
Molecular Basis of Oligoubiquitin-Dependent Internalization of Membrane Proteins in Mammalian Cells
16
Molecular Basis of Oligoubiquitin-Dependent Internalization of Membrane Proteins in Mammalian Cells Herve Barriere 1 , Csilla Nemes 1 , Delphine Lechardeur 1 , Mina Khan-Mohammad, Klaus Fruh 2 and Gergely L. Lukacs 1 1 Program in Cell and Lung Biology, Hospital for Sick Children Research Institute and Department of Laboratory Medicine and Pathobiology, University of Toronto, 555 University Avenue, Toronto, Ontario, Canada, M5G 1X8 2 Molecular Microbiology and Immunology, Oregon Health and Science University, Vaccine and Gene Therapy Institute, 505 NW 185th Ave., Beaverton, OR, 97006, USA *Corresponding author: G. L. Lukacs, [email protected] Ubiquitination induced down-regulation of cell surface proteins by internalization and lysosomal targeting plays a fundamental role in cell physiology and patho- genesis of diseases. The molecular basis of a single ubiquitin (Ub) as an autonomous endocytic signal, the widely accepted mechanism, however, remains elusive in higher eukaryotes. Using Ub containing reporter proteins without signalling abilities, we present evidence that only multiple Ub moieties, linked either covalently or assembled as oligomers with an intact interface for recognition by Ub-interacting motifs (UIMs), are recog- nized by the endocytic machinery in vivo and associate with a subset of Ub-binding clathrin adaptors in vitro. Genetic and pharmacological approaches show that internalization of plasma membrane proteins harbouring multiple Ub moieties is clathrin-dependent, but caveolin- independent. Functional assays demonstrate the cargo- dependent involvement of eps15/15R and epsin, UIM containing clathrin adaptors, in the endocytosis of model proteins, CD4 and the activated b 2 -adrenergic receptor complex, containing polymeric or oligomeric Ub. These results provide a paradigm for the clathrin- mediated uptake of ubiquitinated membrane proteins in mammalian cells, requiring the assembly of multiple UIM–Ub interactions to overcome the low affinity bind- ing of mono-Ub to UIM. Key words: caveolin, CD4, clathrin, endocytosis, eps15, epsin, UIM, b 2 -adrenergic receptor, ubiquitin Received 29 September 2005, revised and accepted for publication 29 November 2005, published on-line 12 January 2006 Post-translational attachment of ubiquitin (Ub) to polypep- tides has a fundamental role in modulating the plasma membrane protein composition. Polyubiquitination prevents the delivery of non-native membrane proteins by signalling their proteasome-dependent degradation at the endoplas- mic reticulum (ER) or shunting them for vacuolar/lysoso- mal proteolysis from post-ER compartments (1,2). While ubiquitination serves as the primary endocytic signal for the down-regulation of plasma membrane proteins in yeast (3,4,5), a variety of short peptide motifs [e.g. YXXF, where F is a bulky hydrophobic residue and (D/ E)XXXL(L/I)] have been established as internalization motifs in mammalian cells (1). Recently, monoubiquitination also emerged as a recognition signal in the retrieval of plasma membrane receptors and transporters from the cell sur- face in higher eukaryotes (3,4,8–10). The notion that mono-Ub represents an autonomous endocytic signal in mammalian cells was primarily based on the internalization activity of fusion proteins, containing a Ub moiety and the truncated epidermal growth factor (EGF), platelet-derived growth factor (PDGF) and the IL-2 receptor (5,8,10). Considering that these receptors have intrinsic homo- and/or hetero-oligomerization propensity, as well as the fused Ub moiety may have been subjected to cycles of ubiquitination-deubiquitination at the plasma membrane/ endosomes (11), the presentation of oligomeric Ub adduct(s) to the endocytic machinery cannot be ruled out. Molecular recognition of Ub can be achieved by Ub- receptors that contain Ub-binding module(s), such as Ub- interacting motif (UIM), Ub associated, coupling of Ub to endoplasmic reticulum associated degradation, Ub E2 var- iant and Npl4 Zn finger domains (3,12). It is assumed that decoding of mono-Ub as an endocytic signal is mediated by modular adaptor molecules (e.g. epsin and eps15/ eps15R) (4,13,14), harbouring multiple UIMs and other functional domains that trigger coat proteins assembly and subsequent vesicular budding similar to cargo sorting at the trans Golgi network and early endosomes (6,15,16). Intriguingly, while the preferential association of poly Ub chain to the conserved UIM of the S5a subunit of protea- some correlates well with the proteasome-mediated degradation of polyubiquitinated targets, the negligible binding of mono-Ub to the UIMs of eps15 and epsin is at variance with their presumed role in monoubiquitinated cargo recruitment to clathrin-coated pits/vesicles (CCP/ CCV) (13,17–19). Multiple internalization pathways have been invoked in the uptake of ubiquitinated proteins from the cell surface. While several plasma membrane proteins that undergo oligo/polyubiquitination-induced down-regulation (e.g. Traffic 2006; 7: 282–297 # 2006 The Authors Blackwell Munksgaard doi: 10.1111/j.1600-0854.2006.00384.x 282
Transcript of Molecular Basis of Oligoubiquitin-Dependent Internalization of Membrane Proteins in Mammalian Cells

Molecular Basis of Oligoubiquitin-DependentInternalization of Membrane Proteins inMammalian Cells
Herve Barriere1, Csilla Nemes1, DelphineLechardeur1, Mina Khan-Mohammad, KlausFruh2 and Gergely L. Lukacs1
1Program in Cell and Lung Biology, Hospital for SickChildren Research Institute and Department of LaboratoryMedicine and Pathobiology, University of Toronto, 555University Avenue, Toronto, Ontario, Canada, M5G 1X82Molecular Microbiology and Immunology, Oregon Healthand Science University, Vaccine and GeneTherapy Institute, 505 NW 185th Ave., Beaverton, OR,97006, USA*Corresponding author: G. L. Lukacs, [email protected]
Ubiquitination induced down-regulation of cell surfaceproteins by internalization and lysosomal targetingplays a fundamental role in cell physiology and patho-genesis of diseases. The molecular basis of a singleubiquitin (Ub) as an autonomous endocytic signal, thewidely accepted mechanism, however, remains elusivein higher eukaryotes. Using Ub containing reporterproteins without signalling abilities, we present evidencethat only multiple Ub moieties, linked either covalentlyor assembled as oligomers with an intact interface forrecognition by Ub-interacting motifs (UIMs), are recog-nized by the endocytic machinery in vivo and associatewith a subset of Ub-binding clathrin adaptors in vitro.Genetic and pharmacological approaches show thatinternalization of plasma membrane proteins harbouringmultiple Ub moieties is clathrin-dependent, but caveolin-independent. Functional assays demonstrate the cargo-dependent involvement of eps15/15R and epsin, UIMcontaining clathrin adaptors, in the endocytosis ofmodel proteins, CD4 and the activated b2-adrenergicreceptor complex, containing polymeric or oligomericUb. These results provide a paradigm for the clathrin-mediated uptake of ubiquitinated membrane proteinsin mammalian cells, requiring the assembly of multipleUIM–Ub interactions to overcome the low affinity bind-ing of mono-Ub to UIM.
Key words: caveolin, CD4, clathrin, endocytosis, eps15,epsin, UIM, b2-adrenergic receptor, ubiquitin
Received 29 September 2005, revised and accepted forpublication 29 November 2005, published on-line 12January 2006
Post-translational attachment of ubiquitin (Ub) to polypep-
tides has a fundamental role in modulating the plasma
membrane protein composition. Polyubiquitination prevents
the delivery of non-native membrane proteins by signalling
their proteasome-dependent degradation at the endoplas-
mic reticulum (ER) or shunting them for vacuolar/lysoso-
mal proteolysis from post-ER compartments (1,2). While
ubiquitination serves as the primary endocytic signal for
the down-regulation of plasma membrane proteins in
yeast (3,4,5), a variety of short peptide motifs [e.g.
YXXF, where F is a bulky hydrophobic residue and (D/
E)XXXL(L/I)] have been established as internalization motifs
in mammalian cells (1). Recently, monoubiquitination also
emerged as a recognition signal in the retrieval of plasma
membrane receptors and transporters from the cell sur-
face in higher eukaryotes (3,4,8–10). The notion that
mono-Ub represents an autonomous endocytic signal in
mammalian cells was primarily based on the internalization
activity of fusion proteins, containing a Ub moiety and the
truncated epidermal growth factor (EGF), platelet-derived
growth factor (PDGF) and the IL-2 receptor (5,8,10).
Considering that these receptors have intrinsic homo-
and/or hetero-oligomerization propensity, as well as the
fused Ub moiety may have been subjected to cycles of
ubiquitination-deubiquitination at the plasma membrane/
endosomes (11), the presentation of oligomeric Ub
adduct(s) to the endocytic machinery cannot be ruled out.
Molecular recognition of Ub can be achieved by Ub-
receptors that contain Ub-binding module(s), such as Ub-
interacting motif (UIM), Ub associated, coupling of Ub to
endoplasmic reticulum associated degradation, Ub E2 var-
iant and Npl4 Zn finger domains (3,12). It is assumed that
decoding of mono-Ub as an endocytic signal is mediated
by modular adaptor molecules (e.g. epsin and eps15/
eps15R) (4,13,14), harbouring multiple UIMs and other
functional domains that trigger coat proteins assembly
and subsequent vesicular budding similar to cargo sorting
at the trans Golgi network and early endosomes (6,15,16).
Intriguingly, while the preferential association of poly Ub
chain to the conserved UIM of the S5a subunit of protea-
some correlates well with the proteasome-mediated
degradation of polyubiquitinated targets, the negligible
binding of mono-Ub to the UIMs of eps15 and epsin is at
variance with their presumed role in monoubiquitinated
cargo recruitment to clathrin-coated pits/vesicles (CCP/
CCV) (13,17–19).
Multiple internalization pathways have been invoked in the
uptake of ubiquitinated proteins from the cell surface.
While several plasma membrane proteins that undergo
oligo/polyubiquitination-induced down-regulation (e.g.
Traffic 2006; 7: 282–297# 2006 The Authors
Blackwell Munksgaard doi: 10.1111/j.1600-0854.2006.00384.x
282

CD4, major histocompatibility complex (MHC) class I,
b2-adrenergic receptor (b2-AR) and ENaC) are internalized
via the CCP and CCV, activated EGF receptor (EGFR) and
PDGF receptor (PDGFR) appear to be targeted for endo-
cytosis via caveolin- and clathrin-dependent mechanisms
(4,20–22). These observations, jointly, indicate that neither
the structural basis nor the molecular machinery respon-
sible for the recognition and internalization of ubiquitinated
plasma membrane proteins is fully understood in mamma-
lian cells (9,23).
To gain more insight into this process, we utilized model
proteins harbouring Ub as their primary endocytic signal.
Using a panel of CD4 chimeras, as well as other ubiquiti-
nated membrane proteins, in combination with functional,
biochemical and genetic techniques, we present data sug-
gesting that polymeric or oligomeric Ub is recognized as
an endocytic signal by a subset of UIM containing clathrin
adaptor molecules in higher eukaryotes.
Results
Chimeric membrane proteins to study the endocytic
activity of Ub
To examine the role of Ub as an endocytic signal, first we
used CD4, a monomeric type-I membrane protein, as a
reporter molecule. CD4 is targeted to the plasma mem-
brane by default and has been extensively used to deline-
ate sorting signals, and its truncated form lacks signalling
ability (1). Ub recognition by the endocytic sorting machin-
ery was optimized by creating linkers with various lengths
between the CD4 reporter molecule and the fused Ub
(Figure 1A). The cytoplasmic tail (43 amino acid residues)
Ii0
10
20
30
40
50
IiT IiT-Ub
% In
tern
aliz
ed
CD4T
Ub
CD4Tl
Ub Ub V26G
0
10
20
30
40
CD4
Ub–
mockCD4Tlwt AllR ∆GG
α-actin
– Ub
α-CD4
– –
CD4Tl CD4Tl-Ub CD4Tl-UbAllR
% In
tern
aliz
ed
NCD4
CD4T-Ub
CD4Tl-Ub
CD4-Ub
Ii
IiT-Ub
TMA D
E
B
C
C
CD4TCC-Ub
C N
CD4T
CD4Tl
CD4
IiT
53 kDa
Figure 1: Ubiquitin (Ub) fusion confers internalization activity to type-I and type-II plasma membrane proteins. (A) Schematic
representations of model proteins containing Ub fused to CD4 or invariant chain (Ii). The cytoplasmic tail of CD4T, CD4Tl and CD4 is drawn
to scale. The Ub linker (black line, 21 amino acid residues) and the additional spacer (orange line) are indicated and specified in the
Materials and Methods. To preserve the distance between Ub and the plasma membrane, we replaced the flexible linker by the coiled-
coiled tetramerization domain (CC) of the Kir6.2 channel (small circles) in CD4TCC-Ub. (B) Plasma membrane expression of CD4 chimeras.
