
Motor imagery practice may compensate for the slowdown of sensorimotor processes induced by...
11
ORIGINAL ARTICLE Motor imagery practice may compensate for the slowdown of sensorimotor processes induced by short-term upper-limb immobilization Aurore Meugnot • Nounagnon Frutueux Agbangla • Yves Almecija • Lucette Toussaint Received: 10 April 2014 / Accepted: 24 May 2014 Ó Springer-Verlag Berlin Heidelberg 2014 Abstract Recently, it has been demonstrated that senso- rimotor representations are quickly updated following a brief period of limb non-use. The present study examined the potential of motor imagery practice (MIP) and inves- tigated the role of motor imagery instructions (kinesthetic vs. visual imagery) to counteract the functional impairment induced by sensorimotor restriction. The participants were divided into four groups. Three groups wore a splint on their left hand for 24 h. Prior to the splint removal, two of the three groups performed 15 min of MIP, with kines- thetic or visual modalities (KinMIP and VisMIP groups, respectively). The third group did not practice motor imagery (NoMIP group). Immediately after the splint removal, the participants were assessed using a hand lat- erality task known for evaluating sensorimotor processes. A fourth group served as the control (i.e., without immo- bilization and MIP). The main results showed slower left- hand response times for the immobilized NoMIP group compared with the controls. Importantly, faster response times for the left-hand stimuli appeared for the KinMIP groups only compared with the NoMIP group. No differ- ence between the four groups was observed for the right- hand stimuli. Overall, these results highlighted the soma- totopic effect of limb non-use on the efficiency of senso- rimotor processes. Importantly, the slowdown of the sensorimotor processes induced by 24 h of sensorimotor deprivation may be counteracted by a kinesthetic MIP, whereas no beneficial effect appeared with visual imagery. We discuss the importance of imagery modalities for sen- sorimotor reactivation. Introduction Motor imagery (MI) is ‘‘a dynamic state during which the representation of a given motor act is internally rehearsed within working memory without any overt motor output’’ (Decety & Gre `zes, 1999). Psychophysical and brain imaging studies have demonstrated that movement simu- lation and movement execution involve similar sensori- motor representations. Behavioral experiments have shown that motor imagery preserved the spatiotemporal charac- teristics of the actual actions (Cerritelli, Maruff, Wilson, & Currie, 2000; Decety & Jeannerod, 1995) and obeyed the same joint limits (Petit, Pegna, Mayer, & Hauert, 2003) or inertial and gravitational constraints (Papaxanthis, Pozzo, Skoura, & Schiappati, 2002). Neuroimaging studies have revealed an overlap between the brain regions activated during MI and the actual movement (Gerardin et al., 2000; Guillot et al., 2009; Hanakawa, Dimyan, & Hallett, 2008; Solodkin, Hlustik, Chen, & Small, 2004). Another similarity emerged from studies which recorded autonomic nervous system activity, with similar increases in ventila- tion and systolic blood pressure (Wang & Morgan, 1992) or in electrodermal and thermovascular responses (Guillot & Collet, 2005) during simulation and the actual execution of movements. Overall, these findings allowed to us to consider MI as the cognitive level of actions (Jeannerod, 2001), and MI tasks appeared as a relevant means to highlight the central functioning of the sensorimotor system. MI tasks are often used in clinical studies, with patients suffering from somatic motor disorders, to provide information on the link A. Meugnot Á N. F. Agbangla Á Y. Almecija Á L. Toussaint (&) Universite ´ de Poitiers, Centre de Recherches sur la Cognition et l’Apprentissage, CeRCA, CNRS/UMR 7295, 5 rue The ´odore Lefebvre, 86000, Poitiers, France e-mail: [email protected] URL: http://cerca.labo.univ-poitiers.fr 123 Psychological Research DOI 10.1007/s00426-014-0577-1
-
Upload
univ-poitiers -
Category
Documents
-
view
2 -
download
0
Transcript of Motor imagery practice may compensate for the slowdown of sensorimotor processes induced by...

ORIGINAL ARTICLE
Motor imagery practice may compensate for the slowdownof sensorimotor processes induced by short-term upper-limbimmobilization
Aurore Meugnot • Nounagnon Frutueux Agbangla •
Yves Almecija • Lucette Toussaint
Received: 10 April 2014 / Accepted: 24 May 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract Recently, it has been demonstrated that senso-
rimotor representations are quickly updated following a
brief period of limb non-use. The present study examined
the potential of motor imagery practice (MIP) and inves-
tigated the role of motor imagery instructions (kinesthetic
vs. visual imagery) to counteract the functional impairment
induced by sensorimotor restriction. The participants were
divided into four groups. Three groups wore a splint on
their left hand for 24 h. Prior to the splint removal, two of
the three groups performed 15 min of MIP, with kines-
thetic or visual modalities (KinMIP and VisMIP groups,
respectively). The third group did not practice motor
imagery (NoMIP group). Immediately after the splint
removal, the participants were assessed using a hand lat-
erality task known for evaluating sensorimotor processes.
A fourth group served as the control (i.e., without immo-
bilization and MIP). The main results showed slower left-
hand response times for the immobilized NoMIP group
compared with the controls. Importantly, faster response
times for the left-hand stimuli appeared for the KinMIP
groups only compared with the NoMIP group. No differ-
ence between the four groups was observed for the right-
hand stimuli. Overall, these results highlighted the soma-
totopic effect of limb non-use on the efficiency of senso-
rimotor processes. Importantly, the slowdown of the
sensorimotor processes induced by 24 h of sensorimotor
deprivation may be counteracted by a kinesthetic MIP,
whereas no beneficial effect appeared with visual imagery.
We discuss the importance of imagery modalities for sen-
sorimotor reactivation.
Introduction
Motor imagery (MI) is ‘‘a dynamic state during which the
representation of a given motor act is internally rehearsed
within working memory without any overt motor output’’
(Decety & Grezes, 1999). Psychophysical and brain
imaging studies have demonstrated that movement simu-
lation and movement execution involve similar sensori-
motor representations. Behavioral experiments have shown
that motor imagery preserved the spatiotemporal charac-
teristics of the actual actions (Cerritelli, Maruff, Wilson, &
Currie, 2000; Decety & Jeannerod, 1995) and obeyed the
same joint limits (Petit, Pegna, Mayer, & Hauert, 2003) or
inertial and gravitational constraints (Papaxanthis, Pozzo,
Skoura, & Schiappati, 2002). Neuroimaging studies have
revealed an overlap between the brain regions activated
during MI and the actual movement (Gerardin et al., 2000;
Guillot et al., 2009; Hanakawa, Dimyan, & Hallett,
2008; Solodkin, Hlustik, Chen, & Small, 2004). Another
similarity emerged from studies which recorded autonomic
nervous system activity, with similar increases in ventila-
tion and systolic blood pressure (Wang & Morgan, 1992)
or in electrodermal and thermovascular responses (Guillot
& Collet, 2005) during simulation and the actual execution
of movements.
Overall, these findings allowed to us to consider MI as
the cognitive level of actions (Jeannerod, 2001), and MI
tasks appeared as a relevant means to highlight the central
functioning of the sensorimotor system. MI tasks are often
used in clinical studies, with patients suffering from
somatic motor disorders, to provide information on the link
A. Meugnot � N. F. Agbangla � Y. Almecija � L. Toussaint (&)
Universite de Poitiers, Centre de Recherches sur la Cognition et
l’Apprentissage, CeRCA, CNRS/UMR 7295, 5 rue Theodore
Lefebvre, 86000, Poitiers, France
e-mail: [email protected]
URL: http://cerca.labo.univ-poitiers.fr
123
Psychological Research
DOI 10.1007/s00426-014-0577-1

between a cortical structure and its functional outcome. In
studies with apraxic patients, Sirigu and collaborators
highlighted the contribution of the parietal cortex in the
generation of action representations (Sirigu, Daprati, Pra-
dat-Diehl, Franck, & Jeannerod, 1999; Sirigu et al., 1996).
