
Prostate Cancer: Basic Mechanisms and Therapeutic ... - ANME
Upload
independentCategory
view
4download
0
INTRODUCTION
CONVINCING EVIDENCE suggests that aberrant regulationof reactive oxygen species (ROS) production and re-
moval is involved in the development of cancer (3 23) Anti-oxidant enzymes scavenge ROS and inhibit the tumor cellmalignant phenotype (24) Manganese superoxide dismutase(MnSOD) is an antioxidant enzyme and a nuclear-encodedmitochondrial matrix protein MnSOD converts superoxidethe f irst ROS generated from one-electron reduction of oxy-gen from cellular respiratory chain to hydrogen peroxide (H2O2)
which is further metabolized to water by catalase or glutathioneperoxidase (24 32) A variety of the tumor cells examinedthus far have been found to exhibit a decrease or loss ofMnSOD expression (3 11 23 31 39) Forced overexpressionof MnSOD inhibits malignant transformation and suppressestumor growth in a variety cancer cells both in vitro and invivo (1 15 19 24) This collection of evidence suggests thatMnSOD is a new type of tumor suppressor gene (3 1223ndash25 29) However the molecular mechanisms by whichMnSOD suppresses the malignant phenotype are still unclearPrevious studies suggest that MnSOD may exert its tumor
677
1Free Radical and Radiation Biology Program 2Department of Radiation Oncology 3Holden Cancer Center University of Iowa Iowa City IA52242
4Vertex Pharmaceuticals Inc Cambridge MA 021395Department of Pharmacology and Toxicology Bone Marrow Transplant Program and the Arizona Cancer Center University of Arizona Tuc-
son AZ 85724
Original Research Communication
MnSOD Up-Regulates Maspin Tumor Suppressor GeneExpression in Human Breast and Prostate Cancer Cells
HONG DUAN1 HANNAH J ZHANG1 JI-QIN YANG4 LARRY W OBERLEY1ndash3
BERNARD W FUTSCHER5 and FREDERICK E DOMANN1ndash3
ABSTRACT
Manganese superoxide dismutase (MnSOD) is an antioxidant enzyme with tumor suppressor activity how-ever the molecular mechanisms of MnSOD antitumor effects remain unclear We hypothesized that MnSODactivity in cancer cells might cause downstream changes in the expression of other tumor suppressor genes Todetermine whether maspin a tumor suppressor gene that inhibits breast cancer cell invasion and metastasismight be a target of MnSOD we forced MnSOD expression in several human breast and prostate cancer celllines by adenovirus-mediated gene transfer and measured maspin mRNA expression Forced expression ofMnSOD caused maspin mRNA to accumulate in a dose-dependent manner in both human breast and prostatecancer cells Normal p53 was not necessary to mediate the effect of MnSOD because MnSOD up-regulatedmaspin in cells that harbor wild-type p53 and in cells that harbor mutant p53 Moreover the effects of MnSODon maspin were not due to demethylation of the maspin promoter Analyses of maspin promoter activity tran-scriptional run-on and mRNA stability showed that maspin mRNA stability was the major mechanism formaspin up-regulation by MnSOD Our findings identify a mechanism underlying MnSOD antitumor effectsand provide evidence to support MnSOD as a genetic therapy in the treatment of human breast and prostatecancers Antioxid Redox Signal 5 677ndash688
ANTIOXIDANTS amp REDOX SIGNALINGVolume 5 Number 5 2003copy Mary Ann Liebert Inc
suppressor activity through modulation of redox-sensitive on-cogenes and transcription factors (13) Nevertheless directevidence that relates MnSOD overexpression to other cellphenotype-related genes especially tumor suppressor genesis beginning to be addressed but still lacking (10) Li et alpreviously reported that transfection of MnSOD cDNA intoMCF-7 breast cancer cells reactivated mRNA expression ofthe tumor suppressor gene maspin (16) a bona fide tumorsuppressor gene in human breast and prostate cancer cells(34 36) More recently Zou et al reported a rapid and robustinduction of maspin expression in prostate cancer cells(LNCaP DU145 and PC3) and breast tumor cell MCF-7 fol-lowing wild-type p53 expression from an adenovirus p53 ex-pression vector (41) Based on these studies we hypothesizedthat overexpression of MnSOD may up-regulate maspin ex-pression not only in breast cancer cells but also in prostatecancer cells and functional p53 might be a factor that worksbetween MnSOD and maspin Our results indicate that forcedMnSOD overexpression induces expression of maspin in amanner that appears independent of p53 status
Maspin (mammary serine protease inhibitor) is a 42-kDaprotein and a unique member of the serpin family (27 40)Maspin is abundantly expressed in normal human mammaryand prostate epithelial cells but down-regulated in primarymammary and prostate cancer cells and lost in advanced can-cer cell lines (27 35 40) Forced expression of maspin hasbeen found to inhibit breast and prostate tumor formation cellmotility invasion and metastasis (28 40) Recently it wasfound that maspin works as an effective inhibitor of angio-genesis (37) Maspin has been classified as a class II tumorsuppressor gene because no genetic mutations deletions orrearrangements have been found in maspin in association withits loss of expression We previously found that silencing ofmaspin in breast cancer cells was closely associated with itspromoter methylation status (6) and that this methylation pat-tern was comparable to that in normal cell types that werenegative for maspin expression (8) We were interested to de-termine whether the effect of MnSOD on maspin expressionin breast tumor cells was mediated through demethylation ofthe maspin promoter Therefore we measured the maspin pro-moter methylation status before and after Ad-MnSOD (re-combinant adenovirus containing MnSOD cDNA) infectionin human breast cancer MDA-MB-435 cells which is a maspin-negative cell line and contains a highly methylated maspinpromoter
The balance between mRNA synthesis and degradation de-termines the steady-state level of an individual mRNA speciesDifferential maspin expression in normal and carcinoma-derived prostate and mammary epithelial cells has been shownto be due to differences in transcriptional control (34 35) Toexplore the mechanism(s) involved in the MnSOD effect onmaspin mRNA accumulation we studied the effect of MnSODon maspin transcription initiation rate in vitro by nuclear run-on analysis and maspin promoter activity by promoter-reporteranalysis Recently the posttranscriptional regulation of mRNAstability has emerged as an important control mechanism ofgene expression Therefore we also assessed the effect ofMnSOD on maspin mRNA stability Our results indicated thatincreasing maspin mRNA stability but not transcriptional ac-tivity contributes to up-regulation of maspin by MnSOD
MATERIALS AND METHODS
Cell culture
Human breast cancer cell lines MDA-MB-435 MDA-MB-231 ZR-75ndash1 MCF-7 and normal immortalized humanmammary epithelial cell MCF-10A were obtained from Amer-ican Type Culture Collection (Rockville MD USA) MDA-MB-435 and MDA-MB-231 cells were maintained in RPMI1640 containing 10 fetal bovine serum supplemented with50 microgml gentamycin ZR-75ndash1 was cultured in RPMI 1640with 10 fetal bovine serum supplemented with 100 unitsmlpenicillin and 50 microgml streptomycin MCF-10A cells weregrown in mammary epithelial growth medium (Clonetics Walk-ersville MD USA) supplemented with 100 ngml choleratoxin (CalbiochemndashNovabiochem Corp La Jolla CA USA)MCF-7 cells were cultured in Eaglersquos minimum essential mediumcontaining nonessential amino acids 1 mM sodium pyruvateand 10 fetalbovine serum MCF-7 cell derived stable trans-fectants SOD15 Mn1 Mn40 and neomycin transfection con-trol cell line Neo were maintained in the mediumsupple-mented with 400 microgml geneticin (Life Technologies IncGaithersburg MD USA) Prostate cancer cell lines LNCaPPC3 and DU145 were kindly provided by Dr Michael Cohenat the University of Iowa These three prostate cancer cellswere maintained in RPMI 1640 medium containing 10 fetalbovine serum supplemented with 100 unitsml penicillin and50 microgml streptomycin All the cells were cultured in a 5CO295 humidified air environment at 37degC Cultures werefed with fresh medium two or three times per week
Adenovirus infections
Replication-deficient recombinant type 5 adenovirus con-taining human MnSOD cDNA (Ad-MnSOD) or bacteria LacZcDNA (Ad-LacZ) were from Gene Transfer Vector Core inthe University of Iowa A CMV promoter was used to driveexpression of all the cDNAs Cells were subcultured in 60-mm dishes (05ndash1 3 106 cellsdish) the day before infectionin normal growth medium On the day of the infections cellswere washed twice in prewarmed 13 phosphate-buffered sa-line (PBS) and infected with adenovirus in 25 ml of serum-free medium for 24 h Cells were washed twice with 13 PBSand returned to full growth medium After 48 h of recoveryinfected cells were used for the analysis of MnSOD and maspinprotein expression maspin mRNA expression or maspin pro-moter methylation status
