
Mechanistic kinetic model for symmetric carboligations using benzaldehyde lyase

Eur. J. Biochem.255, 2352245 (1998) FEBS1998
Interactions between serine acetyltransferase andO-acetylserine (thiol) lyasein higher plantsStructural and kinetic properties of the free and bound enzymes
Michel DROUX, Marie-Line RUFFET, Roland DOUCE and Dominique JOB
Laboratoire Mixte CNRS/Rhoˆne-Poulenc (UMR041), Rhone-Poulenc Agrochimie, Lyon, France
(Received13 February1998) 2 EJB 98 0220/4
The last steps of cysteine synthesis in plants involve two consecutive enzymes. The first enzyme,serine acetyltransferase, catalyses the acetylation ofL-serine in the presence of acetyl-CoA to formO-acetylserine. The second enzyme,O-acetylserine (thiol) lyase, convertsO-acetylserine toL-cysteine inthe presence of sulfide. We have, in the present work, over-produced inEscherichia coliharboring varioustype of plasmids, either a plant serine acetyltransferase or this enzyme with a plantO-acetylserine (thiol)lyase. The free recombinant serine acetyltransferase (subunit mass of 34 kDa) exhibited a high propensityto form high-molecular-mass aggregates and was found to be highly unstable in solution. However, theseaggregates were prevented in the presence ofO-acetylserine (thiol) lyase (subunit mass of 36 kDa). Underthese conditions homotetrameric serine acetyltransferase associated with two molecules of homodimericO-acetylserine (thiol) lyase to form a bienzyme complex (molecular mass<300 kDa) called cysteinesynthase containing 4 mol pyridoxal 5′-phosphate/mol complex.O-Acetylserine triggered the dissociationof the bienzyme complex, whereas sulfide counteracted the action ofO-acetylserine. Protein2proteininteractions within the bienzyme complex strongly modified the kinetic properties of plant serine acetyl-transferase: there was a transition from a typical Michaelis-Menten model to a model displaying positivekinetic co-operativity with respect to serine and acetyl-CoA. On the other hand, the formation of thebienzyme complex resulted in a very dramatic decrease in the catalytic efficiency of boundO-acetylserine(thiol) lyase. The latter enzyme behaved as if it were a structural and/or regulatory subunit of serineacetyltransferase. Our results also indicated that bound serine acetyltransferase produces a build-up ofO-acetylserine along the reaction path and that the full capacity for cysteine synthesis can only be achievedin the presence of a large excess of freeO-acetylserine (thiol) lyase. These findings contradict the widelyheld belief that such a bienzyme complex is required to channel the metabolite intermediateO-acetylser-ine.
Keywords:cysteine synthase; serine acetyltransferase;O-acetylserine (thiol) lyase; protein2protein in-teractions; cysteine synthesis in plants.
In metabolic networks, a fine tuning of enzyme activities is verted into the final reaction product (P). Here, accumulation ofI might be prevented through properly adjusted concentrationsrequired to maintain the rates at which useful end-products are
produced with regard to those at which they are needed, but also of E2 over E1, so that, at steady state, the net rate of I transforma-tion through E2 at least equals that of synthesis through E1. Thisto avoid accumulation of toxic internal metabolites and/or to
achieve efficient trapping of highly unstable reaction intermedi- situation is analogous to that encountered in enzyme assays withauxiliary enzymes [1].ates [123]. Considering the simple case of two enzymes catalys-
ing consecutive reactions, this might be achieved essentially ac- There exists, however, in the literature at least one examplesuggesting that catalysis of consecutive reactions by distinct en-cording to two extreme types of mechanisms. In the first, as
depicted in model1 of Fig. 1, the two enzymes E1 and E2 associ- zymes may proceed through a different mechanism. This situa-tion relates to the two plant enzymes that are involved, as theirate permanently, so that the reaction intermediate I (i.e. the prod-
uct of E1 and substrate of E2) can be directly transferred (e.g. bacterial counterparts [4], in the final steps of cysteine biosyn-thesis. The first enzyme in the pathway, serine acetyltransferasethrough a channeling mechanism) from the active site of E1 to
that of E2. In the second, depicted in model 2 of Fig.1, the two (EC 2.3.1.30), catalyses the acetylation ofL-serine in the pres-ence of acetyl-CoA, thus yielding the carbon precursorO-ace-enzymes do not associate, even transiently, so that I is released
into solution and must reassociate with E2 before to being con- tylserine (Eqn1). The second enzyme,O-acetylserine (thiol) ly-ase (EC 4.2.99.8), convertsO-acetylserine toL-cysteine in the
Correspondence toM. Droux, Laboratoire Mixte CNRS/Rhoˆne- presence of free or bound sulfide (Eqn 2) [5].Poulenc, (UMR041) Rhone-Poulenc Agrochimie,14220, rue Pierre
Serine1 acetyl-CoA→ O-Acetylserine1 CoASH (1)Baizet, F-69263, Lyon cedex 9, FranceFax: 133 04 72 85 22 97. O-Acetylserine1 sulfide→ Cysteine1 acetate . (2)E-mail : [email protected]
Both enzymes have been extensively studied in bacteria andEnzymes.SerineO-acetyltransferase (EC 2.3.1.30) ; O-acetylserine(thiol) lyase (EC 4.2.99.8). it is now well established that, in procaryotes, homotetrameric

236 Droux et al. (Eur. J. Biochem. 255)
teine biosynthesis in plants. Thus, we previously demonstratedthe existence of excess ofO-acetylserine (thiol) lyase over serineacetyltransferase molecules in all plant cell compartments whereprotein synthesis occurs, namely cytosol, mitochondria and chlo-roplast [9,14, 15]. Also, in vitro assays conducted with purespinach chloroplast serine acetyltransferase indicated that a largeexcess ofO-acetylserine (thiol) lyase over serine acetyltransfer-ase molecules is needed to achieve optimal cysteine synthesisfrom L-serine, acetyl CoA and sulfide [10]. These featuresthereby suggest that the two enzymes may function in plantsaccording to model 2 of Fig.1. Alternative possibilities to ac-count for these findings would be that the rather difficult purifi-cation procedure used to isolate the plant serine acetyltransferase[10] only led to some altered form of this enzyme, which mightbe unable to establish specific interactions with the correspond-ing O-acetylserine (thiol) lyase, and/or thatO-acetylserine(thiol) lyase plays additional, yet unknown, roles in higher plantsthan in the last step of cysteine synthesis. It is established, how-ever, thatO-acetylserine does not accumulate in plant tissuesand that this molecule has no other known function in plantsother than to serve as a substrate for cysteine synthesis [16]. Therecent cloning of both serine acetyltransferase [12, 17219] andO-acetylserine (thiol) lyase [15, 20222] plant cDNAs offers thepossibility to overproduce the plant enzymes in recombinantFig. 1. Theoretical models for cysteine synthesis involving serine ace-bacteria and to purify them in much larger quantities and pre-tyltransferase and O-acetylserine (thiol) lyase.E1 stands for serine
acetyltransferase and E2 for O-acetylserine (thiol) lyase. The following sumably in more native states than from plant tissues.possibilities are shown: model1 or direct transfer mechanism, refers to The aim of this present work is (a) to evaluate the efficiencytrue intermediate channeling ; model 2, assumes a free diffusion mecha-of a plant cysteine-synthase complex in catalysis of cysteine for-nism; model 3, assumes release of theO-acetylserine intermediate (I) in mation and (b) to compare the kinetic properties of the free andthe bulk solution and synthesis of cysteine (P) both from the bienzymebound enzymes.complex and the freeO-acetylserine (thiol) lyase acting as an auxiliaryenzyme.
MATERIALS AND METHODS
Construction of expression plasmids.A cDNA encodingserine acetyltransferase associates with two molecules of homo-dimericO-acetylserine (thiol) lyase to form a bienzyme complex an isoform ofA. thalianaserine acetyltransferase of 315 amino-
acid residues and with a molecular mass of 34 330 Da [18] wascalled cysteine synthase [6, 7]. The activity of the complexmight be subjected to metabolic regulation since accumulation used to generate two types of plasmids. In the first, referred to
as pET9d-SAT, the plant cDNA was cloned as a 942-bpNcoI2of O-acetylserine causes this complex to dissociate into one mol-ecule of serine acetyltransferase and two molecules ofO-ace- BamHI fragment into the pET-9d vector (Novagen). In the sec-
ond, referred to as pET29a-SAT, the NcoI2BamHI insert wastylserine (thiol) lyase whereas sulfide prevents complex dissoci-ation [7]. The role of specific protein2protein interactions in the cloned into pET-29a, an expression vector designed to generate
fusion proteins owing to a15 amino-acid leader S-Tag pre-se-formation of bacterial cysteine synthase has been well illustratedby the discovery of a mutant form ofSalmonella typhimurium quence that allows affinity purification of recombinant proteins
(Novagen). The plasmids, carrying the kanamycin resistance,serine acetyltransferase. This mutation alters both the interac-tions with O-acetylserine (thiol) lyase and the catalytic proper- were used to transformEscherichia coliBL21(DE3)pLysS cells
(Novagen) or to co-transform these cells in the presence ofties of each of the two enzymes within the functional com-plex [8]. pET3d-OASTL.The latter carries the ampicillin resistance and
encodes for the mature form of spinach chloroplastO-acetylser-The mechanism of cysteine synthesis in plants is less clear.On the one hand, recent work suggests that serine acetyltransfer- ine (thiol) lyase [5].
