
Molecular cloning and characterization of wheat calreticulin ...
13
Journal of Experimental Botany, Vol. 59, No. 4, pp. 739–751, 2008 doi:10.1093/jxb/erm369 This paper is available online free of all access charges (see http://jxb.oxfordjournals.org/open_access.html for further details) RESEARCH PAPER Molecular cloning and characterization of wheat calreticulin (CRT) gene involved in drought-stressed responses Xiao-Yun Jia 1,2 , Chong-Yi Xu 1 , Rui-Lian Jing 1, *, Run-Zhi Li 2 , Xin-Guo Mao 1 , Ji-Ping Wang 1,2 and Xiao-Ping Chang 1 1 The National Key Facility for Crop Gene Resources and Genetic Improvement, Key Laboratory of Crop Germplasm and Biotechnology, Ministry of Agriculture, Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China 2 Center for Agricultural Biotechnology, Shanxi Agricultural University, Taigu 030801, China Received 12 October 2007; Revised 5 December 2007; Accepted 13 December 2007 Abstract Calreticulin (CRT) is a highly conserved and ubiqui- tously expressed Ca 2+ -binding protein in multicellular eukaryotes. CRT plays a crucial role in many cellular processes including Ca 2+ storage and release, protein synthesis, and molecular chaperone activity. To eluci- date the function of CRTs in plant responses against drought, a main abiotic stress limiting cereal crop production worldwide, a full-length cDNA encoding calreticulin protein namely TaCRT was isolated from wheat (Triticum aestivum L.). The deduced amino acid sequence of TaCRT shares high homology with other plant CRTs. Phylogenetic analysis indicates that TaCRT cDNA clone encodes a wheat CRT3 isoform. Southern analysis suggests that the wheat genome contains three copies of TaCRT. Subcellular locations of TaCRT were the cytoplasm and nucleus, evidenced by transient expression of GFP fused with TaCRT in onion epidermal cells. Enhanced accumulation of TaCRT transcript was observed in wheat seedlings in response to PEG-induced drought stress. To investi- gate further whether TaCRT is involved in the drought- stress response, transgenic plants were constructed. Compared to the wild-type and GFP-expressing plants, TaCRT-overexpressing tobacco (Nicotiana bentham- iana) plants grew better and exhibited less wilt under the drought stress. Moreover, TaCRT-overexpressing plants exhibited enhanced drought resistance to water deficit, as shown by their capacity to maintain higher WUE (water use efficiency), WRA (water retention ability), RWC (relative water content), and lower MDR (membrane damaging ratio) (P <0 01) under water- stress conditions. In conclusion, a cDNA clone encod- ing wheat CRT was successfully isolated and the results suggest that TaCRT is involved in the plant response to drought stress, indicating a potential in the transgenic improvements of plant water-stress. Key words: Calreticulin, drought resistance, gene expression, physiological responses, TaCRT, wheat (Triticum aestivum L.). Introduction Calreticulin (CRT) is an abundant Ca 2+ -binding protein that ubiquitously expresses in all of the multicellular eukaryotes investigated (Coppolino and Dedhar, 1998; Michalak et al., 1999). CRT was first detected in the endoplasmic reticulum (ER) of rabbit skeletal muscle (Ostwald and MacLennan, 1974) and later was cloned from rabbit (Fliegel et al., 1989) and mouse (Smith and Koch, 1989). Besides its main location in ER (Opas et al., 1996), CRT has been found to reside in the nuclear envelope (Napier et al., 1995), the spindle apparatus of the dividing cells (Denecke et al., 1995), the cell surface (Gardai et al., 2005), and the plasmodesmata (Laporte et al., 2003; Chen D et al., 2005), indicating that CRT is essential for normal cell function. * To whom correspondence should be addressed. E-mail: [email protected] Abbreviations: CRT, calreticulin; ER, endoplasmic reticulum; PEG, polyethylene glycol; GFP, green fluorescent protein; MDR, membrane damaging ratio; PEBV, pea early browning virus; qRT-PCR, quantitative real-time PCR; RWC, relative water content; TaCRT, calreticulin of Triticum aestivum; WRA, water retention ability; WUE, water use efficiency; WT, wild type. ª 2008 The Author(s). This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/2.0/uk/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. Downloaded from https://academic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Molecular cloning and characterization of wheat calreticulin ...

Journal of Experimental Botany, Vol. 59, No. 4, pp. 739–751, 2008
doi:10.1093/jxb/erm369This paper is available online free of all access charges (see http://jxb.oxfordjournals.org/open_access.html for further details)
RESEARCH PAPER
Molecular cloning and characterization of wheat calreticulin(CRT) gene involved in drought-stressed responses
Xiao-Yun Jia1,2, Chong-Yi Xu1, Rui-Lian Jing1,*, Run-Zhi Li2, Xin-Guo Mao1, Ji-Ping Wang1,2
and Xiao-Ping Chang1
1 The National Key Facility for Crop Gene Resources and Genetic Improvement, Key Laboratory of CropGermplasm and Biotechnology, Ministry of Agriculture, Institute of Crop Sciences, Chinese Academy ofAgricultural Sciences, Beijing 100081, China2 Center for Agricultural Biotechnology, Shanxi Agricultural University, Taigu 030801, China
Received 12 October 2007; Revised 5 December 2007; Accepted 13 December 2007
Abstract
Calreticulin (CRT) is a highly conserved and ubiqui-
tously expressed Ca2+-binding protein in multicellular
eukaryotes. CRT plays a crucial role in many cellular
processes including Ca2+ storage and release, protein
synthesis, and molecular chaperone activity. To eluci-
date the function of CRTs in plant responses against
drought, a main abiotic stress limiting cereal crop
production worldwide, a full-length cDNA encoding
calreticulin protein namely TaCRT was isolated from
wheat (Triticum aestivum L.). The deduced amino acid
sequence of TaCRT shares high homology with other
plant CRTs. Phylogenetic analysis indicates that
TaCRT cDNA clone encodes a wheat CRT3 isoform.
Southern analysis suggests that the wheat genome
contains three copies of TaCRT. Subcellular locations
of TaCRT were the cytoplasm and nucleus, evidenced
by transient expression of GFP fused with TaCRT in
onion epidermal cells. Enhanced accumulation of
TaCRT transcript was observed in wheat seedlings in
response to PEG-induced drought stress. To investi-
gate further whether TaCRT is involved in the drought-
stress response, transgenic plants were constructed.
Compared to the wild-type and GFP-expressing plants,
TaCRT-overexpressing tobacco (Nicotiana bentham-
iana) plants grew better and exhibited less wilt under
the drought stress. Moreover, TaCRT-overexpressing
plants exhibited enhanced drought resistance to water
deficit, as shown by their capacity to maintain higher
WUE (water use efficiency), WRA (water retention
ability), RWC (relative water content), and lower MDR
(membrane damaging ratio) (P <0�01) under water-
stress conditions. In conclusion, a cDNA clone encod-
ing wheat CRT was successfully isolated and the
results suggest that TaCRT is involved in the plant
response to drought stress, indicating a potential in
the transgenic improvements of plant water-stress.
Key words: Calreticulin, drought resistance, gene expression,
physiological responses, TaCRT, wheat (Triticum aestivum L.).
Introduction
Calreticulin (CRT) is an abundant Ca2+-binding proteinthat ubiquitously expresses in all of the multicellulareukaryotes investigated (Coppolino and Dedhar, 1998;Michalak et al., 1999). CRT was first detected in theendoplasmic reticulum (ER) of rabbit skeletal muscle(Ostwald and MacLennan, 1974) and later was clonedfrom rabbit (Fliegel et al., 1989) and mouse (Smith andKoch, 1989). Besides its main location in ER (Opas et al.,1996), CRT has been found to reside in the nuclearenvelope (Napier et al., 1995), the spindle apparatus ofthe dividing cells (Denecke et al., 1995), the cell surface(Gardai et al., 2005), and the plasmodesmata (Laporteet al., 2003; Chen D et al., 2005), indicating that CRT isessential for normal cell function.
* To whom correspondence should be addressed. E-mail: [email protected]: CRT, calreticulin; ER, endoplasmic reticulum; PEG, polyethylene glycol; GFP, green fluorescent protein; MDR, membrane damaging ratio;PEBV, pea early browning virus; qRT-PCR, quantitative real-time PCR; RWC, relative water content; TaCRT, calreticulin of Triticum aestivum; WRA, waterretention ability; WUE, water use efficiency; WT, wild type.