Western blot analyses
Cells were scrape-harvested and sonicated in 005 M potas-sium phosphate buffer (pH 78) with two bursts of 20 s eachat maximum power using a Vibra Cell Sonicator equippedwith an ice water-chilled cup horn tip After centrifugation ofthe lysates protein concentrations of the supernatants weredetermined by the Bio-Rad protein assay (Bio-Rad Laborato-ries Hercules CA USA) Ten micrograms (for MnSOD) or20 microg (for maspin) of denatured protein samples was sepa-rated on a 125 sodium dodecyl sulfatendashpolyacrylamide gelelectrophoresis (SDS-PAGE) and subsequently transferred tonitrocellulose membranes For MnSOD immunoblotting blots
678 DUAN ET AL
were sequentially incubated with primary rabbit antiserumagainst human kidney MnSOD and secondary anti-rabbit IgGconjugated with horseradish peroxidase (HRP) as previouslydescribed (33) For maspin immunoblotting blots were se-quentially incubated with mouse monoclonal antibody againsthuman maspin (dilution 1500 Pharmingen San Diego CAUSA) and secondary anti-rabbit IgG conjugated with HRPThe HRP-conjugated antibodies were visualized by exposureof the blot to enhanced chemiluminescence staining (Amer-sham Arlington IL USA) and bands were revealed by ex-posure of x-ray f ilm
Northern blot analysis for maspin mRNA
Total cellular RNA was isolated using GibcoBRL TotalRNA Isolation Reagent (Life Technologies Inc) Twenty mi-crograms of total cellular RNA from each cell line was elec-trophoresed on an agarose gel containing formaldehyde andtransferred to a nylon membrane Nucleic acids were cross-linked to the membranes by UV irradiation and the membraneswere incubated in prehybridization solution (50 formamide103 Denhardtrsquos solution 10 dextran sulfate and 200 microgmlsalmon sperm DNA) for 12 h at 42degC A KpnI-XhoI restric-tion fragment containing a ~11-kb partial human maspin cDNAwas gel-isolated and labeled with [32P]-dCTP using a randomprimed DNA labeling kit following the instructions providedby the supplier (Roche Molecular Biochemicals Indianapo-lis IN USA) The denatured cDNA probe was added to theprehybridization solution and incubated with the blots for 18h at 42degC The membranes were washed twice in 13 salinendashsodium citrate buffer (SSC)1 SDS at 65degC for 20 min Thelabeled mRNA signal was detected and quantified by a Typhoon8600 variable Mode Imager (Molecular Dynamics Sunny-vale CA USA) Each blot was then stripped and rehybridizedwith a glyceraldehyde-3-phosphate dehydrogenase (GADPH)probe to confirmequal loading and transfer of RNA
Isolation of genomic DNA and sodium bisulfitegenomic sequencing of maspin promoter
Cells were treated with DNA lysis buffer (containing 100mM NaCl 10 mM Tris-HCl 50 mM EDTA 1 SDS) andharvested into a 15-ml centrifuge tube Genomic DNA wasthen isolated and purified using routine phenolchloroformisoamyl procedure Five micrograms of genomic DNA wasmodified with sodium bisulfite under conditions previouslydescribed (5) The maspin promoter was amplified from thebisulfite-modified DNA by nested PCR reactions using primersspecific to the bisulfite-modified sequence of the maspinpromoter The first-round forward primer 5rsquo-AAA AGA ATGGAG ATT AGA GTA TTT TTT GTG-3rsquo and reverse primer5rsquo-CCT AAA ATC ACA ATT ATC CTA AAA AAT A-3rsquo al-lowed the amplification of the fragment from 2283 to +181relative to the transcription start site PCR amplifications wereperformed under the following conditions 94degC for 4 min 5cycles of 94degC for 1 min 56degC for 2 min 72degC for 3 minthen 35 cycles of 94degC for 30 s 56degC for 2 min 72degC for 15min and a final extension of 72degC for 10 min One microliterof first PCR product was further used as template in a nestedPCR amplification The nested forward and reverse primers5rsquo-GAA ATT TGT AGT GTT ATT ATT ATT ATA-3rsquo and 5rsquo-
AAA AAC ACA AAA ACC TAA ATA TAA AAA-3rsquo wereused to generate the fragment from 2237 to +133 PCR am-plifications were performed under the following conditions94degC for 4 min 30 cycles of 94degC for 30 s 56degC for 1 min72degC for 15 min and a final extension of 72degC for 10 minFresh nested PCR products were gel-isolated and cloned intothe TA Original vector according to the manufacturerrsquos instruc-tions (Invitrogen Carlsbad CA USA) Twenty-five positiverecombinants from MDA-MB-435 cells and 10 positive re-combinants from adenovirus-infected cells were isolated andplasmids were prepared using a Qiaprep Spin Plasmid Miniprepkit (Qiagen Valencia CA USA) according to the manufac-turerrsquos instructions and sequenced on an ABI automated DNAsequencer The number of methylated CpG at each site wasdivided by the number of clones analyzed to yield the per-centage of methylation
MnSOD expression plasmids maspin reportergene construct and transient transfections
pcDNA3-MnSOD expression vector which contains thefull-length human MnSOD cDNA driven by a CMV promoterwas constructed as described (33) The maspin promoter re-porter construct (pGL3-MP) was made by fusing the maspingene 5rsquo-flanking region from 2396 to +131 relative to thetranscription start site to the pGL3Basic vector (PromegaMadison WI USA) The identity and orientation of the in-sert were verified by DNA sequencing Two micrograms ofeach pGL3-MP pcDNA3-MnSOD was used for transient trans-fections One microgram of pCMV-b-galactosidase vector wasused as a control for normalizing variations in transfection ef-ficiencies within and between experiments The pcDNA3 orpGL3-Basic vectors wereadded to normalize the total amountof DNA transfected in each assay Transfection was done in5 3 105 MDA-MB-435 cells and DU145 cells by using 20 microlof SuperFect transfection reagent (Qiagen) After 4 h of incu-bation in 1 ml of full growth medium transfection mixturewas removed and cells were allowed to recover in full mediumfor an additional 48 h Plasmid DNA was transferred into ZR-75-1 cells by electroporation Ten micrograms of pGL3-MPor pGL3-Basic with 5 microg of pCMV-b-galactosidase was trans-ferred into 5 3 107 cells by a single electric pulse at 260 Vand 1180 microF ZR-75-1 cells were grown in 5 ml of growthmedium in a 60-mm dish after transfection Four hours laterculture medium was changed Cells were grown in fresh mediumfor 48 h All the transfected cells were harvested and lysedusing 200 microl of passive lysis buffer (Promega) The resultantcell lysate was used for luciferase reporter gene assay and b-galactosidase activity assay
Promoter activity assay
Luciferase activity of 20 microl of cell lysate was measuredafter addition of 100 microl of luciferase assay reagent followingthe manufacturerrsquos protocol (Promega) Luciferase activity wasnormalized to b-galactosidase activity from the same amountof extract in each sample The relative maspin promoter activ-ity in each cell type was represented as a fold increase rela-tive to that of pGL3-Basic vector The mean and standard de-viation from multiple plates in two independent transfectionexperiments were used as one datapoint
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 679
Nuclear run-on assay for maspin mRNAin vitro transcription
Five million PC3 cells were infected with 100 MOI (mul-tiplicity of infection) Ad-MnSOD or Ad-LacZ for 24 h andrecovered in full growth medium for another 24 h Cells wereharvested by scraping into 13 PBS and centrifugation at1500 rpm for 2 min The resulting cell pellet was resuspendedin 1 ml of hypotonic buffer [10 mM KCl 10 mM Tris-HCl(pH 75) 15 mM MgCl2 03 M sucrose 025 NP-40 1 mMdithiothreitol and 05 mM phenylmethylsulfonyl fluoride] onice for 15 min and gently disrupted in a Dounce homogenizerLysed cells were centrifuged at 1500 rpm for 15 min and nu-clear pellet was immediately resuspended in 200 microl of tran-scription reaction buffer [50 mM Tris-HCl (pH 75) 01 mMammonium sulfate 18 mM dithiothreitol 18 mM MnCl2 2 microlof RNasin 300 microM each of ATP CTP and GTP and 100 microCiof [32P]-UTP] Transcription was carried out for 30 min atroom temperature and terminated by incubation with 5 microl ofRNase-free DNase and 2 microl of tRNA (20 mgml) for 15 minTwenty microliters of 10 SDS and 2 microl of proteinase K (01mgml) were added and the incubation continued for 45 minat room temperature RNA was isolated using the RNeasy kit(Qiagen) according to the manufacturerrsquos protocol Equalamounts of radioactivity (~3 3 106 cpmml) were used to hy-bridize dot blots with denatured 2 microg of maspin and GAPDHcDNA in hybridization mixture [583 SSC 58 formamide012 SDS 12 mM EDTA 12 mM Tris-HCl (pH 75) 473
Denhardtrsquos 04 mgml yeast tRNA and 04 mgml shearedssDNA] for 72 h Blots were washed twice at 65 degC for 1 h 32Psignals were visualized and quantitated by a PhosphorImager
Maspin mRNA stability assay
