
Morphological Transitions Governed by Density Dependence and Lipoxygenase Activity in Aspergillus...
12
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2008, p. 5674–5685 Vol. 74, No. 18 0099-2240/08/$08.000 doi:10.1128/AEM.00565-08 Copyright © 2008, American Society for Microbiology. All Rights Reserved. Morphological Transitions Governed by Density Dependence and Lipoxygenase Activity in Aspergillus flavus † S. Horowitz Brown, 1 R. Zarnowski, 2 W. C. Sharpee, 1 and N. P. Keller 1,2 * Department of Plant Pathology 1 and Department of Medical Microbiology and Immunology, 2 University of Wisconsin—Madison, 1550 Linden Dr., Madison, Wisconsin 53706-1598 Received 9 March 2008/Accepted 10 July 2008 Aspergillus flavus differentiates to produce asexual dispersing spores (conidia) or overwintering survival structures called sclerotia. Results described here show that these two processes are oppositely regulated by density-dependent mechanisms and that increasing the cell density (from 10 1 to 10 7 cells/plate) results in the lowest numbers of sclerotial and the highest numbers of conidial. Extract from spent medium of low-cell- density cultures induced a high-sclerotium-number phenotype, whereas high-cell-density extract increased conidiation. Density-dependent development is also modified by changes in lipid availability. Exogenous linoleic acid increased sclerotial production at intermediate cell densities (10 4 and 10 5 cells/plate), whereas oleic and linolenic acids inhibited sclerotium formation. Deletion of Aflox encoding a lipoxygenase (LOX) greatly diminished density-dependent development of both sclerotia and conidia, resulting in an overall increase in the number of sclerotia and a decrease in the number of conidia at high cell densities (>10 5 cells/plate). Aflox mutants showed decreased linoleic acid LOX activity. Taken together, these results suggest that there is a quorum-sensing mechanism in which a factor(s) produced in dense cultures, perhaps a LOX-derived metabolite, activates conidium formation, while a factor(s) produced in low-density cultures stimulates sclerotium formation. Aspergillus flavus is a cosmopolitan, soilborne, filamentous fungus that frequently infects oil-rich seeds of several crop species. Concerns about the association of this organism with plant products, particularly corn, cotton, peanuts, and tree nuts, center on its ability to produce aflatoxins, which are carcinogenic polyketide secondary metabolites (3, 13). In ad- dition, A. flavus is second only to Aspergillus fumigatus as a cause of human invasive aspergillosis (22). Asexual spores (conidia) are the primary disseminating mechanism of A. flavus and serve as the major inoculum source. A. flavus also differentiates to produce sclerotia, which are compact masses of mycelia that overwinter and germinate to produce either additional hyphae or conidia (13). Sclerotia are hypothesized to be degenerate sexual structures and may represent a vestige of cleistothecium production (16). Sclerotia allow A. flavus to survive under harsh conditions and to out- compete other organisms for substrates in the soil or in plants (13). Infected plant tissue, such as corn kernels, cobs, and leaf tissue, can remain in the soil and support the fungus until the following season, when newly exposed mycelium or sclerotia can give rise to conidial structures, thus producing the primary inoculum for the next infection cycle (13). In the genetic model organism Aspergillus nidulans, which does not produce sclerotia but does produce cleistothecia, the ratio of cleistothecial production (and production of asco- spores) to conidial production is balanced by oxylipin availabil- ity. Oxylipins represent a vast and diverse family of secondary metabolites that originate from the oxidation or further con- version of unsaturated fatty acids. In A. nidulans endogenous oleic acid-, linoleic acid-, and likely linolenic acid-derived oxy- lipins, collectively called “psi factors” (precocious sexual in- ducer), influence the development of cleistothecia and conidia (6, 7, 8, 9, 51). The proportion of these different oxylipins has been postulated to regulate the ratio of asexual development to sexual development in A. nidulans. Such a “balancing” role for oxylipins was genetically supported by the results of dele- tion of the dioxygenases responsible for psi factor production. Loss of ppoB (psi factor-producing oxygenase B) yielded an increased-conidium, decreased-cleistothecium phenotype (52), whereas loss of both ppoA and ppoC resulted in the opposite phenotype (51). Additionally, exogenous applications of oxy- lipins resulted in phenotypes that mimicked the ppo mutant phenotypes in A. nidulans and also altered sclerotial and conid- ial production in A. flavus (7). Results of these studies sug- gested a possible quorum-driven, cell density-dependent phe- nomenon in morphological transitions in Aspergillus species (14, 50). Cell density-dependent regulatory networks in micro- organisms generally control processes that involve cell-cell in- teractions, such as group motility and the formation of multi- cellular structures leading to differentiation processes (1, 25). Most cell density studies have centered on quorum sensing in bacteria (25). The swarm motility of microorganisms such as Vibrio parahaemolyticus and Proteus mirabilis is a multicellular behavior dependent on cell density (1). Likewise, the marine luminescent bacterium Vibrio fischeri uses the LuxR and LuxI proteins for autoinduction of luminescence (15). However, cell density-dependent phenomena are not limited to prokaryotes. Dictyostelium discoideum, a simple eukaryote, exhibits quorum- * Corresponding author. Mailing address: 3476 Microbial Science Building, Department of Medical Microbiology and Immunology and Department of Plant Pathology, UW—Madison, 1550 Linden Dr., Mad- ison, WI 53706. Phone: (608) 262-9795. Fax: (608) 262-8418. E-mail: [email protected]. † Supplemental material for this article may be found at http://aem .asm.org/. Published ahead of print on 25 July 2008. 5674
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Morphological Transitions Governed by Density Dependence and Lipoxygenase Activity in Aspergillus...

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2008, p. 5674–5685 Vol. 74, No. 180099-2240/08/$08.00�0 doi:10.1128/AEM.00565-08Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Morphological Transitions Governed by Density Dependence andLipoxygenase Activity in Aspergillus flavus�†
S. Horowitz Brown,1 R. Zarnowski,2 W. C. Sharpee,1 and N. P. Keller1,2*Department of Plant Pathology1 and Department of Medical Microbiology and Immunology,2
University of Wisconsin—Madison, 1550 Linden Dr., Madison, Wisconsin 53706-1598
Received 9 March 2008/Accepted 10 July 2008
Aspergillus flavus differentiates to produce asexual dispersing spores (conidia) or overwintering survivalstructures called sclerotia. Results described here show that these two processes are oppositely regulated bydensity-dependent mechanisms and that increasing the cell density (from 101 to 107 cells/plate) results in thelowest numbers of sclerotial and the highest numbers of conidial. Extract from spent medium of low-cell-density cultures induced a high-sclerotium-number phenotype, whereas high-cell-density extract increasedconidiation. Density-dependent development is also modified by changes in lipid availability. Exogenouslinoleic acid increased sclerotial production at intermediate cell densities (104 and 105 cells/plate), whereasoleic and linolenic acids inhibited sclerotium formation. Deletion of Aflox encoding a lipoxygenase (LOX)greatly diminished density-dependent development of both sclerotia and conidia, resulting in an overallincrease in the number of sclerotia and a decrease in the number of conidia at high cell densities (>105
cells/plate). Aflox mutants showed decreased linoleic acid LOX activity. Taken together, these results suggestthat there is a quorum-sensing mechanism in which a factor(s) produced in dense cultures, perhaps aLOX-derived metabolite, activates conidium formation, while a factor(s) produced in low-density culturesstimulates sclerotium formation.
Aspergillus flavus is a cosmopolitan, soilborne, filamentousfungus that frequently infects oil-rich seeds of several cropspecies. Concerns about the association of this organism withplant products, particularly corn, cotton, peanuts, and treenuts, center on its ability to produce aflatoxins, which arecarcinogenic polyketide secondary metabolites (3, 13). In ad-dition, A. flavus is second only to Aspergillus fumigatus as acause of human invasive aspergillosis (22).
Asexual spores (conidia) are the primary disseminatingmechanism of A. flavus and serve as the major inoculumsource. A. flavus also differentiates to produce sclerotia, whichare compact masses of mycelia that overwinter and germinateto produce either additional hyphae or conidia (13). Sclerotiaare hypothesized to be degenerate sexual structures and mayrepresent a vestige of cleistothecium production (16). Sclerotiaallow A. flavus to survive under harsh conditions and to out-compete other organisms for substrates in the soil or in plants(13). Infected plant tissue, such as corn kernels, cobs, and leaftissue, can remain in the soil and support the fungus until thefollowing season, when newly exposed mycelium or sclerotiacan give rise to conidial structures, thus producing the primaryinoculum for the next infection cycle (13).
In the genetic model organism Aspergillus nidulans, whichdoes not produce sclerotia but does produce cleistothecia, theratio of cleistothecial production (and production of asco-
spores) to conidial production is balanced by oxylipin availabil-ity. Oxylipins represent a vast and diverse family of secondarymetabolites that originate from the oxidation or further con-version of unsaturated fatty acids. In A. nidulans endogenousoleic acid-, linoleic acid-, and likely linolenic acid-derived oxy-lipins, collectively called “psi factors” (precocious sexual in-ducer), influence the development of cleistothecia and conidia(6, 7, 8, 9, 51). The proportion of these different oxylipins hasbeen postulated to regulate the ratio of asexual developmentto sexual development in A. nidulans. Such a “balancing” rolefor oxylipins was genetically supported by the results of dele-tion of the dioxygenases responsible for psi factor production.Loss of ppoB (psi factor-producing oxygenase B) yielded anincreased-conidium, decreased-cleistothecium phenotype (52),whereas loss of both ppoA and ppoC resulted in the oppositephenotype (51). Additionally, exogenous applications of oxy-lipins resulted in phenotypes that mimicked the ppo mutantphenotypes in A. nidulans and also altered sclerotial and conid-ial production in A. flavus (7). Results of these studies sug-gested a possible quorum-driven, cell density-dependent phe-nomenon in morphological transitions in Aspergillus species(14, 50). Cell density-dependent regulatory networks in micro-organisms generally control processes that involve cell-cell in-teractions, such as group motility and the formation of multi-cellular structures leading to differentiation processes (1, 25).
