
Purification and characterization of an alkaline pectin lyase from Aspergillus flavus
9
1 3 Eur Food Res Technol (2014) 239:127–135 DOI 10.1007/s00217-014-2198-8 ORIGINAL PAPER Purification and characterization of an alkaline pectin lyase produced by a newly isolated Brevibacillus borstelensis (P35) and its applications in fruit juice and oil extraction Nazan Demir · Hayrunnisa Nadaroglu · Yasar Demir · Ceyhun Isik · Esen Taskin · Ahmet Adiguzel · Medine Gulluce Received: 7 January 2014 / Revised: 6 March 2014 / Accepted: 10 March 2014 / Published online: 29 March 2014 © Springer-Verlag Berlin Heidelberg 2014 the purified enzyme. In the course of the laboratory trials, it was demonstrated that PNL from B. borstelensis (P35) could be successfully applied to the production and clarifi- cation of fruit juice and oil extraction. Keywords Pectin lyase · Purification · Brevibacillus borstelensis (P35) · Pectin · Juice clarification · Oil extraction Introduction Pectic polymers (chain of 1,4-linked-α-D-galacturonic acid and methoxylated derivatives), an important cell wall component in dicotyledonous plants, are a heteropolysac- charide composed of α-1,4-linked galacturonate chains. Enzymes that hydrolyze pectic substances are known as pectinases [1, 2]. Pectinases are the group of enzymes involved in depolymerisation of the pectic polymers. This group of enzymes consists of pectin esterases (PEs) (E.C 3.1.1.11), polygalacturonases (PGs) (E.C 3.2.1.15), pectate lyases (PLs) (E.C 4.2.2.2) and pectin lyases (PNLs) (E.C 4.2.2.10) [3]. Pectinases are biotechnologically important because they have potential applications in clarification of fruit juices, textile and plant fiber processing, coffee and tea fermentation, virus purification, treatment of industrial wastewater containing pectinacious materials, and paper manufacturing [4, 5]. There are studies related to PNLs from microorganisms but there are scanty reports of their presence in plants and animals also [6]. PNLs are mainly produced by fungal gen- era Aspergillus, Penicillium and Fusarium but there are a few reports on bacterial and yeast PNLs. In recent years, thermophilic bacteria have been a great center of interest due to their thermostable enzymes and Abstract An alkaline pectin lyase (PNL) (EC 4.2.2.10) secreted by Brevibacillus borstelensis P35 (GenBank Number: FJ417406) was purified using ammonium sul- fate fractionation, anion exchange chromatography on DEAE-cellulose and gel filtration chromatography on Sephadex G-150. The pH and temperature optima of the enzyme were found to be 8.0 and 60 °C. The enzyme does not loose activity up to 60 °C if exposed for 1 h. The val- ues of K m and V max of the enzyme were 0.625 mg/mL and 126.32 s −1 , respectively. The molecular weight was found to be 36 ± 01 kDa. The presence of 10 mM concentration of Ca 2+ , Cu 2+ , Mn 2+ , Mg 2+ , Zn 2+ , Hg 2+ , Fe 2+ and EDTA, L-cystein, ascorbic acid significantly enhanced the PNL of N. Demir (*) · Y. Demir · C. Isik Department of Chemistry, Faculty of Sciences, Mugla University, 48000 Mugla, Turkey e-mail: [email protected] H. Nadaroglu Department of Food Technology, Erzurum Vocational Training School, Ataturk University, 25240 Erzurum, Turkey e-mail: [email protected] Y. Demir Department of Chemistry, Faculty of Science, University of Mehmet Akif Ersoy, 15000 Burdur, Turkey E. Taskin Faculty of Education, Elementary Science Education, Bayburt University, 69100 Bayburt, Turkey A. Adiguzel Department of Molecular Biology and Genetic, Faculty of Science, Ataturk University, 25240 Erzurum, Turkey M. Gulluce Department of Biology, Faculty of Arts and Sciences, Ataturk University, 25240 Erzurum, Turkey
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Purification and characterization of an alkaline pectin lyase from Aspergillus flavus

1 3
Eur Food Res Technol (2014) 239:127–135DOI 10.1007/s00217-014-2198-8
ORIgInal PaPER
Purification and characterization of an alkaline pectin lyase produced by a newly isolated Brevibacillus borstelensis (P35) and its applications in fruit juice and oil extraction
Nazan Demir · Hayrunnisa Nadaroglu · Yasar Demir · Ceyhun Isik · Esen Taskin · Ahmet Adiguzel · Medine Gulluce
Received: 7 January 2014 / Revised: 6 March 2014 / accepted: 10 March 2014 / Published online: 29 March 2014 © Springer-Verlag Berlin Heidelberg 2014
the purified enzyme. In the course of the laboratory trials, it was demonstrated that Pnl from B. borstelensis (P35) could be successfully applied to the production and clarifi-cation of fruit juice and oil extraction.
Keywords Pectin lyase · Purification · Brevibacillus borstelensis (P35) · Pectin · Juice clarification · Oil extraction
Introduction
Pectic polymers (chain of 1,4-linked-α-d-galacturonic acid and methoxylated derivatives), an important cell wall component in dicotyledonous plants, are a heteropolysac-charide composed of α-1,4-linked galacturonate chains. Enzymes that hydrolyze pectic substances are known as pectinases [1, 2]. Pectinases are the group of enzymes involved in depolymerisation of the pectic polymers. This group of enzymes consists of pectin esterases (PEs) (E.C 3.1.1.11), polygalacturonases (Pgs) (E.C 3.2.1.15), pectate lyases (Pls) (E.C 4.2.2.2) and pectin lyases (Pnls) (E.C 4.2.2.10) [3]. Pectinases are biotechnologically important because they have potential applications in clarification of fruit juices, textile and plant fiber processing, coffee and tea fermentation, virus purification, treatment of industrial wastewater containing pectinacious materials, and paper manufacturing [4, 5].
