
Frequent detection of human cytomegalovirus in neuroblastoma: A novel therapeutic target?
11
Frequent detection of human cytomegalovirus in neuroblastoma: A novel therapeutic target? Nina Wolmer-Solberg 1 , Ninib Baryawno 2 , Afsar Rahbar 1 , Dieter Fuchs 1 , Jenny Odeberg 1 , Chato Taher 1 , Vanessa Wilhelmi 1 , Jelena Milosevic 2 , Abdul-Aleem Mohammad 1 , Tommy Martinsson 3 , Baldur Sveinbj€ ornsson 2,4 , John Inge Johnsen 2 , Per Kogner 2 and Cecilia S€ oderberg-Naucl er 1 1 Department of Medicine, Center for Molecular Medicine, Karolinska University Hospital in Solna, Karolinska Institutet, Stockholm, Sweden 2 Department of Women’s and Children’s Health, Childhood Cancer Research Unit, Karolinska Institutet, Stockholm, Sweden 3 The Sahlgrenska Academy at the University of Gothenburg, Genomics Core Facility, G€ oteborg 4 Immunology Research Group, Faculty of Medicine, University of Troms€ o, Troms€ o, Norway Neuroblastoma is the most common and deadly tumor of childhood, where new therapy options for patients with high-risk disease are highly warranted. Human cytomegalovirus (HCMV) is prevalent in the human population and has recently been implicated in different cancer forms where it may provide mechanisms for oncogenic transformation, oncomodulation and tumor cell immune evasion. Here we show that the majority of primary neuroblastomas and neuroblastoma cell lines are infected with HCMV. Our analysis show that HCMV immediate-early protein was expressed in 100% of 36 primary neuroblas- toma samples, and HCMV late protein was expressed in 92%. However, no infectious virus was detected in primary neuroblas- toma tissue extracts. Remarkably, all six human neuroblastoma cell lines investigated contained CMV DNA and expressed HCMV proteins. HCMV proteins were expressed in neuroblastoma cells expressing the proposed stem cell markers CD133 and CD44. When engrafted into NMRI nu/nu mice, human neuroblastoma cells expressed HCMV DNA, RNA and proteins but did not produce infectious virus. The HCMV-specific antiviral drug valganciclovir significantly reduced viral protein expression and cell growth both in vitro and in vivo. These findings indicate that HCMV is important for the pathogenesis of neuroblastoma and that anti-viral therapy may be a novel adjuvant treatment option for children with neuroblastoma. Neuroblastoma is an embryonal tumor that arises in the sympathetic nervous system, including the adrenal medulla, sympathetic ganglia, and paraganglia. These tumors are the most common and deadly tumors of childhood, and clinically and biologically heterogeneous—one favorable subset is prone to spontaneous apoptosis or differentiation over time with no or little therapy—whereas most neuroblastomas are metasta- sizing high-risk tumors that are difficult to cure with current treatment options. 1 Except for a small minority of patients with specific germline mutations of the anaplastic lymphoma kinase gene, 2 there is no clear evidence for a familial predis- position to neuroblastoma. Thus, modifiable risk factors, including viruses as an additional triggering factor, may be important in the etiology of these tumors. Relapse of neuroblastoma has been related to active infec- tion with human cytomegalovirus (HCMV). 3 Viral particles in urine and HCMV-specific antibodies and DNA in the blood at time of diagnosis have been reported. 3,4 In long- term in vitro culture, neuroblastoma cells persistently infected with HCMV exhibit enhanced Mycn expression, tumor Key words: human cytomegalovirus, neuroblastoma, valganciclovir Additional Supporting Information may be found in the online version of this article. The first two authors contributed equally to this work and share first and second authorship. Last two authors contributed equally to this work and share senior authorship. Grant sponsor: Torsten and Ragnar S€ oderbergs Stiftelse and Ragnar S€ oderbergs Foundation; Grant sponsor: The Swedish Children’s Cancer Foundation; Grant numbers: PROJ12/115 and PROJ10/121; Grant sponsor: The Swedish Cancer Society; Grant number: 100637; Grant sponsor: The Swedish Research Council; Grant numbers: K2010-56X-12615-13-3 and K2009-56P-20937-03-1; Grant sponsors: Stichting af Jochnicks Foundation, BILTEMA Foundation, Sten A. Olssons Foundation, the M€ arta and Gunnar V. Philipson Foundation, The Mary Bev e Foundation, The Hans and M€ arit Rausing Charitable Fund, The Dðmman Foundation, Swedish Society for Medical Research (SLS), Goljes Memory Foundation, Magnus Bergvalls Foundation, Swedish Society for Medical Research (SSMF) and Tore Nilsons Foundation. DOI: 10.1002/ijc.28265 History: Received 4 Dec 2012; Accepted 24 Apr 2013; Online 10 May 2013 Correspondence to: Cecilia S€ oderberg-Naucl er, CMM L8:03, Group of Cellular and Molecular Immunology, Karolinska Institutet, Karolin- ska University Hospital, SE-171 76 Stockholm, Sweden, Tel.: 146-8-517-79896, Fax: 146-8-31-3147, E-mail: [email protected] Infectious Causes of Cancer Int. J. Cancer: 133, 2351–2361 (2013) V C 2013 UICC International Journal of Cancer IJC
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Frequent detection of human cytomegalovirus in neuroblastoma: A novel therapeutic target?

Frequent detection of human cytomegalovirus inneuroblastoma: A novel therapeutic target?
Nina Wolmer-Solberg1, Ninib Baryawno2, Afsar Rahbar1, Dieter Fuchs1, Jenny Odeberg1, Chato Taher1, Vanessa Wilhelmi1,
Jelena Milosevic2, Abdul-Aleem Mohammad1, Tommy Martinsson3, Baldur Sveinbj€ornsson2,4, John Inge Johnsen2,
Per Kogner2 and Cecilia S€oderberg-Naucl�er1
1 Department of Medicine, Center for Molecular Medicine, Karolinska University Hospital in Solna, Karolinska Institutet, Stockholm, Sweden2 Department of Women’s and Children’s Health, Childhood Cancer Research Unit, Karolinska Institutet, Stockholm, Sweden3 The Sahlgrenska Academy at the University of Gothenburg, Genomics Core Facility, G€oteborg4 Immunology Research Group, Faculty of Medicine, University of Troms€o, Troms€o, Norway
Neuroblastoma is the most common and deadly tumor of childhood, where new therapy options for patients with high-risk
disease are highly warranted. Human cytomegalovirus (HCMV) is prevalent in the human population and has recently been
implicated in different cancer forms where it may provide mechanisms for oncogenic transformation, oncomodulation and
tumor cell immune evasion. Here we show that the majority of primary neuroblastomas and neuroblastoma cell lines are
infected with HCMV. Our analysis show that HCMV immediate-early protein was expressed in 100% of 36 primary neuroblas-
toma samples, and HCMV late protein was expressed in 92%. However, no infectious virus was detected in primary neuroblas-
toma tissue extracts. Remarkably, all six human neuroblastoma cell lines investigated contained CMV DNA and expressed
HCMV proteins. HCMV proteins were expressed in neuroblastoma cells expressing the proposed stem cell markers CD133 and
CD44. When engrafted into NMRI nu/nu mice, human neuroblastoma cells expressed HCMV DNA, RNA and proteins but did not
produce infectious virus. The HCMV-specific antiviral drug valganciclovir significantly reduced viral protein expression and cell
growth both in vitro and in vivo. These findings indicate that HCMV is important for the pathogenesis of neuroblastoma and
that anti-viral therapy may be a novel adjuvant treatment option for children with neuroblastoma.