Expression and purification of recombinant serine acetyl-ase andO-acetylserine (thiol) lyase may functionin vivo accord-ing to model1 in Fig. 1, that is, they associate to form a bien-transferase.Bacterial strains were grown at 37°C in Luria-Ber-
tani medium [1% (mass/vol.) trypton, 0.5% (mass/vol.) yeastzyme complex analogous to bacterial cysteine synthase. Experi-mental evidence favoring this hypothesis includes the elution, extract and 0.5% (mass/vol.) NaCl] supplemented with
200µg · ml21 kanamycin, or with both 200µg · ml21 kanamycinupon gel-filtration chromatography, of a complex composed ofserine acetyltransferase andO-acetylserine (thiol) lyase from and ampicillin for the co-transformed cells, for12 h at180 rpm.
These cultures were used to inoculate 500 ml fresh medium, inspinach chloroplasts [9,10], the purification of serine acetyl-transferase with firmly boundO-acetylserine (thiol) lyase inAl- the presence of the respective antibiotics at100 µg · ml21, in a
Belco flask, and grown at 37°C (180 rpm). WhenA600 reachedlium tuberosum[11], the reconstitution of a plant cysteine syn-thase from recombinant watermelon serine acetyltransferase and 0.6,1 mM isopropyl β-thiogalactopyranoside was added, and
growth was continued for 4 h at 37°C. To ensure high expres-O-acetylserine (thiol) lyase [12] and the characterization of theinteractions between serine acetyltransferase andO-acetylserine sion of active recombinant, pyridoxal-5′-phosphate-dependent
O-acetylserine (thiol) lyase, the growth medium also contained(thiol) lyase fromArabidopsis thalianawith a yeast two-hybridsystem [13]. 10 µM pyridoxine and15 µM thiamin. Thiamin is required be-
cause pyridoxine inhibits endogenous thiamin biosynthesis [15].On the other hand, other results do not favor the idea that acysteine-synthase complex, if it exists, plays a major role in cys- Cells were collected by centrifugation (2500g, 20 min), resus-

237Droux et al. (Eur. J. Biochem. 255)
pended in 25 mM Hepes/HCl pH 7.5,1mM EDTA-Na2, 1 mM 40-ml linear gradient of 020.4 M NaCl in the same buffer (flowdithiothreitol, 5 mM 6-aminohexanoic acid and1mM benzami- rate, 0.5 ml · min21; 1-ml fractions). The recombinant andE.dine [with addition of 0.1% (by vol.) Triton X-100 when indi- coli O-acetylserine (thiol) lyases eluted as a sharp peak atcated], and disrupted by sonication at 4°C (Vibra-cell disruptor). 0.22 M and 0.17 M NaCl, respectively.Lysates were centrifuged at 40000g for 30 min and the superna- Purification of the recombinant serine acetyltransferase wastants stored at280°C. achieved by enzymatic cleavage as described above for the cys-
The lysate ofE. coli cells harboring the pET-9d-SATplasmid teine-synthase complexes.(275 mg protein) was supplemented with ammonium sulfate to Enzyme assays.O-acetylserine (thiol) lyase activity was50% saturation. The resulting precipitate (120 mg protein) was measured as described [5] in a volume of100 µl containingdissolved in 25 mM Hepes/HCl pH 7.5,1mM EDTA-Na2 and 50 mM Hepes/HCl pH 7.5,1mM dithiothreitol,10 mM O-ace-150 mM NaCl (buffer A) and desalted on a Sephadex G25 col-tylserine and 2.5 mM sulfide. Cysteine was determined accord-umn equilibrated in buffer A. This extract was applied to a Frac-ing to the method of Gaitonde [24]. To evaluateO-acetylserinetogel EMD-650(M) column (1.6315 cm; Merck) equilibrated in (thiol) lyase activity in crude extracts of transformed bacteria,buffer A and connected to a Pharmacia FPLC system. Proteinsthe extracts were first incubated in the presence of10 mM O-were eluted using a 020.7 M NaCl gradient in buffer A. The acetylserine to ensure cysteine-synthase complex dissociationmajor part of serine acetyltransferase activity, corresponding toprior to conducting the enzyme assay as above. For steady-statethe recombinant enzyme, eluted in the 2002280 mM NaCl frac- analyses of bound and freeO-acetylserine (thiol) lyase, the con-tions. A minor part, corresponding to the endogenous bacterialcentration ofO-acetylserine varied over 0220 mM at a constantactivity, eluted at 350 mM NaCl. The fractions containing theconcentration of 2.5 mM Na2S. Alternatively, that of Na2S variedhighest specific activity from the first peak were pooled, concen-over 025 mM at a constant concentration of10 mM O-ace-trated using a Filtron 30K device (Amicon), and applied to atylserine. One unit (U) of enzyme activity corresponded to theblue-Sepharose CL-6B column (1.638 cm, Pharmacia) equili- amount of enzyme catalysing the formation of1 µmolbrated in 20 mM Hepes/HCl pH 6.8,1mM EDTA-Na2 and cysteine · min21 at 25°C.150 mM NaCl (buffer B). After washing with 20 ml 0.5 M Serine acetyltransferase was assayed for the production ofNaCl, then with 20 ml1M NaCl, proteins were eluted by pas-O-acetylserine after derivatization witho-phthalaldehyde, fol-sage of 40 ml 2 M NaCl in buffer B, concentrated as above, andlowed by reverse-phase chromatography using a high-perfor-submitted to a gel-filtration chromatography on Superdex S200mance liquid chromatography system (HPLC). Reactions were(Pharmacia) equilibrated in buffer B supplemented with 0.25 Mcarried out at 25°C in a final volume of 100µl containingNaCl. Active fractions were concentrated and stored at 5°C in 50 mM Hepes/HCl pH 7.5,1mM acetyl-CoA,10 mM L-serine20 mM Hepes/HCl, pH 7.5,1mM EDTA-Na2 and 250 mM and an appropriate amount of enzyme. Reactions were initiatedNaCl. Activity measurements were performed immediately afterwith acetyl-CoA, and stopped by adding 50µl 20% (mass/vol.)enzyme purification, because the purified recombinant enzymetrichloroacetic acid followed by centrifugation at12 000g forwas found to be extremely unstable, loosing enzyme activity5 min. An aliquot of the supernatant (502100 µl) was mixedwith a half life of about 4 h. with 500µl of a solution ofo-phthalaldehyde (54 mgo-phthalal-
Purification of cysteine synthase and of component en- dehyde dissolved in1ml ethanol, 9 ml 400 mM borate/NaOHzymes.Soluble protein extracts (approx. 240 mg protein) werepH 9.5, and 0.2 ml 2-mercaptoethanol) and brought to1ml withprepared as above from lysates ofE. coli cells harboring the water. An aliquot of this reaction medium (10250 µl) was in-pET29a-SATplasmid (with or without the pET3d-OASTLvec- jected onto an AccQ.TagTM C18 column (3.93150 mm, 10-nmtor), and incubated with 2 ml S-protein AgaroseTM (Novagen) for particle size, Millipore Waters) connected to a HPLC system30 min at 25°C with gentle shaking to ensure optimal binding of
using a Waters 717 Plus autosampler. Mobile phases used forthe fusion protein to the matrix. Following a wash with 20 mMelution of theo-phthalaldehyde derivatives were as follows : A,
Tris/HCl pH 7.5, 150 mM NaCl and 0.1% (by vol.) Triton85 mM sodium acetate to pH 4.5 with acetic acid, 6% (by vol.)X-100, the cysteine-synthase complexes were recovered by en-acetonitrile; B, 60% acetonitrile in H2O. The elution protocolzymatic cleavage with biotinylated thrombin (25 U) for 2 h at(30°C, 1 ml · min21) employed a linear gradient as follows: ini-25°C in 3 ml buffer containing 20 mM Tris/HCl pH 8.4,tial, 25% B in A; 028 min, 25270% B in A, and re-equilibra-150 mM NaCl, 2.5 mM CaCl2 (Novagen). Biotinylated thrombintion for 5 min in 25% B in A before new injection. Fluorescencewas removed by a streptavidin capture on Streptavidin2Agaroseof theo-phthalaldehyde adducts was monitored at 455 nm upon(Novagen) according to the manufacturer’s instructions. The elu-excitation at 340 nm using a SFM25 fluorimeter (Kontron). Theate was concentrated to1 ml by ultrafiltration (YM10 mem-retention times forL-serine andL-O-acetylserine derivativesbrane, Amicon), and submitted to a gel-filtration chromatogra-were respectively 2.3 min and 5.8 min. Amino-acids were quan-phy on a PD10 column (Pharmacia) equilibrated in 25 mMtified by measuring peak areas using the 450-MT2 softwareHepes/HCl pH 7.5,1mM EDTA-Na2 and 250 mM NaCl.(Kontron): 1 unit area corresponded to 3.6 pmolL-O-acetylser-Matrix-bound cysteine-synthase complexes were also sub-ine. One unit (U) of activity corresponded to the amount of en-mitted to a dissociation step prior to thrombin treatment withzyme catalysing the formation of1 µmol O-acetylserine · min21.10 ml wash buffer containing10 mM O-acetylserine. This re-For steady-state kinetic analyses theL-serine concentration var-sulted in the specific elution of either the recombinant or theied over 0220 mM at a constant concentration of acetyl-CoAbacterialO-acetylserine (thiol) lyase. At this stage the resolved(2 mM), and that of acetyl-CoA over 025 mM at a constantenzyme eluted with boundO-acetylserine in a Schiff base inter-concentration of serine (10 mM).mediate betweenA-aminoacrylate and the pyridoxal 5′-phos-
Serine acetyltransferase was also assayed by measuring itsphate cofactor [5, 23]. Following addition of excess sulfide tocapacity to promote cysteine synthesis in the presence of variedconvert the excessO-acetylserine into cysteine, the preparationsconcentrations ofL-serine and acetyl-CoA, and a constant con-were desalted using a PD10 column equilibrated in 50 mMcentration (2.5 mM) of sulfide. This assay is referred to as theHepes/HCl pH 7.5, and applied to a Mono Q HR5/5 columncysteine-synthase assay [25]. Here,O-acetylserine (thiol) lyase(Pharmacia) equilibrated in 50 mM Hepes/HCl pH 7.5,1 mM(either that present in the assayed extracts or the purified recom-EDTA-Na2, at room temperature. After the column had been
washed with 5 ml equilibration buffer, it was eluted with a binant enzyme) was used as a coupling enzyme. Cysteine pro-

238 Droux et al. (Eur. J. Biochem. 255)
duced was measured as described [24], with1 U activity corre-sponding to1 µmol cysteine · min21 at 25°C.
