
Cannflavins from hemp sprouts, a novel cannabinoid-free hemp food product, target microsomal...
8
PharmaNutrition 2 (2014) 53–60 Contents lists available at ScienceDirect PharmaNutrition j o u r n a l h o m e p a g e : w w w . e l s e v i e r . c o m / l o c a t e / p h a n u Cannflavins from hemp sprouts, a novel cannabinoid-free hemp food product, target microsomal prostaglandin E 2 synthase-1 and 5-lipoxygenase Oliver Werz a, * , Julia Seegers b , Anja Maria Schaible a , Christina Weinigel a , Dagmar Barz c , Andreas Koeberle a , Gianna Allegrone d , Federica Pollastro d , Lorenzo Zampieri d , Gianpaolo Grassi e , Giovanni Appendino d, * a Department of Pharmaceutical/Medicinal Chemistry, Institute of Pharmacy, University of Jena, Philosophenweg 14, D-07743 Jena, Germany b Department for Pharmaceutical Analytics, Pharmaceutical Institute, University of T¨ ubingen, Auf der Morgenstelle 8, D-72076 Tuebingen, Germany c Institute of Transfusion Medicine, Jena University Hospital, 07743 Jena, Germany d Dipartimento di Scienze del Farmaco, Universit` a del Piemonte Orientale, Largo Donegani 2, 28100 Novara, Italy e Consiglio per le Ricerca e la sperimentazione in Agricoltura, Centro di Ricerca per le Colture Industriali, CRA, CIN, Viale G. Amendola 82, 45100 Rovigo, Italy a r t i c l e i n f o Article history: Received 8 April 2014 Received in revised form 12 May 2014 Accepted 12 May 2014 Keywords: Hemp Plant-derived food Cannflavins Inflammation Microsomal prostaglandin E2 synthase 1 5-Lipoxygenase a b s t r a c t Hemp seeds are of great nutritional value, containing all essential amino acids and fatty acids in sufficient amount and ratio to meet the dietary human demand. Hemp seeds do not contain cannabinoids, and because of their high contents of ω -3 fatty acids, are enjoying a growing popularity as a super-food to beneficially affect chronic inflammation. Seeds also lack the typical phenolics of hemp leaves and inflorescences, but we found that sprouting, while not triggering the production of cannabinoids, could nevertheless induce the production of the anti-inflammatory prenylflavonoids cannflavins A and B. This effect was especially marked in Ermo, a cannabinoid-free variety of Cannabis sativa L. Microsomal prostaglandin E 2 synthase (mPGES-1) and 5-lipoxygenase (5-LO) were identified as the molecular targets of cannflavins A and B, solving an almost three-decade old uncertainty on the mechanism of their the anti-inflammatory activity. No change on the fatty acid profile was observed during sprouting, and the presence of lipophilic flavonoids combines with the high concentration of ω -3 essential acids to qualify sprouts from Ermo as a novel anti-inflammatory hemp food product worth considering for mass production and commercial development. c 2014 Elsevier B.V. All rights reserved. 1. Introduction The nutritional virtues of hemp seeds can hardly be underesti- mated, since, virtually unique between food plants, they contain all essential amino acids and fatty acids in sufficient amount and ratio to meet the human demand [1]. Unsurprisingly, hemp was one of the first “grain” plants domesticated by man [2], and, starting from the na- tive Central Asia, hemp cultivation spread all over the world because of its multipurpose use as a source of nutrition and fiber as well as a medicinal/ritual plant [2]. Per se, hemp seeds do not contain cannabi- noids, but, just like the seeds from poppy or peppers, their surface can be contaminated by compounds present in the plant matrix in which they are hosted, eventually becoming a source of trace amounts of them (alkaloids for poppy seeds, capsaicinoids for hot pepper seeds, cannabinoids for hemp seeds) [1]. Because of regulatory limits on the contents of psychoactive cannabinoids in hemp products, hemp seeds are produced from strains bred for low tetrahydrocannabinol (THC) * Corresponding authors. E-mail addresses: [email protected] (O. Werz) [email protected] (G. Appendino). contents that would perform very poorly as sources of drug mate- rial [1]. Currently, hemp seeds are mostly used as staple in certain animals, and especially for some birds too, but hemp seed products (whole or dehulled hemp grains, hemp oil, hemp flour, hemp protein powder, hemp substitute milk, hemp tofu, hemp butter) are enjoying a growing popularity in human nutrition, where they have attained a super-food status because of a combination of healthy fatty acids composition, high contents of proteins (20–25%) and vitamins, and, last but not least, they have a pleasant nutty taste also [3]. Hemp seeds contain up to 35% of oil, with an 80% concentration of polyun- saturated acids, and a ca. 3:1 ratio between linoleic (LA, ω -6)- and α-linolenic (ALA, ω -3) acids [3]. Hemp oil is one of the very few edi- ble oils that contains significant amounts (up to 7%) of γ-linolenic acid (GLA) and stearidonic acid (SDA, 18:4n3) (up to 1.5%). GLA and SDA attenuate the formation of inflammatory prostanoids from arachi- donic acid [4], overall making hemp oil unique in terms of capacity to increase tissue concentration of n-3 polyunsaturated fatty acids and to decrease the production of inflammatory eicosanoids. Because of the anti-inflammatory profile of their fatty acid composition, hemp seed food products could therefore beneficially affect pathological conditions characterized by chronic inflammation, as suggested, for 2213-4344/$ - see front matter c 2014 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.phanu.2014.05.001
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cannflavins from hemp sprouts, a novel cannabinoid-free hemp food product, target microsomal...

PharmaNutrition 2 (2014) 53–60
Contents lists available at ScienceDirect
PharmaNutrition
j o u r n a l h o m e p a g e : w w w . e l s e v i e r . c o m / l o c a t e / p h a n u
Cannflavins from hemp sprouts, a novel cannabinoid-free hemp
food product, target microsomal prostaglandin E 2
synthase-1 and
5-lipoxygenase
Oliver Werz
a , * , Julia Seegers b , Anja Maria Schaible
a , Christina Weinigel a , Dagmar Barz
c , Andreas Koeberle
a , Gianna Allegrone
d , Federica Pollastro
d , Lorenzo Zampieri d , Gianpaolo Grassi e , Giovanni Appendino
d , *
a Department of Pharmaceutical / Medicinal Chemistry, Institute of Pharmacy, University of Jena, Philosophenweg 14, D-07743 Jena, Germany b Department for Pharmaceutical Analytics, Pharmaceutical Institute, University of T ubingen, Auf der Morgenstelle 8, D-72076 Tuebingen, Germany c Institute of Transfusion Medicine, Jena University Hospital, 07743 Jena, Germany d Dipartimento di Scienze del Farmaco, Universit a del Piemonte Orientale, Largo Donegani 2, 28100 Novara, Italy e Consiglio per le Ricerca e la sperimentazione in Agricoltura, Centro di Ricerca per le Colture Industriali, CRA, CIN, Viale G. Amendola 82, 45100 Rovigo, Italy
a r t i c l e i n f o
Article history:
Received 8 April 2014
Received in revised form 12 May 2014
Accepted 12 May 2014
Keywords:
Hemp
Plant-derived food
Cannflavins
Inflammation
Microsomal prostaglandin E 2 synthase
1
5-Lipoxygenase
a b s t r a c t
Hemp seeds are of great nutritional value, containing all essential amino acids and fatty acids in sufficient
amount and ratio to meet the dietary human demand. Hemp seeds do not contain cannabinoids, and because
of their high contents of ω -3 fatty acids, are enjoying a growing popularity as a super-food to beneficially
affect chronic inflammation. Seeds also lack the typical phenolics of hemp leaves and inflorescences, but we
found that sprouting, while not triggering the production of cannabinoids, could nevertheless induce the
production of the anti-inflammatory prenylflavonoids cannflavins A and B. This effect was especially marked
in Ermo, a cannabinoid-free variety of Cannabis sativa L. Microsomal prostaglandin E 2 synthase (mPGES-1)
and 5-lipoxygenase (5-LO) were identified as the molecular targets of cannflavins A and B, solving an almost
three-decade old uncertainty on the mechanism of their the anti-inflammatory activity. No change on the
fatty acid profile was observed during sprouting, and the presence of lipophilic flavonoids combines with the
high concentration of ω -3 essential acids to qualify sprouts from Ermo as a novel anti-inflammatory hemp
food product worth considering for mass production and commercial development. c © 2014 Elsevier B.V. All rights reserved.