Most cell density studies have centered on quorum sensingin bacteria (25). The swarm motility of microorganisms such asVibrio parahaemolyticus and Proteus mirabilis is a multicellularbehavior dependent on cell density (1). Likewise, the marineluminescent bacterium Vibrio fischeri uses the LuxR and LuxIproteins for autoinduction of luminescence (15). However, celldensity-dependent phenomena are not limited to prokaryotes.Dictyostelium discoideum, a simple eukaryote, exhibits quorum-
* Corresponding author. Mailing address: 3476 Microbial ScienceBuilding, Department of Medical Microbiology and Immunology andDepartment of Plant Pathology, UW—Madison, 1550 Linden Dr., Mad-ison, WI 53706. Phone: (608) 262-9795. Fax: (608) 262-8418. E-mail:[email protected].
† Supplemental material for this article may be found at http://aem.asm.org/.
� Published ahead of print on 25 July 2008.
5674

sensing behavior. When an adequate bacterial food source ispresent, Dictyostelium cells live as unicellular amoebae anddivide by fission. However, when food is scarce, Dictyosteliumcells enter a developmental cycle that begins with the aggre-gation of cells that behave as a multicellular organism (31).Much research has also focused on quorum sensing in thefungus Candida albicans. In this organism at least two mole-cules, including the oxygenated lipid farnesol, regulate quorumsensing, yeast-to-hypha transitions, and biofilm formation (11,38, 41). Density-dependent developmental transitions havebeen found in other fungi; for example, Ceratocystis ulmi growsas a yeast at high densities but as filaments at low densities(26). The switch can be attenuated by chemical inhibition oflipoxygenase (LOX) activity (28), indicating that oxylipins havea possible role in quorum sensing in C. ulmi.
Here we examined cell density transitions in A. flavus, fo-cusing on conidial and sclerotial development of the fungusgrown at different cell densities. We found that both of theseprocesses are regulated by density-dependent mechanisms andthat increasing the cell density (from 101 to 107 cells/plate)resulted in the lowest numbers of sclerotia, yet the highestnumbers of conidia. Extracts from spent medium of cultureswith low cell densities stimulated sclerotial formation, andextracts from spent medium of cultures with high cell densitiesstimulated conidial formation, suggesting that there is a quo-rum-sensing mechanism in A. flavus. Furthermore, these den-sity-dependent phenomena are influenced by lipid modifiers asboth exogenous fatty acid application and loss of a putativeoxylipin-generating LOX, encoded by Aflox, affect the conidium-to-sclerotium switch.
MATERIALS AND METHODS
Fungal strains and growth conditions. The A. flavus strains used and gener-ated in this study are shown in Table 1. All strains used for physiological studieswere prototrophic, and the strains were grown at 29°C and maintained on glucoseminimal medium (GMM) (47) unless otherwise indicated. Cultures were growneither in the dark inside cardboard boxes that provided continuous darkness orin the presence of continuous white light by using an incubator equipped with aGeneral Electric 15-W broad-spectrum fluorescent light bulb (F15T12CW) po-sitioned 50 cm from the agar surface.
Assay for conidial and sclerotial density dependence. For analysis of sclerotialdensity dependence, 10 ml of GMM with 1.6% agar and 2% sorbitol was overlaidwith 3 ml of GMM with 0.7% agar and 2% sorbitol containing 101 to 107 conidiaof an appropriate strain in a petri plate (60 by 15 mm). The plates were incubatedat 29°C under continuous dark conditions for 7 days. To accurately visualize thesclerotia, plates were sprayed thoroughly with 70% ethanol in water to removeconidia and aerial mycelia. Either the exposed sclerotia then were counted witha dissecting scope at a magnification of �4 or the sclerotia were collected andlyophilized and sclerotium production was determined gravimetrically (mg [dryweight] per plate). Conidia were counted on day 3 or 7. To accurately countconidia, three 1-cm plugs were homogenized in 3 ml of water containing 0.01%
Tween 80, diluted 1:10, and counted with a hemocytometer at a magnification of�400. For each conidial concentration, conidial and sclerotial counts were de-termined by using four replicates, and tests were repeated three times in order toexamine reproducibility. Spore suspensions were freshly prepared for each strainand replication.
Spent medium bioassay. For preparation of cell extracts from spent mediumof A. flavus wild-type strain NRRL 3357 cultures, 30 petri plates (100 by 15 mm),each containing 25 ml of GMM supplemented with 1.6% agar plus 2% sorbitol,were overlaid with 10 ml of GMM supplemented with 0.7% agar plus 2% sorbitolcontaining 103 or 107 conidia/plate. Thirty uninoculated agar plates were used asthe control. All 90 plates were incubated at 29°C under continuous dark condi-tions for 5 days, after which the cultures were extracted with ethyl acetate asfollows. For convenience, the contents of 10 plates were extracted at a time. Theagar and fungus in 10 plates were removed using a spatula, placed in a largeblender with 250 ml of sterile distilled H2O, and blended for 10 s. The maceratewas transferred into a 4-liter beaker, and the procedure described above wasrepeated for the remaining 20 plates, 10 at a time, for each treatment. The totalmacerate was placed into the fume hood, and 2 volumes of ethyl acetate wasadded. The macerate with the ethyl acetate was stirred with a magnetic stirrer for15 min, allowing the layers to separate. The organic layer was then removed usinga 100-ml glass pipette and placed into a 1-liter evaporating flask (Rotavapor-RE;Brinkmann, Switzerland). The dry extract was resuspended in a few milliliters ofethyl acetate and transferred into a glass vial. The vial was covered with foil (toprotect the contents from light), and the extract was again evaporated andresuspended in 2.5 ml ethanol, topped with nitrogen gas, and kept at �80°C untilit was used.
Extracts of spent medium were added to new cultures by spreading 100-�lportions of 1:5, 1:10, or 1:100 dilutions (in ethanol) from the 2.5-ml ethanolextracts from the plates containing 103 or 107 conidia/plate and medium extractsobtained as described above on petri plates (60 by 15 mm) containing 10 ml ofGMM supplemented with 1.6% agar plus 2% sorbitol. A water control was alsoincluded. The plates were allowed to dry for 15 min and then overlaid with 3 mlof GMM supplemented with 0.7% agar plus 2% sorbitol containing 101 to 107
conidia. This yielded a total of 112 petri plates for the entire experiment. Cul-tures were allowed to grow at 29°C under continuous dark conditions for 7 days.To accurately visualize the sclerotia, plates were sprayed thoroughly with 70%ethanol in water to remove conidia and aerial mycelia. The exposed sclerotiathen were collected and lyophilized, and sclerotium production was determinedgravimetrically (mg [dry weight] per plate).
Numbers of conidia were determined only for the plates containing 106 and107 conidia/plate treated with the low-density (103 conidia/plate), high-density(107 conidia/plate), and medium extracts at 3 days. Conidia were counted byusing the method described above for the assay for conidial and sclerotial densitydependence using four replicates for each combination. The entire experimentwas repeated twice in order to examine reproducibility. Extracts from low- andhigh-density plates and medium extract were freshly prepared for each experi-ment.
Fatty acid assay. Linoleic acid, oleic acid (Cayman Chemical, Ann Arbor, MI),arachidonic acid (TCI America, Portland, OR), and linolenic acid (MP Biomedi-cals, Solon, OH) were dissolved in ethanol at concentrations of 0.1 and 1 mg/60�l and then applied to and dried on 12.5-mm-diameter filter paper disks. A filterpaper disk treated with 60 �l of ethanol was used as the solvent control. Afterdrying, the disks were laid on agar surfaces after plates were inoculated with 101
to 107 conidia/plate. Cultures were incubated in the dark inside cardboard boxes(resulting in continuous darkness) for 7 days. To accurately visualize the sclero-tia, plates were sprayed thoroughly with 70% ethanol in water to remove conidiaand aerial mycelia. Sclerotia were collected and freeze-dried, and sclerotiumproduction was determined gravimetrically (mg [dry weight] per plate). All of thefatty acid tests with wild-type strain NRRL 3357 were performed by using threereplicates, the entire experiment was repeated twice, and similar results wereobtained.
Nucleic acid analysis. Extraction of DNA from fungi, restriction enzymedigestion, gel electrophoresis, blotting, hybridization, and probe preparationwere performed by using standard methods (46). To examine Aflox expression,cultures of wild-type strain NRRL 3357 and complemented strains TSHB3.1Cand TSHB3.5C were cultured in shaken liquid GMM containing 2% yeast extractfor 48 h at 29°C and 250 rpm. Equal amounts of mycelium were then removed,placed in liquid GMM containing 2% sorbitol, and grown for an additional 2 hwith shaking 250 rpm at 29°C; this was followed by harvesting. Three separaterepetitions of this experiment yielded similar results. Aflox expression was alsoassessed by growing strain NRRL 3357, �lox strain TSHB2.39, and strainTSHB3.5C in liquid GMM containing 2% sorbitol for 24 h at 29°C and 220 rpmand then transferring the mycelia to filter paper on solid GMM containing 2%
TABLE 1. A. flavus strains used in this study
Strain(s)a Genotype Reference
NRRL 3357 Wild type 41NRRL 3357.5 pyrG� 41TJSPara17 pyrG; pyrG� This studyTSHB2.32, TSHB2.39,
and TSHB2.40�Aflox::pyrG; pyrG� This study
TSHB3.5C, TSHB3.1C,and TSHB3.2(3)C
Aflox::phleomycin; �Aflox::pyrG;pyrG�
This study
a Strains whose designations begin with T are original transformants.
VOL. 74, 2008 QUORUM SENSING IN A. FLAVUS 5675

sorbitol. Total RNA was extracted at the time of the shift (zero time) and 6, 12,24, and 48 h after the shift to solid culture. Two separate repetitions of thisexperiment yielded similar results.
Total RNAs were extracted from lyophilized mycelia using Trizol reagent(Invitrogen, Carlsbad, CA). RNA (10 to 30 �g) was separated on a 1.3% form-aldehyde gel. RNA was transferred to a Hybond membrane (Amersham Bio-science, Piscataway, NJ) by capillary action. RNA transcript analysis was per-formed using random primer radiolabeled DNA fragments (46) derived fromgenomic PCR products. Aflox expression studies were performed with an Aflox-specific DNA probe (a 1.07-kb PCR fragment) which was generated fromgenomic DNA using the following primers: loxF (5�-AGGCCCGTATGAGCAGTTGAGT-3�) and loxR (5�-GTTACCTTTACGGCTCCCCTCT-3�). An actin-specific DNA probe (a 0.5-kb PCR fragment) was generated from genomic DNAusing the following primers: ActinF (5�-ACAGTCCAAGCGTGGTATCC-3�)and ActinR (5�-GAAGCGGTCTGAATCTCCTG-3�). Signals were detectedwith a Phosphorimager-SI (Molecular Dynamics). Nucleotide sequences wereanalyzed and compared using Sequencher (Gene Codes) and ClustalW (www.ebi.ac.uk/clustalw/) programs (12).