Neuroblastoma is an embryonal tumor that arises in thesympathetic nervous system, including the adrenal medulla,sympathetic ganglia, and paraganglia. These tumors are themost common and deadly tumors of childhood, and clinicallyand biologically heterogeneous—one favorable subset is proneto spontaneous apoptosis or differentiation over time with noor little therapy—whereas most neuroblastomas are metasta-sizing high-risk tumors that are difficult to cure with currenttreatment options.1 Except for a small minority of patientswith specific germline mutations of the anaplastic lymphoma
kinase gene,2 there is no clear evidence for a familial predis-position to neuroblastoma. Thus, modifiable risk factors,including viruses as an additional triggering factor, may beimportant in the etiology of these tumors.
Relapse of neuroblastoma has been related to active infec-tion with human cytomegalovirus (HCMV).3 Viral particlesin urine and HCMV-specific antibodies and DNA in theblood at time of diagnosis have been reported.3,4 In long-term in vitro culture, neuroblastoma cells persistently infectedwith HCMV exhibit enhanced Mycn expression, tumor
Key words: human cytomegalovirus, neuroblastoma, valganciclovir
Additional Supporting Information may be found in the online version of this article.
The first two authors contributed equally to this work and share first and second authorship. Last two authors contributed equally to this
work and share senior authorship.
Grant sponsor: Torsten and Ragnar S€oderbergs Stiftelse and Ragnar S€oderbergs Foundation; Grant sponsor: The Swedish Children’s Cancer
Foundation; Grant numbers: PROJ12/115 and PROJ10/121; Grant sponsor: The Swedish Cancer Society; Grant number: 100637; Grant
sponsor: The Swedish Research Council; Grant numbers: K2010-56X-12615-13-3 and K2009-56P-20937-03-1; Grant sponsors: Stichting af
Jochnicks Foundation, BILTEMA Foundation, Sten A. Olssons Foundation, the M€arta and Gunnar V. Philipson Foundation, The Mary Bev�e
Foundation, The Hans and M€arit Rausing Charitable Fund, The Dðmman Foundation, Swedish Society for Medical Research (SLS), Goljes
Memory Foundation, Magnus Bergvalls Foundation, Swedish Society for Medical Research (SSMF) and Tore Nilsons Foundation.
DOI: 10.1002/ijc.28265
History: Received 4 Dec 2012; Accepted 24 Apr 2013; Online 10 May 2013
Correspondence to: Cecilia S€oderberg-Naucl�er, CMM L8:03, Group of Cellular and Molecular Immunology, Karolinska Institutet, Karolin-
ska University Hospital, SE-171 76 Stockholm, Sweden, Tel.: 146-8-517-79896, Fax: 146-8-31-3147, E-mail: [email protected]
Infectious
Cau
sesof
Can
cer
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC
International Journal of Cancer
IJC

growth, invasiveness, and metastasis.5,6 About 20 to 30% ofneuroblastomas exhibit MYCN amplification, which is associ-ated with aggressive, poorly differentiated tumors that pro-gress rapidly and have a poor prognosis.1
HCMV infects 70 to 90% of the world’s population, andis found in 0.5 to 2.2% of newborns, making it the mostcommon congenital viral infection. Symptoms of a primaryHCMV infection are generally mild or asymptomatic inimmunocompetent individuals but may be life-threatening inimmunocompromised patients such as transplant recipientsand AIDS patients. The virus is spread through all bodily flu-ids and establishes a life-long latent/persistent infection.Reactivation from latency appears to be triggered by inflam-mation, which the virus can itself initiate by inducing cyto-kine and chemokine production and by enhancingleukotriene and prostaglandin synthesis, which induce expres-sion of 5-lipoxygenase and cyclooxygenase-2 (COX-2). Werecently demonstrated that HCMV positive tumor cellsexpress COX-2 in medulloblastoma tumors. Anti-viral ther-apy in combination with a COX-2 inhibitor resulted in a72% reduction in tumor growth.7
The prevalence of HCMV is surprisingly high in severalforms of cancer, including brain tumors and cancers of thecolon, breast, and prostate,8–12 and rhabdomyosarcoma.13
The active viral infection is restricted to tumor cells andpresent in over 90% of these tumor types; healthy tissuesfrom the same patients are consistently HCMV nega-tive.8,9,11,12 Low levels of HCMV infection in glioblastomas isassociated with improved survival,14,15 which suggest a roleof HCMV in tumor progression. HCMV is not considered tobe oncogenic, but it may exert oncomodulatory effects thatcontribute to cancer development.16 The 230-kb genome ofHCMV has 252 open reading frames was believed to encodeapproximately 180 proteins. Of these, only about 50 areknown to be essential for viral replication. Thus, mostHCMV proteins fulfill other functions in the viral life cycle.Through sophisticated mechanisms, viral proteins control cel-lular and immunological functions relevant to tumor biologyand immune surveillance.17 HCMV proteins control cellulardifferentiation, proliferation, and epigenetic functions, induce
migration and angiogenesis, inhibit apoptosis, and controlimmunological functions that help virus-infected tumor cellsto avoid detection and elimination by the immune systemand to support efficient viral replication. In this study, wereport a high prevalence of HCMV in human neuroblastomasamples, cell lines, and xenografts. Inhibition of viral replica-tion with ganciclovir significantly inhibited tumor growthboth in vitro and in vivo, suggesting anti-HCMV treatmentas a novel adjuvant therapeutic option for neuroblastomapatients.
Material and MethodsHuman neuroblastoma tumor tissue
Tumor tissue was collected from 40 patients, 36 with neuro-blastoma including three non-tumor adrenal gland samples,and four with ganglioneuroma undergoing surgery at AstridLindgren Children’s Hospital, Stockholm, Sweden. Ethicalapproval was obtained from the Karolinska University Hospi-tal Research Ethics Committee. Parents to eligible pediatricneuroblastoma patients gave informed consent to donatetumor tissue to be used for research purpose (approval no.2009/1369-31/1, 03-736). Ethical permission to examine thesetumor samples specifically for CMV was also obtained(approval no. 2008/628-31/2). The tissue was snap frozen inliquid nitrogen and stored at –80�C. For immunohistochem-istry, tissues were thawed, fixed in paraformaldehyde, embed-ded in paraffin, sectioned at 5 lm, and placed on SuperfrostPlus glass slides. Frozen tumor tissue from 24 patients waskept at –80�C until extraction of DNA, or RNA extraction(16 samples).
Cell cultures
Human neuroblastoma cell lines (SK-N-BE(2), SK-N-AS, SK-N-DZ, Kelly and IMR-32) and human chronic myelogenousleukemia cells (K562) were cultured in RPMI 1640 medium(Gibco BRL, Sundbyberg, Sweden). SH-SY5Y neuroblastomacells and human fibroblasts were grown in MEM medium(Life Technologies, Paisley, Scotland). All media were supple-mented with 10% fetal calf serum (Sigma-Aldrich, Solna,Sweden), 2 mM L-glutamine (Gibco BRL), and 100 U/ml
What’s new?