1. Introduction
The nutritional virtues of hemp seeds can hardly be underesti-
mated, since, virtually unique between food plants, they contain all
essential amino acids and fatty acids in sufficient amount and ratio
to meet the human demand [ 1 ]. Unsurprisingly, hemp was one of the
first “grain” plants domesticated by man [ 2 ], and, starting from the na-
tive Central Asia, hemp cultivation spread all over the world because
of its multipurpose use as a source of nutrition and fiber as well as a
medicinal / ritual plant [ 2 ]. Per se , hemp seeds do not contain cannabi-
noids, but, just like the seeds from poppy or peppers, their surface can
be contaminated by compounds present in the plant matrix in which
they are hosted, eventually becoming a source of trace amounts of
them (alkaloids for poppy seeds, capsaicinoids for hot pepper seeds,
cannabinoids for hemp seeds) [ 1 ]. Because of regulatory limits on the
contents of psychoactive cannabinoids in hemp products, hemp seeds
are produced from strains bred for low tetrahydrocannabinol (THC)
* Corresponding authors.
E-mail addresses: [email protected] (O. Werz)
[email protected] (G. Appendino).
2213-4344/ $ - see front matter c © 2014 Elsevier B.V. All rights reserved.
http://dx.doi.org/10.1016/j.phanu.2014.05.001
contents that would perform very poorly as sources of drug mate-
rial [ 1 ]. Currently, hemp seeds are mostly used as staple in certain
animals, and especially for some birds too, but hemp seed products
(whole or dehulled hemp grains, hemp oil, hemp flour, hemp protein
powder, hemp substitute milk, hemp tofu, hemp butter) are enjoying
a growing popularity in human nutrition, where they have attained
a super-food status because of a combination of healthy fatty acids
composition, high contents of proteins (20–25%) and vitamins, and,
last but not least, they have a pleasant nutty taste also [ 3 ]. Hemp
seeds contain up to 35% of oil, with an 80% concentration of polyun-
saturated acids, and a ca. 3:1 ratio between linoleic (LA, ω -6)- and
α-linolenic (ALA, ω -3) acids [ 3 ]. Hemp oil is one of the very few edi-
ble oils that contains significant amounts (up to 7%) of γ-linolenic acid
(GLA) and stearidonic acid (SDA, 18:4n3) (up to 1.5%). GLA and SDA
attenuate the formation of inflammatory prostanoids from arachi-
donic acid [ 4 ], overall making hemp oil unique in terms of capacity to
increase tissue concentration of n -3 polyunsaturated fatty acids and
to decrease the production of inflammatory eicosanoids. Because of
the anti-inflammatory profile of their fatty acid composition, hemp
seed food products could therefore beneficially affect pathological
conditions characterized by chronic inflammation, as suggested, for

54 O. Werz et al. / PharmaNutrition 2 (2014) 53–60
i
d
c
t
h
i
t
s
v
b
a
a
m
p
p
t
S
o
s
t
p
l
e
o
p
B
I
f
n
a
a
a
i
d
2
2
c
m
f
5
o
2
h
v
v
i
i
n
t
a
nstance, by a clinical study on atopic dermatitis [ 5 ], and various anec-
otal reports [ 5 ]. By decreasing platelet aggregation, hemp seed oil
ould also reduce cardiovascular risk [ 6 ].
In the context of hemp seed food, little attention has been given
o sprouts, despite their possibility to expand the nutritional use of
emp not only in culinary terms, but also phytochemically, exploit-
ng the expression of new metabolic pathways induced by germina-
ion. Germination-associated changes in the phytochemical profile of
eeds are well documented [ 7 ], and have spurred the commercial de-
elopment of sprouts enriched in specific phytochemicals, as shown
y sulforaphane-rich broccoli sprouts [ 8 ]. Quantitative changes in the
mount of specific phytochemicals have also been reported, as well
s a general remodeling of the whole profile of specific secondary
etabolites. Thus, the triterpenoid profile of pumpkin seeds is com-
letely changed by sprouting, with the replacement of multiflorane
-aminobenzoate triterpenoid esters with bryonolic acid [ 9 ], a potent
riterpene inducer of anti-inflammatory and antioxidant genes [ 10 ].
ince the production of cannabinoid is associated with the presence
f specific glandular hairs typical of foliar and floral hemp tissues [ 11 ],
prouting was unlikely to induce their synthesis or accumulation. On
he other hand, hemp is a prolific producer of structurally unique
henolics (cannflavins, cannabispiranes, canniprenes), present in the
eaves and flowers, but absent in the seeds [ 12 ]. Since beneficial prop-
rties have been associated to these compounds [ 13 ], the induction
f their production by sprouting might, in principle, adds value to the
roduct.
In this context, we have focused on cannflavin A (CVA, 1 ) and
(CVB, 2 ), a pair of prenylated flavonoids unique to hemp [ 14 ].
nhibition of prostanoid formation has previously been demonstrated
or cannflavins [ 15 ], implying anti-inflammatory activity that might
icely complement one of the ω -3 fatty acids present in the seeds,
nd overall increasing the value of hemp seed products as an
nti-inflammatory nutritional support. Critical to this project was
lso the clarification of the mechanism(s) by which cannflavins
nhibit prostanoid synthesis, an issue that, despite almost three-
ecade of studies on these compounds [ 15 ], is still unresolved.
. Materials and methods
.1. General phytochemical procedures
Silica gel 60 (70–230 mesh) and RP-18 used for gravity column
hromatography were purchased from Macherey-Nagel (D uren, Ger-
any). Aluminum-coated Merck 60 F254 (0.25 mm) plates were used
or TLC, visualizing the spots by UV inspection and / or staining with
% H 2 SO 4 in ethanol and heating. Flash chromatography was carried
ut in a Biotage apparatus SP1.
.2. Plant material
Hemp seeds were either purchased (Finola) or obtained from in
ouse cultivations at CRA, Rovigo, (Ermo, Carma and a THC-containing
ariety), where also the aerial parts of the variety Ermo were har-
ested. Ermo is a monoecious medium-size ( ca . 2 m high when grown
n temperature climate) hemp variety. It blooms relatively early, and
s derived from the Ermes variety, with whom Ermo shares the pin-
atifidus shape of the leaves, a rare feature in hemps. Ermo is resistant
o pests as well as parasite plants like those from the genus Orobanche ,
nd shows a good tolerance to salinity.
2.3. Production of sprouts
Seeds (50 g) were washed in running water, sterilized with a short
(5 min) treatment with hydrogen peroxide, and then rinsed with
running water. Germination was carried out in a VitaSeed domestic
equipment for 5 days under dark, with changing the water every day.
The sprouted seeds were washed, and, after removing their tegument
manually, were dried in an oven at 50 ◦C for 2 days.
2.4. Isolation of cannflavin A from C. sativa var. Ermo
Powdered dried plant material (900 g) was twice extracted with
acetone (ratio plant–solvent 1:9), to afford, after evaporation 32 g
(3.5%) of a dark oil, that was dissolved in the minimal amount of
methanol and adsorbed on a bed of RP18 silica gel (160 g). Vacuum-
filtration with methanol afforded a de-pigmented and de-waxed frac-
tion (12 g) as a brownish gum, next purified by gravity column chro-
matography on silica gel using petroleum ether–EtOAc as eluent (from
7:3 to 5:5). A first crop of CFA( 1 ) (80 mg) was obtained from the more
polar fractions of the column. The mother liquors (880 mg) were fur-
ther purified by flash-chromatography on silica gel, using petroleum
ether–EtOAc 6:4 as eluent to afford a fraction that crystallized to pro-
vide 15 further mg of CFA( 1 ). The mother liquors (280 mg) contained
additional amounts of CFA( 1 ) that, however, could not be induced
to crystallize and were impure for the presence of other phenolics,
including CFB( 2 ). The overall yield of crystalline CFA( 1 ) was 95 mg
(0.011% on dried plant material). By NMR-inspection of the mother
liquors, an estimated similar amount of CFA( 1 ) was present in the
mother liquors.
2.5. Synthesis of cannflavin B
The published protocol [ 14 ] was used without any significant mod-
ification.
2.6. Determination of the lipid contents of seeds and sprouts
The overall amounts of lipids was estimated by exhaustive hexane
extraction. Oven-dried (heating in a ventilated oven at 50 ◦C for 48 h)
powdered seeds and sprouts (10 g each) were extracted with hexane
at room temperature (3 × 100 mL). Evaporation of the pooled extracts
afforded an oily residue, that, when analyzed by NMR, only showed
the signal of triglycerides. Triplicate analyses on seeds and sprouts
from Ermo gave a lipid contents of 36 ± 1.5% for the seeds, and
31 ± 1.2% for the sprouts.