There are studies related to Pnls from microorganisms but there are scanty reports of their presence in plants and animals also [6]. Pnls are mainly produced by fungal gen-era Aspergillus, Penicillium and Fusarium but there are a few reports on bacterial and yeast Pnls.
In recent years, thermophilic bacteria have been a great center of interest due to their thermostable enzymes and
Abstract an alkaline pectin lyase (Pnl) (EC 4.2.2.10) secreted by Brevibacillus borstelensis P35 (genBank number: FJ417406) was purified using ammonium sul-fate fractionation, anion exchange chromatography on DEaE-cellulose and gel filtration chromatography on Sephadex g-150. The pH and temperature optima of the enzyme were found to be 8.0 and 60 °C. The enzyme does not loose activity up to 60 °C if exposed for 1 h. The val-ues of Km and Vmax of the enzyme were 0.625 mg/ml and 126.32 s−1, respectively. The molecular weight was found to be 36 ± 01 kDa. The presence of 10 mM concentration of Ca2+, Cu2+, Mn2+, Mg2+, Zn2+, Hg2+, Fe2+ and EDTa, l-cystein, ascorbic acid significantly enhanced the Pnl of
n. Demir (*) · Y. Demir · C. Isik Department of Chemistry, Faculty of Sciences, Mugla University, 48000 Mugla, Turkeye-mail: [email protected]
H. nadaroglu Department of Food Technology, Erzurum Vocational Training School, ataturk University, 25240 Erzurum, Turkeye-mail: [email protected]
Y. Demir Department of Chemistry, Faculty of Science, University of Mehmet akif Ersoy, 15000 Burdur, Turkey
E. Taskin Faculty of Education, Elementary Science Education, Bayburt University, 69100 Bayburt, Turkey
a. adiguzel Department of Molecular Biology and genetic, Faculty of Science, ataturk University, 25240 Erzurum, Turkey
M. gulluce Department of Biology, Faculty of arts and Sciences, ataturk University, 25240 Erzurum, Turkey

128 Eur Food Res Technol (2014) 239:127–135
1 3
industrial importance [7, 8]. The study was carried out in order to determine the phenotypic and genotypic charac-terization of Brevibacillus borstelensis (P35) isolated from Van City Ercis Town Hasanabdal hot spring water, Turkey [9]. To verify the systematic position of B. borstelensis (P35), a study of morphological and physiological char-acteristics, cellular fatty acids analysis (FaMEs), rep-PCR (REP and BOX-PCR) and 16S rRna sequence analysis were undertaken by Savas et al. [9].
In this paper, we report the production of extra cellular Pnls (E.C. 4.2.2.10) produced by B. borstelensis (P35) using solid-state fermentation. Pnl from B. borstelensis (P35) was purified and characterized. We also determined the action of Pnl in fruit juice and oil extraction. This is the first report on the production, purification and charac-terization of a Pnl from B. borstelensis (P35).
Materials and methods
Materials
DEaE-cellulose, pectin (ED 9.4 %) from citrus fruit, dithiothreitol (DTT), thiobarbituric acid (TBa), Sepha-dex g150, the electrophoresis equipment and reagents were purchased from Sigma Chemical Co. (St. louis, MO, USa). pgEM-T vector system was maintained from Promega Corp., Southampton, United Kingdom, and aBI PRISM cycle sequencing kit was maintained from Macro-gen, Korea. The other reagents were of analytical grade.
Isolation of the thermophilic bacteria
The water sample was taken from Van City Ercis Town Hasanabdal hot spring water, Turkey. after the collec-tion, it (1 ml) was first inoculated into 10 ml of nutrient broth (nB) medium at 55–60 °C for 24–48 h, with shaken at 150 rpm. Then, by enrichment culturing 3–4 phases was transferred into nutrient agar (na)-containing Petri dishes; colonies that were thought to be different than each other were purified. The purity of the test isolate was assessed using colony morphology and microscopy. Isolated and purified bacterial strains were stored in nutrient broth con-taining 15 % glycerol at −86 °C for further studies [9].
Biochemical and physiological characterizations of the test strain
The temperature range for growth was determined by incu-bating the isolate from 30 to 85 °C. The effect of naCl on the thermophilic bacterial growth was studied in nB medium containing 2.0, 3.0, 4.0, 5.0, 8.0, 10.0 % (w/v) naCl. The pH dependence of growth was tested in the pH
range of 4.0–11.0 in nutrient broth medium. The gram reac-tion was investigated in the presence of catalase, oxidase and nitrate reduction according to the methods described by Fahy and Hayward [10].
Extraction of bacterial Dna from pure culture
as previously stated by adiguzel [11], the extraction of the total genomic Dna from bacteria sample was performed through applying a modified method.
PCR amplification, cloning and sequencing of 16S rRna
The 16S rRna (1,400 bp) gene was amplified from puri-fied genomic Dna with primers UnI16S-l (5′-aTTCTa gagTTTgaTCaTggCTTCa) corresponded to positions 11–26 of the Escherichia coli 16S rRna and the reverse primer, UnI16S-R (5′-aTggTaCCgTgTgaCgggCg gTgTTgTa), corresponded to the complement of posi-tions 1411–1393 of E. coli 16S rRna. amplicons were cloned into a pgEM-T vector system, and 16S rRna gene sequences were determined with applied Biosystems model 373a Dna sequencer using the aBI PRISM cycle sequencing kit [12, 13].
