
Abnormal sensorimotor control, but intact force field adaptation, in multiple sclerosis subjects...
14
http://msj.sagepub.com Multiple Sclerosis DOI: 10.1177/1352458507085068 2008; 14; 330 originally published online Jan 21, 2008; Mult Scler Maura Casadio, Vittorio Sanguineti, Pietro Morasso and Claudio Solaro no clinical disability Abnormal sensorimotor control, but intact force field adaptation, in multiple sclerosis subjects with http://msj.sagepub.com/cgi/content/abstract/14/3/330 The online version of this article can be found at: Published by: http://www.sagepublications.com can be found at: Multiple Sclerosis Additional services and information for http://msj.sagepub.com/cgi/alerts Email Alerts: http://msj.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.co.uk/journalsPermissions.nav Permissions: http://msj.sagepub.com/cgi/content/refs/14/3/330 SAGE Journals Online and HighWire Press platforms): (this article cites 28 articles hosted on the Citations at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.com Downloaded from
Transcript of Abnormal sensorimotor control, but intact force field adaptation, in multiple sclerosis subjects...

http://msj.sagepub.com
Multiple Sclerosis
DOI: 10.1177/1352458507085068 2008; 14; 330 originally published online Jan 21, 2008; Mult Scler
Maura Casadio, Vittorio Sanguineti, Pietro Morasso and Claudio Solaro no clinical disability
Abnormal sensorimotor control, but intact force field adaptation, in multiple sclerosis subjects with
http://msj.sagepub.com/cgi/content/abstract/14/3/330 The online version of this article can be found at:
Published by:
http://www.sagepublications.com
can be found at:Multiple Sclerosis Additional services and information for
http://msj.sagepub.com/cgi/alerts Email Alerts:
http://msj.sagepub.com/subscriptions Subscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.co.uk/journalsPermissions.navPermissions:
http://msj.sagepub.com/cgi/content/refs/14/3/330SAGE Journals Online and HighWire Press platforms):
(this article cites 28 articles hosted on the Citations
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

RESEARCH PAPER Multiple Sclerosis 2008; 14: 330–342
© 2008 SAGE Publications 10.1177/1352458507085068Los Angeles, London, New Delhi and Singapore
Abnormal sensorimotor control, but intact force field adaptation, in multiple sclerosis subjects with no clinical disability
Maura Casadio1,2, Vittorio Sanguineti1, Pietro Morasso1 and Claudio Solaro3
In MS subjects with no clinical disability, we assessed sensorimotor organization and their ability toadapt to an unfamiliar dynamical environment. Eleven MS subjects performed reaching movementswhile a robot generated a speed-dependent force field. Control and adaptation performance werecompared with that of an equal number of control subjects. During a familiarization phase, when therobot generated no forces, the movements of MS subjects were more curved, displayed greater andmore variable directional errors and a longer deceleration phase. During the force field phase, bothMS and control subjects gradually learned to predict the robot-generated forces. The rates of adap-tation were similar, but MS subjects showed a greater variability in responding to the force field.These results suggest that MS subjects have a preserved capability of learning to predict the effectsof the forces, but make greater errors when actually using such predictions to generate movements.Inaccurate motor commands are then compensated later in the movement through an extra amountof sensory-based corrections. This indicates that early in the disease MS subjects have intact adaptivecapabilities, but impaired movement execution. Multiple Sclerosis 2008; 14: 330–342.http://msj.sagepub.com
Key words: motor adaptation; motor control; multiple sclerosis; robot therapy
Introduction
In the early stages of MS, about 85% of the sub-jects show a relapsing–remitting course, that is,acute phases followed by clinical recovery.Relapses are often followed by partial or completefunctional recovery. In about 40% of the patients,recovery is incomplete when evaluated after sixmonths, with a residual deficit of 0.5 or moreunits of the expanded disability status scale(EDSS) score [1].
Brain plasticity is one potential contributor tothe recovery of function after tissue damage, and
may limit the functional consequences of axonalloss. Clinical recovery is determined by many fac-tors, such as increased expression of sodium chan-nels, recruitment of silent pathways, remyelination.Cortical and subcortical reorganization may takeplace as well, thus playing a role in limiting theimpact of tissue damage. Magnetic resonance imag-ing (MRI) and, even more, functional MRI (fMRI)studies have provided important insights on themechanisms of tissue damage and functional recov-ery in MS subjects. Functional MRI studies in MSsubjects with a relapsing–remitting course andminimal disability have shown that in simple motor
1 Department of Informatics, Systems and Telematics and Research Centre for Neuroscience and Neuroengineering,University of Genoa, via Opera Pia 13, Genoa 16145, Italy2 Foundation ‘Don Gnocchi’, via Cisa Vecchia, 19038 Sarzana SP, Italy3 Department of Neurology, Hospital PA Micone, Genoa, ItalyAuthor for correspondence: Professor Vittorio Sanguineti, Department of Informatics, Systems and Telematics andResearch Centre for Neuroscience and Neuroengineering, University of Genoa, via Opera Pia 13, Genoa 16145, Italy.E-mail: [email protected] 18 April 2007; revised 30 July 2007; accepted 5 August 2007
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

Abnormal sensorimotor control in MS subjects with no clinical disability 331
http://msj.sagepub.com Multiple Sclerosis 2008; 14: 330–342
tasks, these subjects recruit areas that (in healthysubjects) are typically associated to more complextasks [2]. A recent study provided evidence thatboth inter- and intra-hemispheric motor reorgani-zation occur in MS [3]. These findings point at acortical and subcortical reorganization aimed atcounteracting demyelination and/or neurodegenera-tion [4]. From the behavioral point of view, reorgani-zation may lead to the development of compensatorystrategies.
Although compensatory strategies may leaveperformance relatively unaffected, they may beunveiled by experimental and analytical tech-niques that aim at investigating the mechanismsunderlying sensorimotor control and adaptation.Robotic devices that were originally developed tostudy motor learning in normal subjects can beused to assess the motor functionality of neurolog-ical patients. Recently, an experimental protocolthat had been widely used to investigate motorlearning capabilities [5] was applied to subjectswith degenerative cerebellar atrophy [6,7]. Theresults showed that these subjects completely losttheir adaptation capability.
Recently, it has been shown that MS subjectswith little or no disability have subtle problems ingait and balance [8] and in controlling arm move-ments [9]. Multiple sclerosis subjects with mild dis-ability also displayed a reduced ability to improvetheir performance with exercise in a visual trackingtask [10]. However, it is unclear whether the abilityof adapting to an unfamiliar environment is pre-served in MS subjects with no disability and/or whatis the origin of these problems.