Other experiments examined the function of the basal
ganglia in both inner and behavioral states of action by
comparing the performance of parkinsonian patients and
healthy participants in actual and imagined sequential fin-
ger movements (Dominey, Decety, Brousolle, Chazot, &
Jeannerod, 1994). In other cases, MI studies were used to
better understand complex pathologies, such as schizo-
phrenia (de Vignemont et al., 2006), focal hand dystonia
(Fiorio, Tinazzi, & Aglioti, 2006), anosognosia for hemi-
plegia (Jenkinson, Edelstyn, & Ellis, 2009), developmental
coordination disorder in children (Wilson et al., 2004) and
Asperger syndrome (Conson et al., 2013).
Recently, MI tasks were also used in studies of func-
tional brain plasticity that explored the effects of peripheral
perturbations on sensorimotor representations, such as limb
amputation (Curtze, Otten, & Postema, 2010; Malouin
et al., 2009; Nico, Daprati, Rigal, Parsons, & Sirigu, 2004),
chronic deafferentation (ter Horst, Cole, van Lier, &
Steenbergen, 2012), congenitally absent limb (Funk &
Brugger, 2008) and visual loss and disuse (Malouin et al.,
2009). Overall, these studies have shown that kinesthetic
afferent signals are crucial to maintain efficient sensori-
motor representation and implicitly use internal motor
representation. Importantly, ter Horst et al. (2012) revealed
that long-term loss of kinesthetic afferents may be com-
pensated by visual information. In order to compensate for
the chronic loss of proprioceptive afferents, the deaffe-
rented patient used visual information to construct a rep-
resentation of his body, highlighting the multimodal nature
of body representation. More interested by transient
peripheral perturbations, some authors examined the effect
of regional anesthesia (Silva et al., 2011) or short-term
limb non-use (Meugnot, Almecija, & Toussaint, 2014;
Toussaint & Meugnot, 2013) on sensorimotor representa-
tion. Toussaint and collaborators (Meugnot et al., 2014;
Toussaint & Meugnot, 2013) specifically explored the
effect of short-term hand immobilization (24 or 48 h delay)
on motor simulation processes in healthy participants. The
authors clearly demonstrated that motor imagery processes
(i.e., mental rotation of body stimuli) were affected by left-
or right-hand immobilization, whereas visual imagery
processes were not affected (i.e., mental rotation of non-
body stimuli). For the mental rotation of body stimuli, the
immobilization-induced effects have been demonstrated by
the slower response times for the stimuli that depicted the
immobilized hand. Toussaint and collaborators suggested
that the limb non-use slowed down the corresponding
sensorimotor processes. This quick update of sensorimotor
representations induced by the decrease of input/output
signals could be the origin of motor performance distur-
bances reported in other experiments on movement accu-
racy (Huber et al., 2006; Moisello et al., 2008) and
movement kinematics (Bassolino, Bove, Jacono, Fadiga, &
Pozzo, 2012).
Motor imagery practice (MIP), ‘‘the act of repeating the
imagined movements several times with the intention of
learning a new ability or perfecting a known skill’’ (Jack-
son et al., 2001), has been investigated in both motor
learning and rehabilitation domains. The benefits of MIP in
improving or recovering motor skills have been widely
highlighted in sports (Driskell, Copper, & Moran, 1994;
Feltz & Landers, 1983; Guillot & Collet, 2008), motor
learning (Gentili, Han, Schweighofer, & Papaxanthis,
2010; Jackson, Lafleur, Malouin, Richards, & Doyon,
2003; Toussaint & Blandin, 2010) and rehabilitation pro-
grams (Braun et al., 2013; Dunsky, Dickstein, Marcovitz,
Levi, & Deutsch, 2008; Malouin, Jackson, & Richards,
2013; Malouin, Richards, Durand, & Doyon, 2008). Exe-
cution facilitation following MIP was attributed to the
rehearsal of the corresponding brain structures and the
solicitation of the same sensorimotor representations that
are used in physical practice (Decety & Grezes, 1999;
Jeannerod, 2001). This suggestion is particularly supported
by neurophysiological data that showed MIP produced a
similar reorganization of the motor cortex compared with
physical practice (Jackson et al., 2003; Lacourse, Turner,
Randolph-Orr, Schandler, & Cohen, 2004; Pascual-Leone
et al., 1995). Therefore, MIP appears to be an effective tool
for the maintenance of sensorimotor schemes, especially
when physical execution is not possible.
The benefit of MIP following short-term reduction of
proprioceptive information is actually not known. In this
study, we specifically investigated whether MIP could be
useful to counteract the slowdown of the sensorimotor
processes induced by the decrease in the input/output sig-
nal processing during 24 h of left-arm immobilization.
Moreover, we investigated the importance of the imagery
instructions on the sensorimotor reactivation by focusing
on the visual vs. kinesthetic motor imagery. Motor imagery
based on the visual modality (i.e., visual motor imagery)
requires the self-visualization of the movement, and indi-
viduals are instructed to focus on the spatial components of
the action (i.e., visualization of space, size, amplitude or
form of the movements). Motor imagery based on the
kinesthetic modality (i.e., kinesthetic motor imagery)
requires an individual to ‘‘feel’’ the movement, to mentally
perceive the muscle contractions, including the force and
the effort perceived during the movement and to focus on
the temporal component of the movements (rhythm, speed,
duration). Neural data have demonstrated that kinesthetic
motor imagery had a greater association with the
Psychological Research
123

sensorimotor system compared with visual motor imagery,
which was predominately activated by the visual associa-
tion cortex (Guillot et al., 2009; Ruby & Decety, 2003;
Solodkin et al., 2004; Stinear, Byblow, Steyvers, Levin, &
Swinnen, 2006). Therefore, the authors recommended the
use of a kinesthetic MIP rather than a visual MIP for
reactivating motor function within rehabilitation programs
(Jackson et al., 2001; Mulder, 2007; Stinear et al., 2006).
However, until now, no study has investigated whether the
motor imagery instructions have specific effects in the
functional rehabilitation domain (Braun et al., 2013).
Moreover, behavioral studies in sport or laboratory situa-
tions currently remain controversial. Many studies have
emphasized the benefits associated with kinesthetic MIP in
skill learning, but differences existed depending on the task
constraints (Fery, 2003; Guillot, Collet, & Dittmar, 2004;
Hardy & Callow, 1999; White and Hardy, 1995). Recently,
Toussaint and Blandin (2010) demonstrated the impact of a
previously stored sensory-specific movement representa-
tion (visual vs. kinesthetic) on the efficiency of MIP, which
favored either visual or kinesthetic motor imagery
instructions. The authors reported the dominance of visual
information over kinesthetic information when participants
executed hand movements with visual feedback. In that
specific case, a subsequent visual MIP promoted motor
learning, whereas no benefit of a kinesthetic MIP was
observed. In contrast, when participants experienced the
task without vision (the eyes closed), their performance
increased after a subsequent kinesthetic MIP, whereas no
benefit of a visual MIP appeared. Consequently, in the
present experiment, the sensory-specific condition of the
task experienced before MIP was accurately controlled to
test the potential of kinesthetic and/or visual MIP to
counteract the functional impairment induced by 1 day of
sensorimotor restriction.
In the present experiment, to examine whether MIP may
reactivate the sensorimotor processes that have been slo-
wed down by immobilization and whether kinesthetic or
visual imagery may be favored, four groups of participants
were used: a control group and three left-hand immobili-
zation groups. Two of the three immobilized groups were
subjected to a MIP with either visual (VisMIP group) or
kinesthetic imagery (KinMIP group) during the immobili-
zation (immediately prior to the splint removal), whereas
the third group did not practice motor imagery (NoMIP
group). All participants were required to judge whether a
hand figure displayed at the center of the screen was a left
or a right hand. As emphasized in the literature, we
expected slower response times for the left (immobilized)
hand identification in the immobilized NoMIP group
compared with the control group. The VisMIP and KinMIP
groups should allow us to examine the effect of motor
imagery instructions on sensorimotor reactivation. In
agreement with neuroimaging studies, if the use of bodily
information is crucial to reactivate the sensorimotor sys-
tem, a kinesthetic imagery practice (KinMIP group) should
further compensate for the slowdown of the sensorimotor
processes compared with a visual imagery practice
(VisMIP group), especially for stimuli that depict the
immobilized hand. In contrast, regarding the multimodal
nature of action representation, we may argue that senso-
rimotor memories might recover vividness by working on
their visual component. In this case, the VisMIP group
should improve its ability to identify the hand immobilized
stimuli similar to the KinMIP group.