Deletion and complementation of the A. flavus lox gene. The Aflox gene wasidentified by a TBLASTX search based on a putative A. fumigatus arachidonic5-lipoxygenase (accession no. XP_746463), which was used as a query sequence.Only one lox sequence was found, as confirmed by Southern analysis. The Afloxdeletion construct pSHB2, including the Aspergillus parasiticus pyrG marker geneand Aflox flanking sequences, was constructed using the following method. First,the modified primers 5� flank loxF SalI (5�-CCAGGTGTCGACAAAAAGGAGAAAGGAAGCAAA-3�) and 5� flank loxR EcoRI (5�-TCCTTCGAGTACACGTAGGGAATTCAGCCCG-3�) were used to PCR amplify a 1.07-kb flankingregion at the 5� untranslated region of the Aflox open reading frame using A.flavus genomic DNA as the template. The resulting amplified SalI-EcoRI PCRfragment was subcloned into pJW24 harboring the A. parasiticus pyrG cassette(5), yielding the vector pSHB1. Next, the modified primers 3� Flank loxF XbaI(5�-TTATTGTCTAGAGATGTCTTCGAGATTTGAACC-3�) and 3� FlankloxR SacI (5�-ATATTGTCATGTTTGGGACGGAGCTCTAATTG-3�) wereused to PCR amplify a 1.3-kb flanking region at the 3� end of the predicted Afloxopen reading frame. The amplified XbaI-SacI 3� flanking region was furtherligated into XbaI-SacI-digested plasmid pSHB1, generating plasmid pSHB2. Thefinal deletion vector, pSHB2, was used as a template to PCR amplify a fragmentcontaining the Aflox 5� flanking region, a pyrG selectable marker, and the Aflox3� flanking region with the following primers: Aflox nested F (5�-CAAGAGCAGTAGCAGCAGAAGG-3�) and Aflox nested R (5�-GGGCCCACTCACAACGTATCAT-3�). The PCR product was used to transform A. flavus NRRL 3357.5(pyrG�) to create Aflox deletion strains. Fungal transformation was performedessentially as described by Miller et al. (36). The Aflox deletion strains (�Aflox)were obtained by a double-crossover event, exchanging the pyrG selectablemarker gene for the Aflox coding region (see Fig. 4A). Southern analysis wasused to confirm that there was a single gene replacement event in TSHB2.39,TSHB2.40, and TSHB2.32 (Fig. 4B) with an Aflox full-length probe which wasobtained by performing PCR with primers Aflox.comp F (5�-GCTGATATTCCGTCCAGTTCG-3�) and Aflox.comp R (5�-CTGGATTTGTCATCGTGCAG-3�). Signals were detected with a Phosphorimager-SI (Molecular Dynamics).Complementation of the �lox transformant TSHB2.39 was achieved using thevector pSHB3. Plasmid pSHB3 was created by inserting a 3.09-kb fragmentcontaining the predicted promoter, coding sequence, and termination cassette ofAflox into pBC-Phleo. Plasmid pBC-Phleo carries the phleomycin resistancecassette in which the ble gene is under control of the A. nidulans gpdA promoterand the Saccharomyces cerevisiae CYC1 terminator (21).
Phylogenetic analysis. Conservation of LOX proteins was searched usingBLASTP. Hits for putative fungal, plant, and mammalian LOXs were aligned,and a phylogenetic tree was created using the ClustalW program. Pairwise scoresfor the amino acid sequences were calculated by dividing the number of identitiesin the best alignment by the number of residues compared (gap positions wereexcluded). The phylogenetic tree was calculated based on the multiple alignment,and the distances between the amino acid sequences in the alignment were thenused by the TreeView software program (12) to construct the tree shown in Fig.S1 in the supplemental material. The GenBank accession numbers of the se-quences of other fungal, plant, and mammalian LOXs which were used togenerate the tree are as follows: A. fumigatus, XP_746463 and XP_746844;Neosartorya fischeri, XP_001262545; Gaeumannomyces graminis, AAK81883;Magnaporthe grisea, XP_362938; Botryotinia fuckeliana, XP_001550612; Gib-berella moniliformis, AAW21637; Neurospora crassa, CAD37061; Chaetomiumglabosum, XP_001225066; Homo sapiens 12-LOX, NP_001130; soybean,1Y4K_A; Zea mays, NP_001105975; and Oryza sativa, NP_001055143.
Lipid extraction and fatty acid analysis. Wild-type strain NRRL 3357, �loxstrain TSHB2.39, and complemented strain TSHB3.5C were grown in 10 ml ofliquid GMM containing 2% sorbitol at a density of 105 conidia/plate understationary conditions at 29°C in the dark. Mycelial mats collected after 120 h ofgrowth were frozen in liquid nitrogen and ground with a mortar and pestle.Ground mycelial samples were then transferred to beakers and soaked in 30 mlof a chloroform-methanol mixture (1:1, vol/vol). Lipids were extracted for 48 hat �20°C. Next, the extraction mixtures were filtered through filter paper withlow porosity (Fisher Scientific) and transferred to 50-ml centrifuge tubes (Pyrex).Ten microliters of 0.97% KCl was then added to each tube, thoroughly mixed,and left to stand for 1 h. To separate the organic and aqueous phases, the tubeswere centrifuged (300 � g, 5 min), and the top aqueous layer and the interphasewere removed with a Pasteur pipette. The organic layer containing extractedlipids was washed with 2.0 ml of 0.97% KCl, and the top separated phase wasagain removed. The remaining chloroform extracts were dried with a stream ofnitrogen, and the solid lipid residues were resuspended in 1 ml of chloroform andtransferred into dark-glass vials. Fatty acids were converted into correspondingmethyl ester derivatives in 2% sulfuric acid in methanol (Sigma). Prepared fattyacid methyl esters were extracted with n-hexane. All solvents contained 5 mgliter�1 butylated hydroxytoluene as an antioxidant. Fatty acid methyl esters wereidentified by gas chromatography using a Hewlett-Packard 5890 equipped with acapillary column coated with DB-225 (length, 30 m; 0.25 mm; internal diameter,0.25 �m; Agilent Technologies, Inc., Wilmington, DE). The column temperaturewas kept at 70°C for 1 min and increased to 180°C at a rate of 20°C min�1 andthen to 220°C at a rate of 3°C min�1. Then the temperature was kept at 220°Cfor 15 min. The injector and detector temperatures were set at 250°C, and theinjection port temperature was set at 300°C. Peaks were identified by comparingretention times with the retention times of a set of authentic fatty acid standardsprovided by Supelco. The abundance of fatty acids was calculated from relativepeak areas.
Cell protein extraction and determination of LOX activity. Wild-type strainNRRL 3357, �lox strain TSHB2.39, and complemented strains TSHB3.5C,TSHB3.1C, and TSHB3.2(3)C were cultured in shaken liquid GMM containing2% yeast extract for 48 h at 29°C and 250 rpm, and equal amounts of myceliumwere then transferred into liquid GMM containing 2% sorbitol, allowed to growfor an additional 2 h with shaking at 250 rpm at 29°C, and harvested. Mycelialballs were immediately collected and transferred into 2-ml screw-top plastictubes containing chloroform-washed glass beads (diameter, �106 �m; Sigma, St.Louis, MO) and 1 ml of phosphate-saline buffer (pH 7.2). The mycelia werehomogenized mechanically using a Mini Beadbeater-8 (Biospec Products Inc.,Bartlesville, OK) for 1.5 min, followed by 3 min of cooling on ice. After this, theslurry was centrifuged (10,000 � g, 5 min, 4°C), and cell homogenates weretransferred into fresh Eppendorf tubes. Protein contents of cell extracts weredetermined using a bicinchoninic acid protein assay kit (Pierce Biotechnology,Rockford, IL) with bovine serum albumin as a standard. LOX was assayedspectrophotometrically using a modified method of Axelrod et al. (2). The 10mM substrate stock mixture contained 157.2 �l of linoleic acid and 157.2 �l ofTween 20 dissolved in 10 ml of water. The solution was thoroughly mixed, 1 mlof 1 M NaOH was added, and the volume was subsequently adjusted to 50 mlwith water. Prior to the assay, the substrate stock mixture was diluted withphosphate-saline buffer (1:4) to obtain a final linoleic acid concentration of 2.5mM. The solution was then bubbled with a 95% oxygen-5% CO2 mixture for 20min and allowed to equilibrate for 15 min before use. For the LOX assay, 100 �lof crude cell extract was added to 500 �l of the working substrate solution, mixedwell, and incubated in the dark for 20 min at room temperature. After this, thereaction mixture was extracted with 700 �l of a chloroform-methanol mixture(1:1, vol/vol) and vortexed, and the top aqueous layer and the interphase wereremoved after centrifugation (300 � g, 5 min). The organic layer was washed with200 �l of 0.97% KCl, and the top separated phase was again removed. Theremaining chloroform extracts were dried with a stream of nitrogen, and the solidlipid residues were resuspended in 500 �l of chloroform and transferred to a1-cm-path-length quartz cuvette. The formation of conjugated diene was mea-sured at 234 nm with a Beckman DU530 Life Science UV/VIS spectrophotom-eter. One unit of specific enzymatic activity was defined a change in absorbanceat 234 nm of 1 milli-absorbance unit (mAU) per mg of extracted proteins underthe assay conditions.
Statistical analysis. Data were analyzed using the JMP software package(version 3.2.6; SAS Institute, Inc., Cary, NC). Mean values for sclerotial andconidial formation affected by different conidium concentrations or strains werecompared using the least significant difference and the Tukey-Kramer multiple-comparison test (P � 0.05). Significantly different mean values are indicatedbelow.
5676 HOROWITZ BROWN ET AL. APPL. ENVIRON. MICROBIOL.
Nucleotide sequence accession number. Nucleotide sequence data reportedhere have been deposited in the GenBank database under accession numberEU486993.