Production of pectic enzyme by solid-state fermentation
The solid-state fermentation medium described by Tara-gano and Pilosof [14] consisted of 5 g wheat bran, 1 % citrus pectin (DE 9.4 %, Sigma Chemical Co) and 10 ml salt mixture that composed of 0.14 % (nH4)2SO4, 0.02 % MgCl2 and 0.02 % K2HPO4 was used to inoculate Erlen-meyer flasks (250 ml). The final moisture content of the medium was approximately as 67 %. The flasks were steri-lized for 20 min at 115 °C, inoculated and then incubated at 50 °C for 3 days. The fermented material was mixed with 60 ml of 1 % naCl and centrifuged (4 °C, 10,000 rpm and 30 min). The Pnl enzyme activity was measured into supernatant [14].
Pectin lyase purification
The crude extract was produced from B. borstelensis (P35) and dialyzed against the sodium phosphate buffer (50 mM, pH 8.0).
Purification of pectin lyase enzyme
The crude extract was centrifuged, and precipitates were removed. For the purification of the Pnl enzyme, the fol-lowing procedure was implemented [4, 5].
Pectin lyase was purified from the crude extract in three steps. Firstly, it was partially purified by precipitation % in

129Eur Food Res Technol (2014) 239:127–135
1 3
(nH4)2SO4. Secondly, ion exchange chromatography on DEaE-Sephadex was used, and lastly, Sephacryl S 200 was used.
The collapse of (nH4)2SO4 was done from 0 to 90 % in supernatant with the internals of 0–10, 10–20, 20–30, 30–40, 40–50, 50–60, 60–70, 70–80 and 80–90. Signifi-cant activity was not observed below at a range 0–60 % (nH4)2SO4. The majority of activity was found in the 60–80 % precipitate.
Solid (nH4)2SO4 was added to the supernatant to increase the concentration of (nH4)2SO4 from 60 % the fraction to 80 %. after mixing it in an ice-bath for 1 h with magnetic stirring, it was centrifuged (10.000×g, 30 min, and 4 °C). The supernatant was discarded, and the precipi-tate was dissolved in the sodium phosphate buffer (50 mM, pH 8.0), and dialyzed against the same buffer.
Ion exchange chromatography
The dialyzed solution was subjected to an anion exchange chromatography using a DEaE-cellulose fast flow col-umn. This column size was 3.0 × 70 cm and bed volume 120 ml. Column was washed with a 300-ml sodium phos-phate buffer (50 mM, pH 8.0). Then, the bound proteins were eluted with a sodium phosphate buffer (50 mM, pH 8.0), using a linear gradient of 0–0.4 M naCl. Eluates were taken into tubes at the 15 ml/h flow rate. Protein elution was monitored spectrophotometrically by measuring the absorbance at 280 nm. activity was measured using pectin from citrus fruits as the assay substrate (see below). after dialyzing the eluents against 50 mM phosphate buffer (pH 8.0), the eluents were collected. The fractions showing Pnl activity were pooled, concentrated and saved for fur-ther analysis [15].
gel filtration chromatography
a glass column was packed with Sephacryl S 200 (35 × 1.5 cm, bed volume 60 ml). The concentrated sam-ple was loaded onto this column, and the elution of the pro-teins was done using Tris–HCl buffer (50 mM, pH 8). Frac-tions were collected at 3 ml volume, and the absorbance of the samples for protein content and Pnl activity was found by the methods described earlier. The Pnl-positive frac-tions were pooled. Then, purified enzyme was lyophilized for obtained concentrated enzyme and saved for further analysis [15].
Pectin lyase activity
The principle of the activity determination is based on measuring the absorption of colored derivatives which was obtained by reaction of unsaturated uronic acid ester and
thiobarbituric acid at 550 nm [16]. Briefly, enzyme solu-tion was incubated with pectin from citrus fruits (1 %, w/v) previously prepared in 50 mM sodium phosphate buffer, pH 8.0 at 60 °C for 30 min. after the addition of 50 μl of naOH (1 n), the mixture was shaken, then heated at 80 °C in a water bath for 5 min and cooled. after the addition of 600 μl of HCl (1 n) to acidify the medium, the yellow color disappeared.
When HCl was added to reaction mixture, yellow color was disappeared because of acidic medium. The solution was then shaken and was finally added 500 μl of a solution (0.04 M) of thiobarbituric acid. The tube was incubated a second time at 80 °C for 5 min in a water bath. The solu-tion was briefly cooled in an ice-water bath, and absorb-ance was measured at 550 nm. Spectrophotometry was performed on a Spekol 13000 spectrophotometer (analytic jena ag, germany) with 1-cm quartz cells.
Protein concentration
Protein concentration was determined spectrophotometri-cally (absorbance at 280 nm) as well as by Bradford’s method [17], using bovine serum albumin (BSa) as the standard.
SDS polyacrylamide gel electrophoresis
SDS-PagE was performed using 3 and 10 % acrylamide concentrations for the stacking and running gels, respec-tively, each of them containing 0.1 % SDS as described by laemmli to check the homogeneity of the enzyme and determine the molecular weight [18]. a 20 μg of sample was applied to the electrophoresis medium. gels were stained for 1.5 h in 0.1 % Coomassie brilliant blue R-250 in 50 % methanol and 10 % acetic acid and then destained with several changes of the same solvent without the dye. Standard protein was consisted of alcohol dehydrogenase, 150 kDa; muscle phosphorylase b, 97.4 kDa; bovine serum albumin, 66 kDa; egg ovalbumin, 45 kDa; and carbonic anhydrase, 29 kDa. The electrophoretic pattern was photo-graphed (Fig. 3).