2.7. HPLC–MS determination of cannflavins in sprouts and seeds
Dried and powdered sprouts or seeds (1 g) were extracted with
acetone (25 mL) under stirring. After 10 min the solution was fil-
tered, and the plant residue was washed with 10 mL acetone, and
re-extracted a second time with acetone. After filtration and wash-
ing, the pooled filtrates were evaporated, and the oily residue was
analyzed by HPLC–MS. A Surveyor HPLC on line with an LCQ DECA
XP Plus (Thermo Finnigan, San Jos e, USA) Ion Trap mass spectrom-
eter equipped with an ESI source was employed. Separations were
performed on an analytical Luna 5 μm C18(2), 150 mm × 2.0 mm
(Phenomenex, Torrance, CA) protected with a C18-Security Guard
cartridge, 4 mm × 2.0 mm (Phenomenex). The injection volume was
10 μL. The mobile phase components were: A = formic acid 0.2%; B =
acetonitrile; the cannflavins were eluted according to the following
linear gradient: A:B (50:50) for 2 min, then A:B (0:100) over 22 min
at flow rate of 0.4 mL / min. The ESI-MS and ESI-MS / MS spectra of
the cannflavins were recorded using direct infusion of each reference
compounds. Data were acquired in positive and negative MS total ion
scan mode (mass scan range: m / z 100–1000) and in negative MS / MS

O. Werz et al. / PharmaNutrition 2 (2014) 53–60 55
product ion scan mode; the normalized collision energy (nce %) was
optimized for each precursor ion selected: m / z 367, 44%; m / z 435,
45%. LC–ESI-MS / MS in single reaction monitoring (SRM) modalities
were applied to the selected precursor ions, following the conditions
set during the infusion analysis. The parent / daughter ion transitions
m / z 367 → 352 and m / z 367 → 309 were selected for the identifica-
tion and quantification of CFB( 2 ) while the transitions m / z 435 → 420
and m / z 435 → 351 were selected for the identification and quan-
tification of CFA( 1 ). The calibration curves, obtained by injecting six
different concentration levels of CFB( 2 ) and CFA( 1 ) standard solu-
tions and analyzed in triplicate, showed a good linearity in the range
of 5–1000 μg / L. The LOD was determined by the analysis of samples
of known concentrations and found to be 1 μg / L and LOQ found to
be 4 μg / L for both CFB( 2 ) and CFA( 1 ). All samples were diluted and
analyzed in triplicate according to this method.
2.8. HS-SPME / GC determination of cannabinoids in sprouts
Powdered sprouts (10 mg) were placed directly in 15-mL
headspace vial in presence of 1 mL of NaOH 1 M, 300 mg of Na 2 CO 3
and 1 μL of THC-d6 solution (100 μg / mL in ethanol) as internal stan-
dard [ 16 ]. The mixture was stirred (1000 rpm) for 40 min at 90 ◦C and
under these conditions, the PDMS-100 μm SPME fiber (Supelco Bella-
fonte, PA, USA) was exposed to the headspace of the vial to absorb the
analytes [ 16 ]. The analytes retained on the fiber were thermally des-
orbed by exposure in the chromatograph injector for 10 min. Analyses
were performed using a Trace GC gas chromatograph coupled with a
Polaris Q ion trap mass spectrometer (Thermo Finnigan, San Jos e, USA)
operating in electron ionization mode. Chromatographic separation
was achieved using an RTX-5MS capillary column (30 m × 0.25 mm
i.d., thickness 0.25 μm, Restek GmbH, Germany) kept at 150 ◦C for
2 min then ramped to 250 ◦C at 5 ◦C / min followed by a hold at 250 ◦C
for 10 min. Helium was used as carrier gas at 1.0 mL min
–1 . The injec-
tor temperature was 250 ◦C and it was operated at splitless mode for
2 min. The manifold source and the transfer line temperatures were
set to 250 ◦C and 270 ◦C, respectively. The MS was used in tandem
MS / MS mode to improve sensitivity and selectivity of the method.
The selected parent / daughter ion transitions were m / z 231 → 174,
m / z 295 → 223, m / z 299 → 231 and m / z 320 → 278, respectively for
cannabidiol (CBD), cannabinol (CBN), THC and THC-d6.
Quantification limits (LOQ) were fixed at 0.1 ng / mg for CBD,
0.1 ng / mg for CBN and 5.0 ng / mg for THC. In all sprout samples
cannabinoids were below the threshold of detection.
2.9. Comparative determination of fatty acid methyl esters in seeds and
sprouts from the hemp variety Ermo
The fatty acid methyl esters were prepared from hexane extracts
of the plant material according to literature [ 17 ], and GC–MS analysis
was performed using a Trace GC gas chromatograph coupled with a
Polaris Q ion trap mass spectrometer (Thermo Finnigan, San Jos e, USA)
operating in electron ionization mode. Chromatographic separation
was performed with an RTX-5MS capillary column (30 m × 0.25
mm i.d., thickness 0.25 μm, Restek GmbH, Germany). The program
temperature was: 2 min at 150 ◦C then 150–270 ◦C at 3 ◦C / min.
The manifold source and the transfer line temperatures were set to
250 ◦C and 270 ◦C, respectively, and the split injector was maintained
at 270 ◦C. Helium was used as carrier gas at 1.0 mL min –1 and
split ratio was used 1:50. The mass spectrometer was operated in
electron impact (70 eV) mode and full scan (33–350 m / z ) spectra
were recorded.
2.10. Cells and cell isolation
Human peripheral blood was taken from fastened (12 h) healthy
donors who had not taken any anti-inflammatory drugs during the
last 10 days, with venipuncture in heparinized tubes (16 IE heparin /mL blood). The blood was centrifuged at 4000 × g for 20 min at 20 ◦C
for the preparation of leukocyte concentrates (University Hospital
Jena, Germany). Leukocyte concentrates were subjected to dextran
sedimentation and centrifugation on Nycoprep cushions (PAA Lab-
oratories, Linz, Austria). For isolation of platelets, the supernatants
of the Nycoprep cushion centrifugation were mixed with phosphate-
buffered saline (PBS) pH 5.9 (3:2 v / v), centrifuged (2100 × g , 15 min,
20 ◦C), and the pelleted platelets were resuspended in PBS pH 5.9 /0.9% NaCl (1:1, v / v). Washed platelets were finally resuspended in PBS
pH 7.4 and 1 mM CaCl 2 . For isolation of polymorphonuclear leuko-
cytes (PMNL), contaminating erythrocytes of pelleted PMNL were
lysed by hypotonic lysis. PMNL were washed twice in ice-cold PBS
and finally resuspended in PBS pH 7.4 containing 1 mg / mL glucose
and 1 mM CaCl 2 (PGC buffer) (purity > 96–97%). Monocytes were
isolated from the peripheral blood mononuclear cell fraction, which
was obtained after Nycoprep cushion centrifugation of leukocyte con-
centrates, by adherence for 1.5 h at 37 ◦C to culture flasks (Greiner,
Nuertingen, Germany), cell density was 2 × 10 7 cells / mL RPMI 1640
medium containing 2 mM l -glutamine and 100 U / mL penicillin and
100 μg / mL streptomycin, which gave a purity of > 85%, defined by
forward- and side-light scatter properties and detection of the CD14
surface molecule by flow cytometry (BD FACS Calibur, Heidelberg,
Germany). Monocytes were finally resuspended in ice-cold PBS plus
1 mg / mL glucose or in PGC buffer.
For analysis of acute cytotoxicity of test compounds during pre-
incubation periods, the viability of PMNL was analyzed by light mi-
croscopy and trypan blue exclusion. For the analysis of cytotoxicity
of monocytes, the MTT assay was used. In brief, cells (100 μL cell
suspension corresponding to 2 × 10 5 monocytes) were incubated
for 30 min at 37 ◦C with vehicle (0.3% DMSO) or test compounds.
After 30 min, 20 μL of a sterile filtered solution of 3-(4,5-dimethyl-2-
thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) in PBS (5 mg /mL) were added and samples were incubated for 4 h at 37 ◦C. Then,
100 μL of 10% SDS in 20 mM HCl (pH 4.5; SDS lysis buffer) were added
and samples were shaken for 15 h at RT in the dark and the absorp-
tion was measured at 570 nm. Results are reported as percentage of
viable cells as compared to vehicle control. A statistically significant
impairment of MTT reduction to formazan was considered cytotoxic.
2.11. Determination of 5-lipoxygenase products in intact cells
For the determination of 5-LO products in intact PMNL (5 × 10 6 )
or monocytes (2 × 10 6 ), cells were resuspended in 1 mL PGC buffer,
preincubated for 15 min at 37 ◦C with test compounds or vehicle (0.3%
DMSO), and incubated for 10 min at 37 ◦C with the indicated stim-
uli. Thus, the Ca 2 + -ionophore A23187 (2.5 μM) was added with or
without 20 μM AA and 10 min later the reaction was stopped on ice
by addition of 1 mL of methanol. 30 μL 1 N HCL and 500 μLPBS,
and 200 ng prostaglandin B1 were added and the samples were
subjected to solid phase extraction on C18-columns (100 mg, UCT,
Bristol, PA, USA). 5-LO products (LTB 4 , trans-isomers, 5-H(p)ETE),
and the cyclooxygenase (COX)-1 product 12( S )-hydroxy-5- cis -8,10-
trans-heptadecatrienoic acid (12-HHT), respectively, were analyzed
by HPLC and quantities calculated on the basis of the internal stan-
dard PGB1. Cysteinyl-LTs C 4 , D 4 and E 4 were not detected (amounts
were below detection limit), and oxidation products of LTB 4 were not
determined.