In this study, we asked whether sensorimotoradaptation in MS subjects might contribute tomaintaining a normal level of motor performance.To this purpose, we investigated the ability of MSsubjects with no disability to adapt to velocity-dependent force fields. We also asked whether suchadaptation differs from control subjects either in itsoutcome or in its dynamics.
Materials and Methods
Subjects
Eleven subjects with clinically definite relapsing–remitting MS, according to Poser criteria [11], par-ticipated in this study (three males, eight females).Their mean age was 33.4 years (range 24–44) andthe mean disease duration was five years (range1–15). Patients were recruited at the Department ofNeurology of the Hospital ‘P Antero Micone’ and atthe Department of Neurosciences, Ophthalmologyand Genetics, University of Genova. The inclusioncriteria were: (i) EDSS [12] less than or equal to 1
and (ii) presence of neurological signs only, but nosigns or symptoms at upper limbs and consequently,‘normal’ score for the ‘arm’ portion of the Scripps’neurological rating scale [13] for the sensory, motorand cerebellar systems. The exclusion criteria were:(i) relapses within the last three months; (ii) treat-ment with corticosteroids within the previous threemonths; (iii) mini mental state examination(MMSE) less than 24 and (iv) symptomatic oculo-motor signs or visual acuity less than 8/10. Thesame neurologist examined all the patients.
The performance of these subjects was comparedwith 11 controls – six males, five females, age 29.3(range 22–41) – with no previous history of neuro-logical dysfunction. Demographic and clinicaldetails for each subject are listed in Table 1.
The research conforms to the ethical standardslaid down in the 1964 Declaration of Helsinki thatprotects research subjects. Each subject signed aconsent form that conforms to these guidelines.
Experimental apparatus and task
We used a planar robotic manipulandum (Braccio diFerro, see Figure 1), specifically designed for robottherapy and for the evaluation of motor controland motor adaptation.
The manipulandum – technical details arereported in Casadio et al. (2006) [14] – has a80 � 40 cm elliptic workspace, and is actuated bytwo direct-drive brushless motors, mounted proxi-mally to minimize inertia and frictional forces. Thestructure is extremely rigid, and robot geometryhas been optimized for uniformity of the manipu-lability index and the force/torque ratio over thewhole workspace. The force available at the handlein all directions is �50 N (continuous) and �200 N(peak). The rotations of the motors are measuredby optical encoders, which allow to estimate theposition of the handle with a spatial resolution�0.1 mm over the whole workspace. The controlarchitecture consists of an inner current loop
Table 1 Multiple sclerosis subjects
Disease Subject Age (y) Sex Hand duration (y) EDSS
P1 34 F R 4 0P2 32 F R 4 1P3 44 F R 3 0P4 38 F R 15 1P5 45 F R 5 0P6 27 F R 5 1P7 32 M R 3 0P8 37 F R 6 0P9 22 F R 7 1P10 32 M R 2 0P11 24 M R 1 0
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

332 M Casadio et al.
Multiple Sclerosis 2008; 14: 330–342 http://msj.sagepub.com
(16 kHz), and an outer impedance control loop(1 kHz).
We used a speed-dependent force field adapta-tion paradigm [5,7], which has been widely used tostudy sensorimotor co-ordination in unperturbedreaching movements and the adaptation inducedby an unfamiliar artificial dynamic environment.
Subjects were seated, with their torso and wristrestrained by means of suitable holders and graspedthe handle of the manipulandum with their domi-nant hand. A low-friction sled on the horizontalsurface of a table supported the forearm. The heightof the seat was adjusted so that the arm could bekept horizontally at the level of the shoulder.Therefore, only shoulder and elbow could moveand motion was restricted to the horizontal plane.The position of the seat was also adjusted in sucha way that, with the hand pointing at the centerof the workspace, the elbow joint was anteflexedabout 90° and the shoulder was horizontallyabducted about 45°.
The task consisted of reaching movements ineight different directions, starting from the samecenter position. The targets were presented on a 19�LCD computer screen, placed in front of the sub-jects, about 1 m away, at eye level. Targets were dis-played as green circles (diameter 2 cm) against ablack background. The current position of the handin the workspace was also continuously displayed,
as a yellow circle (diameter 0.4 cm). The nominalamplitude of the movements (distance of the tar-gets from the center position) was 10 cm.
The sequence of target presentations alternatedthe central target and one of four peripheral targets,(corresponding to the directions: 0°, 45°, 90° and135°) generated in random order. As in Diedrichsenet al. (2005) [15], the endpoint of each movementwas used as the starting point for the subsequentmovement. This corresponded to four additionaldirections (180°, 225°, 270° and 315°) for the returnmovements.
In order to decrease movement variability, thesubjects were encouraged to keep an approximatelyconstant timing. We set the desired duration to500 � 50 ms. If the estimated duration was insidethis range, a positive feedback/reward to the subject(a pleasant sound) was provided. If the measure wasbelow or above that range, no sound was generatedand the color of the target was changed to eitherwhite or red, respectively.
We also informed the subjects that reaction timewas not important – they could wait as long as theywanted after target appearance before starting eachmovement – but when ready, they had to performa single, rapid movement toward the target.
The experiment was organized into target sets,each consisting of a sequence of target presentationsin which each peripheral target occurred 12 times,for a total 12 � 4 � 48 center-out movements, plusthe corresponding 48 return movements.
The experimental protocol included threephases: (i) null field, in which the robot generatesno force (five target sets); (ii) force field, in whichthe force field was turned on (five target sets) and(iii) aftereffect (two target sets).
Each set lasted approximately 5 min, and thesubjects were allowed to rest between one set andanother. The null-field phase had the purpose ofestablishing a background level of performance.During the force field phase, the manipulandumgenerated a velocity-dependent curl field [7,16], inwhich perturbing forces were perpendicular to theinstantaneous movement direction and had a mag-nitude proportional to the hand speed. More specif-ically, the force applied to the hand was generated as:
(1)
where b is a viscous coefficient. We chose a value,b � 13 N/m/s, which is large enough to induce adetectable adaptation, but small enough to preventmuscle fatigue. This value corresponds to peakforces of 4–6 N at the hand. The hand velocity vec-tor, x, was calculated online from the rotations ofthe joints, measured by the encoders.
Figure 1 Experimental set-up, seen from above: it showsthe wrist holder, the arm weight support, the screen in frontof the subject and the four-bar linkage of the robot.
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

Abnormal sensorimotor control in MS subjects with no clinical disability 333
http://msj.sagepub.com Multiple Sclerosis 2008; 14: 330–342
During the field sets, we inserted ‘catch trials’,that is, trials in which the force field was unexpect-edly turned off. The probability of catch trials wasset to 1/6, corresponding to two catch trials perdirection per target set.
