
Modulation of Nav1.5 by ?1- and ?3-subunit co-expression in mammalian cells
10
ION CHANNELS, TRANSPORTERS Seong-Hoon Ko Paul W. Lenkowski Hwa C. Lee J. Paul Mounsey Manoj K. Patel Modulation of Na v 1.5 by b1- and b3-subunit co-expression in mammalian cells Received: 9 April 2004 / Revised: 6 July 2004 / Accepted: 8 September 2004 / Published online: 28 September 2004 Ó Springer-Verlag 2004 Abstract Cardiac sodium channels (Na v 1.5) comprise a pore-forming a-subunit and auxiliary b-subunits that modulate channel function. In the heart, b1 is expressed throughout the atria and ventricles, whilst b3 is present only in the ventricles and Purkinje fibers. In view of this expression pattern, we determined the effects of b3 and b1 co-expression alone, and in combination, on Na v 1.5 stably expressed in Chinese hamster ovary cells. The current/voltage relationship was shifted À5 mV with either b1 or b3 co-expression alone and À10 mV with co-expression of both b1 and b3. In addition, b3 and b1/b3 co-expression accelerated macroscopic current decay. There were significant hyperpolarizing shifts in equilib- rium gating relationships with co-expression of b1 and b3 alone and in combination. Co-expression of b1/b3 together resulted in a greater hyperpolarizing shift in channel availability, and an increase in the slopes of equilibrium gating relationships. Co-expression of b3 and b1/b3, but not b1, slowed recovery from inactiva- tion at À90 mV. Development of inactivation at À70 and À50 mV was accelerated by b-subunit co-expression alone and in combination. b-Subunit co-expression also reduced the late Na current measured at 200 ms. In conclusion, b-subunits modulate Na v 1.5 gating with important differences between co-expression of b1 and b3 alone and b1/b3 together. Keywords Sodium channels b-Subunits Electrophysiology Introduction Voltage-gated sodium (Na) channels are at the root of cellular excitability and play a fundamental role in the generation and conduction of cardiac and neuronal ac- tion potentials. They comprise of a pore-forming a-subunit, which is sufficient to induce Na currents in heterologous expression systems, and a variable number of auxiliary b-subunits that modulate the gating prop- erties of the channel [1]. To date, nine a-subunit isoforms have been cloned [2], each of which has a distinct tissue distribution. Four b-subunit isoforms have been cloned, b1[3], b2[4], b3[5] and b4[6]. The properties of cardiac Na channel a-subunits are modulated by b1 and b3 in heterologous expression systems [2], whilst b2 is thought to have no functional role in the heart [7]. b-Subunits are glycoproteins with a single, trans- membrane-spanning segment, a large immunoglobulin- like extracellular domain, and a small intracellular peptide [3, 8]. The b1-subunit is non-covalently asso- ciated with the a-subunit, and shares significant homology with b3. It has been suggested that they are structurally distinct members of a single functional class of auxiliary subunits with different kinetic effects on channel function [5]. Co-expression of b3 with cardiac Na channels in Xenopus oocytes results in en- hanced current amplitude, a depolarizing shift in stea- dy-state inactivation and enhanced recovery from inactivation, whereas in the same expression system b1 co-expression increases current amplitude and enhances recovery from inactivation, but has no effect on steady- state gating [9]. However, the slow mode of Na channel gating, frequently observed when using Xenopus oocytes as the expression system, is considered to be abnormal and is not observed when mammalian cells are used as expression systems. In addition, mamma- S.-H. Ko P. W. Lenkowski M. K. Patel (&) Department of Anesthesiology, University of Virginia Health System, Box 800710, Charlottesville, Virginia 22908-0710, USA E-mail: [email protected] Tel.: +1-434-9249693 Fax: +1-434-9242105 H. C. Lee Department of Biomedical Engineering, University of Virginia Health System, Charlottesville, Virginia 22908, USA J. P. Mounsey Department of Internal Medicine (Cardiovascular Division), University of Virginia Health System, Charlottesville, Virginia 22908, USA Pflugers Arch – Eur J Physiol (2005) 449: 403–412 DOI 10.1007/s00424-004-1348-4
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Modulation of Nav1.5 by ?1- and ?3-subunit co-expression in mammalian cells

ION CHANNELS, TRANSPORTERS
Seong-Hoon Ko Æ Paul W. Lenkowski Æ Hwa C. Lee
J. Paul Mounsey Æ Manoj K. Patel
Modulation of Nav1.5 by b1- and b3-subunitco-expression in mammalian cells
Received: 9 April 2004 / Revised: 6 July 2004 / Accepted: 8 September 2004 / Published online: 28 September 2004� Springer-Verlag 2004
Abstract Cardiac sodium channels (Nav1.5) comprise apore-forming a-subunit and auxiliary b-subunits thatmodulate channel function. In the heart, b1 is expressedthroughout the atria and ventricles, whilst b3 is presentonly in the ventricles and Purkinje fibers. In view of thisexpression pattern, we determined the effects of b3 andb1 co-expression alone, and in combination, on Nav1.5stably expressed in Chinese hamster ovary cells. Thecurrent/voltage relationship was shifted �5 mV witheither b1 or b3 co-expression alone and �10 mV withco-expression of both b1 and b3. In addition, b3 and b1/b3co-expression accelerated macroscopic current decay.There were significant hyperpolarizing shifts in equilib-rium gating relationships with co-expression of b1 andb3 alone and in combination. Co-expression of b1/b3together resulted in a greater hyperpolarizing shift inchannel availability, and an increase in the slopes ofequilibrium gating relationships. Co-expression of b3and b1/b3, but not b1, slowed recovery from inactiva-tion at �90 mV. Development of inactivation at �70and �50 mV was accelerated by b-subunit co-expressionalone and in combination. b-Subunit co-expression alsoreduced the late Na current measured at 200 ms. Inconclusion, b-subunits modulate Nav1.5 gating withimportant differences between co-expression of b1 andb3 alone and b1/b3 together.