To measure the maspin mRNA stability PC3 cells were in-cubated with 10 microgml actinomycin D (Sigma Chemical CoSt Louis MO USA) for the indicated times To study theeffect of MnSOD on maspin mRNA stability cells were in-fected with 100 MOI Ad-MnSOD or Ad-LacZ for 24 h andrecovered for 24 h before the treatment with the same dose ofactinomycin D Total cellular mRNA was harvested at eachtime point and subjected to northern blot analysis
Statistical analysis
Maspin promoter activity data were expressed as the meansof two independent experiments plusmn SD Statistical significanceamong data groups was determined by one-way ANOVA andfurther two-sample t test at p lt 005 level using S-PLUS 2000
RESULTS
Ad-MnSOD increased MnSOD protein expressionin human breast and prostate cancer cells
Adenovirus has proven to be a powerful gene delivery ve-hicle for the expression of transgenes In this study we used atype 5 recombinant adenovirus vector containing the full-lengthhuman MnSOD cDNA driven by a CMV promoter (Ad-MnSOD) to infect human breast and prostate cancer cells
Similarly Ad-LacZ was used as the infection control To ver-ify the expression of MnSOD protein in the studied cells weinfected one breast cancer cell line MDA-MB-435 and oneprostate cancer cell line DU145 with Ad-MnSOD and mea-sured MnSOD protein expression by western blot analysisThe SDS-PAGE western blot results are shown in Fig 1 Asseen in Fig 1 Ad-MnSOD infection led to MnSOD expres-sion detectable as low as 10 MOI and abundantly at 100 MOIin both of the two cell types compared with controls Ad-LacZ as high as 100 MOI did not induce MnSOD protein ex-pression We also observed a band with a higher molecularweight of MnSOD in DU145 cells This is likely the MnSODprotein precursor which has an extra 24 amino acids that areresponsible for the transportation of the precursor to the mito-chondria (30) Previous immunohistochemical studies havedemonstrated the correct mitochondrial localization of thisadenovirally expressed MnSOD (14)
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells
To determine whether overexpression of MnSOD couldup-regulate maspin we performed northern blot analyses tomeasure maspin mRNA expression in a variety of untreatedAd-LacZ-infected and Ad-MnSOD-infected human breast andprostate cancer cells The northern blot results from human breastcancer cells MDA-MB-435 and MDA-MB-231 are shown inFig 2A and C The results from human prostate cancer cellsDU145 PC3 and LNCaP are shown in Fig 3A B and C re-spectively As shown in Fig 2 we observed a dose-dependentincrease of maspin mRNA expression in MDA-MB-435 andMDA-MB-231 cells infected with 50ndash250 MOI Ad-MnSODImportantly MnSOD expression also induced expression of
680 DUAN ET AL
FIG 1 Overexpression of MnSOD protein in Ad-MnSOD-infected human breast cancer cell MDA-MB-435 and humanprostate cancer cell DU145 Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Ten mi-crograms of whole cell lysate was separated by SDS-PAGE andanalyzed by immunoblotting with rabbit antiserum againsthuman MnSOD
the maspin protein in MDA-MB-231 cells as determined bywestern blot analysis shown in Fig 2B The induced maspinmRNA transcript was ~30 kb determined by its location rela-tive to the 18S and 28S RNA Infection with Ad-LacZ did notinduce maspin expression Interestingly in Ad-MnSOD-infectedhuman prostate cancer cells we observed two maspin tran-scripts corresponding to 30 kb and 12 kb respectively Care-ful inspection of the 3rsquo-untranslated region (3rsquo-UTR) of maspincDNA revealed two potential poly(A) signals located at nu-cleotides 1239 to 1244 and nucleotides 2551 to 2555 (Fig3C) relative to the transcription start site These two poly(A)signals apparently lead to the generation of the shorter 12-kband longer 30-kb maspin transcripts Between the two tran-scripts the smaller 12-kb transcript was much more induciblethan the 30-kb transcripts as seen in the northern blots Themechanisms of this change will be discussed later from theperspective of altered mRNA stability
MnSOD up-regulated maspin mRNA expression in human breast and prostate cancer cellsindependent of the methylation status of the maspin promoter
Our previous study showed that aberrant CpG methylationof maspin promoter participated in the silencing of maspin inmost of the studied breast carcinoma cell lines (6) To findout whether the effect of MnSOD on up-regulation of maspinmRNA in human breast cancer cells is through demethylatingthe maspin promoter we examined the cytosine methylationstatus of the maspin 5rsquo regulatory region from 2237 to +133in MDA-MB-435 cells before and after adenovirus infection
To increase the amplification efficiency of bisulfite-modifiedDNA we performed nested PCR reactions The two pairs ofprimers in the maspin promoter region are shown above thearrows in Fig 4A This amplified region contains 19 CpGdinucleotides that are potential sites for cytosine methylationThe bisulfite genomic sequencing results of maspin promotermethylation status in the untreated MDA-MB-435 cell Ad-LacZ-infected cell and Ad-MnSOD-infected cell are shownin Fig 4BndashD As seen in Fig 4B nearly all the 19 CpGs arehypermethylated in MDA-MB-435 cells Neither Ad-LacZ norAd-MnSOD infection changed the maspin promoter methyla-tion pattern under the same conditions that provided maximalmaspin reactivation Therefore the effect of MnSOD on maspinexpression is not due to the demethylation of maspin pro-moter in MDA-MB-435 cells
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 681
FIG 2 Ad-MnSOD infection reactivates maspin mRNA(A) and protein (B) expression in human breast cancer cellsMDA-MB-231 Ad-MnSOD infection also reactivates maspinmRNA expression in MDA-MB-435 human breast carcinomacells (C) Cells were infected with Ad-LacZ or Ad-MnSOD for24 h and recovered for 48 h Twenty micrograms of total RNAwas used for northern blot analysis of maspin mRNA expres-sion GAPDH mRNA expression was used as RNA loadingand transfer control Twenty micrograms of total cellular pro-tein was used for western blot analysis of maspin protein ex-pression
FIG 3 Ad-MnSOD infection up-regulated maspin mRNAexpression in human prostate cancer cells DU145 (A) PC3(B) and LNCaP (C) Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Twenty micrograms oftotal RNA was put to northern blot analysis of maspin mRNAexpression GAPDH mRNA expression was used as RNA load-ing and transfer control (D) Schematic representation of maspin3rsquo-UTR showing the two poly(A) signals
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells independent ofcellular p53 status
Zou et al reported that p53 up-regulated maspin expres-sion in both prostate and breast cancer cells including all threeprostate cancer cells we used here and one breast cancer cell
MCF-7 (41) Thus it is reasonable to hypothesize that MnSODup-regulated maspin in a p53-dependent manner To clarifywhether this was the case we examined the p53 status of thecells that were studied here and the MCF-7 cell which wasstudied previously (16) based on the published literature Re-sults of this analysis are summarized in Table 1 Among thethree breast cancer cell lines MCF-7 harbors wild-type p53
682 DUAN ET AL
FIG 4 Ad-MnSOD did not change the maspin promoter methylation status in human breast cancer MDA-MB-435 cells(A) DNA sequence of maspin 5rsquo regulatory region Two pairs of arrows indicate the four primers for nest PCR The 19 CpG po-tential sites for cytosine methylation are underlined Cells were infected with 100 MOI Ad-LacZ or Ad-MnSOD for 24 h and re-covered for 48 h The cytosine methylation status of maspin in untreated MDA-MB-435 cells Ad-LacZ-infected cells and Ad-MnSOD-infected cells is shown in BndashD
A
B
C
D
whereas MDA-MB-435 and MDA-MB-231 cells harbor mu-tant p53 Among the prostate cancer cell lines LNCaP har-bors wild-type p53 expression whereas DU145 has a mutantp53 expression and PC3 is a p53 null cell line Therefore theup-regulation effect of MnSOD on maspin mRNA expressionwas independent of cellular p53 status
MnSOD did not influence maspin promoter activityor mRNA transcription rate in PC3 cells
Several studies have shown that maspin promoter activityis abolished or down-regulated in human breast and prostatecancer cells compared with their normal counterpart (34 35)To elucidate whether the effect of MnSOD is due to activationof the maspin promoter we measured maspin promoter activ-ity in transient and stable MnSOD overexpression modelsThe maspin promoter luciferase reporter construct pGL3-MPconsists of the 5rsquo regulatory region from 2396 to +131 rela-tive to the maspin transcription start (Fig 5A) This regionwhich contains the binding sites of Ets p53 and AP-1 hasbeen characterized to be necessary and sufficient for the maspintransactivation (34) To test the function of this construct wefirst transfected it into an immortalized but nontumorigenichuman breast epithelial cell line MCF-10A that constitutivelyexpresses a high level of maspin As shown in Fig 5B maspinpromoter activity in MCF-10A cells was higher than the ac-tivity of pGL3-Basic construct Compared with the promoteractivity in normal MCF-10A cells the maspin promoter wasless active in human breast cancer cells MDA-MB-435 andZR-75-1 To determine whether MnSOD up-regulated themaspin promoter we cotransfected pGL3-MP and an MnSODexpression vector pcDNA3-MnSOD into MDA-MB-435 andDU145 cells As shown in Fig 5C MnSOD had no effect onmaspin promoter activity in either of the two cell lines We alsomeasured maspin promoter activity in MnSOD stable trans-fectants SOD15 Mn1 and Mn40 which were derived fromMCF-7 human breast cancer cells (33) The MnSOD enzymeactivities in the stable transfectants are 19 62 and 230 Umgof protein for SOD15 Mn1 and Mn40 respectively How-ever no significant changes were observed in maspin promoteractivity among the three MnSOD overexpressing cell lines(Fig 5D)