Movement analysis
Hand trajectories were sampled at 100 Hz andsmoothed by using a sixth order Savitzky–Golayfilter with a 170 ms window (cut-off frequency:~11 Hz), which was also used to estimate the subse-quent time derivatives of the trajectory. Movementonset was defined as the first time when speedexceeded 10% of the maximum speed. Likewise,end of movement was defined as the first timewhen speed fell below 10% of the maximum speed.This criterion allows to identify primary move-ments. Usually, late adjustments to the endpoint(secondary submovements) were not observed andare not analysed here.
For each movement, we estimated the timeelapsed between, respectively, movement onset andthe time when speed reaches a peak (accelerationduration) and between this same time and that ofmovement termination (deceleration duration).
We also estimated the aiming error as the differ-ence between the target direction and the actualmovement direction, estimated from the trajectoryin the early phase of the movement. The aimingerror was calculated within two different time inter-vals after movement onset: 100 ms and 300 ms.During the first 100 ms, movements can be assumedto be under open-loop control; therefore, we tookthe 100-ms aiming error as an indicator of the per-formance of the feed-forward component of con-trol. As regards the 300-ms aiming error, we took itas a measure of lateral deviation from the straightline (i.e., an indicator of path curvature) [7].
The 100-ms aiming error has a ‘systematic’ com-ponent, related to target direction, which reflectsthe inability to account for the anisotropy of arm(and robot) inertia and a random component,which reflects the variability of the performancefrom trial to trial. To distinguish among theseeffects, for each movement direction we took boththe average aiming error (the ‘systematic’ compo-nent) and its SD (the ‘random’ component).
To test for changes in motor performance duringthe null-field phase, we carried out a three-wayrepeated-measures ANOVA, by looking at theeffects of disease (control versus MS), movementdirection (0°, 45°, 90°, 135°, 180°, 225°, 270°, 315°)and time (early versus late target set of the nullphase) for all the above quantities, namely 100-msaiming error (systematic and random part), acceler-ation and deceleration duration and 300-ms aiming
error. To test whether the differences (if any)among control and MS subjects were still present atthe end of familiarization, we looked (contrastanalysis) at the effect of disease at the end of famili-arization (late target set).
Finally, to assess the adaptation capability of MSsubjects in comparison to controls, the same analy-sis was repeated for the force field phase.
Analysis of force field adaptation
To test for adaptation, during each phase of theexperiment we looked at the time course of eachindicator. In particular, for each phase we looked atthe changes of each indicator between the first andthe last target set.
Subjects may react to the perturbations intro-duced by the curl field in different ways: (1) theymay just ignore them, and accept the distortionintroduced by the field, a viable alternative as thiskind of perturbation does not specifically affect theachievement of the target; (2) they may resist theperturbation by increasing their hand stiffness (e.g.,by co-contracting the muscles), without changingthe underlying motor commands and (3) they maycompensate the perturbation by predicting it,through the formation of a suitable internal model.The latter option implies that subjects modify theirmotor commands.
The first alternative can be ruled out if we candemonstrate that the movements made under thecurl field do change over repetitions, a finding thatis well established for healthy subjects [5], butneeds to be demonstrated for MS patients. The sec-ond alternative can be ruled out by observing thecatch trials. If the observed adaptation to the forcefield were due to a mere increase in hand stiffness,catch trials should display no deviation. In con-trast, if error reduction is a consequence of anadapted feed-forward control, the unexpected dis-appearance of the field in the catch trials shouldresult in errors in the opposite direction. In thisstudy, we used the following definition of a learn-ing index (LI), originally proposed by Criscimagna-Hemminger et al. (2003) [16] and used by Smithet al. (2005) [7]:
(2)
where yf and yc are the 300-ms aiming errors in thefield trials and in the catch trials, respectively.Learning index is zero (no learning) if the error incatch trials is zero; it is one if the error in field tri-als is zero. Aiming error measures were adjusted forany bias that may have been present during the last
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

334 M Casadio et al.
Multiple Sclerosis 2008; 14: 330–342 http://msj.sagepub.com
null-field set. Therefore, errors in a field set alwaysrefer to variations with respect to the errors in thelate null set.
To test for differences in the LI of MS subjectsand controls, we focused on the late value of the LI,and ran a one-way ANOVA with disease (controlversus MS) as factor.
State–space model of adaptation
Recently, it has been suggested [17] that sensorimo-tor adaptation is better characterized in terms of itstemporal evolution rather than just in terms of itsoutcome. A recent theory [18–20] describes how, inforce field adaptation, the effect of errors experi-enced in a movement in one specific direction canbe related to the previous history of experiencedmovements.
Let us denote the error on trial n as y(n). Thismay be any measure of movement inaccuracy, forinstance the 300-ms aiming error. We may expressthis quantity as a function of the force perturba-tion, f(n), and of an internally stored ‘motor com-mand’ that – for that direction – would compensatethe perturbation, zd(n), where d is the movementdirection in that trial. The observed temporal evo-lution of y(n) can be modeled by a simple linearstate–space model:
(3)
The parameter Dd indicates how much angularerror results from a perturbation in direction d ina naive participant (i.e., with zd(n) � 0). This para-meter may be related to the hand compliance (theinverse of stiffness), and corresponds to the contri-bution of co-contraction to the response to forcefield perturbation. The quantity r(n) is an outputor ‘execution’ noise term, which accounts for theportion of the observed movement error that is notpredicted by the model.
The state equation (Equation 3, top) states thatat each trial the experienced error, y(n), modifiesthe internal state z(n) – that is, the internallystored ‘motor command’ for each movement direc-tion – by an amount that is determined by the rateof adaptation, B. The vector quantity q(n) is astate–space or ‘adaptation’ noise term, whichaccounts for the portion of state update that doesnot depend on the previous error. Note that the stateequation (Equation 3, top) is an eight-dimensionalvector equation (because there are eight movementdirections) whereas the output equation (Equation 3,bottom) is scalar.
On every trial, the state vector is updated for allpossible movement directions – in other words,movements in one particular direction contributeto the adaptation of the internal model in alldirections – but only the state component that cor-responds to that particular movement directioncontributes to the error at that trial. Therefore, B isan 8 � 8 matrix, where the element bij denotes thecontribution of experiencing an error in direction jon the internally stored motor command for direc-tion i. Each column of B represents the error gener-alization function, that is, how an error in onedirection contributes to adaptation of the internalmodel in all directions. Further details about themodel are reported in Appendix A.
To test for changes in MS subjects and control,for each model parameter we ran a one-wayANOVA with disease (control/MS) as factor.