Relapse and invasiveness of neuroblastoma, a frequently fatal cancer of early childhood, may be linked to the presence of
human cytomegalovirus (HCMV), one of the most common congenital viral infections known. In this study, HCMV was observed
in primary neuroblastoma tumors and in six neuroblastoma cell lines. Although no infectious virus was isolated from tumors,
the HCMV-specific drug valganciclovir significantly reduced viral protein expression and tumor cell growth both in vitro and in
vivo. The results suggest that HCMV may be important in the pathogenesis of neuroblastoma and that antiviral therapy may
represent a possible future treatment option for affected children. We have shown that all examined primary neuroblastoma
tumors and six neuroblastoma cell lines were infected with HCMV, but no infectious virus was isolated from tumors. The
HCMV-specific drug Valganciclovir significantly reduced viral protein expression and tumor cell growth in vitro and in vivo.
Thus, HCMV may be important in the pathogenesis of neuroblastoma and anti-viral therapy may provide a novel treatment
option for children with neuroblastoma.
Infectious
Cau
sesof
Can
cer
2352 Human cytomegalovirus in neuroblastoma
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC

penicillin/streptomycin (Gibco BRL) at 37�C in 5% CO2/95%air. The authenticity of the cell lines was verified by usingthe STR DNA profiling test (Genetica DNA Laboratories,Cincinnati). The latest verification was performed in Septem-ber 2010.
Virus isolation
Frozen neuroblastoma tissue (n 5 8) and neuroblastoma cellxenografts (of six cell lines) was homogenized by ultra soni-cation. Homogenate (0.5–1 ml) suspended in serum freemedium and added to human fibroblasts, grown in six-wellplates to a confluency of 50 to 70%. After 24 hr low serummedium was added. Half of the medium was exchanged twiceweekly and after 2 weeks the cells were washed in PBS, fixedin cold methanol:acetone 1:1 for 10 min in room temperatureand stained for IEA.
Chemicals
Ganciclovir (Cymvene, Roche, Basel, Switzerland) and val-ganciclovir (Valcyte; Roche, Stockholm, Sweden) were dis-solved in water according to manufacturer’s guidelines. Alldrugs were further diluted in Opti-MEM (Gibco BRL) to thedesired in vitro concentration.
Detection of HCMV in tumor specimens
For detailed methods regarding detection of HCMV proteins,DNA and RNA in tumor tissues, please see SupportingInformation.
Clonogenic assay
Tumor cell growth was assessed with in vitro clonogenicassays as described.18 After surface attachment, cells weretreated with 150 or 300 lM ganciclovir in Opti-MEM (LifeTechnologies, Paisley, Scotland) for 48 hrs. The medium wasthen replaced with drug-free complete medium. When con-trol plates reached sufficient number of colonies, cells werefixed in formaldehyde and stained with Giemsa (GIBCOBRL). The number of colonies consisting of >75 cells with50% plating efficiency were counted manually. For each treat-ment combination, the surviving fraction was calculated asthe ratio of the mean plating efficiency of treated cells to thatof untreated control cells.
Cell viability assay
Cell viability was measured using a colorimetric assay for 96-well plates with 2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium monosodium salt (WST-1)reagent (Roche). Neuroblastoma cells were seeded at a con-centration of 1,250 cells per well, allowed to attach for 24 hand treated with ganciclovir daily for 7 days at the concen-trations 300, 600, 900 or 1,800 lM. After 7 days, 10 ll ofWST-1 were added and cells were incubated for an addi-tional 3 hr. Cytotoxicity of ganciclovir at different concentra-tions was expressed as percentage of viable cells compared tocontrols.
Xenografts and in vivo administration of valganciclovir
Female NMRI nu/nu mice (Taconic Laboratories, Ejby, Den-mark) 4 to 6 weeks of age were maintained at five mice percage and were given sterile water and food ad libitum. Eachmouse was subcutaneously injected with SH-SY5Y neuroblas-toma cells (20 3 106) in both hind legs. Treatment wasstarted when palpable tumors reached a volume of 0.15 ml.Mice were randomized to receive either no treatment (n 5
10) or valganciclovir at 20 mg/kg (n 5 7) or 30 mg/kg (n 5
7). Valganciclovir was administered twice daily through agastric feeding tube. Tumors were measured daily, and tumorvolume was calculated as (width)2 3 length 3 0.44. Tumorvolume index was calculated as the measured volume dividedby the volume measured at start of treatment. Tumor weightwas recorded at autopsy, after which a part of the tumorwere fixed in formaldehyde and a part was frozen.
All animal experiments were approved by the regionalethics committee for animal research (nos. N231/05 andN304/08) in accordance with animal protection regulations(SFS 1988:534) and regulations of the Swedish NationalBoard for Laboratory Animals (SFS 1988:541). A total of 55mice were used in this experiment.
Statistical analysis
All statistical analyses were performed with GraphPad Prismsoftware (GraphPad Software, San Diego, CA). For in vitrostudies, the t-test (two-tailed) was used to determine whetherthe mean of a single sample differed significantly from con-trol. Tumor growth and tumor weight was analyzed by MannWhitney test. p < 0.05 was considered significant.
ResultsHigh prevalence of HCMV in primary neuroblastoma
tumors
The link between HCMV and neuroblastoma was first pro-posed by Nigro et al., who observed a correlation between anactive HCMV infection and neuroblastoma patients whoexcreted HCMV in urine at the time of clinical diagnosis.3,19
Here, we examined primary tumor biopsies from 36 childrenwith neuroblastoma for HCMV. All 36 expressed HCMVimmediate-early (IE) protein, and 33 (92%) expressed lateprotein; IE and late proteins were widely expressed in tumorcells (Fig. 1 and Table 1). The number of positive cells wasestimated manually, and samples were graded according topercentage of cells infected with IE and late proteins (Fig. 1iand Table 1). Patients of all ages and genetic subsets andclinical risk groups were included20 and all containedHCMV. We also investigated four samples from differenti-ated ganglioneuromas and three samples from healthy adre-nal tissues, two of which from children with neuroblastoma.All these samples were also positive for HCMV DNA andprotein expression (Table 1). In the lack of an HCMV nega-tive neuroblastoma tissue sample, we included an HCMVnegative glioblastoma tumor sample as a control for the
Infectious
Cau
sesof
Can
cer
Wolmer-Solberg et al. 2353
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC

immunohistochemical staining (Figs. 1d and 1h). Expressionof HCMV IE (Fig. 1k) and late proteins was also demon-strated in two fresh neuroblastoma samples by flow cytome-try, which revealed HCMV proteins in 14% of cells in onesample and 54% of cells in the other. We next performedwestern blot analyses of seven frozen primary neuroblastomasamples using an antibody to the IE proteins IE72 and IE86(clone 810R) and found that IE72 but not IE86 wereexpressed in all primary neuroblastoma examined (Fig. 1l).However, we detected a band that is predicted to be IE55,which is a splice variant of IE86, in seven of seven samples(Fig. 1l). DNA was extracted from 24 frozen neuroblastomaspecimens; 22/24 were positive for HCMV IE and/or pp150genes by PCR (data not shown). Sixteen of 16 frozen sampleswere positive for HCMV RNA (data not shown). In five sam-ples, HCMV IE DNA PCR products were sequenced andfound to contain the HCMV genome, which all were dis-tinctly different from four laboratory isolates (Merlin,AD169, TB40E and VR1814) (data not shown) and excludedthe possibility of contamination. However, viral cultures fromeight frozen samples were consistently negative (data notshown), implying defective HCMV replication in neuroblas-tomas, or very low virus presence, but the possibility of fail-ure to grow the tumor associated virus by conventionaltechniques for virus isolation can not be excluded. Theseobservations suggest that HCMV proteins rather than pro-duction of infectious virus may be involved in the pathogene-sis of neuroblastoma.
