
Two casein kinase 1 isoforms are differentially expressed in Trypanosoma cruzi

Journal of Nkurochemrstry Raven Press. Ltd., New York (d 1990 International Society for Neurochemistry
Neuroblastoma Differentiation Involves the Expression of Two Isoforms of the a-Subunit of Go
Philippe Brabet, Colette Pantaloni, Marc Rodriguez, Jean Martinez, Joel Bockaert, and Vincent Homburger
Centre CNRS-INSERM de Pharrnacologie et Endocrinologie, Montpellier, France
Abstract: The regulation of GTP-binding proteins (G pro- teins) was examined during the course of differentiation of neuroblastoma N 1 E- I 15 cells. N 1 E- I I5 cell membranes possess three Bordetella pertussis toxin (PTX) substrates as- signed to a-subunits (Ga) of Go (a G protein of unknown function) and “Ci (a G protein inhibitory to adenylate cy- c1ase)-like” proteins and one substrate of Vibrio cholerae toxin corresponding to an a-subunit of G, (a G protein stimulatory to adenylate cyclase). In undifferentiated cells, only one form of Goa was found, having a PI of 5.8. Goa content increased by approximately twofold from the undifferentiated state to 96 h of cell differentiation. This is mainly due to the ap- pearance of another Goa form having a PI of 5.55. Both Goa isoforms have similar sizes on sodium dodecyl sulfate-poly- acrylamide gels, are recognized by polyclonal antibodies to
bovine brain Goa, are ADP-ribosylated by PTX, and are co- valently myristylated in whole N 1 E- I 15 cells. In addition, imrnunofluorescent staining of N 1 E- 1 15 cells with Goa an- tibodies revealed that association of Goa with the plasma membrane appears to coincide with the expression of the most acidic isoform and morphological cell differentiation. In contrast, the levels ofboth Gia and Gsa did not significantly change, whereas that of the common @-subunit increased by - 30% over the same period. These results demonstrate spe- cific regulation of the expression of Goa during neuronal dif- ferentiation, Key Words: ADP ribosylation-G,a protein- Neuroblastoma cells. Brabet P. et al. Neuroblastoma differ- entiation involves the expression of two isoforms of the a- subunit of Go. J. Neurochem. 54, 1310-1320 (1990).
Go [a GTP-binding protein (G protein) of unknown function] is heterotrimeric, composed of a-, 6-, and y- subunits (Ga, GP, and Gy, respectively), and was first purified from bovine brain (Neer et al., 1984; Sternweis and Robishaw, 1984). The G proteins represent a fam- ily of highly homologous proteins on the basis of the apparent similarities in molecular weight, amino acid sequence, structure, and function. The a-subunits, which have similar but distinct molecular weights, bind guanine nucleotides, and possess sites for bacterial toxin-catalyzed ADP-ribosylation, confer specificity for each G protein, whereas tightly associated P- and y- subunits are believed to be interchangeable between all the G proteins (Birnbaumer et al., 1987; Neer and Clapham, 1988). Goa has a relative molecular weight (M,) of 39,000 and is ADP-ribosylated by Bordetella
Received June 14, 1989; revised manuscript received August 31, 1989; accepted September 7, 1989.
Address correspondence and reprint requests to Dr. V. Hornburger at Centre CNRS-INSERM de Pharmacologie et Endocrinologie, rue de la Cardonille, 34094 Montpellier Cedex 2, France.
Abbreviations used: BSA, bovine serum albumin; CTX, Cholera toxin; DMSO, dimethyl sulfoxide; DTT, dithiothreitol: FCS, fetal calf serum; G protein, GTP-binding protein; Ga, G& and Gy, a-, p-, and y-subunit, respectively, of GTP-binding protein; GABA, y-
pertussis toxin (PTX) like some other G proteins: the a-subunit of the G protein inhibitory to adenylate cy- clase (Gia; M, = 40,000 and 4 1,000) and transducin (T; retinal photoreceptor G protein). The a-subunit of the G protein stimulatory to adenylate cyclase (G,a; two species of M, 42,000-52,000) is a substrate of Cholera toxin (CTX)-catalyzed ADP-ribosylation.
Numerous immunochernical and biochemical in- vestigations have clearly indicated that the tissue dis- tribution of G,a is essentially restricted to nervous and endocrine systems (Gierschik et al., 1986; Homburger et al., 1987; Toutant et al., 1987; Asano et al., 1988~; Brabet et al., 1988a,h), a result suggesting that this pro- tein may participate in secretory mechanisms. Re- cently, mRNA for Goa has been reported to be abun- dant in the CNS but was not detected in peripheral
aminobutyric acid; Gi and G,, GTP-binding proteins inhibitory and stimulatory, respectively, to adenylate cyclase; GiaW and Gia4, , a- subunit of GTP-binding protein inhibitory to adenylate cyclase having a relative molecular weight of 40,000 and 41,000, respectively; Go, GTP-binding protein of unknown function; IEF, isoelectrkfocusing; M,, relative molecular weight; NEM, N-ethylmaleimide; PAGE, polyacrylamide gel electrophoresis; PBS, phosphate-buffered saline; PTX, Bordetella pertussis toxin; SDS, sodium dodecyl sulfate; T, transducin (retinal photoreceptor GTP-binding protein).
1310

NEURONAL DIFFERENTIATION AND G,a ISOFORMS 1311
tissues (Brann et al., 1987). In contrast, the Goa subunit and its mRNA have been identified immunologically and by Northern blot analysis in rat heart (Luetje et al., 1988).
Our finding about the extrasynaptic localization of Go observed at the ultrastructural level in cultured neurons suggested that this protein was probably re- sponsible for a transduction occurring outside of mor- phologically differentiated synapses (Gabrion et al., 1988). Functional reconstitution experiments indicated that purified Go interacts with muscarinic cholinergic, y-aminobutyric acid B (GABAB), a,-adrenergic, and D,-dopamine receptors (Asano et al., 1985; Cerione et al., 1986; Kurose et al., 1986; Senogles et al., 1987). To date, a possible function for Go is the inhibition of voltage-sensitive Ca2+ channel activity (Hescheler et al., 1987; Ewald et al., 1988; Harris-Warrick et al., 1988); and the activation of some neuronal K+ chan- nels (VanDongen et al., 1988).
Recently, the presence of multiple forms of PTX substrates and in particular two Goa forms has been demonstrated in bovine brain (Brabet et al., 19886; Goldsmith et al., 1988). Although some modulations of the amounts of G proteins have been reported during cell differentiation (Watluns et al., 1987; Mullaney et al., 1988) and brain and heart development (Asano et al., 19886), little is known of the regulation of isoforms of these G proteins in these systems. In this study, we investigated the regulation of the G proteins and the temporal nature of Go expression in neuroblastoma N1E-115 cells induced to differentiate by addition of dimethyl sulfoxide (DMSO). The morphological dif- ferentiation of this clonal cell line N 1 E- 1 15 is accom- panied by several events characteristic of neuronal maturation (Kimhi et al., 1976). We report that during the course of differentiation with DMSO, there is a twofold increase in the amount of Goa, whereas Gia and G,a were not affected. More interesting is that this increase in the amount of G,a is mainly due to the expression of an acidic isoform of Goa (PI 5.55). In undifferentiated neuroblastoma cells, only one isoform of Goa was present, having a PI of 5.8, whereas both isoforms were present in differentiated neuroblastoma cells. Both forms of G,a are ADP-ribosylated by PTX, recognized by our anti-Goa antibody, and myristylated.
MATERIALS AND METHODS
Bovine brain G proteins Purification of G proteins from bovine brain membranes
was performed as previously reported (Homburger et al., 1987) according to the procedure initially described by Sternweis and Robishaw ( 1 984).
Culture conditions and biosynthetic labeling Cerebral cortex from 14 or 15 Swiss Albino mouse fetuses
was used to generate primary cultures of neurons and as- troglial cells under conditions previously described (Brabet et al., 1988a). Murine neuroblastoma NIE-115 cells were cultured in optiMEM 1 medium (GIBCO) supplemented with
10% (vol/vol) fetal calf serum (FCS) and 50 units/ml of pen- icillin plus streptomycin (GIBCO). Cells were grown in 100- mm-diameter Falcon dishes in a humidified atmosphere of 10% C02/90% air at 37°C. Cells were subcultured weekly by removing the medium, incubating for 10 rnin in Versene ( I : 5,000) isotonically buffered saline solution, and then resus- pending in 10 ml of growth medium. The cells were then replated at a density of 2-4 X lo5 cells/dish, and medium was changed on day 3 and each day thereafter. On day 3, the cells were maintained in growth medium or induced to dif- ferentiate for various times in the presence of 0.5% FCS and I .5% DMSO (Fluka). For biosynthetic labeling, cells were incubated for 4 h in methionine-free medium (GIBCO) sup- plemented with 20 pCi/ml of [35S]methionine (1,000 Ci/ mmol; New England Nuclear) or in Dulbecco's modified Ea- gle's medium (Boehringer) supplemented with 5 mM pyru- vate and 40-50 pCi/ml of [3H]myristic acid (22 Ci/mmol; New England Nuclear).