To explore further whether MnSOD modulates maspin mRNAexpression at the transcriptional level we measured the maspin
mRNA in vitro transcription rate The nuclear run-on assayresults in the Ad-MnSOD- and Ad-LacZ-infected PC3 cellsare shown in Fig 6 Again we detected an almost equal amountof the newly transcribed maspin mRNA signal in the two groupsThis result indicates that MnSOD has no effect on the rate oftranscriptional initiation of maspin as determined by this invitro transcription assay
MnSOD increased maspin mRNA stability in PC3 cells
Much attention has been focused on the transcriptional reg-ulation of maspin in normal and cancer cells Thus far an ef-fect of maspin mRNA stability has not been explored In thisstudy we examined maspin mRNA stability in PC3 cells whichhave the most abundant endogenous maspin mRNA expres-sion among the cancer cells we used here We inhibited newmaspin mRNA synthesis by treating the cells with 10 microgmlactinomycin D and measured the residual levels of the 30-kband 12-kb maspin transcripts at various time points A repre-sentative northern blot result is shown in Fig 7A Quantita-tive analysis of the maspin half-life based on two independentexperiments is shown in Fig 7B It shows that the half-life ofthe 30-kb transcript was ~20 h and the half-life of 12-kbtranscript was much longer ~30ndash36 h
To determine whether the effect of MnSOD on the up-regulation of maspin mRNA was due to the modulation ofmaspin mRNA stability we infected the PC3 cells with 100MOI Ad-LacZ and Ad-MnSOD then treated cells with actin-omycin D and assessed maspin mRNA levels at each timepoint The northern blot results are shown in Fig 8A Quanti-tative analysis of the maspin mRNA remaining as a functionof time is shown in Fig 8B for the 30-kb transcript and Fig8C for the 12-kb transcript These data suggest that MnSODincreased the stability of both transcripts compared with theAd-LacZ infection control For the 30-kb transcript the resid-ual maspin mRNA dropped to 30 within 36 h In contrastthe 12-kb transcript consistently remained at nearly 80 from12 to 36 hours Therefore the stabilizing effect of MnSODexpression on the 12-kb transcript was more significant thanthat on the 30-kb transcript Taking into account that the 12-kb transcript has a longer half-life it might explain why weobserved a significant accumulation of 12-kb transcript butnot the 30-kb transcript after MnSOD overexpression
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 683
TABLE 1 OVERVIEW OF MASPIN PROMOTER METHYLATION AND P53STATUS IN HUMAN BREAST AND PROSTATE CANCER CELL LINES
Cell types p53 status Promoter methylation status (6)
Breast cancer cellsMDA-MB-435 mt-p53 +MDA-MB-231 mt-p53 +MCF-7 wt-p53 +
Prostate cancer cellsPC3 p53(22) 2DU145 mt-p53 2LNCaP wt-p53 2
mt mutant wt wild type
DISCUSSION
Tumor cells have often been found to display lower levelsof antioxidant enzyme expression than their normal counter-parts For example tumor cells are nearly always low in MnSODand catalase activity and usually low in copperzinc superox-ide dismutase activity whereas glutathione peroxidase activ-ity is variable (3 25 33) MnSOD is a nuclear-encoded mito-chondrial matrix protein and serves a protective function by
684 DUAN ET AL
FIG 5 Maspin promoter activity in human breast andprostate cancer cells (A) Schematic representation of maspinpromoter construct with the important cis-elements labeled inthe shaded areas (B) Maspin promoter activity was down-regulated in human breast cancer cells MCF-10A and MDA-MB-435 cells were transiently transfected with 2 microg of maspinpromoter construct pGL3-MP or pGL3-Basic together with 1microg of pCMV-b-galactosidase as internal control ZR-75ndash1 cellswere transfected by electroporation Ten micrograms of pGL3-MP or pGL3-basic was transfected into 5 3 107 cells with 5 microgof pCMV-b-galactosidase as the internal control The relativeluciferase activity normalized by b-galactosidase activity wasrepresented as the fold increase compared with pGL3-Basicvector activity in each cell types (C) MnSOD did not influencemaspin promoter activity in human breast cancer MDA-MB-435 cells and human prostate cancer DU145 cells Two micro-grams of pcDNA3-MnSOD expression plasmid was cotrans-fected with 2 microg of pGL3-MP The relative maspin promoteractivity in the control cells was designated as 100 (D)Maspin promoter activity in MnSOD transfected cells was rep-resented as a percentage increase compared with control Re-sults shown here are the means and standard deviations fromtwo independent transfection experiments
FIG 6 Influence of MnSOD on the maspin mRNA invitro transcription rate in PC3 cells Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h total RNA was isolated and put into in vitro tran-scription analysis [32P]-UTP-labeled mRNA samples were hy-bridized with slot-blotted 2 microg of maspin and GAPDH cDNA32P signals were detected and quantified by PhosphorImager
FIG 7 Maspin mRNA stability in human prostate cancerPC3 cells (A) Northern blot analysis for maspin mRNA stabil-ity assay Cells were treated with 10 microgml actinomycin DTotal RNA was harvested at the times indicated and processedfor northern blot analysis The mRNA signals were quantifiedby PhosphorImager (B) The remaining maspin mRNA tran-scripts normalized to GAPDH loading control were plottedversus time
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL

suppressor activity through modulation of redox-sensitive on-cogenes and transcription factors (13) Nevertheless directevidence that relates MnSOD overexpression to other cellphenotype-related genes especially tumor suppressor genesis beginning to be addressed but still lacking (10) Li et alpreviously reported that transfection of MnSOD cDNA intoMCF-7 breast cancer cells reactivated mRNA expression ofthe tumor suppressor gene maspin (16) a bona fide tumorsuppressor gene in human breast and prostate cancer cells(34 36) More recently Zou et al reported a rapid and robustinduction of maspin expression in prostate cancer cells(LNCaP DU145 and PC3) and breast tumor cell MCF-7 fol-lowing wild-type p53 expression from an adenovirus p53 ex-pression vector (41) Based on these studies we hypothesizedthat overexpression of MnSOD may up-regulate maspin ex-pression not only in breast cancer cells but also in prostatecancer cells and functional p53 might be a factor that worksbetween MnSOD and maspin Our results indicate that forcedMnSOD overexpression induces expression of maspin in amanner that appears independent of p53 status
Maspin (mammary serine protease inhibitor) is a 42-kDaprotein and a unique member of the serpin family (27 40)Maspin is abundantly expressed in normal human mammaryand prostate epithelial cells but down-regulated in primarymammary and prostate cancer cells and lost in advanced can-cer cell lines (27 35 40) Forced expression of maspin hasbeen found to inhibit breast and prostate tumor formation cellmotility invasion and metastasis (28 40) Recently it wasfound that maspin works as an effective inhibitor of angio-genesis (37) Maspin has been classified as a class II tumorsuppressor gene because no genetic mutations deletions orrearrangements have been found in maspin in association withits loss of expression We previously found that silencing ofmaspin in breast cancer cells was closely associated with itspromoter methylation status (6) and that this methylation pat-tern was comparable to that in normal cell types that werenegative for maspin expression (8) We were interested to de-termine whether the effect of MnSOD on maspin expressionin breast tumor cells was mediated through demethylation ofthe maspin promoter Therefore we measured the maspin pro-moter methylation status before and after Ad-MnSOD (re-combinant adenovirus containing MnSOD cDNA) infectionin human breast cancer MDA-MB-435 cells which is a maspin-negative cell line and contains a highly methylated maspinpromoter