ª 2008 The Author(s).This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/2.0/uk/) whichpermits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

Extensive studies of mammalian CRTs have elucidateda number of key physiological functions, including theregulation of Ca2+ homeostasis and Ca2+-dependent signalpathways (Michalak et al., 2002; Gelebart et al., 2005),molecular chaperone activity in the folding of manyproteins (Denecke et al., 1995; Williams, 2006), modula-tion of nuclear-hormone receptor-mediated gene expres-sion (Burns et al., 1994), control of cell adhesion (Opaset al., 1996), and integrin-dependent Ca2+ signalling at theextra-ER sites in mammalian cells (Coppolino et al.,1997; Krause and Michalak, 1997). In addition, CRTappears to play a role in the immune system (Guo et al.,2002) and apoptosis. For example, CRT-dependent shap-ing of Ca2+ signalling was found to be a criticalcontributor to the modulation of the T cell adaptiveimmune response (Porcellini et al., 2006). Surface CRTmediates muramyl dipeptide-induced RK13 cell apoptosisthrough activating the apoptotic pathway (Chen D et al.,2005).CRT is highly conserved in eukaryotic cells, which is
indicated by sequence analysis on the deduced aminoacids of the known CRT cDNA clones from severalmammalian species (Fliegel et al., 1989; Smith and Koch,1989) and other organisms including nematode (Smith,1992a), fruit fly (Smith, 1992b), marine snail (Kennedyet al., 1992), clawed frog (Treves et al., 1992), rainbowtrout (Stephen et al., 2004), and Cotesia rubecula (Zhanget al., 2006). The protein contains three distinct structuraland functional domains with loosely defined boundaries:the nearly neutral N-domain, the proline-rich P-domain,and the polyacidic C-domain. CRT also has an N-terminalsignal peptide sequence and an ER retention motif inthe C-domain. The P-domain is responsible for the high-affinity and low-capacity Ca2+ binding while theC-domain is responsible for the low-affinity and high-capacity Ca2+ binding. Within the P-domain, there are twotypes of triplicate repeated motifs that are highly con-served among various animal species. However, theC-domain is less conserved than other domains of CRT(Michalak et al., 1992, 1999). Four amino acid residues(Glu239, Asp241, GLu243, and Trp244) at the tip of the‘extended arm’ of the P-domain are critical in thechaperone function of CRT (Martin et al., 2006). Atpresent, no structural information is available for theC-domain which is involved in Ca2+ storage in the lumenof the ER (Nakamura et al., 2001).More recently, the multifunctional roles of CRT in plant
cellular events are rapidly emerging areas of study in plantbiology. CRT has been identified in a few plant species,although functional analysis lags that undertaken in theanimal system. The first indication that plants containcalreticulin-like proteins came from the purified Ca2+-binding proteins of spinach leaves (Menegazzi et al.,1993). Those proteins showed high homology to CRTsequences of mammals. Subsequently, CRT cDNA clones
were isolated from Arabidopsis (Huang et al., 1993),barley (Chen et al., 1994), pea (Hassan et al., 1995),tobacco (Denecke et al., 1995), maize (Napier et al.,1995; Dresselhaus et al., 1996), Brassica rapa (Lim et al.,1996), Ricinus communis L. (Coughlan et al., 1997), andrice (Li and Komatsu, 2000). As a Ca2+ sensor andmolecular chaperone within the ER, plant CRT shares thesame structural domain features and basic functionsidentified for animal CRTs (Coughlan et al., 1997; Li andKomatsu, 2000; Wyatt et al., 2002). Plant CRT is highlyexpressed during mitosis in tobacco (Denecke et al.,1995), embryogenesis of barley (Chen et al., 1994),Nicotiana plumbaginifolia (Borisjuk et al., 1998), andmaize (Dresselhaus et al., 1996), and in flower tissues(Nelson et al., 1997) including sperm cells (Williamset al., 1997), pollen tubes as well as anthers (Nardi et al.,2006). Calreticulin in plant cells has been shown to playa role in regeneration (Li and Komatsu, 2000; Jin et al.,2005), pollen–pistil interaction (Lenartowska et al., 2002),and cell-to-cell transport via the plasmodesmata (Baluskaet al., 1999; Laporte et al., 2003; Chen MH et al., 2005).Increasing evidence also indicates that this protein isinvolved in the plant response to a variety of stress-mediated stimuli, for example, pathogen-related signallingmolecules (Jaubert et al., 2002), gravistimulation (Heilmannet al., 2001), and other stress factors (Sharma et al.,2004). However, the precise mechanism of CRT functionin these plant cell processes, particularly in regulatingstress response, remains to be fully ascertained.Wheat, with a huge and very complex genome, is an
important cereal crop in the world. Drought is a majorconstraint to wheat production worldwide. Understandingthe molecular basis of drought-stress response is highlyrequired for wheat genetic improvement of droughttolerance. Little information is available for the wheatCRT gene and its functions, although a wheat calreticulin-like sequence was reported (GI 56606826). The presentstudy aims to investigate and clarify the function of CRTin the drought-stress response of wheat. The isolation andcharacterization of a full-length cDNA encoding calreticu-lin-like protein (designated as TaCRT) which is highlyexpressed in the wheat seedlings subjected to water-stress,is reported here. In addition, TaCRT’s genomic organiza-tion, subcellular localization, and its mRNA expressionpattern during wheat seedling response to PEG-induceddrought stress is described. The role of TaCRT in the plantresponse to drought stress is demonstrated by its effectswhen expressed in transgenic tobacco plants.
Materials and methods
Plant materials and water-stress experiments
Wheat (Triticum aestivum L.) genotype ‘Hanxuan 10’ with a higherdrought-tolerant phenotype was used in this study. After being
740 Jia et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

sterilized with 75% ethanol and washed with sterilized water, thewheat seeds were germinated and cultured with water in a controlled-growth chamber (2061 �C, 12 h light/dark cycle). Seedlings at thetwo-leaf stage (9-d-old) were treated by PEG-6000 (–0.5 MPa)solution, which, in pilot experiments, had been shown to constitutesignificant stress at this developmental stage. The treated plantswere stressed in the PEG-6000 solution for 1, 3, 6, 12, 24, 48, and72 h, respectively. The control seedlings were watered as normaland without PEG treatment. Leaf samples were collected from theseedlings at different time points and frozen quickly with liquidnitrogen and stored at –70 �C for RNA isolation and otheranalysis.Four genotypes of wheat and its relatives were used in Southern
hybridizations: Hanxuan 10, hexaploid (AABBDD) and threediploid genotypes including T. urartu (AA) accession No.1010004, Ae. speltoides (SS, closely related to the BB genome)accession No. IcAG 400046, and Ae. tauschii (DD) accession No.PH1878. Leaves from pot-grown plants were sampled for DNAisolation.Tobacco plants (Nicotiana benthamiana) were grown in pots filled
with vermiculite in a controlled-growth chamber with 12 h photo-period, 25 �C, 70% humidity and 45 lmol�m�2�s�1 illumination.
Cloning of full-length TaCRT cDNA
Total RNA was extracted from the leaf samples using the TrizolReagent (Tianwei) as described in the manufacturer’s instructions.Based on the candidate EST of a TaCRT from the cDNA libraryestablished in our laboratory (Pang et al., 2007), the putative full-length TaCRT cDNA was obtained by in silico cloning. Using totalRNA isolated from the leaves of 12 h PEG-treated wheat seedlingsas templates, the first-strand cDNA synthesis was performed withM-MLV Reverse Transcriptase reagent (Invitrogen) according tothe manufacturer’s instructions. To obtain full-length TaCRTcDNA, two primers, 5#-ACCACCACTTCCTCGTCTC-3# (sense)and 5#-TTCCCTCACACGAGACAAG-3# (antisense) weredesigned based on the lateral flanking sequence of ORF of theputative complete CRT cDNA.A total of 20 ll of the RT-PCR reaction mixture contained 1 ll
of 10-fold-diluted cDNAs (0.2 lg), 0.2 lM of each primer, 1.5 mMMgCl2, and 0.4 mM dNTPs, 13 buffer and 0.1 ll of proof readingPfu Pyrobest polymerase (5 U ll�1) (Takara). Amplification wasconducted in a DNA thermocycler (GeneAmp PCR System 9600,Applied Biosystems) using 30 cycles of 2 min at 94 �C, 1 min at61 �C, and 2 min at 72 �C; final extension at 72 �C for 10 min. Asa control, a Tubulin gene fragment was amplified using senseprimer 5#-AGAACACTGTTGTAAGGCTCAAC-3# and antisenseprimer 5#-GAGCTTTACTGC CTCGAACATGG-3# under condi-tions similar to those described for TaCRT, except for annealing at50 �C and 21 cycles.The RT-PCR generated a 1446 bp DNA fragment namely
TaCRT. This DNA fragment was subcloned into pGEM-T easyVector system (Tianwei) and introduced in E. coli JM109 accordingto the manufacturer’s recommendations. The plasmid DNA isolatedfrom the positive E. coli cells was digested with EcoRI, and theinserted DNA (TaCRT cDNA) was sequenced in both directionsusing the BigDye Terminator method on an autosequencer (ModelXL3730, Applied Biosystems).Database searches of the nucleotide and deduced amino acid
sequences were performed through the NCBI/GenBank/BlastSequence alignment and similarity among species were determinedby the megAlign program in DNAStar. Signal sequence waspredicted with SignalP (http://genome.cbs.dtu.dk/services/SignalP).The functional region and activity sites were identified usingPROSITE (http://expasy.hcuge.ch/sprot/ prosite.html) and SMARTmotif search program (http://coot.embl-heidelberg.de/SMART).