RESULTS
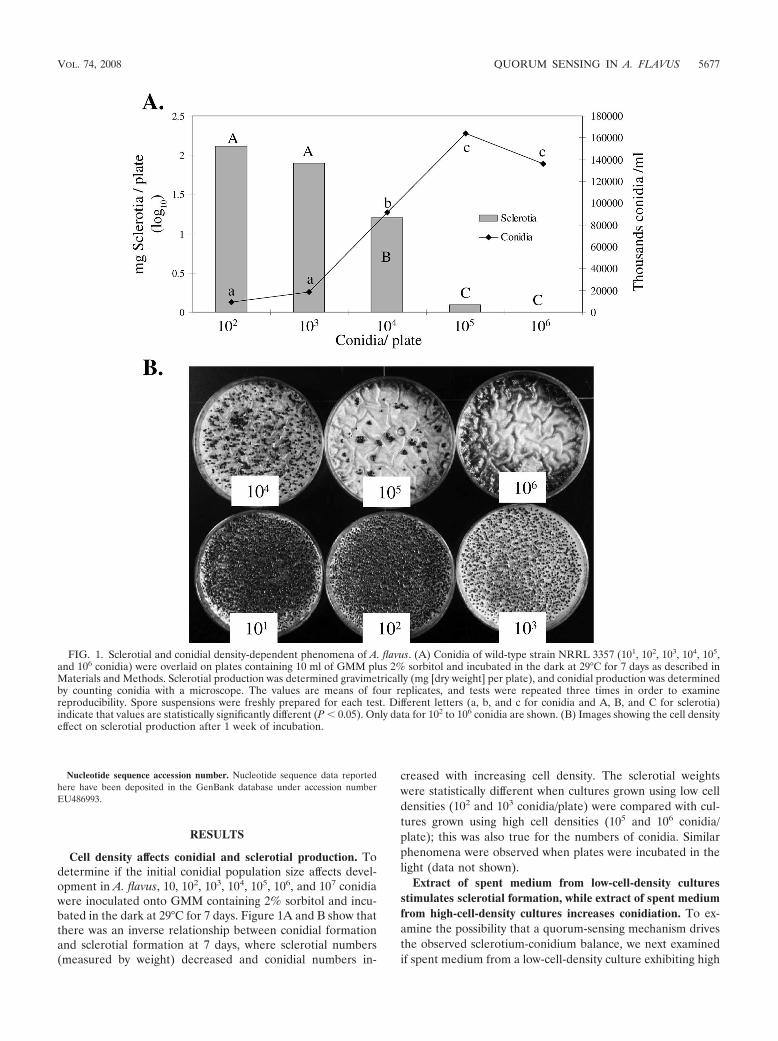
Cell density affects conidial and sclerotial production. Todetermine if the initial conidial population size affects devel-opment in A. flavus, 10, 102, 103, 104, 105, 106, and 107 conidiawere inoculated onto GMM containing 2% sorbitol and incu-bated in the dark at 29°C for 7 days. Figure 1A and B show thatthere was an inverse relationship between conidial formationand sclerotial formation at 7 days, where sclerotial numbers(measured by weight) decreased and conidial numbers in-
creased with increasing cell density. The sclerotial weightswere statistically different when cultures grown using low celldensities (102 and 103 conidia/plate) were compared with cul-tures grown using high cell densities (105 and 106 conidia/plate); this was also true for the numbers of conidia. Similarphenomena were observed when plates were incubated in thelight (data not shown).
Extract of spent medium from low-cell-density culturesstimulates sclerotial formation, while extract of spent mediumfrom high-cell-density cultures increases conidiation. To ex-amine the possibility that a quorum-sensing mechanism drivesthe observed sclerotium-conidium balance, we next examinedif spent medium from a low-cell-density culture exhibiting high
FIG. 1. Sclerotial and conidial density-dependent phenomena of A. flavus. (A) Conidia of wild-type strain NRRL 3357 (101, 102, 103, 104, 105,and 106 conidia) were overlaid on plates containing 10 ml of GMM plus 2% sorbitol and incubated in the dark at 29°C for 7 days as described inMaterials and Methods. Sclerotial production was determined gravimetrically (mg [dry weight] per plate), and conidial production was determinedby counting conidia with a microscope. The values are means of four replicates, and tests were repeated three times in order to examinereproducibility. Spore suspensions were freshly prepared for each test. Different letters (a, b, and c for conidia and A, B, and C for sclerotia)indicate that values are statistically significantly different (P � 0.05). Only data for 102 to 106 conidia are shown. (B) Images showing the cell densityeffect on sclerotial production after 1 week of incubation.
VOL. 74, 2008 QUORUM SENSING IN A. FLAVUS 5677
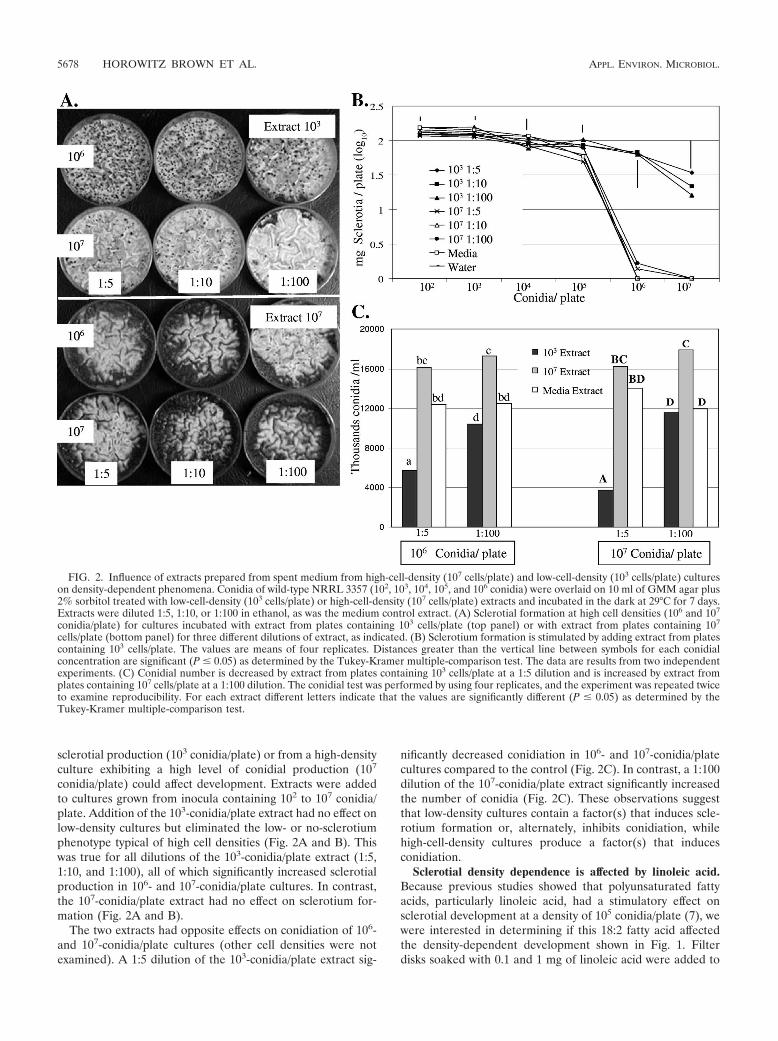
sclerotial production (103 conidia/plate) or from a high-densityculture exhibiting a high level of conidial production (107
conidia/plate) could affect development. Extracts were addedto cultures grown from inocula containing 102 to 107 conidia/plate. Addition of the 103-conidia/plate extract had no effect onlow-density cultures but eliminated the low- or no-sclerotiumphenotype typical of high cell densities (Fig. 2A and B). Thiswas true for all dilutions of the 103-conidia/plate extract (1:5,1:10, and 1:100), all of which significantly increased sclerotialproduction in 106- and 107-conidia/plate cultures. In contrast,the 107-conidia/plate extract had no effect on sclerotium for-mation (Fig. 2A and B).
The two extracts had opposite effects on conidiation of 106-and 107-conidia/plate cultures (other cell densities were notexamined). A 1:5 dilution of the 103-conidia/plate extract sig-
nificantly decreased conidiation in 106- and 107-conidia/platecultures compared to the control (Fig. 2C). In contrast, a 1:100dilution of the 107-conidia/plate extract significantly increasedthe number of conidia (Fig. 2C). These observations suggestthat low-density cultures contain a factor(s) that induces scle-rotium formation or, alternately, inhibits conidiation, whilehigh-cell-density cultures produce a factor(s) that inducesconidiation.
Sclerotial density dependence is affected by linoleic acid.Because previous studies showed that polyunsaturated fattyacids, particularly linoleic acid, had a stimulatory effect onsclerotial development at a density of 105 conidia/plate (7), wewere interested in determining if this 18:2 fatty acid affectedthe density-dependent development shown in Fig. 1. Filterdisks soaked with 0.1 and 1 mg of linoleic acid were added to
FIG. 2. Influence of extracts prepared from spent medium from high-cell-density (107 cells/plate) and low-cell-density (103 cells/plate) cultureson density-dependent phenomena. Conidia of wild-type NRRL 3357 (102, 103, 104, 105, and 106 conidia) were overlaid on 10 ml of GMM agar plus2% sorbitol treated with low-cell-density (103 cells/plate) or high-cell-density (107 cells/plate) extracts and incubated in the dark at 29°C for 7 days.Extracts were diluted 1:5, 1:10, or 1:100 in ethanol, as was the medium control extract. (A) Sclerotial formation at high cell densities (106 and 107
conidia/plate) for cultures incubated with extract from plates containing 103 cells/plate (top panel) or with extract from plates containing 107
cells/plate (bottom panel) for three different dilutions of extract, as indicated. (B) Sclerotium formation is stimulated by adding extract from platescontaining 103 cells/plate. The values are means of four replicates. Distances greater than the vertical line between symbols for each conidialconcentration are significant (P � 0.05) as determined by the Tukey-Kramer multiple-comparison test. The data are results from two independentexperiments. (C) Conidial number is decreased by extract from plates containing 103 cells/plate at a 1:5 dilution and is increased by extract fromplates containing 107 cells/plate at a 1:100 dilution. The conidial test was performed by using four replicates, and the experiment was repeated twiceto examine reproducibility. For each extract different letters indicate that the values are significantly different (P � 0.05) as determined by theTukey-Kramer multiple-comparison test.
5678 HOROWITZ BROWN ET AL. APPL. ENVIRON. MICROBIOL.

cultures, and the production of sclerotia was assessed after 6days. No effects were observed at either low cell densities (10to 103 conidia/plate) or high cell densities (106 and 107 conidia/plate), but addition of 1 mg linoleic acid resulted in a signifi-cant increase in sclerotium weight at intermediate cell densities(104 and 105 conidia/plate) compared to the control (Fig. 3Aand B). Sclerotial weight increased 1.4-fold for the 104-conidia/plate cell population and 1.7-fold for the 105-conidia/plate cellpopulation. These changes effectively shifted the density-de-pendent curve toward increased sclerotial production at inter-mediate cell densities.