Another aim of the present experiment was to examine
whether the effect of sensorimotor deprivation varied
according to the accuracy requirement of the simulated
movements. A previous study conducted by Gentilucci,
Benuzzi, Bertolani, Daprati, and Gangitano (2000) showed
that the type of grip influenced the recognition of hand
laterality, with the response times increasing with the
increase in precision grip (small sphere vs. large sphere).
Moreover, they observed similar grip-precision effects for
both the simulated and the actual grasping movements. The
authors suggested that to recognize hand laterality, the
participants used a motor hand representation that retrieved
the main aspect of the motor act. Therefore, in the present
experiment, we presented the participants with pictures of
the hand grasping a small or a large sphere to determine
whether the effect of sensorimotor deprivation varied
according to the precision of the simulated movements.
Methods
Participants
Fifty-two university students (18–26 years, mean
age = 19.7 years, 27 males) completed the experiment. All
participants were self-reported right-handers and non
ambidextrous. The participants were divided into four
groups of 13 participants: a control group (mean
age = 20 years, 9 males) and three left-hand immobiliza-
tion groups. Two of these 3 groups performed a MIP with
either visual (VisMIP, mean age = 20 years, 6 males) or
kinesthetic imagery instructions (KinMIP, mean
age = 19.5 years, 6 males) during the immobilization,
whereas the third group did not practice motor imagery
(NoMIP, mean age = 19 years, 6 males). The participants
were healthy, had normal or corrected-to-normal vision and
no history of motor or neurological disorders. The local
ethics committee approved the study protocol. All partici-
pants provided their written informed consent prior to their
inclusion in this study. Before testing, they were naıve to
the aims of the experiment.
Psychological Research
123

Materials and tasks
The participants were seated *60 cm in front of a com-
puter screen, with their hands resting palm down on their
thighs. All participants were asked to identify, as accu-
rately and as quickly as possible, whether a hand figure
displayed at the center of the computer screen corre-
sponded to a right or a left hand (Fig. 1). Each trial began
when a fixation cross was displayed on the center of the
screen for 500 ms. Then, a hand figure was presented and
remained visible until the participant provided his/her
response. The participants provided a verbal response
(‘‘left’’/‘‘right’’). The E-Prime 2.0 software package (Psy-
chology Software Tools Inc., Pittsburgh, USA) was used to
present the stimuli and record the participants’ response
times via a microphone connected to the computer. For
each trial, the experimenter wrote the participants’
responses on a data sheet.
The stimuli consisted of pictures of right or left hands
and forearms (see Gentilucci et al., 2000 for similar
material). They were presented on a rectangular
(12 9 9.44 cm) dark background. They were horizontally
located and could be presented in two directions. The fin-
gers were located either on the right (right-oriented hand)
or on the left (left-oriented hand). The hands held a sphere
that was one of two sizes (‘‘small’’ or ‘‘large’’). The
diameters of the small and large spheres were 2 and 14 cm,
respectively (see Fig. 1). Eight stimuli (2 hands 9 2
grips 9 2 directions) were used in the present experiment.
Procedure
We used a rigid splint (model DONJOY ‘‘Comfort Digit’’,
DJO, Surrey, UK) to immobilize the left wrist and three
fingers (index, middle and ring fingers) of the participants
for the three immobilization groups for 24 h. Their
immobilized arm was placed in a sling to ensure that the
participants kept their hand at rest as much as possible
during the day of immobilization. The participants were
instructed to never remove the splint and to keep the left-
hand at rest during the immobilization period. Back in our
lab, none of the participants reported having removed the
splint when we asked them. The fourth group served as the
control (i.e., without immobilization).
Before the splint removal, two of the three immobilized
groups participated in a motor imagery session for 15 min,
whereas the third immobilized group did not practice motor
imagery (NoMIP). The first two groups differed with
respect to the imagery instructions.1 The participants who
practiced visual imagery (VisMIP) had to imagine them-
selves producing specific hand and fingers movements,
which focused on visual information. The participants who
practiced kinesthetic imagery (KinMIP) were asked to feel
themselves producing specific hand and finger movements,
which focused on bodily information. MIP was divided
into three series of 5 min. In each series, the participants
repeated four motor imagery exercises with the immobi-
lized (left) hand, after having physically executed it with
the non-immobilized (right) hand (with vision for the
VisMIP group, without vision for the KinMIP group; see
Toussaint & Blandin, 2010 for the importance of the sen-
sory-specific experience prior to motor imagery). In the
first exercise, the participants simulated finger tapping
movements (i.e., alternatively touched the thumb starting
with the index, middle, ring and little fingers and then in
the opposite direction). The second and third exercises
consisted of adduction/abduction and flexion/extension
movements of the wrist, respectively. In the fourth exer-
cise, they imagined successively clenching and unclen-
ching their hand. In each exercise, the participants
simulated *25 hand and finger movement repetitions with
the immobilized hand. The participants kept their eyes
closed during each mental exercise. The experimenter
ensured that the participants kept their eyes closed during
each mental exercise, by watching them continuously. The
participants were instructed to say ‘‘top’’ when they started
and finished a repetition. Chronometry of the imagined
movement was used to gauge the engagement in the MIP
training and to control that all participants practiced the
motor imagery for *15 min.
The hand identification task was performed immediately
after the splint removal for the immobilized groups. For all
participants (control, NoMIP, VisMIP and KinMIP
groups), the task was divided into two phases. During the
first familiarization phase, the participants were shown 8
randomly presented trials (2 hands 9 2 grips 9 2 direc-
tions). No time constraint was imposed during the famil-
iarization phase. During the second experimental phase, the
participants were shown 6 blocks of 32 trials (i.e., 192 trials
Left-oriented hand Right-oriented hand
Small sphere
Large sphere
Fig. 1 Illustration of the right-hand stimuli used in the present
experiment. Mirror images were used for the left-hand stimuli
1 Note that visual imagery is less motor than kinesthetic imagery.
Psychological Research
123
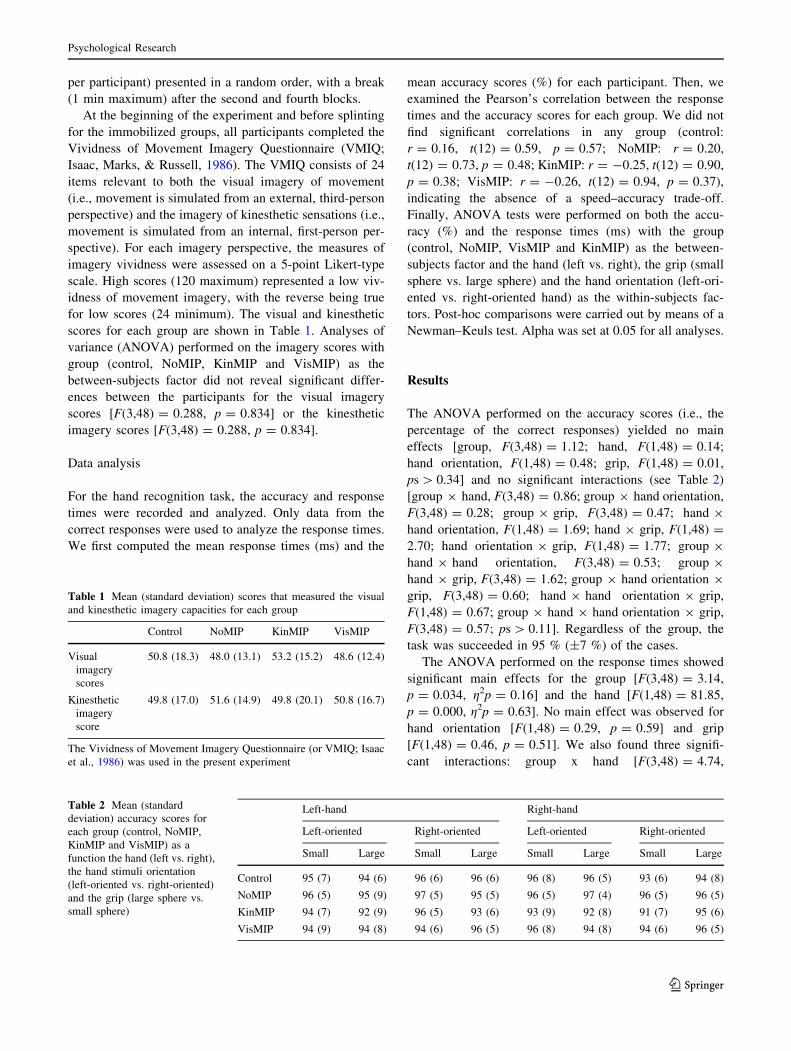
per participant) presented in a random order, with a break
(1 min maximum) after the second and fourth blocks.