Subcellular localization of TaCRT
A full-length cDNA clone of TaCRT was inserted between theupstream GFP and the downstream constitutive CaMV 35S pro-moter in a pJIT163-GFP expression vector for constructing a 35S-TaCRT-GFP fusion protein. Restriction sites HindIII (5#) and SalI(3#) were added to the 5# and 3# ends of the coding region by thePCR method. The PCR product obtained was digested with HindIIIand SalI, and then ligated into the HindIII and SalI sites of thepJIT163-GFP plasmid polylinker to create a recombinant plasmidpJIT163-TaCRT-GFP for expressing the fusion protein. Thepositive plasmids were confirmed by restriction analysis, and furtherverified by sequencing. The recombinant plasmid pJIT163-TaCRT-GFP was transformed into living onion epidermal cells by GeneGun(Helios�) according to the instruction manual. The subcellularlocation of the TaCRT was detected by monitoring the transientexpression of GFP in onion epidermal cells. The transformed cellswere incubated in Murashige and Skoog (MS) medium at 28 �C for24–48 h and then were observed with a laser scanning confocalmicroscope (Olympus FV500) and the images obtained wererecorded automatically. The recombinant pJIT163-TaCRT-GFPplasmid and the control pJIT163-GFP plasmid were bombardedinto 20 onion epidermal segments, respectively. Twenty fluorescentcells of these segments were observed with identical results.
Southern blot analysis
Genomic DNA of hexaploid wheat and its diploid relatives wasextracted according to the procedure described by Stewart and Via(1993). Aliquots of genomic DNA (15 lg) was digested overnightwith EcoRI, EcoRV, and HindIII, respectively. The digested DNAwas fractionated in 0.8% agarose gel followed by blotting to nylonmembranes (Hybond N+, Amersham) overnight in 203 SSC. AfterUV cross-linking, the membrane was hybridized to a [a_32P]dCTP-labelled full-length TaCRT cDNA probe in 503 Denhardt’s solutionat 65 �C. The membrane was washed with 23 SSC, 0.1% SDS;0.23 SSC, 0.1% SDS, and 0.13 SSC, 0.1% SDS for 15 min at65 �C, respectively. The hybridized blot was exposed to a phosphorscreen (Kodak-K) for 2 d and visualized with the Molecular ImagerFX (Bio-Rad).
Northern blot analysis
Total RNA of each sample was isolated from the leaf tissue of thewheat plants stressed by PEG-6000 (–0.5 MPa) solution using theTrizol Reagent methods as described above. Twenty micrograms oftotal RNA from each sample was separated in 1.5% agarose gelswith 2% formaldehyde followed by transferring to nylon mem-branes (Hybond N+, Amersham) overnight in 203 SSC. The blottedmembranes were subsequently prehybridized for 1 h at 65 �C in503 Denhardt’s solution with the addition of 500 ll of 10 mg ml�1
of denatured salmon DNA (Promega) and then hybridized withthe [a_32P]dCTP-labelled full-length TaCRT cDNA probe under thesame conditions overnight. Washing and autoradiography of thehybridized membranes was performed as described for the Southernblot.
Quantitative real-time PCR
Treated with deoxyribonuclease I, the RNA samples used fornorthern blot also served as templates for the first-strand cDNAsynthesis by the Superscript First-Strand Synthesis System kit(Invitrogen). Quantitative real-time PCR (qRT-PCR) was performedin triplicate with an ABI PRISM� 7000 system using the SYBRGreen PCR master mix kit (Applied Biosystems) according to the
Cloning and functional analysis of TaCRT 741
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

manufacturer’s instructions. The relative amount of gene expressionwas calculated using the expression of Tubulin as internal standard.Oligonucleotides of qRT-PCR primers were as following: TaCRTforward primer 5#-GAAGCCCCCCAAATCTT-3# and reverseprimer 5#-CCTCACACGAGACAAGAAACAC-3#. Tubulin for-ward primer 5#- TGTGCCCCGTGCTGTTCTTATG-3# and reverseprimer 5#- CCCTTGGCCCAGTTGTTACCC-3#.The relative quantity of gene expression was detected using 2–DDCT
method (Livak et al., 2001). DDCT¼(CT,Target–CT,Tubulin)Time x –(CT,Target–CT,Tubulin)Time 0. The CT (cycle threshold) values for boththe target and internal control genes were means of triplicateindependent PCRs. Time x is any treated time point (1, 3, 6, 12, 24,48, or 72 h) and Time 0 represents the untreated time (0 h).
Construction of expression vector of pCAPE2-TaCRT and
Agrobacterium-mediated transformation of tobacco plants
PEBV (the tobravirus pea early browning virus) has already beendeveloped as an expression vector for the reporter gene GFP in bothP. sativum and N. benthamiana (MacFarlane and Popovich, 2000).The PEBV-based pCAPE vector system including two plasmids,pCAPE1 (assistant plasmid) and pCAPE2-GFP (control plasmid)were used to construct a plasmid vector to express TaCRT intobacco plants. For developing a vector pCAPE2 to express TaCRT,GFP in pCAPE2-GFP was replaced by the ORF sequence ofTaCRT. The restriction sites of SacI and SalI were introduced in the5# and 3# ends of the ORF sequence of TaCRT cDNA by PCR withforward primer 5#-AGCGAGCTCACCACCACTTCCTCGTCTC-3# and reverse primer 5#-TATCGTCGACTCAGTGGTGGTGAT-GATGGTGTCATGGTAGTCATC-3#. The underlined sequences ofthe primers correspond to the SacI and SalI restriction sites,respectively. In the reverse primer, the nucleotide sequences of sixhistidines (boxed region) were added before the stop codon (TGA)so that the expressed protein could have a His tag, allowingvalidation by a commercial His-tag monoclonal antibody if theexogenous TaCRT gene was expressed in the host N. benthamiana.The PCR product of TaCRT and the vector pCAPE2-GFP wereboth cleaved with SacI and SalI. The digested vector pCAPE2 andthe PCR fragment of TaCRT were then ligated together to generatea new recombinant vector pCAPE2-TaCRT.Assistant vector pCAPE1, control vector pCAPE2-GFP, and the
recombinant vector pCAPE2-TaCRT were transformed into Agro-bacterium tumefaciens strain GV3101 by electroporation (GenePulser II, Bio-Rad) as described by Shen and Forde (1989),respectively. The transformed agrobacteria were grown overnight at28 �C under kanamycin and rifampicin selection. The positiveclones were confirmed by restriction analysis, plasmidic PCR, andsequencing. An individual clone was grown in 20 ml of Luria Broth(LB) liquid medium including 0.1 M 2-morpholinoethanesulphonicacid (MES), 0.2 mM acetosyringone (As), 100 lg ml�1 rifampicinand 50 lg ml�1 kanamycin at 28 �C for 16–18 h with shaking (230rpm). At OD600�2.0, the bacteria were collected by centrifugation(7 500 rpm, 15 min). Cells were resuspended in about 20 mlinfiltration medium (0.1 M MES, 2 mM As, 0.2 M MgCl2), andadjusted to a final OD600 of 2.0, and then incubated at roomtemperature for 3 h without shaking. Agrobacterium culturescarrying pCAPE2-GFP and pCAPE2-TaCRT were mixed, respec-tively, with Agrobacterium carrying pCAPE1 at a ratio of 1:1, andinfiltrated into the abaxial side of the third pair of leaves on 5-week-old tobacco plants using a 5 ml syringe without needle. Twentyplants in each of three independent experiments were inoculated,respectively. The positive transgenic plants were confirmed bydetecting GFP fluorescence of pCAPE2-GFP transformants, PCRand RT-PCR assays of pCAPE2-TaCRT transformants. At least58 tobacco plants expressing pCAPE2-TaCRT were obtained andsubsequently analysed for their phenotypes.
Western blot analysis
Protein isolation and quantification were performed using methodsadapted from Laemmli et al. (1970) and Bradford (1976). Twentymicrograms of protein from each sample, including His-taggedTaCRT protein and control protein, were electrophoreticallyseparated in two pieces of 12.5% SDS–PAGE gels with Bio-RadMini-Protean gel rigs following the procedure described by Kyse-Anderson (1984). One of the gels was stained with CoomassieBrilliant Blue G250. The other was subsequently transferredelectrophoretically onto a nitrocellulose membrane (pore size:0.45 lm) using a semi-dry transfer blotter (Bio-Rad) in transferbuffer TBST (2 mM TRIS, 192 mM glycine, 20% methanol, and0.1% SDS). The membranes were blocked in TBS buffer with 3%BSA for 1 h and thereafter blotted with a commercial His-tagmonoclonal antibody for 3 h at a 1:2000 dilution. After extensivewashing, the bound primary antibody was detected with a horserad-ish peroxidase-conjugated goat antimouse IgG secondary antibodyusing the ECL technique according to the manufacturer’s protocol(Amersham).
