mean of the three fractionsimmediately before administration of test compound. Data wereanalysed using Statview (SAS Institute) by repeated measuresanalysis of variance (ANOVA), and if significant effects were found,followed by Fischer's PLSD post hoc test.
Total immobility time in the tail suspension tests are expressed asmedians with ranges and were analysed by the non-parametricMann–Whitney U test.
For all tests, Pb0.05 was considered significant.
11 102.3 4.5 101.3 5.0 117.3 8.733 150.9c 7.0 161.4c 3.9 136.6c 6.5100 204.4c 10.1 242.6c 10.6 167.0c 9.8300 255.8c 2.5 297.9c 7.8 201.6c 12.0
HVA 3.7 103.6 5.0 93.2 1.1 91.7 1.711 121.2b 4.7 112.5 7.5 113.2 6.433 167.9c 4.0 172.1c 4.5 142.9c 5.1
100 238.4c 7.5 245.3c 8.3 185.2c 7.9300 283.2c 6.2 283.1c 8.1 222.1c 9.3
3-MT 3.7 127.7a 4.8 109.8 19.8 n.d. n.d.11 108.4 9.3 97.5 22.0 n.d. n.d.33 124.7 9.0 105.2 11.3 n.d. n.d.
100 109.9 4.7 95.1 9.8 n.d. n.d.300 83.4 11.8 67.9a 6.6 n.d. n.d.
2.7. Drugs
Pridopidine HCl salt (molecular weight 317.88) was synthesizedinternally at NeuroSearch Sweden AB. MK-801 (dizocilpine) anddesipramine HCl was obtained from Sigma-Aldrich. D-amphetaminewas obtained from AKL (Sweden). All compounds were dissolved inphysiological saline (0.9 wt.%/volume NaCl) and injected in a volumeof 5 ml/kg, with the exception of the tail suspension tests in whichagents were injected in a volume of 10 ml/kg.
5-HT 3.7 93.7 2.9 94.2 5.3 92.1 3.911 96.9 2.2 95.5 2.9 98.2 0.933 95.6 3.1 91.1 2.7 95.2 2.5
100 95.1 1.9 88.2a 3.1 93.6 2.3300 94.4 1.9 89.3a 2.6 97.6 3.0
5-HIAA 3.7 103.0 3.4 102.2 3.3 100.3 1.511 102.9 2.3 103.3 4.8 97.1 3.533 106.7 4.1 102.7 3.2 101.5 3.1
100 110.7a 3.1 105.9 2.8 105.9 3.2300 109.6 3.2 106.4 1.2 106.9 2.6
3-MT, 3-methoxytyramine; 5-HIAA, 5-hydroxyindoleacetic acid; 5-HT, 5-hydroxytryptamine; DOPAC, 3,4-dihydroxyphenylalanine; HVA, homovanillic acid; n.d., not determined. aPb0.05, bPb0.01 and cPb0.001.
3. Results
3.1. Ex vivo brain monoaminergic biochemistry
The main effects observed in rats were dose-dependent increasesof the dopamine metabolites DOPAC and HVA in the cortical, striataland limbic regions (Fig. 1; Table 1). In the striatum, DOPAC increasedto approximately 300% of the vehicle control at 300 μmol/kg s.c. TheED50 value for increase in striatal DOPAC was calculated to be81 μmol/kg of pridopidine (Table 2). Pridopidine induced decreases indopamine, noradrenaline and 5-HT in ex vivo brain tissue. Thedecreases in 5-HT were less prominent and reached significanceonly in the striatum, where it decreased to 88% of vehicle group at100 μmol/kg s.c. (Table 1).
Fig. 1. Effects of pridopidine on DOPAC. Pridopidine increases 3,4-dihydroxyphenyla-lanine (DOPAC) in the limbic forebrain, striatum and cortex of male Sprague–Dawleyrats with no pretreatment. Doses are given in μmol/kg and were administered s.c.64 min before sacrificing the animals. Data are mean±S.E.M. ***Pb0.001 vs controls.Number of animals per dose treatment group: vehicle: n=9; 3.7 and 11 μmol/kg:n=4; 33 and 100 μmol/kg: n=9; 300 μmol/kg: n=5.
3.2. Microdialysis
The main effects of pridopidine were dose-dependent increases inthe extracellular levels of dopamine, noradrenaline, DOPAC, HVA, 3-MT and 5-HIAA in the nucleus accumbens, dorsal striatum andprefrontal cortex; by contrast, 5-HT increased significantly (up to126% of the vehicle control) only in prefrontal cortex (Fig. 2, Table 3).At 50 μmol/kg of pridopidine, dopamine increased significantly to194%, 173% and 212% of the vehicle control, and noradrenaline to157%, 300% and 209% of the vehicle control in the nucleus accumbens,dorsal striatum and prefrontal cortex, respectively.
Table 2Summary of ED50 calculations for increase in striatal 3,4-dihydroxyphenylalanine, andfor reductions in drug-induced hyperactivity and spontaneous activity.
Neurochemical/behaviouralvariable in rat
Pridopidine ED50
(μmol/kg s.c.)95% confidenceinterval
Increase of DOPAC in striatum 81 71–99Reduction of D-amphetaminehyperactivity (15–60 min)
54 33–72
Reduction of MK-801hyperactivity (15–60 min)
40 14–65
Reduction of spontaneouslocomotor activity (15–60 min)
n.c. (no reduction) n.c. (no reduction)
Reduction of spontaneouslocomotor activity (0–15 min)
318 248–566
DOPAC, 3,4-dihydroxyphenylalanine; n.c., not calculated.