At the beginning of the experiment and before splinting
for the immobilized groups, all participants completed the
Vividness of Movement Imagery Questionnaire (VMIQ;
Isaac, Marks, & Russell, 1986). The VMIQ consists of 24
items relevant to both the visual imagery of movement
(i.e., movement is simulated from an external, third-person
perspective) and the imagery of kinesthetic sensations (i.e.,
movement is simulated from an internal, first-person per-
spective). For each imagery perspective, the measures of
imagery vividness were assessed on a 5-point Likert-type
scale. High scores (120 maximum) represented a low viv-
idness of movement imagery, with the reverse being true
for low scores (24 minimum). The visual and kinesthetic
scores for each group are shown in Table 1. Analyses of
variance (ANOVA) performed on the imagery scores with
group (control, NoMIP, KinMIP and VisMIP) as the
between-subjects factor did not reveal significant differ-
ences between the participants for the visual imagery
scores [F(3,48) = 0.288, p = 0.834] or the kinesthetic
imagery scores [F(3,48) = 0.288, p = 0.834].
Data analysis
For the hand recognition task, the accuracy and response
times were recorded and analyzed. Only data from the
correct responses were used to analyze the response times.
We first computed the mean response times (ms) and the
mean accuracy scores (%) for each participant. Then, we
examined the Pearson’s correlation between the response
times and the accuracy scores for each group. We did not
find significant correlations in any group (control:
r = 0.16, t(12) = 0.59, p = 0.57; NoMIP: r = 0.20,
t(12) = 0.73, p = 0.48; KinMIP: r = -0.25, t(12) = 0.90,
p = 0.38; VisMIP: r = -0.26, t(12) = 0.94, p = 0.37),
indicating the absence of a speed–accuracy trade-off.
Finally, ANOVA tests were performed on both the accu-
racy (%) and the response times (ms) with the group
(control, NoMIP, VisMIP and KinMIP) as the between-
subjects factor and the hand (left vs. right), the grip (small
sphere vs. large sphere) and the hand orientation (left-ori-
ented vs. right-oriented hand) as the within-subjects fac-
tors. Post-hoc comparisons were carried out by means of a
Newman–Keuls test. Alpha was set at 0.05 for all analyses.
Results
The ANOVA performed on the accuracy scores (i.e., the
percentage of the correct responses) yielded no main
effects [group, F(3,48) = 1.12; hand, F(1,48) = 0.14;
hand orientation, F(1,48) = 0.48; grip, F(1,48) = 0.01,
ps [ 0.34] and no significant interactions (see Table 2)
[group 9 hand, F(3,48) = 0.86; group 9 hand orientation,
F(3,48) = 0.28; group 9 grip, F(3,48) = 0.47; hand 9
hand orientation, F(1,48) = 1.69; hand 9 grip, F(1,48) =
2.70; hand orientation 9 grip, F(1,48) = 1.77; group 9
hand 9 hand orientation, F(3,48) = 0.53; group 9
hand 9 grip, F(3,48) = 1.62; group 9 hand orientation 9
grip, F(3,48) = 0.60; hand 9 hand orientation 9 grip,
F(1,48) = 0.67; group 9 hand 9 hand orientation 9 grip,
F(3,48) = 0.57; ps [ 0.11]. Regardless of the group, the
task was succeeded in 95 % (±7 %) of the cases.
The ANOVA performed on the response times showed
significant main effects for the group [F(3,48) = 3.14,
p = 0.034, g2p = 0.16] and the hand [F(1,48) = 81.85,
p = 0.000, g2p = 0.63]. No main effect was observed for
hand orientation [F(1,48) = 0.29, p = 0.59] and grip
[F(1,48) = 0.46, p = 0.51]. We also found three signifi-
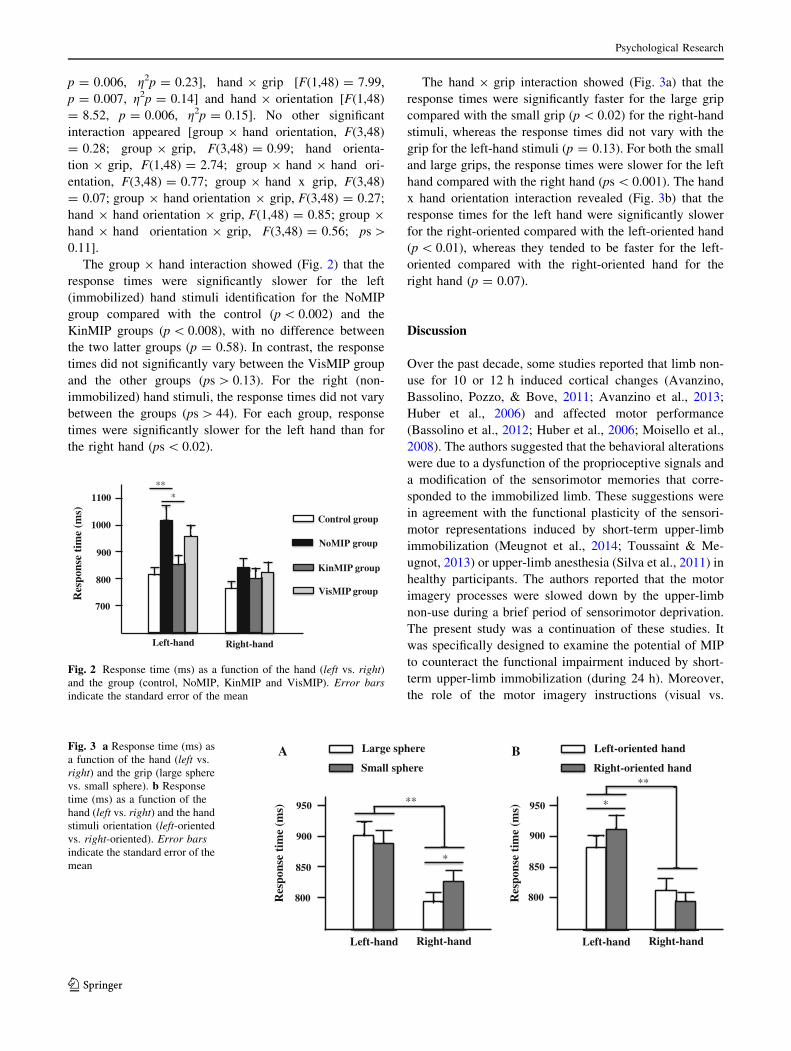
cant interactions: group x hand [F(3,48) = 4.74,
Table 1 Mean (standard deviation) scores that measured the visual
and kinesthetic imagery capacities for each group
Control NoMIP KinMIP VisMIP
Visual
imagery
scores
50.8 (18.3) 48.0 (13.1) 53.2 (15.2) 48.6 (12.4)
Kinesthetic
imagery
score
49.8 (17.0) 51.6 (14.9) 49.8 (20.1) 50.8 (16.7)
The Vividness of Movement Imagery Questionnaire (or VMIQ; Isaac
et al., 1986) was used in the present experiment
Table 2 Mean (standard
deviation) accuracy scores for
each group (control, NoMIP,
KinMIP and VisMIP) as a
function the hand (left vs. right),
the hand stimuli orientation
(left-oriented vs. right-oriented)
and the grip (large sphere vs.