Results
Figure 2 shows typical hand trajectories recordedduring the adaptation process (early and latephases, respectively), with respect to the differentexperimental conditions (null field, force field andafter-effect). The figure suggests that the overallmotor and adaptation patterns are similar in con-trol and MS subjects, but there might be detectableperformance differences.
Multiple sclerosis subjects display subtle butdetectable abnormalities
To assess whether and how the motor performanceof MS subjects differs from that of controls, we ini-tially focused on the null-field trials (N trials), whensubjects had to familiarize with the robotic device,but the latter generated no forces.
The early portion of the movements can be char-acterized in terms of the spatial distribution of thepeak hand acceleration, for the different targetdirections. Previous studies with normal subjects[21] reported a systematic directional variation ofpeak acceleration, possibly due to an inaccurateaccount of the anisotropy of arm inertia. In ourexperiment, we found qualitatively similar varia-tions in peak acceleration in MS subjects and con-trols. In addition, MS subjects displayed a muchgreater trial-by-trial variability; see Figure 3A.
To quantify these differences, we initiallyfocused on the 100-ms aiming error (both system-atic and random components), which is largelydetermined by peak acceleration. As regards theeffect of disease, we found that in MS subjects boththe systematic and, even more, the random compo-nent are significantly greater than in controls;
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

Abnormal sensorimotor control in MS subjects with no clinical disability 335
http://msj.sagepub.com Multiple Sclerosis 2008; 14: 330–342
respectively, F(1,20) � 19.441, P � 0.00027 andF(1,20) � 29.099, P � 0.00003. In other words, inMS subjects the directional errors are greater andmore variable than in controls.
We also found that in MS subjects the accelera-tion and, even more, the deceleration portionsof the movements last significantly longer thanin controls [F(1,20) � 8.1135, P � 0.00993 andF(1,20) � 16.090, P � 0.00069, respectively]. Pathcurvature (i.e., the 300-ms aiming error) too is sig-nificantly greater in MS subjects [F(1, 20) � 13.429,P � 0.00154].
In summary, the movements of MS subjects thattend to start in the wrong direction are slower,more curved and display more variability.
We also analysed the effect of time to assesswhether subjects modified their motor perform-ance while getting familiar with the task and theexperimental conditions – for example, the inertialand friction forces of the manipulandum. Wefound that, over time, movements significantlydecrease the 100-ms aiming error – both systematic[F(1,20) � 13.818, P � 0.0014) and random[F(1,20) � 6.6194, P � 0.01816], components; thedurations of the acceleration [F(1,20) � 35.436,P � 0.00001) and of the deceleration phases[F(1,20) � 46.280, P � 0.00001], and the lateraldeviation [F(1,20) � 54.663, P � 0.0001].
We then looked at the disease � time interac-tion. A significant interaction would be the evi-dence that the rate of familiarization differs in MSsubjects and controls. We found no significantinteraction for the 100-ms aiming error (systematicpart) and for acceleration duration. In contrast, MSsubjects displayed a greater decrease in aimingerror variability [F(1,20) � 5.8762, P � 0.02495]and in the duration of the deceleration phase [F(1, 20) � 10.528, P � 0.00406); see Figure 3C.
Figure 3 Subjects display directional errors in the early partof the movement. (A) Spatial distribution of peak accelera-tion in XY space, for two typical control (top) and MSsubjects (bottom). Scale bar: 2 m/s2; (B) Directional depend-ence of the systematic and random component of the 100-ms aiming error, for Control (gray) and MS (black) subjects;Thin and thick lines indicate, respectively, early and latefamiliarization (null field) phase. Scales: � 25 deg and (C)Time change of the systematic (left) and random (right)components of the 100-ms aiming error, during the familiar-ization phase. Error bars denote SE.Figure 2 Typical trajectories toward the eight targets
(center-target distance � 10 cm) in the different phases ofthe experiment. From Top to Bottom: null field (early andlate trials); force field (early and late trials) and early aftereffects. Gray traces denote presence of forces; black tracesindicate no forces. Black traces in the force field phasedenote the catch trials. Control subject (left column); MSsubjects (right column). For clarity of display, trajectorieshave been shifted to originate from the same central position.
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

336 M Casadio et al.
Multiple Sclerosis 2008; 14: 330–342 http://msj.sagepub.com
We also looked at the differences between con-trol and MS subjects at the end of familarization(late target set of the null phase). We found that MSsubjects still have a greater 100-ms aiming error(both systematic – P � 0.011 – and random compo-nents – P � 0.0023) than controls. Moreover, theirmovements are still more curved (P � 0.039) andhave a prolonged deceleration phase (P � 0.004)whereas the durations of the acceleration phase areno longer different.
This means that, while all subjects graduallyimprove their performance, MS subjects display agreater improvement in trajectory repeatability anda greater decrease in the duration of the decelera-tion portion of the movement.
As regards the effect of direction, all subjects dis-play a significant direction dependence in the 100-msaiming error – both systematic [F(7,140) � 25.169,P � 0.00001] and random parts [F(7,140) � 43.481,P � 0.00001], which is consistent with that of thepeak acceleration; see Figures 3A and 3B. A similardependence was found for acceleration duration[F(7,140) � 58.304, P � 0.00001) and, to a lesserextent, deceleration duration [F(7,140) � 12.354,P � 0.00001). For no indicator direction dependencewas not found to be significantly affected by disease.
In addition, in their movements MS subjectsdisplay a greater variability. Moreover, even thoughMS subjects display a greater improvement, at theend of the familiarization phase there is still adiscrepancy in the performance of MS subjects andcontrols (see Figure 3C).