Molecular weight determination by gel filtration
a molecular weight column (3 × 70 cm) was prepared with Sephacryl S-100. The column was equilibrated with the buffer (0.05 M na2HPO4, 1 mM DTT, pH 7) until the absorbance was zero at 280 nm. The standard pro-tein solution (bovine serum albumin, 66 kDa; egg oval-bumin, 45 kDa; pepsin, 34 kDa; trypsinogen, 24 kDa; β-lactoglobulin and lysozyme, 14 kDa) was added to the column. The purified Pnl enzyme was added into the col-umn separately and then eluted under the same conditions.

130 Eur Food Res Technol (2014) 239:127–135
1 3
The flow rate through the column was 20 ml/h. The elution volume was compared with standard proteins [19].
Optimum temperature
The optimum temperature was determined by carrying out the enzyme assay in buffer (50 mM sodium phosphate buffer, pH 8) at 10–90 °C, respectively, for 5 min for Pnl activity. In each case, the substrate was preincubated at the different temperatures.
Optimum pH
The enzyme activity was assayed with 0.05 M acetate (pH 4–5), 0.05 M phosphate (pH 6–8) and 0.05 M carbonate buffer (pH 9–11, for pH values of 2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0 and 11.0, respectively).
Thermal stability
Thermal stability was also determined as described for 1–24 h time periods. Enzyme samples were taken periodi-cally and assayed to determine the residual enzyme activity using phosphate buffer (50 mM sodium phosphate buffer, pH 8) as described above.
Effect of various metal ions on the pectin lyase activity
To determine whether the effects of various metal ions (10 mM Cu2+, Mg2+, Hg2+, Fe2+ and Zn2+) on the Pnl activity were measured at 5 mM concentrations of them. Each inhibitor solution was prepared at 100 mM concentra-tion, and 50, 100, 150, 200 or 250 μl of each solution was added in a cuvette-containing 250 μl of enzyme solution. Its total volume was adjusted to 750 μl with a buffer solu-tion. a control assay of the purified enzyme activity was measured without inhibitors, and the resulting activity was taken as 100 %. The effect of each agent was determined by measuring the enzyme activity using pectin from citrus fruits as a substrate [16].
Production of fruit juice
Peach, cherry, grape, apple, strawberry and orange were used for preparing fruit juice. The fruits were firstly care-fully washed, then dried in air and homogenized. To 10 g of fruit sample homogenate, 2 ml of enzyme solution was added (in control experiment, 2 ml of distilled water was added). The same experiment was done using 2 ml of crude extract of B. borstelensis (P35). Fruit homogenates were treated with enzyme for 1 h, at 60 °C and at a natural pH value. In the course of the experiment, a volume of the juice was filtered through a paper filter for 15 min using
vacuum. Pectin content was hydrolyzed in fruits. Then, the volume of fruit juice was measured, and yield of juice was calculated [20].
Oil extraction
Pumpkin seed, nut, sun flower, ground nut, corn and olive were used for oil extraction. The oil seeds were firstly care-fully washed, then dried in air and homogenized. To 10 g of oil seeds sample homogenate, 2 ml of enzyme solu-tion was added (in control experiment, 10 ml of distilled water was added) with a mixing rate of 150 rpm for 1 h, at 60 °C and at pH 8.0 value, and the increase in oil yield was measured as compared to control (including without enzyme). The same experiment was done using 2 ml of crude homogenate [21]. When pectin structure of oil seed was hydrolyzed, oil flow cloud be facilitated from seed.
Statistical analysis
all of the tests were conducted in triplicate for determina-tion of the Pnl activities of samples. Data were expressed as means ± standard errors. Statistical analyses were performed using SPSS version 10.0 software (SPSS Inc., Chicago, Il., USa), and the significant differences were determined with a 95 % confidence interval (p < 0.05) using Tukey’s test.
Results and discussion
Isolation and identification of the test isolate
To verify the systematic position of this bacterium, a study of morphological, biochemical and physiological charac-teristics and 16S rRna analysis were undertaken.
The isolate (P35) was gram, catalase, oxidase, nitrate reduction-positive and endospore forming, mobile rod, and amylase negative from the water sample of Van City Ercis Town Hasanabdal hot spring water in the provinces of Erzurum, Turkey.
as a result of pH and salt tests, it was determined that for P35 isolate, the optimum pH is 7.5–8.5, and all of this bacteria propagate well in the range of salt concentration of 2–5 %. at the end of temperature tests, it was determined that for all of the isolate, the optimum temperature is 56 °C. These met the criteria of thermophilic bacteria, which grow at temperatures above 50 °C [22].
The 16S rRna gene sequence determined for strain P35 was about 1,398 nucleotides long. afterward, as the result of the blast study performed by utilizing the database in the genbank, the test isolate (genbank numbers of P35 is FJ417406) was ascertained to bear a resemblance to B. bor-stelensis at a powerful rate of (≥99 %).