The balance between mRNA synthesis and degradation de-termines the steady-state level of an individual mRNA speciesDifferential maspin expression in normal and carcinoma-derived prostate and mammary epithelial cells has been shownto be due to differences in transcriptional control (34 35) Toexplore the mechanism(s) involved in the MnSOD effect onmaspin mRNA accumulation we studied the effect of MnSODon maspin transcription initiation rate in vitro by nuclear run-on analysis and maspin promoter activity by promoter-reporteranalysis Recently the posttranscriptional regulation of mRNAstability has emerged as an important control mechanism ofgene expression Therefore we also assessed the effect ofMnSOD on maspin mRNA stability Our results indicated thatincreasing maspin mRNA stability but not transcriptional ac-tivity contributes to up-regulation of maspin by MnSOD
MATERIALS AND METHODS
Cell culture
Human breast cancer cell lines MDA-MB-435 MDA-MB-231 ZR-75ndash1 MCF-7 and normal immortalized humanmammary epithelial cell MCF-10A were obtained from Amer-ican Type Culture Collection (Rockville MD USA) MDA-MB-435 and MDA-MB-231 cells were maintained in RPMI1640 containing 10 fetal bovine serum supplemented with50 microgml gentamycin ZR-75ndash1 was cultured in RPMI 1640with 10 fetal bovine serum supplemented with 100 unitsmlpenicillin and 50 microgml streptomycin MCF-10A cells weregrown in mammary epithelial growth medium (Clonetics Walk-ersville MD USA) supplemented with 100 ngml choleratoxin (CalbiochemndashNovabiochem Corp La Jolla CA USA)MCF-7 cells were cultured in Eaglersquos minimum essential mediumcontaining nonessential amino acids 1 mM sodium pyruvateand 10 fetalbovine serum MCF-7 cell derived stable trans-fectants SOD15 Mn1 Mn40 and neomycin transfection con-trol cell line Neo were maintained in the mediumsupple-mented with 400 microgml geneticin (Life Technologies IncGaithersburg MD USA) Prostate cancer cell lines LNCaPPC3 and DU145 were kindly provided by Dr Michael Cohenat the University of Iowa These three prostate cancer cellswere maintained in RPMI 1640 medium containing 10 fetalbovine serum supplemented with 100 unitsml penicillin and50 microgml streptomycin All the cells were cultured in a 5CO295 humidified air environment at 37degC Cultures werefed with fresh medium two or three times per week
Adenovirus infections
Replication-deficient recombinant type 5 adenovirus con-taining human MnSOD cDNA (Ad-MnSOD) or bacteria LacZcDNA (Ad-LacZ) were from Gene Transfer Vector Core inthe University of Iowa A CMV promoter was used to driveexpression of all the cDNAs Cells were subcultured in 60-mm dishes (05ndash1 3 106 cellsdish) the day before infectionin normal growth medium On the day of the infections cellswere washed twice in prewarmed 13 phosphate-buffered sa-line (PBS) and infected with adenovirus in 25 ml of serum-free medium for 24 h Cells were washed twice with 13 PBSand returned to full growth medium After 48 h of recoveryinfected cells were used for the analysis of MnSOD and maspinprotein expression maspin mRNA expression or maspin pro-moter methylation status
Western blot analyses
Cells were scrape-harvested and sonicated in 005 M potas-sium phosphate buffer (pH 78) with two bursts of 20 s eachat maximum power using a Vibra Cell Sonicator equippedwith an ice water-chilled cup horn tip After centrifugation ofthe lysates protein concentrations of the supernatants weredetermined by the Bio-Rad protein assay (Bio-Rad Laborato-ries Hercules CA USA) Ten micrograms (for MnSOD) or20 microg (for maspin) of denatured protein samples was sepa-rated on a 125 sodium dodecyl sulfatendashpolyacrylamide gelelectrophoresis (SDS-PAGE) and subsequently transferred tonitrocellulose membranes For MnSOD immunoblotting blots
678 DUAN ET AL
were sequentially incubated with primary rabbit antiserumagainst human kidney MnSOD and secondary anti-rabbit IgGconjugated with horseradish peroxidase (HRP) as previouslydescribed (33) For maspin immunoblotting blots were se-quentially incubated with mouse monoclonal antibody againsthuman maspin (dilution 1500 Pharmingen San Diego CAUSA) and secondary anti-rabbit IgG conjugated with HRPThe HRP-conjugated antibodies were visualized by exposureof the blot to enhanced chemiluminescence staining (Amer-sham Arlington IL USA) and bands were revealed by ex-posure of x-ray f ilm
Northern blot analysis for maspin mRNA
Total cellular RNA was isolated using GibcoBRL TotalRNA Isolation Reagent (Life Technologies Inc) Twenty mi-crograms of total cellular RNA from each cell line was elec-trophoresed on an agarose gel containing formaldehyde andtransferred to a nylon membrane Nucleic acids were cross-linked to the membranes by UV irradiation and the membraneswere incubated in prehybridization solution (50 formamide103 Denhardtrsquos solution 10 dextran sulfate and 200 microgmlsalmon sperm DNA) for 12 h at 42degC A KpnI-XhoI restric-tion fragment containing a ~11-kb partial human maspin cDNAwas gel-isolated and labeled with [32P]-dCTP using a randomprimed DNA labeling kit following the instructions providedby the supplier (Roche Molecular Biochemicals Indianapo-lis IN USA) The denatured cDNA probe was added to theprehybridization solution and incubated with the blots for 18h at 42degC The membranes were washed twice in 13 salinendashsodium citrate buffer (SSC)1 SDS at 65degC for 20 min Thelabeled mRNA signal was detected and quantified by a Typhoon8600 variable Mode Imager (Molecular Dynamics Sunny-vale CA USA) Each blot was then stripped and rehybridizedwith a glyceraldehyde-3-phosphate dehydrogenase (GADPH)probe to confirmequal loading and transfer of RNA
Isolation of genomic DNA and sodium bisulfitegenomic sequencing of maspin promoter
Cells were treated with DNA lysis buffer (containing 100mM NaCl 10 mM Tris-HCl 50 mM EDTA 1 SDS) andharvested into a 15-ml centrifuge tube Genomic DNA wasthen isolated and purified using routine phenolchloroformisoamyl procedure Five micrograms of genomic DNA wasmodified with sodium bisulfite under conditions previouslydescribed (5) The maspin promoter was amplified from thebisulfite-modified DNA by nested PCR reactions using primersspecific to the bisulfite-modified sequence of the maspinpromoter The first-round forward primer 5rsquo-AAA AGA ATGGAG ATT AGA GTA TTT TTT GTG-3rsquo and reverse primer5rsquo-CCT AAA ATC ACA ATT ATC CTA AAA AAT A-3rsquo al-lowed the amplification of the fragment from 2283 to +181relative to the transcription start site PCR amplifications wereperformed under the following conditions 94degC for 4 min 5cycles of 94degC for 1 min 56degC for 2 min 72degC for 3 minthen 35 cycles of 94degC for 30 s 56degC for 2 min 72degC for 15min and a final extension of 72degC for 10 min One microliterof first PCR product was further used as template in a nestedPCR amplification The nested forward and reverse primers5rsquo-GAA ATT TGT AGT GTT ATT ATT ATT ATA-3rsquo and 5rsquo-
AAA AAC ACA AAA ACC TAA ATA TAA AAA-3rsquo wereused to generate the fragment from 2237 to +133 PCR am-plifications were performed under the following conditions94degC for 4 min 30 cycles of 94degC for 30 s 56degC for 1 min72degC for 15 min and a final extension of 72degC for 10 minFresh nested PCR products were gel-isolated and cloned intothe TA Original vector according to the manufacturerrsquos instruc-tions (Invitrogen Carlsbad CA USA) Twenty-five positiverecombinants from MDA-MB-435 cells and 10 positive re-combinants from adenovirus-infected cells were isolated andplasmids were prepared using a Qiaprep Spin Plasmid Miniprepkit (Qiagen Valencia CA USA) according to the manufac-turerrsquos instructions and sequenced on an ABI automated DNAsequencer The number of methylated CpG at each site wasdivided by the number of clones analyzed to yield the per-centage of methylation
MnSOD expression plasmids maspin reportergene construct and transient transfections
pcDNA3-MnSOD expression vector which contains thefull-length human MnSOD cDNA driven by a CMV promoterwas constructed as described (33) The maspin promoter re-porter construct (pGL3-MP) was made by fusing the maspingene 5rsquo-flanking region from 2396 to +131 relative to thetranscription start site to the pGL3Basic vector (PromegaMadison WI USA) The identity and orientation of the in-sert were verified by DNA sequencing Two micrograms ofeach pGL3-MP pcDNA3-MnSOD was used for transient trans-fections One microgram of pCMV-b-galactosidase vector wasused as a control for normalizing variations in transfection ef-ficiencies within and between experiments The pcDNA3 orpGL3-Basic vectors wereadded to normalize the total amountof DNA transfected in each assay Transfection was done in5 3 105 MDA-MB-435 cells and DU145 cells by using 20 microlof SuperFect transfection reagent (Qiagen) After 4 h of incu-bation in 1 ml of full growth medium transfection mixturewas removed and cells were allowed to recover in full mediumfor an additional 48 h Plasmid DNA was transferred into ZR-75-1 cells by electroporation Ten micrograms of pGL3-MPor pGL3-Basic with 5 microg of pCMV-b-galactosidase was trans-ferred into 5 3 107 cells by a single electric pulse at 260 Vand 1180 microF ZR-75-1 cells were grown in 5 ml of growthmedium in a 60-mm dish after transfection Four hours laterculture medium was changed Cells were grown in fresh mediumfor 48 h All the transfected cells were harvested and lysedusing 200 microl of passive lysis buffer (Promega) The resultantcell lysate was used for luciferase reporter gene assay and b-galactosidase activity assay