Kinetic data analysis.With all methods used, enzyme activ-ities were shown to be dependent on the presence of all requiredsubstrates, and were linear with respect to the amount of proteinassayed, testifying adherence to steady-state conditions. The ki-netic data were fitted to the appropriate rate equations by non-linear regression analysis using the KaleidagraphTM programme(Abelbeck Software) and a Macintosch LC475 computer. Forthe characterization of the interactions between serine acetyl-transferase andO-acetylserine (thiol) lyase, as measured by thecysteine-synthase assay, the data were analysed in terms of aserine acetyltransferaseO-acetylserine (thiol) lyase equilibrium,and Eqn. (3) was used.
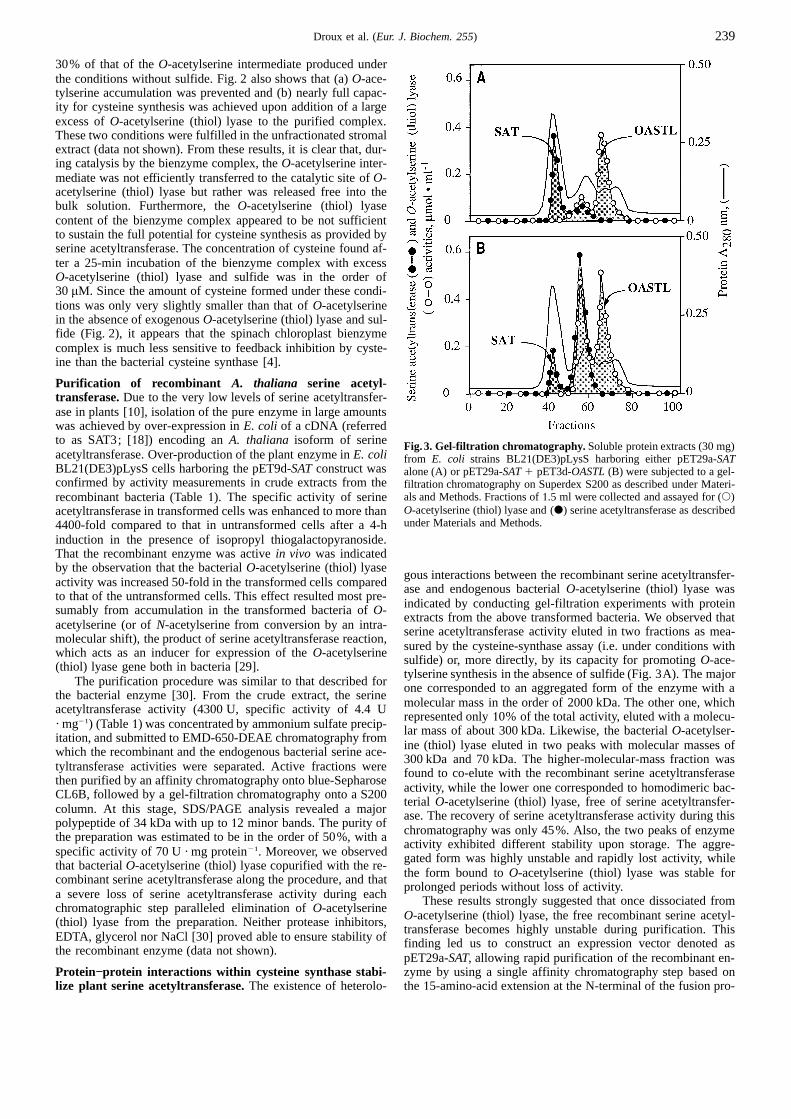
Activity 5Fig.2. Time courses ofO-acetylserine and cysteine accumulation.The spinach chloroplast bienzyme complex was prepared as reported [9].Vmax
([SAT] 1 [L] 1Kd) 2 !([SAT] 1 [L] 1 Kd)2 2 4[SAT] [L]
2[SAT] O-Acetylserine and cysteine accumulation were measured as describedunder Materials and Methods in the presence of10 mM L-serine and(3)2 mM acetyl-CoA (O-acetylserine) and10 mM L-serine, 2 mM acetyl-CoA and 2.5 mM sulfide (cysteine). (h) O-Acetylserine accumulationwhere [SAT] and [L] are the total concentrations of serine ace-in the absence of sulfide; (m) O-acetylserine or (d) cysteine formed intyltransferase andO-acetylserine (thiol) lyase (subunits), respec-the presence of 2.5 mM sulfide; (r) capacity for cysteine and (e) O-tively, Kd is the equilibrium dissociation constant for the en-acetylserine formed in the presence of a 300-fold excess of purified re-
counter complex andVmax represents the maximum activity for combinant spinach chloroplastO-acetylserine (thiol) lyase over thecysteine synthesis achieved at saturation. bound O-acetylserine (thiol) lyase, and 2.5 mM sulfide. The maximal
Steady-state rate data obtained by varying substrate concen-concentration of cysteine that was reached in the complete assay was of30µM.trations were fitted using the Michaelis-Menten model [1]. Al-
ternatively, when a significant deviation from this model wasobserved, the Hill equation (Eqn 4) was used:
Table 1. Expression of recombinantArabidopsis thalianaserine ace-v 5
Vm[S]h
11 F S
K Gh
(4) tyltransferase in Escherichia colicells transformed with pET9d-SAT,pET29a-SAT, or with pET29a-SAT plus pET3d-OASTL. Serine ace-tyltransferase andO-acetylserine (thiol) lyase activities were measuredin crude desalted extracts fromE. coli strain BL21(DE3)pLysS. Theconstructions of overexpression plasmids (pET9d-SAT, pET29a-SAT) arewhere the Hill constant,h, was calculated from the rate (v) ver-described under Materials and Methods. The construction of expressionsus total substrate concentration.plasmid pET3d-OASTLwas described in [5]. WhenA600 reached 0.6,Gel-filtration chromatography. Protein extracts were frac-1mM isopropyl thiogalactopyranoside was added and growth was con-tionated by gel-filtration chromatography using a Superdex S200tinued for 4 h at 37°C. Enzyme activities were then measured in bacterialcolumn (Pharmacia), equilibrated with 20 mM Tris/HCl pH 7.5,lysates as described under Materials and Methods.
1mM EDTA-Na2, 1mM dithiothreitol and 200 mM NaCl, andconnected to a Pharmacia FPLC system. Elution was performedPlasmid Time Activity ofat room temperature (flow rate,1 ml · min21 ; 1-ml fractions) [9].
serine acetyl- O-acetylserine (thiol)Protein determination. Protein was measured by thetransferase lyasemethod of Bradford [26] using the Biorad Protein assay reagent
with γ-globulin as the standard, and when working with purifiedh µmol min21 mg protein21
protein by measuring the absorbance at 205 nm [27]. The con-centrations of plant and bacterialO-acetylserine (thiol) lyase
pET9d-SAT 0 0.001 0.09were determined from the absorbance at 409 nm or 412 nm, re- 4 4.4 3.8spectively, using their respective absorption coefficient [5, 7].
pET29a-SAT 0 0.001 0.009Electrophoretic and western-blot analyses.SDS/PAGE4 4.2 3.1was performed at room temperature in SDS/polyacrylamide slab
pET29a-SAT1 0 0.002 0.2gels (15315 cm) containing a10220% (mass/vol.) acrylamidepET3d-OASTL 4 11 9.0gradient, according to Chua [28]. Western-blot analyses were
conducted using antibodies raised against purified spinach chlo-roplastO-acetylserine (thiol) lyase, as described previously [9].