Physiological analysis of tobacco plants expressing
TaCRT under drought stress
Tobacco plants expressing pCAPE2-TaCRT and pCAPE2-GFP,respectively, were cultured under normal conditions for 2 weeksbefore exposure to drought stress. When GFP fluorescence wasobserved in tobacco roots, drought stress was imposed bywithholding water in a growth chamber (25 �C, 50–60% relativehumidity, continuously illuminated at 45 lmol�m�2�s�1) untila lethal effect of dehydration was observed on most of the controlplants (wild type). Ten independent plants from each sample groupof three replicates were examined for all of the followingphysiological indexes.Leaf relative water content (RWC) was estimated according to
the method of Turner (1981). RWC(%)¼(fresh weight–dry weight)/(turgid weight–dry weight)3100.Water retention ability (WRA) was measured according to the
formula: WRA(%)¼(desiccated weight–dry weight)/(fresh weight–dry weight)3100. The leaves with the same age were taken andweighed (fresh weight), then desiccated for 24 h under controlledconditions (65% relative humidity and 25 �C), and weighed again(desiccated weight). The leaves were finally oven-dried overa period of 24 h at 90 �C to a constant weight (dry weight).Water use efficiency (WUE) was calculated with the formula:
WUE(%)¼net photosynthetic rate (Pn)/transpiration rate (Tr)3100.Fully expanded leaves with the same age were selected formeasuring Pn and Tr by the Li-6400 photosynthesis system (Li-CorInc.) according to the manufacturer’s instructions.Leaf membrance damage rate (MDR) was determined according
to the method of Sairam (1994) with a few modifications. Aconductivity meter (DDS-1, YSI) was used to measure the MDR ofthe sample. MDR(%)¼initial electrical conductivity/electrical con-ductivity after being boiled3100.
Results
Cloning of calreticulin cDNA
Previously, RNA from 12 h PEG-treated wheat (cv.Hanxuan 10) seedling was used to construct a cDNAlibrary by the method of suppression subtractive hybrid-ization (SSH). A total of 1833 high quality ESTsincluding 133 known function ESTs were obtained from
742 Jia et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

the cDNA library (Pang et al., 2007). A 535 bp EST,which is highly homologous to CRT genes, was chosen asa query probe for in silico cloning. Based on the sequencedata of wheat dbEST, a 1680 bp extended sequence wasobtained. Subsequently, a 1446 bp cDNA fragment wasamplified from the drought-treated wheat seedlings byRT-PCR using a pair of primers designed from theextended sequence. This fragment was fully sequencedand identified as a wheat CRT (TaCRT) cDNA clone(GenBank accession no. EF452301).
Characterization of TaCRT cDNA sequence
The TaCRT cDNA clone was 1446 bp in length, whichconsists of a 35 bp 5#-untranslated region (UTR), a 115bp 3# UTR, and a 1296 bp open reading frame (ORF).The ORF of TaCRT encodes 431 deduced amino acidresidues with a calculated molecular mass of 50.573 kDaand a predicted pI of 6.84. Like other known plant CRTs,the deduced coding sequence of TaCRT exhibits the samezonal characteristics of the three-domain structure typicalof animal CRTs (Michalak et al., 1992, 1999). The threedomains of TaCRT are the globular N-domain (residues34 to 218), the proline-rich central domain (P-domain)(residues 219 to 319) which is folded into an ‘extendedarm’, and the polyacidic C-domain (residues 320 to 431)(Fig. 1A).The highly conserved N-terminal domain was nearly
neutral with a 33-amino-acid-long signal sequence. Themature peptide of TaCRT has 398 amino acids anda molecular mass of 46.866 kDa. The N-domain alsocontained two well-conserved CRT family signaturemotifs of KFEQKIECGGGYVKLM and LMFGPDICGdescribed as CRT motif 1 and CRT motif 2 (Fig. 1A). Inthe N-domain, the conserved cysteines (Cys) are residues119, 151, and 177, of which the latter two are believed toform a disulphide bond, involved in folding of the nativeprotein. These amino acids correspond to Cys120 andCys146 of the bovine brain calreticulin, which wereinvolved in forming an intramolecular disulphide bridge inthe intact calreticulin (Matsuoka et al., 1994). A similarCys position was also found in the amino acid sequence ofbarley calreticulin (Cys136 and Cys162) (Chen et al., 1994).The N-domain is followed by the central P-domain,
which is a highly conserved proline-rich region andcontains 17 of the total 27 proline residues. Within thisregion, there are two types of triplicate repeats (A and B)that are highly conserved among CRTs from variousanimal and plant species. Repeat A has the structure ofPXXIXDXDAXKPEDWDE while the consensus se-quence for repeat B is GXWKPPXIXNPXYX (Fig. 1A).These two triplicate repeats are also highly conserved butnot invariant. The two types of repeats could enableTaCRT to form the extended hairpin loop configuration asdescribed previously in other CRTs (Ellgaard et al., 2001).
In front of the C-domain, there are 30 short hydropho-bic residues (amino acid 320 to 349) (Fig. 1A). Thepredicted C-terminal domain of TaCRT is less conserved,but highly acidic. 32% of acidic acids in TaCRT are foundin this domain, which is common to this protein family.The C-domain terminates with the amino acid motifHDEL, an ER-retention sequence.To investigate the sequence homology of TaCRT to
other known CRTs, a rooted phylogenetic tree wasestablished (Fig. 1B) based on 14 CRT protein sequencesfrom eight species including wheat, Arabidopsis, rice,maize, barley, tobacco, human, and mouse. As shown inFig. 1B, these CRTs were clustered into two distinctgroups, plant CRT and animal CRT groups. In plantCRTs, the CRT1 and CRT2 isoforms formed a subgroupdistinct from the CRT3 subgroup. TaCRT showed thehighest identity with rice CRT3 by 88%, and wasclustered into the plant CRT3 isoform subgroup whileanother wheat CRT-like protein (AY836753) belonged tothe plant CRT1/2 isoform subgroup. The isoform-specificclades of plant CRTs were similar when an analogousanalysis was performed by corresponding CRT nucleotidesequences (data not shown). All these bioinformaticsanalyses of TaCRT clearly indicate that this cDNA cloneencodes a wheat CRT protein (e.g. wheat CRT3 isoform).
Genomic organization of the TaCRT gene
To study the genomic organization and the copy numberof the TaCRT gene in hexaploid wheat, Southern blotanalysis of genomic DNA was performed using the full-length sequence of TaCRT cDNA as a probe (Fig. 2). Thehybridization patterns reveal that TaCRT is most likely toexist as three-copy genes in the genomes of hexaploidwheat, as three-hybridized bands were always evident ineach lane of genomic DNA digested with EcoRI, EcoRV,and HindIII, respectively (Fig. 2, lane 1). Similarly, the Bgenome contains at least two copies of this gene for eachof the three enzymes resulting in two bands in the blot(Fig. 2, lane 3). In the A and D genomes, there was onlyone-hybridized band of TaCRT digested by EcoRI orHindIII restriction (Fig. 2, lanes 2, 4), while two copieswere generated by EcoRV (Fig. 2, lanes 2, 4) probablycaused by a site in the intron. This analysis indicates thatthe TaCRT gene has three copies in hexaploid wheatgenomes and one or two copies in its ancestors.
Subcellular localization of the TaCRT protein
To address the subcellular localization of TaCRT in livingcells, a construct containing TaCRT fused in-frame withthe GFP (TaCRT::GFP) driven by the CaMV 35Spromoter was transiently expressed in living onionepidermal cells. As shown in Fig. 3, confocal microscopicexamination revealed that cells transferred with the un-conjugated GFP (control) exhibited a diffused distribution
Cloning and functional analysis of TaCRT 743
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

Fig. 1. Sequence alignment of TaCRT and other plant CRTs. (A) An alignment is shown for the deduced amino acid sequence of CRT from wheat,barley, rice, maize, Arabidopsis, and tobacco. Residues that were identical in all these proteins are shown in a black background. Dashes indicatedgaps introduced for optimal alignment. The numbers on the left indicate the amino acid position. Dashed lines indicate the predicted signal peptide.The conserved CRT family signature motifs 1 and 2 common to nearly all the CRTs are underlined. Solid triangles represent the positions of threehighly conserved Cys residues. A double-underlined region indicates the hydrophobic sequence. The A and B triplicate repeats are shown by boldunderlining. The ER-retention sequence is indicated. The approximate positions of the three domains (N, P, and C) are indicated with arrows. (B)Rooted phylogenetic tree based on the sequence alignment, including CRT1/2, and CRT3 protein sequences from rice; CRT1, CRT2, and CRT3 fromArabidopsis and maize, respectively; two mammalian CRT sequences from human and mouse. Two distinct isoform groups are presented in differentshades of grey. Accession numbers are indicated in parentheses. Abbreviations: Ta, Triticum aestivum; Hv, Hordeum vulgare; Os, Oryza sativa; Zm,Zea mays; At, Arabidopsis thaliana; Nt, Nicotiana tabacum; Hs, Homo sapiens; Mm, Mus musculus.
744 Jia et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

of green fluorescence throughout the cell (Fig. 3A). Bycontrast, when fused with the TaCRT, the GFP signal wasconfined to the cytoplasm and nucleus (Fig. 3B), suggest-ing that TaCRT is distributed in the cytoplasmic andnuclear compartments.
Expression pattern of TaCRT in response toPEG-induced drought stress
The cloning of TaCRT cDNA was based on an ESTsequence in the cDNA library constructed from 12 hwater-stressed seedlings of wheat, suggesting that theTaCRT gene might be expressed in response to droughtstress. To reveal the temporal expression pattern ofTaCRT in wheat seedling plants subjected to droughtstress, two-leaf seedlings (9-d-old) were stressed in thePEG-6000 (–0.5 MPa) solution for different periods oftime. The control was wheat seedlings watered as normalin the same condition.Northern blot and qRT-PCR both revealed similar
expression patterns as shown in Fig. 4 with triplicateindependent experiments, respectively. No expression ofthe TaCRT gene was detected in the control plants (CK).The TaCRT transcript began to appear in the seedlings 1 hafter stress treatment and peaked at 12 h. However, thelevel of the transcript decreased gradually in the treatedplants from 12 h to 72 h under the stress. Even at 72 hafter the stress treatment, TaCRT expression is about2-fold more than that at 1 h in the PEG-treated wheatseedlings (Fig. 4A). A similar expression pattern wasobtained by qRT-PCR using the RNA samples from at
least 25 seedlings for each time period (Fig. 4B). Clearly,TaCRT expression was significantly induced by PEGstress, suggesting that CRT may function as an importantchaperone protein for wheat drought-stress response.