small sphere)
Left-hand Right-hand
Left-oriented Right-oriented Left-oriented Right-oriented
Small Large Small Large Small Large Small Large
Control 95 (7) 94 (6) 96 (6) 96 (6) 96 (8) 96 (5) 93 (6) 94 (8)
NoMIP 96 (5) 95 (9) 97 (5) 95 (5) 96 (5) 97 (4) 96 (5) 96 (5)
KinMIP 94 (7) 92 (9) 96 (5) 93 (6) 93 (9) 92 (8) 91 (7) 95 (6)
VisMIP 94 (9) 94 (8) 94 (6) 96 (5) 96 (8) 94 (8) 94 (6) 96 (5)
Psychological Research
123
p = 0.006, g2p = 0.23], hand 9 grip [F(1,48) = 7.99,
p = 0.007, g2p = 0.14] and hand 9 orientation [F(1,48)
= 8.52, p = 0.006, g2p = 0.15]. No other significant
interaction appeared [group 9 hand orientation, F(3,48)
= 0.28; group 9 grip, F(3,48) = 0.99; hand orienta-
tion 9 grip, F(1,48) = 2.74; group 9 hand 9 hand ori-
entation, F(3,48) = 0.77; group 9 hand x grip, F(3,48)
= 0.07; group 9 hand orientation 9 grip, F(3,48) = 0.27;
hand 9 hand orientation 9 grip, F(1,48) = 0.85; group 9
hand 9 hand orientation 9 grip, F(3,48) = 0.56; ps [0.11].
The group 9 hand interaction showed (Fig. 2) that the
response times were significantly slower for the left
(immobilized) hand stimuli identification for the NoMIP
group compared with the control (p \ 0.002) and the
KinMIP groups (p \ 0.008), with no difference between
the two latter groups (p = 0.58). In contrast, the response
times did not significantly vary between the VisMIP group
and the other groups (ps [ 0.13). For the right (non-
immobilized) hand stimuli, the response times did not vary
between the groups (ps [ 44). For each group, response
times were significantly slower for the left hand than for
the right hand (ps \ 0.02).
The hand 9 grip interaction showed (Fig. 3a) that the
response times were significantly faster for the large grip
compared with the small grip (p \ 0.02) for the right-hand
stimuli, whereas the response times did not vary with the
grip for the left-hand stimuli (p = 0.13). For both the small
and large grips, the response times were slower for the left
hand compared with the right hand (ps \ 0.001). The hand
x hand orientation interaction revealed (Fig. 3b) that the
response times for the left hand were significantly slower
for the right-oriented compared with the left-oriented hand
(p \ 0.01), whereas they tended to be faster for the left-
oriented compared with the right-oriented hand for the
right hand (p = 0.07).
Discussion
Over the past decade, some studies reported that limb non-
use for 10 or 12 h induced cortical changes (Avanzino,
Bassolino, Pozzo, & Bove, 2011; Avanzino et al., 2013;
Huber et al., 2006) and affected motor performance
(Bassolino et al., 2012; Huber et al., 2006; Moisello et al.,
2008). The authors suggested that the behavioral alterations
were due to a dysfunction of the proprioceptive signals and
a modification of the sensorimotor memories that corre-
sponded to the immobilized limb. These suggestions were
in agreement with the functional plasticity of the sensori-
motor representations induced by short-term upper-limb
immobilization (Meugnot et al., 2014; Toussaint & Me-
ugnot, 2013) or upper-limb anesthesia (Silva et al., 2011) in
healthy participants. The authors reported that the motor
imagery processes were slowed down by the upper-limb
non-use during a brief period of sensorimotor deprivation.
The present study was a continuation of these studies. It
was specifically designed to examine the potential of MIP
to counteract the functional impairment induced by short-
term upper-limb immobilization (during 24 h). Moreover,
the role of the motor imagery instructions (visual vs.
Control group
VisMIP group
KinMIP group
NoMIP group
Res
pons
eti
me
(ms)
900
800
700
Left-hand Right-hand
1000
***1100
Fig. 2 Response time (ms) as a function of the hand (left vs. right)
and the group (control, NoMIP, KinMIP and VisMIP). Error bars
indicate the standard error of the mean
Res
pons
eti
me
(ms)
900
850
800
Left-hand Right-hand
950
Large sphere
Small sphere
*
**
Res
pons
eti
me
(ms)
900
850
800
Left-hand Right-hand
950
Left-oriented hand
Right-oriented hand
*
**
A BFig. 3 a Response time (ms) as
a function of the hand (left vs.
right) and the grip (large sphere
vs. small sphere). b Response
time (ms) as a function of the
hand (left vs. right) and the hand
stimuli orientation (left-oriented
vs. right-oriented). Error bars
indicate the standard error of the
mean
Psychological Research
123

kinesthetic) in reactivating the sensorimotor processes
(assessed using a motor imagery task) was of particular
interest in this experiment.
Short-term upper-limb immobilization
and the slowdown of the sensorimotor processes
As previously observed for the hand mental rotation tasks
(Meugnot et al., 2014; Toussaint & Meugnot, 2013), the
results of the present experiment showed that the immo-
bilization-induced effects became widespread in the hand-
grasping object recognition. One day of sensorimotor
restriction for the NoMIP group induced a slowdown of the
hand-stimuli recognition (i.e., slower response times) that
corresponded to the immobilized hand only compared with
the performance of the participants who did not undergo
the immobilization procedure (the control group). The
immobilization-induced effects provided evidence for the
sensorimotor representations involvement in solving the
hand-grasping object task and confirmed the embodied
nature of the motor imagery processes (Meugnot et al.,
2014). The involvement of the sensorimotor representa-
tions when identifying hand laterality was confirmed in the
present experiment by the slower identification processes
for the left (non-dominant) hand compared with the right
hand in the four groups (see also Gentilucci, Daprati, &
Gantigano, 1998; Ionta & Blanke, 2009; Ni Choisdealbha,
Brady, & Maguiness, 2011; Parsons, 1987, 1994, for sim-
ilar results). The greater expertise of the dominant hand
(vs. the non-dominant hand), which is associated with a
finer sensorimotor representation, is at the origin of the
difference in the response times between the hands for both
actual and simulated movements.
Did the immobilization-induced effects vary as a function
of the accuracy requirement of the simulated movements
(i.e., small vs. large sphere grasping)? In the present exper-
iment, the results indicated that left-hand non-use (i.e., the
non-dominant hand) was not the appropriate way to answer
this question. Contrary to Gentilucci et al. (2000), who
reported an increase in the response times for the small
sphere compared with the large sphere for both left and right
hands, we observed a grip-precision effect for the right hand
only. In our experiment, grasping small spheres was more
time-consuming compared with grasping large spheres for
the dominant hand only. This lower engagement of the
sensorimotor processes when identifying the left-hand
stimuli was also suggested by the influence of the spatial
orientation of the hand pictures on the screen. The effect of
the spatial incompatibility, which was more important for the
left-hand stimuli, suggested its greater dependence on the
visual context. The overall results for the right- and left-hand
differences indicate that the use of a motor imagery strategy
to solve the hand laterality task was stronger in identifying
the dominant hand compared with the non-dominant hand.
Therefore, these unexpected results show that immobilizing
the left non-dominant hand of the participants was not the
right solution to determine whether the effects of sensori-
motor deprivation varied according to the precision of the
simulated movements. Note, however, that the sensorimotor
processes remained involved in the identification of the left-
hand stimuli. As reported at the beginning of this section,
24 h of left-hand immobilization effectively led to the
slowdown of the left-hand movement simulation, without
affecting the motor imagery processes for the right-hand.
Motor imagery practice and the reactivation
of the sensorimotor processes
The major aim of the present experiment was to examine
the potential role of MIP in reactivating the sensorimotor
processes that have been slowed down by short-term upper-
limb immobilization. Importantly, we questioned the
influence of the motor imagery instructions (visual vs.
kinesthetic) in counteracting the deleterious effect of 24 h
of sensorimotor deprivation (i.e., left-hand immobilization).
The results of the present experiment showed that the
response times were faster for the left-hand identification
for the participants who practiced 15 min of kinesthetic
motor imagery (KinMIP group) compared with the par-
ticipants who did not practice imagery (NoMIP group).