To assess if other fatty acids that differ in saturation, chainlength, and the position of double bonds affect the density-dependent development similar to linoleic acid, filter diskscontaining linoleic, oleic (18:1), linolenic (18:2), and arachi-donic (20:4) acids were added to cultures grown at densities of104 and 105 conidia/plate. In contrast to linoleic acid, none ofthese fatty acids stimulated sclerotial formation, and oleic andlinolenic acids had the opposite effect at a density of 105
conidia/plate (that is, they depressed sclerotial formation) (Fig.3C). All studies were compared to the ethanol treatment, asethanol by itself stimulated sclerotial production at a density of105 conidia/plate (Fig. 3C).
Identification of A. flavus LOX and sequence homology. BLASTsearches of the A. flavus genome (http://www.aspergillusflavus.org/genomics) with the A. fumigatus putative arachidonate 5-lipoxy-genase (accession no. XP_746463) revealed the presence ofonly one gene encoding a LOX homolog, which was designatedAflox. The Aflox gene is located on chromosome 3 (GenBankaccession no. EU486993). Based on the predicted annotation,Aflox encodes a 574-amino-acid protein after a four-intronsplicing event of mRNA. When the predicted amino acid se-quence of AfLOX was subjected to a BLASTP search, theprogram reported homology with the consensus sequences ofthe LOX family (Pfam 00305; “LOX motif,” His-X4-His-X4-His-X17-His-X8-His; http://www.ncbi.nlm.nih.gov/Structure/cdd/).The iron atom in LOX is bound by four ligands, three of whichare histidine residues. Six histidines are conserved in all LOXsequences, and five of them are clustered in a stretch of 40amino acids and are important for LOX activity. Residues thatact as iron coordination ligands in plant (48) and mammalian(17, 23) LOXs are very well conserved and are present inAfLOX. When the AfLOX sequence was used as a query withthe Protein Data Base, the highest scores were the scores forpredicted and known fungal LOXs, followed by mammalianand plant LOXs. The ClustalW software program was used toalign A. flavus LOX with several fungal, plant, and mammalianLOXs, and a tree was created by using the TreeView softwareprogram (see Fig. S1A in the supplemental material). Thepredicted AfLOX exhibited 75 and 77% sequence identity witharachidonate 5-lipoxygenase of A. fumigatus (accession no.XP_746463) and N. fischeri LOX (accession no. XP_001262545), respectively, and 46% identity with manganeseLOX of G. graminis (accession no. AAK81883). Unlike the G.graminis manganese LOX (49), AfLOX is predicted to havethe more common nonheme iron at its catalytic center. Thehomology of AfLOX to the LOX family included homology inthe region that contains two His residues found in � helix 9 inthe characteristic 30-amino-acid sequence WLLAK-X15-H-X4-H-X3-E (27). However, the sequence of AfLOX contained
only 29 amino acids, WLFAK-X14-H(310)-X4-H(315)-X3-E, asdid the N. fischeri (accession no. XP_001262545) and A. fu-migatus (accession no. XP_746463) LOXs (see Fig. S1B in thesupplemental material). Residues important in plant andmammalian LOX catalysis are conserved in AfLOX and otherputative fungal LOXs (see Fig. S1B in the supplemental ma-terial). A number of residues were completely conserved infungal LOXs.
Identification of A. flavus �lox mutants. The LOX gene of A.flavus was deleted by homologous recombination by replacingAflox with A. parasiticus pyrG in a pyrG1 mutant (NRRL3357.5), as shown in Fig. 4A. A PCR screen of 100 transfor-mants for integration of pyrG at the Aflox locus was employed.Transformants showing replacement of Aflox by pyrG by PCRwere then confirmed by Southern blot analysis, in which ex-pected hybridization band patterns for a gene replacementevent were identified in several transformants (Fig. 4B). Threetransformants with Aflox deleted that had identical genotypesas determined by Southern analysis, TSHB2.32, TSHB2.39,and TSHB2.40 (Fig. 4B), were used for physiological analysis.The following two strains were used as controls: the prototrophA. flavus wild-type strain NRRL 3357 and TJSpara17, whichwas obtained by transforming NRRL 3357.5 to prototrophywith pJW24 containing the A. parasiticus pyrG gene alone.Extensive physiological testing showed that the two controlstrains have the same phenotype, as demonstrated below (alsodata not shown).
Complementation of Aflox in the �lox background was con-firmed by Southern analysis to determine plasmid copy num-ber. Strains TSHB3.5C and TSHB.2(3)C harbored at least twocopies of Aflox (Fig. 4B) (extra bands were observed for boththese strains), while TSHB3.1C contained a single-copy inser-tion. These strains were examined to determine if copy numberhad an effect on Aflox expression, LOX activity, and fungaldevelopment.
Density-dependent development is minimized in �lox strains.Because inhibition of LOX activity by chemical inhibitors hasbeen shown to affect fungal morphological shifts (28), includ-ing those in Aspergillus (A. M. Calvo and N. P. Keller, unpub-lished data), we thought that it was possible that the density-dependent development demonstrated for the wild type couldbe affected in lox mutants. The relative ability of �lox strainsTSHB2.32, TSHB2.39, and TSHB2.40 to form sclerotia andconidia was determined using the conditions described abovefor Fig. 1. Conidial production and sclerotial production by theA. flavus wild-type NRRL 3357 and TJSpara17 control strainsand complemented strains TSHB3.1C and TSHB3.5C weresimilarly regulated by cell density. However, the pattern ofdevelopment was greatly skewed in the �lox strains. The mostobvious phenotype was the greatly increased production ofsclerotia in all �lox strains at high cell densities (Fig. 5A and5B). Unlike the the sclerotial production by the wild-type con-trols, the sclerotial production was only minimally reduced athigh inoculum concentrations (105 to 107 conidia/plate). Thisprofile was nearly identical to the profile observed after addi-tion of extract from spent medium from 103-conidia/plate cul-tures (Fig. 2B). Complemented strains TSHB3.1C andTSHB3.5C had a wild-type phenotype, thus confirming that theeffects on sclerotial formation were due to deletion of the loxgene (Fig. 5B). Conidial production was also examined in these
VOL. 74, 2008 QUORUM SENSING IN A. FLAVUS 5679
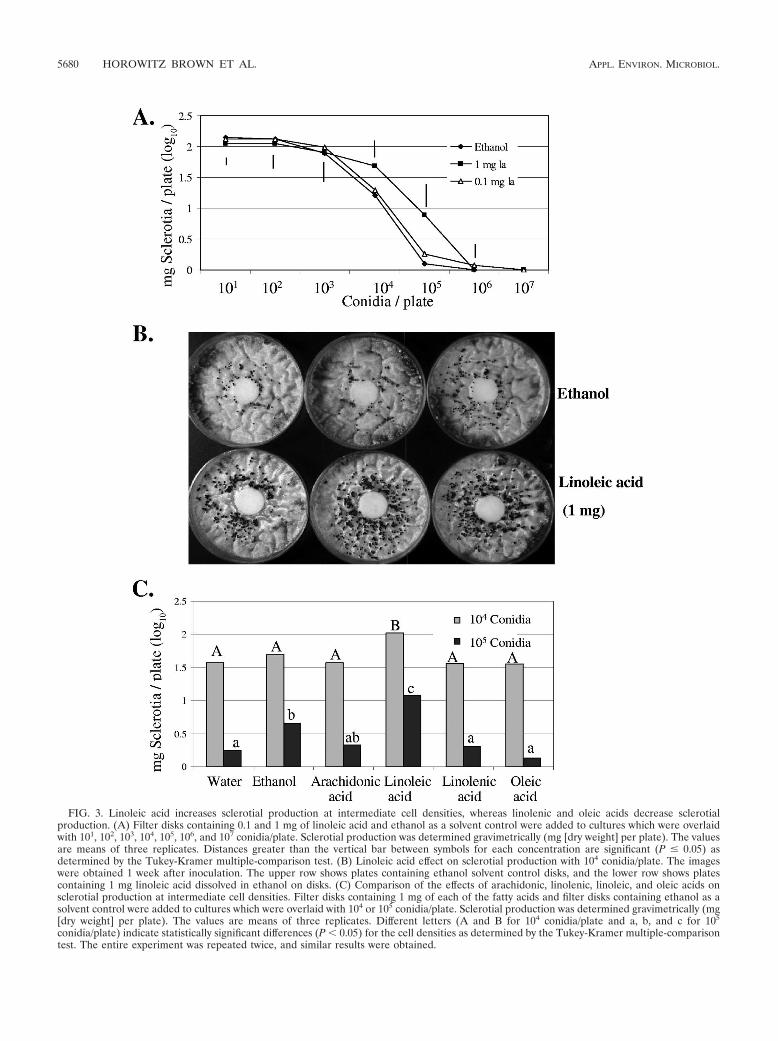
FIG. 3. Linoleic acid increases sclerotial production at intermediate cell densities, whereas linolenic and oleic acids decrease sclerotialproduction. (A) Filter disks containing 0.1 and 1 mg of linoleic acid and ethanol as a solvent control were added to cultures which were overlaidwith 101, 102, 103, 104, 105, 106, and 107 conidia/plate. Sclerotial production was determined gravimetrically (mg [dry weight] per plate). The valuesare means of three replicates. Distances greater than the vertical bar between symbols for each concentration are significant (P � 0.05) asdetermined by the Tukey-Kramer multiple-comparison test. (B) Linoleic acid effect on sclerotial production with 104 conidia/plate. The imageswere obtained 1 week after inoculation. The upper row shows plates containing ethanol solvent control disks, and the lower row shows platescontaining 1 mg linoleic acid dissolved in ethanol on disks. (C) Comparison of the effects of arachidonic, linolenic, linoleic, and oleic acids onsclerotial production at intermediate cell densities. Filter disks containing 1 mg of each of the fatty acids and filter disks containing ethanol as asolvent control were added to cultures which were overlaid with 104 or 105 conidia/plate. Sclerotial production was determined gravimetrically (mg[dry weight] per plate). The values are means of three replicates. Different letters (A and B for 104 conidia/plate and a, b, and c for 105
conidia/plate) indicate statistically significant differences (P � 0.05) for the cell densities as determined by the Tukey-Kramer multiple-comparisontest. The entire experiment was repeated twice, and similar results were obtained.