Promoter activity assay
Luciferase activity of 20 microl of cell lysate was measuredafter addition of 100 microl of luciferase assay reagent followingthe manufacturerrsquos protocol (Promega) Luciferase activity wasnormalized to b-galactosidase activity from the same amountof extract in each sample The relative maspin promoter activ-ity in each cell type was represented as a fold increase rela-tive to that of pGL3-Basic vector The mean and standard de-viation from multiple plates in two independent transfectionexperiments were used as one datapoint
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 679
Nuclear run-on assay for maspin mRNAin vitro transcription
Five million PC3 cells were infected with 100 MOI (mul-tiplicity of infection) Ad-MnSOD or Ad-LacZ for 24 h andrecovered in full growth medium for another 24 h Cells wereharvested by scraping into 13 PBS and centrifugation at1500 rpm for 2 min The resulting cell pellet was resuspendedin 1 ml of hypotonic buffer [10 mM KCl 10 mM Tris-HCl(pH 75) 15 mM MgCl2 03 M sucrose 025 NP-40 1 mMdithiothreitol and 05 mM phenylmethylsulfonyl fluoride] onice for 15 min and gently disrupted in a Dounce homogenizerLysed cells were centrifuged at 1500 rpm for 15 min and nu-clear pellet was immediately resuspended in 200 microl of tran-scription reaction buffer [50 mM Tris-HCl (pH 75) 01 mMammonium sulfate 18 mM dithiothreitol 18 mM MnCl2 2 microlof RNasin 300 microM each of ATP CTP and GTP and 100 microCiof [32P]-UTP] Transcription was carried out for 30 min atroom temperature and terminated by incubation with 5 microl ofRNase-free DNase and 2 microl of tRNA (20 mgml) for 15 minTwenty microliters of 10 SDS and 2 microl of proteinase K (01mgml) were added and the incubation continued for 45 minat room temperature RNA was isolated using the RNeasy kit(Qiagen) according to the manufacturerrsquos protocol Equalamounts of radioactivity (~3 3 106 cpmml) were used to hy-bridize dot blots with denatured 2 microg of maspin and GAPDHcDNA in hybridization mixture [583 SSC 58 formamide012 SDS 12 mM EDTA 12 mM Tris-HCl (pH 75) 473
Denhardtrsquos 04 mgml yeast tRNA and 04 mgml shearedssDNA] for 72 h Blots were washed twice at 65 degC for 1 h 32Psignals were visualized and quantitated by a PhosphorImager
Maspin mRNA stability assay
To measure the maspin mRNA stability PC3 cells were in-cubated with 10 microgml actinomycin D (Sigma Chemical CoSt Louis MO USA) for the indicated times To study theeffect of MnSOD on maspin mRNA stability cells were in-fected with 100 MOI Ad-MnSOD or Ad-LacZ for 24 h andrecovered for 24 h before the treatment with the same dose ofactinomycin D Total cellular mRNA was harvested at eachtime point and subjected to northern blot analysis
Statistical analysis
Maspin promoter activity data were expressed as the meansof two independent experiments plusmn SD Statistical significanceamong data groups was determined by one-way ANOVA andfurther two-sample t test at p lt 005 level using S-PLUS 2000
RESULTS
Ad-MnSOD increased MnSOD protein expressionin human breast and prostate cancer cells
Adenovirus has proven to be a powerful gene delivery ve-hicle for the expression of transgenes In this study we used atype 5 recombinant adenovirus vector containing the full-lengthhuman MnSOD cDNA driven by a CMV promoter (Ad-MnSOD) to infect human breast and prostate cancer cells
Similarly Ad-LacZ was used as the infection control To ver-ify the expression of MnSOD protein in the studied cells weinfected one breast cancer cell line MDA-MB-435 and oneprostate cancer cell line DU145 with Ad-MnSOD and mea-sured MnSOD protein expression by western blot analysisThe SDS-PAGE western blot results are shown in Fig 1 Asseen in Fig 1 Ad-MnSOD infection led to MnSOD expres-sion detectable as low as 10 MOI and abundantly at 100 MOIin both of the two cell types compared with controls Ad-LacZ as high as 100 MOI did not induce MnSOD protein ex-pression We also observed a band with a higher molecularweight of MnSOD in DU145 cells This is likely the MnSODprotein precursor which has an extra 24 amino acids that areresponsible for the transportation of the precursor to the mito-chondria (30) Previous immunohistochemical studies havedemonstrated the correct mitochondrial localization of thisadenovirally expressed MnSOD (14)
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells
To determine whether overexpression of MnSOD couldup-regulate maspin we performed northern blot analyses tomeasure maspin mRNA expression in a variety of untreatedAd-LacZ-infected and Ad-MnSOD-infected human breast andprostate cancer cells The northern blot results from human breastcancer cells MDA-MB-435 and MDA-MB-231 are shown inFig 2A and C The results from human prostate cancer cellsDU145 PC3 and LNCaP are shown in Fig 3A B and C re-spectively As shown in Fig 2 we observed a dose-dependentincrease of maspin mRNA expression in MDA-MB-435 andMDA-MB-231 cells infected with 50ndash250 MOI Ad-MnSODImportantly MnSOD expression also induced expression of
680 DUAN ET AL
FIG 1 Overexpression of MnSOD protein in Ad-MnSOD-infected human breast cancer cell MDA-MB-435 and humanprostate cancer cell DU145 Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Ten mi-crograms of whole cell lysate was separated by SDS-PAGE andanalyzed by immunoblotting with rabbit antiserum againsthuman MnSOD
the maspin protein in MDA-MB-231 cells as determined bywestern blot analysis shown in Fig 2B The induced maspinmRNA transcript was ~30 kb determined by its location rela-tive to the 18S and 28S RNA Infection with Ad-LacZ did notinduce maspin expression Interestingly in Ad-MnSOD-infectedhuman prostate cancer cells we observed two maspin tran-scripts corresponding to 30 kb and 12 kb respectively Care-ful inspection of the 3rsquo-untranslated region (3rsquo-UTR) of maspincDNA revealed two potential poly(A) signals located at nu-cleotides 1239 to 1244 and nucleotides 2551 to 2555 (Fig3C) relative to the transcription start site These two poly(A)signals apparently lead to the generation of the shorter 12-kband longer 30-kb maspin transcripts Between the two tran-scripts the smaller 12-kb transcript was much more induciblethan the 30-kb transcripts as seen in the northern blots Themechanisms of this change will be discussed later from theperspective of altered mRNA stability
MnSOD up-regulated maspin mRNA expression in human breast and prostate cancer cellsindependent of the methylation status of the maspin promoter
Our previous study showed that aberrant CpG methylationof maspin promoter participated in the silencing of maspin inmost of the studied breast carcinoma cell lines (6) To findout whether the effect of MnSOD on up-regulation of maspinmRNA in human breast cancer cells is through demethylatingthe maspin promoter we examined the cytosine methylationstatus of the maspin 5rsquo regulatory region from 2237 to +133in MDA-MB-435 cells before and after adenovirus infection
To increase the amplification efficiency of bisulfite-modifiedDNA we performed nested PCR reactions The two pairs ofprimers in the maspin promoter region are shown above thearrows in Fig 4A This amplified region contains 19 CpGdinucleotides that are potential sites for cytosine methylationThe bisulfite genomic sequencing results of maspin promotermethylation status in the untreated MDA-MB-435 cell Ad-LacZ-infected cell and Ad-MnSOD-infected cell are shownin Fig 4BndashD As seen in Fig 4B nearly all the 19 CpGs arehypermethylated in MDA-MB-435 cells Neither Ad-LacZ norAd-MnSOD infection changed the maspin promoter methyla-tion pattern under the same conditions that provided maximalmaspin reactivation Therefore the effect of MnSOD on maspinexpression is not due to the demethylation of maspin pro-moter in MDA-MB-435 cells
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 681
FIG 2 Ad-MnSOD infection reactivates maspin mRNA(A) and protein (B) expression in human breast cancer cellsMDA-MB-231 Ad-MnSOD infection also reactivates maspinmRNA expression in MDA-MB-435 human breast carcinomacells (C) Cells were infected with Ad-LacZ or Ad-MnSOD for24 h and recovered for 48 h Twenty micrograms of total RNAwas used for northern blot analysis of maspin mRNA expres-sion GAPDH mRNA expression was used as RNA loadingand transfer control Twenty micrograms of total cellular pro-tein was used for western blot analysis of maspin protein ex-pression