Detection of HCMV in neuroblastoma cell lines and
neuroblastoma xenografts
We next analyzed six neuroblastoma cell lines (SH-SY5Y,SK-N-BE(2), SK-N-AS, SK-N-DZ, Kelly, and IMR-32) andfound that all six were positive for HCMV DNA by PCR(Supporting Information Table 1). Fluorescence in situhybridization (FISH) with an HCMV probe (Fig. 2a and Sup-porting Information Table 1) also showed HCMV DNA inall six cell lines. Human chronic myelogenous leukemia cells
(K562) and human fibroblasts (MRC-5) infected with HCMVAD169 served as negative and positive controls, respectively(Fig. 2a). The location of viral DNA differed in neuroblas-toma cells compared to HCMV-infected fibroblasts; viralDNA was located in the periphery of the nucleus in tumorcells but was centrally located (sometimes in owl’s eye forma-tion) in infected fibroblasts.
Flow cytometric analysis of all cell lines further demon-strated HCMV IE antigen and pp65 expression in 0 to 84%of the cells (data not shown and Figs. 2b and 2c). The num-ber of protein-positive cells varied considerably between celllines and at different sampling times; within each cell line,the number of positive cells identified with antibodies againstIE or pp65 was relatively constant at individual samplingoccasions (Figs. 2b and 2c). At least once over 9 months,IMR-32, Kelly, and SY-SH5Y were negative for both HCMVIE antigen and pp65 by flow cytometry (n 5 2–7 samplingoccasions for individual cell lines and was generally low afterthawing of frozen cell lines), which implies a high variationof HCMV protein expression in neuroblastoma cell lines overtime. We hypothesized that the consistent presence ofHCMV DNA and proteins in long-term cultures of neuro-blastoma cell lines may be linked to viral persistence in acancer stem cell or a tumor initiating cell. No clear markerhas been defined for tumor initiating cells in neuroblastomatumors. We double stained neuroblastoma cell lines forHCMV IEA and CD133 or CD44; two potential stem cellmarkers that could be used for double staining with HCMVproteins. Only SK-N-DZ demonstrated high CD133 expres-sion (84%), whereof 32% were double positive for CD133and HCMV IEA (Fig. 2c). CD44 was present on 23% of SK-N-BE(2), 32% of IMR-32, 2.2% of SH-SY5Y, 69% of SK-N-AS and 4.1% of SK-N-SH cells. Double immunofluorescencerevealed that 14% of SK-N-BE(2), 29% of IMR-32, 2% ofSH-SY5Y, 34% of SK-N-AS and 4% of SK-N-SH co-expressed HCMV IE and CD44 (representative examples inFig. 2c). Double staining for CD133 and HCMV IE
Table 1. HCMV protein expression in neuroblastic tumors
Protein expression IE proteins/L proteins
Diagnosis nAge1 (mo) median;range Sex M/F 0 11 21 31 41
Neuroblastoma2 36 18; 0–136 20/16 0/3 9/20 11/5 5/4 11/4
Low/intermediate Risk3 26 14; 0–123 14/12 0/1 6/14 6/4 5/4 9/3
High risk3 10 23; 12–136 6/4 0/2 3/6 5/1 0/0 2/1
Ganglioneuroma2 4 20; 10–137 2/2 0/1 2/ 0/1 1/0 0/0
Healthy adrenal6 3 24; 12–25 1/2 0/0 1/1 1/2 1/0 0/0
1Age in completed months at diagnosis (median; range).2According to INSS diagnostic criteria (International Neuroblastoma Staging System, Brodeur et al., 1993).20
3According to INRG criteria (International Neuroblastoma Risk grouping, Cohn et al., 2009).21
4Metastatic stage M and age >18 months, or MYCN-amplification.5Benign peripheral neuroblastic tumor.6Non-tumoral adrenal medulla.
Infectious
Cau
sesof
Can
cer
2354 Human cytomegalovirus in neuroblastoma
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC

demonstrated a few IE-positive cells expressing CD133 in fro-zen neuroblastoma tissue samples (Fig. 2d). CD44 stainingwas not successful in tissue samples.
To further investigate whether neuroblastoma cell linescould be super-infected with HCMV and establish a produc-tive infection, we infected three neuroblastoma cell lines;Kelly; SK-N-AS and SH-SY5Y with VR1814, MOI 5, and col-lected samples for different analyses of CMV transcripts, pro-teins, and infectious virus at 1 and 7 days post-infection(dpi). All three cell lines were susceptible to infection; anddemonstrated very low HCMV protein expression and tran-script levels in cultures not superinfected by the virus (Sup-porting Information Figs. 1a and 1b). IE transcripts weredetected at 1 dpi and approximately 20 to 25% of cells wereCMV IE positive at 1 dpi. We collected both cells and super-natants of HCMV infected SK-N-AS and SH-SY5Y at 7 dpi,which were positive for HCMV IE and gB DNA (SupportingInformation Table 2) and transferred either supernatants or
cells/supernatants to the same corresponding uninfected cells.The following day, the cells were collected for IE transcriptanalyses or stained for IE protein expression. We observed adose depended increase of IE transcripts in both cell lines(Supporting Information Table 2), and higher frequency ofHCMV IE positive cells in neuroblastoma cultures infectedwith cells and supernatants (data not shown). Thus, HCMVcan establish a productive infection in neuroblastoma celllines at low levels, and the infection appeared to be mainlycell associated.