Keywords Sodium channels Æ b-Subunits ÆElectrophysiology
Introduction
Voltage-gated sodium (Na) channels are at the root ofcellular excitability and play a fundamental role in thegeneration and conduction of cardiac and neuronal ac-tion potentials. They comprise of a pore-forminga-subunit, which is sufficient to induce Na currents inheterologous expression systems, and a variable numberof auxiliary b-subunits that modulate the gating prop-erties of the channel [1]. To date, nine a-subunit isoformshave been cloned [2], each of which has a distinct tissuedistribution. Four b-subunit isoforms have been cloned,b1 [3], b2 [4], b3 [5] and b4 [6]. The properties of cardiacNa channel a-subunits are modulated by b1 and b3 inheterologous expression systems [2], whilst b2 is thoughtto have no functional role in the heart [7].
b-Subunits are glycoproteins with a single, trans-membrane-spanning segment, a large immunoglobulin-like extracellular domain, and a small intracellularpeptide [3, 8]. The b1-subunit is non-covalently asso-ciated with the a-subunit, and shares significanthomology with b3. It has been suggested that they arestructurally distinct members of a single functionalclass of auxiliary subunits with different kinetic effectson channel function [5]. Co-expression of b3 withcardiac Na channels in Xenopus oocytes results in en-hanced current amplitude, a depolarizing shift in stea-dy-state inactivation and enhanced recovery frominactivation, whereas in the same expression system b1co-expression increases current amplitude and enhancesrecovery from inactivation, but has no effect on steady-state gating [9]. However, the slow mode of Na channelgating, frequently observed when using Xenopusoocytes as the expression system, is considered to beabnormal and is not observed when mammalian cellsare used as expression systems. In addition, mamma-
S.-H. Ko Æ P. W. Lenkowski Æ M. K. Patel (&)Department of Anesthesiology,University of Virginia Health System, Box 800710,Charlottesville, Virginia 22908-0710, USAE-mail: [email protected].: +1-434-9249693Fax: +1-434-9242105
H. C. LeeDepartment of Biomedical Engineering,University of Virginia Health System,Charlottesville, Virginia 22908, USA
J. P. MounseyDepartment of Internal Medicine (Cardiovascular Division),University of Virginia Health System, Charlottesville,Virginia 22908, USA
Pflugers Arch – Eur J Physiol (2005) 449: 403–412DOI 10.1007/s00424-004-1348-4

lian expression systems are thought to reflect moreaccurately the channel kinetics observed in native cells[10, 11, 12, 13].
The b1-subunit is thought to interact with the P-loopof domain IV of the Na channel a-subunit [14, 15], andthe similarities between b1 and b3 suggest the possibilitythat b3 may also bind in this region. Support for thisidea comes from the observation that two of the threeamino acids essential for this interaction with b1 [16] arepresent also in b3 [5].
b1 and b3 are co-expressed in many tissues [17, 18]. Inthe heart, b1 is expressed throughout both atria andventricles, whereas b3 expression is limited to the ventri-cles and Purkinje fibers [9]. In view of this expressionpatternwe set out to determine the effects of co-expressingb1 and b3 alone and in combination on cardiac Nachannels to test the idea that this differential expressionpattern might have functional consequences in myocar-dium. Co-expression of b1 and b3 alone modulated Nachannel gating, whilst co-expression of b1/b3 in combi-nation, a scenario that could exist in the ventricle andPurkinje fibers, but not in the atrium, had a more pro-found effect than co-expression of either b-subunit alone.
Materials and methods
Transient transfection and cell culture
Chinese hamster ovary cells (CHO-K1) stably expressinghuman Nav1.5 were grown in DMEM/F12 media (In-vitrogen, Carlsbad, Calif., USA) supplemented with10% fetal bovine serum, penicillin (100 U/ml), strepto-mycin (100 lg/ml) and G418 (500 lg/ml; Sigma,St. Louis, Mo., USA). Transient transfection of rat b1and rat b3 both engineered into pcDNA3.1 were carriedout either alone or in combination (b1/b3) together withCD8 (0.2 lg/ml DNA) using Lipofectamine 2000 (Invi-trogen). The cells expressing b1 and/or b3 were grown ina humidified atmosphere of 5% CO2 and 95% air at37 �C. Two days after transfection, positive cells wereidentified by immunobeads (CD8-Dynabeads, Dynal,Norway). To ensure that expression of the b-subunitswas not limiting, cells were transfected with a range ofDNA concentrations (2, 4, or 8 lg/ml). There were nosystematic differences in the Na channel gating at anyb-subunit concentration suggesting that expression isalready in excess at 2 lg/ml DNA.
Patch-clamp electrophysiology
Sodium currents were recorded from CHO cells stablyexpressing Nav1.5 using the whole-cell configuration ofthe patch-clamp recording technique and an appropriateamplifier (Axopatch 200B, Axon Instruments, UnionCity, Calif., USA). CHO cells were selected because theydo not express b1 or b3 subunits constitutively [13]. Inaddition, although there is an endogenous Na current, it
amounted to less than 10 pA/pF in amplitude with acurrent/voltage (I/V) peak at +20 mV (n=5). All volt-age protocols were applied using appropriate software(pCLAMP v. 8, Axon) and a digital interface (Digidata1322A, Axon). Currents were amplified, low-pass fil-tered (2 kHz), and sampled at 33 kHz.
Borosilicate glass pipettes (0.6–1.0 MW) were filledwith (mM) CsCl 130, MgCl2 1, MgATP 5, BAPTA 10,HEPES 5 (pH adjusted to 7.4 with CsOH). Cells wereplated on glass cover-slips and superfused with (mM)NaCl 130, KCl 4, CaCl2 1, MgCl2 5, HEPES 5, andglucose 5 (pH adjusted to 7.4 with NaOH). Cells withmore than five beads attached were selected for record-ing. On establishing the whole-cell patch clamp, accessresistance was 1–1.5 MW and was further reduced byseries resistance compensation of 75%. Cells were heldat �80 mV for 7 min because equilibrium gating hadreached steady state after this period (n=10). Data fromcells with a current amplitude of more than 8 nA, orcells that showed evidence of poor voltage control asreflected by the shape of the I/V curve, were excludedfrom the study. All experiments were performed at roomtemperature (20–22 �C).
Voltage protocols
I/V relationships were determined with 20-ms voltagepulses from �80 to +60 mV in steps of 5 mV from aholding potential of �120 mV at 2-s intervals. Con-ductance as a function of voltage was derived from theI/V relationship as described previously [19]. Channelavailability as a function of voltage was determinedusing a two-pulse protocol, from a holding potential of�120 mV and test potentials between �130 mV and�20 mV (1 s duration) as previously described [19].Equilibrium gating plots were fitted to the Boltzmannfunction: g=1/(1+exp[(V�V1/2)/k]), where g is con-ductance, V1/2 the voltage of half-maximal inactivation,and k the slope factor.
Recovery from inactivation was assessed over 1–15,800 ms at test potentials of �110, �100, �90, �80,and �70 mV using a two-pulse protocol. Channels wereinactivated by a pre-pulse to 0 mV (holding potential�120 mV) for 100 ms. Development of inactivation wasdetermined at �70 and �50 mV over 1–15,800 ms usinga two-pulse protocol. The second pulse to +20 mV for20 ms assessed the proportion of channels inactivated.In each case currents were normalized and time con-stants (s) for both recovery and development of inacti-vation obtained using a double exponential function:y=A1[1�exp(�t/s1)]+A2[1�exp(�t/s2)], where t is thetime, A the current amplitude and the subscripts 1 and 2indicate the fast and slow gating modes respectively.