The chimeras were visualized by indirect immunostaining and confocal fluorescence microscopy using anti-CD4 antibody in transiently
transfected, non-permeabilized COS-7 cells. Bar: 10 mm. (C) Immunoblot analysis of CD4Tl, CD4Tl-Ub, CD4Tl-UbAllR and CD4Tl-UbnGG
expression in transiently transfected COS-7 cells. Equal amounts of cell lysates were separated by SDS-PAGE and visualized by enhanced
chemiluminescence (ECL) assay with polyclonal anti-CD4 antibody. Actin was visualized as loading control. Black triangle, CD4Tl; grey
triangle, CD4Tl-Ub variants. (D) Ub fusion confers internalization signal to CD4 variants. Endocytosis rates of the indicated chimeras were
measured by the antibody-uptake assay in transiently transfected COS-7 cells at 37 �C for 5 min as described in Materials and Methods.
Internalization rates are expressed as percentage of CD4 antibody uptake in 5 min relative to the initial amount at the cell surface
(mean � SE, n ¼ 3–4). Experiments were performed in triplicates. Separate experiments verified that antibody uptake is linear for 5 min.
(E) Ub fusion confers internalization signal to the truncated IiT. Internalization rates of the indicated construct were measured by the anti-Ii
antibody uptake as described in Materials and Methods (mean � SEM, n ¼ 3–4).
Oligoubiquitin Recognition as Endocytic Signal
Traffic 2006; 7: 282–297 283

of CD4 was preserved in the CD4-Ub chimera, while it
was truncated at the membrane plane and replaced with a
flexible 20 or 40 amino acid linker in CD4T-Ub and CD4Tl-
Ub, respectively (Figure 1A) as described in Materials and
Methods. Immunostaining and immunoblotting demon-
strated the expression of CD4Tl, CD4T and CD4, as well
as their Ub fusions at plasma membrane and in the lysates
of COS-7 cells (Figure 1B,C and data not shown).
The endocytosis of chimeras was measured by anti-CD4
antibody uptake assays in transiently transfected COS-7
cells. After binding the anti-CD4 antibody at 4 �C, internal-ization was resumed at 37 �C for 3–5 min, and the amount
of cell surface-remaining antibody was quantified by radio-
active or fluorescence assay as described in Materials and
Methods. Ub fusion accelerated the internalization of CD4Tl
from 9.6 � 0.9 to 41.9 � 2.6% in 5 min, while it was less
effective upon fusion to the CD4 and CD4T, monitored by
anti-CD4 antibody uptake (Figure 1D). The endocytic rates
of CD4Tl-Ub were similar in CHO, HeLa and HEK293 cells
(data not shown). Inducing Ub unfolding by replacing Val26
with Gly (CD4Tl-UbV26G) abolished the internalization of the
chimera (Figure 1D), implying that the native conformation
of Ub is a prerequisite for its recognition by the endocytic
machinery, as observed in yeast (5).
Ub was also recognized as an endocytic signal in type II
membrane protein (24). Attachment of Ub to the truncated
invariant chain (IiT), a homo-trimeric type-II membrane
protein lacking its di-Leu-based endocytic signal, increased
the internalization rate of IiT (6.9 � 2.0%/5 min) by seven-
fold (IiT-Ub, 49.9 � 4.2%/5 min) (Figure 1E).
Mono-Ub is not recognized efficiently by the
endocytic machinery as an internalization signal
in mammalian cells
To assess whether a single Ub moiety constitutes an
autonomous internalization signal, we prevented Ub-
chain synthesis by replacing all the seven Lys residues
with Arg (UbAllR) or deleting the carboxy-terminal Gly
residues (UbnGG) in Ub (25). While CD4Tl-UbnGG
uptake was similar to the CD4Tl-Ub, the internalization of
both CD4Tl-UbAllR (10.0 � 2.1%/5 min) and IiT-UbAllR
(8.6 � 0.5%/5 min) was reduced to the level of the
respective reporter molecule (CD4Tl, 9.6 � 0.9%/5 min;
and IiT, 6.9 � 2.0%/5 min) (Figure 2A). Although the
steady-state expression of the CD4Tl and CD4Tl-UbAllR
was fourfold to fivefold higher, saturation of the internal-
ization machinery can account for their impeded uptake.
Reducing the cell surface density of CD4Tl by fourfold did
not influence its endocytosis rates (data not shown). The
slow signal-independent endocytosis of CD4Tl and CD4Tl-
UbAllR is likely mediated by bulk membrane flow (26) and
was comparable to that observed for CD4Tl in COS-7 and
CHO cells (Figure 2A,2H). These observations suggest
that monomeric Ub cannot be recognized as an autono-
mous endocytic signal in mammalian cells, a conclusion
that was reinforced by subsequent results.
Next, we tested whether reinsertion of a single Lys into the
UbAllR is sufficient to render endocytic activity to the CD4Tl-
UbAllR chimera. Reverting Arg48 into Lys residue comple-
tely restored the CD4Tl-UbAllR internalization rates to the
CD4Tl-Ub level, likely by providing an acceptor site for Ub
conjugation (Figure 2B). The endocytic activity was partially
recovered by Lys63, but not by Lys6 or Lys29 reinsertion
into UbAllR (Figure 2B). In line with the redundancy of the
branching site for Ub chain elongation, replacement of Lys48
with Arg in wild-type Ub caused only partial inhibition of the
CD4Tl-UbK48R internalization (approximately 37%), while
neither Lys63Arg, nor Lys29Arg mutations had an inhibitory
effect (Figure 2B). These observations suggest that Lys48 is
preferential, but not the exclusive acceptor site for the first
isopeptide bond formation in CD4Tl-UbAllR. Thus incorpor-
ation of UbK48R in fusion proteins does not preclude the
possibility of Ub conjugation. Tandem fusion of two Ubs
(CD4Tl-2Ub) or three Ubs (CD4Tl-3Ub) significantly acceler-
ated the initial uptake rates compared with CD4Tl-Ub, sup-
porting the notion that Ub multimerization facilitates cargo
uptake (Figure 2C). In the latter measurements, the inter-
nalization time was reduced from 5 to 3 min to assess the
rapid initial rates of endocytosis.
Biochemical evidence for the ubiquitination of CD4Tl-Ub
was obtained by determining the apparent molecular
mass and Ub conjugation of CD4Tl-Ub. Cell surface resi-
dent and endocytosed reporter molecules were selec-
tively immunoprecipitated following in vivo labelling the
cells with anti-CD4 antibody to preclude the isolation of
ubiquitinated adducts from the ER. Immunoprecipitates
were isolated by protein G sepharose and probed with
anti-CD4 antibody. While CD4Tl was represented by an
approximately 52 kDa polypeptide corresponding to its
predicted mass (Figure 2D, lane 3), CD4Tl-Ub appeared
as a doublet with molecular masses of approximately
58 kDa, representing CD4Tl-Ub and with MW of approxi-
mately 65–66 kDa, likely reflecting monoubiquitinated
adduct of CD4Tl-Ub (Figure 2D, lane 4). Both of the
mono- and di-Ub containing CD4Tl-Ub polypeptides were
recognized by anti-Ub antibody (Figure 2D, lower panel). In
addition, polyubiquitination of CD4Tl-Ub, but not CD4Tl or
CD4Tl-UbAllRnGG, could be observed, indicated by the
high molecular weight ladder formation in the precipitates
(Figure 2D, lane 4, lower panel and data not shown).
Polymeric Ub chain and multimers of mono-Ub serve
as endocytic signals
If mono-Ub cannot be recognized by the endocytic
machinery, non-covalently linked multimers of UbAllR
representing multiple monoubiquitination (8) or oligo-
merization of monoubiquitinated native cargo molecules
(27) may serve as an efficient internalization signal.
Tetramerization of CD4Tl-UbAllR was induced by inserting
the tetramerization coiled-coil (CC) domain of the Kir6.2
potassium channel between the membrane-spanning seg-
ment of CD4 and the UbAllR moiety (CD4TCC-UbAllR
Figure 1A) (28). The ability of the CC domain to induce
Barriere et al.
284 Traffic 2006; 7: 282–297
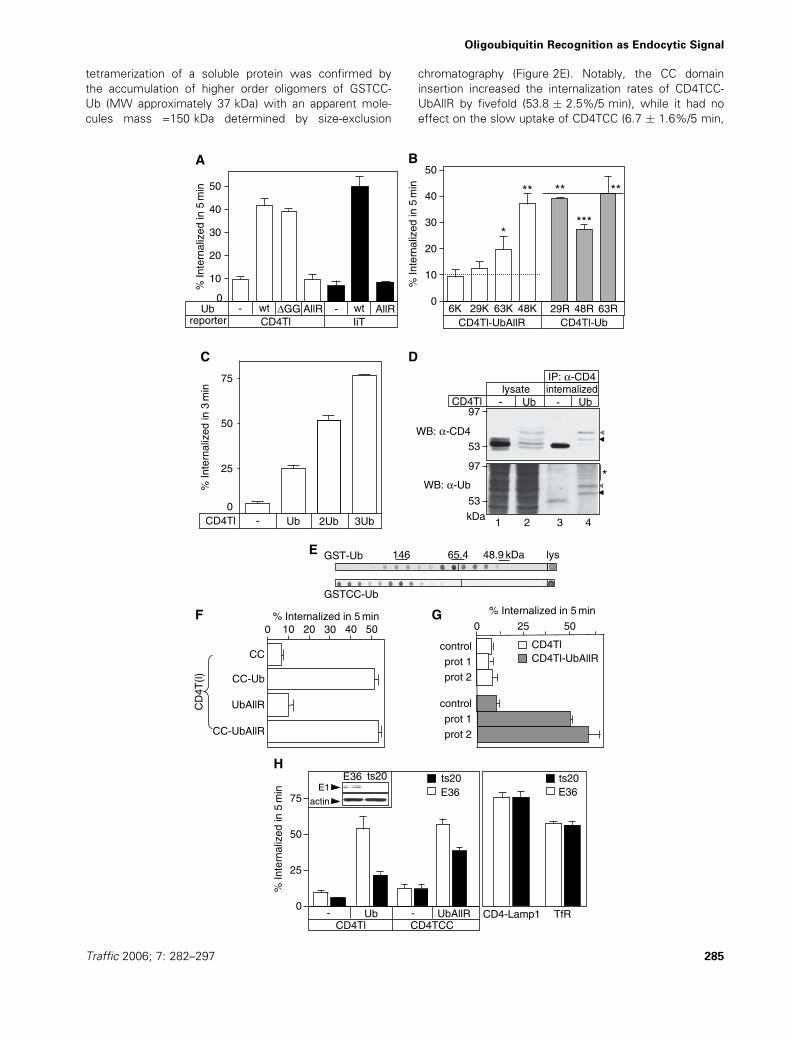
tetramerization of a soluble protein was confirmed by
the accumulation of higher order oligomers of GSTCC-
Ub (MW approximately 37 kDa) with an apparent mole-
cules mass =150 kDa determined by size-exclusion
chromatography (Figure 2E). Notably, the CC domain
insertion increased the internalization rates of CD4TCC-
UbAllR by fivefold (53.8 � 2.5%/5 min), while it had no
effect on the slow uptake of CD4TCC (6.7 � 1.6%/5 min,
B
D
E
F G
H
A
C
- wt ∆GG AllR wtIiT
AllR0
10
40
50
20
30
CD4TlreporterUb -
0
10
40
50
20
30*
63R48RCD4Tl-UbAllR
29RCD4Tl-Ub
***
29K 63K 48K6K
UbCD4Tl 2Ub 3Ub-
25
75
% In
tern
aliz
ed in
3 m
in
50
0
97
53
lysate internalizedIP: α-CD4
CD4Tl - Ub - Ub97
53WB: α-CD4
WB: α-Ub*
kDa1 2 3 4
actin
ts20E1
E36
CD4TlUb CD4-Lamp1
0
25
50
75
ts20E36
ts20E36
CD4TCCUbAllR- - TfR
65.4 48.9 kDa lys
GSTCC-Ub
146GST-Ub
% Internalized in 5 min
CC
CC-Ub
UbAllR
CC-UbAllR
0 10 40 5020 30
CD
4T(l)
CD4TlCD4Tl-UbAllR
0 25 50
control
control
% Internalized in 5 min
% In
tern
aliz
ed in
5 m
in
% In
tern
aliz
ed in
5 m
in ** ****
prot 1
prot 2
prot 1
prot 2
% In
tern
aliz
ed in
5 m
in
Oligoubiquitin Recognition as Endocytic Signal
Traffic 2006; 7: 282–297 285

Figure 2F). As a complementary approach to induce
CD4Tl-UbAllR oligomerization, antibody-mediated cross-
linking was applied. Cell surface bound anti-CD4 antibody
was crosslinked by secondary antibody at 4 �C, and inter-
nalization was measured by two different protocols. Non-
covalent crosslinking increased the internalization of
CD4Tl-UbAllR, but not CD4Tl, by fivefold at 37 �C(Figure 2G). These results not only rule out that conforma-
tional defect of UbAllR accounts for the defective endocy-
tosis of CD4Tl-UbAllR but, more importantly, demonstrate
that the internalization machinery can recognize Ub multi-
mers in covalently linked Ub chain, as well as in oligomeric
complexes.