Preparation of membrane fractions Cells grown in Falcon dishes were scraped off in 0.9% NaCl
and centrifuged for 10 rnin at 300 g. The cell pellets were resuspended in I ml of ice-cold lysing buffer containing 20 mM Tris-HC1 (pH 7.5), 3 m M EDTA, 5 mg/ml of soybean trypsin inhibitor, and 0.1 mM fresh phenylmethylsulfonyl fluoride, homogenized at 0°C in a Potter-Elvehjem homog- enizer, and then centrifuged for 15 min at 40,000 g. Mem- brane pellets were homogenized in the same buffer at a final protein concentration of 1-2 mgjml and stored in liquid ni- trogen before use. The term membrane is used in the text to design this crude membrane fraction.
Bacterial toxin labeling ADP-ribosylation with PTX (kindly provided by Dr. A.
Garcia-Sainz, Department of Bioenergktica, Universidad Nacional Autonomh de Mexico) was carried out as previously described (Brabet et al., 19886) for 60 rnin at 30°C under the following conditions: 70 mM Tris-HC1 (pH 7.5), 1 mM EDTA, 0.5 pM NAD, 0.5-1 pCi of [a-32P]NAD, I mMATP, 100 p M GTP, 10 mM thymidine, 0.1 mM MgCI2, 1 mg/ml of L-myristyl phosphatidylcholine, 10 mM nicotinamide, 25 mM dithiothreitol (DTT), and 5 pg of activated PTX/60 pl total volume. For [32P]ADP-ribosylation of membrane pro- teins with CTX (Sigma), membranes (50 pg of protein) were incubated for 60 rnin at 37°C in a volume of20 pl containing 100 mMpotassium phosphate buffer(pH 7.5), 10 mMMgC12, 1 mA4 ATP, 10 mM thymidine, 10 mM arginine, 5 mM isoniazide, 100 p M GTP, 2-3 pCi of [cI-~'P]NAD, and 60 pg/ml of CTX preactivated by incubation with 20 mM DTT for 15 min at 30°C. The reaction was stopped by adding 2% sodium dodecyl sulfate (SDS) plus 100 pg/ml of bovine serum albumin (BSA) followed by precipitation with 10% trichloracetic acid for 2 h at 0°C.
One- and two-dimensional electrophoretic analysis and immunoblotting
For one-dimensional analysis, samples were solubilized in 50 mM Tris-HC1 (pH 6 4 , 5% SDS, and 50 mM DTT for 5 rnin at 90"C, cooled, mixed with 100 mMN-ethylmaleimide (NEM), and allowed stand for 15 rnin at room temperature. Then, Laemmli sample buffer (Laemmli, 1970) was added, and an aliquot was loaded onto 10% polyacrylamide gels for SDS-polyacrylamide gel electrophoresis (PAGE). Gels con- taining ADP-ribosylated proteins were stained with Coo- massie Brilliant Blue (R250), dried, and exposed to Kodak
J. Neurochem., Vol. 54, No. 4, 1990

1312 P. BRABET ET AL.
X AR5 film in an x-ray exposer holder with no intensifying screen. For immunoblotting, proteins were transferred onto nitrocellulose sheets at a voltage of 20 V overnight. Subse- quent procedures were performed according to the method previously reported (Brabet et al., 1988~). Strips of blots or gels that displayed bands on the autoradiogram were ex- cised, and the amount of radioactivity associated with G pro- tein subunits was quantified in y- and (kcintillation counters, respectively.
Two-dimensional analysis consisted of isoelectrofocusing (IEF) and SDS-PAGE in the first and second dimension, respectively. IEF gels were prepared according to the pro- cedure of O’Farrell ( 1975) with a mixture of 2% ampholines at a ratio of 4: I of pH 5-7 and pH 3.5- 10. Before application onto vertical IEF tube gels, samples were solubilized in 9.2 M urea, 4% Nonidet P-40, and 0.5 mM DTT, alkylated with 9 mM NEM for 15 min at room temperature, and then di- luted in a buffer containing 9.2 M urea, 4% ampholines (Pharmacia), 4% B-mercaptoethanol, and 4% Nonidet P-40. The established gradient in pH was linear between 5.0 and 6.4. Then, vertical IEF tube gels were either blotted as de- scribed by Otey et al. (1986) or laid onto the top of 1.5-mm 10% polyacrylamide gels to further separate proteins in the second dimension. Nitrocellulose sheets that contained membrane proteins stained with [3H]myristic acid were sub- jected to fluorography with 2,5-diphenyloxazole/toluene.
Antisera and immunoprecipitation Production, characterization, and affinity purification of
polyclonal rabbit antibodies raised against bovine brain Go& and Go have previously been described (Homburger et al., 1987; Brabet et al., 1988a). To produce anti-G,a, the pep- tide immunogen Gln-Leu-Gln-Lys-Asp-Lys-Gln-Val-Tyr- Arg-Ala-NH2, representing a specific amino acid sequence of Gsa, was covalently linked via the maleimido-thiol pro- cedure (Blache et al., 1988). Thiol groups were grafted on the BSA molecule according to the method of Klotz and Heiney (1962) by reaction of the protein with an excess of S-acetylmercaptosuccinic anhydride followed by hydrolysis of the thioester linkage with hydroxylamine. The BSA-thiol derivative was allowed to react in situ with the Na-maleoyl- p- Ala- Gln- Leu-Gln - Lys- Asp- Lys-Gln - Val -Tyr- Arg- Ala- NH,toproduce theBSA-peptideconjugate. ThisNa-maleoyl- /3- Ala-Gln - Leu-Gln - Lys- Asp- Lys- Gln - Val- Tyr- Arg- Ala- NH2 was synthesized in solution, purified by HPLC [C18 column, Beckman ODS 5 pm, 10 X 250 mm; solvent, 0.05 M ammonium acetate (pH 6.5) and methanol (40:60 vol/ vol); flow rate, 3 ml/min; detection at 220 nm], and identified by 360-MHz ‘H nuclear magnetic resonance spectroscopy. Its purity was ascertained by TLC in different solvent systems and by HPLC. (Experimental details of this synthesis will be reported elsewhere.)
Immunoprecipitation was performed using anti-G,a an- tibodies affinity-purified on an Na-maleoyl-peptide-thiopro- pyl Sepharose 6B column, followed by a 10% (wt/vol) sus- pension of fixed Staphylococcus aureus cells (Immuno-pre- cipitin; BRL). Membrane proteins [32P]ADP-ribosylated with CTX were solubilized for 3 min at 100°C with 1% SDS in 50 mM sodium phosphate buffer (pH 7.5) containing 1 mM DTT. Four volumes of buffer A 1.25X [12.5 mM sodium phosphate (pH 7.5), 190 mM NaCl, 1.25 mM DTT, 1.25% Nonidet P-40, and 0.25% SDS] was added to the soluble frac- tions, which were then reacted with 5 pl of anti-G,a antibodies for 16 h at 4°C and with 30 p1 of S. aureiis suspension for 60 min at room temperature. Pelleted precipitates were then
washed three times with buffer A and subjected to SDS- PAGE.
Immunofluorescent detection of G,a This was performed as previously described (Brabet et al.,
1988~). In brief, cells cultured on Thermanox glass cov- erslips (Miles, Norgerville, IL, U.S.A.) were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS; pH 7.4) for 30 min at room temperature, and permeabilized with 0.05% Triton X-100 in PBS for 5 min. Coverslips were then incubated with affinity-purified anti-G,,a antibodies ( 10-20 pg/ml) in PBS containing 150 mM NaCl and 0.2% gelatin for 2 h at 37”C, washed three times with the same buffer, and exposed to goat antibody conjugated to rhodamine for 60 min at room temperature. After extensive washing with PBS, coverslips were mounted in Moviol 4.88 (Hoechst) mounting medium and visualized with a Leitz Dialux 20 equipped with interferential filters.
RESULTS
Status of FTX substrates during N1E-115 cell differentiation
In addition to G,a, PTX ADP-ribosylated several distinct “Gia-like” proteins, which have been detected by molecular cloning, immunological and biochemical studies, and are now designated as the Gi 1 a, Gi2a, and Gi3a subunits (M, = 40,000-4 1,000) (Itoh et al., 1988; Mumby et al., 1988).