2.12. Expression and purification of human recombinant 5-LO
Escherichia coli MV1190 was transformed with pT3-5-LO plasmid,
and recombinant 5-LO protein was expressed at 27 ◦C as described
[ 18 ]. Cells were lysed in 50 mM triethanolamine / HCl pH 8.0, 5 mM

56 O. Werz et al. / PharmaNutrition 2 (2014) 53–60
E
s
c
4
t
5
2
t
i
s
2
w
m
d
a
2
d
i
t
h
f
1
h
1
i
c
m
p
w
w
t
(
i
a
2
C
(
l
5
p
w
w
s
a
u
r
2
p
i
f
1
1
DTA, soybean trypsin inhibitor (60 μg / mL), 1 mM phenylmethane-
ulfonyl fluoride, and lysozyme (500 μg / mL), homogenized by soni-
ation (3 × 15 s), and centrifuged at 40,000 × g for 20 min at 4 ◦C. The
0,000 × g supernatant (S40) was applied to an ATP-agarose column
o partially purify 5-LO as described previously [ 18 ]. Semi-purified
-LO was immediately used for activity assays.
.13. Determination of 5-LO activity in cell-free assay
Aliquots of semi-purified 5-LO were diluted with ice-cold PBS con-
aining 1 mM EDTA, and 1 mM ATP was added. Samples were pre-
ncubated with the test compounds as indicated. After 10 min at 4 ◦C,
amples were pre-warmed for 30 s at 37 ◦C, and 2 mM CaCl 2 plus
0 μM AA was added to start 5-LO product formation. The reaction
as stopped after 10 min at 37 ◦C by the addition of 1 mL ice-cold
ethanol, and the formed metabolites were analyzed by RP-HPLC as
escribed [ 18 ]. 5-LO products include the all-trans isomers of LTB 4
nd 5(S)-hydro(pero)xy-6- trans -8,11,14- cis -eicosatetraenoic acid.
.14. Preparation of crude mPGES-1 in microsomes of A549 cells and
etermination of PGE 2 synthase activity
Preparations of A549 cells and determination of mPGES-1 activ-
ty were performed as described previously [ 19 ]. In brief, cells were
reated with 1 ng / mL Il-1 β for 48 h at 37 ◦C, 5% CO 2 . Cells were
arvested and sonicated and the homogenate was subjected to dif-
erential centrifugation at 10,000 × g for 10 min and 174,000 × g for
h at 4 ◦C. The pellet (microsomal fraction) was resuspended in 1 mL
omogenization buffer (0.1 M potassium phosphate buffer, pH 7.4,
mM phenylmethanesulfonyl fluoride, 60 μg / mL soybean trypsin
nhibitor, 1 μg / mL leupeptin, 2.5 mM glutathione, and 250 mM su-
rose), and the total protein concentration was determined. Microso-
al membranes were diluted in potassium phosphate buffer (0.1 M,
H 7.4) containing 2.5 mM glutathione. Test compounds or vehicle
ere added, and after 15 min at 4 ◦C reaction (100 μL total volume)
as initiated by addition of PGH 2 at the indicated concentration. Af-
er 1 min at 4 ◦C, the reaction was terminated using stop solution
100 μL; 40 mM FeCl 2 , 80 mM citric acid, and 10 μM 11 β-PGE 2 ) as
nternal standard. PGE 2 was separated by solid-phase extraction and
nalyzed by RP-HPLC as described, previously [ 19 ].
.15. Activity assays of isolated COX-1 and COX-2
Inhibition of the activities of isolated ovine COX-1 and human
OX-2 was performed as described [ 19 ]. Briefly, purified COX-1
ovine, 50 units) or COX-2 (human recombinant, 20 units) were di-
uted in 1 mL reaction mixture containing 100 mM Tris buffer pH 8,
mM glutathione, 5 μM hemoglobin, and 100 μM EDTA at 4 ◦C and
re-incubated with the test compound for 5 min. Samples were pre-
armed for 60 s at 37 ◦C, and AA (5 μM for COX-1, 2 μM for COX-2)
as added to start the reaction. After 5 min at 37 ◦C, the reaction was
topped, PGB 1 as standard added and 12-HHT was extracted and then
nalyzed by HPLC. Indomethacin (20 μM) and celecoxib (5 μM) were
sed as well-recognized reference inhibitors of COX-1 and of COX-2,
espectively, to control the assays.
.16. Determination of the COX-1-derived product 12-HHT in human
latelets
Freshly isolated human platelets (10 8 / mL PGC buffer) were pre-
ncubated with the test compounds for 15 min at 37 ◦C and stimulated
or 10 min at 37 ◦C with 5 μM AA. The COX reaction was stopped after
0 min at 37 ◦C by addition of 1 mL ice-cold methanol and the formed
2-HHT was analyzed by HPLC as described [ 19 ].
2.17. Determination of the activity of isolated human recombinant
cPLA 2 α in a cell-free assay
The cPLA 2 α coding sequence was cloned from pVL1393 plasmid
(kindly provided by Dr. Wonhwa Cho, University of Illinois at Chicago)
into pFastBacTM HT A containing a 6 × his-tag coding sequence. The
recombinant plasmid was transformed into D10BacTM E. coli. Sf9 cells
were transfected with recombinant bacmid DNA using Cellfectin
®
Reagent and the generated baculovirus was amplified. Overexpres-
sion of His-tagged cPLA 2 in baculovirus-infected Sf9 cells and isolation
using Ni-NTA agarose beads was performed as described [ 20 ]. Mul-
tilamellar vesicles (MLVs) were prepared by drying 1-palmitoyl-2-
arachidonyl-sn-glycero-3-phosphocholine (PAPC) and 1-palmitoyl-
2-oleoyl-sn-glycerol (POG) in a ratio of 2:1 (in chloroform) under
nitrogen in glass vials. After addition of 20 mM Tris buffer (pH 7.4)
containing 134 mM NaCl and 1 mg / mL fatty acid free BSA, the MLV
suspension was disrupted by several freeze–thaw cycles (liquid nitro-
gen) and then extruded 11 times with a mini-extruder (Avanti Polar
Lipids Inc.) through a polycarbonate membrane (100 nm pore diame-
ter) at room temperature (above transition temperature of the lipids)
to produce LUV (large unilamellar vesicles). Final total concentration
of lipids was 250 μM in 200 μL. Test compounds and 1 mM CaCl 2 were added to the vesicles, and the reaction was started by addi-
tion of 500 ng his-tagged cPLA 2 (in 10 μL buffer). After 1 h at 37 ◦C,
1.6 mL CH 3 OH was added, and AA was extracted by RP-18 solid phase
extraction. Following derivatization with p-anisidinium chloride, the
resulting derivate was analyzed by RP-HPLC at 249 nm as described
[ 20 ].
2.18. DPPH assay
The radical scavenger capability was assessed by measuring the
reduction of the stable free radical 2,2-diphenyl-1-picrylhydrazyl
(DPPH) [ 21 ] with slight modifications. Briefly, 100 μL of test com-
pound in ethanol (corresponding to 0.5, 2.5, and 5 nmol) was added to
100 μL of a solution of the stable free radical diphenylpicrylhydrazyl
(DPPH) in ethanol (50 μM, corresponding to 5 nmol), buffered with
acetate to pH 5.5, in a 96-well plate. The absorbance was recorded at
520 nm (Victor plate reader, PerkinElmer) after 30 min incubation un-
der gentle shaking in the dark. Ascorbic acid and l -cysteine were used
as reference compounds. All analyses were performed in triplicates.
2.19. Statistics
Data are expressed as mean ± S.E. IC 50 values were graphically
calculated from averaged measurements at 4–5 different concentra-
tions of the compounds using SigmaPlot 9.0 (Systat Software Inc.,
San Jose, USA). Statistical evaluation of the data was performed by
one-way ANOVA followed by a Bonferroni or Tukey–Kramer post hoc
test for multiple comparisons respectively. A p value < 0.05 (*) was
considered significant.