If Ub-chain synthesis is required for CD4Tl-Ub endocyto-
sis, inactivation of the cellular ubiquitination machinery
should impede this process. Down-regulation of the
thermo-sensitive E1 Ub-activating enzyme was achieved
by exposing the transiently transfected ts20 cells to 40 �C(29). Inactivation of the E1 enzyme inhibited CD4Tl-Ub
endocytosis by 63%, compared with that measured in
E36 cells containing the wild-type E1 enzyme
(Figure 2H). The incomplete inhibition of endocytosis
may be explained by the residual activity of E2/E3 after
E1 inactivation. Considering that putative components of
the Ub-dependent internalization machinery are suscepti-
ble to monoubiquitination (e.g. eps15/15R and epsin)
(13,19,30), which may influence cargo recognition, we
also assessed the impact of E1 ablation on the activity of
the Ub-specific internalization machinery. To accomplish
this, we used the tetrameric CD4TCC-UbAllR as cargo,
because this chimera is not susceptible to ubiquitination
at the cell surface (Figure 2F). E1 down-regulation has
less-pronounced inhibition on the CD4TCC-UbAllR
endocytosis (43%) as compared with that on CD4Tl-Ub
uptake (63%, Figure 2H), lending credence to the hypoth-
esis that ubiquitination of the endocytic machinery and/or
the elevated cytoplasmic Ub concentration can influence
cargo recognition efficiency at the cell surface. Notably, E1
inactivation failed to alter the endocytosis of the CD4-
Lamp1 chimera and the transferrin receptor (TfR)
(Figure 2H). This suggests that recognition of the Tyr-
based endocytic motifs by the AP-2-dependent internaliza-
tion pathway is independent of the ubiquitination machin-
ery activity.
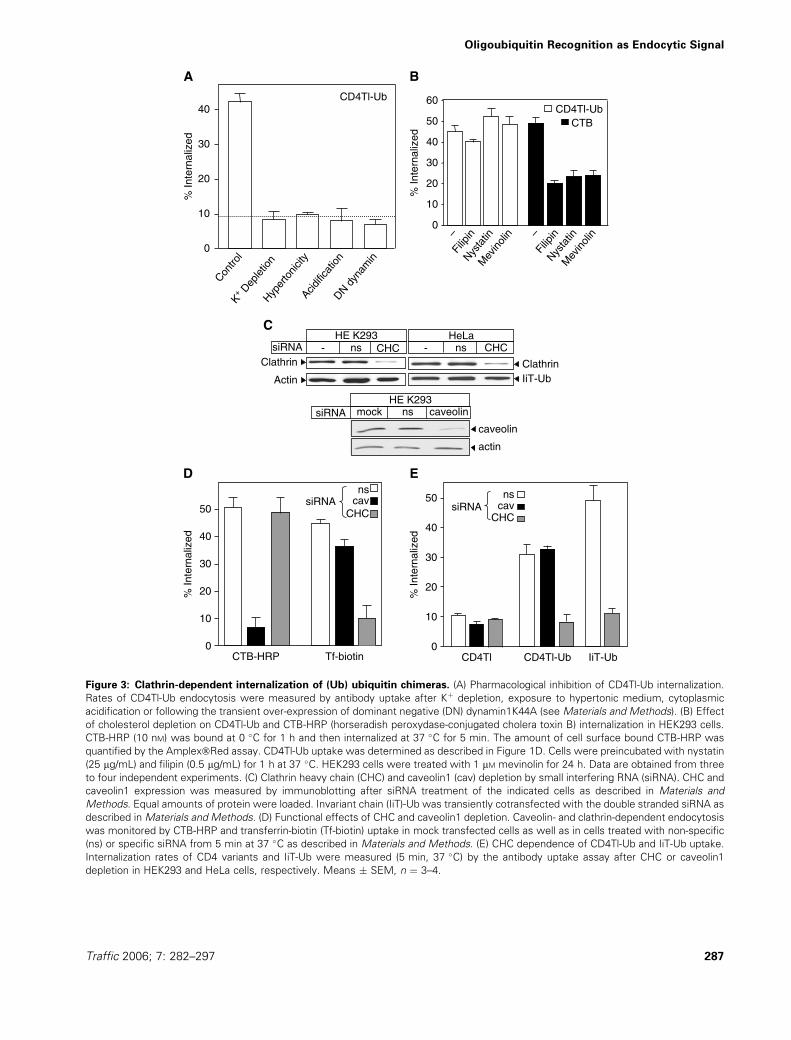
Clathrin-dependent endocytosis of ubiquitinated
chimeras
To unveil the identity of the internalization pathway of
ubiquitinated membrane proteins, we used Ub chimeras
without signalling ability. Established inhibitors of CCV
formation, such as hypertonicity, Kþ-depletion or cytoplas-
mic acidification abolished the Ub-dependent internaliza-
tion of the CD4Tl-Ub, similar to the dominant negative
dynamin1K44A overexpression (Figure 3A). The effect of
dynamin1K44A cannot be attributed to defective caveo-
some formation, because blocking caveosome-mediated
endocytosis by cholesterol depletion with filipin, nystatin
or mevinolin had no effect on CD4Tl-Ub uptake
(Figure 3B). These drugs efficiently reduced the cellular
cholesterol content, indicated by the impeded cholera
toxin B (CTB) uptake (Figure 3B), a cargo of caveolae-
mediated internalization (31). To validate these results
and rule out possible crosstalk of inhibitors, we deter-
mined CD4Tl-Ub endocytosis in clathrin heavy chain
(CHC) or caveolin1-depleted cells. Delivery of small inter-
fering RNA (siRNA) resulted in >80% loss of CHC and
caveolin1 content in HEK293 cells (Figure 3C) and caused
Figure 2: Monomeric ubiquitin (Ub) is not sufficient to signal internalization. (A) In-frame fusion of UbAllR is not recognized as an
internalization signal in type-I and -II plasma membrane proteins. Endocytosis rates of the indicated CD4 and invariant chain chimeras were
determined by the antibody uptake assay in transiently transfected COS-7 cells. (B) Oligo-ubiquitination is required for efficient internalization of the
CD4Tl-UbAllR chimera. Endocytosis rates of the indicated chimeras were determined as in panel (A) (means � SEM, n ¼ 3). For comparison,
dashed and solid lines indicate the internalization rates of CD4Tl-UbAllR and CD4Tl-Ub, respectively. The significance of change was calculated by
two-tailed unpaired t-test and indicated as follows: *, p ¼ 0.01 for R63K versus CD4Tl-UbAllR; **, p > 0.3 for R48K, K29R or K63Rmutants versus
CD4TlUb and ***, p < 0.01 for K48R mutant versus CD4Tl. (C) The internalization rates are proportional to the number of Ub moieties fused to
CD4Tl. Endocytosis of CD4Tl harbouring mono-, di- or tri-Ub moieties weremeasured in transiently transfected COS-7 cells. Antibody uptake rates
were calculated after 3 min of internalization. (D) Ubiquitination of CD4Tl-Ub. Cell surface resident and endocytosed CD4Tl and CD4Tl-Ub were
selectively labelled by incubating stably transfectedHEK293 cellswith CD4 antibody for 30 min at 37 �C. CD4-Ab complexeswere immunoisolated
on protein G. Imunoprecipitates and lysates were probed with anti-CD4 (top panel) and anti-Ub antibodies (bottom panel). CD4Tl-Ub and its
monoubiquitinated version are marked by black and grey arrow head, respectively. Asterisk denotes polyubiquitinated adducts of CD4Tl-Ub,
undetectable for CD4Tl-Ub. (E) Oligomerization tendency of the coil-coiled (CC) tetramerization domain containing GST-Ub. Bacterial extracts,
expressing GST-Ub or GSTCC-Ub were fractioned by size-exclusion chromatography. Fractions were analysed by dot-blotting using anti-GST Ab
and chemiluminescence (ECL). (F) The tetramerization domain (CC) restores the CD4Tl-UbAllR internalization activity. Endocytosis rates of
CD4TCC, CD4TCC-Ub, CD4TCC-UbAllR and CD4Tl-UbAllR were measured by antibody uptake in COS-7 cells (means � SE, n ¼ 3–4). (G)
Oligomerization of CD4Tl-UbAllR by antibody cross-linking induces internalization, measured in COS-7. Following primary and secondary antibody
binding (prot. 1) or primary, biotinylated secondary antibody and streptavidin-horseradish peroxydase (HRP) cross-linking (prot. 2) on ice, internaliza-
tion was resumed for 5 min at 37 �C. Data are expressed as percentage of uptake of the initial amount of antibody or streptavidin-HRP, measured
as described in Materials and Methods (means � SE, n ¼ 3). (H) Active ubiquitination machinery is required for efficient internalization of
ubiquitinated reportermolecules, but not for transferrin receptor (TfR) or the CD4-Lamp1 chimera. Internalization ratesweremeasured in transiently
transfected ts20 or E36 cells, after incubation at 40 �C for 2 h to down-regulate the E1 enzyme. CD4-Lamp1 and TfR uptakewas determined under
the same conditions using CD4 Ab and Tf-biotin, respectively, as detailed inMaterials andMethods. Inset: down-regulation of the thermo-sensitive
E1 enzyme was monitored by immunoblotting after 2 h of incubation at 40 �C with anti-E1 antibody and ECL.
Barriere et al.
286 Traffic 2006; 7: 282–297
0
K+ D
eple
tion
Hyper
toni
city
Acidific
atio
nDN d
ynam
in
–Fi
lipin
Nysta
tinM
evin
olin –
Filip
inNys
tatin
Mev
inol
in
Contro
l
10
20
30
% In
tern
aliz
ed
% In
tern
aliz
ed
40 CD4Tl-UbCD4Tl-Ub
CTB
0
10
20
30
40
50
60
10
20
30
% In
tern
aliz
ed
% In
tern
aliz
ed40
0
50
CTB-HRP Tf-biotin
siRNAns
cavCHC
10
20
30
0CD4Tl-UbCD4Tl
siRNAns
cavCHC
IiT-Ub
40
50
Clathrin
Actin
siRNA ns CHC ns CHC
Clathrin
A B
C
D E
IiT-Ub
- -
siRNA mock ns caveolin
actin
caveolin
HE K293 HeLa
HE K293
Figure 3: Clathrin-dependent internalization of (Ub) ubiquitin chimeras. (A) Pharmacological inhibition of CD4Tl-Ub internalization.
Rates of CD4Tl-Ub endocytosis were measured by antibody uptake after Kþ depletion, exposure to hypertonic medium, cytoplasmic
acidification or following the transient over-expression of dominant negative (DN) dynamin1K44A (see Materials and Methods). (B) Effect
of cholesterol depletion on CD4Tl-Ub and CTB-HRP (horseradish peroxydase-conjugated cholera toxin B) internalization in HEK293 cells.
CTB-HRP (10 nM) was bound at 0 �C for 1 h and then internalized at 37 �C for 5 min. The amount of cell surface bound CTB-HRP was
quantified by the Amplex�Red assay. CD4Tl-Ub uptake was determined as described in Figure 1D. Cells were preincubated with nystatin
(25 mg/mL) and filipin (0.5 mg/mL) for 1 h at 37 �C. HEK293 cells were treated with 1 mM mevinolin for 24 h. Data are obtained from three
to four independent experiments. (C) Clathrin heavy chain (CHC) and caveolin1 (cav) depletion by small interfering RNA (siRNA). CHC and
caveolin1 expression was measured by immunoblotting after siRNA treatment of the indicated cells as described in Materials and
Methods. Equal amounts of protein were loaded. Invariant chain (IiT)-Ub was transiently cotransfected with the double stranded siRNA as
described in Materials and Methods. (D) Functional effects of CHC and caveolin1 depletion. Caveolin- and clathrin-dependent endocytosis
was monitored by CTB-HRP and transferrin-biotin (Tf-biotin) uptake in mock transfected cells as well as in cells treated with non-specific
(ns) or specific siRNA from 5 min at 37 �C as described in Materials and Methods. (E) CHC dependence of CD4Tl-Ub and IiT-Ub uptake.