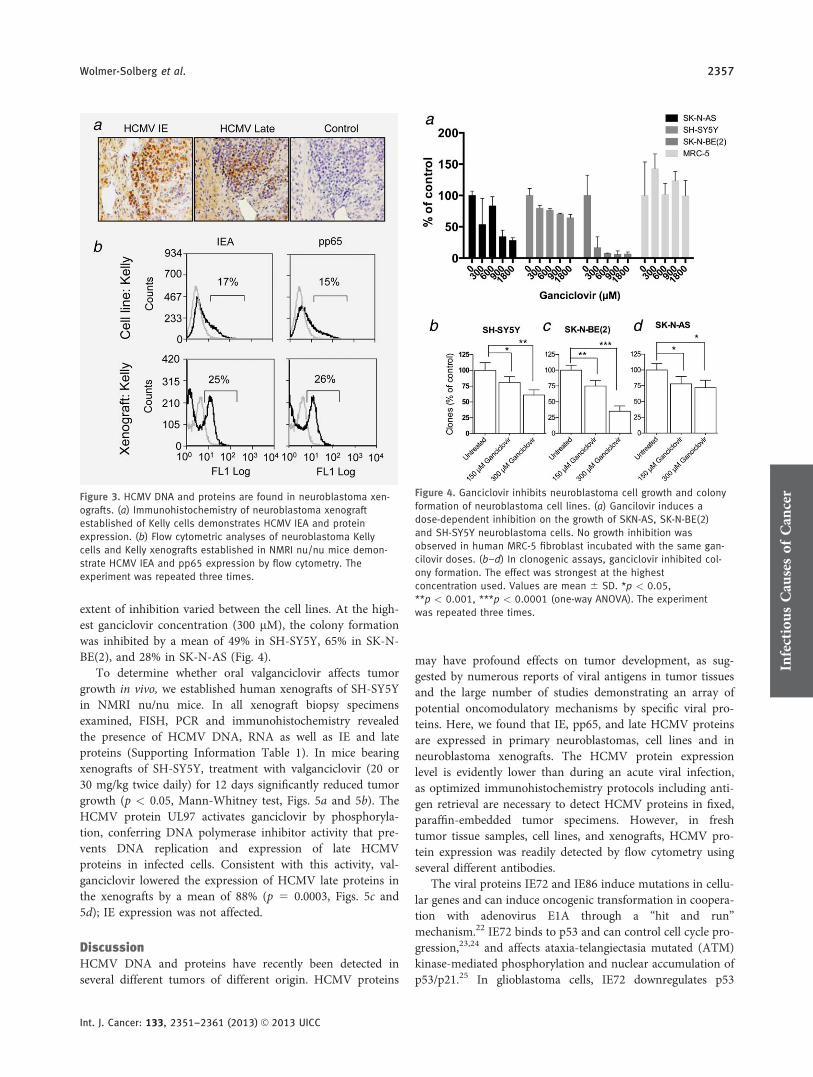
HCMV DNA, RNA and IE and late proteins were alsodetected by PCR and immunohistochemistry in xenografts ofSH-SY5Y, SK-N-BE(2), SK-N-AS, SK-N-DZ, Kelly, and IMR-32 xenografts (Supporting Information Table 1, Kelly demon-strated in Fig. 3a). When Kelly cells were xenografted subcu-taneously in NMRI nu/nu mice, HCMV IE and pp65 proteinexpression slightly increased as determined by flow cytometryin xenografts compared to the cells in culture (Fig. 3b).
Figure 1. HCMV proteins are detected in 100% of human neuroblastoma samples. (d, h) HCMV-IE staining in tissue section obtained from a
GBM patient. Neuroblastoma samples were analyzed by immunohistochemistry for IE (a, e) and L (b, f) HCMV proteins and a-actin as iso-
type control (c, g). Original magnifications: 203 (a–d) and 403 (e–h). (i) IE proteins were found in 100% of analyzed neuroblastoma tis-
sues (n 5 36/36), and late proteins were found in 92% (n 5 33). (j) Stained tissues were graded 0–4 depending on the percentage of
positive cells. (k) Flow cytometric analyses of a primary neuroblastoma tumor reveal HCMV IEA protein expression in a majority of the cells
(n 5 2, one representative example is shown using the 810R clone detecting both the HCMV IE55, IE72 and IE86 proteins). (I) Western blot
analyses of seven frozen neuroblastoma tumor samples (lane 1–7) and control (C) using the using an HCMV IE specific antibody (810R
clone) recognizing IE72, IE86 and IE55. Marker sizes are indicated on right. Bars a–d; 800 lm. e–h; 100 lm. Infectious
Cau
sesof
Can
cer
Wolmer-Solberg et al. 2355
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC

Ganciclovir inhibits neuroblastoma growth in vitro and in
vivo
Since HCMV DNA and proteins were detected in neuroblas-toma cell lines, we tested the effects of ganciclovir on cellgrowth and the clonogenic capacity of neuroblastoma cell
lines SH-SY5Y, SK-N-BE(2), and SK-N-AS. Both neuroblas-toma cell growth and colony formation was significantlyinhibited (Fig. 4), whereas no growth inhibitory effects wereobserved when human MRC-5 fibroblasts were incubatedwith the same doses of gancilovir (Fig. 4). However, the
Figure 2. HCMV DNA and proteins are found in neuroblastoma cell lines. (a) FISH analysis demonstrates HCMV DNA in SH-SY5Y cells and
SK-N-AS cells. The viral DNA was located in the periphery of the cell nucleus in neuroblastoma cells compared to a central location in
HCMV AD169 infected MRC-5 fibroblasts. K562 cells provide a negative control demonstrating no detection of HCMV DNA. (b) Flow cytomet-
ric analyses of the neuroblastoma cells SK-N-DZ, SH-SY5Y and Kelly revealed different levels of expression of HCMV IE and pp65 proteins
(n 5 2–7 sampling occasions for individual cell lines). (c) Neuroblastoma cells as exemplified by SK-N-DZ cells co-expressed HCMV IEA and
the stem cell markers CD133 or CD44. The experiment was repeated three times. Left panels indicate isotype IgG control. Middle panels
demonstrate CMV IE single staining and right panels double staining of HCMV IEA and CD133 (SK-N-DZ) or CD44 (IMR-32). (d) Double stain-
ing of a primary neuroblastoma tissue sample section with HCMV IE (green) and CD133 specific antibodies (red) revealed some double pos-
itive cells indicated with arrows, DAPI positive cells in blue.
Infectious
Cau
sesof
Can
cer
2356 Human cytomegalovirus in neuroblastoma
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC
extent of inhibition varied between the cell lines. At the high-est ganciclovir concentration (300 lM), the colony formationwas inhibited by a mean of 49% in SH-SY5Y, 65% in SK-N-BE(2), and 28% in SK-N-AS (Fig. 4).
To determine whether oral valganciclovir affects tumorgrowth in vivo, we established human xenografts of SH-SY5Yin NMRI nu/nu mice. In all xenograft biopsy specimensexamined, FISH, PCR and immunohistochemistry revealedthe presence of HCMV DNA, RNA as well as IE and lateproteins (Supporting Information Table 1). In mice bearingxenografts of SH-SY5Y, treatment with valganciclovir (20 or30 mg/kg twice daily) for 12 days significantly reduced tumorgrowth (p < 0.05, Mann-Whitney test, Figs. 5a and 5b). TheHCMV protein UL97 activates ganciclovir by phosphoryla-tion, conferring DNA polymerase inhibitor activity that pre-vents DNA replication and expression of late HCMVproteins in infected cells. Consistent with this activity, val-ganciclovir lowered the expression of HCMV late proteins inthe xenografts by a mean of 88% (p 5 0.0003, Figs. 5c and5d); IE expression was not affected.
DiscussionHCMV DNA and proteins have recently been detected inseveral different tumors of different origin. HCMV proteins
may have profound effects on tumor development, as sug-gested by numerous reports of viral antigens in tumor tissuesand the large number of studies demonstrating an array ofpotential oncomodulatory mechanisms by specific viral pro-teins. Here, we found that IE, pp65, and late HCMV proteinsare expressed in primary neuroblastomas, cell lines and inneuroblastoma xenografts. The HCMV protein expressionlevel is evidently lower than during an acute viral infection,as optimized immunohistochemistry protocols including anti-gen retrieval are necessary to detect HCMV proteins in fixed,paraffin-embedded tumor specimens. However, in freshtumor tissue samples, cell lines, and xenografts, HCMV pro-tein expression was readily detected by flow cytometry usingseveral different antibodies.