Data analysis
Data analysis was performed using appropriate software(Clampfit v. 8, Axon; Origin v. 5, Microcal Software,
404

Northampton, Mass., USA; Sigmastat, Jandel Scientific,Calif., USA; Excel, Microsoft, Redmond, Wash., USA).The significance of the differences between the peak I/Vrelationship (Fig. 1C), the voltage dependence of currentdecay (Fig. 1D), the voltage dependence of time-to-peakmacroscopic current (Fig. 1E), and inactivation kinetics(Figs. 3 and 4) were tested using a non-parametricmultivariate rank-sum test [19, 20] that consists ofassigning ranks to the measured parameters for eachcell, summing the ranks for each cell, and comparing thesums by one-way ANOVA. For groups that were sig-nificantly different from each other, a Tukey test wasused for pairwise comparisons. Standard one-wayANOVA followed by Tukey’s post hoc test was used todetermine significance of differences in Fig. 1B andFig. 2. Data are presented as means±SEM; P<0.05was considered significant.
Results
Effects of b-subunit co-expression on whole-cellNa currents
Figure 1A shows families of whole-cell Na currents.Co-expression of b1, b3 or both b1/b3 subunits incombination with Nav1.5 had no obvious effect on thecurrent amplitude, but the decay of the macroscopiccurrent appeared to be enhanced. The pooled dataconfirmed these trends. There was no detectable effecton peak current density (Fig. 1B), although the peak ofthe I/V relationship (Fig. 1C), i.e., the voltage at whichcurrent was maximal, was shifted in a hyperpolarizingdirection by �5 mV for b1 or b3 co-expression alone,and by �10 mV for b1/b3 together (P<0.05 for all
Fig. 1A–B b-Subunitmodulation of the current/voltage (I/V) relationship.A Families of whole-cell Nacurrents evoked in Chinesehamster ovary (CHO) cellsexpressing the Na+ channela-subunit Nav1.5 alone orco-transfected with b1, b3, andboth b1/b3 in combination.Currents were elicited bydepolarizing steps (20 ms) froma holding potential of �120 mVto test pulses of �80 to+70 mV in 5 mV increments.B Current density, assessed asthe peak of the I/V relationshipfor each cell divided by cellcapacitance, was unaffected byb-subunit co-expression.C Normalized peak I/Vrelationship showing significanthyperpolarizing shifts withb-subunit co-expression.Means±SEM, n=15–27 cells.D The decay of macroscopiccurrents were fitted to doubleexponential functions and thefaster time constant (s1) isshown as a function ofpotential. E Time-to-peak ofmacroscopic current as afunction of potential. In D andE, means±SEM, n=10 foreach point
405
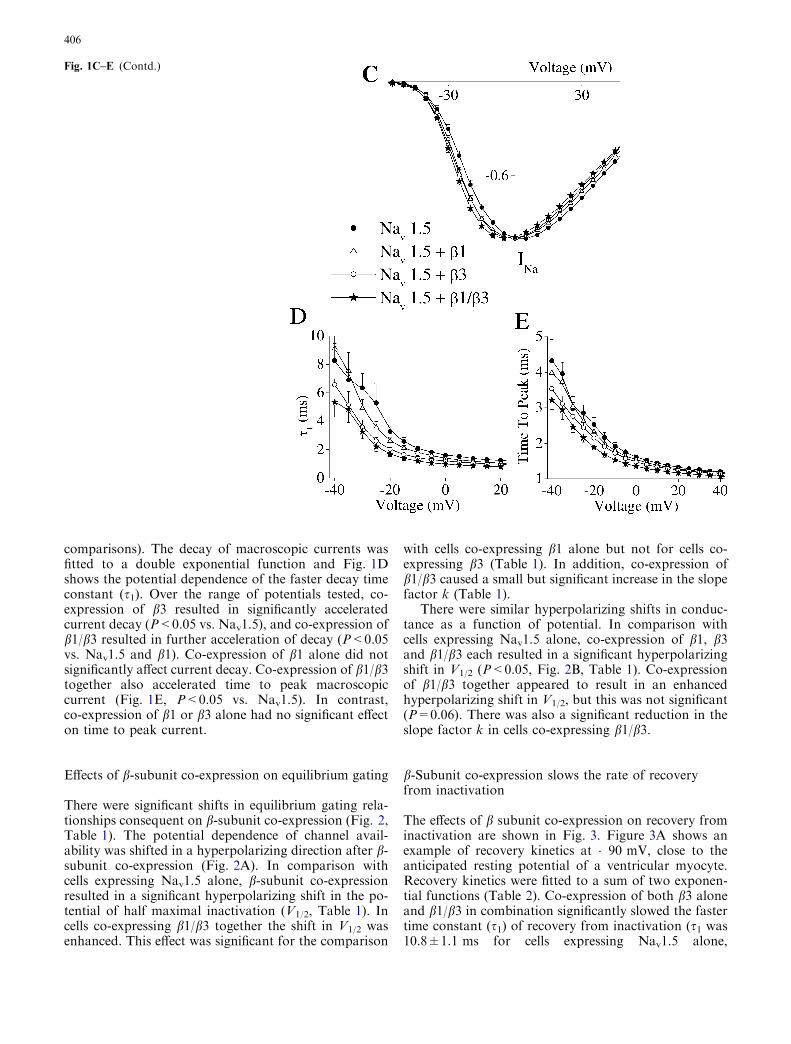
comparisons). The decay of macroscopic currents wasfitted to a double exponential function and Fig. 1Dshows the potential dependence of the faster decay timeconstant (s1). Over the range of potentials tested, co-expression of b3 resulted in significantly acceleratedcurrent decay (P<0.05 vs. Nav1.5), and co-expression ofb1/b3 resulted in further acceleration of decay (P<0.05vs. Nav1.5 and b1). Co-expression of b1 alone did notsignificantly affect current decay. Co-expression of b1/b3together also accelerated time to peak macroscopiccurrent (Fig. 1E, P<0.05 vs. Nav1.5). In contrast,co-expression of b1 or b3 alone had no significant effecton time to peak current.