Heterologous overexpression analysis of TaCRT intobacco plants
In order to verify TaCRT function, a virus-inducedexpression construct of TaCRT was developed (Fig. 5).Agrobacterium cultures carrying pCAPE2-GFP (Fig. 5B)and pCAPE2-TaCRT (Fig. 5C) were mixed respectivelywith pCAPE1 (Fig. 5A) at a ratio of 1:1 for thetransformation of host tobacco plants. The transformedtobacco plants containing both pCAPE2-GFP andpCAPE1 were used as positive controls, while wild-typetobacco plants were used as blank controls. A total of 70and 65 independent transgenic lines expressing pCAPE2-TaCRT or pCAPE2-GFP were generated, respectively.Green fluorescence was clearly and strongly detected inthe root tissues of tobacco plants containing pCAPE2-GFP at 17 d of post-inoculation (Fig. 6). The resultindicated that the agro-inoculation was successful.The pCAPE2-TaCRT transformants were identified with
tissue PCR amplification (Fig. 7A). The TaCRT fragmentswere apparently obtained from 58 of the pCAPE2-TaCRTtransformants, but not from the wild-type plants and thecontrol plants transformed with pCAPE2-GFP. TheTaCRT transcripts in leaves of tobacco were detected withRT-PCR (Fig. 7B). Again, the target TaCRT fragment wasonly obtained from the pCAPE2-TaCRT transformants.To testify further if TaCRT protein was expressed
correctly, a western blot was performed. As described inthe Materials and methods, 20 lg of total protein fromeach sample was electrophoretically separated on SDS-PAGE and transferred to a nitrocellulose membrane.Using the commercial His-tag monoclonal antibody, a pro-tein band which was almost consistent with the predictedmolecular mass of TaCRT, was apparently detected in thepCAPE2-TaCRT inoculated tobacco plants, but not fromthe wild-type plants (Fig. 7C). The immunoblot resultfurther showed that TaCRT was overexpressed in thetransformed tobacco plants.
Phenotype of the TaCRT-overexpressing tobaccoplants under drought stress
TaCRT-expressing plants exhibited no obvious phenotypicdifference from the GFP-expressing and WT plants underwell-watered conditions as revealed by four physiologicaltraits related to plant water status, including MDR, WUE,RWC, and WRA (F-test, P¼0.05). To examine theperformance of the TaCRT-expressing tobacco plantsunder drought stress, water was withheld from TaCRTand GFP transgenic, as well as WT tobacco, plants after
Fig. 2. Southern blot analysis of TaCRT. The ABD genome DNA ofhexaploid wheat cv. Hanxuan 10 (lane 1), A genome DNA of T. urartu(lane 2), S genome DNA of Ae. speltoides (lane 3) closely related to theB genome and D genome DNA of Ae. tauschii (lane 4) were digestedovernight with the restriction enzymes indicated. The full-lengthsequence of TaCRT cDNA was used as a probe. M: The origin andlambda DNA/HindIII-digest size markers (MBI Fermentas) are in-dicated in kilobase (kb) pairs on the left.
Cloning and functional analysis of TaCRT 745
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022
confirmation of successful transformation and expression.In the first week of drought treatments, both transgenicand WT tobacco plants showed a similar phenotype.However, after 3 weeks of drought treatment, phenotypicdifferences were clearly observed among the TaCRTtransgenic and other plants (Fig. 8A). Both WT and GFPtransgenic plants displayed severe leaf-wilting, while only40% of the TaCRT transgenic plants showed slight leaf-wilting, which demonstrated less wilting than the corre-sponding WT and GFP transgenic plants. GFP transgenicplants were very similar to WT plants in performance,indicating that the GFP had no effect on functionalexpression of TaCRT in tobacco plants. The resultsindicated that TaCRT was involved in the response towater stress and its over-expression may contribute todrought tolerance to some extent.The tissue samples from the TaCRT-expressing tobacco
plants and WT plants were also analysed for detectingtheir cellular-physiological status under the water stress.On day 22 of drought stress, MDR, WUE, RWC, andWRA were measured in WT and TaCRT transgenicplants. Corresponding to the previous phenotypic results,leaves of TaCRT transgenic plants displayed a lowerMDR, higher WUE and WRA at statistically significantlevel (P <0�01) compared with the wild-type plants (Fig.
8B). The transgenic plants also showed a slightly higherRWC although the increase was not significant. In plantcells, lower MDR, and higher WUE, RWC, and WRAwere believed to be responsible for stronger toleranceagainst osmotic stress. Therefore, analysis of thesecellular-physiological data again indicated that TaCRTtransgenic plants performed better than WT and the
Fig. 3. Subcellular localization of the TaCRT in onion epidermal cells. Cells were bombarded with construct carrying GFP (A) or TaCRT::GFP (B),respectively as described in the Materials and methods. GFP and TaCRT::GFP fusion proteins were transiently expressed under the control of CaMV35S promoter in onion epidermal cells and observed with a laser scanning confocal microscope. Images were taken in a dark field for greenfluorescence (1, 4) while the outlook of cell (2, 5) and the combination (3, 6) were photographed in a bright field.
Fig. 4. Expression patterns of TaCRT in response to PEG-modeled drought stress. (A) Northern blot analysis. (B) qRT-PCR analysis. Two-leafseedlings of common wheat cv. Hanxuan 10 were treated with –0.5 MPa PEG-6000 for 1–72 h. Total RNAs isolated from untreated (CK or 0 h) andthe stressed leaf tissue at different time period were probed with [a-32P]dCTP-labelled full-size TaCRT cDNA. The lower panel shows ethidiumbromide-stained 18 S rRNA to quantify equal RNA loading. Hybridization band were visualized by phosphoimager. Graphs show the mean 6SD ofresults from three independent experiments.
Fig. 5. The T-DNA region of PEBV-based binary vectors. 35SP,CaMV 35S promoter; CP, the coat protein coding region of PEBV; T,the NOS terminator; RB and LB, the left and right borders of theT-DNA region, respectively. (A) pCAPE1 containing full-length cDNAof PEBV RNA-1 with an intron inserted to stabilize the plasmid inbacteria. (B) pCAPE2-GFP containing full-length cDNA of PEBVRNA-2 with the GFP coding sequence. (C) pCAPE2-TaCRT witha full-length cDNA of TaCRT inserted into the RNA2 cDNA.
746 Jia et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

overexpression of TaCRT gene could enhance plantdrought tolerance.
Discussion
In the present study, the cloning of a full-length cDNA ofTaCRT containing an ORF of 1296 bp and encodinga protein of 431 amino acids and one termination codonfrom wheat seedlings has been described. The deducedamino acid sequence alignment (Fig. 1A) indicates thatTaCRT has high sequence identity (54–88%) with otherplant CRTs, whereas the similarity between animal andplant calreticulins is lower, only about 45%. Mammalianand plant CRTs form their own distinct clusters (Fig. 1B).The similarities among these 14 CRTs have been furtherconfirmed by their nucleotide sequence alignment and
phylogenetic analyses (data not shown). TaCRT hasa higher sequence identity with rice CRT3, maize CRT3,and Arabidopsis CRT3, respectively. It indicates thatTaCRT cDNA clone encodes a wheat CRT3 isoform. Theresult of sequence analysis showed an early duplicationevent in wheat genomes, from which the CRT1 (or CRT2if present) and CRT3 isoforms derive. This early di-vergence of CRTs into two groups (CRT1/CRT2 andCRT3) was also found in Arabidopsis, rice, maize, andBrassica rapa (Persson et al., 2003).Although plant CRTs have different isoforms, several
conserved regions exist in their sequences. Like otherplant CRTs, the deduced amino acid sequence of TaCRTalso contains the three typical domain organization (Fig.1A) proposed for animal CRTs (Michalak et al., 1999). Inaddition, the putative TaCRT protein consists of conser-vative amino acid residues involved in two well-
Fig. 6. Confirmation of the successful transformation by monitoring GFP expression. Image of the root epidermal cells from tobacco plantsinoculated with pCAPE2-GFP. After 17 d post-inoculation, the roots of pCAPE2-GFP transformed plants were observed under a laser scanningconfocal microscope. The photographs were taken in a dark field for green fluorescence of GFP (A); the outlook of the cells (B), and the combination(C) were photographed in a light field. Scale bar¼80 lm.