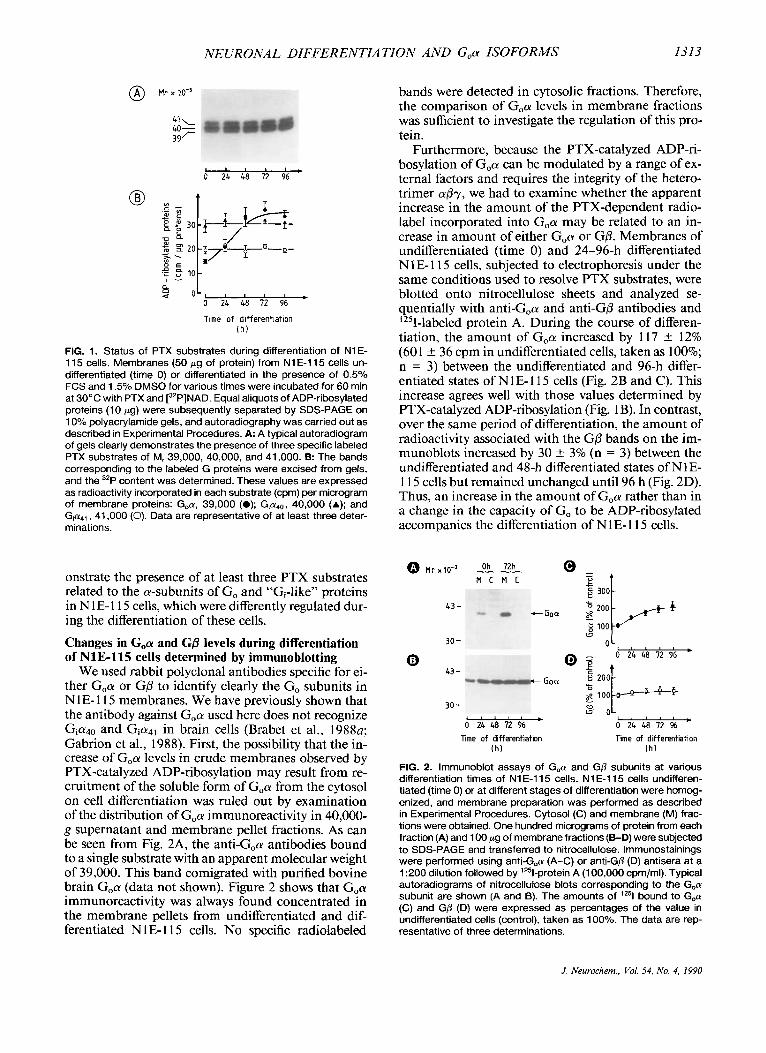
We thus attempted to examine first the status of these PTX substrates of N 1 E- 1 1 5 neuroblastoma cells in the undifferentiated state and during differentiation in- duced by 1.5% (vol/vol) DMSO. As shown in Fig. IA, incubation of membrane preparations from undiffer- entiated cells (time 0) with preactivated PTX and [a- 32P]NAD led to the incorporation of specific radioac- tivity into three G a polypeptides of M, 39,000,40,000, and 41,000, which were clearly separated by SDS- PAGE after reduction and alkylation with NEM. These polypeptides are referred to here as Goa, Gia40, and Gia4,, respectively. During the time course of cell dif- ferentiation, the labeling of G,a steadily increased (Fig. 1A). Thus, there was approximately twice as much label incorporated into G,a (from 159 k 7 cpm/l0 pg of protein in undifferentiated cells to 333 k 15 cpm/l0 p g in 96-h differentiated cells; n = 3). Whereas relatively slight differences were noted in the labeling of Cia polypeptides from a unique experiment (Fig. lA), no significant difference was quantified in the incorpora- tion of radioactivity into Gia40 (from 293 f 25 to 3 12 +- 30 cpm; n = 3) and Gia4, (from 197 k 15 to 195 f 5 cpm; n = 3) when a series of experiments were performed (Fig. 1B). The evolution of percentages of the radioactivity incorporated in each PTX substrate relative to the total radioactivity incorporated in the three substrates indicated clearly that GiadO represented the major PTX substrate in undifferentiated N 1 E- 1 15 cells (44.6 k 0.7%; n = 6) in comparison with Gia4, (31.4 f 0.8%; n = 6) and Goa (24.1 k 0.6%; n = 6) , whereas Goa became the major one (44.2 k 1.2%, n = 6) in 96-h differentiated cells. These results dem-
J. Neurochem., Val. 54, No. 4, 1990
NEURONAL DIFFERENTIATION AND G,a ISOFORI.1;S 1313
@ Mr x
4' k
3 9 r 40 -
I " " . 0 24 48 ?2 96
Time of dif ferentiat ion l h )
FIG. 1. Status of PTX substrates during differentiation of N1 E- l 15 cells. Membranes (50 pg of protein) from N1 E-115 cells un- differentiated (time 0) or differentiated in the presence of 0.5% FCS and 1.5% DMSO for various times were incubated for 60 min at 30°C with PTX and [32P]NAD. Equal aliquots of ADP-ribosylated proteins (10 rg) were subsequently separated by SDS-PAGE on 10% polyacrylamide gels, and autoradiography was carried out as described in Experimental Procedures. A: A typical autoradiogram of gels clearly demonstrates the presence of three specific labeled PTX substrates of M, 39,000, 40,000, and 41,000. B: The bands corresponding to the labeled G proteins were excised from gels, and the 32P content was determined. These values are expressed as radioactivity incorporated in each substrate (cpm) per microgram of membrane proteins: Goa, 39,000 (0); G,ado, 40,000 (A); and G,a4,, 41,000 (0). Data are representative of at least three deter- minations.
onstrate the presence of at least three PTX substrates related to the a-subunits of Go and "Gi-like" proteins in N 1 E- 1 15 cells, which were differently regulated dur- ing the differentiation of these cells.
Changes in G,a and GP levels during differentiation of N1E-115 cells determined by immunoblotting
We used rabbit polyclonal antibodies specific for ei- ther G,a or GP to identify clearly the Go subunits in N 1E- 1 1 5 membranes. We have previously shown that the antibody against G,a used here does not recognize
and Gia41 in brain cells (Brabet et al., 1988~; Gabrion et al., 1988). First, the possibility that the in- crease of G,a levels in crude membranes observed by PTX-catalyzed ADP-ribosylation may result from re- cruitment of the soluble form of G,a from the cytosol on cell differentiation was ruled out by examination of the distribution of G,a immunoreactivity in 40,000- g supernatant and membrane pellet fractions. As can be seen from Fig. 2A, the anti-G,a antibodies bound to a single substrate with an apparent molecular weight of 39,000. This band comigrated with purified bovine brain G,a (data not shown). Figure 2 shows that G,a immunoreactivity was always found concentrated in the membrane pellets from undifferentiated and dif- ferentiated N 1 E- 1 15 cells. No specific radiolabeled
bands were detected in cytosolic fractions. Therefore, the comparison of G,a levels in membrane fractions was sufficient to investigate the regulation of this pro- tein.
Furthermore, because the PTX-catalyzed ADP-ri- bosylation of G,a can be modulated by a range of ex- ternal factors and requires the integrity of the hetero- trimer spy, we had to examine whether the apparent increase in the amount of the PTX-dependent radio- label incorporated into G,a may be related to an in- crease in amount of either G,a or GP. Membranes of undifferentiated (time 0) and 24-96-h differentiated N1 E- 1 15 cells, subjected to electrophoresis under the same conditions used to resolve PTX substrates, were blotted onto nitrocellulose sheets and analyzed se- quentially with anti-G,a and anti-GP antibodies and '"I-labeled protein A. During the course of differen- tiation, the amount of G,a increased by 117 +- 12% (601 * 36 cpm in undifferentiated cells, taken as 100%; n = 3) between the undifferentiated and 96-h differ- entiated states of NlE-115 cells (Fig. 2B and C). This increase agrees well with those values determined by PTX-catalyzed ADP-ribosylation (Fig. 1 B). In contrast, over the same period of differentiation, the amount of radioactivity associated with the GO bands on the im- munoblots increased by 30 1- 3% (n = 3) between the undifferentiated and 48-h differentiated states of N1 E- l 15 cells but remained unchanged until 96 h (Fig. 2D). Thus, an increase in the amount of G,a rather than in a change in the capacity of Go to be ADP-ribosylated accompanies the differentiation of N 1 E- 1 1 5 cells.