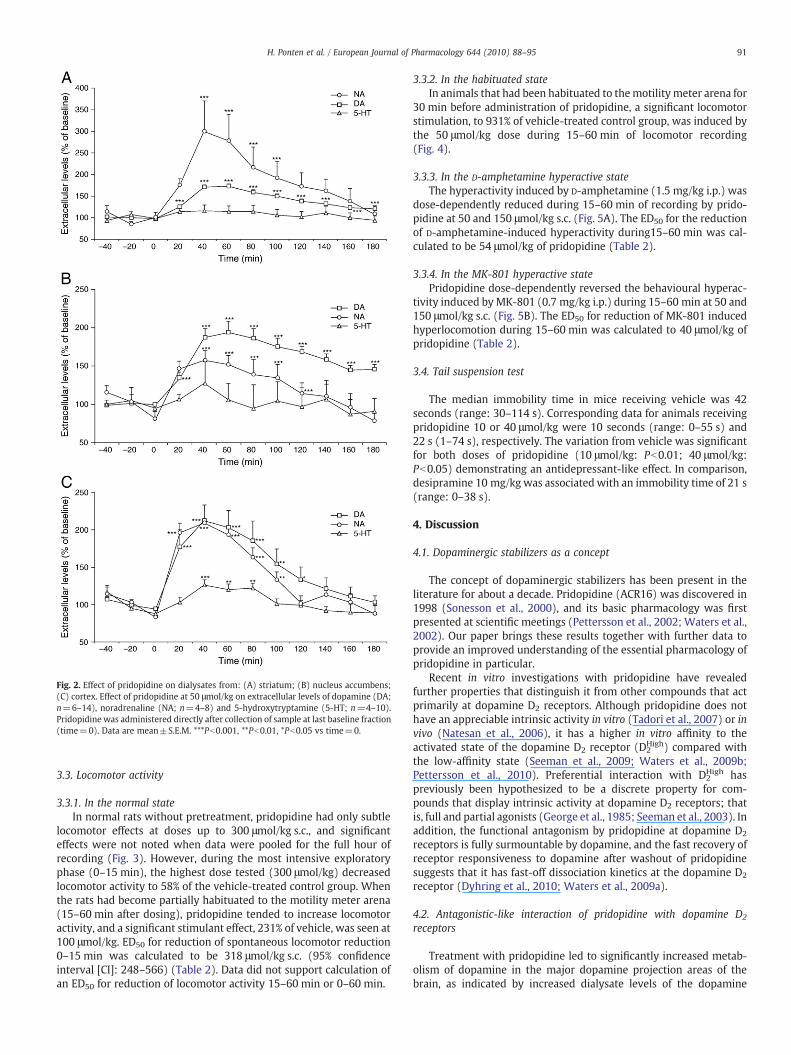
Fig. 2. Effect of pridopidine on dialysates from: (A) striatum; (B) nucleus accumbens;(C) cortex. Effect of pridopidine at 50 μmol/kg on extracellular levels of dopamine (DA;n=6–14), noradrenaline (NA; n=4–8) and 5-hydroxytryptamine (5-HT; n=4–10).Pridopidine was administered directly after collection of sample at last baseline fraction(time=0). Data are mean±S.E.M. ***Pb0.001, **Pb0.01, *Pb0.05 vs time=0.
91H. Ponten et al. / European Journal of Pharmacology 644 (2010) 88–95
3.3. Locomotor activity
3.3.1. In the normal stateIn normal rats without pretreatment, pridopidine had only subtle
locomotor effects at doses up to 300 μmol/kg s.c., and significanteffects were not noted when data were pooled for the full hour ofrecording (Fig. 3). However, during the most intensive exploratoryphase (0–15 min), the highest dose tested (300 μmol/kg) decreasedlocomotor activity to 58% of the vehicle-treated control group. Whenthe rats had become partially habituated to the motility meter arena(15–60 min after dosing), pridopidine tended to increase locomotoractivity, and a significant stimulant effect, 231% of vehicle, was seen at100 μmol/kg. ED50 for reduction of spontaneous locomotor reduction0–15 min was calculated to be 318 μmol/kg s.c. (95% confidenceinterval [CI]: 248–566) (Table 2). Data did not support calculation ofan ED50 for reduction of locomotor activity 15–60 min or 0–60 min.
3.3.2. In the habituated stateIn animals that had been habituated to themotility meter arena for
30 min before administration of pridopidine, a significant locomotorstimulation, to 931% of vehicle-treated control group, was induced bythe 50 μmol/kg dose during 15–60 min of locomotor recording(Fig. 4).
3.3.3. In the D-amphetamine hyperactive stateThe hyperactivity induced by D-amphetamine (1.5 mg/kg i.p.) was
dose-dependently reduced during 15–60 min of recording by prido-pidine at 50 and 150 μmol/kg s.c. (Fig. 5A). The ED50 for the reductionof D-amphetamine-induced hyperactivity during15–60 min was cal-culated to be 54 μmol/kg of pridopidine (Table 2).
3.3.4. In the MK-801 hyperactive statePridopidine dose-dependently reversed the behavioural hyperac-
tivity induced by MK-801 (0.7 mg/kg i.p.) during 15–60 min at 50 and150 μmol/kg s.c. (Fig. 5B). The ED50 for reduction of MK-801 inducedhyperlocomotion during 15–60 min was calculated to 40 μmol/kg ofpridopidine (Table 2).