3. Results
3.1. Analytical characterization of hemp seeds and sprouts
Seeds and sprouts from three varieties of fiber hemp (Finola,
Carma, Ermo) and a drug-type THC-rich variety were characterized in
terms of contents of cannabinoids and cannflavins. The quantification
of cannabinoids from fatty matrixes requires special methods, due to
the difficulty of their recovery. Using a method specifically developed
for the quantification of cannabinoids in hemp oil [ 16 ], we found that
in dehulled sprouts the concentration of the three major cannabinoids
contained in the inflorescences of the corresponding plants (THC, CBD
and CBC) was below detection (0.1 ng / mg for CBD, 0.1 ng / mg for CBN
and 5.0 ng / mg for THC). Also the concentration of cannflavins in seeds
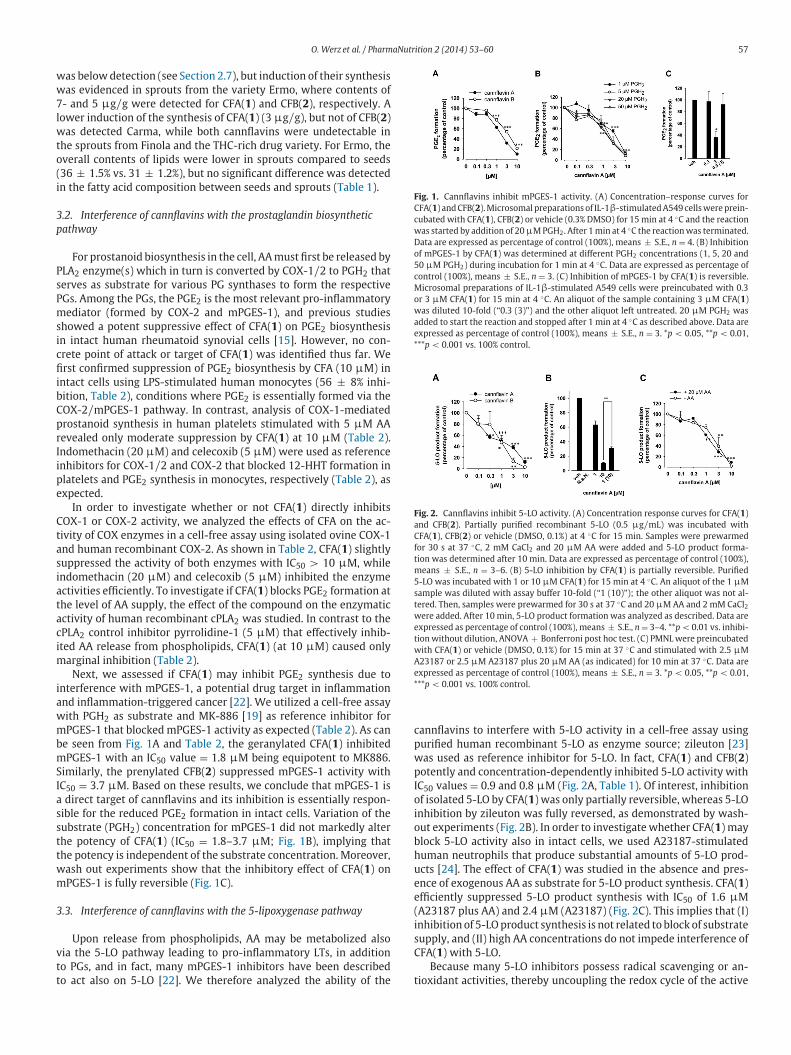
O. Werz et al. / PharmaNutrition 2 (2014) 53–60 57
Fig. 1. Cannflavins inhibit mPGES-1 activity. (A) Concentration–response curves for
CFA( 1 ) and CFB( 2 ). Microsomal preparations of IL-1 β-stimulated A549 cells were prein-
cubated with CFA( 1 ), CFB( 2 ) or vehicle (0.3% DMSO) for 15 min at 4 ◦C and the reaction
was started by addition of 20 μM PGH 2 . After 1 min at 4 ◦C the reaction was terminated.
Data are expressed as percentage of control (100%), means ± S.E., n = 4. (B) Inhibition
of mPGES-1 by CFA( 1 ) was determined at different PGH 2 concentrations (1, 5, 20 and
50 μM PGH 2 ) during incubation for 1 min at 4 ◦C. Data are expressed as percentage of
control (100%), means ± S.E., n = 3. (C) Inhibition of mPGES-1 by CFA( 1 ) is reversible.
Microsomal preparations of IL-1 β-stimulated A549 cells were preincubated with 0.3
or 3 μM CFA( 1 ) for 15 min at 4 ◦C. An aliquot of the sample containing 3 μM CFA( 1 )
was diluted 10-fold (“0.3 (3)”) and the other aliquot left untreated. 20 μM PGH 2 was
added to start the reaction and stopped after 1 min at 4 ◦C as described above. Data are
expressed as percentage of control (100%), means ± S.E., n = 3. * p < 0.05, ** p < 0.01,
*** p < 0.001 vs. 100% control.
Fig. 2. Cannflavins inhibit 5-LO activity. (A) Concentration response curves for CFA( 1 )
and CFB( 2 ). Partially purified recombinant 5-LO (0.5 μg / mL) was incubated with
CFA( 1 ), CFB( 2 ) or vehicle (DMSO, 0.1%) at 4 ◦C for 15 min. Samples were prewarmed
for 30 s at 37 ◦C, 2 mM CaCl 2 and 20 μM AA were added and 5-LO product forma-
tion was determined after 10 min. Data are expressed as percentage of control (100%),
means ± S.E., n = 3–6. (B) 5-LO inhibition by CFA( 1 ) is partially reversible. Purified
5-LO was incubated with 1 or 10 μM CFA( 1 ) for 15 min at 4 ◦C. An aliquot of the 1 μM
sample was diluted with assay buffer 10-fold (“1 (10)”); the other aliquot was not al-
tered. Then, samples were prewarmed for 30 s at 37 ◦C and 20 μM AA and 2 mM CaCl 2 were added. After 10 min, 5-LO product formation was analyzed as described. Data are
expressed as percentage of control (100%), means ± S.E., n = 3–4. ** p < 0.01 vs. inhibi-
tion without dilution, ANOVA + Bonferroni post hoc test. (C) PMNL were preincubated
with CFA( 1 ) or vehicle (DMSO, 0.1%) for 15 min at 37 ◦C and stimulated with 2.5 μM
A23187 or 2.5 μM A23187 plus 20 μM AA (as indicated) for 10 min at 37 ◦C. Data are
expressed as percentage of control (100%), means ± S.E., n = 3. * p < 0.05, ** p < 0.01,
*** p < 0.001 vs. 100% control.
was below detection (see Section 2.7 ), but induction of their synthesis
was evidenced in sprouts from the variety Ermo, where contents of
7- and 5 μg / g were detected for CFA( 1 ) and CFB( 2 ), respectively. A
lower induction of the synthesis of CFA( 1 ) (3 μg / g), but not of CFB( 2 )
was detected Carma, while both cannflavins were undetectable in
the sprouts from Finola and the THC-rich drug variety. For Ermo, the
overall contents of lipids were lower in sprouts compared to seeds
(36 ± 1.5% vs. 31 ± 1.2%), but no significant difference was detected
in the fatty acid composition between seeds and sprouts ( Table 1 ).
3.2. Interference of cannflavins with the prostaglandin biosynthetic
pathway
For prostanoid biosynthesis in the cell, AA must first be released by
PLA 2 enzyme(s) which in turn is converted by COX-1 / 2 to PGH 2 that
serves as substrate for various PG synthases to form the respective
PGs. Among the PGs, the PGE 2 is the most relevant pro-inflammatory
mediator (formed by COX-2 and mPGES-1), and previous studies
showed a potent suppressive effect of CFA( 1 ) on PGE 2 biosynthesis
in intact human rheumatoid synovial cells [ 15 ]. However, no con-
crete point of attack or target of CFA( 1 ) was identified thus far. We
first confirmed suppression of PGE 2 biosynthesis by CFA (10 μM) in
intact cells using LPS-stimulated human monocytes (56 ± 8% inhi-
bition, Table 2 ), conditions where PGE 2 is essentially formed via the
COX-2 / mPGES-1 pathway. In contrast, analysis of COX-1-mediated
prostanoid synthesis in human platelets stimulated with 5 μM AA
revealed only moderate suppression by CFA( 1 ) at 10 μM ( Table 2 ).
Indomethacin (20 μM) and celecoxib (5 μM) were used as reference
inhibitors for COX-1 / 2 and COX-2 that blocked 12-HHT formation in
platelets and PGE 2 synthesis in monocytes, respectively ( Table 2 ), as
expected.
In order to investigate whether or not CFA( 1 ) directly inhibits
COX-1 or COX-2 activity, we analyzed the effects of CFA on the ac-
tivity of COX enzymes in a cell-free assay using isolated ovine COX-1
and human recombinant COX-2. As shown in Table 2 , CFA( 1 ) slightly
suppressed the activity of both enzymes with IC 50 > 10 μM, while
indomethacin (20 μM) and celecoxib (5 μM) inhibited the enzyme
activities efficiently. To investigate if CFA( 1 ) blocks PGE 2 formation at
the level of AA supply, the effect of the compound on the enzymatic
activity of human recombinant cPLA 2 was studied. In contrast to the
cPLA 2 control inhibitor pyrrolidine-1 (5 μM) that effectively inhib-
ited AA release from phospholipids, CFA( 1 ) (at 10 μM) caused only
marginal inhibition ( Table 2 ).
Next, we assessed if CFA( 1 ) may inhibit PGE 2 synthesis due to
interference with mPGES-1, a potential drug target in inflammation
and inflammation-triggered cancer [ 22 ]. We utilized a cell-free assay
with PGH 2 as substrate and MK-886 [ 19 ] as reference inhibitor for
mPGES-1 that blocked mPGES-1 activity as expected ( Table 2 ). As can
be seen from Fig. 1 A and Table 2 , the geranylated CFA( 1 ) inhibited
mPGES-1 with an IC 50 value = 1.8 μM being equipotent to MK886.
