
Blow-up collocation solutions of nonlinear homogeneous Volterra integral equations
Upload
independentCategory
view
1download
0
Modelo de Lotka-Volterra
Alfonso de Miguel
July 1, 2014
Abstract
En este trabajo se plantea un estudio basico de modelos matematicos usadostıpicamente en el estudio de ecologıa y dinamica de poblaciones. Daremos un repasoal archiconocido modelo de Lotka-Volterra desde el punto de vista de la teorıa desistemas dinamicos (determinista). A continuacion se trabajara su deduccion yanalisis considerando los procesos elementales del sistema (procesos de salto) paradeducir una ecuacion maestra del sistema. Compararemos el modelo determinista yeste ultimo, basado en el analisis de las fluctuaciones internas.
1 Introduccion.
La incursion de la fısica y de la matematica en terrenos no convencionales es desde hacelargo tiempo una barrera mas que superada y el uso de herramientas tıpicas de estoscampos tan fundamentales ha de ser siempre bienvenido por su capacidad de aportarnuevos puntos de vista, metodos y conclusiones. La generalidad que ofrece la teorıa desistemas dinamicos, ası como la teorıa de procesos estocasticos, nos permite modelardiversos fenomenos caracterısticos de cada campo de estudio y dotarlos de un cuerpomatematico y analıtico, con el fin de lograr una comprension mas profunda y una mayorcapacidad de prediccion y analisis del fenomeno en cuestion.
En este trabajo me centro en el llamado modelo o ecuaciones de Lotka-Volterra [1],de amplio uso y estudio en ecologıa, biologıa y dinamica de poblaciones en general. Dadosu atractivo, el modelo aparece en numerosos textos y artıculos, bien sea enfocando elanalisis desde el punto de vista de la teorıa de sistemas dinamicos (estabilidad, bifurca-ciones, etc.) o como posible modelo descriptivo de sistemas depredador-presa o de otrotipo de relaciones entre elementos de un ecosistema o sistema de caracter mas general [2].Tambien existen, por supuesto, textos dedicados a un estudio estocastico del modelo,bien sea desde el punto de vista de las fluctuaciones internas del sistema o por causasde perturbaciones externas [3]. Siguiendo la filosofıa del curso lo propio aquı es enfocarla cuestion desde esta ultima perspectiva.
1

Lo que aquı se pretende es realizar un estudio basico del modelo de Lotka-Volterraen dos dimensiones. En la primera seccion se presenta el sistema dinamico y se repasanlas caracterısticas basicas del modelo: construccion del modelo, significado, soluciones,puntos fijos y estabilidad. En la segunda seccion entramos a considerar ya la cuestionestocastica: tomando el modelo como consecuencia de la ocurrencia de una serie de pro-cesos elementales de variable discreta llegaremos a la deduccion de su ecuacion maestra,abordaremos su solucion estacionaria y de ahı deduciremos las ecuaciones macroscopicasde Lotka-Volterra. Compararemos los resultados entre el modelo determinista y el mod-elo basado en las fluctuaciones intrınsecas.
2 Modelos de dinamica poblacional: Lotka-Volterra.
2.1 Construccion de modelos.
Vamos a mostrar o, mejor dicho, a construir el modelo de Lotka-Volterra de depredador-presa paso a paso para ver su razon de ser como modelo descriptivo (y muy simplificado)de la dinamica de poblaciones de un ecosistema 1.
Sea entonces un ecosistema donde habita una unica especie y cuya poblacion (den-sidad) en el habitat designamos por la variable x. Parece razonable postular que laevolucion en la poblacion x de la especie presa en un tiempo t+ dt sera proporcional altamano de la poblacion en t, x(t), estableciendo la igualdad un multiplicador r tal que
dx
dt= rx, (1)
donde uno puede definir r como el numero de nacimientos menos el numero de muertesde miembros de la especie (crecimiento natural), esto es: r es la tasa neta de crecimientoper capita. Este modelo para r > 0 recibe el nombre de modelo de Malthus [6]y, consecuentemente, implica un crecimiento exponencial de la especie; lo cual parecea todas luces irreal. El modelo logıstico o modelo de Verlhust, tambien de granpopularidad, ofrece una vision mas realista introduciendo el concepto o efecto de com-petencia entre miembros de una propia especie en lucha por los recursos limitados queel ecosistema ofrezca. El modelo logıstico se leerıa ası
dx
dt= r
(1− x
K
)x, (2)
donde K es la denominada capacidad de carga 2. La novedad del modelo logıstico es queintroduce un termino cuadratico negativo en x que actua como regulador de la poblacion,
1Para ver una exposicion historica del desarrollo del modelo puede consultarse [4], [5]2La capacidad de carga de una especie biologica en un ambiente es el tamano maximo de poblacion
que el ambiente puede soportar indefinidamente en un periodo determinado, teniendo en cuenta elalimento, agua, habitat, y otros elementos necesarios disponibles en ese ambiente. Mas detalles: http:
//es.wikipedia.org/wiki/Capacidad_de_carga
2