Internalization rates of CD4 variants and IiT-Ub were measured (5 min, 37 �C) by the antibody uptake assay after CHC or caveolin1
depletion in HEK293 and HeLa cells, respectively. Means � SEM, n ¼ 3–4.
Oligoubiquitin Recognition as Endocytic Signal
Traffic 2006; 7: 282–297 287

near complete elimination of TfR and CTB uptake, respec-
tively (Figure 3D). Importantly, down-regulation of CHC,
but not caveolin1, suppressed CD4Tl-Ub endocytosis to
basal level (Figure 3E). Similar results were obtained with
the IiT-Ub chimera (Figure 3E).
Ubiquitinated reporter was excluded from detergent-resistant
membranes, because CD4Tl-Ub (and CD4Tl), but not CTB
binding to ganglioside 1 (GM1) was eliminated by TritonX-
100 extraction at 4 �C, visualized by fluorescencemicroscopy
(Figure 4A,B). Immunostaining confirmed that CD4Tl-Ub was
excluded from plasma membrane rafts, while it was partially
co-localized with the TfR (Figure 4C,D). These data, collec-
tively, reinforce the notion that ubiquitinated cargoes without
signalling ability are excluded from cholesterol-rich rafts and
preferentially targeted for clathrin-dependent endocytosis.
Biochemical interaction of clathrin adaptors with
ubiquitinated cargo
It has been postulated that eps15, eps15R and/or epsin
may play critical role in the recruitment of ubiquitinated
cargo into CCP and CCV because these adaptors (i)
harbour multiple UIMs (1); (ii) bind directly or indirectly to
CHC and AP-2 (16); (iii) have been localised to CCP and
CCV by electron microscopy (22,32) and (iv) their ortholo-
gues are implicated in the down-regulation of ubiquitinated
plasma membrane proteins in Drosophila melanogaster
(33) and yeast (14). Biochemical association of potential
clathrin adaptors, involved in the recognition of ubiquiti-
nated plasma membrane proteins, was examined next.
Binding of eps15, eps15R and epsin1 to GST-Ub fusion pro-
teins was measured by pull-down assays after incubating the
recombinant proteinswith HeLa cytosol. Although these adap-
tors were unable to bind to GST-Ub (and GST), significant
fraction of the lysate was associated with GST-2Ub and GST-
3Ub (Figure 5A). Misfolding of GST-Ub cannot account for
its inability to bind to the adaptor proteins, because Hrs
(34) and the E1 Ub-activating enzyme were bound to
mono-Ub (data not shown). In light of the dimerization
propensity of GST (Figure 2E), it is plausible to assume
that presentation of oligomeric Ub represents the efficient
binding partner for these adaptors by increasing their avid-
ity to Ub containing complex. This notion was further
supported by the inability of recombinant eps15 to bind
to mono-, di- or tri-Ub, but efficiently associate with oligo-
Ub, containing four or more Ub residues conjugated via
K63 or K48 (Figure 5B). These observations, together with
the preferential poly Ub binding of UIMs of eps15 and
epsin1 (13,18,19,35), are consistent with our functional
data, and the hypothesis that multivalent substrate – adap-
tor interactions are necessary to overcome the low affinity
binding of mono-Ub to various UIM domains (Kd approxi-
mately 0.1–1 mM) (17,36,37). The AP-2 association with
GST-2Ub and GST-3Ub is, presumable, indirect and
mediated by eps15/15R or epsin1 (Figure 5A), because
the AP-2 complex has eps15 and epsin1-binding sites,
but lacks Ub-binding domain (38).
Association of eps15 with the Ub chimera was demon-
strated by coimmunoprecipitation of eps15 with CD4Tl-
3Ub, but not with CD4Tl in transiently transfected COS-7
cells (Figure 5C and data not shown). We were unable to
detect eps15 binding to CD4Tl-Ub. Combination of rapid
deubiquitination, decreased binding affinity of adaptors to
and the low expression level of the oligo-ubiquitinated
CD4Tl-Ub may explain the lack of eps15 detection in
CD4Tl-Ub precipitates.
Functional relevance of eps15/15R and epsin1 in the
endocytosis of ubiquitinated cargo
The functional relevance of eps15, eps15R, epsin1 and
AP-2a in ubiquitinated cargo endocytosis was evaluated
after depletion of these adaptors by siRNA. Immunoblot
analysis revealed that the UIM adaptors were reduced by
73–76%, while AP-2a was depleted by approximately
84% in HEK293 cells (Figure 6A,B). While eps15,
eps15R and epsin1 depletion caused 73, 84 and 73%
inhibition of the Ub-dependent internalizaion of CD4Tl-
Ub, respectively (Figure 6C), TfR endocytosis was dimin-
ished by approximately 55% (Figure 6D). Inhibition of TfR
uptake is consistent with the role of these adaptors as
accessory proteins in CCV formation (39,40). However, a
more dramatic difference was observed in the sensitivity
to AP-2a depletion. While AP-2a siRNA treatment had only
marginally inhibited (approximately 11%) CD4Tl-Ub uptake
(Figure 6C), TfR endocytosis was attenuated by approxi-
mately 75% (Figure 6D). Notably, internalization of the
oligomeric CD4TCC-UbAllR was attenuated comparable
to that of CD4Tl-Ub in CHC, eps15 and epsin knockdown
cells (Figure 6E), implying that an overlapping set of UIM
containing endocytic adaptors can recognize cargo mole-
cules tagged with oligo-Ub chain(s) or oligomeric Ub.
These results also suggest that ubiquitinated plasma
membrane proteins may represent a diverse group of
ligands that can be recruited via clathrin-dependent, but
AP-2-independent internalization mechanism, similar to
the LDL-R family (16). Although GGA3, a Golgi/endosome
adaptor, contributes to the sorting of ubiquitinated cargo
at Golgi and endosomal compartments (15), GGA3
appears to have no role in the recognition of the Ub
chimeras at the plasma membrane in HEK293 cells.
The established biochemical interaction network of eps15/
15R, epsin and AP-2 adaptors suggest that ubiquitinated
cargo recruitment to CHC may proceed by eps15/eps15R
and AP-2, or epsin1 in cargo and cell type-dependent
manner (5,9,23,41) (Figure 7). In line with this hypothesis,
AP-2a and epsin depletion together inhibited CD4Tl-Ub
uptake by approximately 90%, while their individual ablation
reduced cargo uptake by approximately 11 and 73%, respec-
tively (Figure 6C). Furthermore, epsin1 and eps15 siRNA
combination inhibited CD4Tl-Ub internalization by approxi-
mately 90% (data not shown), suggesting that they have
partially overlapping function in ubiquitinated cargo uptake
(23,39). This phenomenon is reminiscent of the functional
co-operativity of the yeast eps15 and epsin orthologues
Barriere et al.
288 Traffic 2006; 7: 282–297

CTB CD4Tl-Ub
Con
trol
Ext
ract
edC
ontr
olE
xtra
cted
Overlay
CTB CD4Tl OverlayA
B
C
D
CD4Tl CTB
CD4Tl-Ub CTB
Overlay
Overlay
TfR
CD4Tl-Ub
CD4Tl Overlay
TfR Overlay
Figure 4: CD4Tl-ubiquitin (Ub) is excluded
from cholesterol-rich rafts. (A–B) Detergent
extractability of (A) CD4Tl and (B) CD4Tl-Ub.
COS-7 cells were transiently transfected with
CD4Tl or CD4Tl-Ub. Cells were stained with
cholera toxin B (CTB)-FITC, monoclonal anti-
CD4 and TRITC-conjugated anti-mouse antibo-
dies. Extraction of non-raft associated mem-
brane proteins was performed in 1% TritonX-
100 (0 �C, 15 min) before fixation. Non-treated
cells were used as controls. Optical sections of
representative cells were obtained by laser con-
focal fluorescence microscopy (LCFM) and pro-
jected images are shown. Note that the laser
light intensity was increased by 10-fold to visua-
lize the small amount of CD4 variants remaining
after TritonX-100 extraction. Bar: 10 mm. (C)
CD4Tl-Ub does not co-localize with raft micro-
domains, enriched in GM1. COS-7 cells were
transfected with CD4Tl or CD4Tl-Ub and
patched with mouse anti-CD4 antibody fol-
lowed by anti-mouse TRITC-conjugated anti-
body as described in Materials and Methods.
GM1 containing lipid rafts were labelled with
CTB-FITC. Boxed areas are enlarged and
depicted in the right column. (D) Partial co-loca-
lization of CD4Tl-Ub and transferrin receptor
(TfR). HEK293 cells were transiently trans-
fected with CD4Tl or CD4Tl-Ub. Cells, plated
on poly Lys-coated coverslips, were incubated
with sheep anti-CD4 and mouse anti-Tf antibo-
dies for 1 h at 12 �C, followed by incubation
with the relevant FITC- and TRITC-conjugated
secondary antibody under the same conditions
and imaged by LCFM.
Oligoubiquitin Recognition as Endocytic Signal
Traffic 2006; 7: 282–297 289

(Ede1p and Ent1/2p) (14). Complete restoration of the ubiqui-
tinated a-factor receptor endocytosis in the background of
ent1D/ent2D/ede1D strain could be only achieved upon com-
plementation with both plasmid-borne Ent1p and Ede1 (14).
Because epsin as well as eps15/15R are susceptible
to monoubiquitination and oligomerization in vivo
(13,19,30,42), multiple UIM and Ub interactions may
stabilize the Ub-adaptor network. While the precise
mechanism of this process remains to be discovered, it
is plausible to assume that cargo binding itself may alter
the avidity of UIM–Ub interactions. Inactivation of the E1
Ub-activating enzyme in ts20 cells may interfere with the
formation of the UIM-Ub adaptor network and therefore
contributes to the observed inhibition of ubiquitinated
cargo uptake (Figure 2H).
To demonstrate the functional relevance of UIM contain-
ing adaptors in the endocytosis of Ub chimeras, we
mutated the UIM-binding interface of Ub. These experi-
ments were performed on CD4TCC-UbAllR to avoid the
incorporation of wt Ub into the oligo-Ub chain of CD4Tl-
Ub. Based on structural studies and mutagenesis, the
interface of the UIM-Ub complexes is largely formed by
the hydrophobic residues of 44Ile, 8Leu and 70Val of Ub
(17,37). Replacement of 44Ile (UbAllR-1A) or 8Leu and
70Val (UbAllR-2A) with Ala inhibited CD4TCC-UbAllR
uptake by approximately 74% (Figure 8A), implying an
essential role of Ub–UIM interactions in ubiquitinated
cargo endocytosis. Notably, the Ile44Ala mutation similarly
prevented the antibody-induced uptake of oligomeric
CD4Tl-Ile44AlaUbAllR (data not shown), while it had a mod-
est effect on CD4Tl-Ub internalization (Figure 8A). The latter
observation is consistent with oligo-Ub chain formation in
vivo, incorporating wt Ub moieties readily recognizable by
the relevant endocytic adaptors. Combining the three point
mutations (UbAllR-3A) had no additive effect, suggesting
that residual binding of UIM adaptors and/or alternative
adaptors may be also involved in the internalization process
(Figure 8A).