FIG 3 Ad-MnSOD infection up-regulated maspin mRNAexpression in human prostate cancer cells DU145 (A) PC3(B) and LNCaP (C) Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Twenty micrograms oftotal RNA was put to northern blot analysis of maspin mRNAexpression GAPDH mRNA expression was used as RNA load-ing and transfer control (D) Schematic representation of maspin3rsquo-UTR showing the two poly(A) signals
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells independent ofcellular p53 status
Zou et al reported that p53 up-regulated maspin expres-sion in both prostate and breast cancer cells including all threeprostate cancer cells we used here and one breast cancer cell
MCF-7 (41) Thus it is reasonable to hypothesize that MnSODup-regulated maspin in a p53-dependent manner To clarifywhether this was the case we examined the p53 status of thecells that were studied here and the MCF-7 cell which wasstudied previously (16) based on the published literature Re-sults of this analysis are summarized in Table 1 Among thethree breast cancer cell lines MCF-7 harbors wild-type p53
682 DUAN ET AL
FIG 4 Ad-MnSOD did not change the maspin promoter methylation status in human breast cancer MDA-MB-435 cells(A) DNA sequence of maspin 5rsquo regulatory region Two pairs of arrows indicate the four primers for nest PCR The 19 CpG po-tential sites for cytosine methylation are underlined Cells were infected with 100 MOI Ad-LacZ or Ad-MnSOD for 24 h and re-covered for 48 h The cytosine methylation status of maspin in untreated MDA-MB-435 cells Ad-LacZ-infected cells and Ad-MnSOD-infected cells is shown in BndashD
A
B
C
D
whereas MDA-MB-435 and MDA-MB-231 cells harbor mu-tant p53 Among the prostate cancer cell lines LNCaP har-bors wild-type p53 expression whereas DU145 has a mutantp53 expression and PC3 is a p53 null cell line Therefore theup-regulation effect of MnSOD on maspin mRNA expressionwas independent of cellular p53 status
MnSOD did not influence maspin promoter activityor mRNA transcription rate in PC3 cells
Several studies have shown that maspin promoter activityis abolished or down-regulated in human breast and prostatecancer cells compared with their normal counterpart (34 35)To elucidate whether the effect of MnSOD is due to activationof the maspin promoter we measured maspin promoter activ-ity in transient and stable MnSOD overexpression modelsThe maspin promoter luciferase reporter construct pGL3-MPconsists of the 5rsquo regulatory region from 2396 to +131 rela-tive to the maspin transcription start (Fig 5A) This regionwhich contains the binding sites of Ets p53 and AP-1 hasbeen characterized to be necessary and sufficient for the maspintransactivation (34) To test the function of this construct wefirst transfected it into an immortalized but nontumorigenichuman breast epithelial cell line MCF-10A that constitutivelyexpresses a high level of maspin As shown in Fig 5B maspinpromoter activity in MCF-10A cells was higher than the ac-tivity of pGL3-Basic construct Compared with the promoteractivity in normal MCF-10A cells the maspin promoter wasless active in human breast cancer cells MDA-MB-435 andZR-75-1 To determine whether MnSOD up-regulated themaspin promoter we cotransfected pGL3-MP and an MnSODexpression vector pcDNA3-MnSOD into MDA-MB-435 andDU145 cells As shown in Fig 5C MnSOD had no effect onmaspin promoter activity in either of the two cell lines We alsomeasured maspin promoter activity in MnSOD stable trans-fectants SOD15 Mn1 and Mn40 which were derived fromMCF-7 human breast cancer cells (33) The MnSOD enzymeactivities in the stable transfectants are 19 62 and 230 Umgof protein for SOD15 Mn1 and Mn40 respectively How-ever no significant changes were observed in maspin promoteractivity among the three MnSOD overexpressing cell lines(Fig 5D)
To explore further whether MnSOD modulates maspin mRNAexpression at the transcriptional level we measured the maspin
mRNA in vitro transcription rate The nuclear run-on assayresults in the Ad-MnSOD- and Ad-LacZ-infected PC3 cellsare shown in Fig 6 Again we detected an almost equal amountof the newly transcribed maspin mRNA signal in the two groupsThis result indicates that MnSOD has no effect on the rate oftranscriptional initiation of maspin as determined by this invitro transcription assay
MnSOD increased maspin mRNA stability in PC3 cells
Much attention has been focused on the transcriptional reg-ulation of maspin in normal and cancer cells Thus far an ef-fect of maspin mRNA stability has not been explored In thisstudy we examined maspin mRNA stability in PC3 cells whichhave the most abundant endogenous maspin mRNA expres-sion among the cancer cells we used here We inhibited newmaspin mRNA synthesis by treating the cells with 10 microgmlactinomycin D and measured the residual levels of the 30-kband 12-kb maspin transcripts at various time points A repre-sentative northern blot result is shown in Fig 7A Quantita-tive analysis of the maspin half-life based on two independentexperiments is shown in Fig 7B It shows that the half-life ofthe 30-kb transcript was ~20 h and the half-life of 12-kbtranscript was much longer ~30ndash36 h
To determine whether the effect of MnSOD on the up-regulation of maspin mRNA was due to the modulation ofmaspin mRNA stability we infected the PC3 cells with 100MOI Ad-LacZ and Ad-MnSOD then treated cells with actin-omycin D and assessed maspin mRNA levels at each timepoint The northern blot results are shown in Fig 8A Quanti-tative analysis of the maspin mRNA remaining as a functionof time is shown in Fig 8B for the 30-kb transcript and Fig8C for the 12-kb transcript These data suggest that MnSODincreased the stability of both transcripts compared with theAd-LacZ infection control For the 30-kb transcript the resid-ual maspin mRNA dropped to 30 within 36 h In contrastthe 12-kb transcript consistently remained at nearly 80 from12 to 36 hours Therefore the stabilizing effect of MnSODexpression on the 12-kb transcript was more significant thanthat on the 30-kb transcript Taking into account that the 12-kb transcript has a longer half-life it might explain why weobserved a significant accumulation of 12-kb transcript butnot the 30-kb transcript after MnSOD overexpression
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 683
TABLE 1 OVERVIEW OF MASPIN PROMOTER METHYLATION AND P53STATUS IN HUMAN BREAST AND PROSTATE CANCER CELL LINES
Cell types p53 status Promoter methylation status (6)
Breast cancer cellsMDA-MB-435 mt-p53 +MDA-MB-231 mt-p53 +MCF-7 wt-p53 +
Prostate cancer cellsPC3 p53(22) 2DU145 mt-p53 2LNCaP wt-p53 2
mt mutant wt wild type
DISCUSSION
Tumor cells have often been found to display lower levelsof antioxidant enzyme expression than their normal counter-parts For example tumor cells are nearly always low in MnSODand catalase activity and usually low in copperzinc superox-ide dismutase activity whereas glutathione peroxidase activ-ity is variable (3 25 33) MnSOD is a nuclear-encoded mito-chondrial matrix protein and serves a protective function by
684 DUAN ET AL
FIG 5 Maspin promoter activity in human breast andprostate cancer cells (A) Schematic representation of maspinpromoter construct with the important cis-elements labeled inthe shaded areas (B) Maspin promoter activity was down-regulated in human breast cancer cells MCF-10A and MDA-MB-435 cells were transiently transfected with 2 microg of maspinpromoter construct pGL3-MP or pGL3-Basic together with 1microg of pCMV-b-galactosidase as internal control ZR-75ndash1 cellswere transfected by electroporation Ten micrograms of pGL3-MP or pGL3-basic was transfected into 5 3 107 cells with 5 microgof pCMV-b-galactosidase as the internal control The relativeluciferase activity normalized by b-galactosidase activity wasrepresented as the fold increase compared with pGL3-Basicvector activity in each cell types (C) MnSOD did not influencemaspin promoter activity in human breast cancer MDA-MB-435 cells and human prostate cancer DU145 cells Two micro-grams of pcDNA3-MnSOD expression plasmid was cotrans-fected with 2 microg of pGL3-MP The relative maspin promoteractivity in the control cells was designated as 100 (D)Maspin promoter activity in MnSOD transfected cells was rep-resented as a percentage increase compared with control Re-sults shown here are the means and standard deviations fromtwo independent transfection experiments
FIG 6 Influence of MnSOD on the maspin mRNA invitro transcription rate in PC3 cells Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h total RNA was isolated and put into in vitro tran-scription analysis [32P]-UTP-labeled mRNA samples were hy-bridized with slot-blotted 2 microg of maspin and GAPDH cDNA32P signals were detected and quantified by PhosphorImager