es decir, suprime el crecimiento exponencial incontrolado del modelo de Malthus. Porsupuesto, esta correccion no es la unica manera de describir la saturacion o limitacional crecimiento de una poblacion y dependiendo del problema concreto la modelizacionpuede requerir una funcion diferente [6]; es simplemente un punto de partida. Tampocohay nada que limite a la tasa de crecimiento r como un factor constante, sino que puededepender de la propia poblacion o de otros factores. Ası que, desde una perspectiva masgeneral, uno puede escribir la evolucion en la poblacion de una especie como
dx
dt= ρ(x)x. (3)
Consideremos la situacion donde tenemos una especie depredadora pero no hay presa.En ausencia de especies inferiores en la cadena de alimentaria se supone una extincionexponencial a un ritmo µ, tal que
dy
dt= −µy, (4)
donde y es la densidad de miembros de la especie depredadora.
Ahora introduzcamos ambas especies en el mismo ecosistema. Bajo la premisa deque la relacion entre ambas es de presa-depredador, la evolucion dinamica del numerode miembros de estas especies ya no dependera unicamente de la propia especie sino decomo sea la interaccion con la rival. De este modo, la presencia del depredador tendraun efecto negativo en la tasa de crecimiento per capita de la presa, mientras que eldepredador basa su crecimiento neto en tanto se alimente de la presa. Denotaremos porφ(x, y) a la tasa de desaparicion per capita de la presa debido a la depredacion y a latasa de crecimiento del depredador debido al consumo per capita de la presa como σ(x).Por lo tanto, de manera general, se tendra este sistema de ecuaciones diferenciales
x
x= ρ(x)− φ(x, y), (5)
y
y= σ(x)− µ, (6)
Se asume que tanto φ como σ son funciones suaves de sus argumentos. El coeficienteque acompana a φ(x, y) es negativo, ya que la interaccion presa-depredador sobre lapresa se traduce en un perjuicio, en un decrecimiento del numero de miembros de estaespecie. Por contra, para el depredador esta interaccion es beneficiosa en terminos de sucrecimiento y σ(x) va con signo positivo3 Reajustando los terminos, podrıamos expresardefinitivamente el modelo como
dx
dt= x[ρ(x)− φ(x, y)] = F (x, y), (7)
3De manera general estas funciones, φ(x, y) y σ(x) no tienen porque ser iguales aunque se trate de lamisma interaccion presa-depredador. De todos modos, en lo posterior se asumiran iguales.
3

dy
dt= y[σ(x)− µ] = G(x, y). (8)
que es una version general de modelo de Lotka-Volterra bidimensional de depredador-presa con crecimiento logıstico de la presa. Uno puede bajo esta generalidad, y conintencion de refinar el modelo, seguir aportando condiciones logicas o esperables a satis-facer para las funciones φ(x, y) y σ(x) (vease por ejemplo [7]), pero opto ya por centrarla cuestion en un modelo concreto que sera sobre el que luego haremos el trabajo es-tocastico.
2.2 Lotka-Volterra de presa-depredador.
El modelo concreto de trabajo es este Lotka-Volterra de presa-depredador4:
dx
dt= rx− αyx, (9)
dy
dt= βyx− µy (10)
Donde r es la tasa de crecimiento natural per capita (como en la ecuacion (1)); ω1 es laconstante de acoplo presa-depredador; notese que
φ(x, y) = αy,
σ(x) = βx.
En el caso de la tasa de desaparicion per capita de la presa, funcion φ(x, y), esta esproporcional a la poblacion de depredadores en un tiempo t; el factor α se suele denom-inar eficiencia de captura y es una medida del efecto del depredador en la tasa decrecimiento per capita de la presa. El producto αx representa la llamada respuestafuncional del depredador, esta describe como la tasa de captura de presas viene afec-tada por la abundancia de presas 5. Por otro lado, la tasa de crecimiento del depredadordebido al consumo per capita de la presa, funcion σ(x), es proporcional a la poblacion depresas en un tiempo t. La constante β recibe el nombre de eficiencia de conversion,esto es, la habilidad de los depredadores para convertir presa en tasa de crecimientoper capita adicional para la poblacion de depredadores. El producto βx se denominatambien respuesta numerica de la poblacion depredadora.
Resumamos, para que queden claras las hipotesis del modelo:
- En ausencia de depredeadores, las presas tienen un comportamiento malthusiano.
4Este modelo es el propuesto orginalmente por Volterra en 1926, para explicar las oscilaciones de losbancos de pesca en el mar Adriatico (y por Lotka, independientemente en 1920).
5Existen distintos tipos de respuesta funcional con el fin de modelar el sistema de acuerdo a unasexigencias mas realistas y concretas, vease [8]. En el caso de la ecuacion (9), esta es lineal con x.
4