complex does not contain freeO-acetylserine (thiol) lyase mole-cules], as obtained by gel-filtration chromatography [9] (Fig. 2).RESULTSIn the presence of sulfide,O-acetylserine first rapidly accumu-lated and then continued to increase at a slower rate throughoutTime course of the overall reaction catalysed by the native
bienzyme complex.Serine acetyltransferase andO-acetylserine the remainder of the time course. Likewise, synthesis of cysteinedisplayed a considerable lag. Furthermore, these data show that,(thiol) lyase activities were measured from a purified fraction of
the chloroplast bienzyme complex composed of firmly bound following a 25-min incubation, the amount of cysteine accumu-lated under the conditions with sulfide only represented aboutserine acetyltransferase andO-acetylserine (thiol) lyase [i.e. this
239Droux et al. (Eur. J. Biochem. 255)
30% of that of theO-acetylserine intermediate produced underthe conditions without sulfide. Fig. 2 also shows that (a)O-ace-tylserine accumulation was prevented and (b) nearly full capac-ity for cysteine synthesis was achieved upon addition of a largeexcess ofO-acetylserine (thiol) lyase to the purified complex.These two conditions were fulfilled in the unfractionated stromalextract (data not shown). From these results, it is clear that, dur-ing catalysis by the bienzyme complex, theO-acetylserine inter-mediate was not efficiently transferred to the catalytic site ofO-acetylserine (thiol) lyase but rather was released free into thebulk solution. Furthermore, theO-acetylserine (thiol) lyasecontent of the bienzyme complex appeared to be not sufficientto sustain the full potential for cysteine synthesis as provided byserine acetyltransferase. The concentration of cysteine found af-ter a 25-min incubation of the bienzyme complex with excessO-acetylserine (thiol) lyase and sulfide was in the order of30 µM. Since the amount of cysteine formed under these condi-tions was only very slightly smaller than that ofO-acetylserinein the absence of exogenousO-acetylserine (thiol) lyase and sul-fide (Fig. 2), it appears that the spinach chloroplast bienzymecomplex is much less sensitive to feedback inhibition by cyste-ine than the bacterial cysteine synthase [4].
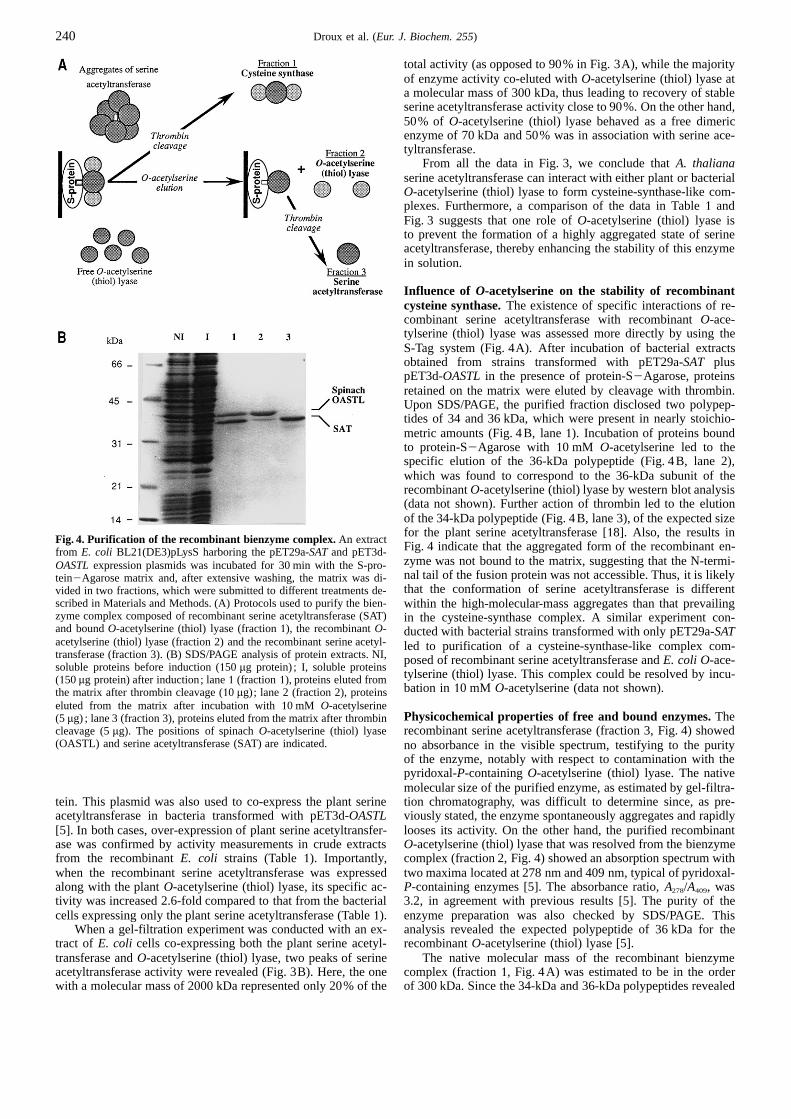
Purification of recombinant A. thaliana serine acetyl-transferase.Due to the very low levels of serine acetyltransfer-ase in plants [10], isolation of the pure enzyme in large amountswas achieved by over-expression inE. coli of a cDNA (referredto as SAT3; [18]) encoding anA. thaliana isoform of serine Fig.3. Gel-filtration chromatography. Soluble protein extracts (30 mg)acetyltransferase. Over-production of the plant enzyme inE. coli from E. coli strains BL21(DE3)pLysS harboring either pET29a-SATBL21(DE3)pLysS cells harboring the pET9d-SATconstruct was alone (A) or pET29a-SAT1 pET3d-OASTL(B) were subjected to a gel-confirmed by activity measurements in crude extracts from thefiltration chromatography on Superdex S200 as described under Materi-
als and Methods. Fractions of1.5 ml were collected and assayed for (s)recombinant bacteria (Table1). The specific activity of serineO-acetylserine (thiol) lyase and (d) serine acetyltransferase as describedacetyltransferase in transformed cells was enhanced to more thanunder Materials and Methods.4400-fold compared to that in untransformed cells after a 4-h
induction in the presence of isopropyl thiogalactopyranoside.That the recombinant enzyme was activein vivo was indicatedby the observation that the bacterialO-acetylserine (thiol) lyase
gous interactions between the recombinant serine acetyltransfer-activity was increased 50-fold in the transformed cells comparedase and endogenous bacterialO-acetylserine (thiol) lyase wasto that of the untransformed cells. This effect resulted most pre-
indicated by conducting gel-filtration experiments with proteinsumably from accumulation in the transformed bacteria ofO-extracts from the above transformed bacteria. We observed that
acetylserine (or ofN-acetylserine from conversion by an intra-serine acetyltransferase activity eluted in two fractions as mea-molecular shift), the product of serine acetyltransferase reaction,
sured by the cysteine-synthase assay (i.e. under conditions withwhich acts as an inducer for expression of theO-acetylserinesulfide) or, more directly, by its capacity for promotingO-ace-(thiol) lyase gene both in bacteria [29].tylserine synthesis in the absence of sulfide (Fig. 3A). The major
The purification procedure was similar to that described forone corresponded to an aggregated form of the enzyme with athe bacterial enzyme [30]. From the crude extract, the serine
molecular mass in the order of 2000 kDa. The other one, whichacetyltransferase activity (4300 U, specific activity of 4.4 Urepresented only10% of the total activity, eluted with a molecu-
· mg21) (Table1) was concentrated by ammonium sulfate precip-lar mass of about 300 kDa. Likewise, the bacterialO-acetylser-itation, and submitted to EMD-650-DEAE chromatography from
ine (thiol) lyase eluted in two peaks with molecular masses ofwhich the recombinant and the endogenous bacterial serine ace-300 kDa and 70 kDa. The higher-molecular-mass fraction was
tyltransferase activities were separated. Active fractions werefound to co-elute with the recombinant serine acetyltransferasethen purified by an affinity chromatography onto blue-Sepharose
activity, while the lower one corresponded to homodimeric bac-CL6B, followed by a gel-filtration chromatography onto a S200terial O-acetylserine (thiol) lyase, free of serine acetyltransfer-
column. At this stage, SDS/PAGE analysis revealed a majorase. The recovery of serine acetyltransferase activity during thispolypeptide of 34 kDa with up to12 minor bands. The purity of
chromatography was only 45%. Also, the two peaks of enzymethe preparation was estimated to be in the order of 50%, with aactivity exhibited different stability upon storage. The aggre-
specific activity of 70 U · mg protein21. Moreover, we observed gated form was highly unstable and rapidly lost activity, whilethat bacterialO-acetylserine (thiol) lyase copurified with the re-
the form bound toO-acetylserine (thiol) lyase was stable forcombinant serine acetyltransferase along the procedure, and thatprolonged periods without loss of activity.
a severe loss of serine acetyltransferase activity during each These results strongly suggested that once dissociated fromchromatographic step paralleled elimination ofO-acetylserine
O-acetylserine (thiol) lyase, the free recombinant serine acetyl-(thiol) lyase from the preparation. Neither protease inhibitors,transferase becomes highly unstable during purification. This
EDTA, glycerol nor NaCl [30] proved able to ensure stability offinding led us to construct an expression vector denoted asthe recombinant enzyme (data not shown).
pET29a-SAT, allowing rapid purification of the recombinant en-zyme by using a single affinity chromatography step based onProtein−protein interactions within cysteine synthase stabi-
lize plant serine acetyltransferase.The existence of heterolo- the15-amino-acid extension at the N-terminal of the fusion pro-
240 Droux et al. (Eur. J. Biochem. 255)
total activity (as opposed to 90% in Fig. 3A), while the majorityof enzyme activity co-eluted withO-acetylserine (thiol) lyase ata molecular mass of 300 kDa, thus leading to recovery of stableserine acetyltransferase activity close to 90%. On the other hand,50% of O-acetylserine (thiol) lyase behaved as a free dimericenzyme of 70 kDa and 50% was in association with serine ace-tyltransferase.