Multiple sclerosis subjects have an intact ability to adapt to force fields
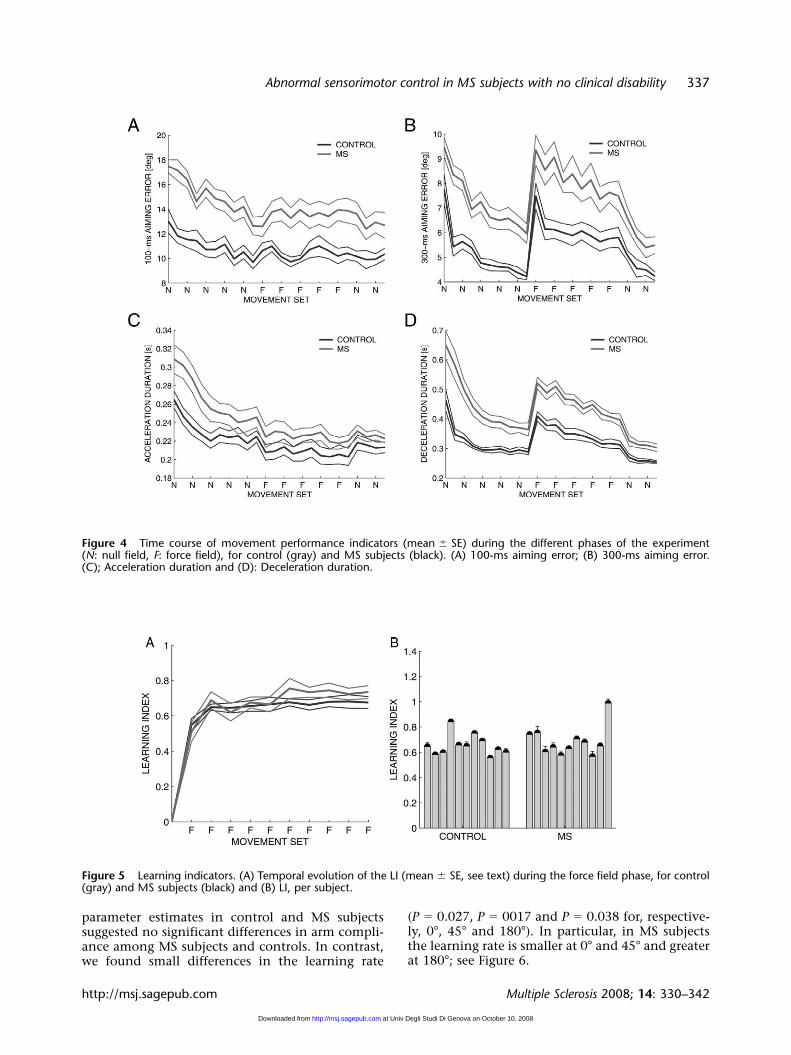
The qualitative observation of the trajectories inthe different phases of the experiment (see Figure 2)suggests that MS subjects are still capable of adapt-ing to this unfamiliar dynamic environment. Alook at the time course of the performance indica-tors suggests that early exposition to the force fieldinduces a sudden increase in deceleration durationand lateral deviation. However, these changes aregradually compensated during the force fieldphase.
When the force field is turned off, these sameindicators undergo a second abrupt change in theopposite direction, which gradually vanishes dur-ing the last (wash-out) phase. They eventuallyrecover the magnitudes observed at the end of thebaseline phase; see Figure 4.
A very different behavior is observed in the 100-ms aiming error and in acceleration duration,which both decrease monotonically along all thephases of the experiment. In the initial portion ofthe movement, hand speed is small, and therefore
the speed-dependent force field has little effect onthe 100-ms aiming error. However, its monotonicdecrease over the whole experiment suggests thatsubjects keep adapting to the unfamiliar dynamicsof the robot arm over the whole experiment, irrespec-tive of the presence/absence of the force field.
Statistical analysis of the adaptation capability ofMS subjects in comparison to controls resulted in sig-nificant effects of disease for the 100-ms aiming error[F(1,20) � 8.0521, P � 0.010173], the 300-ms aimingerror [F(1,20) � 7.9221, P � 0.01071], the decelera-tion duration [F(1,20) � 16.211, P � 0.000661], butnot in the acceleration duration.
The effect of disease in the field trials confirmsthe findings related to the null-field phase: MS sub-jects tend to make greater directional errors, andtheir movements have a prolonged decelerationphase.
Moreover, we found a significant effect of time fordeceleration duration [F(1,20) � 67.839, P � 0.0001)and the 300-ms aiming error [F(1,20) � 11.128,P � 0.00329], but not in the 100-ms aiming errorand in the acceleration duration. In no indicatorswe found significant interactions between diseaseand time.
This means that both MS and control subjectseffectively adapt to the force field. The effect of theforce field does not affect the early phase of themovement much, when speed is lower. This is whythere is no adaptation in the 100-ms aiming errorand in the acceleration duration. Moreover, thelack of significant interaction between disease andtime suggests that there are no differences inMS subjects and controls in the amount of theiradaptation.
In other words, in spite of the differences inmotor performance, MS subjects and controls donot differ significantly in their ability to adapt andin their rate of adaptation. This is also evident fromFigure 4. This result is further confirmed whenlooking at the rate of adaptation for each individ-ual subject measured by the LI defined in Equation2. Statistical analysis shows that there is no signifi-cant difference in comparing the two populationsof subjects. Figure 5 confirms in graphical termsthat the temporal evolution of the 300-ms aimingerror in the field trials and catch trials, as well asthat of the LI, is almost the same in MS subjects andcontrols.
Trial-by-trial adaptation
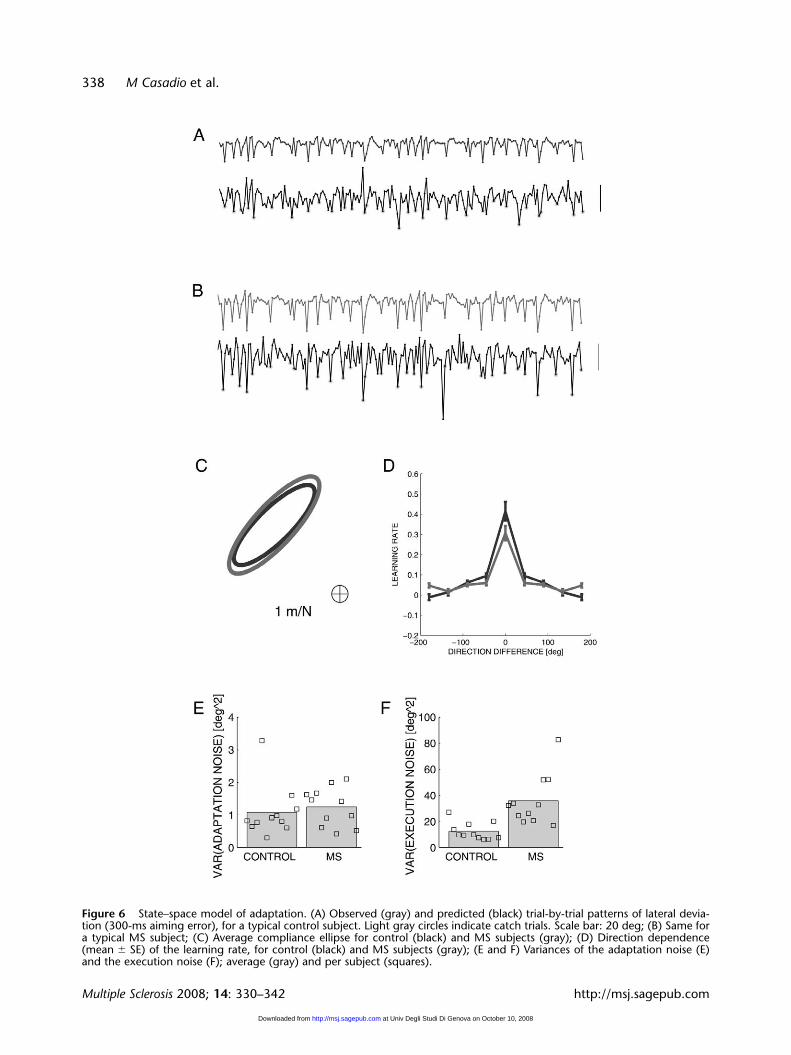
The state–space model provides additional informa-tion on the way adaptation evolves in time. Foreach subject, we estimated the model parameters(compliance, learning rate, variance of the adapta-tion and execution noise). A comparison of the
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from
Abnormal sensorimotor control in MS subjects with no clinical disability 337
http://msj.sagepub.com Multiple Sclerosis 2008; 14: 330–342
Figure 5 Learning indicators. (A) Temporal evolution of the LI (mean � SE, see text) during the force field phase, for control(gray) and MS subjects (black) and (B) LI, per subject.