- El efecto de los depredadores es reducir el numero de presas de forma proporcionala ambas poblaciones.
- En ausencia de presas, los depredadores se extinguen exponencialmente.
- Las presas contribuyen a hacer crecer la poblacion de depredadores de forma propor-cional a su propia poblacion. Esto es una simplificacion considerable de algun mecanismocomplejo que convertirıa carne de presas en hijos de depredadores.
Evidentemente estamos en una situacion muy simplista. Aparte de la no inclusionde terminos logısticos o algun otro tipo de saturacion para modelar un crecimientocontrolado de las poblaciones, se obvian otros detalles importantes como el hecho de quese asume que los depredadores son insaciables y, ademas, cuantas mas presas hay, mascomen. Tambien se da por hecho que todos los depredadores tienen las mismas opcionesde atrapar a sus presas. Hay modelos con difusion, donde se introduce una dependenciaespacial, que intentan aplacar estas cuestiones.
- Puntos fijos: Las nulclinas del sistema son aquellas curvas de pendiente nula, esdecir
0 = x(r − αy) y 0 = y(βx− µ) →Nulclinas de x:
y =r
αy x = 0. (11)
Nulclinas de y:
x =µ
βy y = 0. (12)
La interseccion de las nulclinas de x e y nos dan los puntos fijos del sistema, ası quetenemos:
(x01, y01) = (0, 0), (13)
(x02, y02) =
(µ
β,r
α
). (14)
La primera solucion implica la extincion total de ambas especies. La segunda significauna situacion de coexistencia de ambas especies. Aparte uno tiene la solucion (∞, 0)que acaece en ausencia de depredadores como condicion inicial, y(0) = 0.
- Trayectorias y estabilidad: Para estudiar la estabilidad del sistema es precisolinealizar el sistema de ecuaciones diferenciales en un entorno de los puntos fijos. Pormedio de la evaluacion del jacobiano
J(x, y) =
r − αy −αx
βy βx− µ
, (15)
en los puntos fijos, extraemos los autovalores asociados para conocer la estabilidad decada uno. Para (0, 0) el espectro de J es
σ(J(0, 0)) = r,−µ, (16)
5
Para el sistema presa-depredador r > 0 y µ > 0 y por lo tanto tenemos un autovalorpositivo y otro negativo, estamos ante un punto-silla y por tanto un punto fijo in-estable. Calculando los autovectores asociados se puede ver que las soluciones crecenexponencialmente para x(y) y decrecen exponencialmente para Para (µβ ,
rα) el espectro
de J es
σ
(J
(µ
β,r
α
))= ±i√rµ. (17)
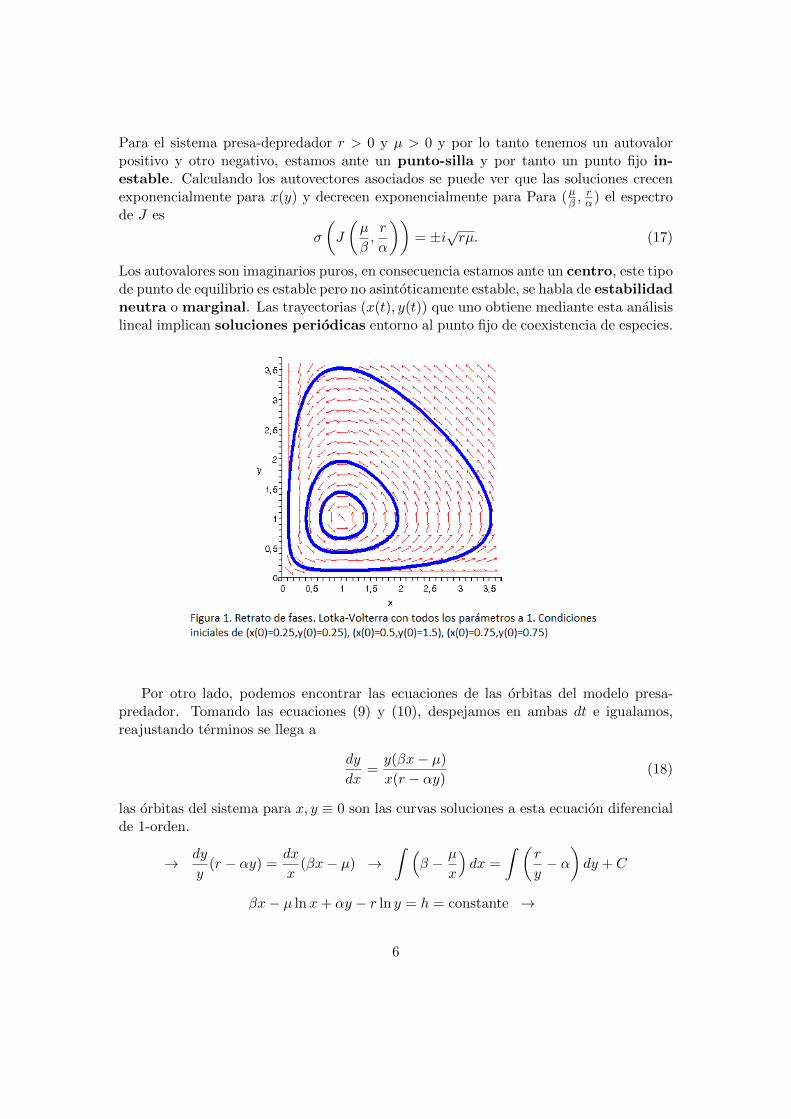
Los autovalores son imaginarios puros, en consecuencia estamos ante un centro, este tipode punto de equilibrio es estable pero no asintoticamente estable, se habla de estabilidadneutra o marginal. Las trayectorias (x(t), y(t)) que uno obtiene mediante esta analisislineal implican soluciones periodicas entorno al punto fijo de coexistencia de especies.
Por otro lado, podemos encontrar las ecuaciones de las orbitas del modelo presa-predador. Tomando las ecuaciones (9) y (10), despejamos en ambas dt e igualamos,reajustando terminos se llega a
dy
dx=y(βx− µ)
x(r − αy)(18)
las orbitas del sistema para x, y ≡ 0 son las curvas soluciones a esta ecuacion diferencialde 1-orden.
→ dy
y(r − αy) =
dx
x(βx− µ) →
∫ (β − µ
x
)dx =
∫ (r
y− α
)dy + C
βx− µ lnx+ αy − r ln y = h = constante →
6

→ eβx
x−µeαy
y−r= H(x, y) = H = constante. (19)
Las orbitas del sistema son la familia de curvas definidas por la ecuacion anterior6 Lafuncion H(x, y) es constante a lo largo de las trayectorias (x(t), y(t)) solucion del sistemade ecuaciones diferenciales de Lotka-Volterra (9-10). La ecuacion (19) muestra quetenemos una integral o constante del movimiento. De hecho, el sistema de Lotka-Volterra puede plantearse como un sistema dinamico Hamiltoniano [9],[10], siendo elpropio H(x, y) el Hamiltoniano de este sistema.
- Conservacion de los promedios: Supongase condiciones iniciales positivasx(0) = x0 > 0 e y(0) = y0 > 0. Si T es el periodo de una orbita cerrada que pasapor (x0, y0), entonces
x
x= r − αy → lnx(t)− lnx(0) =
∫ T
0[r − αy(t)]dt → 0 = rT − α
∫ T
0y(t)dt,
usando la propiedad de periodicidad de las orbitas. Ası que el valor medio sobre unperiodo T es
〈y〉T =1
T
∫ T
0y(t)dt =
r
α, (20)
6Para ver una demostracion de que estamos ante una solucion periodica vease con mas detalle [].
7