From all the data in Fig. 3, we conclude thatA. thalianaserine acetyltransferase can interact with either plant or bacterialO-acetylserine (thiol) lyase to form cysteine-synthase-like com-plexes. Furthermore, a comparison of the data in Table1 andFig. 3 suggests that one role ofO-acetylserine (thiol) lyase isto prevent the formation of a highly aggregated state of serineacetyltransferase, thereby enhancing the stability of this enzymein solution.
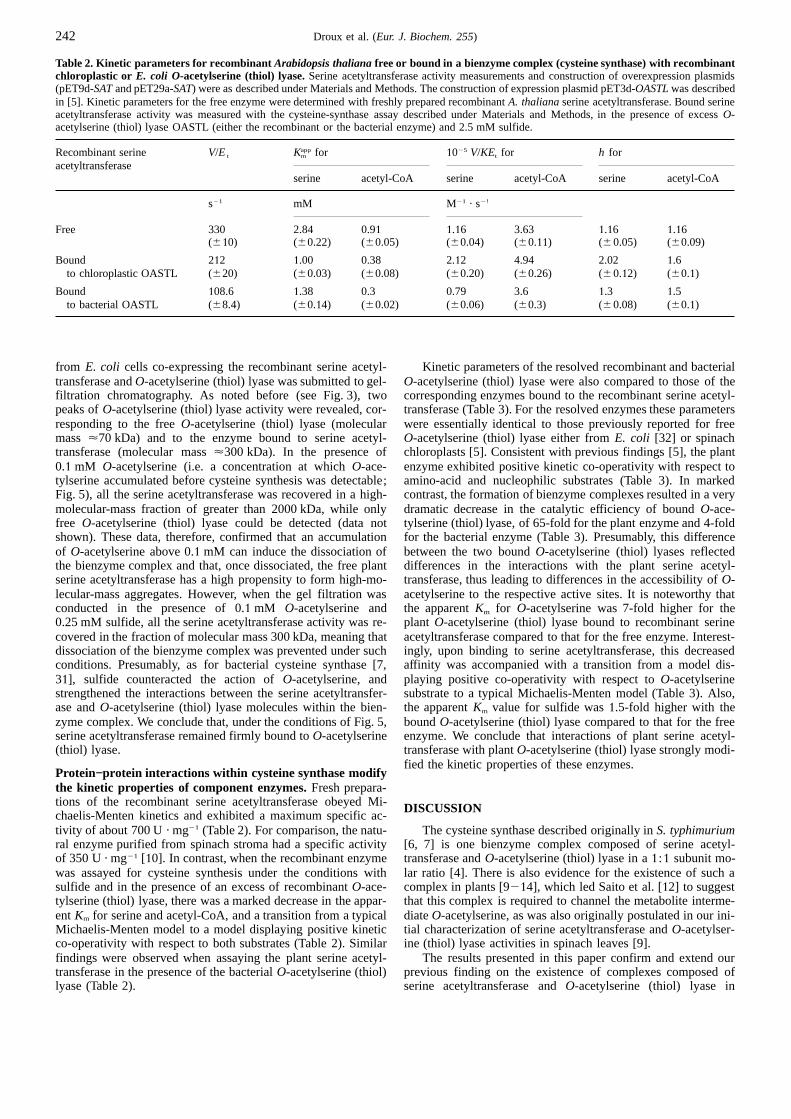
Influence of O-acetylserine on the stability of recombinantcysteine synthase.The existence of specific interactions of re-combinant serine acetyltransferase with recombinantO-ace-tylserine (thiol) lyase was assessed more directly by using theS-Tag system (Fig. 4A). After incubation of bacterial extractsobtained from strains transformed with pET29a-SAT pluspET3d-OASTLin the presence of protein-S2Agarose, proteinsretained on the matrix were eluted by cleavage with thrombin.Upon SDS/PAGE, the purified fraction disclosed two polypep-tides of 34 and 36 kDa, which were present in nearly stoichio-metric amounts (Fig. 4B, lane1). Incubation of proteins boundto protein-S2Agarose with10 mM O-acetylserine led to thespecific elution of the 36-kDa polypeptide (Fig. 4B, lane 2),which was found to correspond to the 36-kDa subunit of therecombinantO-acetylserine (thiol) lyase by western blot analysis(data not shown). Further action of thrombin led to the elutionof the 34-kDa polypeptide (Fig. 4B, lane 3), of the expected sizefor the plant serine acetyltransferase [18]. Also, the results in
Fig. 4. Purification of the recombinant bienzyme complex.An extractFig. 4 indicate that the aggregated form of the recombinant en-from E. coli BL21(DE3)pLysS harboring the pET29a-SATand pET3d-zyme was not bound to the matrix, suggesting that the N-termi-OASTLexpression plasmids was incubated for 30 min with the S-pro-nal tail of the fusion protein was not accessible. Thus, it is likelytein2Agarose matrix and, after extensive washing, the matrix was di-that the conformation of serine acetyltransferase is differentvided in two fractions, which were submitted to different treatments de-
scribed in Materials and Methods. (A) Protocols used to purify the bien-within the high-molecular-mass aggregates than that prevailingzyme complex composed of recombinant serine acetyltransferase (SAT)in the cysteine-synthase complex. A similar experiment con-and boundO-acetylserine (thiol) lyase (fraction1), the recombinantO- ducted with bacterial strains transformed with only pET29a-SATacetylserine (thiol) lyase (fraction 2) and the recombinant serine acetyl-led to purification of a cysteine-synthase-like complex com-transferase (fraction 3). (B) SDS/PAGE analysis of protein extracts. NI,posed of recombinant serine acetyltransferase andE. coli O-ace-soluble proteins before induction (150 µg protein) ; I, soluble proteins
tylserine (thiol) lyase. This complex could be resolved by incu-(150µg protein) after induction; lane1 (fraction1), proteins eluted frombation in10 mM O-acetylserine (data not shown).the matrix after thrombin cleavage (10 µg); lane 2 (fraction 2), proteins
eluted from the matrix after incubation with10 mM O-acetylserinePhysicochemical properties of free and bound enzymes.The(5 µg) ; lane 3 (fraction 3), proteins eluted from the matrix after thrombin
cleavage (5µg). The positions of spinachO-acetylserine (thiol) lyase recombinant serine acetyltransferase (fraction 3, Fig. 4) showed(OASTL) and serine acetyltransferase (SAT) are indicated. no absorbance in the visible spectrum, testifying to the purity
of the enzyme, notably with respect to contamination with thepyridoxal-P-containingO-acetylserine (thiol) lyase. The nativemolecular size of the purified enzyme, as estimated by gel-filtra-tion chromatography, was difficult to determine since, as pre-tein. This plasmid was also used to co-express the plant serine
acetyltransferase in bacteria transformed with pET3d-OASTL viously stated, the enzyme spontaneously aggregates and rapidlylooses its activity. On the other hand, the purified recombinant[5]. In both cases, over-expression of plant serine acetyltransfer-
ase was confirmed by activity measurements in crude extractsO-acetylserine (thiol) lyase that was resolved from the bienzymecomplex (fraction 2, Fig. 4) showed an absorption spectrum withfrom the recombinantE. coli strains (Table1). Importantly,
when the recombinant serine acetyltransferase was expressed two maxima located at 278 nm and 409 nm, typical of pyridoxal-P-containing enzymes [5]. The absorbance ratio,A278/A409, wasalong with the plantO-acetylserine (thiol) lyase, its specific ac-
tivity was increased 2.6-fold compared to that from the bacterial 3.2, in agreement with previous results [5]. The purity of theenzyme preparation was also checked by SDS/PAGE. Thiscells expressing only the plant serine acetyltransferase (Table1).