3.4. Tail suspension test
The median immobility time in mice receiving vehicle was 42seconds (range: 30–114 s). Corresponding data for animals receivingpridopidine 10 or 40 μmol/kg were 10 seconds (range: 0–55 s) and22 s (1–74 s), respectively. The variation from vehicle was significantfor both doses of pridopidine (10 μmol/kg: Pb0.01; 40 μmol/kg:Pb0.05) demonstrating an antidepressant-like effect. In comparison,desipramine 10 mg/kg was associated with an immobility time of 21 s(range: 0–38 s).
4. Discussion
4.1. Dopaminergic stabilizers as a concept
The concept of dopaminergic stabilizers has been present in theliterature for about a decade. Pridopidine (ACR16) was discovered in1998 (Sonesson et al., 2000), and its basic pharmacology was firstpresented at scientific meetings (Pettersson et al., 2002; Waters et al.,2002). Our paper brings these results together with further data toprovide an improved understanding of the essential pharmacology ofpridopidine in particular.
Recent in vitro investigations with pridopidine have revealedfurther properties that distinguish it from other compounds that actprimarily at dopamine D2 receptors. Although pridopidine does nothave an appreciable intrinsic activity in vitro (Tadori et al., 2007) or invivo (Natesan et al., 2006), it has a higher in vitro affinity to theactivated state of the dopamine D2 receptor (D2
High) compared withthe low-affinity state (Seeman et al., 2009; Waters et al., 2009b;Pettersson et al., 2010). Preferential interaction with D2
High haspreviously been hypothesized to be a discrete property for com-pounds that display intrinsic activity at dopamine D2 receptors; thatis, full and partial agonists (George et al., 1985; Seeman et al., 2003). Inaddition, the functional antagonism by pridopidine at dopamine D2
receptors is fully surmountable by dopamine, and the fast recovery ofreceptor responsiveness to dopamine after washout of pridopidinesuggests that it has fast-off dissociation kinetics at the dopamine D2
receptor (Dyhring et al., 2010; Waters et al., 2009a).
4.2. Antagonistic-like interaction of pridopidine with dopamine D2
receptors
Treatment with pridopidine led to significantly increased metab-olism of dopamine in the major dopamine projection areas of thebrain, as indicated by increased dialysate levels of the dopamine
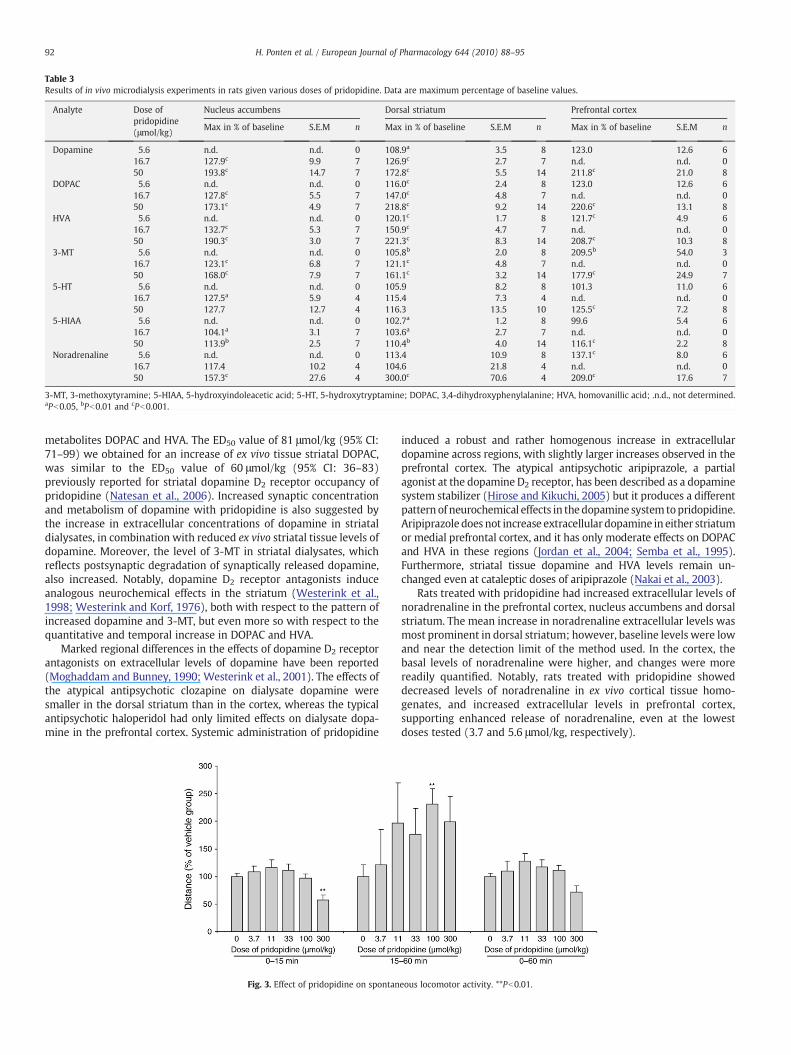
Table 3Results of in vivo microdialysis experiments in rats given various doses of pridopidine. Data are maximum percentage of baseline values.