Moreover, although imagery abilities were similar for the
KinMIP and VisMIP groups (i.e., ‘‘clear and vivid’’ or
‘‘fairly clear and vivid’’, as determined by the VMIQ
scores), a visual imagery practice did not improve the
performance of the immobilized participants, whereas a
kinesthetic imagery practice did improve the performance.
In accordance with neuroimaging (Guillot et al., 2009;
Solodkin et al., 2004) and behavioral studies (Fery, 2003;
Hardy & Callow, 1999; Toussaint & Blandin, 2010), this
finding supports a neuronal and functional distinction
between kinesthetic and visual imagery, with kinesthetic
imagery having a greater association with the sensorimotor
system compared with visual imagery, which would pre-
dominantly activate the visual association cortex (Guillot
et al., 2009). These results clearly indicated that a kines-
thetic MIP was particularly effective in reactivating the
sensorimotor representations of the immobilized hand,
thereby canceling the slowdown of the motor imagery
processes induced by sensorimotor deprivation. As previ-
ously suggested (Moisello et al., 2008; Toussaint & Me-
ugnot, 2013) and in line with the theory of internal models
(Wolpert & Flanagan, 2001), it is likely that the acute
decrease in the sensory and motor flow into the CNS during
the 24 h of limb non-use disrupted the sensorimotor
memories, thereby affecting the ability of the forward
processes to accurately form the movement predictions.
Psychological Research
123

Indeed, the central representation theory (Jeannerod, 2001)
states that motor imagery relies on the same cognitive
processes as motor execution. During simulation, the for-
ward models would provide sensory and motor information
similar to that of the physical execution to mentally predict
the movement (i.e., in the laterality task, the future states
and sensory consequences of hand matching the stimulus
on the computer screen). During the period of limb non-
use, the processing of the input/output in the contralateral
area was greatly reduced, and the sensorimotor represen-
tations were no longer updated. Thus, the left-hand inner
memories would be less vivid and less easily retrieved by
the forward models after immobilization. However, during
a kinesthetic MIP, proprioceptive and motor processing
would be progressively reactivated to simulate left-hand
movements. The motor planning routines would progres-
sively recover their initial level of functioning.
The present experiment showed that kinesthetic MIP is
particularly effective in reactivating the motor imagery pro-
cesses following short-term limb non-use. However, no clear
conclusion can be drawn with regard to visual MIP. The
results did not reveal a significant difference in hand recog-
nition between the VisMIP and KinMIP groups, as well as
between the VisMIP and NoMIP groups. These ambiguous
results could be due to several possibilities. First, we cannot
exclude the possibility that participants who performed a
visual MIP still relayed on residual proprioceptive informa-
tion. Indeed, before practicing Visual MIP, participants exe-
cuted right hand movements with both visual and
proprioceptive information. However, the residual of propri-
oceptive information could contribute to sensorimotor reac-
tivation without being sufficient enough to induce a significant
improvement compared with the KinMIP group. Second, the
benefit of visual MIP might necessitate more times to reacti-
vate the sensorimotor system following a brief period of
sensorimotor deprivation compared with kinesthetic MIP.
Visual motor imagery is known to predominately activate the
visual association cortex, whereas kinesthetic motor imagery
has stronger association with the sensorimotor system (Guillot
et al., 2009). It may be possible that the lesser association of
visual motor imagery with the sensorimotor system, combined
with the slowdown of the sensorimotor processes due to
immobilization, decreases the potential benefit of visual
imagery. Whether a longer visual MIP will effectively help to
reactivate the sensorimotor system following a relatively short
period of limb immobilization is a relevant question that must
be clarified in the future.
Conclusion
Overall, the results of the present experiment showed for
the first time that a kinesthetic MIP was efficient in
counteracting the deleterious effect of short-term upper-
limb immobilization on motor imagery processes. When
movement is not possible, the kinesthetic imagery rehearsal
would allow an individual to maintain the recursive pro-
cessing of sensory and motor inflow into the system and
prevent the disruption of sensorimotor memories. To our
knowledge, only one study has investigated the potential of
kinesthetic imagery in modulating the brain changes fol-
lowing short-term limb non-use (Bassolino, Campanella,
Bove, Pozzo, & Fadiga, 2013). The authors used trans-
cranial magnetic stimulation to assess the effect of short-
term immobilization (10 h) on the excitability of the pri-
mary motor cortex contralateral to the limb immobilization
(cM1). Moreover, they examined the effect of a kinesthetic
MIP during the period of sensorimotor restriction to pre-
vent the cortical depression. Surprisingly, they observed no
compensatory effect and concluded that motor imagery
was not effective in modulating brain plasticity during
inactivity. In fact, the contribution of the cM1 during motor
imagery remains controversial, as neuroimaging studies did
not systematically report the activation of the cM1 during
motor imagery (De Vries & Mulder, 2007; Guillot, Di
Rienzo, MacIntyre, Moran, & Collet, 2012). In contrast,
there is substantial evidence that kinesthetic motor imagery
involves many others brain structures (i.e., the ventral and
dorsal parts of the premotor cortex, the supplementary
motor area, the inferior parietal lobule; Gerardin et al.,
2000; Guillot et al., 2009; Munzert, Lorey, & Zentgraf,
2009; Solodkin et al., 2004). Therefore, the lack of benefit
of kinesthetic imagery practice on the cM1 excitability
does not necessarily indicate that MIP is not a reliable tool
for reactivating the sensorimotor system during limb non-
use. In fact, the results of the present experiment support
the use of kinesthetic MIP.
The present study supports the potential use of a kin-
esthetic MIP in a rehabilitation program as a means of
promoting the recovery of motor function, especially when
physical practice is not possible. In fact, the potential of MI
in rehabilitation is now well accepted, even though no
definite conclusions on the effects of mental practice can be
drawn at this time because the evidence base is relatively
small (Braun et al., 2013; Malouin et al., 2013). In the
specific case of limb disuse, to our knowledge, only two
studies have examined the potential of mental practice on
functional recovery (Crews & Kamen, 2006; Stenekes,
Geertzen, Nicolai, De Jong, & Mulder, 2009). However, in
these studies, the positive effect of motor imagery on motor
performance following at least 1 week of immobilization
was rather weak or non-significant. Further research on the
use of kinesthetic motor imagery in injury rehabilitation
(i.e., with immobilized patients after the effects of ortho-
pedic injury, such as sprain, fracture or surgical interven-
tion) would be of interest. One cannot exclude the
Psychological Research
123

possibility that a kinesthetic MIP is effective only when
sensorimotor deprivation is transient. As reported in studies
examining the effect of long-term loss of kinesthetic
afferents on the sensorimotor representation (ter Horst
et al., 2012), the availability of kinesthetic afferent signals
is crucial to incorporate bodily information into the
imagery processes. Therefore, when patients are immobi-
lized longer because of injury, a visual MIP could be more
effective to construct a body representation than a kines-
thetic MIP. Studies in healthy individuals have shown that
motor imagery use in addition to physical practice allows a
reduction in the amount of physical practice required to
attain a given level of motor performance (Allami, Pau-
lignan, Brovelli, & Boussaoud, 2008; Pascual-Leone et al.,
1995). The challenge over the coming decades will be to
ensure the field is readily amenable to integrating MIP into
the current standard of care used in clinical practice.
References
Allami, L., Paulignan, Y., Brovelli, A., & Boussaoud, D. (2008).
Visuo-motor learning with combination of different rates of
motor imagery and physical practice. Experimental Brain
Research, 184, 105–113. doi:10.1007/s00221-007-1086-x.
Avanzino, L., Bassolino, M., Pozzo, T., & Bove, M. (2011). Use-
dependent hemispheric balance. Journal of Neuroscience, 31,
3423–3428. doi:10.1523/JNEUROSCI.4893-10.2011.
Avanzino, L., Pelosin, E., Abbruzzese, G., Bassolino, M., Pozzo, T.,
& Bove, M. (2013). Shaping motor cortex plasticity through
proprioception. Cerebral Cortex,. doi:10.1093/cercor/bht139.
Bassolino, M., Bove, M., Jacono, M., Fadiga, L., & Pozzo, T. (2012).
Functional effect of short-term immobilization: kinematic
changes and recovery on reaching-to-grasp. Neuroscience, 215,
127–134. doi:10.1016/j.neuroscience.2012.04.019.