5680 HOROWITZ BROWN ET AL. APPL. ENVIRON. MICROBIOL.

strains. Again similar to the results obtained for 103-conidia/plate extracts, the �lox strains produced significantly fewer(P � 0.05) conidia at high cell densities (�105 conidia/plate).Complemented strains TSHB3.1C (Fig. 5C) and TSHB3.5C(data not shown) showed wild-type growth.
Expression of Aflox. Aflox expression was examined in A.flavus wild-type strain NRRL 3357, �lox strain TSHB2.39, andcomplemented strains TSHB3.1C and TSHB3.5C. Aflox is alow-expression gene under the conditions used as expressionwas not detected in wild-type strain NRRL 3357 (Fig. 6A, lanesa and b, and Fig. 6B, lanes a, b, g, and h) or, as expected, in the�lox strain (Fig. 6A, lanes c and d, and Fig. 6B, lanes c, d, k,and l). However, expression of Aflox was observed in the com-plemented strains. While complemented strain TSHB3.1C(harboring a single-copy insertion) expressed a clear transcript(Fig. 6A, lanes e and f), complemented strain TSHB3.5C (har-boring at least two copies of Aflox and likely more) produceda greatly increased transcript and also produced an additionalsmaller band (Fig. 6B, lanes e, f, k, and l and data not shown).This small band was not observed for single-copy strainTSHB3.1C.
LOX activity and fatty acid analysis. LOX enzymes canutilize many fatty acids (39), and linoleic acid is one of thepreferred substrates (33, 43, 49), as well as a common lipidconstituent of Aspergillus (6). Therefore, cell fractions ofwild-type strain NRRL 3357, �lox strain TSHB2.39, and thecomplemented strains were examined for linoleic acid LOXactivity. Table 2 shows that the LOX activity of the �loxstrain (6.5 6.8 1 mAU) was less than that of either thewild-type strain or the complemented strains. The minimalLOX activity detected in �lox strain TSHB2.39 likely re-sulted from trace activity and may have represented back-ground or nonspecific peroxidation. There was no significantdifference in LOX activity between the wild-type strain(97.1 8.7 mAU) and the complemented strains. However,the two multicopy complemented strains, TSHB3.5C (82.9 7.5mAU) and TSHB3.2(3)C (85.6 8.6 mAU), had slightly lowerLOX activity than the single-copy complemented strain,TSHB3.1C (113.2 8.1 mAU). This may have reflected theputative degradation of the transcript in multicopy strains(Fig. 6B).
Because oxylipin mutants of A. nidulans have altered fatty
FIG. 4. Deletion of the LOX gene in A. flavus. (A) Diagram of the Aflox coding sequence and how it was replaced with the A. parasiticus pyrGselectable marker by homologous recombination to generate the �lox mutants. (B) Southern analysis of wild-type strain NRRL 3357, �lox mutantsTSHB2.39 and TSHB2.40, and complemented strains TSHB3.5C, TSHB3.1C, and TSHB3.2(3)C. Genomic DNA was digested with ApaI, NcoI,and PstI to determine Aflox deletion in the �lox strains or plasmid copy number in the complemented strains. The expected hybridization bandpatterns for ApaI digestion were a single 3.9-kb band for the wild-type strain, 3.1- and 2.2-kb bands for the �lox strains, and an additional upperband at �9.5-kb for complemented strains TSHB3.1C and TSHB3.2(3)C. The expected hybridization band patterns for NcoI digestion were 4.1-,1.3-, and 0.77-kb bands for the wild-type strain, 4.5- and 1.2-kb bands for the �lox strains, and an additional ca. 2.4-kb band for the complementedstrains. The expected hybridization band patterns for PstI were 1.9- and 3.3-kb bands for the wild-type strain, 2.4- and 3.2-kb bands for the �loxstrains, and additional 5.1- and 4.3-kb bands for an Aflox copy insertion in the complemented strains. Note the extra copies of Aflox inTSHB3.2(3)C and TSHB3.5C (indicated by asterisks). A 3.9-kb PCR product containing Aflox and the 5� and 3� flanking regions was used as alox probe (indicated by the arrow in panel A).
VOL. 74, 2008 QUORUM SENSING IN A. FLAVUS 5681

acid contents (52), we next examined any possible effects of the�lox mutation on fatty acid biosynthesis. The mycelial fattyacid contents of the �lox and TSHB3.5C strains were com-pared to that of wild-type strain NRRL 3357 using gas chro-matography analysis of mycelia grown under dark conditions at29°C for 5 days (Table 3). Whereas deletion of Aflox did notgreatly alter the overall fatty acid profile for saturated andunsaturated fatty acids, there were some small effects on indi-
FIG. 5. Effect of Aflox deletion on sclerotial and conidial density-dependent regulation. Conidia of wild-type NRRL 3357 and TJSPara17control strains, �lox strains TSHB2.32, TSHB2.39, and TSHB2.40, and complemented strains TSHB3.5C and TSHB3.1C (102, 103, 104, 105, 106,and 107 conidia) were overlaid on 10 ml of GMM plus 2% sorbitol and incubated in the dark at 29°C for 7 days. (A) Sclerotial formation after1 week for densities of 105 to 107 conidia/plate. (B) Sclerotium production increased at high population densities in �lox strains. Each data pointis the mean of three replicates. Distances greater than the vertical bar at each concentration are significant (P � 0.05) as determined by theTukey-Kramer multiple-comparison test. (C) Conidial production decreased at high population densities in �lox strains. Conidial production at densitiesof 102, 104, 105, and 106 cells/plate was determined after 3 days. For each of the fungal strains, the conidial test was performed by using five replicates.Different letters indicate that values are significantly different (P � 0.05) as determined by the Tukey-Kramer multiple-comparison test.
FIG. 6. Aflox expression studies. (A) Aflox expression in liquidcultures of wild-type strain NRRL 3357, �lox strain TSHB2.39, andcomplemented strain TSHB3.1C assessed as described in Materialsand Methods. Three separate repetitions of the experiment yieldedsimilar results. The upper panel was probed with a 1.07-kb internal PCRfragment of Aflox. The middle panel was probed with actin, and rRNAstained with ethidium bromide to indicate RNA loading is shown in lowerpanel. (B) Aflox is degraded in multicopy strain TSHB3.5C. Liquid cul-tures of strain NRRL 3357, �lox strain TSHB2.39, and strain TSHB3.5Cwere transferred to solid medium to induce conidiophore formation asdescribed in the text. Aflox expression in the complemented strainTSHB3.5C was accompanied by a small transcript(s), as indicated by thelower arrow at 12 h (lanes a to f) and 24 h (lanes g to l) after the shift tosolid culture. Two separate repetitions of the experiment yielded similarresults. The upper panel was probed with a 1.07-kb internal PCR frag-ment of Aflox, and rRNA stained with ethidium bromide to indicate RNAloading is shown in the lower panel.
TABLE 2. LOX activities of mycelial extracts from A. flavuswild-type strain NRRL 3357, �lox strain TSHB2.39,
and complemented strains TSHB3.5C, TSHB3.1C,and TSHB3.2(3)C
Samplea LOX enzymaticactivity (U)b
NRRL 3357 (wild type) ........................................................ 97.1 8.7TSHB2.39 (�lox) .................................................................... 6.5 6.8TSHB3.5C (complemented lox) ........................................... 82.9 7.5TSHB3.1C (complemented lox) ...........................................113.2 8.1TSHB3.2(3)C (complemented lox) ...................................... 85.6 8.6
a Total proteins were extracted from NRRL 3357, TSHB2.39, TSHB3.5C,TSHB3.1C, and TSHB3.2(3)C, and LOX activity was assayed as described in thetext.
b One unit of enzymatic specific activity was defined a change in absorbance at234 nm of 1 mAU per mg of extracted proteins under the assay conditions used.The data are the means standard errors from three experiments.
5682 HOROWITZ BROWN ET AL. APPL. ENVIRON. MICROBIOL.

vidual fatty acids. Some of the fatty acid differences were sim-ilar in the deletion strain and the complemented strain andlikely are not significant for the developmental processes de-scribed here. Unique to the deletion strain were slight in-creases in the levels of palmitic acid (16:0) and oleic acid (18:1)and a decrease in the level of linolenic acid (18:3). Similar toother Aspergillus species, linoleic acid, whose levels were equiv-alent in all strains, is the major constituent of A. flavus fattyacids and comprises 38% of the total fatty acid content in A.flavus mycelia.
DISCUSSION
In this study cell density was found to have a profound effecton morphological transitions in A. flavus; low cell densitiesyielded primarily sclerotial cultures, and high cell densitiesyielded conidial cultures. Cell density alteration of develop-ment has been observed in other fungi, including C. albicans,C. ulmi, Histoplasma capsulatum, and S. cerevisiae (25, 26, 28,32). These four organisms are dimorphic yeasts, and much ofthe literature describes cell density effects on the yeast-to-hypha switch; e.g., C. ulmi develops as a budding yeast at highdensities and as filaments at low densities (26). Studies of C.albicans and S. cerevisiae have identified quorum-sensing mol-ecules governing this switch. In the former organism farnesolblocks the filament-to-yeast transition, whereas tyrosol pro-motes the yeast-to-filament switch; all of this is density depen-dent (11, 44). Two aromatic alcohols, phenylethanol and tryp-tophol, induce filamentation in S. cerevisiae (10). Although amolecule that conditions the C. ulmi switch has not been iden-tified, chemical inhibition of LOX activity promotes the myce-lium-to-yeast conversion in C. ulmi (28). The latter data sug-gest that an oxylipin signal is involved in cell density-dependentphenomena in this fungus.
Cell density regulates development transitions in A. flavus.While filamentous fungi do not exhibit yeast growth patterns,they do develop several tissue types. Members of the genusAspergillus display several morphologies, including vegetativehyphal growth, asexual sporulation (conidia produced on con-idiophores), and in some species sclerotium or cleistotheciumformation. Several studies have indicated that sclerotia or cleis-tothecia are produced under conditions different than thosefavoring conidiophore formation. Conidiophore production isinduced by light, and sclerotia and cleistothecia are producedmore abundantly in the dark (5, 6, 7, 37). In A. flavus, circadianrhythms regulate both sclerotial and conidial formation, andconidial production is separated from sclerotial production
(19; and N. P. Keller and D. Bell-Pedersen, unpublished re-sults). Here we show that A. flavus sclerotial formation andconidial formation are regulated in opposite ways by cell den-sity (Fig. 1). High cell densities result in pure conidial cultures,whereas populations with high sclerotial densities are sup-ported at low cell densities.