Analyte Dose ofpridopidine(μmol/kg)
Nucleus accumbens Dorsal striatum Prefrontal cortex
Max in % of baseline S.E.M n Max in % of baseline S.E.M n Max in % of baseline S.E.M n
Dopamine 5.6 n.d. n.d. 0 108.9a 3.5 8 123.0 12.6 616.7 127.9c 9.9 7 126.9c 2.7 7 n.d. n.d. 050 193.8c 14.7 7 172.8c 5.5 14 211.8c 21.0 8
DOPAC 5.6 n.d. n.d. 0 116.0c 2.4 8 123.0 12.6 616.7 127.8c 5.5 7 147.0c 4.8 7 n.d. n.d. 050 173.1c 4.9 7 218.8c 9.2 14 220.6c 13.1 8
HVA 5.6 n.d. n.d. 0 120.1c 1.7 8 121.7c 4.9 616.7 132.7c 5.3 7 150.9c 4.7 7 n.d. n.d. 050 190.3c 3.0 7 221.3c 8.3 14 208.7c 10.3 8
3-MT 5.6 n.d. n.d. 0 105.8b 2.0 8 209.5b 54.0 316.7 123.1c 6.8 7 121.1c 4.8 7 n.d. n.d. 050 168.0c 7.9 7 161.1c 3.2 14 177.9c 24.9 7
5-HT 5.6 n.d. n.d. 0 105.9 8.2 8 101.3 11.0 616.7 127.5a 5.9 4 115.4 7.3 4 n.d. n.d. 050 127.7 12.7 4 116.3 13.5 10 125.5c 7.2 8
5-HIAA 5.6 n.d. n.d. 0 102.7a 1.2 8 99.6 5.4 616.7 104.1a 3.1 7 103.6a 2.7 7 n.d. n.d. 050 113.9b 2.5 7 110.4b 4.0 14 116.1c 2.2 8
Noradrenaline 5.6 n.d. n.d. 0 113.4 10.9 8 137.1c 8.0 616.7 117.4 10.2 4 104.6 21.8 4 n.d. n.d. 050 157.3c 27.6 4 300.0c 70.6 4 209.0c 17.6 7
3-MT, 3-methoxytyramine; 5-HIAA, 5-hydroxyindoleacetic acid; 5-HT, 5-hydroxytryptamine; DOPAC, 3,4-dihydroxyphenylalanine; HVA, homovanillic acid; .n.d., not determined.aPb0.05, bPb0.01 and cPb0.001.
92 H. Ponten et al. / European Journal of Pharmacology 644 (2010) 88–95
metabolites DOPAC and HVA. The ED50 value of 81 μmol/kg (95% CI:71–99) we obtained for an increase of ex vivo tissue striatal DOPAC,was similar to the ED50 value of 60 μmol/kg (95% CI: 36–83)previously reported for striatal dopamine D2 receptor occupancy ofpridopidine (Natesan et al., 2006). Increased synaptic concentrationand metabolism of dopamine with pridopidine is also suggested bythe increase in extracellular concentrations of dopamine in striataldialysates, in combination with reduced ex vivo striatal tissue levels ofdopamine. Moreover, the level of 3-MT in striatal dialysates, whichreflects postsynaptic degradation of synaptically released dopamine,also increased. Notably, dopamine D2 receptor antagonists induceanalogous neurochemical effects in the striatum (Westerink et al.,1998; Westerink and Korf, 1976), both with respect to the pattern ofincreased dopamine and 3-MT, but even more so with respect to thequantitative and temporal increase in DOPAC and HVA.
Marked regional differences in the effects of dopamine D2 receptorantagonists on extracellular levels of dopamine have been reported(Moghaddam and Bunney, 1990; Westerink et al., 2001). The effects ofthe atypical antipsychotic clozapine on dialysate dopamine weresmaller in the dorsal striatum than in the cortex, whereas the typicalantipsychotic haloperidol had only limited effects on dialysate dopa-mine in the prefrontal cortex. Systemic administration of pridopidine
Fig. 3. Effect of pridopidine on spontan
induced a robust and rather homogenous increase in extracellulardopamine across regions, with slightly larger increases observed in theprefrontal cortex. The atypical antipsychotic aripiprazole, a partialagonist at the dopamine D2 receptor, has been described as a dopaminesystem stabilizer (Hirose and Kikuchi, 2005) but it produces a differentpattern of neurochemical effects in thedopamine system topridopidine.Aripiprazole does not increase extracellular dopamine in either striatumor medial prefrontal cortex, and it has only moderate effects on DOPACand HVA in these regions (Jordan et al., 2004; Semba et al., 1995).Furthermore, striatal tissue dopamine and HVA levels remain un-changed even at cataleptic doses of aripiprazole (Nakai et al., 2003).
Rats treated with pridopidine had increased extracellular levels ofnoradrenaline in the prefrontal cortex, nucleus accumbens and dorsalstriatum. The mean increase in noradrenaline extracellular levels wasmost prominent in dorsal striatum; however, baseline levels were lowand near the detection limit of the method used. In the cortex, thebasal levels of noradrenaline were higher, and changes were morereadily quantified. Notably, rats treated with pridopidine showeddecreased levels of noradrenaline in ex vivo cortical tissue homo-genates, and increased extracellular levels in prefrontal cortex,supporting enhanced release of noradrenaline, even at the lowestdoses tested (3.7 and 5.6 μmol/kg, respectively).
eous locomotor activity. **Pb0.01.