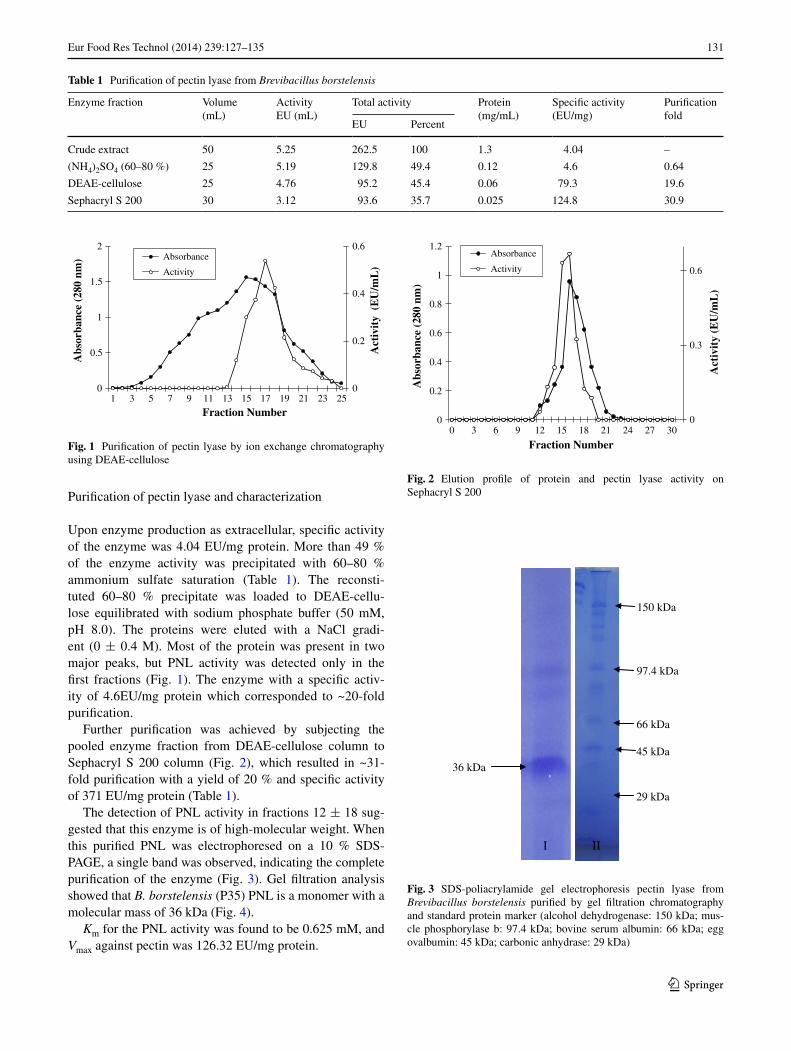
131Eur Food Res Technol (2014) 239:127–135
1 3
Purification of pectin lyase and characterization
Upon enzyme production as extracellular, specific activity of the enzyme was 4.04 EU/mg protein. More than 49 % of the enzyme activity was precipitated with 60–80 % ammonium sulfate saturation (Table 1). The reconsti-tuted 60–80 % precipitate was loaded to DEaE-cellu-lose equilibrated with sodium phosphate buffer (50 mM, pH 8.0). The proteins were eluted with a naCl gradi-ent (0 ± 0.4 M). Most of the protein was present in two major peaks, but Pnl activity was detected only in the first fractions (Fig. 1). The enzyme with a specific activ-ity of 4.6EU/mg protein which corresponded to ~20-fold purification.
Further purification was achieved by subjecting the pooled enzyme fraction from DEaE-cellulose column to Sephacryl S 200 column (Fig. 2), which resulted in ~31-fold purification with a yield of 20 % and specific activity of 371 EU/mg protein (Table 1).
The detection of Pnl activity in fractions 12 ± 18 sug-gested that this enzyme is of high-molecular weight. When this purified Pnl was electrophoresed on a 10 % SDS-PagE, a single band was observed, indicating the complete purification of the enzyme (Fig. 3). gel filtration analysis showed that B. borstelensis (P35) Pnl is a monomer with a molecular mass of 36 kDa (Fig. 4).
Km for the Pnl activity was found to be 0.625 mM, and Vmax against pectin was 126.32 EU/mg protein.
Table 1 Purification of pectin lyase from Brevibacillus borstelensis
Enzyme fraction Volume (ml)
activity EU (ml)
Total activity Protein (mg/ml)
Specific activity (EU/mg)
Purification fold
EU Percent
Crude extract 50 5.25 262.5 100 1.3 4.04 –
(nH4)2SO4 (60–80 %) 25 5.19 129.8 49.4 0.12 4.6 0.64
DEaE-cellulose 25 4.76 95.2 45.4 0.06 79.3 19.6
Sephacryl S 200 30 3.12 93.6 35.7 0.025 124.8 30.9
0
0.2
0.4
0.6
0
0.5
1
1.5
2
1 3 5 7 9 11 13 15 17 19 21 23 25
Act
ivit
y (
EU
/mL
)
Abs
orba
nce
(280
nm
)
Fraction Number
Absorbance
Activity
Fig. 1 Purification of pectin lyase by ion exchange chromatography using DEaE-cellulose
0
0.3
0.6
0
0.2
0.4
0.6
0.8
1
1.2
0 3 6 9 12 15 18 21 24 27 30
Act
ivit
y (E
U/m
L)
Abs
orba
nce
(280
nm
)
Fraction Number
Absorbance
Activity
Fig. 2 Elution profile of protein and pectin lyase activity on Sephacryl S 200
I II
29 kDa
45 kDa
66 kDa
97.4 kDa
150 kDa
36 kDa
Fig. 3 SDS-poliacrylamide gel electrophoresis pectin lyase from Brevibacillus borstelensis purified by gel filtration chromatography and standard protein marker (alcohol dehydrogenase: 150 kDa; mus-cle phosphorylase b: 97.4 kDa; bovine serum albumin: 66 kDa; egg ovalbumin: 45 kDa; carbonic anhydrase: 29 kDa)

132 Eur Food Res Technol (2014) 239:127–135
1 3
Effect of temperature on pectin lyase activity
The enzyme displayed maximal activity at 60 °C (Fig. 5), which is lower than for Thermoascus aurantiacus Pnl that show the highest enzymatic activity at around 65 °C [23, 24]. Increased catalytic activity at higher temperatures, e.g., 50 °C, is industrial significant because activity is not lost due to ther-mal stability during 24 h in a very long incubation time [4, 5]. In previously studies, a lot of Pnl was denaturated to the tertiary structure of enzyme at evaluated temperatures [25]. When this study was consistent with previously studies, it was obtained that this purified Pnl was very thermal stable.