The balancing of sclerotium and conidium levels was remi-niscent of a lipid-mediated cleistothecium-conidium balancediscovered for A. nidulans (52) and, to a certain extent, foranother strain of A. flavus (7), leading to the suggestion that aquorum-driven mechanism may regulate morphological tran-sitions in Aspergillus spp. (50). Supporting this hypothesis,spent medium from low-cell-density, high-sclerotium-densitycultures was found to greatly increase sclerotium formation inhigh-cell-density cultures normally devoid of sclerotia (Fig. 2).Extracts from high-cell-density cultures did not affect sclerotialproduction at any cell density but led to a significant increasein the number of conidia (Fig. 2C). We suggest that high-cell-density extracts contain a factor(s) that induces conidiation, incontrast to the low-cell-density extracts, which contain a fac-tor(s) that stimulates sclerotial biogenesis and/or inhibits con-idiogenesis.
Lipid mediation of morphological transitions. Lipids havebeen implicated repeatedly as signals that regulate fungal re-productive development and growth, and in pathogenic fungithey are involved in signal communication with host cells (4,24, 30, 39). Furthermore, in the model fungus A. nidulans thecleistothecium-conidium balance is governed by oxylipins (51,52), reminiscent of the sclerotium-conidium balance observedhere. The accumulating data support the hypothesis that lipidpools may be involved in cell density transitions in Aspergillus.
Unsaturated fatty acids in particular have been shown toaffect fungal development in several genera. Fatty acids serveas signals to initiate the filamentous growth needed for invad-ing plant tissue in the smut pathogen Ustilago maydis (29), andlinoleic acid has been shown to induce perithecia in N. crassa(40). Interestingly, fatty acid oscillations are circadian regu-lated in N. crassa (45), suggesting that there may be an inter-action between lipid signals and the circadian regulation ofdevelopment in A. flavus observed previously (19). Likewise,Goottwald and Wood (18) reported that some fatty acids in-hibit sporulation of Cladosporium caryigenum. In Aspergillusspp. fatty acid stimulation of sporulation and/or sclerotial andcleistothecial production is dependent on chain length and thepresence of double bonds (7). Polyunsaturated fatty acids hadthe greatest effect on morphology; for example, linoleic acid
TABLE 3. Fatty acid composition of mycelia of A. flavus wild-type strain NRRL 3357, �lox strain TSHB2.39, andcomplemented strain TSHB3.5Ca
Strain% ofb:
14:0 16:0 16:1 18:0 18:1 18:2 18:3
NRRL 3357 2.3 0.7 17.3 0.4 2.7 1.0 10.3 0.9 28.0 0.2 38.0 2.6 1.3 0.0TSHB2.39 0.6 0.1 19.3 0.2 1.3 0.1 9.3 0.4 30.4 0.1 38.7 0.7 0.5 0.1TSHB3.5C 0.4 0.0 18.3 1.7 0.6 0.0 11.7 1.0 29.1 2.0 38.5 1.0 1.4 0.1
a The analysis was carried out with 5-day-old mycelia grown in liquid GMM containing 2% sorbitol at a concentration of 105 conidia/plate under stationary conditionsat 29°C in the dark.
b The values are the means standard errors for three replicates and are the weight percentages of fatty acid methyl esters based on the lyophilized weight of themycelia.
VOL. 74, 2008 QUORUM SENSING IN A. FLAVUS 5683

was shown to induce sclerotial production and decrease conid-ial production in cultures of A. flavus grown using initial celldensities of 105 cells/plate. Our study showed that the sclerotialstimulation by linoleic acid is limited to intermediate cell den-sities (Fig. 3). This effect could be a direct result of use oflinoleic acid as a carbon source and/or an indirect resultthrough metabolic pathways utilizing this fatty acid. In contrastto linoleic acid, oleic and linolenic acids had inhibitory effectson sclerotium formation.
Role of LOX in A. flavus cell density transitions. Unsatur-ated fatty acids can be metabolized by beta-oxidation pathways(35) or used as substrates by various oxygenases, includingLOXs, dioxygenases, and P450 monooxygenases, to generatediverse oxylipins (34, 49). Three key observations led us toexamine the possible role of A. flavus LOX in the cell-density-dependent conidium-to-sclerotium switch. First, chemical in-hibition of LOX interfered with the dimorphic switch in C.ulmi (28). Second, physiological studies of Aspergillus showedthat purified oxylipins could affect morphological develop-ment. For example, the plant linoleic LOX product 9(S)-hy-droperoxy-trans-10-cis-12-octadecadienoic acid (9S-HPODE)induced cleistothecial formation in A. nidulans at levels of 0.01and 0.1 mg but conidial formation at 10-fold-higher concen-trations (7). Third, genetic disruptions of oxylipin-generatingdioxygenases (ppo genes) in A. nidulans greatly altered theconidium-cleistothecium balance in this species; loss of onedioxygenase yielded a nearly pure conidial culture (52), andloss of two other dioxygenases yielded a predominantly cleis-tothecial culture (51). Because analysis of the A. flavus genomeindicated that there are four dioxygenases (ppoA, ppoB, ppoC,and ppoD) and it would be difficult to disrupt all four genes inthis asexual species, we chose to characterize the single, puta-tive LOX gene designated Aflox.
Deletion of Aflox eliminated the switch from a sclerotialculture to a conidial culture (Fig. 5). In the wild type and thecomplemented controls, conidial production significantly in-creased and sclerotial production decreased at cell densitiesbetween 104 and 105 cells/plate. At concentrations of �106
cell/plate, no sclerotia were produced and conidial productionwas maximal. In the �Aflox mutants, sclerotial productiondecreased minimally and conidial production remained low atthese cell densities. This phenotype was very similar to thatobserved when extract from spent medium from a plate con-taining 103 conidia was added to growing cultures (Fig. 2). Thissuggests that some AfLOX-derived oxylipin and/or a down-stream product(s) signals sclerotium-to-conidium transitions inA. flavus. These observations are reminiscent of the concen-tration-dependent effect of the oxylipin 9S-HPODE on A.nidulans cleistothecium-to-conidium shifts (7).
The LOX activity assay showed that there was a significantreduction in linoleic acid-mediated activity in the �Aflox straincompared to the wild-type and complemented strains, suggest-ing that linoleic acid is an AfLOX substrate. However, it is wellestablished that oxylipin-generating enzymes (dioxygenase,LOX, and cyclooxygenase) exhibit activity with more than onesubstrate, and it is possible that AfLOX utilizes other unsat-urated fatty acids as substrates. AfLOX shows the highestlevels of homology to predicted arachidonate 5-lipoxygenasesin other aspergilli, which suggests that arachidonic acid, a fattyacid found at negligible levels in Aspergillus spp., may also be a
substrate for AfLOX. We note that, along with changes inLOX activity, loss of Aflox resulted in slight changes in fattyacid percentages, including a slight increase in the oleic acidcontent and a decrease in the linolenic acid content. However,the developmental effects observed for the �Aflox strain aremore likely due to changes in oxylipin content than to changesin fatty acids, as one would predict that a slight increase in thecontent of oleic acid (which likely has the greatest effect due tothe total percentage of this fatty acid) would reduce sclerotialproduction (Fig. 3C) rather than increase production, the phe-notype of �Aflox. Therefore, we postulate that one or moreAfLOX-generated oxylipins may play a role in quorum sens-ing, as observed from the lack of a morphological shift in the�Aflox strains (Fig. 5). Such AfLOX products, possibly pro-moting conidial development, would be in opposition to thefactor(s) acting in the spent medium from low-cell-density cul-tures (Fig. 2).
The undetectable level of Aflox in the wild-type strain underthe conditions tested (Fig. 6) may reflect the low levels of aputative quorum-sensing molecule needed or may simply re-flect the cultural conditions used. Similarly, the level of expres-sion of ppoB, which is involved in the production of oleicacid-derived oxylipin in A. nidulans, was very low despite aprofound effect on the conidium-to-cleistothecium switch (52).Interestingly, differences in Aflox expression among the differ-ent complemented strains had no effect on conidium-to-scle-rotium transitions. As the LOX activities were similar for all ofthe complemented strains, it is possible that there is somegene-silencing phenomenon, possibly RNA interference activ-ity (20), in multipcopy strains and/or that there is posttran-scriptional regulation of this enzyme.
These findings support the hypothesis that a quorum-sensingmechanism governs morphological shifts in the filamentousfungus A. flavus. The chemical signals integrate cell density anddevelopment by regulating specialized processes such as mor-phogenesis, perhaps as a means of responding effectively tochanges in environmental opportunities. The conidium is thedispersal spore expected to be produced at times of starvation,conditions that may arise with high population densities andcells competing for limited nutrients. The sclerotium is a re-sistant body, establishing a niche for the producing organismand deterring predators through production of toxic metabo-lites (53). A sentinel system alerting the fungus to environ-ments favoring “flight” versus “fight” would be evolutionarilybeneficial for the organism.
ACKNOWLEDGMENTS
We thank Gary A. Payne for access to A. flavus genome data and forproviding strains, James Scott for critical reading of the manuscript,and Jon P. Woods and Stacey Schultz-Cherry for sharing some of theequipment used in this study.
This research was funded by BARD grant FI-384-2006 to S.H.B. andby NSF grant IOB-0544428 (subagreement S060039) to N.P.K.
REFERENCES
1. Allison, C., and C. Hughes. 1991. Bacterial swarming: an example of pro-karyotic differentiation and multicellular behavior. Sci. Prog. 75:403–422.
2. Axelrod, B., T. M. Cheesebrough, and S. Laakso. 1981. Lipoxygenase fromsoybean. Methods Enzymol. 71:441–451.
3. Bhatnagar, D., J. W. Carry, K. Ehrlich, J. Yu, and T. E. Cleveland. 2006.Understanding the genetics of regulation of aflatoxin production and As-pergillus flavus development. Mycopathologia 162:155–166.
4. Brodhagen, M., D. I. Tsitsigiannis, E. Hornung, C. Goebel, I. Feussner, and
5684 HOROWITZ BROWN ET AL. APPL. ENVIRON. MICROBIOL.

N. P. Keller. 2008. Reciprocal oxylipin-mediated cross-talk in the Aspergillus-seed pathosystem. Mol. Microbiol. 67:378–391.
5. Calvo, A. M., J. W. Bok, W. Brooks, and N. P. Keller. 2004. VeA is requiredfor toxin and sclerotial production in Aspergillus parasiticus. Appl. Environ.Microbiol. 70:4733–4739.