Fig. 4. Effect of pridopidine in habituated animals. **Pb0.01.
93H. Ponten et al. / European Journal of Pharmacology 644 (2010) 88–95
The effects on the 5-HT system were qualitatively similar but oflimited magnitude. The atypical antipsychotic olanzapine inducesregional effects on extracellular dopamine, noradrenaline and 5-HTlevels (Li et al., 1998) that are in general similar to those induced bypridopidine in the present study. Indeed, the positive results obtainedwith pridopidine in the tail suspension test may be related to theseincreases in extracellular dopamine and noradenaline. Dopaminergicand noradrenergic agents, including the dual-acting noradrenalineand dopamine re-uptake inhibitors, have demonstrated antidepres-sant activity in the absence of any serotonin function, by treatingsymptoms associatedwith a reduced positive affect (Nutt et al., 2007).With respect to the somewhat similar neurochemical effects induced
Fig. 5. Effect of pridopidine on hyperlocomotoractivity induced by (A) D-amphetamineand (B) MK-801. *Pb0.05, **Pb0.01, ***Pb0.001.
by olanzapine and pridopidine, it is noteworthy that olanzapine isused clinically, in combination with fluoxetine, for the acutetreatment of depressive episodes associated with bipolar I disorderand treatment-resistant major depressive disorder (Croxtall andScott, 2010).
4.3. Behavioural stabilizing effect of pridopidine and the putative role ofdopamine D2 receptor interactions
Environmental stimuli, such as a novel cage, induce an increase ofboth cortical and subcortical extracellular dopamine (Feenstra et al.,2000). In our normal state experiments, the rats were introduced to anovel cage (the motility meter arenas) 4 min after pridopidineadministration. Hence, during the first minutes of these normalstate experiments, locomotor activity and extracellular dopaminewould both be expected to be elevated compared with rats that arehabituated to their environment. In the habituated-state experiments,pridopidine induced a moderate but definite stimulation of motoractivity, whereas stimulation of motor activity was not evident duringthe first 15 min of the normal state experiments. Furthermore, inpreclinical models of hyperlocomation, due to either hyper-dopami-nergia or hypoglutamatergia, pridopidine displayed dose-dependentreductions of locomotor activity. Hence, the effects of pridopidinewere state dependent (see also (Natesan et al., 2006; Pettersson et al.,2010)).
The behaviour stabilizing effect of pridopidine is not explained solelyby the specific interaction with dopamine D2 receptors. In vitro data oninteractions of pridopidine at the receptor level, such as surmountableand fast-off receptor kinetics, may account for the lack of reduction ofspontaneous locomotor activity over a wide dose range. The increase inmotor activity in habituated animalsmay also be regarded as aneffect ofthe blockade of the dopamine D2
High receptors on the presynaptic site,leading to moderately increased levels of dopamine in the synapse,subsequently stimulating postsynaptic dopamine D2 receptors andinducing an increase in motor activity, provided that dopamine D2
receptor affinity is not too high (see (Pettersson et al., 2010)). However,because data support activation of dopamine, noradrenaline andglutamate systems in the cortex by pridopidine, interactions withthese systems may underlie the differences in vivo versus typical,neuroleptic-like dopamineD2 receptor antagonists. Pridopidine, but notthedopamineD2 receptor antagonist haloperidol, significantly inhibitedamphetamine-inducedhyperactivity in dopamineD2 receptor knockoutmice (Svensson et al., 2009). Hence, the reductions of locomotor activityin the hyperactive states could be explained, in part, by the corticalactivation profile displayed by pridopidine. An increase in corticalnoradrenaline (e.g. by alpha2 adrenoceptor blockade) has beensuggested to augment prefrontal dopamine functioning (Svensson,

94 H. Ponten et al. / European Journal of Pharmacology 644 (2010) 88–95
2003) and adjunctive treatment with the alpha2 adrenoceptor antag-onist idazoxan improves antipsychotic efficacy in preclinical models(Hertel et al., 1999; Marcus et al., 2009) compared with dopamine D2
receptor antagonists alone. In vitro, the combination of idazoxan withthe dopamine D2 receptor antagonist raclopride has been shown tocause a dopamine D1 receptor-mediated facilitation of glutamatetransmission in rat prefrontal pyramidal cells (Marcus et al., 2005).
Increased expression of Arc mRNA in rat cortex after administra-tion of pridopidine, interpreted as increased synaptic glutamateactivation of ionotropic NMDA receptors (Waters et al., 2009a),could possibly be dependent primarily on the cortical increase indopamine and noradrenaline. Increased cortical activation wouldstrengthen cortical–striatal feedback regulation of psychomotoractivity (Alexander et al., 1986), leading to behavioural normalizationof hyperactive states (Tekin and Cummings, 2002). Glutamateinfluence on psychomotor functions in rodents is supported by datashowing that LY404039, a glutamate receptor mGlu2/3 agonist,inhibits D-amphetamine-induced hyperactivity without sedativeeffects (Rorick-Kehn et al., 2007) — a similar effect to that seen withpridopidine.
Taken together, these results suggest that the psychomotorstabilizing effects of pridopidine may be dependent on its dualeffects: the functional antagonism of subcortical dopamine transmis-sion and strengthening of cortical glutamate transmission. Theproperties of dopaminergic stabilizers should therefore be regardedas an in vivo phenomenon, dependent on the effects in the cortico–subcortical brain circuitries not revealed at the receptor level in vitro.