Haciendo lo propio para la ecuacion de los depredadores
y
y= βx− µ → ln y(t)− ln y(0) =
∫ T
0[βx(t)− µ]dt → 0 = −µT + β
∫ T
0x(t)dt.
Y analogamente
〈x〉T =1
T
∫ T
0x(t)dt =
µ
β. (21)
El valor mediado de las variables de estado (x(t), y(t)) sobre el periodo de una orbitaarbitraria es igual al valor del punto fijo de coexistencia.
3 Lotka-Volterra a partir de las fluctuaciones internas
El modelo de Lotka-Volterra que hemos revisado en la seccion anterior, y cualquiera desus variaciones mencionadas y referenciadas, son al fin y al cabo sistemas de ecuacionesdiferenciales deterministas y su estudio matematico esta basado en el uso de conceptosy resultados de la teorıa de sistemas dinamicos convencional (ecuaciones diferenciales,teorıa de bifurcaciones, sistemas Hamiltonianos, etc.).
Los modelos tradicionales de ecologıa matematica describen en forma diferencial laevolucion de las densidades. Como se ha visto en la seccion previa, en general, podemos
8

construir estos modelos sin necesidad de conocer al detalle la interaccion entre los indi-viduos que conforman la poblacion. Se supone, no obstante, que cada termino tiene sufundamento en las interacciones puntuales y que representan efectos macroscopicos queinvolucran a muchos individuos, siendo las ecuaciones de evolucion suaves y continuas.Sin embargo, las poblaciones son discretas y las interacciones estan individualizadas.Por medio de un analisis de todos estos procesos de interaccion a escala microscopicadeberıa ser posible la obtencion de las leyes macroscopicas que obtuvimos, surgiendoestas como un lımite macroscopico, un coarse-graining, de los fenomenos observados.Algo que nos ensena la teorıa de fluctuaciones en sistemas dinamicos es esta posibili-dad de obtener ecuaciones deterministas/macroscopicas a partir de una consideracionestocastica del proceso o fenomeno de turno. Es lo que se denomina analizar el sistemadesde el punto de vista de las fluctuaciones internas del sistema.
3.1 Reacciones elementales.
Entremos en materia. Mediante una considerable simplificacion uno puede describir elmodelo de Lotka-Volterra en terminos de reacciones de la siguiente manera:
X +A → 2X, (22)
X + Y → 2Y, (23)
Y → ∅. (24)
Donde cada reaccion se produce a una tasa ki=0,1,2. X e Y siguen designando respec-tivamente a la poblacion de presa y depredador, pero ahora son variables discretas:representan en el numero de miembros de dicha especie. La primera reaccion representael proceso de crecimiento, mas concretamente de autorreplicacion, de la especie presa apartir de una fuente de alimento A que se toma constante7 La segunda reaccion represen-tarıa el proceso de depredacion. La interaccion de una especie presa y una depredadoradan lugar a la desaparicion de una tal presa y al surgimiento de una depredadora8 Porultimo, la reaccion mas realista representa simple y llanamente la muerte de un predador.
Estas reacciones presentadas ocurren con una cierta probabilidad y en consecuencialas variables de estado (X,Y ) ahora son variables estocasticas y su mera especificacionen un instante de tiempo t concreto no aporta la misma informacion que en una situaciondeterminista. Es por ello que debemos trabajar mediante la funcion de probabilidad delsistema: P (X,Y ; t) denotara entonces la probabilidad conjunto de que en un instante de
7Tomar constante la alimentacion de esta especie serıa equivalente a la consideracion de crecimientoexplosivo de la especie a la Malthus. Por otro lado, puede verse aquı que no se plantea ningun tipo dereproduccion sexual ni gestacion sino que sencillamente el organismo X se autorreplica a partir de laingesta de A.
8Como ya mencionamos en la seccion previa, esta hipotesis tambien es muy brusca. Se presuponeuna completa eficiencia y obscuro mecanismo por el cual el depredador transforma a la presa consumidaen un nuevo miembro Y de pleno de derecho.
9