Similarly, the prenylated CFB( 2 ) suppressed mPGES-1 activity with
IC 50 = 3.7 μM. Based on these results, we conclude that mPGES-1 is
a direct target of cannflavins and its inhibition is essentially respon-
sible for the reduced PGE 2 formation in intact cells. Variation of the
substrate (PGH 2 ) concentration for mPGES-1 did not markedly alter
the potency of CFA( 1 ) (IC 50 = 1.8–3.7 μM; Fig. 1 B), implying that
the potency is independent of the substrate concentration. Moreover,
wash out experiments show that the inhibitory effect of CFA( 1 ) on
mPGES-1 is fully reversible ( Fig. 1 C).
3.3. Interference of cannflavins with the 5-lipoxygenase pathway
Upon release from phospholipids, AA may be metabolized also
via the 5-LO pathway leading to pro-inflammatory LTs, in addition
to PGs, and in fact, many mPGES-1 inhibitors have been described
to act also on 5-LO [ 22 ]. We therefore analyzed the ability of the
cannflavins to interfere with 5-LO activity in a cell-free assay using
purified human recombinant 5-LO as enzyme source; zileuton [ 23 ]
was used as reference inhibitor for 5-LO. In fact, CFA( 1 ) and CFB( 2 )
potently and concentration-dependently inhibited 5-LO activity with
IC 50 values = 0.9 and 0.8 μM ( Fig. 2 A, Table 1 ). Of interest, inhibition
of isolated 5-LO by CFA( 1 ) was only partially reversible, whereas 5-LO
inhibition by zileuton was fully reversed, as demonstrated by wash-
out experiments ( Fig. 2 B). In order to investigate whether CFA( 1 ) may
block 5-LO activity also in intact cells, we used A23187-stimulated
human neutrophils that produce substantial amounts of 5-LO prod-
ucts [ 24 ]. The effect of CFA( 1 ) was studied in the absence and pres-
ence of exogenous AA as substrate for 5-LO product synthesis. CFA( 1 )
efficiently suppressed 5-LO product synthesis with IC 50 of 1.6 μM
(A23187 plus AA) and 2.4 μM (A23187) ( Fig. 2 C). This implies that (I)
inhibition of 5-LO product synthesis is not related to block of substrate
supply, and (II) high AA concentrations do not impede interference of
CFA( 1 ) with 5-LO.
Because many 5-LO inhibitors possess radical scavenging or an-
tioxidant activities, thereby uncoupling the redox cycle of the active

58 O. Werz et al. / PharmaNutrition 2 (2014) 53–60
Table 1
Fatty acids composition of seeds and sprouts from the Ermo variety of hemp (each data represents the mean of three replicates ± standard deviation).
Fatty acid (relative %) Seeds Sprouts
Linoleic 57.53 ± 0.2 58.00 ± 0.31
α-Linolenic 24.53 ± 0.14 24.55 ± 0.17
Oleic 9.97 ± 0.06 10.01 ± 0.07
γ- Linolenic 1.08 ± 0.01 1.42 ± 0.03
Palmitic 4.96 ± 0.13 4.18 ± 0.207
Stearic 1.94 ± 0.01 1.85 ± 0.04
Table 2
Effects of CFA( 1 ) on various eicosanoid-forming enzymes and cellular functions of neutrophils and monocytes. CFA( 1 ) or reference inhibitors (at the indicated concentrations) were
added to the respective enzymes or freshly isolated human blood cells 15 min prior induction of the enzyme reaction. Data (means ± S.E., n = 3–5) are expressed as IC 50 values
and as percentage of the inhibition of the enzyme activity vs. the uninhibited vehicle (0.1% DMSO) control.
Enzyme / assay
CFA(1)
IC 50 [ μm];
% inhibition at 10 μM
Reference control,
% inhibition at indic. conc.
COX-1, platelets > 10 μM; 26 ± 2% Indomethacin (20 μM), 95 ± 5%
COX-1, cell-free > 10 μM; 36 ± 2% Indomethacin (20 μM), 81 ± 8%
COX-2, monocytes 8.8 μM; 56 ± 8% Celecoxib (5 μM), 80 ± 5%
COX-2, cell-free > 10 μM; 35 ± 12% Celecoxib (5 μM), 78 ± 8%
cPLA 2 , cell-free > 10 μM; 26 ± 8% Pyrrolidine-1 (5 μM), 79 ± 3%
mPGES-1, cell-free 1.8 μM; 90 ± 1% MK-886 (10 μM), 83 ± 1%
5-LO cell-free 0.9 μM; 88 ± 3% Zileuton (3 μM), 80 ± 4%
5-lo, PMNL, A23187 2.4 μM; 98 ± 2% BWA4C (0.3 μM), 99 ± 2%
5-lo, PMNL, A23187 + AA 1.6 μM; 91 ± 1% BWA4C (0.3 μM), 76 ± 2%
DPPH assay No effect Ascorbate (50 μM), 80 ± 6%
Cell viability no effect staurosporine (3 μM), 80 ± 5%
Fig. 3. Radical scavenging properties of CFA( 1 ). CFA( 1 ) was incubated with 5 nmol
DPPH for 30 min at RT and the absorbance was measured at 520 nm. Ascorbic acid
and l -cysteine were used as controls. Values are given as percentage of control (100%)
mean ± S.E., n = 3–4. *** p < 0.001 vs. 100% control.
s
b
a
w
f
m
b
v
ite iron in 5-LO, we analyzed CFA( 1 ) for its ability to reduce the sta-
le DPPH radical. As shown in Fig. 3 , the antioxidants ascorbic acid
nd l -cysteine (used as positive controls) reduced the DPPH radical
hereas CFA( 1 ) failed in this respect, excluding radical scavenging
eatures of CFA( 1 ).
Finally, we tested if CFA( 1 ) may affect cell viability using human
onocytes that were treated with 10 μM CFA( 1 ) for 24 h. Analysis
y MTT assay revealed no cytotoxicity or detrimental effects on cell
iability ( Table 2 ) excluding unspecific influences in the cell-based
assays. Similarly, neutrophils treated with 10 μM CFA( 1 ) for 1 h still
excluded the dye trypan blue, analyzed by light microscopy, implying
cellular integrity (not shown).
4. Discussion
Before the identification of specific cannabinoid receptors, the in-
hibition of the synthesis of inflammatory PGs, in particular PGE 2 , was
considered a possible mechanism underlying the analgesic properties
of THC [ 25 ]. However, conflicting data were obtained when THC and
other cannabinoids were evaluated in simple assays of COX inhibition,
and the situation was further confused by the observation that ex-
tracts of Cannabis devoid of cannabinoids showed a strong inhibitory
activity on the production of PGs in cultured rheumatoid synovial cells
[ 15 ]. A flavonoid named cannflavin was eventually identified as the
major inhibitor of cellular PG synthesis from cannabinoids-free ex-
tracts of Cannabis [ 15 ]. Cannflavin is actually a mixture of two closely
related prenylated flavonoids, CFA( 1 ) and CFB( 2 ), differing in the na-
ture of the prenyl group on ring A of the flavone ring system (geranyl
in CFA( 1 ) and prenyl in CFB( 2 )) [ 26 ]. Cannflavin C, a third natural
cannflavin previously obtained as a by-product from the synthesis of
CFB( 2 ) [ 14 ], was recently reported from a high-potency THC chemo-
type, and represents a remarkable example of “anticipated” natural
product discovery [ 27 ]. The anti-inflammatory potency of cannflavin
was found to be intermediate between that of aspirin and dexam-
ethasone [ 15 ], but its mechanism of activity has remained elusive.
Cannflavins are minor constituents of C. sativa . During our phyto-
chemical studies on this plant [ 28 –30 ], we found that the ratio be-
tween the two cannflavins and their concentration are rather variable
within the various varieties of the plant. These compounds are diffi-
cult to obtain completely pure by isolation, and tend to co-crystallize
with other phenolics, making their obtaining process tedious. CFB( 2 )
can be conveniently obtained by synthesis [ 14 ], but synthetic CFA( 1 )
is a mixture of stereoisomers. We were therefore delighted to discover
that Ermo, a variety of hemp devoid of cannabinoids, contains signif-
icantly higher amounts of CFA( 1 ) compared to the cannabinoids-rich

O. Werz et al. / PharmaNutrition 2 (2014) 53–60 59
varieties, with minimal contamination from CFB. Owing to the nutri-
tional potential of cannabinoid-free strains of hemp, we wondered if
CFA( 1 ) was also contained in the seeds of the plant. Hemp seeds are
normally devoid of cannflavins, and the same observation was done
on the seeds of Ermo. However, sprouting induced the formation of
both CFA(1) (7 μg / g) CFB( 2 ) (5 μg / g) in this variety, and a lower
induction of the synthesis of CFA( 1 ) (3 μg / g), but not of CFB( 2 ), was
also observed in the variety Carma, despite the presence of both can-
nflavins in the flowerheads of this variety. Neither CFA( 1 ) nor CFB( 2 )
could be detected in the sprouts from the variety Finola and one THC-
rich medicinal variety, that both contain these compounds in their
flowerheads (data not showed). Since the ratio between ω -3 and ω -6
fatty acids was unaffected by germination, the nutritional profile of
sprouts could benefit, compared to the seeds, from their contents of
cannflavins. The estrogenic hop flavonoid 8-prenylnaringenin (8 PN,
4 ) is structurally related to cannflavins, and showed clinical activity
at dosages of 100 μg / day [ 31 ], an amount roughly corresponding
to the CFA( 1 ) contents of 20 g of fresh hemp sprouts. The affinity
of 8 PN for estrogen receptors is roughly two orders of magnitude
higher than that of CFA( 1 ) for mPGES-1 and 5-LO [ 32 ], but, owing to
the long elimination half life of prenylated flavonoids [ 33 ], it does
not seem unrealistic to assume that regular consumption of hemp
sprouts could lead to the attainment of bioactive concentrations of
cannflavins in plasma and tissues.