When a gel-filtration experiment was conducted with an ex- analysis revealed the expected polypeptide of 36 kDa for therecombinantO-acetylserine (thiol) lyase [5].tract of E. coli cells co-expressing both the plant serine acetyl-
transferase andO-acetylserine (thiol) lyase, two peaks of serine The native molecular mass of the recombinant bienzymecomplex (fraction1, Fig. 4A) was estimated to be in the orderacetyltransferase activity were revealed (Fig. 3B). Here, the one
with a molecular mass of 2000 kDa represented only 20% of the of 300 kDa. Since the 34-kDa and 36-kDa polypeptides revealed

241Droux et al. (Eur. J. Biochem. 255)
ing concentrations of recombinantO-acetylserine (thiol) lyase,in the range of 0210 µM subunit (Fig. 5). Reactions were car-ried out in the presence of saturating concentrations ofL-serine(10 mM) and sulfide (2.5 mM). After a1-min incubation, reac-tions were initiated by adding a saturating amount of 2 mM ace-tyl-CoA. Both the extent ofO-acetylserine accumulation and therate of cysteine synthesis were measured. In the absence ofO-acetylserine (thiol) lyase, the rate ofO-acetylserine formationwas maximal and no cysteine production could be detected, tes-tifying to the purity of the recombinant serine acetyltransferase(Fig. 5). The results also indicated that the production of cyste-ine only became significant when a molar excess ofO-acetylser-ine (thiol) lyase over serine acetyltransferase was added. Indeed,maximal production of cysteine was achieved at a concentrationof <2 µM of O-acetylserine (thiol) lyase, which corresponds, ona subunit basis, to a 60-fold excess ofO-acetylserine (thiol) ly-ase over serine acetyltransferase. TheL-cysteine data were ana-lysed in terms of the following equilibrium:
Serine acetyltransferase1 O-acetylserine (thiol) lyase
78889Kd
Cysteine synthase
L-Serine1 acetyl-CoA1 sulfide
–––––––→cysteine synthaseL-Cysteine1 acetate
which can be described by Eqn (3) (see Materials and Methods).Non-linear regression analysis of the experimental data accord-ing to this equation yielded a value of 41 nM for Kd and theFig. 5. Dependence of the rate of cysteine synthesis and extent ofO-best-fit curve shown in Fig. 5 (top). It is worth noting that, foracetylserine accumulation on the concentration of recombinantO-a stoichiometric ratio of serine acetyltransferase andO-ace-acetylserine (thiol) lyase at constant concentration of recombinanttylserine (thiol) lyase subunits (as found in the bienzyme com-serine acetyltransferase.Assays (150 µl) were carried out in the pres-
ence of 38 nM recombinant serine acetyltransferase subunit (fraction 3,plex, see above for the stoichiometry between subunits and thesee Fig. 4),10 mM L-serine, 2.5 mM sulfide,1 mM dithiothreitol and pyridoxal-P content of the bienzyme complex), only 20% of therecombinantO-acetylserine (thiol) lyase (470 U · mg21) whose subunit maximal potential ofL-cysteine synthesis was obtained. Similarconcentration was varied in the indicated range. The reactions were initi-results were obtained with theE. coli O-acetylserine (thiol) lyaseated by the addition of 2 mM acetyl-CoA and incubations were foras the varied enzyme. In this case, the experimental data fitted5 min. After addition of 50µl 20% (mass/vol.) trichloroacetic acid, the
well with a simple hyperbolic equation, yielding aKd value ofsupernatant was divided in two equal fractions used for cysteine andO-112 nM (data not shown).acetylserine determinations as described in Materials and Methods.
We also measured the accumulation ofO-acetylserine underPoints are experimental; the solid line in the top panel is from a fit usingthe conditions of Fig. 5. Here, the behavior was more complexEqn (3) withVmax5 (12.960.18) nmol · min21 ; Kd 5 (4168) nM; c0 5
(37.860.12) nM. The solid line in the bottom panel has no theoreticalthan that seen for cysteine synthesis (Fig. 5, bottom) and thesignificance. data exhibited a biphasic behavior. We interpret these data as
showing that: (a) fresh preparations of free serine acetyltransfer-ase are active inO-acetylserine formation; (b) serine acetyl-transferase andO-acetylserine (thiol) lyase interact to form a
by SDS/PAGE were present in nearly stoichiometric amountscysteine-synthase-like complex [lowO-acetylserine (thiol) lyase(Fig. 4B, lane1), the recombinant bienzyme complex containsconcentration range] that is also active inO-acetylserine forma-four subunits of each serine acetyltransferase andO-acetylserine tion but poorly active in cysteine synthesis, in agreement with(thiol) lyase. The absorption spectrum of the complex exhibitedthe results in Fig. 2 ; (c) additionalO-acetylserine (thiol) lyasetwo maxima at 278 nm and 416 nm. The absorbance ratio,A278/ molecules are required in combination with the bienzyme com-A416, was of the order of 7, which compares well with the value plex to achieve complete utilization of the synthesizedO-ace-of 8 reported for pureSalmonella typhimuriumcysteine synthase tylserine, also in agreement with the results in Fig. 2. In sum-[7]. Assuming an absorption coefficient of 7.9 ·103 M21 cm21 mary, these data fit the idea that serine acetyltransferase pro-for O-acetylserine-(thiol)-lyase-bound pyridoxal-P [7], a content duces a build-up ofO-acetylserine along the reaction path andof approx. 4 mol pyridoxal-P/mol recombinant bienzyme com- that the full capacity for cysteine synthesis can only be achievedplex can be estimated. Similar results were obtained for the het-in the presence of a large excess of exogenousO-acetylserineerologous bienzyme complex composed of recombinant serine (thiol) lyase.acetyltransferase andE. coli O-acetylserine (thiol) lyase. Weconclude that the molecular organization of the plant bienzymeEvidence that the recombinant cysteine-synthase complexcomplex is the same as that reported for bacterial cysteine syn-does not dissociate during catalysis.The data in Fig. 4 indicatethase [7]. that O-acetylserine is capable of dissociating the recombinant
bienzyme complex. We note from the titration curves reportedin Fig. 5 thatO-acetylserine accumulated to substantial amounts,Titration of serine acetyltransferase activity with O-ace-
tylserine (thiol) lyase. The rate of cysteine synthesis was mea- particularly in the low concentration range ofO-acetylserine(thiol) lyase. Thus, one may argue that during catalysis the com-sured at a constant concentration of freshly prepared recombi-
nant serine acetyltransferase (3.8 nM subunit) and with increas- plex will dissociate. To address this question, a protein extract
242 Droux et al. (Eur. J. Biochem. 255)
Table 2. Kinetic parameters for recombinantArabidopsis thalianafree or bound in a bienzyme complex (cysteine synthase) with recombinantchloroplastic or E. coli O-acetylserine (thiol) lyase.Serine acetyltransferase activity measurements and construction of overexpression plasmids(pET9d-SATand pET29a-SAT) were as described under Materials and Methods. The construction of expression plasmid pET3d-OASTLwas describedin [5]. Kinetic parameters for the free enzyme were determined with freshly prepared recombinantA. thalianaserine acetyltransferase. Bound serineacetyltransferase activity was measured with the cysteine-synthase assay described under Materials and Methods, in the presence of excessO-acetylserine (thiol) lyase OASTL (either the recombinant or the bacterial enzyme) and 2.5 mM sulfide.
Recombinant serine V/E t Kappm for 1025 V/KEt for h for
acetyltransferaseserine acetyl-CoA serine acetyl-CoA serine acetyl-CoA
s21 mM M21 · s2!
Free 330 2.84 0.91 1.16 3.63 1.16 1.16(610) (60.22) (60.05) (60.04) (60.11) (60.05) (60.09)
Bound 212 1.00 0.38 2.12 4.94 2.02 1.6to chloroplastic OASTL (620) (60.03) (60.08) (60.20) (60.26) (60.12) (60.1)
Bound 108.6 1.38 0.3 0.79 3.6 1.3 1.5to bacterial OASTL (68.4) (60.14) (60.02) (60.06) (60.3) (60.08) (60.1)
Kinetic parameters of the resolved recombinant and bacterialfrom E. coli cells co-expressing the recombinant serine acetyl-transferase andO-acetylserine (thiol) lyase was submitted to gel-O-acetylserine (thiol) lyase were also compared to those of the
corresponding enzymes bound to the recombinant serine acetyl-filtration chromatography. As noted before (see Fig. 3), twopeaks ofO-acetylserine (thiol) lyase activity were revealed, cor- transferase (Table 3). For the resolved enzymes these parameters
were essentially identical to those previously reported for freeresponding to the freeO-acetylserine (thiol) lyase (molecularmass <70 kDa) and to the enzyme bound to serine acetyl-O-acetylserine (thiol) lyase either fromE. coli [32] or spinach
chloroplasts [5]. Consistent with previous findings [5], the planttransferase (molecular mass<300 kDa). In the presence of0.1 mM O-acetylserine (i.e. a concentration at whichO-ace- enzyme exhibited positive kinetic co-operativity with respect to
amino-acid and nucleophilic substrates (Table 3). In markedtylserine accumulated before cysteine synthesis was detectable;Fig. 5), all the serine acetyltransferase was recovered in a high- contrast, the formation of bienzyme complexes resulted in a very
dramatic decrease in the catalytic efficiency of boundO-ace-molecular-mass fraction of greater than 2000 kDa, while onlyfree O-acetylserine (thiol) lyase could be detected (data not tylserine (thiol) lyase, of 65-fold for the plant enzyme and 4-fold
for the bacterial enzyme (Table 3). Presumably, this differenceshown). These data, therefore, confirmed that an accumulationof O-acetylserine above 0.1 mM can induce the dissociation of between the two boundO-acetylserine (thiol) lyases reflected
differences in the interactions with the plant serine acetyl-the bienzyme complex and that, once dissociated, the free plantserine acetyltransferase has a high propensity to form high-mo- transferase, thus leading to differences in the accessibility ofO-
acetylserine to the respective active sites. It is noteworthy thatlecular-mass aggregates. However, when the gel filtration wasconducted in the presence of 0.1 mM O-acetylserine and the apparentKm for O-acetylserine was 7-fold higher for the
plant O-acetylserine (thiol) lyase bound to recombinant serine0.25 mM sulfide, all the serine acetyltransferase activity was re-covered in the fraction of molecular mass 300 kDa, meaning that acetyltransferase compared to that for the free enzyme. Interest-
ingly, upon binding to serine acetyltransferase, this decreaseddissociation of the bienzyme complex was prevented under suchconditions. Presumably, as for bacterial cysteine synthase [7, affinity was accompanied with a transition from a model dis-
playing positive co-operativity with respect toO-acetylserine31], sulfide counteracted the action ofO-acetylserine, andstrengthened the interactions between the serine acetyltransfer- substrate to a typical Michaelis-Menten model (Table 3). Also,
the apparentKm value for sulfide was1.5-fold higher with thease andO-acetylserine (thiol) lyase molecules within the bien-zyme complex. We conclude that, under the conditions of Fig. 5, boundO-acetylserine (thiol) lyase compared to that for the free
enzyme. We conclude that interactions of plant serine acetyl-serine acetyltransferase remained firmly bound toO-acetylserine(thiol) lyase. transferase with plantO-acetylserine (thiol) lyase strongly modi-
fied the kinetic properties of these enzymes.Protein−protein interactions within cysteine synthase modifythe kinetic properties of component enzymes.Fresh prepara-tions of the recombinant serine acetyltransferase obeyed Mi-DISCUSSIONchaelis-Menten kinetics and exhibited a maximum specific ac-tivity of about 700 U · mg21 (Table 2). For comparison, the natu- The cysteine synthase described originally inS. typhimurium
[6, 7] is one bienzyme complex composed of serine acetyl-ral enzyme purified from spinach stroma had a specific activityof 350 U · mg21 [10]. In contrast, when the recombinant enzyme transferase andO-acetylserine (thiol) lyase in a1:1 subunit mo-
lar ratio [4]. There is also evidence for the existence of such awas assayed for cysteine synthesis under the conditions withsulfide and in the presence of an excess of recombinantO-ace- complex in plants [9214], which led Saito et al. [12] to suggest
that this complex is required to channel the metabolite interme-tylserine (thiol) lyase, there was a marked decrease in the appar-entKm for serine and acetyl-CoA, and a transition from a typical diateO-acetylserine, as was also originally postulated in our ini-
tial characterization of serine acetyltransferase andO-acetylser-Michaelis-Menten model to a model displaying positive kineticco-operativity with respect to both substrates (Table 2). Similar ine (thiol) lyase activities in spinach leaves [9].