Figure 4 Time course of movement performance indicators (mean � SE) during the different phases of the experiment (N: null field, F: force field), for control (gray) and MS subjects (black). (A) 100-ms aiming error; (B) 300-ms aiming error. (C); Acceleration duration and (D): Deceleration duration.
parameter estimates in control and MS subjectssuggested no significant differences in arm compli-ance among MS subjects and controls. In contrast,we found small differences in the learning rate
(P � 0.027, P � 0017 and P � 0.038 for, respective-ly, 0°, 45° and 180°). In particular, in MS subjectsthe learning rate is smaller at 0° and 45° and greaterat 180°; see Figure 6.
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from
338 M Casadio et al.
Multiple Sclerosis 2008; 14: 330–342 http://msj.sagepub.com
Figure 6 State–space model of adaptation. (A) Observed (gray) and predicted (black) trial-by-trial patterns of lateral devia-tion (300-ms aiming error), for a typical control subject. Light gray circles indicate catch trials. Scale bar: 20 deg; (B) Same fora typical MS subject; (C) Average compliance ellipse for control (black) and MS subjects (gray); (D) Direction dependence(mean � SE) of the learning rate, for control (black) and MS subjects (gray); (E and F) Variances of the adaptation noise (E)and the execution noise (F); average (gray) and per subject (squares).
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

Abnormal sensorimotor control in MS subjects with no clinical disability 339
http://msj.sagepub.com Multiple Sclerosis 2008; 14: 330–342
With regards to noise variances, we found no sig-nificant differences in the adaptation noise. Thismeans that the state–space model explains thedynamics of adaptation equally well in MS subjectsand controls. In contrast, we found that MSsubjects display a significantly greater (P � 0.0006)execution noise. This means that sometimes theadaptive response to force fields may be inappropri-ate, and this occurs more often in MS subjects thanin control subjects.
Discussion
We used a robot manipulandum to assess sensori-motor performance and adaptation capability inMS subjects with no disability – for these subjects,functional brain imaging studies suggest evidenceof cortical and subcortical reorganization. Whenthe robot generated no forces, the movements ofMS subjects were more curved, displayed greaterand more variable directional errors and a longerdeceleration phase. During the force field phase,both MS and control subjects gradually learned topredict the robot-generated forces. The rates ofadaptation were similar, but MS subjects showed agreater variability in responding to the force field.
Descending commands are abnormal in MS subjects
Analysis of the motor performance during the null-field trials shows that MS subjects with no disabilitymake greater errors than controls. More specifically,their movements start in the wrong direction. Also,they have a prolonged deceleration phase and aremore curved. This is consistent with previous find-ings [9]. In addition, here we report that MS sub-jects display a greater variability in the direction ofstart of their movements.
The peculiar dependence on direction displayedby the 100-ms aiming error and the accelerationduration may reflect the fact that both MS andcontrol subjects inaccurately account for theanisotropy of the inertia of the arm and the robotsystem. This finding is well established in normalsubjects [21]; MS subjects display a similar pattern,but with a greater error magnitude. In addition,these subjects generate motor commands that arenot only systematically inappropriate, but alsomore variable from trial to trial. Nevertheless, sucherrors do not prevent subjects from reaching thetargets. This is likely because these errors arecompensated, at least in part, by making on-linecorrections, based on the available visual and/orproprioceptive information. This may explain thegreater duration of the deceleration phase.
Can the above abnormalities be due to degrada-tion of performance caused by, for instance, musclefatigue? If this were the case, these effects wouldhave been even greater at the end of the force fieldadaptation phase because under this condition sub-jects are required to generate much greater muscleforces than those needed in the null-field condi-tion. However, this was not the case; rather, theobserved abnormalities point to defective control.
We also found that in both MS subjects and con-trols motor performance improves across repeti-tions. Multiple sclerosis subjects even display agreater improvement in aiming error variabilityand in deceleration duration. This suggests that MSsubjects and controls are equally good at familiariz-ing with the dynamics of the manipulandum, andthat the performance of MS subjects may benefitfrom exercise. However, the latter never achieve theperformance of controls.
These results are also consistent with similarfindings of a deteriorated balance and gait functionin MS subjects with no disability [8].
Multiple sclerosis subjects preserve their ability toadapt to unfamiliar dynamic environments
In spite of these subtle abnormalities in motor co-ordination, we found that the ability to adapt to anunfamiliar dynamic environment is substantiallypreserved in MS subjects. In particular, MS subjectsand controls display similar levels of arm compli-ance, and very little differences in the rates of adap-tation. Therefore, MS subjects effectively learn topredict the force field generated by the robot ratherthan just trying to resist to perturbations by stiffen-ing their arm.
The most significant difference we foundbetween MS subjects and controls is in the extentto which the observed trial-by-trial motor errorsduring force field adaptation are predicted by thestate–space model. Variability in the process offorce field adaptation may come from at least twodifferent sources [15,17]. It may be due to an inabil-ity to learn how to predict the current perturbation(adaptation noise), to an inability to effectively usesuch prediction (execution noise), or both. Wefound that the variance of adaptation noise doesnot differ significantly from controls, whereas thevariance of execution noise is significantly greater.This latter finding suggests that MS subjects arecapable of acquiring an accurate internal represen-tation of the unfamiliar dynamics (curl field), butthey make more errors when actually using suchinternal representation during movement execu-tion. As a consequence, in MS subjects motor com-mands are inappropriate more often than incontrols, and therefore require an extra amount of
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

340 M Casadio et al.
Multiple Sclerosis 2008; 14: 330–342 http://msj.sagepub.com
corrections, which are performed by appropriatelyexploiting the available sensory information. Sucha difficulty in movement execution is confirmed bythe finding of a greater variability observed in the100-ms aiming error. Movement execution prob-lems may also be related to the lack of attentionobserved in MS subjects [22].