6. Calvo, A. M., H. W. Gardner, and N. P. Keller. 2001. Genetic connectionbetween fatty acid metabolism and sporulation in Aspergillus nidulans.J. Biol. Chem. 276:25766–25774.
7. Calvo, A. M., L. L. Hinze, H. W. Gardner, and N. P. Keller. 1999. Sporogeniceffect of polyunsaturated fatty acids on development of Aspergillus spp. Appl.Environ. Microbial. 65:3668–3673.
8. Calvo, A. M., R. A. Wilson, J. W. Bok, and N. P. Keller. 2002. Relationshipbetween secondary metabolism and fungal development. Microbiol. Mol.Biol. Rev. 66:447–459.
9. Champe, S. P., and A. A. El-Zayat. 1989. Isolation of a sexual sporulationhormone from Aspergillus nidulans. J. Bacteriol. 171:3982–3988.
10. Chen, H., and G. R. Fink. 2006. Feedback control of morphogenesis in fungiby aromatic alcohols. Genes Dev. 20:1150–1161.
11. Chen, H., M. Fujita, Q. Feng, J. Clardy, and G. R. Fink. 2004. Tyrosol is aquorum-sensing molecule in Candida albicans. Proc. Natl. Acad. Sci. USA101:5048–5052.
12. Chenna, R., H. Sugawara, T. Koike, R. Lopez, T. J. Gibson, D. G. Higgins,and J. D. Thompson. 2003. Multiple sequence alignment with the Clustalseries of programs. Nucleic Acids Res. 31:3497–3500.
13. Diener, U. L., R. J. Cole, T. H. Sanders, G. A. Payne, L. S. Lee, and M. A.Klich. 1987. Epidemiology of aflatoxin formation by Aspergillus flavus. Annu.Rev. Phytopathol. 25:249–270.
14. Erb-Downward, J. R., and G. B. Huffnagle. 2006. Role of oxylipins and otherlipid mediators in fungal pathogenesis. Future Microbiol. 1:219–227.
15. Fuqua, W. C., S. C. Winans, and E. P. Greenberg. 1994. Quorum sensing inbacteria: the LuxR-LuxI family of cell density-responsive transcriptionalregulators. J. Bacteriol. 176:269–275.
16. Geiser, D. M., W. E. Timberlake, and M. L. Arnold. 1996. Loss of meiosis inAspergillus. Mol. Biol. Evol. 13:809–817.
17. Gillmor, S. A., A. Villasenor, R. Fletterick, E. Sigal, and M. F. Browner.1997. The structure of mammalian 15-lipoxygenase reveals similarity to thelipases and the determinants of substrate specificity. Nat. Struct. Biol.4:1003–1009.
18. Gottwald, T. R., and B. W. Wood. 1984. The effect of fatty acids on growthand sporulation of the pecan scab fungus Cladosporium caryigenum. Myco-logia 76:326–331.
19. Greene, A. V., N. P. Keller, H. Haas, and D. Bell-Pedersen. 2003. A circadianoscillator in Aspergillus spp. regulates daily development and gene expres-sion. Eukaryot. Cell 2:231–237.
20. Hammond, T. M., and N. P. Keller. 2005. RNA silencing in Aspergillusnidulans is independent of RNA dependent RNA polymerases. Genetics169:607–617.
21. He, Z. M., M. S. Price, G. R. Obrian, D. R. Georgianna, and G. A. Payne.2007. Improved protocols for functional analysis in the pathogenic fungusAspergillus flavus. BMC Microbiol. 7:104.
22. Hedayati, M. T., A. C. Pasqualotto, P. A. Warn, P. Bowyer, and D. W.Denning. 2007. Aspergillus flavus: human pathogen, allergen and mycotoxinproducer. Microbiology 153:1677–1692.
23. Hegg, E. L., and L. Que, Jr. 1997. The 2-His-1-carboxylate facial triad—anemerging structural motif in mononuclear non-heme iron(II) enzymes. Eur.J. Biochem. 250:625–629.
24. Herman, R. P. 1998. Oxylipin production and action in fungi and relatedorganisms, p. 115–132. In A. F. Rowley, H. Kuhn, and T. Schewe (ed.),Eicosanoids and related compounds in plants and animals. Princeton Uni-versity Press, Princeton, NJ.
25. Hogan, D. A. 2006. Talking to themselves: autoregulation and quorum sens-ing in fungi. Eukaryot. Cell 5:613–619.
26. Hornby, J. M., S. M. Jacobitz-Kizzier, D. J. McNeel, E. C. Jensen, D. S.Treves, and K. W. Nickerson. 2004. Inoculum size effect in dimorphic fungi:extracellular control of yeast-mycelium dimorphism in Ceratocystis ulmi.Appl. Environ. Microbiol. 70:1356–1359.
27. Hughes, R. K., D. M. Lawson, A. R. Hornostaj, S. A. Fairhurst, and R. Casey.2001. Mutagenesis and modeling of linoleate-binding to pea seed lipoxygen-ase. Eur. J. Biochem. 268:1030–1040.
28. Jensen, E. C., C. Ogg, and K. W. Nickerson. 1992. Lipoxygenase inhibitors
shift the yeast/mycelium dimorphism in Ceratocystis ulmi. Appl. Environ.Microbiol. 58:2505–2508.
29. Klose, J., M. M. Moniz de Sa, and J. W. Kronstad. 2004. Lipid-inducedfilamentous growth in Ustilago maydis. Mol. Microbiol. 52:823–835.
30. Kock, J. L., C. J. Strauss, C. H. Pohl, and S. Nigam. 2003. The distributionof 3-hydroxy oxylipins in fungi. Prostaglandins Other Lipid Mediat. 71:85–96.
31. Kolbinger, A., T. Gao, D. Brock, R. Ammann, A. Kisters, J. Kellermann, D.Hatton, R. H. Gomer, and B. Wetterauer. 2005. A cysteine-rich extracellularprotein containing a PA14 domain mediates quorum sensing in Dictyosteliumdiscoideum. Eukaryot. Cell 4:991–998.
32. Kugler, S., T. Schurtz Sebghati, L. Groppe Eissenberg, and W. E. Goldman.2000. Phenotypic variation and intracellular parasitism by Histoplasma cap-sulatum. Proc. Natl. Acad. Sci. USA 97:8794–8798.
33. Kuribayashi, T., H. Kaise, C. Uno, T. Hara, T. Hayakawa, and T. Joh. 2002.Purification and characterization of lipoxygenase from Pleurotus ostreatus. J.Agric. Food Chem. 50:1247–1253.
34. Liavonchanka, A., and I. Feussner. 2006. Lipoxygenases: occurrence, func-tions and catalysis. J. Plant Physiol. 163:348–357.
35. Maggio-Hall, L. A., R. A. Wilson, and N. P. Keller. 2005. Fundamentalcontribution of beta-oxidation to polyketide mycotoxin production in planta.Mol. Plant Microbe Interact. 18:783–793.
36. Miller, B. L., K. Y. Miller, and W. E. Timberlake. 1985. Direct and indirectgene replacements in Aspergillus nidulans. Mol. Cell. Biol. 5:1714–1721.
37. Mooney, J. L., and L. N. Yager. 1990. Light is required for conidiation inAspergillus nidulans. Genes Dev. 4:1473–1482.
38. Nickerson, K. W., A. L. Atkin, and J. M. Hornby. 2006. Quorum sensing indimorphic fungi: farnesol and beyond. Appl. Environ. Microbiol. 72:3805–3813.
39. Noverr, M. C., J. R. Erb-Downward, and G. B. Huffnagle. 2003. Productionof eicosanoids and other oxylipins by pathogenic eukaryotic microbes. Clin.Microbiol. Rev. 16:517–533.
40. Nukima, M., T. Sassa, M. Ikeda, and K. Takahashi. 1981. Linoleic acidenhances perithecial production in Neurospora crassa. Agric. Biol. Chem.45:2371–2373.
41. Oh, K. B., H. Miyazawa, T. Naito, and H. Matsuoka. 2001. Purification andcharacterization of an autoregulatory substance capable of regulating themorphological transition in Candida albicans. Proc. Natl. Acad. Sci. USA98:4664–4668.
42. Reference deleted.43. Perez-Gilabert, M., I. Sanchez-Felipe, A. Morte, and F. Garcia-Carmona.
2005. Kinetic properties of lipoxygenase from desert truffle (Terfezia claveryiChatin) ascocarps: effect of inhibitors and activators. J. Agric. Food Chem.53:6140–6145.
44. Ramage, G., S. P. Saville, B. L. Wickes, and J. L. Lopez-Ribot. 2002. Inhi-bition of Candida albicans biofilm formation by farnesol, a quorum-sensingmolecule. Appl. Environ. Microbiol. 68:5459–5463.
45. Roeder, P. E., M. L. Sargent, and S. Brody. 1982. Circadian rhythms inNeurospora crassa: oscillations in fatty acids. Biochemistry 21:4909–4916.
46. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
47. Shimizu, K., and N. P. Keller. 2001. Genetic involvement of a cAMP-dependent protein kinase in a G protein signaling pathway regulating mor-phological and chemical transitions in Aspergillus nidulans. Genetics 157:591–600.
48. Skrzypczak-Jankun, E., L. M. Amzel, B. A. Kroa, and M. O. Funk, Jr. 1997.Structure of soybean lipoxygenase L3 and a comparison with its L1 isoen-zyme. Proteins 29:15–31.
49. Su, C., and E. H. Oliw. 1998. Manganese lipoxygenase. Purification andcharacterization. J. Biol. Chem. 273:13072–13079.
50. Tsitsigiannis, D. I., and N. P. Keller. 2007. Oxylipins as developmental andhost-fungal communication signals. Trends Microbiol. 15:109–118.
51. Tsitsigiannis, D. I., T. M. Kowieski, R. Zarnowski, and N. P. Keller. 2004.Endogenous lipogenic regulators of spore balance in Aspergillus nidulans.Eukaryot. Cell 3:1398–1411.
52. Tsitsigiannis, D. I., T. M. Kowieski, R. Zarnowski, and N. P. Keller. 2005.Three putative oxylipin biosynthetic genes integrate sexual and asexual de-velopment in Aspergillus nidulans. Microbiology 151:1809–1821.
53. Wicklow, D. T., B. W. Horn, and R. J. Cole. 1982. Sclerotium production byAspergillus flavus on corn kernels. Mycologia 74:398–403.
VOL. 74, 2008 QUORUM SENSING IN A. FLAVUS 5685




