The results presented in this paper confirm and extend ourfindings were observed when assaying the plant serine acetyl-transferase in the presence of the bacterialO-acetylserine (thiol) previous finding on the existence of complexes composed of
serine acetyltransferase andO-acetylserine (thiol) lyase inlyase (Table 2).

243Droux et al. (Eur. J. Biochem. 255)
Table 3. Kinetic parameters for recombinant chloroplastic orE. coli O-acetylserine (thiol) lyase free or in complex (cysteine synthase) withrecombinant A. thaliana serine acetyltransferase.O-acetylserine (thiol) lyase (lyase) activity was measured as described under Materials andMethods. Bound stands for the bienzyme complexes of either recombinant orE. coli O-acetylserine (thiol) lyase with recombinant serine acetyl-transferase.
OASTL V/Et Kappm for 1025 V/KEt for h for
Ser(Ac) sulfide Ser(Ac) sulfide Ser(Ac) sulfide
s21 mM M21 · s2!
Free (recombinant) 462 0.64 0.37 7.22 12.4 1.7 1.5(616) (60.07) (60.03) (60.25) (60.5) (60.2) (60.1)
Bound (recombinant) 7.1 4.28 0.55 0.017 0.13 1.1 1.9(60.3) (60.53) (60.04) (60.001) (60.01) (60.1) (60.2)
Free (bacterial enzyme) 353 0.64 0.58 5.5 6.3 1.1 1.4(625) (60.04) (60.06) (60.4) (60.2) (60.1) (60.1)
Bound (bacterial enzyme) 88 0.84 1.04 1.05 1.02 1 1(635) (60.10) (60.13) (60.42) (60.17) (60.1) (60.1)
plants. Furthermore, we have obtained new data showing that bienzyme complex to acquire novel catalytic properties as com-pared to those of the free species. Thus, bound serine acetyl-such a complex can form with a plant serine acetyltransferase
and anO-acetylserine (thiol) lyase of either plant or bacterial transferase exhibited both higher apparent affinity and positivekinetic cooperativity for its substrates, acetyl-CoA andL-serine,origin. From their basic physicochemical properties such com-
plexes are reminiscent of those previously documented for the which might offer the possibility of fine regulation of the overallrate of cysteine synthesis in plants. Also, the kinetic propertiesbacterial cysteine synthase [6, 7], exhibiting a molecular mass
in the order of 300 kDa and containing two 72-kDa molecules ofO-acetylserine (thiol) lyase bound to serine acetyltransferasemarkedly differed from those of the free enzyme. Thus, uponof homodimericO-acetylserine (thiol) lyase and one136-kDa
molecule of homotetrameric serine acetyltransferase. In complex formation, the plant enzyme displayed a strongly re-duced activity, with aV/Et for the bound enzyme that was 65-agreement with this finding, the yeast two-hybrid system al-
lowed to assign two domains responsible for protein2protein fold smaller than that for the free enzyme. Furthermore, the ap-parentKm for O-acetylserine was increased about 7-fold and theinteractions inA. thalianaserine acetyltransferase. One is aβ-
sheet region located at the C-terminal domain of the enzyme specificity constants (V/KEt) were strongly reduced (400-fold forO-acetylserine and100-fold for sulfide). Thus, bothO-acetylser-that is involved inO-acetylserine (thiol) lyase binding [13]. We
note that this domain includes the putative transferase domain ine and sulfide have diminished access to the enzyme activesite. Again, these features make unlikely a role of the boundO-involved in the binding ofL-serine and acetyl-CoA substrates
[18], which might account for the differences observed between acetylserine (thiol) lyase in cysteine synthesis to favorO-ace-tylserine channeling. Finally, we observed that the two substrateskinetic parameters of the free andO-acetylserine-(thiol)-lyase-
bound recombinant serine acetyltransferase (Table 2). The other ofO-acetylserine (thiol) lyase exert antagonist effects on thestability of the bienzyme complex. We do not know yet whetheris located in the central part of theA. thaliana serine acetyl-
transferase protein and is responsible for oligomerization of the these molecules bind to theO-acetylserine (thiol) lyase activesite or to effector sites. It is striking that the gain in structuralenzyme [13]. This domain thus differs from that responsible for
interaction withO-acetylserine (thiol) lyase. Our present results and catalytic properties of plant serine acetyltransferase isachieved at the expense ofO-acetylserine (thiol) lyase since,strongly support the hypothesis that these two types of interac-
tions are mutually exclusive so that the presence ofO-acetylser- upon complex formation, the latter enzyme becomes almostcompletely inactive and behaves as if it were a structural and/orine (thiol) lyase favors the formation of a cysteine-synthase-like
complex at the expense of multimers of serine acetyltransferase. regulatory subunit of serine acetyltransferase. This, in turn, im-plies the existence of free, active,O-acetylserine (thiol) lyase toInterestingly, the extent of formation of these multimers varies
with the nature of theO-acetylserine (thiol) lyase, the plantO- achieve efficient production of cysteine synthesis. Thus, the pre-sent observations might have a physiological significance sinceacetylserine (thiol) lyase being more efficient than the bacterial
enzyme in preventing the formation of aggregated plant serine in plants all the serine acetyltransferase activity is recovered asa form bound toO-acetylserine (thiol) lyase, whereas, in con-acetyltransferase.