tiempo t haya X miembros de la especie presa e Y miembros de la especie predadora. Laecuacion maestra que buscamos es una ecuacion que nos informa sobre la evolucion tem-poral de esta probabilidad; esta puede ser obtenida al considerar los procesos elementalesque ocurran en un intervalo de tiempo (t, t+dt) que puedan contribuir a P (X,Y ; t+dt),que son
Suceso elemental 1: La poblacion era (X,Y ) en el instante t y no ocurren cambiosdurante dt. No nacen ni mueren ni presas ni depredadores.
Suceso elemental 2: La poblacion era (X − 1, Y ) en el instante t y una presa sereprodujo (tuvo lugar la reaccion primera, ecuacion (22)) durante dt.
Suceso elemental 3: La poblacion era (X,Y + 1) en el instante t y un depredadormurio (reaccion tercera, ecuacion (24)) durante dt.
Suceso elemental 4: La poblacion era (X + 1, Y − 1) en el instante t y un predadordevoro a una presa (reaccion segunda, ecuacion (23)) durante dt.
El hecho de que ocurra una reaccion u otra es independiente. Ademas, siendo p laprobabilidad de que un suceso ocurra, uno se refiere a 1−p como la probabilidad de queun tal suceso no tome lugar. Con todo esto, la probabilidad asociada al primer sucesoelemental es
(1−Xk0dt)(1−XY k1dt)(1− Y k2dt). (25)
Siendo entoncesXk0dt, (26)
la probabilidad de autorreplicacion o reproduccion de la presa X, que es proporcionalal numero de presas X en el instante t y a la constante de reaccion k0 que vendrıa a serun equivalente de la tasa de crecimiento natural de la especie. Mientras que
XY k1dt, (27)
es la probabilidad de ocurrencia de la interaccion depredador-presa, que es proporcionalal numero de presas X y de depredadores Y en el instante t y a la constante de reaccionk1 (la interaccion esta regida por la misma constante de acoplo desde el punto de vistade ambos). Y por ultimo
Y k2dt, (28)
es la probabilidad de muerte de un depredador, y es proporcional al numero de depredadoresen el instante t y a la constante k2, que medirıa la tasa de mortalidad del predador.
Las probabilidades asociadas al resto de sucesos se construirıan de la misma formaa la que siguen las ecuaciones (26-28), teniendo en cuenta el numero de especies quehabrıa en el instante t considerado.
10

3.2 Ecuacion Maestra y solucion estacionaria.
Sabiendo la forma de las probabilidades de transicion, expresarıamos la probabilidadconjunta P (X,Y, t+ dt) como la siguiente suma de contribuciones
P (X,Y ; t+ dt) = P (X,Y ; t)[1−Xk0dt][1−XY k1dt][1− Y k2dt]+
+P (X − 1, Y ; t)(X − 1)k0dt+
+P (X,Y + 1; t)(Y + 1)k2+
+ P (X + 1, Y − 1)(Y − 1)(X + 1)k1. (29)
Cada sumando esta conformado por la probabilidad de estar en un estado (m,n) por laprobabilidad de transicion de ir de un estado (m,n) a otro (m′, n′). Desechando terminosde orden O(dt2) y superiores, dividiendo por dt y tomando el lımite dt → 0 tenemos laecuacion de evolucion para la probabilidad conjunta de X e Y , la ecuacion maestra9:
∂P (X,Y ; t)
∂t= −(Xk0 +XY k1 + Y k2)P (X,Y ; t)+
+(X − 1)k0P (X − 1, Y ; t)+
+(Y + 1)k2P (X,Y + 1; t)+
+ (Y − 1)k1(X + 1)P (X + 1, Y − 1; t). (30)
Para un manejo mas compacto escribimos esta ecuacion por medio de los operadores detraslacion:
∂P (X,Y ; t)
∂t= (E−1X −1)Xk0P (X,Y ; t)+(EY−1)k2Y P (X,Y ; t)+(EXE
−1Y −1)XY k1P (X,Y ; t).
(31)La bidimensionalidad del sistema complica enormemente, si no la hace imposible, laresolucion de la ecuacion maestra a tiempo t. Tampoco intentando la construccion deuna funcion generatriz facilita el problema. La solucion estacionaria verifica que
∂Pst(X,Y )
∂t= 0. (32)
Ası:
0 = (E−1X − 1)Xk0P (X,Y ; t) + (EY − 1)k2Y P (X,Y ; t) + (EXE−1Y − 1)XY k1P (X,Y ; t).
(33)Extraer de aquı la solucion no parace nada claro tampoco pero recurro a un argumentode Haken [11] (recogido tambien en [12],[13]), basado en dos puntos: Primero suponerel ansantz
Pst(X,Y ) = δX,0δY,0. (34)
9Se ha elegido esta forma de construir la ecuacion maestra para enfatizar la consideracion de losprocesos elementales dados. Una forma mas directa para construir las probabilidades de transicion habrıasido seguir el procedimiento usado para las reacciones quımicas [18], por medio de la combinatoria.
11

Sustituyendose en la ecuacion (33), esta se verifica. Pero, ¿es unica esta solucion? Aquıviene el segundo punto: todos los puntos (X,Y ) del sistema estan conectados por almenos una trayectoria con cualquier otro punto (X ′, Y ′) del sistema, lo cual permitesiempre una posibilidad no nula de caer al punto (0, 0) y este estado es absorbente yestable; una vez se alcanza es imposible recuperarse, no se puede crear vida de la nada.Vease la figura 4.
Ahora, este resultado puede parecer inesperado. Nuestro sistema determinista es-tudiado en la seccion primera ofrecıa dos puntos fijos: la solucion de coexistencia y laextincion total. Aquı no hay ni rastro de la primera y la unica perspectiva es ciertamenteapocalıptica. ¿Cual es la explicacion a esta discrepancia entre el modelo estocastico yel determinista? La solucion de existencia era un estado punto-silla (inestable) en elmodelo determinista, pero la de coexistencia implicaba orbitas cerradas en el espacio defases del sistema y el problema radica en su estabilidad. Ası es, obtuvimos una solucionde estabilidad neutra, esto significa que en presencia de fluctuaciones, hecho inevitableen un sistema estocastico, las trayectorias cerradas van a verse perturbadas en mayor omenor medida y a la vista de la solucion estacionaria obtenida, en t→∞ la probabilidadde que haya una fluctuacion que lleve al sistema a la extincion tiende a uno.
Debido a la naturaleza discreta del sistema estocastico, las fluctuaciones en el numerode especies pueden ser determinantes para la extincion si el numero de especies se va ha-ciendo pequeno. Ademas, este resultado significa que cualquier sistema con un numerofinito de partıculas siempre alcanzara el estado de extincion, pero la escala temporalde extincion puede ser muy grande para sistemas razonablemente grandes [12]. Estacaracterıstica esta ausente en el llamado modelo de campo-medio, la descripcion pu-ramente determinista, ya que las densidades de poblacion pueden ser arbitrariamentepequenas sin que el destino de la poblacion se vea afectado dramaticamente. El puntofijo de coexistencia de especies marginalmente estable se convierte en meta-estable enun sistema estocastico. Si bien las fluctuaciones acabaran extinguiendo al sistema, elestado de coexistencia puede ser de larga duracion. En tiempo finito el sistema puedemostrar una dinamica de oscilaciones erraticas que aleje temporalmente al sistema de la
12