The viral proteins IE72 and IE86 induce mutations in cellu-lar genes and can induce oncogenic transformation in coopera-tion with adenovirus E1A through a “hit and run”mechanism.22 IE72 binds to p53 and can control cell cycle pro-gression,23,24 and affects ataxia-telangiectasia mutated (ATM)kinase-mediated phosphorylation and nuclear accumulation ofp53/p21.25 In glioblastoma cells, IE72 downregulates p53
Figure 3. HCMV DNA and proteins are found in neuroblastoma xen-
ografts. (a) Immunohistochemistry of neuroblastoma xenograft
established of Kelly cells demonstrates HCMV IEA and protein
expression. (b) Flow cytometric analyses of neuroblastoma Kelly
cells and Kelly xenografts established in NMRI nu/nu mice demon-
strate HCMV IEA and pp65 expression by flow cytometry. The
experiment was repeated three times.
Figure 4. Ganciclovir inhibits neuroblastoma cell growth and colony
formation of neuroblastoma cell lines. (a) Gancilovir induces a
dose-dependent inhibition on the growth of SKN-AS, SK-N-BE(2)
and SH-SY5Y neuroblastoma cells. No growth inhibition was
observed in human MRC-5 fibroblast incubated with the same gan-
cilovir doses. (b–d) In clonogenic assays, ganciclovir inhibited col-
ony formation. The effect was strongest at the highest
concentration used. Values are mean 6 SD. *p < 0.05,
**p < 0.001, ***p < 0.0001 (one-way ANOVA). The experiment
was repeated three times.
Infectious
Cau
sesof
Can
cer
Wolmer-Solberg et al. 2357
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC

mRNA and protein expression.26 The HCMV protein IE86 candirectly interact with p53 and inhibit p53-mediated apoptosis.UL44 inhibits the transcriptional activity of p53. In gliomacells, HCMV infection increases proliferation, which is associ-ated with IE72-mediated inhibition of p53 and Rb proteins andactivation of the PI3K/Akt signaling pathway.24 HCMV UL97phosphorylates the Rb protein and thereby prevents apopto-sis,27 and HCMV pp71 induces its degradation by the protea-some (as reviewed in Ref. 28). Other HCMV proteins inhibitdifferentiation, control epigenetic functions, and modulate theimmune system, enabling the virus to avoid recognition andelimination (as reviewed in Ref. 28). Thus, HCMV regulates aplethora of factors that are important for the initiation of aber-rant cell growth, cell transformation and immune surveillance.HCMV infection also results in chromosomal aberrations andchromosome breaks at 1q21 and 1q42,29–31 and virus-infectedcells exhibit impaired DNA repair mechanisms.32 Interestingly,a poor prognostic subset of high-risk neuroblastoma displays aspecific chromosome instability phenotype. Furthermore, chro-mosome 1 deletions are common in neuroblastoma and amarker of poor prognosis,33,34 and copy number variations(CNV) in chromosome 1q21 have recently been linked to thesetumors.35 We therefore scrutinized the extensive Swedishmaterial of neuroblastoma tumors examined in detail withSNP based aCGH as previously described36 including the setsof tumors in this study. However, we were unable to detect the1q21 CNV as described by Diskin et al.35 (data not shown).
Certain HCMV proteins may also be involved in oncogen-esis. The HCMV-encoded chemokine receptor US28, whichis constitutively expressed in virus-infected cells, inducestumor formation in a murine model,37 and transgenic VS28mice with targeted expression of US28 to the intestinal epi-thelium develop intestinal hyperplasia and tumors.37 US28expression in intestinal epithelial cells inhibited glycogensynthase-3b (GSK-3b) function, promoted accumulation ofb-catenin and increased expression of Wnt target genesinvolved in the control of cell proliferation,37 which providea direct molecular link between HCMV US28 and oncogene-sis. US28 also stimulates angiogenesis by inducing expressionof vascular endothelial growth factor (VEGF) via thecyclooxygenase-2 signaling pathway in vivo and in vitro.38,39
US28 induces phosphorylation of STAT3, resulting ininterleukin-6 production, a phenomenon observed in glio-blastomas and linked to patient survival.40 Furthermore,murine CMV infection of Trp53 heterzygote neonatal miceresulted in tumors at high frequency at 9 months of age.Most of the tumors were rhabdomyosarcoma, and were posi-tive for MCMV DNA, RNA and protein.13 Trp53 heterzygotemice develop various spontaneous tumors, but generally after9 months of age. As only 3% of infected mice developedtumors if they were infected at 4 weeks of age, it was specu-lated that MCMV accelerates tumor formation in Trp53heterzygote mice and that preferentially rhabdomyosarcomadeveloped, although some mice also developed lymphomas.This observation support the hypothesis that CMV may
promote tumorogenesis in genetically predisposed individualsand may provide explanations to why only few HCMV car-riers develop rare tumors.
We have further showed that the HCMV protein IE72induces high telomerase activity through a direct interactionwith SP1 binding sites in the hTERT promoter;41 interest-ingly, in primary glioblastomas, only cells positive for HCMVIE expressed hTERT. Induced telomerase activity is a com-mon theme for oncogenic viruses, as increased telomeraseactivity is a prerequisite for tumor cells to divide indefi-nitely.42 Our observations suggest that only HCMV protein–positive glioblastoma cells have an unrestricted ability todivide. In support of this statement, we discovered that thelevel of HCMV infection in malignant glioblastomas has ahigh prognostic value.14,15 Patients with a low-grade infectionin the tumor lived significantly longer than those with ahigh-grade infection.14,15 This finding implies that the virushas a pathogenetic role and is not merely an epiphenomenonalso in malignant glioblastomas.
Owing to the limited number of patients and their hetero-geneity, we were unable to fully evaluate the potential clinicalimpact of HCMV infection in neuroblastomas in this study.However, no clear difference was found between HCMVgrade and different clinical and biological subsets of neuro-blastomas included. Nevertheless, we found a high prevalenceof HCMV in childhood neuroblastomas; all examined pri-mary tumors exhibited HCMV RNA and protein expression.Viral cultures from tumor tissues were negative, and healthytissues around the tumors appeared negative for HCMV pro-teins. However, we also detected HCMV proteins in differen-tiated ganglioneuromas and healthy adrenal tissue samplesobtained from neuroblastoma patients, which imply that thevirus is not only detected in malignant tumors, nor is itrestricted to the tumor in neuroblastoma patients. The find-ing that HCMV proteins are detected in all subsets of neuro-blastoma indicates that additional chromosomal aberrationsand/or epigenetic alterations are important for the biologicalbehavior and clinical outcome. On the other hand, ourresults in vitro and in vivo clearly show that HCMV may bea novel adjuvant therapy for neuroblastomas.
Our observations further suggest that HCMV is present inlong-term cultures; although the HCMV protein expressionlevels varied between cell lines sampled at different times.We suspect that the virus is present in a population of stemcells within the tumor. In support of this possibility, wefound that HCMV proteins were expressed by neuroblastomacells carrying CD133 or CD44, two potential markers defin-ing stem cells within the tumor. In SH-SY5Y xenografts,HCMV protein expression was low as shown by immunohis-tochemistry (Fig. 4c). SH-SY5Y cells had very few cells dou-ble positive for CD44 and CMV IE proteins and the IEexpression varied in culture; 34, 8, 0.1, 0, 0.1, 54, 61% atseven different sampling occasions during 1 year. Weobserved a few weakly HCMV IE positive cells that alsoexpressed CD133 in frozen neuroblastoma samples (Fig. 2d).