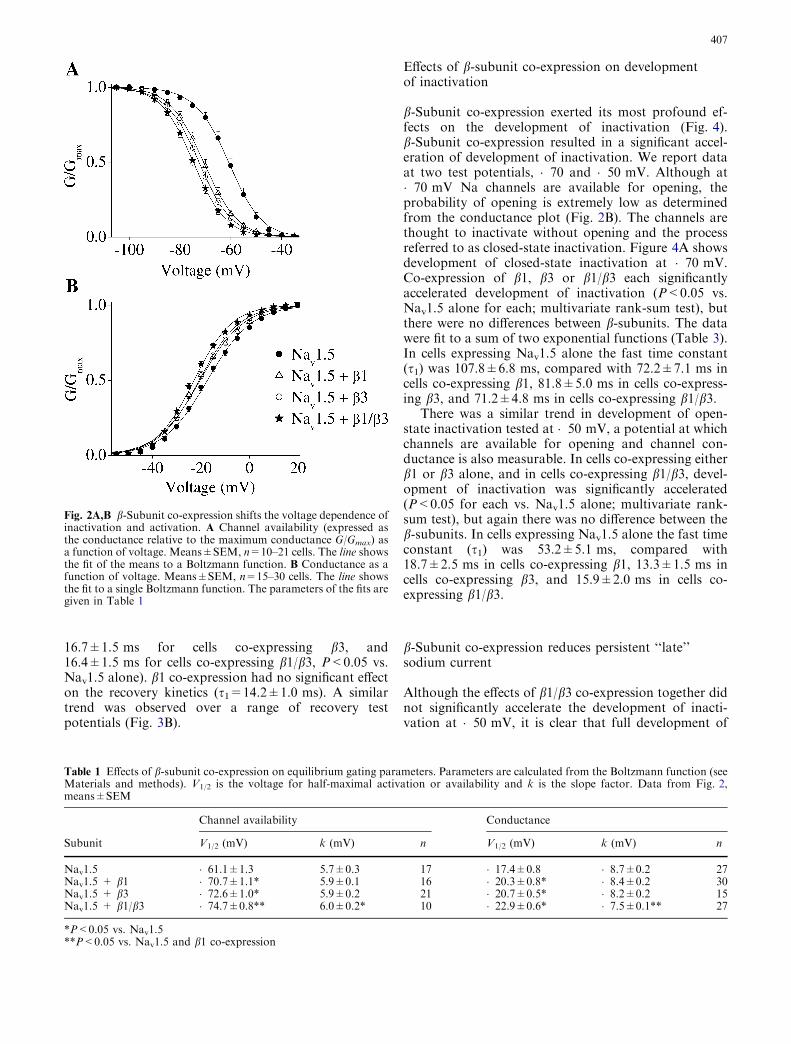
Effects of b-subunit co-expression on equilibrium gating
There were significant shifts in equilibrium gating rela-tionships consequent on b-subunit co-expression (Fig. 2,Table 1). The potential dependence of channel avail-ability was shifted in a hyperpolarizing direction after b-subunit co-expression (Fig. 2A). In comparison withcells expressing Nav1.5 alone, b-subunit co-expressionresulted in a significant hyperpolarizing shift in the po-tential of half maximal inactivation (V1/2, Table 1). Incells co-expressing b1/b3 together the shift in V1/2 wasenhanced. This effect was significant for the comparison
with cells co-expressing b1 alone but not for cells co-expressing b3 (Table 1). In addition, co-expression ofb1/b3 caused a small but significant increase in the slopefactor k (Table 1).
There were similar hyperpolarizing shifts in conduc-tance as a function of potential. In comparison withcells expressing Nav1.5 alone, co-expression of b1, b3and b1/b3 each resulted in a significant hyperpolarizingshift in V1/2 (P<0.05, Fig. 2B, Table 1). Co-expressionof b1/b3 together appeared to result in an enhancedhyperpolarizing shift in V1/2, but this was not significant(P=0.06). There was also a significant reduction in theslope factor k in cells co-expressing b1/b3.
b-Subunit co-expression slows the rate of recoveryfrom inactivation
The effects of b subunit co-expression on recovery frominactivation are shown in Fig. 3. Figure 3A shows anexample of recovery kinetics at �90 mV, close to theanticipated resting potential of a ventricular myocyte.Recovery kinetics were fitted to a sum of two exponen-tial functions (Table 2). Co-expression of both b3 aloneand b1/b3 in combination significantly slowed the fastertime constant (s1) of recovery from inactivation (s1 was10.8±1.1 ms for cells expressing Nav1.5 alone,
Fig. 1C–E (Contd.)
406
16.7±1.5 ms for cells co-expressing b3, and16.4±1.5 ms for cells co-expressing b1/b3, P<0.05 vs.Nav1.5 alone). b1 co-expression had no significant effecton the recovery kinetics (s1=14.2±1.0 ms). A similartrend was observed over a range of recovery testpotentials (Fig. 3B).
Effects of b-subunit co-expression on developmentof inactivation
b-Subunit co-expression exerted its most profound ef-fects on the development of inactivation (Fig. 4).b-Subunit co-expression resulted in a significant accel-eration of development of inactivation. We report dataat two test potentials, �70 and �50 mV. Although at�70 mV Na channels are available for opening, theprobability of opening is extremely low as determinedfrom the conductance plot (Fig. 2B). The channels arethought to inactivate without opening and the processreferred to as closed-state inactivation. Figure 4A showsdevelopment of closed-state inactivation at �70 mV.Co-expression of b1, b3 or b1/b3 each significantlyaccelerated development of inactivation (P<0.05 vs.Nav1.5 alone for each; multivariate rank-sum test), butthere were no differences between b-subunits. The datawere fit to a sum of two exponential functions (Table 3).In cells expressing Nav1.5 alone the fast time constant(s1) was 107.8±6.8 ms, compared with 72.2±7.1 ms incells co-expressing b1, 81.8±5.0 ms in cells co-express-ing b3, and 71.2±4.8 ms in cells co-expressing b1/b3.
There was a similar trend in development of open-state inactivation tested at �50 mV, a potential at whichchannels are available for opening and channel con-ductance is also measurable. In cells co-expressing eitherb1 or b3 alone, and in cells co-expressing b1/b3, devel-opment of inactivation was significantly accelerated(P<0.05 for each vs. Nav1.5 alone; multivariate rank-sum test), but again there was no difference between theb-subunits. In cells expressing Nav1.5 alone the fast timeconstant (s1) was 53.2±5.1 ms, compared with18.7±2.5 ms in cells co-expressing b1, 13.3±1.5 ms incells co-expressing b3, and 15.9±2.0 ms in cells co-expressing b1/b3.