solucion estacionaria, retrasandola considerablemente [14]. Segun se afirma tambien en[14], simulaciones de Monte Carlo muestran que las densidades de poblacion se aproxi-man a las densidades de coexistencia a traves de oscilaciones amortiguadas a partir dela condicion inicial.
Ası las cosas, el destino fatal del ecosistema regido por el modelo de Lotka-Volterraparece inevitable. Y aunque esto puede ocurrir en la naturaleza, la simplificacion extremadel modelo obvia algunas cuestiones que puedan dar un vuelco a este final. Es el caso dela introduccion de espacialidad en el sistema, implicando este una posibilidad de refugio oacogedor de flujos migratorios para la supervivencia. Existen resultados [13],[16],[17] quedemuestran que estas modificaciones permiten de nuevo una situacion de coexistenciaentre especies.
Mas detalles sobre un tratamiento del modelp de Lotka-Volterra a partir de lasfluctuaciones intrınsecas puede verse [15],[3].
3.3 Derivacion de las ecuaciones de Lotka-Volterra.
Como conclusion al estudio estocastico interno de este modelo vamos a deducir las ecua-ciones de Lotka-Volterra macroscopicas (deterministas) utilizadas en la seccion primera.Vamos a trabajar dos metodos, el primero a partir de las ecuaciones de evolucion de losmomentos diferenciales y el segundo desde el punto de vista de la aproximacion de VanKampen.
- Momentos diferenciales: Nos interesa obtener las ecuaciones de evolucion delprimer momento y de la varianza (en su version intensiva):
dnidt
= a1(〈ni〉) +1
2a′′1(〈ni〉)σ2ni
, (35)
dσ2ni
dt= a2(〈ni〉) + 2a′1(〈ni〉)σ2ni
, (36)
donde se ha usado la notacion n1 ≡ x, n2 ≡ y por simplicidad. Necesitamos conocer lasmagnitudes ap, a
′′p (con p = 1, 2) para x e y. Se calculan a partir de
ap(X) =∑ρ
rρXω(X + rρX |X), ap(Y ) =∑ρ
rρY ω(Y + rρY |Y ), (37)
el sumatorio se extiende a todas las reacciones. La variable discreta se relaciona con lacontinua mediante ap(X,Y ) = Ωap(x, y), donde Ω es el volumen del sistema. Estos son:
ap(X) = (−1)pk0AX + (−1)pk1XY → ap(x) = (−1)pk0ax+ (−1)pk1xy, (38)
a′p(X) = (−1)pk0A+ (−1)pk1Y → a′p(x) = (−1)pk0a+ (−1)pk1y, (39)
13

a′′p(X) = 0 → a′′p(x) = 0. (40)
ap(Y ) = (−1)pk1xy + (−1)pk2y → ap(y) = (−1)pk1xy + (−1)pk2y, (41)
a′p(Y ) = (−1)pk1x+ (−1)pk2 → ap(y) = (−1)pk1x+ (−1)pk2, (42)
a′′p(Y ) = 0 → ap(y) = 0. (43)
Conk0 ≡ k0Ω, k1 ≡ k1Ω, k2 ≡ k2. (44)
Sustituyendo adecuadamente los resultados de las ecuaciones (38-43) obtenemos el sis-tema
d〈x〉dt
= k0a〈x〉 − k1〈xy〉, (45)
d〈y〉dt
= k1〈xy〉 − k2〈y〉, (46)
dσ2xdt
= k0a〈x〉+ k1〈xy〉+ 2(k0a− k1〈y〉)σ2x, (47)
dσ2ydt
= k1〈xy〉+ k2〈y〉+ 2(k1〈x〉 − k2)σ2y . (48)
Ahora tenemos que dar un paso ciertamente injustificado: suponer que no hay cor-relacion entre la poblacion de presa y de depredadores, de manera que su covarianza seanula, cov(x, y) = 0 y en consecuencia 〈xy〉 = 〈x〉〈y〉. Entonces, considerando que los val-ores medios se aproximan a los deterministas y redefiniendo r ≡ k0a, α = β ≡ k1, µ ≡ k2,tenemos las ecuaciones de Lotka-Volterra presa-depredador obtenidas a partir delas consideracion de las fluctuaciones internas:
d〈x〉dt
= r〈x〉 − α〈x〉〈y〉, (49)
d〈y〉dt
= α〈x〉〈y〉 − µ〈y〉. (50)
Estas ecuaciones bajo esta aproximacion de correlacion nula es lo que se conoce tambiencomo ecuaciones o aproximacion de campo-medio [15]. Vamos a comprobar que,aunque hayamos obtenido el modelo macroscopico, son un tanto problematicas.
En el estado estacionario encontramos los mismos puntos fijos que en el sistemadeterminista:
(〈x〉st, 〈y〉st) = (0, 0), (〈x〉st, 〈y〉st) = (µ/α, r/α). (51)
Sustituyendo los puntos fijos en las ecuaciones (47-48) y haciendo dσ2x/dt = dσ2y/dt = 0obtenemos la varianza para cada punto estacionario. Para el punto de extincion (0, 0)obtenemos que la varianza es nula
σ2x = 0, σ2y = 0. (52)
14