The role of epsin1 and eps15/15R in clathrin-dependent
internalization of ubiquitinated CD4 and b2-AR
The physiological relevance of Ub-binding adaptors was
validated by establishing their role in the internalization of
ubiquitinated b2-AR/arrestin complex. Because clathrin-
95
51
197
11695
WB:α-CD4
WB:α-eps15
lysate IP: α-CD4
CD4Tl-3Ub– + +
kDa
CD4Tl+– ––
+––
IB: α-Ub200
513729
20
7
poly-Ub (K48)
poly-Ub (K63)
mono-Ub
–
–
–
–
–
–
– – –
–
–
–
–
–
–
–
–
–
+
+
+
+
+
+
+
+
+
GST GST-eps15
Ub
Ub2
Ub3
Ub4
Ub5
Ub6
Ub7
96
kDa
20011596
51
37
29
GSTGST-eps15
α-eps15R
Ponceau
α-AP2
α-GGA3
α-eps15
120100
100120
55
3829
201
A B
C
120
GST – Ub 2Ub 3Ub lys
100 α-epsin
kDa
100120
input
kDa
Figure 5: Biochemical interactions of endocytic adaptors with ubiquitin (Ub) and ubiquitinated cargo. (A) Endocytic adaptor
binding to Ub in vitro. Pull-down assays were performed with HeLa cell extracts and the indicated GST fusion protein containing none,
one, two, or three tandem Ub moieties. Bound polypeptides and cell lysates (Lys) were immunoblotted for eps15, eps15R, epsin1, AP-2aand GGA3. (B) Poly Ub-binding specificity of eps15. GST and GST-eps15 were purified on glutathione beads and visulized by SDS-PAGE
and SimplyBlue Safe Stain (lower panel). To measure Ub binding to eps15, purified GST and GST-eps15 was prebound to glutathione
beads and incubated with K48 or K63 poly Ub chains and bovine mono-Ub. Bound Ub was visualized by immunoblotting with the FK2 anti-
Ub antibody and chemiluminescence (upper panel). Note that tetra-Ub binding to eps15 was revealed by long exposure (not shown). (C)
CD4Tl-3Ub binds to eps15 in vivo. CD4Tl and CD4Tl-3Ub were immunoprecipitated from transiently transfected COS-7 cells. The lysates
and the precipitates were probed with CD4 and eps15-specific antibodies.
Barriere et al.
290 Traffic 2006; 7: 282–297
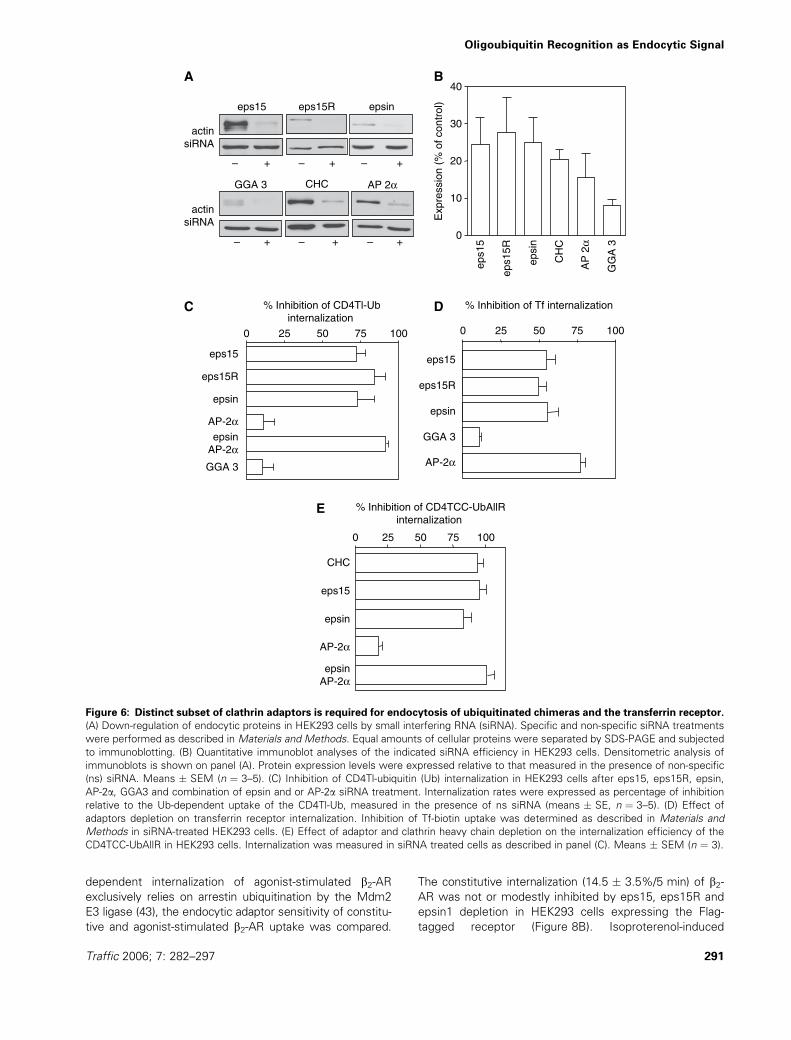
dependent internalization of agonist-stimulated b2-ARexclusively relies on arrestin ubiquitination by the Mdm2
E3 ligase (43), the endocytic adaptor sensitivity of constitu-
tive and agonist-stimulated b2-AR uptake was compared.
The constitutive internalization (14.5 � 3.5%/5 min) of b2-AR was not or modestly inhibited by eps15, eps15R and
epsin1 depletion in HEK293 cells expressing the Flag-
tagged receptor (Figure 8B). Isoproterenol-induced
– + – + – +
– + – + – +0
10
20
30
40
epsi
n
eps1
5
eps1
5R
GG
A 3
CH
C
AP 2α
AP
2α
epsin
eps15
eps15R
GGA 3
AP-2αepsin
AP-2α
AP-2α
epsinAP-2α
epsin
eps15
eps15R
GGA 3
AP-2α
GGA 3 CHC
eps15
A B
C D
E
eps15R epsin
actinsiRNA
actinsiRNA
0 25 50 75 100
% Inhibition of CD4TCC-UbAllRinternalization
eps15
epsin
CHC
0 25 50 75 100
0 25 50 75 100
% Inhibition of CD4Tl-Ubinternalization
% Inhibition of Tf internalization
Exp
ress
ion
(% o
f con
trol
)
Figure 6: Distinct subset of clathrin adaptors is required for endocytosis of ubiquitinated chimeras and the transferrin receptor.
(A) Down-regulation of endocytic proteins in HEK293 cells by small interfering RNA (siRNA). Specific and non-specific siRNA treatments
were performed as described in Materials and Methods. Equal amounts of cellular proteins were separated by SDS-PAGE and subjected
to immunoblotting. (B) Quantitative immunoblot analyses of the indicated siRNA efficiency in HEK293 cells. Densitometric analysis of
immunoblots is shown on panel (A). Protein expression levels were expressed relative to that measured in the presence of non-specific
(ns) siRNA. Means � SEM (n ¼ 3–5). (C) Inhibition of CD4Tl-ubiquitin (Ub) internalization in HEK293 cells after eps15, eps15R, epsin,
AP-2a, GGA3 and combination of epsin and or AP-2a siRNA treatment. Internalization rates were expressed as percentage of inhibition
relative to the Ub-dependent uptake of the CD4Tl-Ub, measured in the presence of ns siRNA (means � SE, n ¼ 3–5). (D) Effect of
adaptors depletion on transferrin receptor internalization. Inhibition of Tf-biotin uptake was determined as described in Materials and
Methods in siRNA-treated HEK293 cells. (E) Effect of adaptor and clathrin heavy chain depletion on the internalization efficiency of the
CD4TCC-UbAllR in HEK293 cells. Internalization was measured in siRNA treated cells as described in panel (C). Means � SEM (n ¼ 3).
Oligoubiquitin Recognition as Endocytic Signal
Traffic 2006; 7: 282–297 291

ubiquitination stimulated the internalization rate of the
b2-AR complex by threefold (42.5 � 2.5%/5 min, data
not shown) (44). The activated receptor endocytosis was
highly sensitive to eps15, eps15R and epsin depletion,
confirming their function as clathrin adaptors in polyubiqui-
tinated b2-AR complex uptake (Figure 8B).
As a second model to demonstrate the physiological rele-
vance of Ub-binding adaptors, we took advantage of the
Ub-dependent down-regulation of cell surface CD4 mole-
cules upon over-expression of viral membrane-anchored
E3 ligases (e.g. M153R) (45,46), or their human ortholo-
gues (e.g. March-IV) (47). Transient co-transfection of
March-IV, a Golgi-associated E3 enzyme (47), with CD4,
but not with CD4-lacking Lys residues in its cytoplasmic
tail, provoked approximately threefold-accelerated interna-
lization (Figure 8C). Co-expression of March-VIII, an E3
ligase, localized to the endosomal and lysosomal compart-
ments (47), had no effect on CD4Tl-Ub endocytosis.
Thus March-IV-mediated ubiquitination is necessary and
sufficient for rapid down-regulation of CD4 from the cell
surface (Figure 8C) (47). The constitutive and Ub-indepen-
dent internalization of CD4 was modestly sensitive to
eps15 and epsin depletion by siRNA (Figure 8D). In con-
trast, March-IV-induced CD4 endocytosis displayed similar
sensitivity to UIM containing adaptor knockdown as
observed for CD4Tl-Ub (Figure 6C). The internalization of
ubiquitinated CD4 was inhibited by 75–87% by eps15 and
epsin1 siRNA, while AP-2a depletion had only 50% block-
ing effect (Figure 8D). These observations with the
approximately 75% inhibition of CD4 internalization by
CHC siRNA (Figure 8D) provide supporting evidence for
the functional relevance of eps15/15R and epsin1 in the
down-regulation of oligo/polyubiquitinated plasma mem-
brane proteins via the clathrin-dependent pathway.
Discussion
Our results highlight three fundamental hallmarks of Ub as
an endocytic signal in mammalian cells. First, using RNA
interference in combination with internalization measure-
ments, we provide evidence for the involvement of the
UIM containing eps15/eps15R and epsin1 adaptors in the
recruitment of ubiquitinated cargo into CCV. The import-
ance of Ub recognition by UIM was demonstrated by
mutagenesis of the UIM-docking surface of the Ub moiety
(Figure 8A). These results, jointly with the observation that
AP-2a depletion caused negligible inhibition of the clathrin-
dependent CD4Tl-Ub uptake but approximately 75% block
of TfR endocytosis, strongly support the proposition that
eps15/15R and epsin1 are functional as clathrin-associated-
sorting proteins for ubiquitinated plasma membrane pro-
teins in higher eukaryotes (1,4,5,16,23).
Second, using morphological and genetic techniques, we
show that Ub attachment to type-I and -II membrane pro-
teins without signalling ability and endocytic motifs can
target reporter molecules for efficient internalization via a
clathrin-dependent and caveolin-independent pathway.
These observations are consistent with an earlier report,
demonstrating that overexpression of a Ub chimera com-
petes with the internalization of membrane proteins har-
bouring Tyr-based endocytic motifs (24). Our results are
also in agreement with data obtained by functional, bio-
chemical and morphological techniques, documenting that
polyubiquitinated CD4, MHC class I and b2-AR/arrestincomplex undergo clathrin-dependent internalization
(48,49). Notably, an alteration of EGFR internalization from
clathrin-dependent to caveolin-dependent has been
reported upon increasing EGF concentration (21). A hier-
archical interplay between Ub- and signalling-dependent-
sorting mechanisms may offer a plausible explanation for
this phenomenon. While the latter may target the activated
EGFR receptor to a detergent-resistant, cholesterol-rich
membrane (50), multiple-monoubiqutination of the receptor
would provide a direct link to the clathrin-dependent uptake
route via Ub-binding clathrin adaptors. The activation state
of the different signalling molecules and their interaction
with the signal-decoding machinery may define the ultimate
route(s) of EGFR endocytosis in a cell type specific manner.
The third characteristic of the Ub-dependent internalization
signal is that a single Ub moiety is insufficient to confer
endocytic activity to cell surface resident reporter
CHC
AP-2
epsin
eps15 eps15R
Ubiquitinated cargo
UIM
Ub
Figure 7: Schematic model for the recruitment of ubiquiti-
nated membrane proteins to clathrin-coated pits/vesicles
(CCP/CCV) based on previously established interactions and
the present work.
Barriere et al.
292 Traffic 2006; 7: 282–297

molecules and by inference to native cargos in mammalian
cells. CD4Tl-Ub internalization was inhibited by E1 inacti-
vation or by replacing the Ub moiety with UbAllR.
Meanwhile, defective endocytosis of CD4Tl-UbAllR was
rescued by the formation of polymeric Ub upon oligomer-
ization of the chimera, or by reintroducing a single Lys
residue into UbAllR. The oligoubiquitination propensity of
CD4Tl-Ub and IiT-Ub cannot be attributed to specific char-
acteristics of the chimeras per se, considering their similar
behaviour in the context of type-I and type-II membrane
proteins. Supporting the notion that oligoubiquitin is recog-
nized as an efficient internalization signal in mammalian
cells, modulator of immune recognition 1 an E3 Ub-ligase
encoded by the Kaposi’s sarcoma-associated herpesvirus
triggers the polyubiquitination and efficient endocytosis of
a Lys-less MHC I molecules, harbouring a single Cys resi-
due in its cytoplasmic tail as the Ub-acceptor site (51).