Fig. 7. Identification of the pCAPE2-TaCRT transformed tobacco plants. (A) PCR analysis of tobacco plants with pCAPE2-TaCRT. M: 200-bpladder; Lane 1, pCAPE2-TaCRT plasmid DNA was used as template (positive control); Lane 2, wild-type tobacco plant genomic DNA was used astemplate (negative control); Lane 3, pCAPE2-GFP transformed tobacco plant; Lanes 4–9, pCAPE2-TaCRT transformed transgenic tobacco plantsharbouring pCAPE2-TaCRT, Lane 7 represents a candidate transformant but no target fragment was detected. (B) Detection of TaCRT transcripts byRT-PCR. M: 200-bp ladder; Lane 1, wild-type plant (control); Lanes 2–6, the PCR positive pCAPE2-TaCRT transformed tobacco plants. Theconstitutively expressed 500 bp Tubulin gene was used as an internal standard. (C) Western blot confirmation of TaCRT protein expression. Left:SDS-PAGE analysis using Coomassie Brilliant Blue G250 staining. Right: immunoblot analysis of TaCRT protein using His-tag monoclonalantibody. M: low molecular mass marker; Lane 1, wild-type tobacco plant (control); Lane 2, pCAPE2-TaCRT transformed tobacco plant.
Cloning and functional analysis of TaCRT 747
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

conserved CRT family signature motifs and two types oftriplicate repeats. The remarkable similarity betweenTaCRT and other known CRTs in both the deducedamino acid sequences and the protein zonal structuresreinforces the viewpoint that calreticulin protein is highlyconserved in all eukaryotic organisms (Chen et al., 1994;Michalak et al., 1999; Persson et al., 2003).As a major Ca2+-binding protein, CRT was originally
detected in the ER (Ostwald and MacLennan, 1974).Sequence analysis reveals that all plant calreticulins so farcloned including TaCRT contain a typical ER retentionsignal HDEL sequence, with the exception of Euglenacalreticulin which has KDEL (Navazio et al., 1998).Consistent with the signal, plant CRT has been locatedprimarily within the lumen of ER. However, CRT wasalso found outside the ER compartment in plant cells,including the cytoplasm of certain cells and nucleus(Dedhar, 1994), nuclear envelope (Napier et al., 1995),spindle apparatus of dividing cells (Denecke et al., 1995),plasmodesmata in root cells (Baluska et al., 1999), the cellsurface (Coppolino and Dedhar, 1998) and Golgi com-partment (Borisjuk et al., 1998). Calreticulin is totallyabsent from the vacuole, the major Ca2+ store in plantcells (Opas et al., 1996). Our GFP transient expressionassay clearly indicates that TaCRT was mainly located inthe cytoplasmic and nuclear compartments in onionepidermal cells (Fig. 3). All these suggest that CRT hasmultiple subcellular locations which might result fromcovalent modification such as phosphorylation or Ca2+-binding (Sharma et al., 2004). The precise mechanism(s)how the protein relocates from the ER to the outside ofthe ER, despite having the ER-retention signal, remains tobe ascertained. The multiple location of TaCRT indicatesthat it may be involved in multiple cellular processes. Thein vivo studies in wheat plants currently being conductedin our laboratory aim to characterize further the in-tracellular distribution of TaCRT.Plant calreticulin does not strictly follow the rule ‘one
protein, one gene’ assessed for its animal counterpart, butis encoded by a low copy-number gene family (Chenet al., 1994; Napier et al., 1995; Coughlan et al., 1997;Nelson et al., 1997; Michalak et al., 1999; Li andKomatsu, 2000). Several investigations have establishedthat plants contain two or multiple CRT isoforms (Chenet al., 1994; Nelson et al., 1997; Persson et al., 2003).Southern blot analysis in this study suggested that theTaCRT was most probably a low-copy gene. Examiningthe genomic architecture of the TaCRT gene in wheat willprovide more information about the intron/exon organiza-tion and evolution of the gene.CRTs have been implicated in plant growth and
development (Menegazzi et al., 1993), where theyregulated calcium signalling, and assisted protein folding(Nardi et al., 2006). It has also been shown that a widerange of developmental and environmental stimuli differ-
entially affect the expression of CRT in plant cells(Borsjuk et al., 1998). Sharma et al. (2004) found theincreased expression of rice CRT under cold stress,suggesting its potential role in regulating plant stressresponses. The growing data support the functionaldiversity of CRT protein in both animal and plantsystems. Here, we are examining TaCRT function inwheat drought-stress responses. Results from our northernblot showed that no TaCRT expression was detected in theunstressed wheat seedlings, and the transcription ofTaCRT was greatly induced by PEG-stress (Fig. 4). Thevaried expression levels of TaCRT in the PEG-stressedseedlings indicated that cells under drought stress respondquickly by elevating the transcription levels. Persson et al.(2003) revealed that different isoforms of CRT inArabidopsis responded differently to applied externalstimuli. CRT1 and CRT2 isoforms showed a slowerinduction under salt or tunicamycin whereas the responseof CRT3 appeared to be more rapid. The CRT2 wasreported in response to both tunicamycin and dithiothrei-tol in Arabidopsis. Although the time and level of theinduced expression of CRTs are different, these reportsclearly demonstrated that CRT expression was increasedunder stress conditions, supporting the up-regulation ofTaCRT described here. The up-regulated expression ofCRTs may be a conserved self-protection mechanismacquired during long-term evolution and should facilitatethe survival of the plants under unfavourable osmoticconditions.To investigate the in vivo role of TaCRT in plant
drought resistance, TaCRT was over-expressed in tobaccoplants by the PEBV-based plant expression vector. Plantvirus-based vectors have proved to be important tools forstudying gene function because of their relatively conve-nient engineering, which does not require the developmentof stable transformants and the interval between cloningand phenotypic analysis is significantly short (Lindboetet al., 2001). The PEBV-based binary vector system (Fig.5) was used as a plant gene expression vector since it hasbeen confirmed as an efficient vector in N. benthamianaand tomato. Moreover, PEBV has already been developedas an expression vector for the reporter gene GFP in N.benthamiana. The host plant used here for the geneexpression was tobacco (N. benthamiana), one of themost-studied hosts for developmental and genetic analysisby far. Via Agrobacterium mediation, a total of 58 and 68,respectively, pCAPE2-TaCRT and pCAPE2-GFP trans-formed plants were successfully obtained. The transgenictobacco plants were successfully confirmed by PCR, RT-PCR assays, and western blot (Fig. 7).MDR, RWC, WUE, and WRA are the typically
physiological indices for the evaluation of droughttolerance and resistance in crop plants. Plants with higherRWC, WUE, WRA, and lower MDR have highertolerance and are more resistant to drought stress.
748 Jia et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022
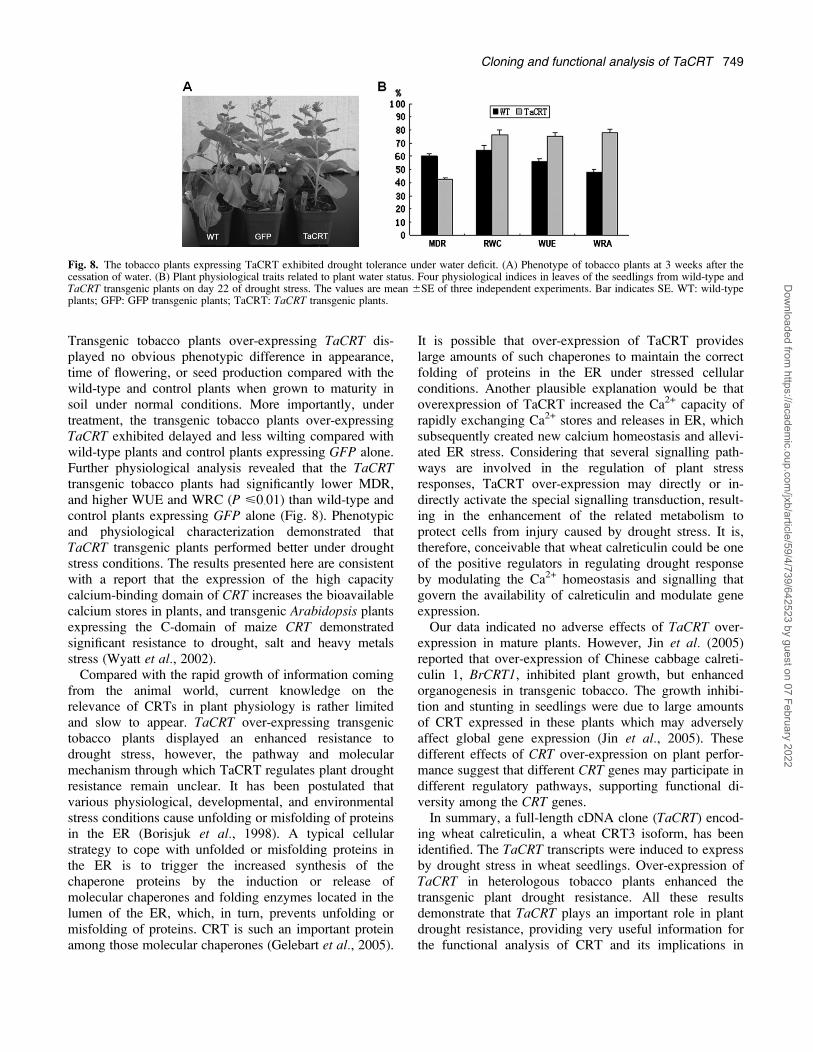
Transgenic tobacco plants over-expressing TaCRT dis-played no obvious phenotypic difference in appearance,time of flowering, or seed production compared with thewild-type and control plants when grown to maturity insoil under normal conditions. More importantly, undertreatment, the transgenic tobacco plants over-expressingTaCRT exhibited delayed and less wilting compared withwild-type plants and control plants expressing GFP alone.Further physiological analysis revealed that the TaCRTtransgenic tobacco plants had significantly lower MDR,and higher WUE and WRC (P <0�01) than wild-type andcontrol plants expressing GFP alone (Fig. 8). Phenotypicand physiological characterization demonstrated thatTaCRT transgenic plants performed better under droughtstress conditions. The results presented here are consistentwith a report that the expression of the high capacitycalcium-binding domain of CRT increases the bioavailablecalcium stores in plants, and transgenic Arabidopsis plantsexpressing the C-domain of maize CRT demonstratedsignificant resistance to drought, salt and heavy metalsstress (Wyatt et al., 2002).Compared with the rapid growth of information coming
from the animal world, current knowledge on therelevance of CRTs in plant physiology is rather limitedand slow to appear. TaCRT over-expressing transgenictobacco plants displayed an enhanced resistance todrought stress, however, the pathway and molecularmechanism through which TaCRT regulates plant droughtresistance remain unclear. It has been postulated thatvarious physiological, developmental, and environmentalstress conditions cause unfolding or misfolding of proteinsin the ER (Borisjuk et al., 1998). A typical cellularstrategy to cope with unfolded or misfolding proteins inthe ER is to trigger the increased synthesis of thechaperone proteins by the induction or release ofmolecular chaperones and folding enzymes located in thelumen of the ER, which, in turn, prevents unfolding ormisfolding of proteins. CRT is such an important proteinamong those molecular chaperones (Gelebart et al., 2005).