Las fluctuaciones en el punto de extincion son nulas lo cual caracteriza al punto (0, 0)como estable y absorbente. Es imposible escapar de el, no hay fluctuacion que saque alsistema de este sumidero. Este resultado vuelve a estar en discrepancia con la descripcionpuramente determinista pero concuerda perfectamente con la prediccion de la solucionestacionaria de la ecuacion maestra. Si ahora evaluamos para el punto de coexistencia(µ/α, r/α) ambos miembros se anulan identicamente y no podemos obtener informacionsobre la varianza. Evidentemente esto no es bueno y es consecuencia de la aproximacionde covarianza cero. Un analisis mas riguroso exigirıa una hipotesis menos restrictivay nos llevarıa a considerar momentos de orden mayor y en consecuencia a ampliar yenrevesar el sistema de ecuaciones diferenciales.
- Aproximacion de Van Kampen: Es muy instructivo aunque algo tedioso tra-bajar con un sistema bidimensional donde desarrollar la aproximacion de Van Kampen.Es lo que aquı voy a hacer. No entrare en detalles sobre la aproximacion (vease [18]).Lo que sı que es preciso remarcar es que aquı se va a utilizar simplemente como formaalternativa para la deduccion de las ecuaciones macroscopicas sin entrar al analisis de laestabilidad, puesto que este metodo tiene una validez local: describe las fluctuaciones dela variable relevante alrededor de su valor medio. Es por tanto inadecuado para estudiarcuestiones de estabilidad relativa, como es el caso de nuestro sistema, donde tenemosdos puntos fijos de diferente estabilidad.
Vamos a obtener de nuevo las ecuaciones de Lotka-Volterra en el orden macroscopico.Siguiendo a van Kampen [19], en un sistema bidimensional suponemos que las variablesrelevantes son resultado de un termino macroscopico del orden del volumen del sistemaΩ y un termino fluctuante, aleatorio, de orden Ω1/2, ası:
X = Ωφ(t) + Ω1/2ξ, (53)
Y = Ωψ(t) + Ω1/2η, (54)
con la relacion entre probabilidades:
P (X,Y ; t) = Π(ξ, η; t). (55)
Ahora Π(ξ, η; t) designa la probabilidad asociada a las nuevas variables aleatorias, lasfluctuaciones respecto al valor macroscopico de X e Y . Los operadores de traslacionpuede escribirse como
EkX ≈ 1 +k√Ω
∂
∂ξ+k2
2Ω
∂2
∂ξ2+ ... , (56)
EkY ≈ 1 +k√Ω
∂
∂η+k2
2Ω
∂2
∂η2+ ... (57)
Entonces, uno parte de la ecuacion maestra escrita en funcion de los operadores detraslacion, ecuacion (31), y reescribe sus miembros de acuerdo a las nuevas transforma-ciones establecidas. Ası, el miembro izquierdo, el de ∂tP (X,Y ; t), pasarıa a ser
∂P (X,Y ; t)
∂t=∂Π(ξ, η; t)
∂t− Ω1/2dφ
dt
∂Π(ξ, η; t)
∂ξ− Ω1/2dψ
dt
∂Π(ξ, η; t)
∂η. (58)
15

El miembro derecho es mas enrevesado y por no cargar el escrito con largos desarrollosexpondre la situacion inicial y final. Los distintos miembros que tenemos para transfor-mar son
k1XY = k1(Ωφ+ Ω1/2ξ)(Ωψ + Ω1/2η) = k1(Ωφψ + Ω1/2φη + Ω1/2ψξ + ξη), (59)
k0AX = k0A(Ωφ+ Ω1/2ξ) = k0a(Ωφ+ Ω1/2ξ), (60)
k2Y = k2(Ωψ + Ω1/2η) = k2(Ωψ + Ω1/2η), (61)
(EXE−1y − 1) = Ω−1/2∂η −Ω−1/2∂ξ +
1
2Ω−1∂ηη −Ω−1∂ξ∂η −
1
2Ω−1∂ξξ +O(Ω−3/2), (62)
Ahora hay que multiplicar estos resultados de acuerdo a como aparecen en la ecuacionmaestra, es decir
(E−1X − 1)k0XP (X,Y ; t), (63)
(EY − 1)k2Y P (X,Y ; t), (64)
(EXE−1Y − 1)k1XY P (X,Y ; t). (65)
Sustituyendo, por supuesto, ademas P (X,Y ; t) por Π(ξ, η; t). Una vez desarolladosestos miembros (63-65) tenemos que cortar el desarrollo y mantener solo el orden Ω0 yΩ1/2. Posteriormente hay que igualar por potencias de igual orden en Ω con el miembroizquierdo de la ecuacion maestra [ecuacion (58)]. El orden Ω0 nos da la ecuacion deFokker-Planck para la distribucion bivariada de probabilidad de las fluctuaciones y elorden Ω1/2 nos da las ecuaciones macroscopicas/deterministas de Lotka-Volterra. Comono se va a hacer un analisis posterior de las fluctuaciones, nos centramos en el ordenΩ1/2. Ası tenemos:
−dφdt
∂Π
∂ξ− dψ
dt
∂Π
∂η= −k0aφ
∂Π
∂ξ+ k2ψ
∂Π
∂η− k1φψ
∂Π
∂η+ k1φψ
∂Π
∂ξ
Separando en dos ecuaciones, una para ∂ξΠ y otra para ∂ηΠ, llegamos definitivamentea las ecuaciones macroscopicas de Lotka-Volterra:
dφ
dt= k0aφ− k1φψ, (66)
dψ
dt= −k0aφ+ k1φψ. (67)
Donde φ es el valor macroscopico asociado a la presa y ψ es el valor macroscopicoasociado al depredador. Evidentemente, los puntos fijos de este sistema de ecuacionesson exactamente los mismos que los obtenidos en los casos previos: sistema deterministay sistema deducido a partir de los momentos. Si uno calcula ahora los valores medios ymomentos de orden superior, 〈ξ〉, 〈η〉, 〈ξ2〉, 〈η2〉, ... llegara a las mismas conclusiones queen el apartado anterior. Las fluctuaciones entorno al punto de extincion son nulas y nopodemos obtener informacion de los momentos de orden superior respecto al punto decoexistencia.
16