Finally, polyubiquitination of the TrkA receptor has been
recently reported to be indispensable for its ligand-induced
internalization (52).
Importantly, our results uncovered the structural diversity
of oligo-Ub recognition as an endocytic signal, represented
by its polymeric and oligomeric forms, in higher eukaryotic
cells. Induction of non-covalent oligomers of CD4Tl-UbAllR
by antibody cross-linking or inducing tetramerization
restored the endocytic activity of the reporter molecule
at least to that of CD4Tl-Ub (Figure 2F). These observa-
tions provide a mechanistic basis for the equally efficient
internalization of numerous oligomeric and monomeric
plasma membrane proteins that are subjected to mono-
ubiquitination, multiple-monoubiquitination and/or polyubi-
quitination [e.g. ROMK1 (27), ENaC (53), EGFR (10,54),
b2-AR/arrestin complex (44), CD4 (47) and MHC I, B7.2
(55)]. The low affinity binding of Ub to a variety of UIMs in
vitro (17,36,37,56) offers a possible explanation for the
inability of mono-Ub to be efficiently recognized as an
autonomous internalization signal in mammalian cells.
This inference is consistent with binding studies, demon-
strating that polymeric Ub, but not the monomeric Ub,
binds to the hetero-oligomeric complexes of eps15
0 10 20 30% Internalized
40 50 60
CD4CD4 Lys-less
March-IV
March-VIII
March-VIII
March-IV
-
-
CD4Tl-Ub1A
UbAllR
A B
C D
UbAllR-1A
UbAllR-2A
UbAllR-3A
0 25% Inhibition
50 75 0 25 50 75
0 25 50 75 100% Inhibition of CD4 internalization
+March-IV –March-IV
eps15epsin
AP-2αCHC
eps15
epsinAP-2α
CHC
CD
4TC
C-
% Inhibition of β2-AR internalization
100
eps15
eps15R
epsin
eps15eps15R
epsin
siRNA
CHCeps15 eps15R epsin
– – + – + – ++
+Isoproterenol
Non-stimulated
Figure 8: Adaptor requirement for efficient internalization of ubiquitinated CD4 and b2-adrenergic receptor (b2-AR). (A)
Recognition surface of ubiquitin (Ub) by endocytic adaptors. COS-7 cells transiently transfected with CD4TCC-UbAllR, CD4TCC-UbAllR-
1A (I44A), CD4TCC-UbAllR-2A (L8A, V70A), CD4TCC-UbAllR-3A (L8A, I44A and V70A) and CD4Tl-Ub1A (I44A). Endocytic rates of
chimeras were determined by antibody uptake (5 min, 37 �C) and expressed as percentage of CD4TCC-UbAllR and CD4Tl-Ub, respec-
tively. (B) Adaptor sensitivity of agonist-stimulated b2-AR internalization. Endocytosis of the Flag-b2-AR was measured in the absence or
presence of 10 mM isoproterenol in small interfering RNA (siRNA)-treated HEK293 cells (5 min, 37 �C) as described in Materials and
Methods. The lower panel illustrates the indicated siRNA efficiency on protein expression in a representative experiment, visualized by
immunoblotting. Non-specific siRNA was used as control. (C) March-IV-mediated ubiquitination accelerates CD4 internalization.
Internalization rates of CD4 and Lys-less CD4 were determined in the absence or presence of heterologously expressed March-IV
(membrane-associated RING-CH) or March-VIII by the antibody uptake assay (5 min, 37 �C) in HEK293 cells. (D) Clathrin heavy chain and
adaptor dependence of ubiquitinated CD4 internalization. HEK293 cells were treated with siRNA and after 48 h co-transfected with CD4
and March-IV or March-VIII. Forty-eight hours later, internalization of CD4 was measured by antibody uptake. Means � SEM, n ¼ 4.
Oligoubiquitin Recognition as Endocytic Signal
Traffic 2006; 7: 282–297 293

isolated from HeLa cell lysates (Figure 5A), or to purified,
recombinant eps15 (Figure 5B) and the UIMs of epsin and
eps15 (19,35). We propose that productive recognition
and uptake of ubiquitinated cargo necessitate polyvalent
Ub–UIM interactions to stabilize the low affinity mono-Ub-
UIM binding. This process could be facilitated by the
oligomerization of eps15/15R and epsin, increasing their
binding avidity to cargo molecules with polymeric Ub moi-
eties and/or by post-translational modifications of adaptors
(13). Association of ubiquitinated cargo may enhance the
affinity of the eps15/15R and epsin adaptors to oligo-Ub as
well as to each other in a co-operative manner. Due to
structural differences, certain Ub-binding motifs may have
preferential binding to oligo-Ub itself, as described for the
C-terminal UIM of the S5a subunit of the proteasome (57)
and the tandem UIMs in epsin and eps15 (19,35).
Uncovering the contribution of these mechanisms to ubi-
quitinated cargo internalization in vivo will require further
experimentations.
Materials and Methods
Cells and transfectionCOS-7, HEK293T, HeLa, CHO E36 cells were grown in Dulbecco’s
Modified Eagle’s Medium (DMEM) containing 10% fetal bovine serum in
thermostated cell culture incubator in 5% CO2 at 37 �C. CHO ts20 cells
were cultured at 32 �C and incubated for the indicated time at 40 �C to
down-regulate the E1 Ub-activating enzyme (29). HEK293 cells stably
expressing CD4Tl or CD4Tl-Ub were selected after transfection with the
pIRES2 expression plasmid (BD Biosciences Inc, Mississauga, ON,
Canada), encoding the respective polypeptide in the presence of 10 mg/mL puromycin. HEK293 cells stably expressing the N-terminally tagged
Flag-b2-AR (58) were kindly provided by Dr Mark von Zastrow (University
California San Francisco, San Francisco, CA, USA). Transient transfection
of COS-7 and HeLa cells was performed with FuGENETM 6 (Roche, Basel,
Switzerland) according to manufacture’s recommendation and analysed
after 48 h.
Plasmid constructionsTo generate the human CD4 chimeras, we fused Ub at the COOH terminus
of CD4, CD4Tl or CD4T via the flexible PRARDPGGGSGGGTGGGSGGG
linker. The linker-flanked Ub was fused in frame to the full-length CD4 in
CD4-Ub, and to the extracellular and transmembrane domain of CD4
(terminated at Phe418) either directly in CD4T-Ub, or via the
SRQEVEVQVEGGSGGGGSG spacer in CD4Tl-Ub (Figure 1A). Mutant Ubs
were generated by PCR mutagenesis, using appropriate templates and
cloned into the XmaI/ClaI sites of CD4Tl-Ub. The cDNA of wt Ub and the
Lys-less Ub (UbAllR) were kindly provided by Dr L. Hicke (North-western
University, Evanston, IL, USA). The cDNAs of R48KUbAllR and
R63KUbAllR were kind gifts of Dr Y. Yarden (The Weizmann Institute of
Science, Rehovot, Israel). The cDNA of UbK29R, UbK48R and UbK63R
were kind gifts of Dr R. Haguenauer-Tsapis (Institute Jacques Monod-
CNRS Universites Paris VI and Paris VII, Paris, France). R6KUbAllR,
R29KUbAllR, UbAllR-1 A (I44A), UbAllR-2 A (L8AV70A) and UbAllR-3 A
(L8AI44AV70A) were generated by overlapping PCR. The cDNA of CD4
containing only Arg residues in its cytoplasmic tail (Lys less) was kindly
provided by Dr U. Schubert (University of Erlangen-Nuremberg, Germany).
GST and CD4T containing the tetramerization CC domain of the potassium
channel Kir6.2 tail were generated by PCR, using the ZZ-pQE60-CC6.2-KKK
plasmid as template, kindly provided by Dr B. Schwappach (ZMBH,
Heidelberg, Germany). The tetramerization domain was attached to the
carboxyl terminus of CD4T (CD4TCC) and to the carboxyl terminus of GST
(GSTCC). Wt or mutant Ub was fused in frame to the CC domain (e.g.
GSTCC-Ub and CD4TCC-Ub). CD4Tl-2Ub and CD4Tl-3Ub were constructed
by subcloning the cDNA encoding the two or three tandem Ub from the
respective pGEX4T plasmids (see below). The dominant negative
dynamin1K44A was a kind gift of Dr S. Schmid (Scripps Institute, CA,
USA). Ii chimeras were engineered by PCR mutagenesis, using the full-
length Ii cDNA (Mammalian Gene Collection, clone BC018726) and sub-
cloned into the pcDNA3.1. The N-terminal 33 amino acid residues of Ii,
containing an internalization signal, were deleted by PCR (IiT) as described
(24). Wt Ub and UbAllR lacking 76GlyGly were fused in frame to the
N-terminus of IiT, yielding IiT-Ub and IiT-UbAllR. All constructs were ver-
ified by DNA sequencing. The construction of CD4-Lamp1 chimera was
described (59).
Recombinant protein purification and pull-down
assayGST fusions, containing one (GST-Ub), two (GST-2Ub) or three (GST-3Ub)
tandem Ub, were constructed by inserting the relevant Ub moieties into
pGEX-4T1. The cDNA for single and multiple Ub fusions were PCR ampli-
fied using pcDNA3.1-2xUb-PL-Bla and pcDNA3.1-3xUb-PL-Bla as tem-
plates (60), containing tandem Ub moieties lacking their C-terminal Gly
residues, kindly provided by Heather Clarke (Aurora Biosciences
Corporation, San Diego, CA, USA). GST-eps15 was obtained by subcloning
hEps15 from pHAT-Eps15, kindly provided by Dr H. Stenmark, into pGEX-
6P1. Protein expression was induced in HB101 cells with 0.3 mM IPTG for
3 h at 37 �C. Recombinant proteins were purified using glutathione-
Sepharose 4B and stored in 150 mM NaCl, 1 mM MgCl2 and 5% glycerol
at �80 �C. For GST and GST-Ub/2U/3Ub pull-down assays, 60 mg recom-
binant proteins were bound to glutathione-Sepharose 4B (150 min, 4 �C),washed three times (150 mM NaCl, 1 mM MgCl2, 0.5 mM EGTA, 20 mM
HEPES/NaOH, 5% glycerol, 0.1% TritonX-100 and pH 7.4) and incubated
with 8 mg HeLa cell lysate (150 min, 4 �C). The beads were washed on a
step-gradient consisting of 17% glycerol and 37% sucrose (0.4 mL), 17%
glycerol (0.2 mL) and 0.2% NP-40 in 150 mM NaCl, 20 mM Hepes/NaOH
pH 7.4, 1 mM MgCl2 (0.2 mL). Bound proteins were eluted with 20 mM
glutathione in 20 mM Tris/HCl, pH 7.4 (25 min, 22 �C).
In the case of GST-Eps15 pull-down, 20 mg purified recombinant proteins
were bound to glutathione beads (120 min at 4 �C), washed three times
(20 mM TRIS/HCl pH 7.4, 150 mM NaCl, 5% glycerol) and incubated with
mono-Ub (50 mg, Sigma, St Louis, MO, USA), polyUb K48-linked chain
(12 mg, Boston Biochem, Cambridge, MA, USA) and polyUb K63-linked
chain (6 mg, Boston Biochem) for 120 min at 4 �C. The beads were washed
as described above, and the bound proteins were eluted with 30 mM
glutathione in 100 mM TRIS/HCl, pH 7.4 (25 min at RT).
Western blotting and immunoprecipitationCD4 (H-370), epsin1 (R-20), eps15 (C-20) antibodies were from Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA CHC (610499, clone 23), caveo-
lin1 (610059, clone pAb), GGA3 (612310, clone 8) and invariant chain Ii
(555612, clone LN2) antibodies were from BD Biosciences, Inc. Anti-E1
enzyme antibody was from Covance, Inc., Berkeley, CA, USA and OKT4
from ATCC, Manassas, VA. Eps15R (AP2160a) antibody was purchased
from ABGENT, Inc (San Diego, CA, USA). For immunoblotting, cells were
lysed in RIPA buffer (150 mM NaCl, 20 mM Tris-HCl, 1% (v/v) TritonX-100,
0.1% (w/v) SDS and 0.5% (w/v) sodium deoxycholate, pH 8.0) supplemen-
ted with protease inhibitors (10 mg/mL leupeptin, 10 mg/mL pepstatin and
1 mM PMSF). Polypeptides were separated by SDS-PAGE, transferred to
nitrocellulose membranes and visualized by enhanced chemiluminescence
Amersham Biosciences, Inc. (Quebec, Canada). Densitometric analyses
were performed with NIH IMAGE 1.62 software as described (59).