b-Subunit co-expression reduces persistent ‘‘late’’sodium current
Although the effects of b1/b3 co-expression together didnot significantly accelerate the development of inacti-vation at �50 mV, it is clear that full development of
Table 1 Effects of b-subunit co-expression on equilibrium gating parameters. Parameters are calculated from the Boltzmann function (seeMaterials and methods). V1/2 is the voltage for half-maximal activation or availability and k is the slope factor. Data from Fig. 2,means±SEM
Channel availability Conductance
Subunit V1/2 (mV) k (mV) n V1/2 (mV) k (mV) n
Nav1.5 �61.1±1.3 5.7±0.3 17 �17.4±0.8 �8.7±0.2 27Nav1.5 + b1 �70.7±1.1* 5.9±0.1 16 �20.3±0.8* �8.4±0.2 30Nav1.5 + b3 �72.6±1.0* 5.9±0.2 21 �20.7±0.5* �8.2±0.2 15Nav1.5 + b1/b3 �74.7±0.8** 6.0±0.2* 10 �22.9±0.6* �7.5±0.1** 27
*P<0.05 vs. Nav1.5**P<0.05 vs. Nav1.5 and b1 co-expression
Fig. 2A,B b-Subunit co-expression shifts the voltage dependence ofinactivation and activation. A Channel availability (expressed asthe conductance relative to the maximum conductance G/Gmax) asa function of voltage. Means±SEM, n=10–21 cells. The line showsthe fit of the means to a Boltzmann function. B Conductance as afunction of voltage. Means±SEM, n=15–30 cells. The line showsthe fit to a single Boltzmann function. The parameters of the fits aregiven in Table 1
407

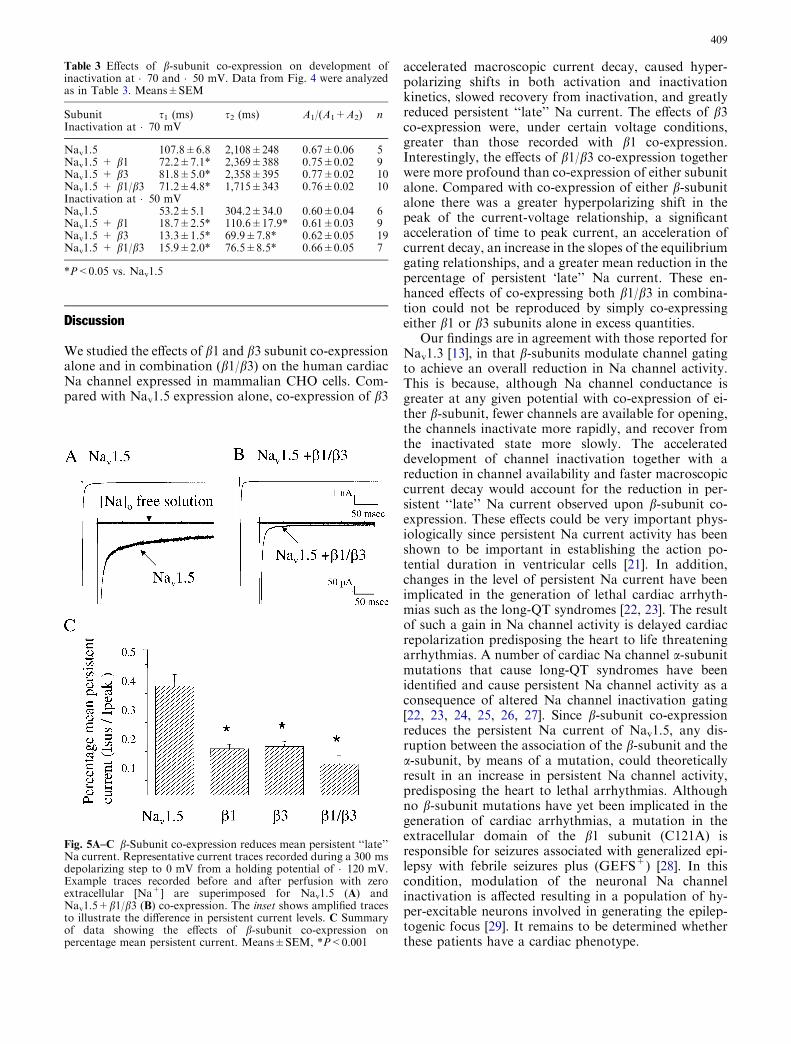
inactivation occurred earlier. In view of this observationand the fact that macroscopic current decay (Fig. 1Aand D) was accelerated by b-subunit co-expression wedetermined the effects of b subunit co-expression onpersistent Na current (Fig. 5). Expression of Nav1.5
alone resulted in a persistent, non-inactivating current at200 ms that amounted to 0.40±0.03% of the peakcurrent (Fig. 5A). This residual ‘‘late’’ current was sig-nificantly reduced upon co-expression of either b1 andb3 alone (0.15±0.01% b1 and 0.16±0.01% b3 respec-tively; P<0.001 vs. Nav1.5 alone). Interestingly, co-expression of b1/b3 together caused a further decrease inthe persistent current to 0.10±0.02% (Fig. 5B;P<0.001 vs. Nav1.5 alone). Mean persistent current isshown in Fig. 5C.
Fig. 3A,B Recovery from inactivation is slowed by b-subunit co-expression. A Effect of b-subunit co-expression on recovery frominactivation (expressed as the current relative to the maximumcurrent I/Imax) at �90 mV. The time course of recovery was fitted(solid lines) to a double exponential function and parameters areshown in Table 2. Recovery at �90 mV is significantly slowed byb3 and b1/b3 co-expression but not b1 co-expression. B Depen-dence of the faster time constant of recovery from inactivation (s1)on test potential. Means±SEM; n=5–18 cells at each potential.*P<0.05 for b3 and b1/b3 vs. Nav1.5
Table 2 Effects of b-subunit co-expression on recovery kinetics at�90 mV. Data from Fig. 3were fitted to a double exponentialfunction (see Materials and methods). s1 and s2 are the time con-stants for the fast and slow gating modes respectively of recovery
and A1 and A2 the corresponding current amplitudes. The pro-portion of channels recovering with the fast time constant wascalculated as A1/(A1+A2). Means±SEM
Fig. 4A,B Development of inactivation is greatly accelerated byb-subunit co-expression. Development of inactivation was deter-mined at �70 mV (n=5–12 cells, A) and �50 mV (n=6–15 cells,B). Means±SEM; the solid lines represent fits to a doubleexponential function
Subunit s1 (ms) s2 (ms) A1/(A1+A2) n
Nav1.5 10.8±1.1 65.0±11.2 0.71±0.05 13Nav1.5 + b1 14.2±1.0 101.6±18.0 0.81±0.02 19Nav1.5 + b3 16.7±1.5* 95.3±11.7 0.84±0.02 21Nav1.5 + b1/b3 16.4±1.5* 94.3±16.9 0.79±0.02 16
*P<0.05 vs. Nav1.5
408
Discussion
We studied the effects of b1 and b3 subunit co-expressionalone and in combination (b1/b3) on the human cardiacNa channel expressed in mammalian CHO cells. Com-pared with Nav1.5 expression alone, co-expression of b3
accelerated macroscopic current decay, caused hyper-polarizing shifts in both activation and inactivationkinetics, slowed recovery from inactivation, and greatlyreduced persistent ‘‘late’’ Na current. The effects of b3co-expression were, under certain voltage conditions,greater than those recorded with b1 co-expression.Interestingly, the effects of b1/b3 co-expression togetherwere more profound than co-expression of either subunitalone. Compared with co-expression of either b-subunitalone there was a greater hyperpolarizing shift in thepeak of the current-voltage relationship, a significantacceleration of time to peak current, an acceleration ofcurrent decay, an increase in the slopes of the equilibriumgating relationships, and a greater mean reduction in thepercentage of persistent ‘late’’ Na current. These en-hanced effects of co-expressing both b1/b3 in combina-tion could not be reproduced by simply co-expressingeither b1 or b3 subunits alone in excess quantities.