43 -
- -- 0 24 48 72 96 0 24 48 12 %
llme of differentiation Time of differentiation lh ) I h l
FIG. 2. lmmunoblot assays of G,a and GO subunits at various differentiation times of N1E-115 cells. N1E-115 cells undifferen- tiated (time 0) or at different stages of differentiation were homog- enized, and membrane preparation was performed as described in Experimental Procedures. Cytosol (C) and membrane (M) frac- tions were obtained. One hundred micrograms of protein from each fraction (A) and 100 pg of membrane fractions (B-D) were subjected to SDS-PAGE and transferred to nitrocellulose. lmmunostainings were performed using anti-(;,a (A-C) or anti-Gp (D) antisera at a 1 :200 dilution followed by '251-protein A (1 00.000 cpmlml). Typical autoradiograms of nitrocellulose blots corresponding to the G,a subunit are shown (A and B). The amounts of lZ5l bound to G,a (C) and GD (D) were expressed as percentages of the value in undifferentiated cells (control), taken as 100%. The data are rep- resentative of three determinations.
J. Neurochern., Vol. 54> No. 4, I990
1314 P. RRABET ET AL.
Quantification of relative levels of G, by immunoblotting using specific antipeptide antibodies
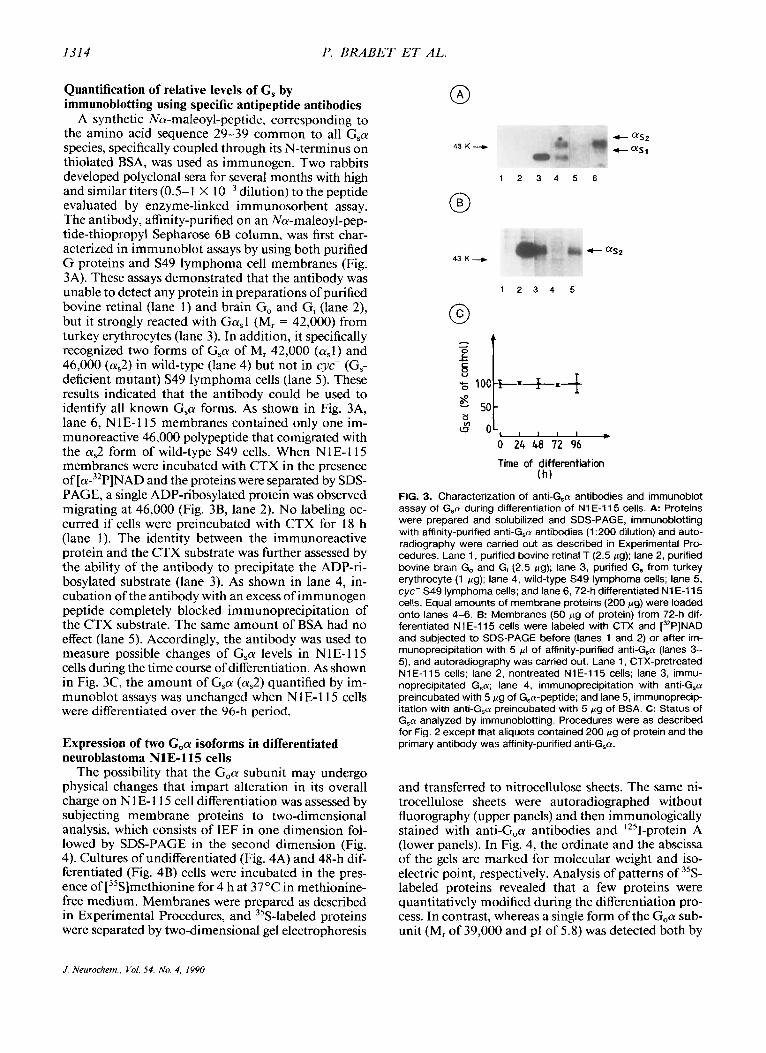
A synthetic Na-maleoyl-peptide, corresponding to the amino acid sequence 29-39 common to all G,a species, specifically coupled through its N-terminus on thiolated BSA, was used as immunogen. Two rabbits developed polyclonal sera for several months with high and similar titers (0.5- I X dilution) to the peptide evaluated by enzyme-linked immunosorbent assay. The antibody, affinity-purified on an Na-maleoyl-pep- tide-thiopropyl Sepharose 6B column, was first char- acterized in immunoblot assays by using both purified G proteins and S49 lymphoma cell membranes (Fig. 3A). These assays demonstrated that the antibody was unable to detect any protein in preparations of purified bovine retinal (lane 1) and brain Go and Gi (lane 2), but it strongly reacted with Ga,l (M, = 42,000) from turkey erythrocytes (lane 3). In addition, it specifically recognized two forms of G,a of M, 42,000 (a,l) and 46,000 (a,2) in wild-type (lane 4) but not in cyc- (G, deficient mutant) S49 lymphoma cells (lane 5). These results indicated that the antibody could be used to identify all known G,a forms. As shown in Fig. 3A, lane 6, NlE-115 membranes contained only one im- munoreactive 46,000 polypeptide that comigrated with the a,2 form of wild-type S49 cells. When NlE-I 15 membranes were incubated with CTX in the presence of [a-32P]NAD and the proteins were separated by SDS- PAGE, a single ADP-ribosylated protein was observed migrating at 46,000 (Fig. 3B, lane 2). No labeling oc- curred if cells were preincubated with CTX for 18 h (lane 1). The identity between the immunoreactive protein and the CTX substrate was further assessed by the ability of the antibody to precipitate the ADP-ri- bosylated substrate (lane 3). As shown in lane 4, in- cubation of the antibody with an excess of immunogen peptide completely blocked immunoprecipitation of the CTX substrate. The same amount of BSA had no effect (lane 5). Accordingly, the antibody was used to measure possible changes of G,a levels in N1E-115 cells during the time course of differentiation. As shown in Fig. 3C, the amount of Gsa (a,2) quantified by im- munoblot assays was unchanged when N 1E-115 cells were differentiated over the 96-h period.
Expression of two G,a isoforms in differentiated neuroblastoma N1E-115 cells
The possibility that the Goa subunit may undergo physical changes that impart alteration in its overall charge on N 1 E- 1 15 cell differentiation was assessed by subjecting membrane proteins to two-dimensional analysis, which consists of IEF in one dimension fol- lowed by SDS-PAGE in the second dimension (Fig. 4). Cultures of undifferentiated (Fig. 4A) and 48-h dif- ferentiated (Fig. 4B) cells were incubated in the pres- ence of [35S]methionine for 4 h at 37°C in methionine- free medium. Membranes were prepared as described in Experimental Procedures, and 35S-labeled proteins were separated by two-dimensional gel electrophoresis
43 K - 1 2 3 4 5 6
+ as2 43 K -.+
1 2 3 4 5
Time of differentiation
FIG. 3. Characterization of anti-(;,a antibodies and immunoblot assay of Gsol during differentiation of N1 E-115 cells. A Proteins were prepared and solubilized and SDS-PAGE, immunoblotting with affinity-purified anti-G,a antibodies (1 :200 dilution) and auto- radiography were carried out as described in Experimental Pro- cedures. Lane 1, purified bovine retinal T (2.5 pg); lane 2, purified bovine brain Go and G, (2.5 pg): lane 3, purified G, from turkey erythrocyte (1 pg); lane 4, wild-type S49 lymphoma cells; lane 5, cyc- S49 lymphoma cells; and lane 6, 72-h differentiated N1E-115 cells. Equal amounts of membrane proteins (200 pg) were loaded onto lanes 4-6. 6: Membranes (50 pg of protein) from 72-h dif- ferentiated N1 E-115 cells were labeled with CTX and [32P]NAD and subjected to SDS-PAGE before (lanes 1 and 2) or after irn- munoprecipitation with 5 pI of affinity-purified anti-G,a (lanes 3- 5), and autoradiography was carried out. Lane 1, CTX-pretreated N1 E-115 cells: lane 2, nontreated N1 E-115 cells; lane 3, immu- noprecipitated G,a; lane 4, immunoprecipitation with anti-G,a preincubated with 5 pg of G,a-peptide; and lane 5, immunoprecip- itation with anti-(;,a preincubated with 5 pg of BSA. C: Status of G,a analyzed by immunoblotting. Procedures were as described for Fig. 2 except that aliquots contained 200 pg of protein and the primary antibody was affinity-purified anti-G,ol.