4.4. Clinical implications of the pharmacological profile
The preclinical profile of pridopidine includes the potential tostabilize motor function not only in hyperactivity, but also in states ofhypoactivity. Furthermore, pridopidine displays antidepressant po-tential, as indicated by the tail suspension test data. It has beensuggested that facilitation of prefrontal cortical dopamine levels,leading to stimulation of dopamine D1 receptors, could attenuatesome cognitive impairments associated with deficits in NMDAreceptor function (Krystal et al., 2005). Hence the increasedextracellular concentrations of dopamine in the prefrontal cortexinduced by pridopidine, in combination with the putative enhancedactivation by pridopidine of synaptic NMDA receptors in the cortex,imply it could be beneficial for restoring impaired cortical activity.Indeed, pridopidine has shown beneficial effects in NMDA receptorantagonist rat models of cognitive and negative symptoms ofschizophrenia (Nilsson et al., 2004; Rung et al., 2005). Huntington'sdisease leads to damage of neurons in both the striatum and thecortex, causing regional hypo- and hyperactivity in the dopaminesystems. Symptoms of HD include severe motor disturbances (lack ofvoluntarymovements as well as presence of involuntarymovements),cognitive impairment and psychiatric characteristics, includingpsychosis and depression. Thus, the in vivo pharmacological-effectprofile of pridopidine suggests it may have significant benefits on themain symptom domains in HD. In an early study in HD, pridopidinewas associated with improvements in voluntary motor performance,with no change in involuntary movement (Tedroff et al., 2009).
Acknowledgements
Ms. Elisabeth Ljung, Ms. Marianne Thorngren andMs. LenaWollterat NeuroSearch Sweden, and Ms. Kirsten Sönniksen, previously atNeuroSearch Sweden are greatly appreciated for expert technicalassistance in the in vivo neurochemical and behavioural studies. CERB,France, performed the tail suspension studies. Oxford PharmaGen-esis™ Ltd provided editorial support to the authors.
References
Alexander, G.E., Delong, M.R., Strick, P.L., 1986. Parallel organization of functionallysegregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 9,357–381.
Carlsson, A., 2000. Focusing on dopaminergic stabilzers and 5HT 2A receptorantagonists. Curr. Opin. CPNS. Investigat. Drugs 2, 22–24.
Carlsson, A., Carlsson, M.L., 2006. A dopaminergic deficit hypothesis of schizophrenia:the path to discovery. Dialogues Clin. Neurosci. 8, 137–142.
Croxtall, J.D., Scott, L.J., 2010. Olanzapine/fluoxetine: a review of its use in patients withtreatment-resistant major depressive disorder. CNS Drugs 24, 245–262.
Dyhring, T., Nielsen, E.O., Sonesson, C., Pettersson, F., Karlsson, J., Svensson, P.,Christophersen, P., Waters, N., 2010. The dopaminergic stabilizers pridopidine(ACR16) and (−)-OSU6162 display dopamine D(2) receptor antagonism and fastreceptor dissociation properties. Eur. J. Pharmacol. 628, 19–26.
Feenstra, M.G., Botterblom, M.H., Mastenbroek, S., 2000. Dopamine and noradrenalineefflux in the prefrontal cortex in the light and dark period: effects of novelty andhandling and comparison to the nucleus accumbens. Neuroscience 100, 741–748.
Frank, S., Jankovic, J., 2010. Advances in the pharmacological management ofHuntington's disease. Drugs 70, 561–571.
George, S.R., Watanabe, M., Di Paolo, T., Falardeau, P., Labrie, F., Seeman, P., 1985. Thefunctional state of the dopamine receptor in the anterior pituitary is in the highaffinity form. Endocrinology 117, 690–697.
Hertel, P., Fagerquist, M.V., Svensson, T.H., 1999. Enhanced cortical dopamine outputand antipsychotic-like effects of raclopride by alpha2 adrenoceptor blockade.Science 286, 105–107.
Hirose, T., Kikuchi, T., 2005. Aripiprazole, a novel antipsychotic agent: dopamine D2receptor partial agonist. J. Med. Invest. 52 (Suppl), 284–290.
Johnson, K.A., Conn, P.J., Niswender, C.M., 2009. Glutamate receptors as therapeutictargets for Parkinson's disease. CNS Neurol. Disord. Drug Targets 8, 475–491.
Jordan, S., Koprivica, V., Dunn, R., Tottori, K., Kikuchi, T., Altar, C.A., 2004. In vivo effectsof aripiprazole on cortical and striatal dopaminergic and serotonergic function. Eur.J. Pharmacol. 483, 45–53.
Kim, D.H., Maneen, M.J., Stahl, S.M., 2009. Building a better antipsychotic: receptortargets for the treatment of multiple symptom dimensions of schizophrenia.Neurotherapeutics 6, 78–85.
Krystal, J.H., Perry Jr., E.B., Gueorguieva, R., Belger, A., Madonick, S.H., Abi-Dargham, A.,Cooper, T.B., Macdougall, L., Abi-Saab, W., D'Souza, D.C., 2005. Comparative andinteractive human psychopharmacologic effects of ketamine and amphetamine:implications for glutamatergic and dopaminergic model psychoses and cognitivefunction. Arch. Gen. Psychiatry 62, 985–994.