Infectious
Cau
sesof
Can
cer
2358 Human cytomegalovirus in neuroblastoma
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC

A link between clinical HCMV infection and neuroblas-toma has been proposed earlier3,19 but was not demonstratedin tumor tissue specimens. HCMV is the most commonpathogen to cause birth defects through congenital infections,which occur in 0.5 to 2% of all live births in the developedworld.43,44 At 1 year of age, 25 to 40% of children are sero-positive for CMV.43 Neuroblastomas consist of poorly orundifferentiated neural cells that are believed to originatefrom a developmental defect and are therefore considered tobe embryonic tumors. We and others have reported thatHCMV infection of neuronal progenitor cells prevents theirdifferentiation.45–47 An early block in differentiation may bean important step in tumor transformation.48 As HCMV caninfect progenitor cells in the developing fetus, it may be anearly step toward tumor transformation in genetically predis-posed individuals.
Metastatic high-risk neuroblastomas, in particular the chro-mosome unstable subset, are often resistant to treatment evenwith potent multimodal therapy. Our results suggest that thisvirus may play a role in neuroblastoma pathogenicity and there-fore represent a therapeutical target in this as well as otherHCMV-positive tumors. We here demonstrate that anti-viraltherapy prevents the growth of neuroblastomas in vitro and invivo. We also recently found that HCMV is highly prevalent inmedulloblastomas and that anti-viral treatment inhibits growthof HCMV-positive medulloblastoma xenografts in immunode-ficient mice.7 In the previous study, we established xenograftsfrom two HCMV-negative cell lines PC-3 established from abone metastasis of a grade IV prostatic adenocarcinoma andBxPC-3 established from a pancreatic adenocarcinoma. In clon-ogenic assays, no significant effects in the clonogenic capacitywere observed when PC-3 or BxPC-3 cells were treated with
Figure 5. Valganciclovir inhibits neuroblastoma growth in mice. (a) Female NMRI nu/nu mice carrying subcutaneous neuroblastoma xeno-
grafts were randomized to receive valganciclovir (20, n 5 7, or 30 mg/kg, n 5 7) twice daily or no treatment (n 5 10). Tumors were meas-
ured every day, and tumor volume was calculated as (width)2 3 length 3 0.44. The tumor volume index was calculated as the measured
volume divided by the volume at the start of treatment. Tumor volume index at day 12 show significant growth reduction in animals treated
with valganciclovir. Tumor volume index values are presented as mean. *p < 0.05 (Mann-Whitney test). (b) Tumor weight recorded at
autopsy correlated with results on tumor volume index. Tumor weight is shown as median. *p < 0.05 (Mann-Whitney test). (c) The number
of cells positive for HCMV IE and late proteins were rather few in the xenografts, yet consistent results for IE and late expression were
obtained. While valganciclovir did not affect the expression of IE proteins, it significantly reduced the expression of late protein in the xeno-
grafts. Values are mean 6 SD (t test). The animal experiment was repeated two times. The bar represents 50 lm.
Infectious
Cau
sesof
Can
cer
Wolmer-Solberg et al. 2359
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC

37.5-, 75- or 150-lM ganciclovir.7 Treatment of NMRI nu/numice carrying established PC-3 or BxPC-3 xenografts with val-ganciclovir (14 mg/kg twice daily po) had no significant effecton tumor growth.7 Although we were unable to identify anHCMV-negative neuroblastoma cell line, our previous observa-tions suggest that the effects of valganciclovir in neuroblastomaare HCMV specific. Furthermore, we have performed theworld’s first clinical study evaluating anti-viral therapy in 42glioblastoma patients randomized to valganciclovir or placebo.Although the study failed its primary endpoint demonstratingno significant difference in tumor volumes at 3 and 6 months,patients who received at least 6 months of valganciclovir treat-ment demonstrate an unexpectedly high overall survival; 50% at2 years at 27% at 4 years.49
HCMV infection of neuroblastoma cells in vitro confersresistance to chemotherapeutic agents such as cisplatinumand etoposide;50 this effect was blocked by ganciclovir, whichtargets HCMV replication. Thus, targeting the virus in tumorcells may decrease potential viral effects on cellular functionsand also increase sensitivity to chemotherapy. Since ganciclo-vir significantly reduced tumor growth both in vitro and invivo, this study indicate that antiviral treatment for HCMVmay be a novel, generally well-tolerated option for treatingchildren with neuroblastoma.
AcknowledgementsWe thank Stephen Ordway for editorial help.
References
1. Brodeur GM. Neuroblastoma: biological insightsinto a clinical enigma. Nat Rev Cancer 2003;3:203–16.
2. Mosse YP, Laudenslager M, Longo L, et al.Identification of ALK as a major familialneuroblastoma predisposition gene. Nature 2008;455:930–5.
3. Nigro G, Schiavetti A, Booth JC, et al.Cytomegalovirus-associated stage 4Sneuroblastoma relapsed stage 4. Med PediatrOncol 1995;24: 200–3.
4. Wertheim P, Voute PA. Neuroblastoma, Wilms’tumor, and cytomegalovirus. J Natl Cancer Inst1976;57:701–3.
5. Scholz M, Blaheta RA, Wittig B, et al.Cytomegalovirus-infected neuroblastoma cellsexhibit augmented invasiveness mediated bybeta1alpha5 integrin (VLA-5). Tissue Antigens2000;55:412–21.
6. Cinatl J, Jr, Vogel JU, Cinatl J, et al. Long-termproductive human cytomegalovirus infection of ahuman neuroblastoma cell line. Int J Cancer1996;65:90–6.
7. Baryawno N, Rahbar A, Wolmer-Solberg N, et al.Detection of human cytomegalovirus inmedulloblastomas reveals a potential therapeutictarget. J Clin Invest 2011;121:4043–55.
8. Cobbs CS, Harkins L, Samanta M, et al. Humancytomegalovirus infection and expression inhuman malignant glioma. Cancer Res 2002;62:3347–50.
9. Harkins L, Volk AL, Samanta M, et al. Specificlocalisation of human cytomegalovirus nucleicacids and proteins in human colorectal cancer.Lancet 2002;360:1557–63.
10. Harkins LE, Matlaf LA, Soroceanu L, et al.Detection of human cytomegalovirus in normaland neoplastic breast epithelium. Herpesviridae2010;1:8.
11. Mitchell DA, Xie W, Schmittling R, et al.Sensitive detection of human cytomegalovirus intumors and peripheral blood of patientsdiagnosed with glioblastoma. Neuro-oncology2008;10:10–18.
12. Samanta M, Harkins L, Klemm K, et al. Highprevalence of human cytomegalovirus in prostaticintraepithelial neoplasia and prostatic carcinoma.J Urol 2003;170:998–1002.
13. Price RL, Bingmer K, Harkins L, et al.Cytomegalovirus infection leads to pleomorphic
rhabdomyosarcomas in Trp531/- mice. CancerRes 2012;72:5669–74.
14. Rahbar A, Stragliotto G, Orrego A, et al. Lowlevels of human cytomegalovirus infection inglioblastoma multiforme associates with patientsurvival—a case-control study. Herpesviridae2012;3:3.
15. Rahbar A, Orrego A, Peredo I, et al. Humancytomegalovirus infection levels in glioblastomamultiforme are of prognostic value for survival. JClin Virol 2013;57:36–42.