CD4 chimeras, confined to the cell surface and endosomes, were immuno-
isolated from transiently transfected COS-7 cells after internalization of
OKT4 anti-CD4 antibody (30 min, 37 �C). Excess antibody was then
removed, and cells were lysed in 150 mM NaCl, 10 mM Tris-Cl, 0.2% NP-
40 and pH 7.4, supplemented with 10 mg/mL pepstatin, 10 mg/mL leupep-
tin, 1 mM PMSF, 10 mM MG132 and 5 mM NEM. Immunocomplexes were
isolated on protein G Sepharose (Sigma). Immunopecipitation of CD4
variants from cell lysates was carried out similarly.
Barriere et al.
294 Traffic 2006; 7: 282–297

Recombinant GST-Ub and GSTCC-Ub production was induced with 0.3 mM
IPTG in HB101 cells. The bacteria were sonicated in 150 mM NaCl, 10 mM
Tris-Cl, 5 mM EGTA, 10 mg/mL leupeptin, 10 mg/mL pepstatin and 1 mM
PMSF. Insoluble materials were sedimented by ultracentrifugation
(110,000 � g, 30 min at 4 �C), and polypeptides were fractionated by
size-exclusion chromatography using a Superdex200 10/300G column
(Amersham Biosciences) with 100 mM Tris-Cl, 150 mM NaCl, 5% glycerol,
5 mM EGTA, 1 mM DTT and pH 7.5 as elution buffer. Fractions were
immunoblotted with the polyclonal anti-GST antibody (Amersham
Biosciences)
Cell surface density and internalization
measurementsThe cell surface density of CD4 chimeras was determined by mouse
monoclonal primary anti-CD4 (RPA-T4 or OKT4, 0 �C, 1 h) and 125I-labelled
goat anti-mouse secondary antibody (3 mCi/mL, Amersham Biosciences,
Inc. (Quebec, Canada), 0 �C, 1 h) binding in PBS supplemented with 1 mM
CaCl2, 1 mM MgCl2 (PBS/Ca2þMg2þ) and 0.5% BSA. Alternatively, the
primary antibody was detected by horseradish peroxydase (HRP)-conju-
gated goat anti-mouse secondary antibody (Amersham Biosciences)
using Amplex�Red (10-acetyl-3,7-dihydroxyphenoxazine, Molecular
Probes, Eugene, OR, USA) as substrate. The fluorescence was measured
by reading with POLARstar OPTIMA (BMG Labtech Inc., Offenburg,
Germany) plate-reader using filters 544 nm for excitation and 590 nm for
emission. Specific binding was calculated by subtracting the signal in the
presence of 10 mg/mL non-immune IgG (Santa Cruz Biotechnology Inc.)
and the respective secondary antibody. Non-specific binding was usually
5–10% of the specific signal.
Following the cell surface binding of anti-CD4 antibody (4 �C for 1 h),
internalization rates of CD4 and CD4 chimeras were calculated from the
loss of cell surface bound anti-CD4 antibody during 5 min internalization at
37 �C and expressed as percentage of radioactivity or fluorescence relative
to that detected initially. Comparable results were obtained with the radio-
active and the fluorescence technique (data not shown). To obtain the
initial uptake rates, we reduced the internalization time to 3 min for
CD4Tl-2Ub and CD4Tl-3Ub at 37 �C (Figure 2C). The internalization rates
of CD4 are not influenced by the valency of the antibody (61) and by the
variations of cell surface expression level of the chimera (see Result).
While the internalization deficient chimeras displayed a fourfold to fivefold
higher cell surface expression level than CD4Tl-Ub, insertion of the tetra-
merization domain decreased the relative cell surface density of the
CD4TCC-Ub and CD4TCC-UbAllR by 40% compared with CD4Tl-Ub.
Chimeras displaying the lowest cell surface expression (CD4Tl-2Ub and
CD4Tl-3Ub) yielded specific signal that was still twofold to threefold higher
than the background signal (data not shown). Antibody uptake assay was
used to determine the internalization of the Ii chimeras and the Flag-b2-ARwith anti-Ii and M2 anti-Flag antibodies, respectively. Tf uptake was mon-
itored after binding of 3 mg/mL Tf-biotin (Molecular Probes, T23363) to
serum-deprived cells in PBS/CM and 0.5% ovalbumin (0 �C, 1 h). The
amount of cell surface associated Tf-biotin before and after internalization
was determined by Amplex�Red, using 0.25 mg/mL streptavidin-HRP (1 h,
0 �C). Endocytosis of CTB was monitored at subsaturating concentration
(10 nM) of HRP-conjugated CTB (C-4672, Sigma) (data not shown).
Endocytosis rates were calculated as described for the CD4 chimeras.
Each internalization experiment was performed in at least triplicates.
CD4 cross-linking at the plasma membraneNon-covalent cross-linking of CD4Tl-UbAllR and CD4Tl was performed by
two protocols in COS-7 cells. Protocol 1 entails sequential binding of
primary anti-CD4 (OKT4, 0 �C, 1 h) and goat anti-mouse Ab (115-005-164,
Jackson ImmunoResearch Laboratory Inc., West Grove, PA, USA 0 �C,1 h). Internalization of CD4-antibody complex was detected by HRP-con-
jugated rabbit anti-goat tertiary antibody (0 �C, 1 h) (305-035-003, Jackson
ImmunoResearch Laboratory Inc.). In Protocol 2, cells were sequentially
incubated with anti-CD4 primary, biotinylated secondary antibody and
finally with streptavidin-HRP (0 �C, 1 h for each incubation). The cell sur-
face density of complexes was measured before and after 5 min of inter-
nalization at 37 �C by Amplex�Red.
Inhibition of clathrin- and caveolae-dependent
endocytosisThree different approaches were employed to inhibit CCV formation in
COS-7 cells as described previously (62). (i) Depletion of the cytosolic Kþ
content was achieved by hypotonic swelling of the cells in 50% diluted
medium, containing 1 mM ouabain (5 min, 37 �C) and then in Kþ-freemedium (100 mM NaCl, 50 mM Hepes, 1 mM MgCl2, 0.1 mM CaCl2,
10 mM glucose, pH 7.3) for 20 min at 37 �C. Internalization was measured
in Kþ-free medium. (ii) Cells were exposed to hypertonic DMEM supple-
mented with 0.3 M sucrose for 30 min at 4 �C, and internalization was
measured in the same medium. (iii) Acidification of the cytosol was accom-
plished with the NH4Cl prepulse method. Cells were incubated in DMEM
supplemented with 40 mM NH4Cl (5 min, 37 �C), transferred to Naþ-freemedium (140 mM N-methyl-D-glucamine-Cl, 10 mM Hepes, 1 mM MgCl2,
0.1 mM CaCl2, 10 mM glucose, pH 7.3). CD4Tl-Ub uptake was monitored in
Naþ-free medium. Cholesterol sequestration was achieved by incubating
HEK293 cells with 25 mg/mL nystatin or 0.5 mg/mL filipin for 1 h at 37 �C(21). Cholesterol synthesis was inhibited with 1 mM mevinolin for 24 h.
SiRNAsiRNA duplexes (Dharmacon Inc. Lafayette, CO, USA) were resuspended in 1X
siRNA Universal Buffer (Dharmacon) at 20 mM concentration. HEK293 cells
were transfected with oligofectamine (Invitrogen, Inc. Burlington, Canada) at
final concentration of 80 nM siRNA according to the manufacture’s recommen-
dation. Depletion of target proteins was optimized by repeated siRNA transfec-
tions with previously validated siRNA as indicated. CHC
(5’CCTGCGGTCTGGAGTCAAC) (63), AP-2a (5’-GAGGTGTGGTACCGAGTCA)
(63), caveolin1 (5’-TGTCTGGGGGCAAATACG) (64) and GGA3 (5’-
AAACGGCTTCCGCATCCTC) (65) knockdown was achieved by transfecting
the respective siRNA twice at 24 h intervals, and experiments were done
after 48 h of the second transfection. Eps15 (5’’-AAACGGAGCTACAGATTAT)
(66), eps15R (5’-GCACTTGGATCGAGATGAG) (66) and epsin1 (5’-
GGAAGACGCCGGAGTCATT) (66) siRNAs were transfected three times with
24 h intervals between each transfection, and experiments were performed
after 48 h of the last transfection. When two siRNAs were used together, the
concentration of individual siRNA was 40 nM. Controls included mock transfec-
tion in the absence of siRNA as well as using 80 nM non-specific (ns) siRNA.
MicroscopyImmunofluorescence localization of CD4 variants was performed using a
mouse anti-CD4 (RPA-T4) antibody (BD Biosciences) and FITC-conjugated
donkey anti-mouse antibodies. Co-localization of CD4Tl-Ub with GM1-
enrichedmembranemicrodomainswas achieved after patching CD4 variants
with OKT4 anti-CD4 antibody (1 h, 12 �C) and then with anti-mouse TRITC-
conjugated antibody (1 h, 12 �C). CTB-FITC (Sigma) was included with the
secondary antibody to visualize rafts. CD4 chimeras and TfRwere co-patched
using polyclonal sheep anti-CD4 and mouse monoclonal anti-TfR (OKT9)
antibody (1 h, 12 �C), followed by a second incubation with FITC-conjugated
anti-sheep and TRITC-conjugated anti-mouse secondary antibodies (1 h,
12 �C) before fixation. Membrane proteins excluded from rafts were
removed with 1% TritonX-100 containing PBS (20 min, 4 �C) prior to fixation.
Images were acquired with a Zeiss LSM510 laser confocal microscope
equipped with a Plan-Achromat 63X/NA 1.4 (Carl Zeiss Microimaging, Inc.,
Jena, Germany).
Statistical analysisExperiments were repeated at least three times. Data, means � SEM.
Significance was assessed by calculating two-tailed p-values at 95% con-
fidence level with unpaired t-test, using the PRISM software (GraphPad
Software, Inc. San Diego, CA, USA).
Acknowledgments
We are indebted to R. Haguenauer-Tsapis, L. Hicke, U. Schubert, S.
Schmid, B. Schwappach, Y. Yarden, H. Stenmark and M. von Zastrow for
generously providing valuable reagents for our studies. H.B was supported
Oligoubiquitin Recognition as Endocytic Signal
Traffic 2006; 7: 282–297 295

in part by Fellowships from the RESTRACOM, Hospital for Sick Children
and The Canadian Cystic Fibrosis Foundation (CCFF). This work was sup-
ported by grants to G.L. from the Canadian Institutes of Health Research
(CIHR), the Premier Research Excellence Award of the Ontario Ministry of
Energy and Education and CCFF. The instrumentation was supported in
part by a Block Term Grant from the Ontario Thoracic Society.
References
1. Bonifacino JS Traub LM. Signals for sorting of transmembrane proteins
to endosomes and lysosomes. Annu Rev Biochem 2003;72:395–447.
2. Ellgaard L Helenius A. Quality control in the endoplasmic reticulum.
Nat Rev Mol Cell Biol 2003;4:181–191.
3. Hicke L Dunn R. Regulation of membrane protein transport by ubiquitin
and ubiquitin-binding proteins. Annu Rev Cell Dev Biol 2003;19:141–172.
4. Dupre S, Urban-Grimal D, Haguenauer-Tsapis R. Ubiquitin and endocy-
tic internalization in yeast and animal cells. Biochim Biophys Acta
2004;1695:89–111.
5. Shih SC, Sloper-Mould KE, Hicke L. Monoubiquitin carries a novel
internalization signal that is appended to activated receptors. EMBO
J 2000;19:187–198.
6. Slagsvold T Stenmark H. The structure of an endosomal protein sorter.
Dev Cell 2004;7:457–458.
7. Aguilar RC, Watson HA, Wendland B. The yeast Epsin Ent1 is recruited
to membranes through multiple independent interactions. J Biol Chem
2003;278:10737–10743.
8. Haglund K, Sigismund S, Polo S, Szymkiewicz I, Di Fiore PP, Dikic I.
Multiple monoubiquitination of RTKs is sufficient for their endocytosis
and degradation. Nat Cell Biol 2003;5:461–466.
9. Di Fiore PP, Polo S, Hofmann K. When ubiquitin meets ubiquitin recep-
tors: a signalling connection. Nat Rev Mol Cell Biol 2003;4:491–497.
10. Mosesson Y, Shtiegman K, KatzM, Zwang Y, Vereb G, Szollosi J, Yarden
Y. Endocytosis of receptor tyrosine kinases is driven by monoubiquityla-
tion, not polyubiquitylation. J Biol Chem 2003;278:21323–21326.