Bassolino, M., Campanella, M., Bove, M., Pozzo, T., & Fadiga, L.
(2013). Training the motor cortex by observing the actions of
others during immobilization. Cerebral Cortex,. doi:10.1093/
cercor/bht190.
Braun, S., Kleynen, M., van Heel, T., Kruithof, N., Wade, D., &
Beurskens, A. (2013). The effects of mental practice in
neurological rehabilitation; a systematic review and meta-
analysis. Frontiers in Human Neuroscience,. doi:10.3389/
fnhum.2013.00390.
Cerritelli, B., Maruff, P., Wilson, P., & Currie, J. (2000). The effect of
an external load on the force and timing components of mentally
represented actions. Behavioral Brain Research, 108, 91–96.
doi:10.1016/S0166-4328(99)00138-2.
Conson, M., Mazzarella, E., Frolli, A., Esposito, D., Marino, N.,
Trojano, L., Massagli, A., Gison, G., Aprea, N., & Grossi, D.
(2013). Motor imagery in asperger syndrome: testing action
simulation by the hand laterality task. PLoS One, 8. doi:10.1371/
journal.pone.0070734.
Crews, R. T., & Kamen, G. (2006). Motor-evoked potentials
following imagery and limb disuse. The International Journal
of Neuroscience, 116, 639–651. doi:10.1080/002074506
00592198.
Curtze, C., Otten, B., & Postema, K. (2010). Effects of lower limb
amputation on the mental rotation of feet. Experimental Brain
Research, 201, 527–534. doi:10.1007/s00221-009-2067-z.
de Vignemont, F., Zalla, T., Posada, A., Louvegnez, A., Koenig, O.,
Georgieff, N., et al. (2006). Mental rotation in schizophrenia.
Consciousness and Cognition, 15, 295–309. doi:10.1016/j.
concog.2005.08.001.
de Vries, S., & Mulder, T. (2007). Motor imagery and stroke
rehabilitation: a critical discussion. Journal of Rehabilitation
Medicine, 39, 5–13. doi:10.2340/16501977-0020.
Decety, J., & Grezes, J. (1999). Neural mechanisms subserving the
percep- tion of human actions. Trends in Cognitive Sciences, 3,
172–178. doi:10.1016/S1364-6613(99)01312-1.
Decety, J., & Jeannerod, M. (1995). Mentally simulated movements
in virtual reality: does Fitts’s law hold in motor imagery?Behavioral Brain Research, 72, 127–134. doi:10.1016/0166-
4328(96)00141-6.
Dominey, P., Decety, J., Brousolle, E., Chazot, J., & Jeannerod, M.
(1994). Motor imagery of a lateralized sequential task is
asymmetrically slowed in hemi-Parkinson’s patients. Neuro-
psychologia, 33, 727–741. doi:10.1016/0028-3932(95)00008-Q.
Driskell, J. E., Copper, C., & Moran, A. (1994). Does mental practice
enhance performance? Journal of Applied Psychology, 79,
481–492. doi:10.1037/0021-9010.79.4.481.
Dunsky, A., Dickstein, R., Marcovitz, E., Levy, S., & Deutsch, J. E.
(2008). Home-based MI training for gait rehabilitation of people
with chronic post-stroke hemiparesis. Archive of Physical
Medicine and Rehabilitation, 89, 1580–1588. doi:10.1016/j.
apmr.2007.12.039.
Feltz, D. L., & Landers, D. M. (1983). The effects of mental practice
on motor skill learning and performance: a meta-analysis.
Journal of Sport & Exercise Psychology, 5, 25–57.
Fery, Y. A. (2003). Differentiating visual and kinesthetic imagery in
mental practice. Canadian Journal of Experimental Psychology,
57, 1–10.
Fiorio, M., Tinazzi, M., & Aglioti, S. M. (2006). Selective impair-
ment of hand mental rotation in patients with focal hand
dystonia. Brain, 129, 47–54. doi:10.1093/brain/awh630.
Funk, M., & Brugger, P. (2008). Mental rotation of congenitally
absent hands. Journal of the International Neuropsychological
Society, 14, 81–89.
Gentili, R., Han, C. E., Schweighofer, N., & Papaxanthis, C. (2010).
Motor learning without doing: trial-by-trial improvement in
motor performance during mental training. Journal of Neuro-
physiology, 104, 774–783. doi:10.1152/jn.00257.2010.
Gentilucci, M., Benuzzi, F., Bertolani, L., Daprati, E., & Gangitano,
M. (2000). Recognising a hand by grasp. Cognitive Brain
Research, 9, 125–135. doi:10.1016/S0926-6410(99)00049-X.
Gentilucci, M., Daprati, E., & Gangitano, M. (1998). Right-handers
and left-handers have different representations of their own
hand. Cognitive Brain Research, 6, 185–192. doi:10.1016/
S0926-6410(97)00034-7.
Gerardin, E., Sirigu, A., Lehericy, S., Poline, J. B., Gaymard, B.,
Marsault, C., et al. (2000). Partially overlapping neural networks
for real and imagined hand movements. Cerebral Cortex, 10,
1093–1104. doi:10.1093/cercor/10.11.1093.
Guillot, Di Rienzo, F., MacIntyre, T., Moran, A., & Collet, C. (2012).
Imagining is not doing but involves specific motor commands: a
review of experimental data related to motor inhibition. Frontiers
in Human Neuroscience, 6, 247. doi:10.3389/fnhum.2012.00247.
Guillot, A., & Collet, C. (2005). Contribution from neurophysiologi-
cal and psychological methods to the study of motor imagery.
Brain Research, 50, 387–397. doi:10.1016/j.brainresrev.2005.09.
004.
Guillot, A., & Collet, C. (2008). Construction of the motor imagery
integrative model in sport: a review and theoretical investigation
of motor imagery use. International Review of Sport and
Exercise Psychology, 1, 31–44. doi:10.1080/1750984070
1823139.
Psychological Research
123

Guillot, A., Collet, C., & Dittmar, A. (2004). Relationship between
visual vs. kinesthetic imagery, field dependence-independence
and complex motor skills. Journal of Psychophysiology, 18,
190–199. doi:10.1027/0269-8803.18.4.190.
Guillot, A., Collet, C., Nguyen, V. A., Malouin, F., Richards, C., &
Doyon, J. (2009). Brain activity during visual versus kinesthetic
imagery: an fMRI study. Human Brain Mapping, 30,
2157–2172. doi:10.1002/hbm.20658.
Hanakawa, T., Dimyan, M. A., & Hallett, M. (2008). Motor planning,
imagery, and execution in the distributed motor network: a time-
course study with functional MRI. Cerebral Cortex, 18,
2775–2788. doi:10.1093/cercor/bhn036.
Hardy, L., & Callow, N. (1999). Efficacy of external and internal
visual imagery perspectives for the enhancement of performance
on tasks in which form is important. Journal of Sport and
Exercise Psychology, 21, 95–112.
Huber, R., Ghilardi, M. F., Massimini, M., Ferrarelli, F., Riedner, B.
A., Peterson, M. J., et al. (2006). Arm immobilization causes
cortical plastic changes and locally decreases sleep slow wave
activity. Nature Neuroscience, 9, 1169–1176. doi:10.1038/
nn1758.
Ionta, S., & Blanke, O. (2009). Differential influence of hands posture
on mental rotation of hands and feet in left and right handers.
Experimental Brain Research, 195, 207–217. doi:10.1007/
s00221-009-1770-0.
Isaac, A., Marks, D. F., & Russell, D. G. (1986). An instrument for
assessing imagery of movement: the Vividness of Movement
Imagery Questionnaire (VMIQ). Journal of Mental Imagery, 10,
23–30.
Jackson, P. L., Lafleur, M. F., Malouin, F., Richards, C., & Doyon, J.
(2001). Potential role of mental practice using motor imagery in
neurologic rehabilitation. Archive of Physical Medicine and
Rehabilitation, 82, 1133–1141. doi:10.1053/apmr.2001.24286.
Jackson, P. L., Lafleur, M. F., Malouin, F., Richards, C. L., & Doyon,
J. (2003). Functional cerebral reorganization following motor
sequence learning through mental practice with motor imagery.