4 Resumen y conclusiones.
En este trabajo se ha hecho un analisis basico del modelo de Lotka-Volterra, de am-plio uso en la teorıa de dinamica de poblaciones. En la primera parte hemos analizadocuestiones basicas propias de la teorıa de sistemas dinamicos determinista. Hemos he-cho enfasis en la edificacion del modelo de Lotka-Volterra de presa-depredador a partirde algun modelo mas simple y de ciertas hipotesis para clarificar su significado. Estu-diamos las soluciones estacionarias del sistema, puntos fijos, y analizamos los tipos desoluciones. El modelo determinista presenta una solucion de coexistencia de especies,matematicamente caracterizada por un punto fijo de estabilidad neutra y dando lugar atrayectorias oscilantes para las poblaciones de presa y depredador. Las otras solucionesimplican la desaparicion de una (el depredador) o de ambas especies, extincion total.Esta solucion es un punto-silla y por tanto inestable. A continuacion se ha propuestola construccion del mismo modelo a partir del tratamiento de las fluctuaciones inter-nas del sistema, es decir, considerando con una cierta probabilidad de ocurrencia losdistintos sucesos elementales que constituyen las interacciones presa-depredador. Se haobtenido la ecuacion maestra para el conjunto de reacciones de Lotka-Volterra y se hahallado la solucion estacionaria. La situacion del estado estacionario respecto al modelodeterminista supone una serie discrepancia: la probabilidad es total para la solucionde extincion total. Las causas son la consideracion de variables discretas en el formal-ismo y la presencia de fluctuaciones importantes que acabaran tarde o temprano conla solucion pseudo-estacionaria de estabilidad neutra, la coexistencia. Finalmente, sehan deducido las ecuaciones macroscopicas de Lotka-Volterra depredador-presa a partirde los momentos diferenciales y de la expansion de van Kampen. El hecho de cuadrarlas ecuaciones obtenidas con las deterministas exige asumir covarianza nula entre presay depredador, una hipotesis injustificada que afecta posteriormente a la obtencion deresultados relativos al punto fijo de coexistencia.
References
[1] Wikipedia, http://en.wikipedia.org/wiki/Lotka%E2%80%93Volterra_equation
[2] Guillermo Abramson, La matematica de los sistemas biologicos. Universidad Na-cional de Cuyo. Centro Atomico Bariloche, Instituto Balseiro y CONICET
[3] H.K. Leung, Stochastic Transients of Noisy Lotka-Volterra Model Chinese Journal ofPhysics. Vol 29, No. 6.
[4] K. Sigmund, Kolmogorov and population dynamics Faculty for Mathematics, Univer-sity of Vienna, and International Institute for Applied Systems Analysis, Austria
[5] G. Belcredi, Modelo presa - depredador
17

[6] L.C.M Miranda, C.A.S. Lima, On the logistic modeling and forecasting of evolution-ary processes: Application to human population dynamics Technological Forecasting& Social Change 77 (2010) 699-771
[7] http://www.ucl.ac.uk/~ucess29/page2/MATH3506Chapters/
/MATH3506Chapter4.pdf
[8] J.H.P. Dawes, M.O. Souza A derivation of Holling’s type I, II and III functionalresponses in predator-prey systems.
[9] C.M. Evans, G.L. Findley Analytic Solutions to the Lotka-Volterra Model For Sus-tained Chemical Oscillations.
[10] S. Baigent Lotka-Volterra Dynamics - An introduction. 2010
[11] H. Haken Synergetics, An Introduction: Nonequilibrium Phase Transitions and Self-Organization in Physics, Chemistry and Biology. Springer-Verlag 1977
[12] U. Dobramysi On the Relaxation Dynamics of Disordered Systems. Dissertation.Doctor of Philosophy in Physics
[13] M. Mobilia, I. T. Georgiev, U. C. Tauber, Phase Transitions and Spatio-TemporalFluctuations in Stochastic Lattice Lotka-Volterra Models. Journal of StatisticalPhysics 128 447-483 (2007).
[14] M. Parker, A. Kamenev Extinction in Lotka-Volterra model 2009.
[15] R. Toral Nonlineal phenomena in biology 2012.
[16] U.C. Tauber Population oscillations in spatial stochastic Lotka-Volterra models: Afield-theoretic perturbational analysis
[17] M.J. Keeling, H.B. Wilson, S.W. Pascala (200) Science 290 1758-1761 y G. MarionStochastic modelling in mathematical biology: Predator-Prey systems and epidemicmodels
[18] J. de la Rubia Fluctuaciones en Sistemas Dinamicos Master en Fısica de SistemasComplejos, UNED
[19] N.G. van Kampen Stochastic Processes in Physics and Chemistry Third edition,2007
18
Copyright © 2022 FDOKUMEN




