Li, X.M., Perry, K.W., Wong, D.T., Bymaster, F.P., 1998. Olanzapine increases in vivodopamine and norepinephrine release in rat prefrontal cortex, nucleus accumbensand striatum. Psychopharmacology (Berl) 136, 153–161.
Loftis, J.M., Janowsky, A., 2003. The N-methyl-D-aspartate receptor subunit NR2B:localization, functional properties, regulation, and clinical implications. Pharmacol.Ther. 97, 55–85.
Marcus, M.M., Jardemark, K.E., Wadenberg, M.L., Langlois, X., Hertel, P., Svensson, T.H.,2005. Combined alpha2 and D2/3 receptor blockade enhances cortical glutama-tergic transmission and reverses cognitive impairment in the rat. Int. J.Neuropsychopharmacol. 8, 315–327.
Marcus, M.M., Wiker, C., Franberg, O., Konradsson-Geuken, A., Langlois, X., Jardemark,K., Svensson, T.H., 2009. Adjunctive alpha2-adrenoceptor blockade enhances theantipsychotic-like effect of risperidone and facilitates cortical dopaminergic andglutamatergic, NMDA receptor-mediated transmission. Int. J. Neuropsychophar-macol. 1–13 (Oct 19 [Epub ahead of print]).
Moghaddam, B., Bunney, B.S., 1990. Acute effects of typical and atypical antipsychoticdrugs on the release of dopamine from prefrontal cortex, nucleus accumbens, andstriatum of the rat: an in vivo microdialysis study. J. Neurochem. 54, 1755–1760.
Nakai, S., Hirose, T., Uwahodo, Y., Imaoka, T., Okazaki, H., Miwa, T., Nakai, M., Yamada, S.,Dunn, B., Burris, K.D., Molinoff, P.B., Tottori, K., Altar, C.A., Kikuchi, T., 2003.Diminished catalepsy and dopamine metabolism distinguish aripiprazole fromhaloperidol or risperidone. Eur. J. Pharmacol. 472, 89–97.
Natesan, S., Svensson, K.A., Reckless, G.E., Nobrega, J.N., Barlow, K.B., Johansson, A.M.,Kapur, S., 2006. The dopamine stabilizers (S)-(−)-(3-methanesulfonyl-phenyl)-1-propyl-piperidine [(−)-OSU6162] and 4-(3-methanesulfonylphenyl)-1-propyl-piperidine (ACR16) show high in vivo D2 receptor occupancy, antipsychotic-likeefficacy, and low potential for motor side effects in the rat. J. Pharmacol. Exp. Ther.318, 810–818.
Nilsson, M., Carlsson, A., Markinhuhta, K.R., Sonesson, C., Pettersson, F., Gullme, M.,Carlsson, M.L., 2004. The dopaminergic stabiliser ACR16 counteracts the behaviouralprimitivization induced by the NMDA receptor antagonist MK-801 in mice:implications for cognition. Prog.Neuropsychopharmacol. Biol. Psychiatry28, 677–685.
Nutt, D., Demyttenaere, K., Janka, Z., Aarre, T., Bourin, M., Canonico, P.L., Carrasco, J.L.,Stahl, S., 2007. The other face of depression, reduced positive affect: the role ofcatecholamines in causation and cure. J. Psychopharmacol. 21, 461–471.
Paxinos, G., Watson, C., 1986. The Rat Brain in Stereotaxic Coordinates, second ed.Academic Press, New York, NY.
Pettersson, F., Ponten, H., Waters, N., Waters, S., Sonesson, C., 2010. Synthesis andevaluation of a set of 4-phenylpiperidines and 4-phenylpiperazines as D2 receptorligands and the discovery of the dopaminergic stabilizer 4-[3-(methylsulfonyl)phenyl]-1-propylpiperidine (huntexil, pridopidine, ACR16). J. Med. Chem. 53,2510–2520.
Pettersson, F., Waters, N.,Waters, E.S., Carlsson, A., Sonesson, C., 2002. The developmentof a new class of dopamine stabilizers. Poster presented at the Society forNeuroscience Annual Conference, Orlando, FL, USA.

95H. Ponten et al. / European Journal of Pharmacology 644 (2010) 88–95
Ponten, H., Sonniksen, K., Abrahamsson, T., Waters, N., Gustafsson, B., Hanse, E., Groc, L.,2005. Behavioral and neurochemical repercussions of hippocampal networkactivity blockade during the neonatal period. Brain Res. Dev. Brain Res. 155, 81–86.
Rorick-Kehn, L.M., Johnson, B.G., Knitowski, K.M., Salhoff, C.R., Witkin, J.M., Perry, K.W.,Griffey, K.I., Tizzano, J.P., Monn, J.A., McKinzie, D.L., Schoepp, D.D., 2007. In vivopharmacological characterization of the structurally novel, potent, selectivemGlu2/3 receptor agonist LY404039 in animal models of psychiatric disorders.Psychopharmacology (Berl) 193, 121–136.
Rung, J.P., Carlsson, A., Markinhuhta, K.R., Carlsson, M.L., 2005. The dopaminergicstabilizers (−)-OSU6162 and ACR16 reverse (+)-MK-801-induced social with-drawal in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 29, 833–839.
Rung, J.P., Rung, E., Helgeson, L., Johansson, A.M., Svensson, K., Carlsson, A., Carlsson, M.L.,2008. Effects of (−)-OSU6162 andACR16onmotor activity in rats, indicating a uniquemechanism of dopaminergic stabilization. J. Neural Transm. 115, 899–908.