Effect of pH on pectin lyase activity
The optimum pH for Pnl activity was 8.0 (Fig. 6), and the enzyme retained over 50 % of the original activity between pH 7.0 and 9. This pH optimum is similar to that previously determined for Pnl from Pythium splendens, Penicillium oxalicum, Cystofilobasidium capitatum and Erwinia aroideae [25–28]. Durability of the enzyme in alkaline pH is an advan-tage in the textile industries. They are used for retting of plant fibers such as ramie, sunn hemp, jute and flax [29–32].
Thermal stability of the purified Pnl enzyme from B. borstelensis was examined in the temperature range of 40–80 °C (Fig. 7). The high optimum temperature of puri-fied Pnl enzyme shows thermal stability of this enzyme. although the enzyme suffered deactivation at higher temperatures, the enzyme still retained about 10 % of its activity at 80 °C after 60 min. The thermal stability of the enzyme indicates excellence of the enzyme for the food, leather, pharmaceutical, detergent industries.
Effect of inhibitors on activity of pectin lyase
among the common enzyme modifiers, tested compounds such as mercury chloride, manganese chloride, copper (II) chloride, iron (II) chloride and barium chloride showed
significantly inhibition for Pnl activity (Table 2). In cont-rast, zinc chloride had no effect on enzyme activity at 5 mM concentration. B. borstelensis Pnl was significantly
y = -0.0199x + 1.915R² = 0.9868
0
0.4
0.8
1.2
1.6
2
0 10 20 30 40 50
Log
MA
Elution Volume (mL)
36 kDa
Fig. 4 Determination of molecular mass by gel filtration
0
0.2
0.4
0.6
0.8
1
1.2
1.4
20 40 60 80 100
Act
ivit
y (E
U/m
L)
Temperature (oC)
Fig. 5 Effect of temperature on the activity of purified pectin lyase from Brevibacillus borstelensis
0
0.5
1
1.5
2
2.5
2 4 6 8 10 12
Act
ivit
y (E
U)
pH
Fig. 6 Effect of pH on the activity of purified pectin lyase from Brev-ibacillus borstelensis
0
20
40
60
80
100
120
0 1 2 3 4 5 6 7 9 11 24
Act
ivity
(E
U)
Time (h)
40 50 60 70 80
Fig. 7 Temperature stability of the purified pectin lyase from Brevi-bacillus borstelensis

133Eur Food Res Technol (2014) 239:127–135
1 3
activated by calcium chloride and magnesium chloride. Similar results have already been reported using various organisms [33–37].
Production of fruit juice
The turbidity and viscosity of the raw juice are mainly because of fruit with high pectin content; therefore, pectin degrading Pnl enzyme has been exploited for clarification of fruit juices, increasing yield and quality characteristics of fruit juices. The results of experiments were applied in yielding of fruit juice from peach, cherry, grape, apple, strawberry and orange are shown in Table 3. It was shown
that the use of purified Pnl increases the juice output and dramatically reduces its viscosity. Pnl treatment for 1 h resulted in almost twofold enhancement in the increas-ing volume of strawberry juice (7–12 ml), grape juice (15–21.5 ml) and apple juice (12–18.5 ml). Enzyme also increased the volume of peach juice (9–10.5 ml), cherry juice (8–10 ml) and orange juice (9–10) in yield of 116.7, 125 and 111.1 %, respectively, as compared to control. The same experiments were performed using crude extract. However, it was determined to be less effective crude extract than purified enzyme in the clarification of fruit juices (Table 3). Compared with control (untreated orange juice) after 5 h of incubation, Demir et al. [4] reported that Pnl treatment of orange, apple and peach juices resulted in the yield of 133, 109 and 105 %, respectively. nadaro-glu et al. [5] reported a clarification yield of 108 and 118 % in the apple and orange juices treated by Pnl induced, respectively, after 5 h of incubation compared with the control. In the present study, percentage turbidity of juices decreased with the application of Pnl and after 2 h fruit juices became almost clear (Table 3). The results indicate the potential usefulness of the P35 pectin lyase in fruit juice industries.