( h )
and transferred to nitrocellulose sheets. The same ni- trocellulose sheets were autoradiographed without fluorography (upper panels) and then immunologically stained with anti-G,a antibodies and '251-protein A (lower panels). In Fig. 4, the ordinate and the abscissa of the gels are marked for molecular weight and iso- electric point, respectively. Analysis of patterns of 35S- labeled proteins revealed that a few proteins were quantitatively modified during the differentiation pro- cess. In contrast, whereas a single form of the G,a sub- unit (M, of 39,000 and pl of 5 .8 ) was detected both by
J. Neurochem., Val. 54. No. 4, 1990
NEURONAL DIFFERENTIATION AND G,a ISOFORMS 1315
w W 2 I
m 0 cn
I 43 g 39 cn
30
Mr x lo-'
94
67
43 39
30
pH b.L 5.8 5.6 5.2 pH 6.4 5.8 5.6 5.2
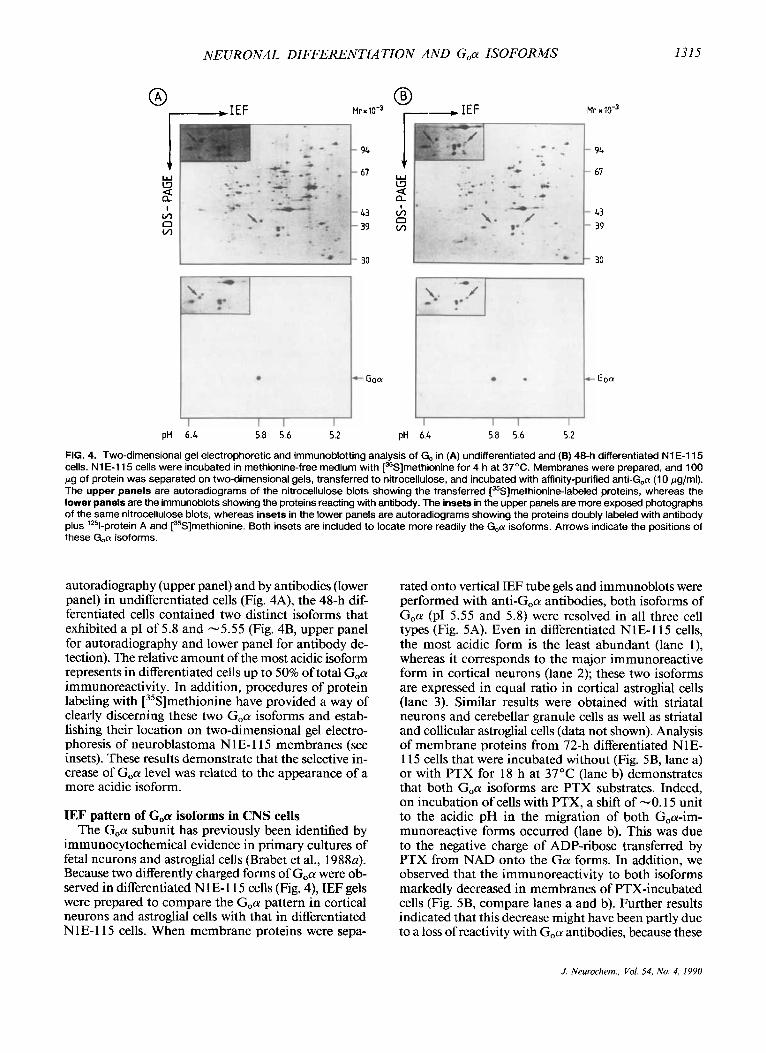
FIG. 4. Two-dimensional gel electrophoretic and immunoblotting analysis of Go in (A) undifferentiated and (B) 48-h differentiated NIE-I 15 cells. N1 E-I15 cells were incubated in methionine-free medium with [35S]methionine for 4 h at 37'C. Membranes were prepared, and 100 pg of protein was separated on two-dimensional gels, transferred to nitrocellulose, and incubated with affinity-purified anti-G,a (1 0 pglml). The upper panels are autoradiograms of the nitrocellulose blots showing the transferred [35S]methionine-labeled proteins, whereas the lower panels are the immunoblots showing the proteins reacting with antibody. The insets in the upper panels are more exposed photographs of the same nitrocellulose blots, whereas insets in the lower panels are autoradiograms showing the proteins doubly labeled with antibody plus '*%protein A and [35S]methionine. Both insets are included to locate more readily the G,a isoforms. Arrows indicate the positions of these G,a isoforms.
autoradiography (upper panel) and by antibodies (lower panel) in undifferentiated cells (Fig. 4A), the 48-h dif- ferentiated cells contained two distinct isoforms that exhibited a PI of 5.8 and -5.55 (Fig. 4B, upper panel for autoradiography and lower panel for antibody de- tection). The relative amount of the most acidic isoform represents in differentiated cells up to 50% of total G,a immunoreactivity. In addition, procedures of protein labeling with [35S]methionine have provided a way of clearly discerning these two G,a isoforms and estab- lishing their location on two-dimensional gel electro- phoresis of neuroblastoma N 1 E- 1 15 membranes (see insets). These results demonstrate that the selective in- crease of G,a level was related to the appearance of a more acidic isoform.
IEF pattern of Goa isoforms in CNS cells The G,a subunit has previously been identified by
immunocytochemical evidence in primary cultures of fetal neurons and astroglial cells (Brabet et al., 1988a). Because two differently charged forms of G,a were ob- served in differentiated N l E- 1 15 cells (Fig. 4), IEF gels were prepared to compare the G,a pattern in cortical neurons and astroglial cells with that in differentiated N 1 E- 1 15 cells. When membrane proteins were sepa-
rated onto vertical IEF tube gels and immunoblots were performed with anti-G,a antibodies, both isoforms of G,a (PI 5.55 and 5.8) were resolved in all three cell types (Fig. 5A). Even in differentiated NIE-115 cells, the most acidic form is the least abundant (lane I ) , whereas it corresponds to the major immunoreactive form in cortical neurons (lane 2); these two isoforms are expressed in equal ratio in cortical astroglial cells (lane 3). Similar results were obtained with striatal neurons and cerebellar granule cells as well as striatal and collicular astroglial cells (data not shown). Analysis of membrane proteins from 72-h differentiated N 1 E- l 15 cells that were incubated without (Fig. 5B, lane a) or with PTX for 18 h at 37°C (lane b) demonstrates that both G,a isoforms are PTX substrates. Indeed, on incubation of cells with PTX, a shift of -0.15 unit to the acidic pH in the migration of both G,a-im- munoreactive forms occurred (lane b). This was due to the negative charge of ADP-ribose transferred by PTX from NAD onto the Ga! forms. In addition, we observed that the immunoreactivity to both isoforms markedly decreased in membranes of PTX-incubated cells (Fig. 5B, compare lanes a and b). Further results indicated that this decrease might have been partly due to a loss of reactivity with Goa antibodies, because these
J. Nrurochem., Vol. 54, No. 4, 1990

1316 P. BRABET ET AL.
A PH 0
c 6.4 --c
c 5.8 - c 5.6 --c - 5.4 ----c
1 2 3 a b
FIG. 5. IEF analysis of Goa isoforms in neuronal and astroglial cells. A: Expression of Goa isoforms in neuronal and astroglial cells. Membranes (50 pg of protein) from 48-h differentiated NIE- 11 5 cells (lane I ) , cortical neurons (lane 2), and cortical astroglial cells (lane 3) were subjected to IEF on tube gels and immunoblotting with affinity-purified anti-G,a (10 pg/ml). The pl values were de- termined by measuring the pH of slices on triplicate IEF tube gels assayed in parallel for each experiment. B: PTX-catalyzed ADP- ribosylation of G,n isoforms detected by IEF. N1 E-115 cells dif- ferentiated for 72 h were cultured without (lane a) or with (lane b) PTX at 1 pg/rnl overnight at 37°C. Membranes (50 pg of protein) were subjected to IEF and immunoblotting with affinity-purified anti-G,a (1 0 pg/ml). These data are representative of at least three IEF patterns that have been replicated with membrane preparations.
antibodies were slightly less efficient in immunopre- cipitating the ADP-ribosylated form than the native form of Goa (data not shown). Furthermore, G,a re- mained intimately associated with the membrane pellet even after incubation of cells with PTX, excluding a loss of Goa during membrane preparation.
Both G0a isoforms contain rnyristate The chemical analysis of lipids covalently bound to
Goa purified from bovine brain has recently shown hydroxylamine-stable amide linkage of myristic acid (Buss et al., 1987; Schultz et al., 1987). The possibility that myristylation could be the posttranslational mod- ification that accounts for the appearance of the most acidic isoform of Goa in differentiated N 1 E- 1 15 cells
I I P H 5.8 5.6
Mrx
f 39
I I 5.8 5.6
was assessed again by two-dimensional analysis of membrane proteins (Fig. 6). Cultures of undifferen- tiated (A and C) and 48-h differentiated (B and D) neuroblastoma N I E- I 15 cells were incubated with [3H]myristic acid for 4 h at 37°C. Direct fluorography of 3H-labeled proteins shows one spot (arrow) at the level of the basic form of Goa in undifferentiated N 1 E- l 15 cells (A) and one additional spot at the level of the acidic form of G,a (second arrow) in differentiated cells (B). Note that two additional highly labeled spots were apparent, having a lower molecular weight than G,a (Fig. 6A and B). The nature of these spots is unknown. The identity of weakly labeled spots marked by arrows with Goa was confirmed by the superposition of these spots with those revealed by anti-G,a antibodies (one in Fig. 6C and two in Fig. 6D). These results were con- sistent with the conclusion that both Goa isoforms contain myristate and then that myristylation is un- likely to discriminate these two isoforms.