It is possible that over-expression of TaCRT provideslarge amounts of such chaperones to maintain the correctfolding of proteins in the ER under stressed cellularconditions. Another plausible explanation would be thatoverexpression of TaCRT increased the Ca2+ capacity ofrapidly exchanging Ca2+ stores and releases in ER, whichsubsequently created new calcium homeostasis and allevi-ated ER stress. Considering that several signalling path-ways are involved in the regulation of plant stressresponses, TaCRT over-expression may directly or in-directly activate the special signalling transduction, result-ing in the enhancement of the related metabolism toprotect cells from injury caused by drought stress. It is,therefore, conceivable that wheat calreticulin could be oneof the positive regulators in regulating drought responseby modulating the Ca2+ homeostasis and signalling thatgovern the availability of calreticulin and modulate geneexpression.Our data indicated no adverse effects of TaCRT over-
expression in mature plants. However, Jin et al. (2005)reported that over-expression of Chinese cabbage calreti-culin 1, BrCRT1, inhibited plant growth, but enhancedorganogenesis in transgenic tobacco. The growth inhibi-tion and stunting in seedlings were due to large amountsof CRT expressed in these plants which may adverselyaffect global gene expression (Jin et al., 2005). Thesedifferent effects of CRT over-expression on plant perfor-mance suggest that different CRT genes may participate indifferent regulatory pathways, supporting functional di-versity among the CRT genes.In summary, a full-length cDNA clone (TaCRT) encod-
ing wheat calreticulin, a wheat CRT3 isoform, has beenidentified. The TaCRT transcripts were induced to expressby drought stress in wheat seedlings. Over-expression ofTaCRT in heterologous tobacco plants enhanced thetransgenic plant drought resistance. All these resultsdemonstrate that TaCRT plays an important role in plantdrought resistance, providing very useful information forthe functional analysis of CRT and its implications in
Fig. 8. The tobacco plants expressing TaCRT exhibited drought tolerance under water deficit. (A) Phenotype of tobacco plants at 3 weeks after thecessation of water. (B) Plant physiological traits related to plant water status. Four physiological indices in leaves of the seedlings from wild-type andTaCRT transgenic plants on day 22 of drought stress. The values are mean 6SE of three independent experiments. Bar indicates SE. WT: wild-typeplants; GFP: GFP transgenic plants; TaCRT: TaCRT transgenic plants.
Cloning and functional analysis of TaCRT 749
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

plant genetic improvement. Further studies, includingcharacterizing the regulation of the signal transductionnetwork that controls TaCRT production, will extend ourunderstanding of the biological role and function of CRTsin plant development and growth as well as the responsesto various biotic and abiotic stresses.
Acknowledgements
This work was supported by the National Basic Research Programof China (2004CB117202) and the National Transgenic PlantsProgram of China (JY03-A-14). The authors thank Dr DaowenWang and Professor Zhensheng Li (Institute of Genetics andDevelopmental Biology, Chinese Academy of Sciences, Beijing,China) for providing tobacco seeds, the PEBV-based pCAPE vectorsystem and pJIT163-GFP expression vector.
References
Baluska F, Samaj J, Napier R, Volkmann D. 1999. Maizecalreticulin localizes preferentially to plasmodesmata in root apex.The Plant Journal 19, 481–488.
Borisjuk N, Sitailo L, Adler K, Malysheva L, Tewes A,Borisjuk L, Manteuffel R. 1998. Calreticulin expression inplant cells: developmental regulation, tissue specificity andintracellular distribution. Planta 206, 504–514.
Bradford MM. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing the principleof protein–dye binding. Analytical Biochemistry 72, 248–254.
Burns K, Duggan B, Atkinson EA, Famulski KS, Nemer M,Bleackley RC, Michalak M. 1994. Modulation of geneexpression by calreticulin binding to the glucocorticoid receptor.Nature 367, 476–480.
Chen D, Texada DE, Duggan C, Liang C, Reden TB,Kooragayala LM, Langford MP. 2005. Surface calreticulinmediates muramyl dipeptide-induced apoptosis in RK13 cells.Journal of Biological Chemistry 280, 22425–22436.
Chen F, Hayes PM, Mulrooney DM, Pan A. 1994. Identificationand characterization of cDNA clones encoding plant calreticulinin barley. The Plant Cell 6, 835–843.
Chen MH, Tian GW, Gafni Y, Citovsky V. 2005. Effects ofcalreticulin on viral cell-to-cell movement. Plant Physiology 138,1866–1876.
Coppolino MG, Dedhar S. 1998. Calreticulin. InternationalJournal of Biochemistry and Cell Biology 30, 553–558.
Coppolino MG, Woodside MJ, Demaurex N, Grinstein S, St-Arnaud R, Dedhar S. 1997. Calreticulin is essential for integrin-mediated calcium signalling and cell adhesion. Nature 386,843–847.
Coughlan SJ, Hastings C, Winfrey Jr RW. 1997. Cloning andcharacterization of the calreticulin gene from Ricinus communisL. Plant Molecular Biology 34, 897–911.
Dedhar S. 1994. Novel functions for calreticulin: interaction withintegrins and modulation of gene expression? Trends in Bio-chemical Sciences 19, 269–271.
Denecke J, Carlsson LE, Vidal S, Hoglund AS, Ek B, vanZeijl MJ, Sinjorgo KM, Palva ET. 1995. The tobacco homologof mammalian calreticulin is present in protein complexes in vivo.The Plant Cell 7, 391–406.
Dresselhaus T, Hagel C, Lorz H, Kranz E. 1996. Isolation ofa full-length cDNA encoding calreticulin from a PCR library of invitro zygotes of maize. Plant Molecular Biology 31, 23–34.
Ellgaard L, Riek R, Herrmann T, Guntert P, Braun D,Helenius A, Wuthrich K. 2001. NMR structure of thecalreticulin P-domain. Proceedings of the National Academy ofSciences, USA 98, 3133–3138.
Fliegel L, Burns K, MacLennan DH, Reithmeier RA,Michalak M. 1989. Molecular cloning of the high affinitycalcium-binding protein (calreticulin) of skeletal muscle sarcoplas-mic reticulum. Journal of Biological Chemistry 264, 21522–21528.
Gardai SJ, McPhillips KA, Frasch SC, Janssen WJ,Starefeldt A, Murphy-Ullrich JE, Bratton DL,Oldenborg PA, Michalak M, Henson PM. 2005. Cell-surfacecalreticulin initiates clearance of viable or apoptotic cells throughtrans-activation of LRP on the phagocyte. Cell 123, 321–334.
Gelebart P, Opas M, Michalak M. 2005. Calreticulin, a Ca2+-binding chaperone of the endoplasmic reticulum. InternationalJournal of Biochemistry and Cell Biology 37, 260–266.
Guo L, Nakamura K, Lynch J, Opas M, Olson EN, Agellon LB,Michalak M. 2002. Cardiac-specific expression of calcineurinreverses embryonic lethality in calreticulin-deficient mouse.Journal of Biological Chemistry 277, 50776–50779.
Hassan AM, Wesson C, Trumble WR. 1995. Calreticulin is themajor Ca2+-storage protein in the endoplasmic reticulum of thepea plant (Pisum sativum). Biochemical and Biophysical Re-search Communications 211, 54–59.
Heilmann I, Shin J, Huang J, Perera IY, Davies E. 2001.Transient dissociation of polyribosomes and concurrent recruit-ment of calreticulin and calmodulin transcripts in gravistimulatedmaize pulvini. Plant Physiology 127, 1193–1203.