Seeman, P., Tallerico, T., Ko, F., 2003. Dopamine displaces [3H]domperidone from high-affinity sites of the dopamine D2 receptor, but not [3H]raclopride or [3H]spiperonein isotonic medium: implications for human positron emission tomography.Synapse 49, 209–215.
Seeman, P., Tokita, K., Matsumoto, M., Matsuo, A., Sasamata, M., Miyata, K., 2009. Thedopaminergic stabilizer ASP2314/ACR16 selectively interacts with D2(High)receptors. Synapse 63, 930–934.
Semba, J., Watanabe, A., Kito, S., Toru, M., 1995. Behavioural and neurochemical effectsof OPC-14597, a novel antipsychotic drug, on dopaminergic mechanisms in ratbrain. Neuropharmacology 34, 785–791.
Sonesson, C., Andersson, B., Waters, S., Waters, N., Tedroff, J., 2000. New modulators ofdopamine neurotransmission. Patent application no. WO146145 A1.
Strange, P.G., 2008. Antipsychotic drug action: antagonism, inverse agonism or partialagonism. Trends Pharmacol. Sci. 29, 314–321.
Svensson, T.H., 2003. Alpha-adrenoceptor modulation hypothesis of antipsychoticatypicality. Prog. Neuropsychopharmacol. Biol. Psychiatry 27, 1145–1158.
Svensson, K.A., Falcone, J.F., Johansson, A.M., Perry, K.W., Fell, M.A., 2009. The actions ofthe dopamine stabilizer ACR16, but not (−)-OSU6162, in behavioral andneurochemical assays are not dependent on the presence of functional dopamineD2 receptors. Poster presented at the Society for Neuroscience Annual Conference,Chicago, IL, USA.
Tadori, Y., Kitagawa, H., Forbes, R.A., McQuade, R.D., Stark, A., Kikuchi, T., 2007. Differencesin agonist/antagonist properties at human dopamine D(2) receptors betweenaripiprazole, bifeprunox and SDZ 208–912. Eur. J. Pharmacol. 574, 103–111.
Tedroff, J., Lindskov Krogh, P., Buusman, A., Rembratt, A., 2009. Pridopidine (ACR16) inHuntington's disease: an update on the MermaiHD and HART studies. Posterpresented at the World Congress on Huntington's Disease, Vancouver, Canada.
Tedroff, J., Sonesson, C.,Waters, N.,Waters, E.S., Carlsson, A., 2004. Apilot studyof thenoveldopamine stabilizer ACR16 in advanced Parkinson's disease. Mov. Disord. 19, S201.
Tekin, S., Cummings, J.L., 2002. Frontal–subcortical neuronal circuits and clinicalneuropsychiatry: an update. J. Psychosom. Res. 53, 647–654.
Waters, N., Carlsson, A., Carlsson, M., Martin, P., Tedroff, J., Kullingsjö, J., Pettersson, F.,Sonesson, C.,Waters, E.S., 2002. Pharmacology of ACR16, a dopamine stabilizer. Posterpresented at the Society for Neuroscience Annual Conference, Orlando, FL, USA.
Waters, N., Lagerkvist, S., Lofberg, L., Piercey, M., Carlsson, A., 1993. The dopamine D3receptor and autoreceptor preferring antagonists (+)-AJ76 and (+)-UH232; amicrodialysis study. Eur. J. Pharmacol. 242, 151–163.
Waters, S., Pettersson, F., Dyhring, T., Sonesson, C., Tedroff, J., Waters, N., Pontén, H.,2009a. Pharmacology of the dopaminergic stabilizer pridopidine (ACR16). Clin.Genet. 76 (Suppl 1), 74 (Abstract D10).
Waters, S., Pettersson, F., Dyhring, T., Sonesson, C., Tedroff, J., Waters, N., Pontén, H.,2009b. Preclinical pharmacology and mode of action of the dopaminergic stabilizerACR16. Poster presented at the 4th Annual Huntington's Disease TherapeuticConference, Cannes, France.
Westerink, B.H., de Boer, P., de Vries, J.B., Kruse, C.G., Long, S.K., 1998. Antipsychoticdrugs induce similar effects on the release of dopamine and noradrenaline in themedial prefrontal cortex of the rat brain. Eur. J. Pharmacol. 361, 27–33.
Westerink, B.H., Kawahara, Y., De Boer, P., Geels, C., De Vries, J.B., Wikstrom, H.V., VanKalkeren, A., Van Vliet, B., Kruse, C.G., Long, S.K., 2001. Antipsychotic drugsclassified by their effects on the release of dopamine and noradrenaline in theprefrontal cortex and striatum. Eur. J. Pharmacol. 412, 127–138.
Westerink, B.H., Korf, J., 1976. Comparison of effects of drugs on dopamine metabolismin the substantia nigra and the corpus striatum of rat brain. Eur. J. Pharmacol. 40,131–136.

















![Synthesis and Evaluation of a Set of 4-Phenylpiperidines and 4-Phenylpiperazines as D 2 Receptor Ligands and the Discovery of the Dopaminergic Stabilizer 4-[3-(Methylsulfonyl)phenyl]-1-propylpiperidine](https://static.fdokumen.com/doc/165x107/6345275cdf19c083b107eeae/synthesis-and-evaluation-of-a-set-of-4-phenylpiperidines-and-4-phenylpiperazines.jpg)