Production of oil extraction
Oily seed samples (10 g) were dispersed in 10 ml of dis-tilled water to make a slurry and stirred at 20 rpm on a mag-netic stirrer. The pH of the dispersed solution was adjusted
Table 2 Effect of some metal ions on pectin lyase from Brevibacillus borstelensis activity
Chemical compounds
Concentration Pectin lyase activity (%)
none – 100
Ca2+ 5 135.5
Mg2+ 5 122.3
Hg2+ 5 1.4
Mn2+ 5 45.8
Zn2+ 5 95.3
Cu2+ 5 82.8
Fe2+ 5 32.2
Ba2+ 5 22.8
Table 3 Production of some fruit juice using pectin lyase from Brev-ibacillus borstelensis
Fruit (5 g) Control Pectin lyase Crude extract
Peach
Volume 9 10.5 10
Yield (%) – 116.7 111.1
Cherry
Volume 8 10 9
Yield (%) – 125 112.5
Grape
Volume 15 21.5 20.5
Yield (%) – 143.3 136.6
Apple
Volume 12 18.5 18.5
Yield (%) – 154.2 154.2
Strawberry
Volume 7 12 10
Yield (%) – 171.4 142.8
Orange
Volume 9 10 11
Yield (%) – 111.1 122.2
Table 4 Production of some fruit juice using pectin lyase from Brev-ibacillus borstelensis
Fruit (5 g) Control Pectin lyase Crude extract
Pumpkin seed
Volume (ml) 0.6 1.1 0.792
Yield (%) – 183.3 132
Nut
Volume (ml) 0.32 1.1 1.1
Yield (%) – 343.8 343.8
Sun flower
Volume (ml) 0.365 1.83 1.53
Yield (%) – 501.4 419.2
Ground nut
Volume (ml) 0.81 1.548 1.0
Yield (%) – 191.1 123.5
Corn
Volume (ml) 0.34 1.2 0.77
Yield (%) – 352.9 126.5
Olive
Volume (ml) 0.85 5.147 1.07
Yield (%) – 605.5 125.9

134 Eur Food Res Technol (2014) 239:127–135
1 3
to 8.0 with 0.1 n HCl. Enzymes were then added in vari-ous oily seed combinations, and the samples were kept at 60 °C for 1 h. The oil was recovered as an upper layer after centrifugation at 10,000×g for 20 min. The upper oil layer was separated and measured. Table 4 was showed that maximum oil recovery (605.5 %) was obtained from olive seed. Enzyme also increased the volume of pumpkin (0.6–1.1 ml), nut (0.32–1.1 ml), sun flower (0.365–1.83), ground nut (0.81–1.548) and corn (0.34–1.2 ml) in yield of 183, 343, 504, 343 and 353 %, respectively, as com-pared to control. all oil extraction experiments were also performed using crude extract. However, it was determined to be less effective crude extract than purified enzyme in the all oil extraction from raw oily seeds (Table 4). It was reported that an improvement of about 2 % on treated olives was obtained on the oil extracted by comparing the yield obtained without and with enzymes [21]. When the pectins were broken by Pnl enzyme, the flow of oil could be facilitated from cells and oil extraction was increasing. The results indicate the potential usefulness of the P35 pec-tin lyase in oil extraction industries.
Conclusions
In this paper, we report the production, purification and char-acterization of extra cellular Pnl (E.C. 4.2.2.10) produced by B. borstelensis. This is the first report on the produc-tion of a Pnl from B. borstelensis. We also determined the action of Pnl in fruit juice production and oil extraction. It was concluded that purified Pnl from B. borstelensis can be useful in process of yielding fruit juice and oil extraction.
Conflict of interest The authors stated that there are no conflicts of interest regarding the publication of this article. Research support played no role in the study design; in the collection, analysis, and interpretation of data; in the writing of the report; or in the decision to submit the report for publication.
Compliance with Ethics Requirements This article does not con-tain any studies with human or animal subjects.
References
1. Serrat M, Bermúdez RC, Villa Tg (2002) Production, purifica-tion, and characterization of a polygalacturonase from a new strain of Kluyveromyces marxianus isolated from coffee wet-pro-cessing wastewater. appl Biochem Biotechnol 97:193–208
2. Yadav S, Yadav PK, Yadav D, Yadav KDS (2008) Purification and characterization of an alkaline extracellular pectin lyase produced by Aspergillus flavus MTCC 7589. Process Biochem 43:547–552
3. alkorta I, garbisu C, liama MJ, Serra Jl (1998) Industrial appli-cation of pectic enzymes: a review. Process Biochem 33:21–28
4. Demir n, nadaroglu H, Tasgın E, adiguzel a, gulluce M (2011) Purification and characterization of a pectin lyase produced by
Geobacillus stearothermophilus (ah22) and fruit juice applica-tion. ann Microbiol 61(4):939–946
5. nadaroglu H, Taskin E, adiguzel a, gulluce M, Demir n (2010) Production of a pectin lyase from Bacillus pumilus (P9), purifi-cation and characterisation and fruit juice application. Rom Bio-technol lett 15(2):5167–5176
6. Whitaker JR (1990) Microbial pectolytic enzymes. In: Fogarty WM, Kelly CT (eds) Microbial enzymes and biotechnology, 2nd edn. Elsevier applied Science, new York, pp 133–176
7. Maugeri Tl, gugliandolo C, Caccamo D, Stackebrandt E (2001) a polyphasic taxonomic study of thermophilic bacilli from shal-low, marine vents. Syst appl Microbiol 24(4):572–587
8. Rahman TJ, Marchant R, Banat IM (2004) Distribution and molecular investigation of highly thermophilic bacteria associated with cool soil environments. Biochem Soc Trans 32(2):209–213
9. Savas S, adiguzel a, Inan K, Ozkan H, gulluce M, Sahin F (2009) Molecular characterization of thermophilic bacteria iso-lated from Van city Ercis town Hasanabdal hot spring. Rom Bio-technol lett 14(3):4445–4454