Flavonoids can modulate the expression of mPGES-1 [ 34 ], but
cannflavins are the first flavonoids having direct inhibitory activity
on this enzyme. Since good inhibitory activity was also reported for
the prenylated phloroglucynol arzanol [ 35 ], a phenolic somewhat re-
sembling the A ring of cannflavins, it is tempting to speculate that
prenylation is important for mPGES-1 inhibition. Because of their
anti-inflammatory activity, the presence of cannflavins might con-
tribute to the nutritional profile of the sprouts. In this context, it was
also important to assure the absence of cannabinoids in the sprouts,
and to evaluate if changes in the lipid profile had occurred during ger-
mination. The concentration of cannabinoids in hemp seeds is very
low, and special extraction protocols have been developed for their
efficient recovery [ 16 ]. By using a seed-specific method of extraction,
we found that traces ( < 1 μg / g) these compounds were only present
in the seed tegument of the cannabinoids-producing varieties. On the
other hand, these compounds were totally absent in the correspond-
ing dehulled material, confirming previous findings, and supporting
the view that the presence of cannabinoids in seeds is the result of
matrix contamination during maturation and not the result of a site-
specific synthesis or accumulation. Furthermore, de novo production
of cannabinoids was not observed during sprouting, confirming their
strict association to specific glandular structures typical of adult plant
tissues [ 11 ]. As expected, cannabinoids were also undetectable in the
sprouts of Ermo.
Plants have the capacity to de novo synthesize sugars from fatty
acids, and, indeed, a decrease of the overall fat contents was observed
between seeds and sprouts of Ermo (from 36 ± 1.5% to 31 ± 1.2%).
On the other hand, limited information exists if certain types of fatty
acids are selectively metabolized during germination. It was there-
fore important to investigate if depletion of ω -3 was occurring during
germination, since this would have eroded the nutritional profile of
sprouts. However, no significant difference was found in the fatty
acids unsaturation profile of seeds and sprouts from Ermo, with the
maintenance of the ca 3:1 ratio between ω -6 and ω -3 essential fatty
acids ( Table 1 ). Significant changes were, however, observed for γ-
linolenic (GLA)- and palmitic acids, both minor constituents, with a ca
30% increase of concentration of GLA (from 1.08% to 1.42%) and a cor-
responding decrease of palmitic acid (from 4.96% to 4.18%) ( Table 1 )
during germination.
The availability of both cannflavins made it possible to investigate
the molecular basis underlying their anti-inflammatory properties,
and we have unequivocally shown that these compounds act as dual
inhibitors of mPGES-1 and 5-LO, two crucial enzymes in the biosyn-
thesis of the pro-inflammatory mediators PGE 2 and LTs, respectively.
Moreover, the direct interference of cannflavins with mPGES-1 and
the weak effects on COX-1 / 2 enzymes suggest mPGES-1 as the tar-
get responsible for repression of cellular PGE 2 synthesis observed
in complex cell-based assay [ 15 ]. Our data show that (a) CFA( 1 ) ex-
erts its inhibitory effects on mPGES-1 and 5-LO in a reversible and
substrate concentration-independent manner, (b) exhibits no radical
scavenger activity (DPPH assay), and (c) fails to substantially inhibit
the related COX-1 and COX-2 enzymes as well as the cPLA 2 , the en-
zyme that provides AA as substrate for PGE 2 and LT formation. In
fact, dual inhibition of mPGES-1 and 5-LO is considered as pharma-
cological strategy in order to intervene with inflammatory diseases
and might be superior over single target interference in terms of ef-
ficacy as well as with respect to lower side effects [ 34 ]. Together, the
potent suppression of mPGES-1 and 5-LO by cannflavins is of con-
siderable anti-inflammatory potential, especially when the intake of
these compounds is associated to a fatty matrix that can both in-
crease their absorption [ 36 ], and potentiate their activity due to its
high contents of ω 3-acids.
5. Conclusions
Sprouting has a general beneficial effect on the digestibility of
seeds by reducing the level of anti-nutritional factors, but changes
in secondary metabolites can also occur, with the potential to en-
rich sprouts with specific phytochemicals. In this context, we have
identified a variety of hemp seeds where production of the anti-
inflammatory lipophilic flavonoids cannflavins is induced by ger-
mination, and have clarified the mechanism by which these com-
pounds inhibit the production of pro-inflammatory prostanoids and
leukotrienes. These data provide a rationale for seriously considering
the commercial development of this novel hemp seed food, optimiz-
ing factors like sprout length, root length, and post-sprouting time.
Conflicts of interest
Ermo is a registered variety of hemp at CPVO by CRA, the employer
of GG.
Layperson ’ s summary
Hemp seeds contain all essential amino acids and lipids
necessary for human nutrition. This, coupled to their anti-
inflammatory profile of unsaturated fatty acids and pleasant
nutty taste, has gained them a “super-food” status in consumers.
Sprouting has been shown to increase the nutritional value of
seeds by reducing the concentration of anti-nutritional factors
and boosting the production of specific phytochemicals, as in
broccoli. We have discovered that sprouting, while not induc-
ing the production of cannabinoids, can trigger the formation
of anti-inflammatory lipophilic flavonoids in some varieties of
hemp. The mechanism of action of these compounds has been
elucidated at the molecular level, providing a rationale for the
development of sprouts as a novel hemp-derived healthfood.

60 O. Werz et al. / PharmaNutrition 2 (2014) 53–60
R
eferences[1] Callaway JC. Hempseed as a nutritional resource: an overview. Euphytica
2004;140:65–72. http://dx.doi.org/10.1007/s10681- 004- 4811- 6 . [2] Russo EB. History of cannabis and its preparations in saga, science, and sobriquet.
Chemistry & Biodiversity 2007;4(8):1614–48. http://dx.doi.org/10.1002/cbdv. 200790144 , 17712811 .
[3] Matthaeus B, Bruehl L. Virgin hemp seed oil: an interesting niche product. European Journal of Lipid Science and Technology 2008;110:655–61. http:
//dx.doi.org/10.1002/ejlt.200700311 .
[4] Johnson MM, Swan DD, Surette ME, Stegner J, Chilton T, Fonteh AN et al. Di- etary supplementation with gamma-linolenic acid alters fatty acid content and
eicosanoid production in healthy humans. Journal of Nutrition 1997;127:1435–44, 9237935 .
[5] Callaway J, Schwab U, Harvima I, Halonen P, Mykkaenen O, Hyvoenen P et al. Efficacy of dietary hempseed oil in patients with atopic dermatitis. Jour-
nal of Dermatological Treatment 2005;16:87–94. http://dx.doi.org/10.1080/
09546630510035832 , 16019622 . [6] Richard MN, Ganguly R, Steigerwald SN, Al-Khalifa A, Pierce GN. Dietary
hempseed reduces platelet aggregation. Journal of Thrombosis and Haemosta- sis: JTH 2007;5:424–5. http://dx.doi.org/10.1111/j.1538-7836.2007.02327.x ,
17155962 . [7] a. Gu Y, Guo Q, Zhang L, Chen Z, Han Y, Gu Z. Physiological and biochemical
metabolism of germinating broccoli seeds and sprouts. Journal of Agricultural
and Food Chemistry 2012;60:209–13. http://dx.doi.org/10.1021/jf203599v , 22142148 ; b. Seo WD, Yuk HJ, Curtis-Long MJ, Jang KC, Lee JH,
Han S-I et al. Effect of the growth stage and cultivar on policosanol profiles of barley sprouts and their adenosine 5 ′ -monophosphate-activated protein ki-
nase activation. Journal of Agricultural and Food Chemistry 2013;61:1117–23. http://dx.doi.org/10.1021/jf3041879 , 23301834 .
[8] Mirmiran P, Bahadoran Z, Hosseinpanah F, Keyzad A, Azizi F. Effects of broccoli
sprout with high sulforaphane concentration on inflammatory markers in type 2 diabetic patients: a randomized double-blind placebo-controlled clinical trial.