Our findings are in agreement with those reported forNav1.3 [13], in that b-subunits modulate channel gatingto achieve an overall reduction in Na channel activity.This is because, although Na channel conductance isgreater at any given potential with co-expression of ei-ther b-subunit, fewer channels are available for opening,the channels inactivate more rapidly, and recover fromthe inactivated state more slowly. The accelerateddevelopment of channel inactivation together with areduction in channel availability and faster macroscopiccurrent decay would account for the reduction in per-sistent ‘‘late’’ Na current observed upon b-subunit co-expression. These effects could be very important phys-iologically since persistent Na current activity has beenshown to be important in establishing the action po-tential duration in ventricular cells [21]. In addition,changes in the level of persistent Na current have beenimplicated in the generation of lethal cardiac arrhyth-mias such as the long-QT syndromes [22, 23]. The resultof such a gain in Na channel activity is delayed cardiacrepolarization predisposing the heart to life threateningarrhythmias. A number of cardiac Na channel a-subunitmutations that cause long-QT syndromes have beenidentified and cause persistent Na channel activity as aconsequence of altered Na channel inactivation gating[22, 23, 24, 25, 26, 27]. Since b-subunit co-expressionreduces the persistent Na current of Nav1.5, any dis-ruption between the association of the b-subunit and thea-subunit, by means of a mutation, could theoreticallyresult in an increase in persistent Na channel activity,predisposing the heart to lethal arrhythmias. Althoughno b-subunit mutations have yet been implicated in thegeneration of cardiac arrhythmias, a mutation in theextracellular domain of the b1 subunit (C121A) isresponsible for seizures associated with generalized epi-lepsy with febrile seizures plus (GEFS+) [28]. In thiscondition, modulation of the neuronal Na channelinactivation is affected resulting in a population of hy-per-excitable neurons involved in generating the epilep-togenic focus [29]. It remains to be determined whetherthese patients have a cardiac phenotype.
Table 3 Effects of b-subunit co-expression on development ofinactivation at �70 and �50 mV. Data from Fig. 4 were analyzedas in Table 3. Means±SEM
Subunit s1 (ms) s2 (ms) A1/(A1+A2) nInactivation at �70 mV
Nav1.5 107.8±6.8 2,108±248 0.67±0.06 5Nav1.5 + b1 72.2±7.1* 2,369±388 0.75±0.02 9Nav1.5 + b3 81.8±5.0* 2,358±395 0.77±0.02 10Nav1.5 + b1/b3 71.2±4.8* 1,715±343 0.76±0.02 10Inactivation at �50 mVNav1.5 53.2±5.1 304.2±34.0 0.60±0.04 6Nav1.5 + b1 18.7±2.5* 110.6±17.9* 0.61±0.03 9Nav1.5 + b3 13.3±1.5* 69.9±7.8* 0.62±0.05 19Nav1.5 + b1/b3 15.9±2.0* 76.5±8.5* 0.66±0.05 7
*P<0.05 vs. Nav1.5
Fig. 5A–C b-Subunit co-expression reduces mean persistent ‘‘late’’Na current. Representative current traces recorded during a 300 msdepolarizing step to 0 mV from a holding potential of �120 mV.Example traces recorded before and after perfusion with zeroextracellular [Na+] are superimposed for Nav1.5 (A) andNav1.5+b1/b3 (B) co-expression. The inset shows amplified tracesto illustrate the difference in persistent current levels. C Summaryof data showing the effects of b-subunit co-expression onpercentage mean persistent current. Means±SEM, *P<0.001
409

Studies designed to determine the subcellular locali-zation pattern of Nav1.5 and b-subunits in cardiacmyocytes have been inconclusive. Nav1.5 has been de-tected in both the intercalated disks and the t-tubules ofthe rat ventricular myocyte [30]. This distribution of thechannel would suggest its co-localization with both b1,which has been shown to be present in both the inter-calated disks and the t-tubules [7, 31] and b3, which isthought to only be present in the t-tubules [31]. How-ever, in a more recent study Nav1.5 localization wasshown to be restricted to the intercalated disks, sug-gesting that it most likely couples only to b1. The preciseco-localization of the Nav1.5, b1 and b3 subunits couldbe important in establishing the electrical activity ofcardiac myocytes. The differential modulation of Nav1.5activity by b-subunit co-expression, along with other ionchannels, may be involved in establishing the heteroge-neity of action potential characteristics throughout theheart since both sodium conductance and b subunitexpression pattern vary with cardiac tissue type andlocation [9, 32]. Action potential upstroke velocity andduration differs very considerably in different myocar-dial layers [33]. The prolonged action potential durationin mid-myocardial cells has been attributed to a largelate sodium conductance in these cells [34]. Persistent Nacurrent density has been shown to be variable in sub-epicardial and subendocardial cells compared with mid-myocardial cells. [21, 35]. Interestingly, there is a small,but significant gradient of b3 mRNA expression acrossthe left ventricular wall in sheep heart with greaterexpression of b3 in the endocardium than in the epi-cardium [9]. This, coupled with the fact that b1 is ex-pressed uniformly throughout the myocardium, couldaccount for smaller Na conductance and persistent Nacurrent density in subendocardial cells.
The effects of b1 and b3 co-expression individually inmammalian cells from this study differ from those seen inXenopus oocytes [9]. These differences are likely to be dueto the expression system selected [13]. In non-mammaliansystems, principally Xenopus oocytes, the modulatoryeffects of b-subunits are very different from the effectsseen in this study using mammalian cells. In Xenopusoocytes [9], co-expression of b3 increases current densitythreefold with no effect on current decay kinetics.Interestingly there was a depolarizing shift in channelavailability as a function of potential and an accelerationof recovery from inactivation. This is in direct contrast tothe present findings in a mammalian expression system.Similarly, the effects of b1-subunit co-expression alsovary with the expression system. For example, innumerous studies in Xenopus oocytes, b1-subunit co-expression has elicited some, all, or none of the followingeffects: enhanced current amplitude, acceleration ofinactivation and recovery from inactivation and reduceduse-dependent decay [9, 36, 37, 38, 39, 40].
Interestingly, differences also exist between variousmammalian cell expression systems. For example, inmammalian HEK 293 cells, co-expression of b1 withNav1.5 causes a small effect on current amplitude and
decay rate, but significant depolarizing shifts in channelavailability as a function of potential [11, 41, 42, 43].Additionally, a small but significant enhancement in therate of recovery from inactivation and entry into inac-tivation have also been observed [41]. In our studies, b1co-expression in CHO cells shifted channel availabilityto hyperpolarized instead of depolarized potentials andslowed recovery from inactivation instead of accelerat-ing recovery. These differences between cell lines couldbe explained by the presence of endogenous b-subunitsin HEK 293 cells [44] in comparison to CHO cells, whichdo not to express endogenous subunits [13]. Althoughthe exact reasons for these differences are not known,factors including variations in expression methods used,cell signaling, membrane properties and other interact-ing proteins could be responsible [11].