Huang L, Franklin AE, Hoffman NE. 1993. Primary structureand characterization of an Arabidopsis thaliana calnexin-likeprotein. Journal of Biological Chemistry 268, 6560–6566.
Jaubert S, Ledger TN, Laffaire JB, Piotte C, Abad P,Rosso MN. 2002. Direct identification of stylet secreted proteinsfrom root-knot nematodes by a proteomic approach. Molecularand Biochemical Parasitology 121, 205–211.
Jin ZL, Hong JK, Yang KA, Koo JC, Choi YJ, Chung WS,Yun DJ, Lee SY, Cho MJ, Lim CO. 2005. Over-expression ofChinese cabbage calreticulin 1, BrCRT1, enhances shoot and rootregeneration, but retards plant growth in transgenic tobacco.Transgenic Research 14, 619–626.
Kennedy TE, Kuhl D, Barzilai A, Sweatt JD, Kandel ER. 1992.Long-term sensitization training in Aplysis leads to an increase incalreticulin, a major presynaptic calcium-binding protein. Neuron9, 1013–1024.
Krause KH, Michalak M. 1997. Calreticulin. Cell 88, 439–443.Kyse-Anderson J. 1984. Semi-dry blotting. Journal of Biochemicaland Biophysical Methods 10, 203–209.
Laporte C, Vetter G, Loudes AM, Robinson DG, Hillmer S,Stussi-Garaud C, Ritzenthaler C. 2003. Involvement of thesecretory pathway and the cytoskeleton in intracellular targetingand tubule assembly of Grapevine fanleaf virus movementprotein in tobacco BY-2 cells. The Plant Cell 15, 2058–2075.
Laemmli UK. 1970. Cleavage of structural proteins during theassembly of head of bacteriophage T4. Nature 227, 680–685.
Lenartowska M, Karas K, Marshall J, Napier R, Bednarska E.2002. Immunocytochemical evidence of calireticulin-like proteinin pollen tubes and styles of Petunia hybrida Hort. Protoplasma219, 23–30.
Li Z, Komatsu S. 2000. Molecular cloning and characterization ofcalreticulin, a calcium-binding protein involved in the regenera-tion of rice cultured suspension cells. European Journal ofBiochemistry 267, 737–745.
Lim CO, Kim HY, Kim MG, Lee SI, Chung WS, Park SH,Hwang I, Cho MJ. 1996. Expressed sequence tags of Chinesecabbage flower bud cDNA. Plant Physiology 111, 577– 588.
750 Jia et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022

Lindboet JA, Fitzmaurice WP, della-Cioppa G. 2001. Virusmediated reprogramming of gene expression in plants. CurrentOpinion in Plant Biology 4, 181–185.
Livak KJ, Schmittgen TD. 2001. Analysis of relative geneexpression data using real-time quantitative PCR and the 2-(DeltaDelta C(T)) method. Methods 25, 402–408.
MacFarlane SA, Popovich AH. 2000. Efficient expression of foreignproteins in roots from tobravirus vectors. Virology 267, 29–35.
Martin V, Groenendyk J, Steiner SS, Guo L, Dabrowska M,Parker JM, Muller-Esterl W, Opas M, Michalak M. 2006.Identification by mutational analysis of amino acid residuesessential in the chaperone function of calreticulin. Journal ofBiological Chemistry 281, 338–346.
Matsuoka K, Seta K, Yamakawa Y, Okuyama T, Shinoda T,Isobe T. 1994. Covalent structure of bovine brain calreticulin.Biochemical Journal 298, 435–442.
Menegazzi P, Guzzo F, Baldan B, Mariani P, Treves S. 1993.Purification of calreticulin-like protein(s) from spinach leaves.Biochemical and Biophysical Research Communications 190,1130–1135.
Michalak M, Milner RE, Burns K, Opas M. 1992. Calreticulin.Biochemical Journal 285, 681–692.
Michalak M, Robert Parker JM, Opas M. 2002. Ca2+ signalingand calcium binding chaperones of the endoplasmic reticulum.Cell Calcium 32, 269–278.
Michalak M, Corbett EF, Mesaeli N, Nakamura K, Opas M.1999. Calreticulin: one protein, one gene, many functions.Biochemical Journal 344, 281–292.
Nakamura K, Zuppini A, Amaudeau S, et al. 2001. Functionalspecialization of calreticulin domains. Journal of Cell Biology154, 961–972.
Napier RM, Trueman S, Henderson J, Boyce JM, Hawes C,Fricker MD, Venis MA. 1995. Purification, sequencing andfunctions of calreticulin from maize. Journal of ExperimentalBotany 46, 1603–1613.
Nardi MC, Feron R, Navazio L, Mariani P, Pierson E, Wolters-Arts M, Knuiman B, Mariani C, Derksen J. 2006. Expressionand localization of calreticulin in tobacco anthers and pollentubes. Planta 223, 1263–1271.
Navazio L, Nardi MC, Pancaldi S, Dainese P, Baldan B,Fitchette-Laine AC, Faye L, Meggio F, Martin W, Mariani P.1998. Functional conservation of calreticulin in Euglena gracilis.Journal of Eukaryot Microbiology 45, 307–313.
Nelson DE, Glaunsinger B, Bohnert HJ. 1997. Abundantaccumulation of the calcium-binding molecular chaperone calre-ticulin in specific floral tissues of Arabidopsis thaliana. PlantPhysiology 114, 29–37.
Opas M, Szewczenko-Pawlikowski M, Jass GH, Mesaeli N,Michalak M. 1996. Calreticulin modulates cellular adhesivenessvia regulation of expression of vinculin. The Journal of CellBiology 135, 1–11.
Ostwald TJ, MacLennan DH. 1974. Isolation of a high affinitycalcium-binding protein from sarcoplasmic reticulum. Journal ofBiological Chemistry 249, 974–979.
Pang XB, Mao XG, JIng RL, Shi JF, Gao T, Chang XP, Li YF.2007. Analysis of gene expression profile responsed to water
stress in wheat (Triticum aestivum L.) seedling. Acta AgronomicaSinica 33, 333–336.
Persson S, Rosenquist M, Svensson K, Galvao R, Boss WF,Sommarin M. 2003. Phylogenetic analyses and expressionstudies reveal two Distinct groups of calreticulin Isoforms inhigher plants. Plant Physiology 133, 1385–1396.
Porcellini S, Traggiai E, Schenk U, Ferrera D, Matteoli M,Lanzavecchia A, Michalak M, Grassi F. 2006. Regulation ofperipheral T cell activation by calreticulin. The Journal ofExperimental Medicine 203, 461–471.
Sairam RK. 1994. Effect of moisture stress on physiologicalactivities of two contrasting wheat genotypes. Indian Journal ofExperimental Biology 32, 594–593.
Sharma A, Isogai M, Yamamoto T, Sakaguchi K, Hashimoto J,Komatsu S. 2004. A novel interaction between calreticulin andubiquitin-like nuclear protein in rice. Plant Cell Physiology 45,684–692.
Shen W, Forde BG. 1989. Efficient transformation of Agro-bacterium spp. by high voltage electroporation. Nucleic AcidsResearch 17, 8385.
Smith MJ, Koch GL. 1989. Multiple zones in the sequence ofcalreticulin (CRP55, calregulin, HACBP), a major calciumbinding ER/SR protein. The EMBO Journal 8, 3581–3586.
Smith MJ. 1992a. A C. elegans gene encoded a protein homolo-gous to mammalian calreticulin. DNA Sequence: The Journal ofDNA Sequencing and Mapping 2, 235–240.
Smith MJ. 1992b. Nucleotide sequence of a Drosophila melano-gaster gene encoding a calreticulin homologue. DNA Sequence:The Journal of DNA Sequencing and Mapping 3, 247–250.
Stewart CN, Via LE. 1993. A rapid CTAB DNA isolationtechnique useful for RAPD fingerprinting and other PCRapplications. Biotechniques 14, 748–750.
Stephen K, Fujiki Kazuhiro, Dixon Brian. 2004. Molecularcloning and characterization of calreticulin from rainbow trout(Oncorhynchus mykiss). Immunogenetics 55, 717–723.
Treves S, Zorzato F, Pozzan T. 1992. Identification of calreticulinisoforms in the central nervous system. Biochemical Journal 287,579–581.
Turner NC. 1981. Techniques and experimental approachesfor the measurement of plant water status. Plant and Soil 58,339–366.
Williams CM, Zhang G, Michalak M, Cass DJ. 1997. Calcium-induced protein protein phosphorylation and changes in levels ofcalmodulin and calreticulin in maize sperm cells. Sexual PlantReproduction 10, 83–88.
Williams DB. 2006. Beyond lectins: the calnexin/calreticulinchaperone system of the endoplasmic reticulum. Journal of CellScience 119, 615–623.
Wyatt SE, Tsou PL, Robertson D. 2002. Expression of thehigh capacity calcium-binding domain of calreticulin increasesbioavailable calcium stores in plants. Transgenic Research 11,1–10.
Zhang G, Schmidt O, Asgari S. 2006. A calreticulin-like proteinfrom endoparasitoid venom fluid is involved in host hemocyteinactivation. Developmental and Comparative Immunology 30,756–764.
Cloning and functional analysis of TaCRT 751
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/4/739/642523 by guest on 07 February 2022




