Journal of Functional Foods 2012;4:837–41. http://dx.doi.org/10.1016/j.jff.2012. 05.012 .
[9] a. Appendino G, Jakupovic J, Belloro E, Marchesini A. Multiflorane triterpenoid esters from pumpkin. An unexpected extrafolic source of PABA. Phytochemistry
1999;51:1021–6. http://dx.doi.org/10.1016/S0031- 9422(98)00748- 1 ; b. Ap- pendino G, Jakupovic J, Belloro E, Marchesini A. Triterpenoid p-aminobenzoates
from the seeds of zucchini. Fitoterapia 2000;71:258–63. http://dx.doi.org/10.
1016/S0367- 326X(99)00168- 9 , 10844164 . [10] Gatbonton-Schwanger JN, Letterio JJ, Tochtrop GP. Bryonolic acid transcriptional
control of anti-inflammatory and antioxidant genes in macrophages in vitro and in vivo. Journal of Natural Products 2012;75:591–8. http://dx.doi.org/10.1021/
np200823p , 22339499 . [11] Kim E-S, Mahlberg PG. Immunochemical localization of tetrahydrocannabi-
nol (THC) in cryofixed glandular trichomes of cannabis (Cannabaceae). Amer-
ican Journal of Botany 1997;84:336–42. http://dx.doi.org/10.2307/2446007 , 21708587 .
[12] Appendino G, Chianese G, Taglialatela-Scafati O. Cannabinoids: occurrence and medicinal chemistry. Current Medicinal Chemistry 2011;18:1085–99. http://dx.
doi.org/10.2174/092986711794940888 , 21254969 . [13] S anchez-Duffhues G, Calzado MA, de Vinuesa AG, Caballero FJ, Ech-Chahad A,
Appendino G et al. Denbinobin, a naturally occurring 1,4-phenanthrenequinone,
inhibits HIV-1 replication through an NF-kappaB-dependent pathway. Bio- chemical Pharmacology 2008;76:1240–50. http://dx.doi.org/10.1016/j.bcp.
2008.09.006 , 18840408 . [14] Minassi A, Giana A, Ech-Chahad A, Appendino G. A regiodivergent synthesis of
ring A C-prenylflavones. Organic Letters 2008;10:2267–70. http://dx.doi.org/10. 1021/ol800665w , 18454537 .
[15] Barrett ML, Gordon D, Evans FJ. Isolation from cannabis sativa L. Of cann-
flavin – a novel inhibitor of prostaglandin production. Biochemical Pharma- cology 1985;34:2019–24. http://dx.doi.org/10.1016/0006- 2952(85)90325- 9 ,
3859295 . [16] Lachenmeier DW, Kroener L, Musshoff F, Burkhard M. Determination of cannabi-
noids in hemp food products by use of headspace solid-phase microextrac- tion and gas chromatography–mass spectrometry. Analytical and Bioanalyti-
cal Chemistry 2004;378:183–9. http://dx.doi.org/10.1007/s00216- 003- 2268- 4 ,
14598006 .
[17] Morrison WR, Smith LM. Preparation of fatty acid methyl esters and dimethy-
lacetals from lipids with boron fluoride–methanol. Journal of Lipid Research 1964;53:600–8, 14221106 .
[18] Fischer L, Szellad D, Radmark O, Steinhilber D, Werz O. Phosphorylation- and stimulus-dependent inhibition of cellular 5-lipoxygenase activity by nonredox-
type inhibitors. FASEB Journal: Official Publication of the Federation of American
Societies for Experimental Biology 2003;17:949–51, 12670876 . [19] Koeberle A, Siemoneit U, Buehring U, Northoff H, Laufer S, Albrecht W et al.
Licofelone suppresses prostaglandin E2 formation by interference with the inducible microsomal prostaglandin E2 synthase-1. Journal of Pharmacology
and Experimental Therapeutics 2008;326:975–82. http://dx.doi.org/10.1124/ jpet.108.139444 , 18550688 .
[20] Hoffmann N, Lopez JJ, Pergola C, Feisst C, Pawelczik S, Jakobsson PJ et al. Hyper-
forin induces Ca2 + -independent arachidonic acid release in human platelets by facilitating cytosolic phospholipase A2 activation through select phospholipid
interactions. Biochimica et Biophysica Acta 2010;1801:462–72 . [21] Blois MS. Antioxidant determinations by the use of a stable free radical. Nature
1958;181:1199–200. http://dx.doi.org/10.1038/1811199a0 .
[22] R admark O, Samuelsson B. Microsomal prostaglandin E synthase-1 and 5- lipoxygenase: potential drug targets in cancer. Journal of Internal Medicine
2010;268:5–14, 20497297 . [23] Carter GW, Young PR, Albert DH, Bouska J, Dyer R, Bell RL et al. 5-Lipoxygenase
inhibitory activity of zileuton. Journal of Pharmacology and Experimental Ther- apeutics 1991;256:929–73, 1848634 .
[24] Pergola C, Dodt G, Rossi A, Neunhoeffer E, Lawrenz B, Northoff H et al. ERK-
mediated regulation of leukotriene biosynthesis by androgens: a molecular basis for gender differences in inflammation and asthma. Proceedings of the National
Academy of Sciences of the United States of America 2008;105:19881–6. http: //dx.doi.org/10.1073/pnas.0809120105 , 19064924 .
[25] Mechoulam R. Looking back at cannabis research. Current Pharmaceutical De- sign 2000;6:1313–22. http://dx.doi.org/10.2174/1381612003399509 .
[26] Barrett ML, Scutt AM, Evans FJ. Cannflavin A and B, prenylated flavones
from Cannabis sativa L. Experientia 1986;42:452–3. http://dx.doi.org/10.1007/ BF02118655 , 3754224 .
[27] Radwan MM, Elsohly MA, Slade D, Ahmed SA, Wilson L, El-Alfy AT et al. Non- cannabinoid constituents from a high potency Cannabis sativa variety. Phyto-
chemistry 2008;69:2627–33. http://dx.doi.org/10.1016/j.phytochem.2008.07. 010 , 18774146 .
[28] Taglialatela-Scafati O, Pagani A, Scala F, De Petrocellis L, Di Marzo V, Grassi G
et al. Cannabimovone, a cannabinoid with a rearranged terpenoid skeleton from
hemp. European Journal of Organic Chemistry 2010;11:2067–72 .
[29] Pagani A, Scala F, Chianese G, Grassi G, Appendino G, Taglialatela-Scafati O. Cannabioxepane, a novel tetracyclic cannabinoid from hemp, Cannabis sativa L.
Tetrahedron 2011;67:3369–73. http://dx.doi.org/10.1016/j.tet.2011.03.062 . [30] Pollastro F, Taglialatela-Scafati O, Allar a M, Munoz E, Di Marzo V, De Petro-
cellis L et al. Bioactive prenylogous cannabinoid from fiber hemp ( Cannabis sativa ). Journal of Natural Products 2011;74:2019–22. http://dx.doi.org/10.
1021/np200500p , 21902175 .
[31] Erkkola R, Vervarcke S, Vansteelandt S, Rompotti P, DeKeukeleire D, Heyer- ick A. A randomized, double-blind, placebo-controlled, cross-over pilot study
on the use of a standardized hop extract to alleviate menopausal discomforts. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology
2010;17:389–96. http://dx.doi.org/10.1016/j.phymed.2010.01.007 , 20167461 . [32] Ikeda K, Arao Y, Otsuka H, Nomoto S, Horiguchi H, Kato S et al. Terpenoids found
in the umbelliferae family act as agonists / antagonists for ER(alpha) and ERbeta:
differential transcription activity between ferutinine-liganded ER(alpha) and ERbeta. Biochemical and Biophysical Research Communications 2002;291:354–
60. http://dx.doi.org/10.1006/bbrc.2002.6446 , 11846412 . [33] Nikoli c D, van Breemen RB. Analytical methods for quantitation of prenylated
flavonoids from hops. Current Analytical Chemistry 2013;9:71–85 . [34] Koeberle A, Werz O. Inhibitors of the microsomal prostaglandin E2 synthase-1
as alternative to non steroidal anti-inflammatory drugs (NSAIDs) – a critical
review. Current Medicinal Chemistry 2009;16:4274–96. http://dx.doi.org/10. 2174/092986709789578178 .
[35] Minassi A, Cicione L, Koeberle A, Bauer J, Laufer S, Werz O et al. A multicom- ponent carba-Betti strategy to alkylidene heterodimers. Total synthesis and
structure–activity relationships of arzanol. European Journal of Organic Chem- istry 2012:772–9 .
[36] Lotito SB, Frei B. Consumption of flavonoid-rich foods and increased plasma
antioxidant capacity in humans: cause, consequence, or epiphenomenon? Free Radical Biology and Medicine 2006;4:1727–46 .




