It is clear from the present study that protein2protein in- trast, most of theO-acetylserine (thiol) lyase is free [9,10, 14,33]. Further work is in progress to define whetherO-acetylserineteractions leading to the formation of cysteine-synthase com-
plexes from plant serine acetyltransferase do not allow a chan- (thiol) lyase molecules exchange between the bienzyme com-plexes and the free pool during catalysis.neling mechanism for theO-acetylserine intermediate (Figs 2
and 5). Instead,O-acetylserine accumulated catalytically with In summary, the present data demonstrate that: (a) a highratio of O-acetylserine (thiol) lyase over serine acetyltransferaserespect to serine acetyltransferase. Only in the presence of ex-
cess exogenousO-acetylserine (thiol) lyase was such an accu- is necessary to solubilize the plant serine acetyltransferase and,upon complex formation, the two enzymes acquire novel cata-mulation of O-acetylserine prevented, conditions which were
also associated with achievement of maximal production ofL- lytic properties as compared to that of the free species; (b) plantO-acetylserine (thiol) lyase bound to serine acetyltransferase iscysteine. This finding contradicts the widely held belief that pro-
tein2protein interactions in bienzyme complexes are an im- mostly inactive in cysteine synthesis; (c) theO-acetylserine in-termediate is released in the bulk solution, and no channeling isportant component of catalysis. Our present results indicate an-
other role for such interactions, allowing both enzymes in the observed; (d) freeO-acetylserine (thiol) lyase serves as an auxil-

244 Droux et al. (Eur. J. Biochem. 255)
11. Nakamura, K. & Tamura, G. (1990) Isolation of serine acetyl-iary enzyme to achieve full capacity for cysteine synthesis.transferase complexed with cysteine synthase fromAllium tubero-These features are depicted in model 3 of Fig.1.sum, Agric. Biol. Chem. 54, 6492656.The observed behavior of the free and bound enzymes in-
12. Saito, K., Yokoyama, H., Noji, M. & Murakoshi, I. (1995) Molecu-volved in cysteine synthesis in plants is not without precedent.lar cloning and characterization of a plant serine acetyltransferase
For example, Avilan et al. [34] and Lebreton et al. [35] recently playing a regulatory role in cysteine biosynthesis from water-reported the kinetic properties of a bienzyme complex from melon,J. Biol. Chem. 270, 16321216326.Chlamydomonas reinhardtiicomposed of two non-consecutive13. Bogdonova, N. & Hell, R. (1997) Cysteine synthesis in plants: pro-enzymes acting in the Benson-Calvin cycle, namely phospho- tein-protein interactions of serine acetyltransferase fromArabi-
dopsis thaliana, Plant J. 11, 2512262.ribulokinase and glyceraldehyde-3-phosphate dehydrogenase.14. Lunn, J. E., Droux, M., Martin, J. & Douce, R. (1990) LocalizationThese authors observed that oxidized free phosphoribulokinase
of ATP-sulfurylase andO-acetylserine (thiol) lyase in spinachwas almost completely devoid of activity. However, when it wasleaves,Plant Physiol. 94, 134521352.bound to glyceraldehyde-3-phosphate dehydrogenase, it dis-
15. Rolland, N., Droux, M., Lebrun, M. & Douce, R. (1993) O-Ace-played significant activity. In striking analogy with our presenttylserine (thiol) lyase from spinach (Spinacia oleraceaL.) leaf:results, the two substrates of phosphoribulokinase, ATP and ri- cDNA cloning, characterization and overexpression inEscheri-
bulose 5-phosphate, altered the interactions between the two en- chia coli of the chloroplast isoform,Arch. Biochem. Biophys. 300,zymes in the bienzyme complex, with ATP destabilizing and 2132222.ribulose 5-phosphate stabilizing the complex. Both these two16. Smith, I. K. & Thompson, J. F. (1969) The synthesis ofO-acetylser-
ine by extracts prepared from higher plants,Biochem. Biophys.examples illustrate, therefore, the potential role of spatial distri-Res. Commun. 35, 9392945.bution of enzyme activities for the regulation of metabolic path-
17. Bogdonova, N., Bork, C. & Hell, R. (1995) Cysteine biosynthesisways.in plants : isolation and functional identification of a cDNA en-Finally, the use of the bienzyme complex (cysteine synthase)coding a serine acetyltransferase fromArabidopsis thaliana,together with free excessO-acetylserine (thiol) lyase in an ap-FEBS Lett. 358, 43247.propriate ratio would be desirable for the purpose of discovering18. Ruffet, M.-L., Lebrun, M., Droux, M. & Douce, R. (1995) Subcellu-
inhibitors of either of these enzymes, or alterating the interac- lar distribution of serine acetyltransferase fromPisum sativumandtions of these enzymes, which might have potential as antibiot- characterization of anArabidopsis thalianaputative cytosolic iso-ics, herbicides, and/or fungicides. form, Eur. J. Biochem. 227, 5002509.
19. Roberts, M. A. & Wray, J. L. (1996) Cloning and characterizationof an Arabidopsis thalianacDNA clone encoding an organellarWe are grateful to Jacques-Henri Julliard for helpful discussions andisoform of serine acetyltransferase,Plant Mol. Biol. 30, 10412to Rick DeRose for critical reading of the manuscript. This work has1049.been conducted under the BioAvenir programme financed by Rhoˆne-
20. Saito, K., Miura, N., Yamazaki, M., Hiramo, H. & Murakoshi, I.Poulenc with the contribution of theMinistere de la Recherche et de(1992) Molecular cloning and bacterial expression of cDNA en-l’Espaceand theMinistere de l’Industrie et du Commerce Exte´rieur.coding a plant cysteine synthase,Proc. Natl Acad. Sci. USA 89,807828092.
21. Hell, R., Bork, C., Bogdonova, N., Frolov, I. & Hauschild, R. (1994)REFERENCES Isolation and characterization of two cDNAs encoding for com-
partment specific isoforms ofO-acetylserine (thiol) lyase from1. Segel, I. H. (1975)Enzymes kinetics. Behavior and analysis of rapidArabidopsis thaliana, FEBS Lett. 351, 2572262.equilibrium and steady-state enzyme systems, John Wiley & Sons,
22. Hesse, H. & Altmann, T. (1995) Molecular cloning of a cysteineNew York.synthase cDNA fromArabidopsis thaliana, Plant Physiol. 108,2. Kacser, H. (1987) Control of metabolism, inThe biochemistry of8512852.plants (Davies, D. D., ed.) vol.11, pp. 39267, Academic Press,
23. McClure, G. D. Jr & Cook, P. F. (1994) Product binding toA-car-San Diego.boxyl subsite results in a conformational change at the active site3. Srere, P.-A. (1987) Complexes of sequential metabolic enzymes,of O-acetylserine sulfhydrylase-A: Evidence from fluorescenceAnnu. Rev. Biochem. 56, 892124.spectroscopy,Biochemistry 33, 167421683.4. Kredich, N. M. (1987) Biosynthesis of cysteine, in Escherichia coli
24. Gaitonde, M. K. (1967) A spectrophotometric method for the directand Salmonella typhimurium: cellular and molecular biologydetermination of cysteine in the presence of other naturally occur-(Neidhardt, F. C., Ingraham, J. L., Low, K. B., Magasanik, M.,ring amino acids,Biochem. J. 104, 6272633.Schaechter, M. & Umberger, H. E., eds) pp. 4192428, American
25. Nakamura, K., Hayama, A., Masada, M., Fukushima, K. & Tamura,Society of Microbiology, Washington DC.G. (1987) Measurement of serine acetyltransferase activity in5. Rolland, N., Ruffet, M.-L., Job, D., Douce, R. & Droux, M. (1996)crude plant extracts by a coupled assay system using cysteineSpinach chloroplast O-acetylserine (thiol) lyase exhibits two cata-synthase,Plant Cell Physiol. 28, 8852891.lytically non-equivalent pyridoxal-5′-phosphate-containing active
26. Bradford, M. M. (1976) A rapid and sensitive methods for the quan-sites,Eur. J. Biochem. 236, 2722282.tification of microgram quantities of proteins utilizing the prin-6. Kredich, N. M. & Tomkins, G. M. (1966) The enzymatic synthesisciple of protein-dye binding,Anal. Biochem. 72, 2482254.of L-cysteine inEscherichia coliandSalmonella typhimurium, J.
27. Scopes, R. K. (1974) Measurements of protein by spectrophotome-Biol. Chem. 241, 495524965.try at 205 nm,Anal. Biochem. 59, 2772282.7. Kredich, N. M., Becker, M. A. & Tomkins, G. M. (1969) Purifica-
28. Chua, N. H. (1980) Electrophoresis analysis of chloroplast proteins,tion and characterization of cysteine synthase, a bifunctional pro-Methods Enzymol. 69, 4342436.tein complex, fromSalmonella typhimurium, J. Biol. Chem. 244,
29. Kredich, N. M. (1992) The molecular basis for positive regulation242822439.of cyspromoters inSalmonella typhimuriumandEscherichia coli,8. Becker, M. A. & Tomkins, G. M. (1969) Pleiotropy in a cysteineMol. Microbiol. 6, 274722753.requiring mutant ofSalmonella typhimuriumresulting from al-
30. Leu, L.-S. & Cook, P. F. (1994) Kinetic mechanism of serine trans-tered protein-protein interactions,J. Biol. Chem. 244, 60232acetylase fromSalmonella typhimurium, Biochemistry 33, 266726030.2671.9. Droux, M., Martin, J., Sajus, P. & Douce, R. (1992) Purification
31. Cook, P. F. & Wedding, R. T. (1977) Initial kinetic characterizationand characterization ofO-acetylserine (thiol) lyase from spinachof the multienzyme complex, cysteine synthetase,Arch. Biochem.chloroplasts,Arch. Biochem. Biophys. 295, 3792390.Biophys. 178, 2932302.10. Ruffet, M.-L., Droux, M. & Douce, R. (1994) Purification and ki-
32. Tai, C.-H., Nalabolu, S. R., Jacobson, T. M., Minter, D. E. & Cook,netic properties of serine acetyltransferase free ofO-acetylserine(thiol) lyase from spinach leaves,Plant Physiol. 104, 5972604. P. F. (1993) Kinetic mechanisms of the A and B isoenzymes of

245Droux et al. (Eur. J. Biochem. 255)
O-acetylserine sulphydrylase fromSalmonella typhimuriumLT-2 dissociation, and reassociation of a phosphoribulokinase-glyceral-dehyde-3-phosphate dehydrogenase complex fromChlamydomo-using the natural and alternative reactants,Biochemistry 32,
643326442. nas reinhardtiichloroplasts,Eur. J. Biochem. 246, 78284.35. Lebreton, S., Gontero, B., Avilan, L. & Ricard, J. (1997) Memory33. Rolland, N., Droux, M. & Douce, R. (1992) Subcellular distribution
of O-acetylserine (thiol) lyase in cauliflower (Brassica oleracea and imprinting effects in multienzyme complexes. II. Kinetics ofthe bienzyme complex fromChlamydomonas reinhardtiiand hys-L.) inflorescence,Plant. Physiol. 98, 9272935.
34. Avilan, L., Gontero, B., Lebreton, S. & Ricard, J. (1997) Memory teretic activation of chloroplast oxidized phosphoribulokinase,Eur. J. Biochem. 246, 85291.and imprinting effects in multienzyme complexes. I. Isolation,
Copyright © 2022 FDOKUMEN




