16. Soroceanu L, Cobbs CS. Is HCMV a tumorpromoter? Virus Res 2011;157:193–203.
17. Soderberg-Naucler C. Does cytomegalovirus playa causative role in the development of variousinflammatory diseases and cancer? J Intern Med2006;259:219–46.
18. Baryawno N, Sveinbjornsson B, Eksborg S, et al.Tumor-growth-promoting cyclooxygenase-2 pros-taglandin E2 pathway provides medulloblastomatherapeutic targets. Neuro Oncol 2008;10:661–74.
19. Nigro G, Adler SP. Cytomegalovirus infectionsduring pregnancy. Curr Opin Obstet Gynecol2011;23:123–8.
20. Brodeur GM, Pritchard J, Berthold F et al.Revisions of the international criteria forneuroblastoma diagnosis, staging and response totreatment. J Clin Oncol 1993;11:1466–77.
21. Cohn SL, Pearson AD, London WB, et al. TheInternational Neuroblastoma Risk Group (INRG)classification system: an INRG Task Force report.J Clin Oncol 2009;27:289–97.
22. Shen Y, Zhu H, Shenk T. Humancytomagalovirus IE1 and IE2 proteins aremutagenic and mediate “hit-and-run” oncogenictransformation in cooperation with the adenovi-rus E1A proteins. Proc Natl Acad Sci USA 1997;94:3341–5.
23. Speir E, Modali R, Huang ES, et al. Potential roleof human cytomegalovirus and p53 interaction incoronary restenosis. Science 1994;265:391–4.
24. Cobbs CS, Soroceanu L, Denham S, et al.Modulation of oncogenic phenotype in humanglioma cells by cytomegalovirus IE1-mediatedmitogenicity. Cancer Res 2008;68:724–30.
25. Castillo JP, Frame FM, Rogoff HA, et al. Humancytomegalovirus IE1-72 activates ataxia telangiec-tasia mutated kinase and a p53/p21-mediatedgrowth arrest response. J Virol 2005;79:11467–75.
26. Lee K, Jeon K, Kim JM, et al. Downregulation ofGFAP, TSP-1, and p53 in human glioblastomacell line, U373MG, by IE1 protein from humancytomegalovirus. Glia 2005;51:1–12.
27. Hume AJ, Finkel JS, Kamil JP, et al.Phosphorylation of retinoblastoma protein byviral protein with cyclin-dependent kinase func-tion. Science 2008;320:797–9.
28. Soderberg-Naucler C. HCMV microinfections ininflammatory diseases and cancer. J Clin Virol2008;41:218–23.
29. Fortunato EA, Dell’Aquila ML, Spector DH.Specific chromosome 1 breaks induced by humancytomegalovirus. Proc Natl Acad Sci USA 2000;97:853–8.
30. Siew VK, Duh CY, Wang SK. Humancytomegalovirus UL76 induces chromosomeaberrations. J Biomed Sci 2009;16:107.
31. Albrecht T, Deng CZ, Abdel-Rahman SZ, et al.Differential mutagen sensitivity of peripheralblood lymphocytes from smokers andnonsmokers: effect of human cytomegalovirusinfection. Environ Mol Mutagen 2004;43:169–78.
32. Luo MH, Rosenke K, Czornak K, et al. Humancytomegalovirus disrupts both ataxiatelangiectasia mutated protein (ATM)- andATM-Rad3-related kinase-mediated DNA damageresponses during lytic infection. J Virol 2007;81:1934–50.
33. Martinsson T, Sjoberg RM, Hedborg F, et al.Deletion of chromosome 1p loci andmicrosatellite instability in neuroblastomasanalyzed with short-tandem repeat polymor-phisms. Cancer Res 1995;55:5681–6.
34. Caron H, van Sluis P, de Kraker J, et al.Allelic loss of chromosome 1p as a predictorof unfavorable outcome in patients withneuroblastoma. New Engl J Med 1996;334:225–30.
35. Diskin SJ, Hou C, Glessner JT, et al. Copynumber variation at 1q21.1 associated withneuroblastoma. Nature 2009;459:987–91.
36. Caren H, Kryh H, Nethander M, et al. High-riskneuroblastoma tumors with 11q-deletion displaya poor prognostic, chromosome instability pheno-type with later onset. Proc Natl Acad Sci USA2010;107:4323–8.
37. Bongers G, Maussang D, Muniz LR, et al. Thecytomegalovirus-encoded chemokine receptorUS28 promotes intestinal neoplasia in transgenicmice. J Clin Invest 2010;120:3969–78.
Infectious
Cau
sesof
Can
cer
2360 Human cytomegalovirus in neuroblastoma
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC

38. Maussang D, Langemeijer E, Fitzsimons CP, et al.The human cytomegalovirus-encoded chemokinereceptor US28 promotes angiogenesis and tumorformation via cyclooxygenase-2. Cancer Res 2009;69:2861–9.
39. Maussang D, Verzijl D, van Walsum M, et al.Human cytomegalovirus-encoded chemokinereceptor US28 promotes tumorigenesis. Proc NatlAcad Sci USA 2006;103:13068–73.
40. Slinger E, Maussang D, Schreiber A, et al.HCMV-encoded chemokine receptor US28mediates proliferative signaling through the IL-6-STAT3 axis. Sci Signal 2010;3:ra58.
41. Straat K, Liu C, Rahbar A, et al. Activation oftelomerase by human cytomegalovirus. J NatlCancer Inst 2009;101:488–97.
42. Opitz OG. Telomeres, telomerase and malignanttransformation. Curr Mol Med 2005;5:219–26.
43. Ahlfors K. IgG antibodies to cytomegalovirus in anormal urban Swedish population. Scand J InfectDis 1984;16:335–7.
44. Forbes BA. Acquisition of cytomegalovirusinfection: an update. Clin Microbiol reviews 1989;2:204–16.
45. Odeberg J, Wolmer N, Falci S, et al. Humancytomegalovirus inhibits neuronal differentiationand induces apoptosis in human neural precursorcells. J Virol 2006;80:8929–39.
46. Odeberg J, Wolmer N, Falci S, et al. Late humancytomegalovirus (HCMV) proteins inhibitdifferentiation of human neural precursor cells intoastrocytes. J Neurosci Res 2007;85:583–93.
47. Han GP, Li L, Kosugi I, et al. Enhancement ofsusceptibility of adult mouse brain tocytomegalovirus infection by infusion of epidermalgrowth factor. J Neurosci Res 2007;85:2981–90.
48. Sanai N, Alvarez-Buylla A, Berger MS. Neuralstem cells and the origin of gliomas. N Engl JMed 2005;353:811–22.
49. Stragliotto G, Rahbar A, Solberg NW, et al. Effects ofvalganciclovir as an add-on therapy in patients withcytomegalovirus-positive glioblastoma: a random-ized, double-blind, hypothesis-generating study. Int JCancer 2013, doi: 10.1002/ijc.28111.
50. Cinatl J, Jr, Cinatl J, Vogel JU, et al. Persistent humancytomegalovirus infection induces drug resistanceand alteration of programmed cell death in humanneuroblastoma cells. Cancer Res 1998;58:367–72.
Infectious
Cau
sesof
Can
cer
Wolmer-Solberg et al. 2361
Int. J. Cancer: 133, 2351–2361 (2013) VC 2013 UICC




