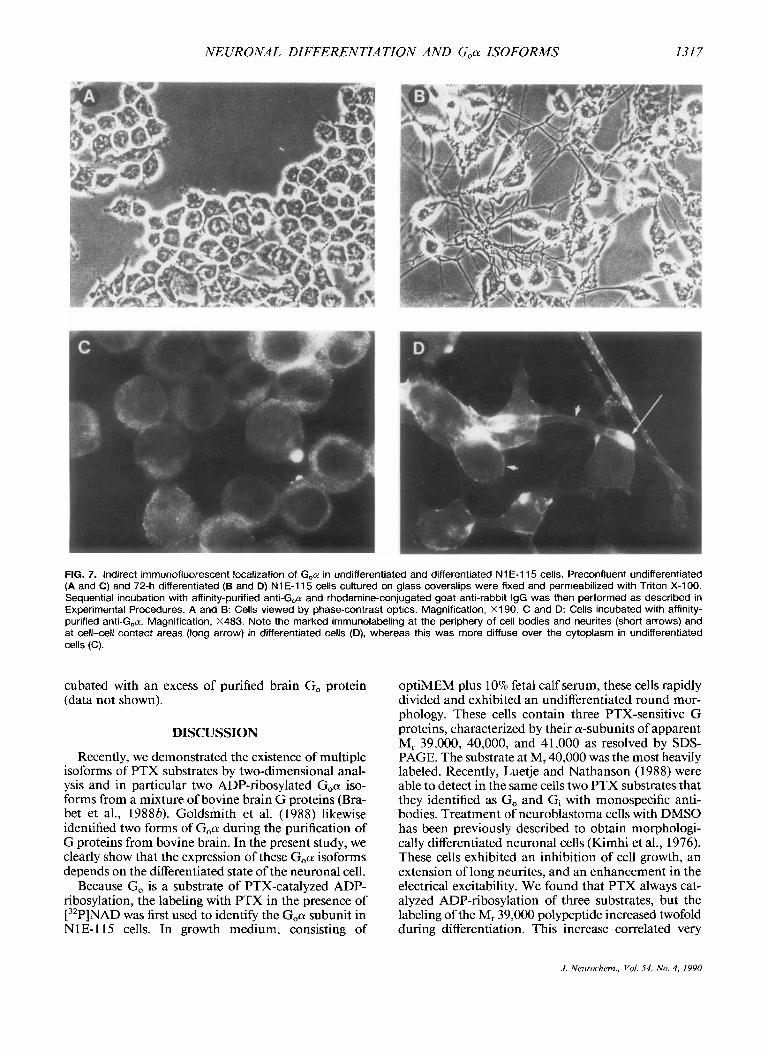
Immunocytolocalization of Go in NlE-115 cells The intracellular localization of Go was assessed with
affinity-purified anti-Goa antibodies by immunoflu- orescent staining of prefixed and permeabilized N 1 E- l 15 cells maintained in growth medium consisting of optiMEM with 10% fetal calf serum (Fig. 7A and C) or induced to differentiate for 72 h in optiMEM with 1.5% DMSO and 0.5% fetal calf serum (Fig. 7B and D). In proliferating N 1 E- 1 15 cells, which have a round cell morphology (A), specific immunolabeling appeared relatively diffuse in the cytoplasmic matrix, whereas nuclei remained unstained (C). The morphological dif- ferentiation of N 1 E- 1 15 cells is known to be associated with cells forming fibers (B). In these differentiated cells, an intense immunofluorescent reaction was especially pronounced at the inner face of the plasma membrane lining the cell body and the neurites (D, short arrows). A reinforcement of the positive reaction was often ob- served in cell-cell or cell-neurite contacts (D, long ar- row). This immunofluorescent pattern was totally ab- sent when anti-G,a antibodies were omitted or prein-
FIG. 6. Myristylation of G,cu isoforms detected by two-dimensional analysis and immunoblotting in neuroblastoma
Mr o- 3 N1 E-115 cells. Undifferentiated (A and C) and 48-h differentiated (B and D) neuroblastoma N1 E-115 cells were in- cubated with [3H]myristic acid for 4 h at 37OC. Membranes were prepared, and 100 pg of proteins was separated on two-dimensional gels, transferred to nitrocellulose, and incubated with af- finity-purified anti-G,a (5 pg/ml). A and B are fluorograms of the relevant por- tion of the nitrocellulose blots showing the transferred [3H]myristic acid-la- beled proteins, and C and D are im- munoblots showing the immunoreac- tive proteins with antibody. Arrows in- dicate the positions of the Goa isoforms.
f 39
c Goa
J. Neurochem., Vol. S4, No. 4. 1990
NEURONAL DIFFERENTIA TION AND G,a ISOFORMS 131 7
FIG. 7. Indirect immunofluorescent localization of G,n in undifferentiated and differentiated N1 E-115 cells. Preconfluent undifferentiated (A and C) and 72-h differentiated (6 and D) N1 E-115 cells cultured on glass coverslips were fixed and permeabilized with Triton X-1 00. Sequential incubation with affinity-purified anti-G,cy and rhodamine-conjugated goat anti-rabbit IgG was then performed as described in Experimental Procedures. A and 8: Cells viewed by phase-contrast optics. Magnification, X190. C and D: Cells incubated with affinity- purified anti-G,n. Magnification, X483. Note the marked immunolabeling at the periphery of cell bodies and neurites (short arrows) and at cell-cell contact areas (long arrow) in differentiated cells (D), whereas this was more diffuse over the cytoplasm in undifferentiated cells (C).
cubated with an excess of purified brain Go protein (data not shown).
DISCUSSION
Recently, we demonstrated the existence of multiple isoforms of PTX substrates by two-dimensional anal- ysis and in particular two ADP-ribosylated Goa iso- forms from a mixture of bovine brain G proteins (Bra- bet et al., 19886). Goldsmith et al. (1988) likewise identified two forms of Goa during the purification of G proteins from bovine brain. In the present study, we clearly show that the expression of these G,a isoforms depends on the differentiated state of the neuronal cell.
Because Go is a substrate of PTX-catalyzed ADP- ribosylation, the labeling with PTX in the presence of [32P]NAD was first used to identify the Goa subunit in NlE-I 15 cells. In growth medium, consisting of
optiMEM plus 10% fetal calf serum, these cells rapidly divided and exhibited an undifferentiated round mor- phology. These cells contain three PTX-sensitive G proteins, characterized by their a-subunits of apparent M, 39,000, 40,000, and 41,000 as resolved by SDS- PAGE. The substrate at M, 40,000 was the most heavily labeled. Recently, Luetje and Nathanson (1988) were able to detect in the same cells two PTX substrates that they identified as Go and Gi with monospecific anti- bodies. Treatment of neuroblastoma cells with DMSO has been previously described to obtain morphologi- cally differentiated neuronal cells (Kimhi et al., 1976). These cells exhibited an inhibition of cell growth, an extension of long neurites, and an enhancement in the electrical excitability. We found that PTX always cat- alyzed ADP-ribosylation of three substrates, but the labeling of the M, 39,000 polypeptide increased twofold during differentiation. This increase correlated very
J. Nrurochem., Vol. 54, No. 4, 1990

1318 P. BRABET ET AL.
well with that of the G,a level assessed by immuno- blotting, whereas the GP level, which is known to modulate PTX- and CTX-catalyzed ADP-ribosylation, was barely affected. This fact was in accordance with the absence of an enhancement of PTX-dependent la- beling of and Gia4, and the absence of a decrease of CTX-dependent labeling of G,a during neuroblas- toma cell differentiation (data not shown). Further- more, immunoblot assays with the G,a-peptide anti- body show that the level of Gsa was unchanged. Thus, the increase in the amount of G,a is one of the selective events associated with the differentiation process in neuroblastoma N 1 E- 1 15 cells. This result agrees with a recent report (Mullaney et al., 1988) on neuroblas- toma X glioma hybrid cells showing a marked increase in the amount of Goa with little alteration in &subunit amount in these cells induced to differentiated by di- butyryl cyclic AMP treatment.