11. McCullough J, Clague MJ, Urbe S. AMSH is an endosome-associated
ubiquitin isopeptidase. J Cell Biol 2004;166:487–492.
12. Hicke L, Schubert HL, Hill CP. Ubiquitin-binding domains. Nat Rev Mol
Cell Biol 2005;6:610–621.
13. Polo S, Sigismund S, Faretta M, Guidi M, CapuaMR, Bossi G, Chen H, De
Camilli P, Di Fiore PP. A single motif responsible for ubiquitin recognition
and monoubiquitination in endocytic proteins. Nature 2002;416:451–455.
14. Shih SC, Katzmann DJ, Schnell JD, Sutanto M, Emr SD, Hicke L.
Epsins and Vps27p/Hrs contain ubiquitin-binding domains that function
in receptor endocytosis. Nat Cell Biol 2002;4:389–393.
15. Bonifacino JS. The GGA proteins: adaptors on the move. Nat Rev Mol
Cell Biol 2004;5:23–32.
16. Traub LM. Sorting it out: AP-2 and alternate clathrin adaptors in endo-
cytic cargo selection. J Cell Biol 2003;163:203–208.
17. Fisher RD, Wang B, Alam SL, Higginson DS, Robinson H, Sundquist
WI, Hill CP. Structure and ubiquitin binding of the ubiquitin-interacting
motif. J Biol Chem 2003;278:28976–28984.
18. Sugiyama S, Kishida S, Chayama K, Koyama S, Kikuchi A. Ubiquitin-
interacting motifs of Epsin are involved in the regulation of insulin-
dependent endocytosis. J Biochem (Tokyo) 2005;137:355–364.
19. Miller SL, Malotky E, O’Bryan JP. Analysis of the role of ubiquitin-
interacting motifs in ubiquitin binding and ubiquitylation. J Biol Chem
2004;279:33528–33537.
20. Chen H De Camilli P. The association of epsin with ubiquitinated cargo
along the endocytic pathway is negatively regulated by its interaction
with clathrin. Proc Natl Acad Sci USA 2005;102:2766–2771.
21. Sigismund S, Woelk T, Puri C, Maspero E, Tacchetti C, Transidico P, Di
Fiore PP, Polo S. Clathrin-independent endocytosis of ubiquitinated
cargos. Proc Natl Acad Sci USA 2005;102:2760–2765.
22. StangE,BlystadFD,KazazicM,BertelsenV,BrodahlT,RaiborgC,StenmarkH,
Madshus IH. Cbl-dependent ubiquitination is required for progression
of EGF receptors into clathrin-coated pits.Mol Biol Cell 2004;15:3591–3604.
23. Wendland B. Epsins: adaptors in endocytosis? Nat Rev Mol Cell Biol
2002;3:971–977.
24. Nakatsu F, Sakuma M, Matsuo Y, Arase H, Yamasaki S, Nakamura N,
Saito T, Ohno H. A Di-leucine signal in the ubiquitin moiety. Possible
involvement in ubiquitination-mediated endocytosis. J Biol Chem
2000;275:26213–26219.
25. Pickart CM. Mechanisms underlying ubiquitination. Annu Rev Biochem
2001;70:503–533.
26. Gruenberg J. The endocytic pathway: a mosaic of domains. Nat Rev
Mol Cell Biol 2001;2:721–730.
27. Lin DH, Sterling H, Wang Z, Babilonia E, Yang B, Dong K, Hebert SC,
Giebisch G, Wang WH. ROMK1 channel activity is regulated by mono-
ubiquitination. Proc Natl Acad Sci USA 2005;102:4306–4311.
28. Yuan H, Michelsen K, Schwappach B. 14-3-3 dimers probe the assem-
bly status of multimeric membrane proteins. Curr Biol 2003;13:638–646.
29. Kulka RG, Raboy B, Schuster R, Parag HA, Diamond G, Ciechanover A,
Marcus M. A Chinese hamster cell cycle mutant arrested at G2 phase
has a temperature-sensitive ubiquitin-activating enzyme, E1. J Biol
Chem 1988;263:15726–15731.
30. Klapisz E, Sorokina I, Lemeer S, Pijnenburg M, Verkleij AJ, van Bergen
en Henegouwen PM. A ubiquitin-interacting motif (UIM) is essential for
Eps15 and Eps15R ubiquitination. J Biol Chem 2002;277:30746–30753.
31. Nichols BJ, Kenworthy AK, Polishchuk RS, Lodge R, Roberts TH,
Hirschberg K, Phair RD, Lippinctt-Schwartz J. Rapid cycling of lipid
raft markers between the cell surface and Golgi complex. J Cell Biol
2001;153:529–541.
32. Tebar F, Sorkina T, Sorkin A, Ericsson M, Kirchhausen T. Eps15 is a
component of clathrin-coated pits and vesicles and is located at the rim
of coated pits. J Biol Chem 1996;271:28727–28730.
33. Wang W Struhl G. Drosophila Epsin mediates a select endocytic path-
way that DSL ligands must enter to activate Notch. Development
2004;131:5367–5380.
34. Sharma M, Pampinella F, Nemes C, Benharouga M, So J, Du K, Bache
KG, Papsin B, Zerangue N, Stenmark H, Lukacs GL. Misfolding diverts
CFTR from recycling to degradation: quality control at early endo-
somes. J Cell Biol 2004;164:923–933.
35. Hawryluk M, Keyel P, Sanjay K, Watkins S, Heuser J, Traub L. Epsin 1 is
a polyubiquitin-selective clathrin-associated sorting protein. Traffic 2006;
7:262–281.
36. Raiborg C, Bache KG, Gillooly DJ, Madshus IH, Stang E, Stenmark H.
Hrs sorts ubiquitinated proteins into clathrin-coated microdomains of
early endosomes. Nat Cell Biol 2002;4:394–398.
37. Swanson KA, Kang RS, Stamenova SD, Hicke L, Radhakrishnan I. Solution
structure of Vps27UIM-ubiquitin complex important for endosomal sorting
and receptor downregulation. EMBO J 2003;22:4597–4606.
38. Traub LM. Common principles in clathrin-mediated sorting at the Golgi and
the plasma membrane. Biochim Biophys Acta 2005;1744:415–437.
39. Chen H, Fre S, Slepnev VI, Capua MR, Takei K, Butler MH, Di Fiore PP,
DeCamilli P. Epsin is an EH-domain-binding protein implicated in
clathrin-mediated endocytosis. Nature 1998;394:793–797.
40. Benmerah A, Lamaze C, Begue B, Schmid SL, Dautry-Varsat A, Cerf-
Bensussan N. AP-2/Eps15 interaction is required for receptor-
mediated endocytosis. J Cell Biol 1998;140:1055–1062.
41. Robinson MS. Adaptable adaptors for coated vesicles. Trends Cell Biol
2004;14:167–174.
42. Cupers P, ter Haar E, Boll W, Kirchhausen T. Parallel dimers and
anti-parallel tetramers formed by epidermal growth factor receptor
pathway substrate clone 15. J Biol Chem 1997;272:33430–33434.
43. Shenoy SK Lefkowitz RJ. Receptor-specific ubiquitination of beta-
arrestin directs assembly and targeting of seven-transmembrane
receptor signalosomes. J Biol Chem 2005;280:15315–15324.
Barriere et al.
296 Traffic 2006; 7: 282–297

44. Shenoy SK, McDonald PH, Kohout TA, Lefkowitz RJ. Regulation of
receptor fate by ubiquitination of activated beta 2-adrenergic receptor
and beta-arrestin. Science 2001;294:1307–1313.
45. Mansouri M, Bartee E, Gouveia K, Hovey Nerenberg BT, Barrett J,
Thomas L, Thomas G, McFadden G, Fruh K. The PHD/LAP-domain
protein M153R of myxomavirus is a ubiquitin ligase that induces the
rapid internalization and lysosomal destruction of CD4. J Virol
2003;77:1427–1440.
46. Shackelford J Pagano JS. Tumor viruses and cell signaling pathways:
deubiquitination versus ubiquitination. Mol Cell Biol 2004;24:5089–5093.
47. Bartee E, Mansouri M, Hovey Nerenberg BT, Gouveia K, Fruh K.
Downregulation of major histocompatibility complex class I by human
ubiquitin ligases related to viral immune evasion proteins. J Virol
2004;78:1109–1120.
48. Lehner PJ, Hoer S, Dodd R, Duncan LM. Downregulation of cell sur-
face receptors by the K3 family of viral and cellular ubiquitin E3 ligases.
Immunol Rev 2005;207:112–125.
49. Lefkowitz RJ Shenoy SK. Transduction of receptor signals by beta-
arrestins. Science 2005;308:512–517.
50. Abulrob A, Giuseppin S, Andrade MF, McDermid A, Moreno M,
Stanimirovic D. Interactions of EGFR and caveolin-1 in human glioblas-
toma cells: evidence that tyrosine phosphorylation regulates EGFR
association with caveolae. Oncogene 2004;23:6967–6979.
51. Cadwell K Coscoy L. Ubiquitination on nonlysine residues by a viral E3
ubiquitin ligase. Science 2005;309:127–130.
52. Geetha T, Jiang J, Wooten MW. Lysine 63 polyubiquitination of the
nerve growth factor receptor TrkA directs internalization and signaling.
Mol Cell 2005;20:301–312.
53. Staub O, Gautschi I, Ishikawa T, Breitschopf K, Ciechanover A, Schild
L, Rotin D. Regulation of stability and function of the epithelial Naþ
channel (ENaC) by ubiquitination. EMBO J 1997;16:6325–6336.
54. Haglund K, Di Fiore PP, Dikic I. Distinct monoubiquitin signals in
receptor endocytosis. Trends Biochem Sci 2003;28:598–603.
55. Coscoy L, Sanchez DJ, Ganem D. A novel class of herpesvirus-encoded
membrane-bound E3 ubiquitin ligases regulates endocytosis of proteins
involved in immune recognition. J Cell Biol 2001;155:1265–1273.
56. Shekhtman A Cowburn D. A ubiquitin-interacting motif from Hrs binds
to and occludes the ubiquitin surface necessary for polyubiquitination
in monoubiquitinated proteins. Biochem Biophys Res Commun
2002;296:1222–1227.
57. Young P, Deveraux Q, Beal RE, Pickart CM, Rechsteiner M.
Characterization of two polyubiquitin binding sites in the 26 S protease
subunit 5a. J Biol Chem 1998;273:5461–5467.
58. Tanowitz M von Zastrow M. A novel endocytic recycling signal that
distinguishes the membrane trafficking of naturally occurring opioid
receptors. J Biol Chem 2003;278:45978–45986.
59. Benharouga M, Haardt M, Kartner N, Lukacs GL. COOH-terminal
truncations promote proteasome-dependent degradation of mature
cystic fibrosis transmembrane conductance regulator from post-Golgi
compartments. J Cell Biol 2001;153:957–970.
60. Stack JH, Whitney M, Rodems SM, Pollok BA. A ubiquitin-based
tagging system for controlled modulation of protein stability. Nat
Biotechnol 2000;18:1298–1302.
61. Pelchen-Matthews A, Armes JE, Marsh M. Internalization and recy-
cling of CD4 transfected into HeLa and NIH3T3 cells. EMBO J
1989;8:3641–3649.
62. Lukacs GL, Segal G, Kartner N, Grinstein S, Zhang F. Constitutive
internalization of cystic fibrosis transmembrane conductance regulator
occurs via clathrin-dependent endocytosis and is regulated by protein
phosphorylation. Biochem J 1997;328 (Pt 2):353–361.
63. Hinrichsen L, Harborth J, Andrees L, Weber K, Ungewickell EJ. Effect
of clathrin heavy chain- and alpha-adaptin-specific small inhibitory
RNAs on endocytic accessory proteins and receptor trafficking in
HeLa cells. J Biol Chem 2003;278:45160–45170.
64. Cho KA, Ryu SJ, Park JS, Jang IS, Ahn JS, Kim KT, Park SC. Senescent
phenotype can be reversed by reduction of caveolin status. J Biol
Chem 2003;278:27789–27795.
65. Puertollano R Bonifacino JS. Interactions of GGA3 with the ubiquitin
sorting machinery. Nat Cell Biol 2004;6:244–251.
66. Huang F, Khvorova A, Marshall W, Sorkin A. Analysis of clathrin-
mediated endocytosis of epidermal growth factor receptor by RNA
interference. J Biol Chem 2004;279:16657–16661.
Oligoubiquitin Recognition as Endocytic Signal
Traffic 2006; 7: 282–297 297




