10. Fahy PC, Hayward aC (1983) Plant bacterial diseases: a diagnos-tic guide. academic Press, Sydney
11. adiguzel a (2006) Molecular characterization of thermophilic bacteria isolated from water samples taken from various thermal plants. PhD thesis, ataturk University, Erzurum, p 130
12. Beffa T, Blanc M, lyon PF, Vogt g, Marchiani M, Fischer Jl, aragno M (1996) Isolation of Thermus strains from hot composts (60 to 80°C). appl Environ Microbiol 62:1723–1727
13. adiguzel a, Ozkan H, Baris O, Inan K, gulluce M (2009) Iden-tification and characterization of thermophilic bacteria isolated from hot springs in Turkey. J Microbiol Methods 79:321–328
14. Taragano VM, Pilosof aMR (1999) application of Doehlert designs for water activity, pH, and fermentation time optimiza-tion for Aspergillus niger pectinolytic activities production in solid-state and submerged fermentation. Enzym Microb Technol 25:411–419
15. Kashyap DR, Chandra S, Kau a, Tewari R (2000) Production, purification and characterization of pectinase from a Bacillus sp. DT7. World J Microbiol Biotechnol 16:277–282
16. nedjma M, Hoffmann n, Belarbi a (2001) Selective and sensi-tive detection of pectin lyase activity using a colorimetric test: application to the screening of microorganisms possessing pectin lyase activity. anal Biochem 291:290–296
17. Bradford MM (1976) Rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principle of protein–dye binding. anal Biochem 72:248–253
18. laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage. nature 22:680–685
19. Whitaker JR (1963) Determination of molecular weight of pro-teins by gel filtration on Sephadex. anal Chem 35:1950–1953
20. Soraes MMCn, Da Silva R, Carmona EC, gomes E (2001) Pec-tinolytic enzyme production by Bacillus species and their poten-tial application on juice extraction. World J Microbiol Biotechnol 17:79–82
21. Hadj-Taieb n, ayadi M, Khlif M, Mrad K, Hassairi I, gargouri a (2006) Fermentor production of pectinases on gruel, a local by-product and their use in olive oil extraction. Enzym Microb Tech-nol 39(5, 4):1072–1076
22. Perry JJ, Staley JT (1997) Taxonomy of eubacteria and archaea. In: Perry JJ, Staley JT (eds) Microbiology: dynamics and diver-sity. Saunders College Publishing, Fort Worth, pp 388–413
23. Martins ES, Silva D, Da Silva R, gomes E (2002) Solid state pro-duction of thermostable pectinases from thermophilic Thermoas-cus aurantiacus. Process Biochem 37:949–954
24. Sinitsyna Oa, Fedorova Ea, Semenova MV, gusakov aV, Sokolova lM, Bubnova TM, Okunev On, Chulkin aM, Vavilova Ea, Vinetsky YP, Sinitsyn aP (2007) Isolation and

135Eur Food Res Technol (2014) 239:127–135
1 3
characterization of extracellular pectin lyase from Penicillium canescens. Biochemistry (Moscow) 72:565–571
25. Kamimiya S, nishiya T, Izaki K, Takahashi H (1974) Purification and properties of a pectin trans-eliminase in Erwinia aroideae formed in the presence of nalidixic acid. agric Biol Chem 38:1071–1078
26. Yadav S, Shastri nV (2007) Purification and properties of an extracellular pectin lyase produced by the strain of Penicillium oxalicum in solid-state fermentation. Indian J Biochem Biophys 44:247–251
27. nakagawa T, nagaoka T, Miyaji T, Tomizuka n (2005) a cold active pectin lyase from psychrophilic and basidiomycetous yeast Cystofilobasidium capitatum strain PPY-1. Biotechnol appl Bio-chem 42:193–196
28. Chen WC, Hsieh HJ, Tseng TC (1998) Purification and character-ization of a pectin lyase from Pythium splendens infected cucum-ber fruits. Bot Bull acad Sin 39:181–186
29. Cao J, Zheng l, Chen S (1992) Screening of pectinase producer from alkalophilic bacteria and study on its potential application in degumming of ramie. Enzyme Microb Technol 14:1013–1016
30. Bruhlmann F, leupin M, Erismann KH, Fiechter a (2000) Enzy-matic degumming of ramie bast fibers. J Biotechnol 76:43–50
31. Henriksson g, akin DE, Rigsby ll, Patel n, Eriksson KEl (1997) Influence of chelating agents and mechanical pretreatment on enzymatic retting of flax. Text Res J 67:829–836
32. Kapoor M, Beg QK, Bhushan B, Singh K, Dadhich KS, Hoondal gS (2001) application of alkaline thermostable polygalacturo-nase from Bacillus sp. Mg-ep-2 in degumming of ramie (Boe-hemeria nivea) and sunn hemp (Crotalaria juncea) bast fibers. Process Biochem 6:803–807
33. nagel CW, Hasegawa S (1968) The isolation and characterization of an 1,4 (4,4-dihydrogalacturonosyl) galacturonate hydrolase. J Food Sci 33:378–382
34. Starr MP, nasumo S (1967) Proteolytic activity of phytopatho-genic xanthomonads. J gen Microbiol 46:425–433
35. Dave Ba, Vaughn RH (1971) Purification and properties of a polygalaturonic acid trans eliminase produced by Bacillus pumi-lus. J Bacteriol 108:166–174
36. Horikoshi K (1972) Production of alkaline enzymes by alkalo-philic microorganisms. Part III. alkaline pectinase of Bacillus no. P-4-n. agric Biol Chem 36:285–293
37. Kelly CT, Fogarty WM (1978) Production and properties of polygalacturonate lyase by an alkalophilic microorganism Bacil-lus sp. RK9. Can J Microbiol 24(10):1164–1172




