We show here, using two-dimensional electropho- retic analysis, that the increase in the amount of G,a is related to the temporal expression of two isoforms of G,a. Thus, only the more basic isoform is detected in undifferentiated neuroblastoma cells, with the more acidic isoform appearing in the differentiated state of these cells. In addition, we show that primary cultures of mature neurons contain almost exclusively the more acidic form of Goa (PI 5.55). In agreement with these data, Goldsmith et al. (1988) have reported that among the two forms of G,a purified from bovine brain, the more acidic form (PI 5.6) was the most abundant. The detection of both isoforms in astroglial cells indicated that their expressions are not restricted to neuronal cells. However, the observations that the G,a isoform proportion quite differs in neurons and astroglial cells from mouse fetuses and that G,a immunoreactivity was associated mostly with the neuronal elements in the adult brains (Worley et al., 1986; Gabrion et al., 1988) suggest that change in Goa isoform expression may occur differently in neurons and astroglial cells.
When these isoforms were separated onto IEF gels, they showed very distinct mobilities, one migrating at PI of 5.8 and the other at a more acidic PI of -5.55. In contrast, both have similar apparent molecular weights on the second dimension, and both are im- munologically indistinguishable on incubation with specific polyclonal G,a antibodies. Furthermore, they are substrates for PTX-catalyzed ADP-ribosylation in whole NlE-115 cells, as revealed by the shift in their migrations due to the negative charge of the ADP-ri- bose. This indicates that in cells, these two isoforms are able to associate with Pr, because the heterotrimeric GaPr is required for PTX-catalyzed ADP-ribosylation. Because two G,a isoforms can coexist in the same cell type, do they result from translation of different mes- sages or from posttranslational modifications? Re- cently, several other investigators (Brann et al., 1987; Jones and Reed, 1987; Luetje et al., 1988) found mul- tiple G,a messages in Northern blot analysis. However, we do not know if these messages were assigned to
multiple G,a isoforms. Ga components have been shown to undergo at least three posttranslational mod- ifications: intrinsic ADP-ribosylation (Jacquemin et al., 1986; Tanuma et al., 1988), phosphorylation (Zick et al., 1986; Watanabe et al., 1988), and myristylation (Buss et al., 1987; Schultz et al., 1987). Intrinsic ADP- ribosylation of one of the two G,a isoforms can be excluded. Indeed, we have shown that both can be ADP-ribosylated by PTX in whole cells, as indicated by the absence of back ADP-ribosylation (data not shown) and by their shift of PI. Metabolic labeling of differentiated N 1 E- 1 15 cells with [3H]myristic acid and two-dimensional analysis revealed that both G,a iso- forms are labeled, a result suggesting that they are co- valently acylated (Fig. 6). Phosphorylation of PTX substrates (the a-subunit of T and Gi) by protein kinase C and A has been reported in vitro and in vivo (Zick et al., 1986; Watanabe et al., 1988), a finding suggesting that G,a may be also a substrate for phosphorylation. The difference in PI between the G,a isoforms (-0.25 unit) is greater than that observed after addition of an ADP-ribose (0.15 unit). Therefore, if the difference in PI between the two isoforms were the result of a phos- phorylation, one would expect that more than two phosphates would be incorporated in the most basic isoform. However, in vitro phosphorylation of G pro- teins generally resulted in < 1 phosphate/mol. Exper- iments to verify this point have to be done. Whatever the nature of the difference between the two isoforms, it is interesting to note that preliminary results indicate that the more acidic isoform has a turnover consider- ably lower than that of the more basic isoform.
Our recent data devoted to the ultrastructural lo- calization of Go in neurons suggested that Goa, asso- ciated or not with &, may bind to cytoplasmic com- partments in addition to its expected distribution under the plasma membrane (Gabrion et al., 1988). Immu- nofluorescence data on N 1E- 1 15 cells confirmed that G,a is located both on the plasma membrane and in the cytoplasmic matrix. However, the lack of immu- noreactivity in the soluble fraction indicates that G,a remains closely attached to a nonsoluble matrix. Moreover, we observed that on differentiation of N 1 E- l 15 cells, localization of G,a is to a great extent shifted from the cytoplasm to the plasma membrane. This main finding suggests that induction of the more acidic form of G,a may correlate with its association to the plasma membrane. However, only specific probes for each isoform would be able to verify this hypothesis.
The differential expression of two distinct isoforms of G,a and their apparently different cellular localiza- tion raise important questions regarding analysis of Go protein function. In neuronal cells, some evidence ex- ists that neurotransmitter receptors can be coupled to K+ and Ca2+ channels through PTX-sensitive G pro- teins. For instance, inhibition of voltage-sensitive Ca” channels has been demonstrated with a2-adrenergic, GABAs, dopaminergic, opiate, and muscarinic ace- tylcholine receptors (Holz et d., 1986; Hescheler et al.,
J. Neurochern.. Val. 54, No. 4, 1990

NEURONAL DIFFERENTIATION AND Goa I S O F O W S 1319
1987; Wanke et al., 1987; Harris-Warrick et al., 1988). In most cases, a role of Go has been proposed. There- fore, it is possible that expression of Go and receptors coupled to voltage-sensitive Ca2+ channels are be under coordinated regulation. Luetje and Nathanson ( 1988) recently reported that inactivation of voltage-sensitive Ca2+ channels, due to chronic membrane depolariza- tion of N1E-115 cells, regulates both muscarinic re- ceptor and G0a density. This treatment had no effect on Gia and GP levels. The results described here suggest that such a regulation may be associated with the expression of isoforms of the Go protein having diverse biochemical characteristics and possibly functions. The significance of this Goa diversity and the exact nature of the isoform remain to be determined.
Acknowledgment: The G,a protein purified from turkey erythrocytes, the bovine retinal T, and S49 lymphoma cells were kindly provided by Drs. M. Heckman, M. Chabre, and B. A. Harris, respectively. We are grateful to Drs. A. Dumuis and M. Sebben for their help in the preparation of primary cultures of mouse fetus brain cells a n d to J. Laur for her skillful technical assistance in the G,a-peptide synthetic pro- cesses. Dr. J. C. Bonnafous is gratefully acknowledged for constructive discussions. We would also like to thank A. Turner-Madeuf, A. Sahuquet, and M. Passama for preparing the manuscript.
REFERENCES Asano T., Ui M., and Ogasawara N. (1985) Prevention ofthe agonist
binding to y-aminobutyric acid B receptors by guanine nucleo- tides and islet-activating protein, pertussis toxin, in bovine ce- rebral cortex. J. Biol. Chein. 260, 12653-12658.
Asano T., Semba R., Kamiya N., Ogasawara N., and Kato K. (1988~) Go, a GTP-binding protein: immunochemical and immunohis- tochemical locahation in the rat. J. Neurochem. 50, 1 164- 1 169.
Asano T., Kamiya N., Semba R., and Kato K. (19886) Ontogeny of the GTP-binding protein Go in rat brain and heart. J. Nrurochem. 51, 1711-1716.
Birnhaumer L., Codina J., Mattera R., Yatani A., Scherer N., Tor0 M., and Brown A. M. (1987) Signal transduction by G proteins. Kidney Inr. 32(Suppl), S14-S37.
Blache P., Kervran A., Martinez J., and Bataille D. (1988) Devel- opment of an oxyntomodulin/glicentin C-terminal radioim- munoassay using a “thiol-maleoyl” coupling method for pre- paring the immunogen. Anal. Biochem. 173, 15 1-1 59.
Brabet P., Dumuis A,, Sebben M., Pantaloni C., Bockaert J., and Homburger V. (1988~) Immunocytochemical localization of the guanine nucleotide-binding protein Go in primary cultures of neuronal and glial cells. J. Neurosci. 8, 701-708.
Brabet P., Pantaloni C., Rouot B., Toutant M., Garcia-Sainz A,, Bockaert J. , and Homburger V. (1988b) Multiple species and isoforms of Bordetella pertussis toxin substrates. Biochem. Bio- phys. Res. Commun. 152, 1185-1 192.
Brann M. R., Collins R. M., and Spiegel A. (1987) Localization of mRNAs encoding the a-subunits of signal-transducing G-pro- teins within rat brain and among peripheral tissues. FEBS Lett.
Buss J. E., Mumby S. M., Casey P. J., Gilman A. G., and Sefton B. M. (1987) Myristoylated a subunits of guanine nucleotide- binding regulatory proteins. Proc. Natl. Acad. Sci. USA 84,7493- 7497.