The experiment presented here allows to dis-criminate between evaluation of motor perform-ance and assessment of adaptation capability (i.e.,the ability to predict the consequences of ouractions). Our results suggest that MS subjects withno disability have problems with movement execu-tion but not with motor adaptation.
Nevertheless, adaptation capability may beaffected in subjects with a more severe impair-ment. Recently, Leocani et al. (2007) [10] reportedthat MS subjects with a more severe impairment(EDSS � 5) display a greater difficulty than con-trols in tracking a moving target, and also displaya smaller improvement with time of their trackingperformance.
Preserved adaptation capability and brain reorganization
We demonstrated that in MS subjects with no dis-ability, in some cases after a long disease duration,the ability to learn new movements is similar tothat of healthy controls. This suggests a preservedpotential for the brain to reorganize. There is anincreasing evidence that cortical reorganizationcontributes to maintain normal levels of functionafter a relapse. Such reorganization would haveimportant implications for the development offuture treatments, aimed at promoting neuronalplasticity and at improving the clinical outcome ofthe disease.
MRI techniques allow to assess the anatomicalneuronal damage and its consequences on corticaladaptation, but provide little information on thecorresponding motor performance and on theresidual ability to adapt. Functional MRI tech-niques have been extensively used to study corticalreorganization in MS subjects. In an fMRI study,Rocca et al. (2005) [2] showed that a repetitivefinger flexion-extension task resulted in the recruit-ment of different brain areas in MS subjects atvarious stages of the disease. With respect to healthycontrols, nondisabled subjects with relapsing–remitting MS (a class of subjects that corresponds tothose tested in the present study) displayedincreased activations in various cortical areas.Moreover, a comparison of performance and fMRIactivations in simple and complex motor tasks [23]suggested that in early-stage MS subjects, fingerflexion-extension leads to the recruitment of brain
areas that in healthy subjects are associated withmore complex tasks (e.g., object manipulation).
Other studies have addressed the contribution ofcortical adaptation to visual recovery after opticneuritis, showing a clear time course: a low fMRIresponse during the acute phase is followed by anincrease in the volume of the activated regions,which suggests a transient adaptive change. Twostudies [24,25] have demonstrated that visual func-tion at baseline was inversely related to optic nervedamage and directly related to the extent ofresponse on fMRI, thus suggesting a dynamic spa-tiotemporal cortical change. These results supportthe notion that if a neural pathway is affected byMS lesions, the corresponding cortical areas aremore largely activated in order to maintain thefinalized activity.
Taken together, these results suggest that in earlyMS a substantial cortical reorganization takes place,possibly triggered by widespread tissue damage.Such reorganization might contribute to maintain-ing normal levels of function. Moreover, such areorganization involves areas that are related to theprocessing of sensorimotor information [23,26].
This is consistent with our findings that MSsubjects achieve a close-to-normal motor functionby performing a greater proportion of microadjust-ments (as witnessed by the greater curvature andduration of the deceleration phase), to compensatefor the partly incorrect descending commands (aswitnessed by the greater aiming errors).
It should be noted that the brain areas whichdisplay extra activation in MS subjects with no dis-ability [2] are consistent with those that were foundto correlate with the processing of motor errors innormal subjects [15], during a task similar to thatdescribed here. This indirectly supports the notionthat extra activation may reflect the need for extracorrections, in the sense that corrections would cor-relate with extra activation in the brain areas relat-ed to error and/or sensory processing, an/or ingreater activation in the areas related to motorcommands.
Fatigue
Fatigue is a major concern with MS subjects. In thedesign of the experimental protocol, we carefullytook this problem into account. Subjects wereallowed to rest between consecutive blocks of trials.However, only one MS subject did and this sup-ports the fact that the task was not fatiguing and infact was well tolerated. Moreover, Figure 4 and thecorresponding statistical analysis shows that therewas no degradation of performance at the end ofthe adaptation phase as compared to the final por-tion of the null phase. This, again, suggests that
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

Abnormal sensorimotor control in MS subjects with no clinical disability 341
http://msj.sagepub.com Multiple Sclerosis 2008; 14: 330–342
subjects displayed no problems in performing thetask in the late phase of the experiment. Indeed, asmall improvement was often observed.
Implications for robot-assisted rehabilitation
Overall, these results suggest that sensorimotoradaptation protocols may represent a useful tool tomeasure the motor performance after the adapta-tion, which follows brain lesions.
Robotic devices are also a promising tool for neu-romotor rehabilitation. They may be used to guideor assist the movements of a patient and in closeinteraction with him, just like a human physicaltherapist [27]. A prerequisite for the success of bothhuman and robot therapy is that patients preservetheir ability to adapt to novel dynamic environ-ments, an ability related to the feed-forward com-ponent of control. Robot therapy would be an idealtool for implementing this strategy, also taking intoaccount that it is possible to integrate in the robottraining sessions motor evaluation primitives thatare essential for monitoring the disease and finelytune the interaction parameters that characterizethe training sessions.
Acknowledgements
This work is partly supported by the ItalianMultiple Sclerosis Foundation (FISM) and by theItalian Ministry of Education, University andResearch (MIUR).
Appendix A. State–space computationalmodel
The type of state–space computational model of thetrial-to-trial changes in motor output described inthe Methods section was previously used to charac-terize the adaptation to curl fields in healthysubjects [18–20], and in subjects with cerebellaratrophy or Huntington’s disease [7].
Our model is similar to the model used in [7],with the only addition of the output andstate–space noise terms. To reduce the number ofparameters, we assumed that arm compliance in thehorizontal plane is described by a symmetric 2 � 2matrix [28]. Since the eight-dimensional compli-ance vector D is completely determined by armcompliance, we constrained D to three free parame-ters. Furthermore, we assumed that the error gener-alization functions (columns of B) are essentiallysymmetric (i.e., the amount of generalization to�45° is essentially the same as generalization to�45°, see [7,18]. Therefore we enforced symmetryon B, which constrained it to five free parameters.
Because the force field was kept constant in alltrials in which it was applied, but the actual forceexperienced varied depending on the movementvelocity, following [20] the force input to themodel, f(n), was represented as a discrete scalar(force field magnitude), and was modeled with val-ues of 0 or 1, corresponding to catch or field trials,respectively. Thus, the compliance computed fromthis model has units of angular error and can beinterpreted as the aiming error directly induced bythe force field. However, this quantity is intrinsical-ly related to the hand compliance matrix, and therelationship between the two is a simple multiplica-tive constant – T/b, where T � 300 ms is the timeafter movement onset, and b � 13 Nm/s is the fieldviscosity, see [7] for more details. We used this con-version to report the compliance ellipses in Figure 6.The noise terms were assumed to be Gaussian andwith zero mean, that is, r(n) ~ N(0,R) andq(n) ~ N(0,Q), and we assumed R � R
2 and Q � IQ
2, which adds another two parameters to themodel (i.e., R
2 and Q2). We used a nonlinear opti-
mization procedure to find B, D, R and Q that max-imize, for every subject, the log-likelihood of theobserved aiming error sequence [17,29].