While b3 and b1 are highly homologous, sharing57% sequence identity [5], the effects of co-expressing b1were less pronounced than those following b3 co-expression. These differences in the effects of the twosubunits could be due to differences within the extra-cellular domain. This domain interacts with the sodiumchannel a-subunit [16] and is the region of greatestvariability between the two auxiliary subunits [5]. In b1,three acidic residues, an aspartic acid flanked by twoglutamic acid residues, are thought to play a major rolein the fast gating of sodium channels [16]. Interestingly,in b3 the aspartic acid residue is replaced by the neutralamino acid proline and this single amino acid changemay account for the different effects of b3 co-expressioncompared with b1 [5]. Alternatively, the differential ef-fects of b3 compared with b1 could be due to differentassociation sites with the pore forming a-subunit. b1 hasbeen studied extensively and its interaction site isthought to be the SS1-SS2 loop between segments 5 and6 of domain IV [14, 15]. Those studies were performedon chimeric channels constructed between Nav1.5 andNav1.4 and also Nav1.5 and Nav1.2. Since similarstudies on b3 have not been performed it is possible that,even with such homology, the interaction site for b3 withthe a-subunit is not the same as for b1. A differentbinding site for b1 and b3 could explain why the greatlyenhanced gating changes associated with co-expressionwith both subunits in combination could not be repro-duced by over expressing either of the subunits in excess.Alternatively, both b-subunits may bind to the a-subunitat the same site either individually or after assembling asdimers resulting in a change in association with the a-subunit and greater modulation of channel gating. Otherpossibilities include a distinct population of a-subunitsassociated with either b1 or b3 subunits.
In essence our data shows first, that b3 is a significantregulator of Nav1.5 in mammalian cells and results insignificant shifts in equilibrium gating and gatingkinetics. Second, the effects of b1 co-expression are lessmarked than the effects of b3 in that b1 co-expressiondoes not accelerate the decay of macroscopic currentssignificantly nor slow recovery from inactivation at�90 mV. Third, the effects of b1/b3 co-expression to-
410

gether are more profound than co-expression of eithersubunit alone, and, finally, b-subunit co-expression re-duces the percentage of persistent ‘‘late’’ Na current.
It remains to be determined whether the subunitsbind at distinct sites on the Na channel, form dimers orcompete for the same binding site. However, since it hasbeen shown that in cardiac ventricles [9] and regions ofthe brain such as the hippocampus [5], both b1 and b3co-exist the differential effects of both subunits could beof fundamental importance for the modulation ofexcitability in these cells. Important differences in themodulatory effects of b-subunits predicate a re-inter-pretation of the significance of b-subunit modulation ofNa channels in the heart.
Acknowledgements We thank Dr. A.O. Grant (Duke University,USA) for providing the Nav1.5 stably transfected CHO cells, Dr.A. Jackson, (University of Cambridge, UK) for the b3-subunitclone, Dr. J. Randall Moorman and Dr. Carl Lynch III for helpfulcomments. This work was supported by a grant from the AmericanHeart Association (Virginia Affiliate; M.K.P and S-H.K) andKorean Science and Engineering Foundation (S-H.K). We alsothank the Cardiovascular Research Center, University of Virginia.We declare that there is no conflict of interest.
References
1. Catterall WA (2000) From ionic currents to molecular mech-anisms: the structure and function of voltage-gated sodiumchannels. Neuron 26:13–25
2. Goldin AL (2001) Resurgence of sodium channel research.Annu Rev Physiol 63:871–894
3. Isom LL, De Jongh KS, Patton DE, Reber BF, Offord J,Charbonneau H, Walsh K, Goldin AL, Catterall WA (1992)Primary structure and functional expression of the beta 1subunit of the rat brain sodium channel, Science 256:839–842
4. Isom LL, Ragsdale DS, De Jongh KS, Westenbroek RE, ReberBF, Scheuer T, Catterall WA (1995) Structure and function ofthe beta 2 subunit of brain sodium channels, a transmembraneglycoprotein with a CAM motif. Cell 83:433–442
5. Morgan K, Stevens EB, Shah B, Cox PJ, Dixon AK, Lee K,Pinnock RD, Hughes J, Richardson PJ, Mizuguchi K, JacksonAP (2000) Beta 3: an additional auxiliary subunit of the volt-age-sensitive sodium channel that modulates channel gatingwith distinct kinetics. Proc Natl Acad Sci USA 97:2308–2313
6. Yu FH, Westenbroek RE, Silos-Santiago I, McCormick KA,Lawson D, Ge P, Ferriera H, Lilly J, DiStefano PS, CatterallWA, Scheuer T, Curtis R (2003) Sodium channel beta4, a newdisulfide-linked auxiliary subunit with similarity to beta2. JNeurosci 23:7577–7585
7. Malhotra DJ, Chen C, Rivolta I, Abriel H, Malhotra R, MatteiLN, Brosius FC, Kass RS, Isom LL (2001) Characterization ofsodium channel a and b subunits in rat and mouse cardiacmyocytes. Circulation 103:1303–1310
8. Isom LL, Catterall WA (1996) Na+ channel subunits and Igdomains. Nature 383:307–308
9. Fahmi AI, Patel M, Stevens EB, Fowden AL, John JE, Lee K,Pinnock R, Morgan K, Jackson AP, Vandenberg JI (2001) Thesodium channel b-subunit SCN3b modulates the kinetics ofSCN5a and is expressed heterogeneously in sheep heart. JPhysiol (Lond) 537:693–700
10. Chen YH, Dale TJ, Romanos MA, Whitaker WRJ, Xie XM,Clare JJ (2000) Cloning, distribution and functional analysis ofthe type III sodium channel from human brain. Eur J Neurosci12:4281–4289
11. Qu Y, Curtis R, Lawson D, Gilbride K, Ge P, DiStefano PS,Silos-Santiago I, Catterall WA, Scheuer T (2001) Differentialmodulation of sodium channel gating and persistent sodiumcurrents by the b1, b2, and b3 subunits. Mol Cell Neurosci18:570–580
12. Meadows L, Malhotra JD, Stetzer A, Isom LL, Ragsdale DS(2001) The intracellular segment of the sodium channel b1subunit is required for its efficient association with the channela subunit. J Neurochem 76:1871–1878
13. Meadows LS, Chen YH, Powell AJ, Clare JJ, Ragsdale DS(2002) Functional modulation of human brain Nam1.3 sodiumchannels, expressed in mammalian cells, by auxiliary b1, b2 andb3 subunits, Neuroscience 114:745–753