Cerione R. H., Regan J. W., Nakata H., Codina J., Benovic J. L., Gierschik P., Somers R. L., Spiegel A. M., Birnbaumer L., Lef- kowitz R. J., and Caron M. G. (1986) Functional reconstitution of the a,-adrenergic receptor with guanine nucleotide regulatory
222, 191-198.
proteins in phospholipid vesicles. J. Biol. Chem. 261, 3901- 3909.
Ewald D. A., Sternweis P. C., and Miller R. J. (1988) Guanine nu- cleotide-binding protein Go-induced coupling of neuropeptide Y receptors to Ca2* channels in sensory neurons. Proc. Natl. Acad. Sci. USA 85, 3633-3637.
Gabrion J., Brabet P., Nguyen Than Dao B., Homburger V., Dumuis A., Sebben M., Rouot B., and Bockaert J . (1988) Ultrastructural localization of the GTP-bindmg protein Go in neurons. Cell. Signal. 1, 107-123.
Gierschik P., Milligan G., Pines M., Goldsmith P., Codina J., Klee W., and Spiegel A. (1986) Use of specific antibodies to quantitate the guanine nucleotide-binding protein Go in brain. Proc. Natl. Acad. Sci. USA 83,2258-2262.
Goldsmith P., Backlund P. S., Rossiter K., Carter A., Milligan G., Unson C. G., and Spiegel A. (1988) Purification of heterotrimeric GTP-binding proteins from brain: identification of a novel form of Go. Biochem. J . 27, 7085-7090.
Harris-Wamck R., Hammond C., Paupardin-Tritsch D., Homburger V., Rouol B., Bockaert J., and Gerschenfeld H. M. (1988) An am subunit of a GTP-binding protein immunologically related to Go mediates a dopamine-induced decrease of Ca2+ current in snail neurons. Neuron 1,27-32.
Hescheler J., Rosenthal W., Trautwein W., and Schultz G. (1987) The GTP-binding protein, Go, regulates neuronal calcium channels. Nature 325, 445-447.
Holz G. G., Rane S. G., and Dunlap K. (1986) GTP-binding protein mediate transmitter inhibition of voltage-dependent calcium channel. Nature 319,670-672.
Homburger V., Brabet P., Audigier Y., Pantaloni C., Bockaert J., and Rouot B. (1987) Immunological localization of the GTP- binding protein Go in different tissues of vertebrates and inver- tebrates. Mol. Pharmacol. 31, 313-319.
Itoh H., Toyama R., Kazasa T.: Tzukamoto T., Matsuoka M., and Kaziro Y. (1988) Presence of three distinct molecular species of Gi protein a subunit. J. Biol. Chem. 263, 6656-6664.
Jacquemin C., Thibout H., Lambert B., and Correze C. (1986) En- dogenous ADP-ribosylation of G, subunit and autonomous reg- ulation of adenylate cyclase. Nature 323, 182-184.
Jones D. T. and Reed R. R. (1987) Molecular cloning of five GTP- hinding protein cDNA species from rat olfactory neuroepithe- lium. J. Biol. Chem. 262, 14241-14249.
Kimhi Y., Palfrey C., Spector I., Barak Y., and Littauer U. Z. (1976) Maturation of neuroblastoma cells in the presence of dimeth- ylsulfoxide. Proc. Natl. Acad. Sci. USA 73, 462-466.
Klotz S. M. and Heiney R. E. ( 1 962) Introduction of sulfhydryl groups into proteins using acetylmercaptosuccinic anhydride. Arch. Biochem. Biophys. 96,605-6 12.
Kurosc H., Katada T., Haga T., Haga K., Ichiyama A., and Ui M. ( I 986) Functional interaction of purified muscarinic receptors with purified inhibitory guanine nucleotide regulatory proteins reconstituted in phospholipid vesicles. J. Biol. Chem. 261,6423- 6428.
Laemmli U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680- 685.
Luetje C. W. and Nathanson N. M. (1988) Chronic membrane de- polarization regulates the level of the guanine nucleotide-binding protein G,a in cultured neuronal cells. J. Neurochem. 50,1775- 1782.
Luetje C:. W., Tietje K. M., Christian J. L., and Nathanson N. M. ( 1988) Differential tissue expression and developmental regu- lation of guanine nucleotide binding proteins and their messenger RNAs in rat heart. J. Biol. Chem. 263, 13357-13365.
Mullaney l., Magee A. I., Unson C. G., and Milligan G. (1988) Dif- ferential regulation of amounts of the guanine-nucleotide-binding proteins Gi and Go in neuroblastoma X glioma hybrid cells in response to dibutyryl cyclic AMP. Biochem. J. 256, 649-656.
Mumby S., Pang I., Gilman A. G., and Sternweis P. C. (1988) Chrc- matographic resolution and immunologic identification of the am and a4, subunits of guanine nucleotide-binding regulatory proteins from bovine brain. J. Biol. Chem. 263, 2020-2026.
J. Neurochem., Vol. 54, No. 4, 1990

1320 P. BRABET ET AL.
Neer E. J . and Clapham D. E. (1988) Roles ofG protein subunits in transmembrane signalling. Nufirre 333, 129- 134.
Neer E. J., Lock J. M., and Wolf L. G. (1984) Purification and prop- erties of the inhibitory guanine nucleotide regulatory unit of brain adenylate cyclase. J. Biol. Chem. 259, 14222-14229.
O'Farrell P. H. ( 1975) High resolution two-dimensional electropho- resis of proteins. J. Bid. c'hem. 250, 4007-402 I .
Otey C. A., Kalnoski M. H., Lessard J. L., and Bulinski J . C. (1986) Immunolocalization of the gamma isoform of nonmuscle actin in cultured cells. J. Cell Biol. 102, 1726-1737.
Schultz A. M., Tsai S., Kung H., Oroszlan S., Moss J., and Vaughan M. (1987) Hydroxylamine-stable covalent linkage of myristic acid in G,u, a guanine nuclcotide-binding protein of bovine brain. Biochem. Biophys. Rcs. Cornrnun. 146. 1234- 1239.
Senogles S. E., Benovic J . L., Amlaiky N., Unson C.. Milligan G., Vinitsky R., Spiegel A. M., and Caron M. G. (1987) The D2- dopamine receptor of anterior pituitary is functionally associated with a pertussis toxin-sensitive guanine nucleotide binding pro- tein. J. B i d . Chern. 262,4860-4867.
Sternweis P. C. and Robishaw J. D. (1984) Isolation of two proteins with high affinity for guanine nucleotides from membranes of bovine brain. J. Bid. Chem. 259, 13806- I38 13.
Tanuma S., Kawashima K., and Endo H. (1988) Eukaryotic mono(ADP-ribosy1)transferase that ADP-ribosylates GTP- binding regulatory G; protein. J . Bid. Chern. 263, 5485-5489.
Toutant M., Aunis D., Bockaert J., Homburger V., and Rouot B. (1987) Presence of three pertussis toxin substrates and Go im-
munoreactivity in both plasma and granule membranes of chro- maffin cells. FEBS Lett. 215, 339-344.
VanDongen A. M. J., Codina J., Olate J., Mattera R., Joho R., Birn- baumer L., and Brown A. M. (1988) Newly identified brain potassium channels gated by the guanine nucleotide binding protein Go. Science 242, 1433-1436.
Wanke E., Ferroni A,, Malgoroli A., Ambrosini A., Pozzan T., and Mcldolesi J. (1987) Activation ofa muscarinic receptor selectively inhibits a rapidly inactivated Ca2+ current in rat sympathetic neurons. Pror. Nutl. Acud. Sci. USA 84, 43 13-43 17.
Watanabe Y., Imaizumi T., Misaki N., lwakura K., and Yoshida H. (1988) Effects of phosphorylation of inhibitory GTP-binding protein by cyclic AMP-dependent protein kinase on its ADP- ribosylation by pertussis toxin, islet-activating protein. E'EBS
Watkins D. C., Northup J. K., and Malbon C. C. (1987) Regulation of G-protein in differentiation. J. Bid. Chem. 262, 1065 1-10657.
Worley P. F., Baraban J . M., Van Dop C., Neer E. J., and Snyder S. H. (1986) Go, a guanine nucleotide binding protein: immu- nohistochemical localization in rat brain resembles distribution of second messenger systems. Proc. Nutl. Acad. Sci. US4 83, 456 1-4565.
Zick Y., Sagi-Eisenberg R., Pines M., Gierschik P., and Spiegel A. M. (1986) Multiple phosphorylation of the 01 subunit of transducin by the insulin receptor kinase and protein kinase C. Proc. Nutl. Acad. Sci. USA 83, 9294-9297.
Lett. 236, 372-374.
J. Neurochem., Vol. 54, No. 4, 1990
Copyright © 2022 FDOKUMEN




