Neuroimage, 20, 1171–1180. doi:10.1016/S1053-8119(03)
00369-0.
Jeannerod, M. (2001). Neural simulation of action: a unifying
mechanism for motor cognition. Neuroimage, 14, 429–439.
doi:10.1006/nimg.2001.0832.
Jenkinson, P. M., Edelstyn, N. M. J., & Ellis, S. J. (2009). Imagining
the impossible: motor representations in anosognosia for hemi-
plegia. Neuropsychologia, 47, 481–488. doi:10.1016/j.neuropsy
chologia.2008.10.004.
Lacourse, M. G., Turner, J. A., Randolph-Orr, E., Schandler, S. L., &
Cohen, M. J. (2004). Cerebral and cerebellar sensorimotor
plasticity following motor imagery-based mental practice of a
sequential movement. Journal of Rehabilitation Research and
Development, 41, 505–524.
Malouin, F., Jackson, P., & Richards, C.L. (2013). Towards the
integration of mental practice in rehabilitation programs. A
critical review. Frontiers in Human Neuroscience. doi:10.3389/
fnhum.2013.00576.
Malouin, F., Richards, C. L., Durand, A., Descent, M., Poire, D.,
Fremont, P., et al. (2009). Effects of practice, visual loss, limb
amputation, and disuse on motor imagery vividness. Neurore-
habilitation Neural Repair, 23, 449–463. doi:10.1177/
1545968308328733.
Malouin, F., Richards, C. L., Durand, A., & Doyon, J. (2008). Clinical
assessment of motor imagery after stroke. Neurorehabilitation
and Neural Repair, 22, 330–340. doi:10.1177/15459
68307313499.
Meugnot, A., Almecija, Y., & Toussaint, L. (2014). The embodied
nature of motor imagery processes highlighted by short-term
limb immobilization. Experimental Psychology, 61, 180–186.
doi:10.1027/1618-3169/a000237.
Moisello, C., Bove, M., Huber, R., Abbruzzese, G., Battaglia, F.,
Tononi, G., et al. (2008). Short-term limb immobilization affects
motor performance. Journal of Motor Behavior, 40, 165–176.
doi:10.3200/JMBR.40.2.165-176.
Mulder, T. (2007). Motor imagery and action observation: cognitive
tools for rehabilitation. Journal of Neural Transmission, 114,
1265–1278. doi:10.1007/s00702-007-0763-z.
Munzert, J., Lorey, B., & Zentgraf, K. (2009). Cognitive motor
processes: the role of motor imagery in the study of motor
representations. Brain Research Review, 60, 306–326. doi:10.
1016/j.brainresrev.2008.12.024.
Ni Choisdealbha, A., Brady, N., & Maguinness, C. (2011). Differing
roles for the dominant and non-dominant hands in the hand
laterality task. Experimental Brain Research, 211, 73–85. doi:10.
1007/s00221-011-2652-9.
Nico, D., Daprati, E., Rigal, F., Parsons, L. M., & Sirigu, A. (2004).
Left and right hand recognition in upper limb amputees. Brain,
127, 120–132. doi:10.1093/brain/awh006.
Papaxanthis, C., Pozzo, T., Skoura, X., & Schiappati, M. (2002). Does
order and timing in performance of imagined and actual
movements affect the motor imagery process? The duration of
walking and writing task. Behavioral Brain Research, 134,
209–215. doi:10.1016/S0166-4328(02)00030-X.
Parsons, L. M. (1987). Imagined spatial transformations of one’s
body. Cognitive Psychology, 19, 178–241. doi:10.1016/0010-
0285(87)90011-9.
Parsons, L. M. (1994). Temporal and kinematic properties of motor
behavioral reflected in mentally simulated action. Journal of
Experimental Psychology: Human Perception and Performance,
20, 709–730. doi:10.1037/0096-1523.20.4.709.
Pascual-Leone, A., Nguyet, D., Cohen, L. G., Brasil-Neto, J. P.,
Cammarota, A., & Hallett, M. (1995). Modulation of muscle
responses evoked by transcranial magnetic stimulation during
the acquisition of new fine motor skills. Journal of Neurophys-
iology, 74, 1037–1045.
Petit, L. S., Pegna, A. J., Mayer, E., & Hauert, C. A. (2003).
Representation of anatomical constraints in motor imagery:
mental rotation of a body segment. Brain and Cognition, 51,
95–101. doi:10.1016/S0278-2626(02)00526-2.
Ruby, P., & Decety, J. (2003). What you believe versus what you
think they believe: a neuroimaging study of conceptual perspec-
tive-taking. European Journal of Neuroscience, 17, 2475–2480.
doi:10.1046/j.1460-9568.2003.02673.x.
Silva, S., Loubinoux, I., Olivier, M., Bataille, B., Fourcade, O., Samii,
K., et al. (2011). Impaired visual hand recognition in Preoper-
ative patients during Brachial Plexus. Anesthesiology, 1,
126–134. doi:10.1097/ALM.0b013e31820164f1.
Sirigu, A., Daprati, E., Pradat-Diehl, P., Franck, N., & Jeannerod, M.
(1999). Perception of self-generated movement following left
parietal lesion. Brain, 122, 1867–1874. doi:10.1093/brain/122.
10.1867.
Sirigu, A., Duhamel, J. R., Cohen, L., Pillon, B., Dubois, B., & Agid,
Y. (1996). The mental representation of hand movements after
parietal cortex damage. Science, 273, 1564–1568.
Solodkin, A., Hlustik, P., Chen, E. E., & Small, S. L. (2004). Fine
modulation in network activation during motor execution and
motor imagery. Cerebral Cortex, 14, 1246–1255. doi:10.1093/
cercor/bhh086.
Stenekes, M. W., Geertzen, J. H., Nicolai, J. P., De Jong, B. M., &
Mulder, T. (2009). Effects of motor imagery on hand function
during immobilization after flexor tendon repair. Archive of
Physical Medicine and Rehabilitation, 90, 553–559. doi:10.
1016/j.apmr.2008.10.029.
Psychological Research
123

Stinear, C. M., Byblow, W. D., Steyvers, M., Levin, O., & Swinnen,
S. P. (2006). Kinesthetic, but not visual, motor imagery
modulates cortico-motor excitability. Experimental Brain
Research, 168, 157–164. doi:10.1007/s00221-005-0078-y.
ter Horst, A. C., Cole, J., van Lier, R., & Steenbergen, B. (2012). The
effect of chronic deafferentation on mental imagery: a case
study. PLoS One, 7, e42742. doi:10.1371/jounal.pone.0042742.
Toussaint, L., & Blandin, Y. (2010). On the role of imagery
modalities on motor learning. Journal of Sports Sciences, 28,
497–504. doi:10.1080/02640410903555855.
Toussaint, L., & Meugnot, A. (2013). Short-term limb immobilization
affects cognitive motor processes. Journal of Experimental
Psychology Learning, Memory, and Cognition, 39, 623–632.
doi:10.1037/a0028942.
Wang, Y., & Morgan, W. P. (1992). The effects of imagery
perspectives on the physiological responses to imagined exer-
cise. Behavioral Brain Research, 52, 167–174. doi:10.1016/
S0166-4328(05)80227-X.
White, A., & Hardy, L. (1995). Use of different imagery perspectives
on the learning and performance of different motor skills. British
Journal of Psychology, 86, 191–216.
Wilson, P. H., Maruff, P., Butson, M., Williams, J., Lum, J., &
Thomas, P. R. (2004). Internal representation of movement in
children with developmental coordination disorder: a mental
rotation task. Developmental Medicine and Child Neurology, 46,
754–759. doi:10.1017/S001216220400129X.
Wolpert, D. M., & Flanagan, J. R. (2001). Motor prediction. Current
Biology, 11, 729–732. doi:10.1016/S0960-9822(01)00432-8.
Psychological Research
123















![Increased [CO 2] does not compensate for negative effects on yield caused by higher temperature and [O 3] in Brassica napus L](https://static.fdokumen.com/doc/165x107/631794e7831644824d038e8a/increased-co-2-does-not-compensate-for-negative-effects-on-yield-caused-by-higher.jpg)