14. Makita N, Bennett PB, George ALJ (1996) Molecular deter-minants of b1 subunit-induced gating modulation in voltage-dependent Na+ channels. J Neurosci 16:7117–7127
15. Qu Y, Rogers JC, Chen SF, McCormick KA, Scheuer T,Catterall WA (1999) Functional roles of the extracellular seg-ments of the sodium channel a subunit in voltage-dependentgating and modulation by b1 subunits. J Biol Chem 274:32647–32654
16. McCormick KA, Isom LL, Ragsdale D, Smith D, Scheuer T,Catterall WA (1998) Molecular determinants of Na+ channelfunction in the extracellular domain of the b1 subunit. J BiolChem 273:3954–3962
17. Shah BS, Stevens EB, Pinnock RD, Dixon AK, Lee K (2001)Developmental expression of the novel voltage-gated sodiumchannel auxiliary subunit b3, in rat CNS. J Physiol (Lond)534:763–776
18. Stevens EB, Cox PJ, Shah BS, Dixon AK, Richardson PJ,Pinnock RD, Lee K (2001) Tissue distribution and functionalexpression of the human voltage-gated sodium channel b3subunit. Pflugers Arch 441:481–488
19. Miller JR, Patel MK, John JE, Mounsey JP, Moorman JR(2000) Contributions of charged residues in a cytoplasmiclinking region to Na channel gating. Biochim Biophys Acta1509:275–291
20. O’Brien PC (1984) Procedures for comparing samples withmultiple endpoints. Biometrics 40:1079–1087
21. Sakmann BF, Spindler AJ, Bryant SM, Linz KW, Noble D(2000) Distribution of a persistent sodium current across theventricular wall in guinea pigs. Circ Res 87:910–914
22. Abriel H, Cabo C, Wehrens XH, Rivolta I, Motoike HK,Memmi M, Napolitano C, Priori SG, Kass RS, Novel ar-rhythmogenic mechanism revealed by a long-QT syndromemutation in the cardiac Na+ channel. Circ Res 88:740–745
23. Wang DW, Yazawa K, George AL Jr, Bennett PB (1996)Characterization of human cardiac Na+ channel mutations inthe congenital long QT syndrome, Proc Natl Acad Sci USA93:13200–13205
24. Chen T, Sheets MF (2002) Enhancement of closed-state inac-tivation in long QT syndrome sodium channel mutation DKPQ.Am J Physiol 283:H966–H975
25. Rivolta I, Abriel H, Tateyama M, Liu H, Memmi M, Vardas P,Napolitano C, Priori SG, Kass RS (2001) Inherited Brugadaand long QT-3 syndrome mutations of a single residue of thecardiac sodium channel confer distinct channel and clinicalphenotypes. J Biol Chem 276:30623–30630
26. Kambouris NG, Nuss HB, Johns DC, Marban E, TomaselliGF, Balser JR (2000) A revised view of cardiac sodium channel‘‘blockade’’ in the long-QT syndrome. J Clin Invest 105:1133–1140
27. Kambouris NG, Nuss HB, Johns DC, Tomaselli GF, MarbanE, Balser JR (1998) Phenotypic characterization of a novellong-QT syndrome mutation (R1623Q) in the cardiac sodiumchannel. Circulation 97:640–644
28. Wallace RH, Wang DW, Singh R, Scheffer IE, George AL Jr,Phillips HA, Saar K, Reis A, Johnson EW, Sutherland GR,Berkovic SF, Mulley JC (1998) Febrile seizures and generalizedepilepsy associated with a mutation in the Na+-channel b1subunit gene SCN1B. Nat Genet 19:366–370
411

29. Moran O, Conti F (2001) Skeletal muscle sodium channel isaffected by an epileptogenic b1 subunit mutation. BiochemBiophys Res Commun 282:55–59
30. Cohen SA (1996) Immunocytochemical localization of rH1sodium channel in adult rat heart atria and ventricle. Presencein terminal intercalated disks. Circulation 94:3083–3086
31. Maier SK, Westenbroek RE, McCormick KA, Curtis R,Scheuer T, Catterall WA (2004) Distinct subcellular localiza-tion of different sodium channel a and b subunits in singleventricular myocytes from mouse heart Circulation 109:1421–1427
32. Antzelevitch C, Sicouri S, Litovsky SH, Lukas A, Krishnan SC,Didiego JM, Gintant GA, Liu DW (1991) Heterogeneity withinthe ventricular wall—electrophysiology and pharmacology ofepicardial, endocardial, and M-cells. Circ Res 69:1427–1449
33. Watanabe T, Rautaharju PM, McDonald TF (1985) Ventric-ular action potentials, ventricular extracellular potentials, andthe ECG of guinea pig. Circ Res 57:362–373
34. Zygmunt AC, Eddlestone GT, Thomas GP, Nesterenko VV,Antzelevitch C (2001) Larger late sodium conductance in Mcells contributes to electrical heterogeneity in canine ventricle.Am J Physiol 281:H689–H697
35. Honen BN, Saint DA (2002) Heterogeneity of the properties ofINa in epicardial and endocardial cells of rat ventricle. Clin ExpPharmacol Physiol 29:161–166
36. Nuss HB, Chiamvimonvat N, Perez-Garcia MT, Tomaselli GF,Marban E (1995) Functional association of the b1 subunit withhuman cardiac (hH1) and rat skeletal muscle (l1) sodiumchannel a subunits expressed in Xenopus oocytes. J Gen Physiol106:1171–1191
37. Baroudi G, Carbonneau E, Pouliot V, Chahine M (2000)SCN5A mutation (T1620M) causing Brugada syndromeexhibits different phenotypes when expressed in Xenopusoocytes and mammalian cells. FEBS Lett 467:12–16
38. Wei J, Wang DW, Alings M, Fish F, Wathen M, Roden DM,George AL Jr (1999) Congenital long-QT syndrome caused bya novel mutation in a conserved acidic domain of the cardiacNa+ channel. Circulation 99:3165–3171
39. Qu Y, Isom LL, Westenbroek RE, Rogers JC, Tanada TN,McCormick KA, Scheuer T, Catterall WA (1995) Modulationof cardiac Na+ channel expression in Xenopus oocytes by b1subunits. J Biol Chem 270:25696–25701
40. Makita N, Bennett PB, George AL (1994) Voltage-gated Na+
channel b1 subunit mRNA expressed in adult human skeletalmuscle, heart, and brain is encoded by a single gene. J BiolChem 269:7571–7578
41. Xiao YF, Wright SN, Wang GK, Morgan JP, Leaf A (2000)Coexpression with b1-subunit modifies the kinetics and fattyacid block of hH1a Na+ channels. Am J Physiol 279:H35–H46
42. An RH, Wang XL, Kerem B, Benhorin J, Medina A, GoldmitM, Kass RS (1998) Novel LQT-3 mutation affects Na+ channelactivity through interactions between a- and b1-subunits. CircRes 83:141–146
43. Wright SN, Wang SY, Xiao YF, Wang GK (1999) State-dependent cocaine block of sodium channel isoforms, chimeras,and channels coexpressed with the b1 subunit. Biophys J 76:33–245
44. Moran O, Conti F, Tammaro P (2003) Sodium channel het-erologous expression in mammalian cells and the role of theendogenous beta 1-subunits. Neurosci Lett 336:175–179
412




