References1. Lublin FD, Baier M, Cutter G. Effect of relapses on
development of residual deficit in multiple sclerosis.Neurology 2003; 61: 1528–32.
2. Rocca MA, Colombo B, Falini A, Ghezzi A,Martinelli V, Scotti G et al. Cortical adaptation inpatients with MS: a cross-sectional functional MRI studyof disease phenotypes. Lancet Neurol 2005; 4: 618–26.
3. Wang J, Hier DB. Motor reorganization in multiplesclerosis. Neurol Res 2007; 29: 3–8.
4. Filippi M, Rocca MA. MRI evidence for multiple scle-rosis as a diffuse disease of the central nervous system. J Neurol 2005; 252(Suppl 5): v16–24.
5. Shadmehr R, Mussa-Ivaldi FA. Adaptive representa-tion of dynamics during learning of a motor task. JNeurosci 1994; 14: 3208–24.
6. Maschke M, Gomez CM, Ebner TJ, Konczak J.Hereditary cerebellar ataxia progressively impairs forceadaptation during goal-directed arm movements. J Neurophysiol 2004; 91: 230–8.
7. Smith MA, Shadmehr R. Intact ability to learn inter-nal models of arm dynamics in Huntington’s disease butnot cerebellar degeneration. J Neurophysiol 2005; 93:2809–21.
8. Martin CL, Phillips BA, Kilpatrick TJ,Butzkueven H, Tubridy N, McDonald E, Galea MP.Gait and balance impairment in early multiple sclerosisin the absence of clinical disability. Mult Scler 2006; 12:620–8.
9. Solaro C, Brichetto G, Casadio M, Roccatagliata L,Ruggiu P, Mancardi GL et al. Subtle upper limbimpairment in asymptomatic multiple sclerosis subjects.Mult Scler 2007; 13: 428–32.
10. Leocani L, Comi E, Annovazzi P, Rovaris M,Rossi P, Cursi M et al. Impaired short-term motorlearning in multiple sclerosis: Evidence from virtualreality. Neurorehabil Neural Repair 2007; 21: 273–8.
11. Poser CM, Paty DW, Scheinberg L, McDonald WI,Davis FA, Ebers GC et al. New diagnostic criteria for
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from

342 M Casadio et al.
Multiple Sclerosis 2008; 14: 330–342 http://msj.sagepub.com
multiple sclerosis: guidelines for research protocols. AnnNeurol 1983; 13: 227–31.
12. Kurtzke JF. Rating neurologic impairment in multiplesclerosis: an expanded disability status scale (EDSS).Neurology 1983; 33: 1444–52.
13. Sipe JC et al. A neurologic rating scale (NRS) for use inmultiple sclerosis. Neurology 1984; 34: 1368–72.
14. Casadio M, Morasso PG, Sanguineti V, Arrichiello V.Braccio di Ferro: a new haptic workstation for neuromo-tor rehabilitation. Technol Health Care 2006; 13: 1–20.
15. Diedrichsen J, Hashambhoy Y, Rane T, Shadmehr R.Neural correlates of reach errors. J Neurosci 2005; 25:9919–31.
16. Criscimagna-Hemminger SE, Donchin O,Gazzaniga MS, Shadmehr R. Learned dynamics ofreaching movements generalize from dominant to non-dominant arm. J Neurophysiol 2003; 89: 168–76.
17. Cheng S, Sabes PN. Modeling sensorimotor learningwith linear dynamical systems. Neural Comput 2006; 18:760–93.
18. Donchin O, Francis JT, Shadmehr R. Quantifyinggeneralization from trial-by-trial behavior of adaptivesystems that learn with basis functions: theory andexperiments in human motor control. J Neurosci 2003;23: 9032–45.
19. Scheidt RA, Dingwell JB, Mussa-Ivaldi FA.Learning to move amid uncertainty. J Neurophysiol 2001;86: 971–85.
20. Thoroughman KA, Shadmehr R. Learning of actionthrough adaptive combination of motor primitives.Nature 2000; 407: 742–7.
21. Gordon J, Ghilardi MF, Cooper SE, Ghez C.Accuracy of planar reaching movements. II. Systematicextent errors resulting from inertial anisotropy. Exp BrainRes 1994; 99: 112–30.
22. Mainero C, Pantano P, Caramia F, Pozzilli C. Brainreorganization during attention and memory tasks inmultiple sclerosis: insights from functional MRI studies.J Neurol Sci 2006; 245: 93–8.
23. Filippi M, Rocca MA, Mezzapesa DM, Ghezzi A,Falini A, Martinelli V et al. Simple and complex move-ment-associated functional MRI changes in patients atpresentation with clinically isolated syndromes sugges-tive of multiple sclerosis. Hum Brain Mapp 2004; 21:108–17.
24. Langkilde AR, Frederiksen JL, Rostrup E,Larsson HB. Functional MRI of the visual cortex andvisual testing in patients with previous optic neuritis.Eur J Neurol 2002; 9: 277–86.
25. Toosy AT, Werring DJ, Bullmore ET, Plant GT,Barker GJ, Miller DH et al. Functional magnetic reso-nance imaging of the cortical response to photic stimu-lation in humans following optic neuritis recovery.Neurosci Lett 2002; 330: 255–9.
26. Filippi M, Rocca MA, Mezzapesa DM, Falini A,Colombo B, Scotti G et al. A functional MRI study ofcortical activations associated with object manipula-tion in patients with MS. Neuroimage 2004; 21:1147–54.
27. Prange GB, Jannink MJ, Groothuis-Oudshoorn CG,Hermens HJ, Ijzerman MJ. Systematic review of theeffect of robot-aided therapy on recovery of the hemi-paretic arm after stroke. J Rehabil Res Dev 2006; 43:171–84.
28. Mussa-Ivaldi FA, Hogan N, Bizzi E. Neural, mechan-ical and geometric factors subserving arm posture inhumans. J Neurosci 1985; 5: 2732–43.
29. Ghahramani Z, Hinton GE. Parameter estimation forlinear dynamical systems. University of Toronto, Toronto,1996.
at Univ Degli Studi Di Genova on October 10, 2008 http://msj.sagepub.comDownloaded from




