FIG 7 Maspin mRNA stability in human prostate cancerPC3 cells (A) Northern blot analysis for maspin mRNA stabil-ity assay Cells were treated with 10 microgml actinomycin DTotal RNA was harvested at the times indicated and processedfor northern blot analysis The mRNA signals were quantifiedby PhosphorImager (B) The remaining maspin mRNA tran-scripts normalized to GAPDH loading control were plottedversus time
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL

were sequentially incubated with primary rabbit antiserumagainst human kidney MnSOD and secondary anti-rabbit IgGconjugated with horseradish peroxidase (HRP) as previouslydescribed (33) For maspin immunoblotting blots were se-quentially incubated with mouse monoclonal antibody againsthuman maspin (dilution 1500 Pharmingen San Diego CAUSA) and secondary anti-rabbit IgG conjugated with HRPThe HRP-conjugated antibodies were visualized by exposureof the blot to enhanced chemiluminescence staining (Amer-sham Arlington IL USA) and bands were revealed by ex-posure of x-ray f ilm
Northern blot analysis for maspin mRNA
Total cellular RNA was isolated using GibcoBRL TotalRNA Isolation Reagent (Life Technologies Inc) Twenty mi-crograms of total cellular RNA from each cell line was elec-trophoresed on an agarose gel containing formaldehyde andtransferred to a nylon membrane Nucleic acids were cross-linked to the membranes by UV irradiation and the membraneswere incubated in prehybridization solution (50 formamide103 Denhardtrsquos solution 10 dextran sulfate and 200 microgmlsalmon sperm DNA) for 12 h at 42degC A KpnI-XhoI restric-tion fragment containing a ~11-kb partial human maspin cDNAwas gel-isolated and labeled with [32P]-dCTP using a randomprimed DNA labeling kit following the instructions providedby the supplier (Roche Molecular Biochemicals Indianapo-lis IN USA) The denatured cDNA probe was added to theprehybridization solution and incubated with the blots for 18h at 42degC The membranes were washed twice in 13 salinendashsodium citrate buffer (SSC)1 SDS at 65degC for 20 min Thelabeled mRNA signal was detected and quantified by a Typhoon8600 variable Mode Imager (Molecular Dynamics Sunny-vale CA USA) Each blot was then stripped and rehybridizedwith a glyceraldehyde-3-phosphate dehydrogenase (GADPH)probe to confirmequal loading and transfer of RNA
Isolation of genomic DNA and sodium bisulfitegenomic sequencing of maspin promoter
Cells were treated with DNA lysis buffer (containing 100mM NaCl 10 mM Tris-HCl 50 mM EDTA 1 SDS) andharvested into a 15-ml centrifuge tube Genomic DNA wasthen isolated and purified using routine phenolchloroformisoamyl procedure Five micrograms of genomic DNA wasmodified with sodium bisulfite under conditions previouslydescribed (5) The maspin promoter was amplified from thebisulfite-modified DNA by nested PCR reactions using primersspecific to the bisulfite-modified sequence of the maspinpromoter The first-round forward primer 5rsquo-AAA AGA ATGGAG ATT AGA GTA TTT TTT GTG-3rsquo and reverse primer5rsquo-CCT AAA ATC ACA ATT ATC CTA AAA AAT A-3rsquo al-lowed the amplification of the fragment from 2283 to +181relative to the transcription start site PCR amplifications wereperformed under the following conditions 94degC for 4 min 5cycles of 94degC for 1 min 56degC for 2 min 72degC for 3 minthen 35 cycles of 94degC for 30 s 56degC for 2 min 72degC for 15min and a final extension of 72degC for 10 min One microliterof first PCR product was further used as template in a nestedPCR amplification The nested forward and reverse primers5rsquo-GAA ATT TGT AGT GTT ATT ATT ATT ATA-3rsquo and 5rsquo-
AAA AAC ACA AAA ACC TAA ATA TAA AAA-3rsquo wereused to generate the fragment from 2237 to +133 PCR am-plifications were performed under the following conditions94degC for 4 min 30 cycles of 94degC for 30 s 56degC for 1 min72degC for 15 min and a final extension of 72degC for 10 minFresh nested PCR products were gel-isolated and cloned intothe TA Original vector according to the manufacturerrsquos instruc-tions (Invitrogen Carlsbad CA USA) Twenty-five positiverecombinants from MDA-MB-435 cells and 10 positive re-combinants from adenovirus-infected cells were isolated andplasmids were prepared using a Qiaprep Spin Plasmid Miniprepkit (Qiagen Valencia CA USA) according to the manufac-turerrsquos instructions and sequenced on an ABI automated DNAsequencer The number of methylated CpG at each site wasdivided by the number of clones analyzed to yield the per-centage of methylation
MnSOD expression plasmids maspin reportergene construct and transient transfections
pcDNA3-MnSOD expression vector which contains thefull-length human MnSOD cDNA driven by a CMV promoterwas constructed as described (33) The maspin promoter re-porter construct (pGL3-MP) was made by fusing the maspingene 5rsquo-flanking region from 2396 to +131 relative to thetranscription start site to the pGL3Basic vector (PromegaMadison WI USA) The identity and orientation of the in-sert were verified by DNA sequencing Two micrograms ofeach pGL3-MP pcDNA3-MnSOD was used for transient trans-fections One microgram of pCMV-b-galactosidase vector wasused as a control for normalizing variations in transfection ef-ficiencies within and between experiments The pcDNA3 orpGL3-Basic vectors wereadded to normalize the total amountof DNA transfected in each assay Transfection was done in5 3 105 MDA-MB-435 cells and DU145 cells by using 20 microlof SuperFect transfection reagent (Qiagen) After 4 h of incu-bation in 1 ml of full growth medium transfection mixturewas removed and cells were allowed to recover in full mediumfor an additional 48 h Plasmid DNA was transferred into ZR-75-1 cells by electroporation Ten micrograms of pGL3-MPor pGL3-Basic with 5 microg of pCMV-b-galactosidase was trans-ferred into 5 3 107 cells by a single electric pulse at 260 Vand 1180 microF ZR-75-1 cells were grown in 5 ml of growthmedium in a 60-mm dish after transfection Four hours laterculture medium was changed Cells were grown in fresh mediumfor 48 h All the transfected cells were harvested and lysedusing 200 microl of passive lysis buffer (Promega) The resultantcell lysate was used for luciferase reporter gene assay and b-galactosidase activity assay
Promoter activity assay
Luciferase activity of 20 microl of cell lysate was measuredafter addition of 100 microl of luciferase assay reagent followingthe manufacturerrsquos protocol (Promega) Luciferase activity wasnormalized to b-galactosidase activity from the same amountof extract in each sample The relative maspin promoter activ-ity in each cell type was represented as a fold increase rela-tive to that of pGL3-Basic vector The mean and standard de-viation from multiple plates in two independent transfectionexperiments were used as one datapoint
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 679
Nuclear run-on assay for maspin mRNAin vitro transcription
Five million PC3 cells were infected with 100 MOI (mul-tiplicity of infection) Ad-MnSOD or Ad-LacZ for 24 h andrecovered in full growth medium for another 24 h Cells wereharvested by scraping into 13 PBS and centrifugation at1500 rpm for 2 min The resulting cell pellet was resuspendedin 1 ml of hypotonic buffer [10 mM KCl 10 mM Tris-HCl(pH 75) 15 mM MgCl2 03 M sucrose 025 NP-40 1 mMdithiothreitol and 05 mM phenylmethylsulfonyl fluoride] onice for 15 min and gently disrupted in a Dounce homogenizerLysed cells were centrifuged at 1500 rpm for 15 min and nu-clear pellet was immediately resuspended in 200 microl of tran-scription reaction buffer [50 mM Tris-HCl (pH 75) 01 mMammonium sulfate 18 mM dithiothreitol 18 mM MnCl2 2 microlof RNasin 300 microM each of ATP CTP and GTP and 100 microCiof [32P]-UTP] Transcription was carried out for 30 min atroom temperature and terminated by incubation with 5 microl ofRNase-free DNase and 2 microl of tRNA (20 mgml) for 15 minTwenty microliters of 10 SDS and 2 microl of proteinase K (01mgml) were added and the incubation continued for 45 minat room temperature RNA was isolated using the RNeasy kit(Qiagen) according to the manufacturerrsquos protocol Equalamounts of radioactivity (~3 3 106 cpmml) were used to hy-bridize dot blots with denatured 2 microg of maspin and GAPDHcDNA in hybridization mixture [583 SSC 58 formamide012 SDS 12 mM EDTA 12 mM Tris-HCl (pH 75) 473
Denhardtrsquos 04 mgml yeast tRNA and 04 mgml shearedssDNA] for 72 h Blots were washed twice at 65 degC for 1 h 32Psignals were visualized and quantitated by a PhosphorImager
Maspin mRNA stability assay
To measure the maspin mRNA stability PC3 cells were in-cubated with 10 microgml actinomycin D (Sigma Chemical CoSt Louis MO USA) for the indicated times To study theeffect of MnSOD on maspin mRNA stability cells were in-fected with 100 MOI Ad-MnSOD or Ad-LacZ for 24 h andrecovered for 24 h before the treatment with the same dose ofactinomycin D Total cellular mRNA was harvested at eachtime point and subjected to northern blot analysis
Statistical analysis
Maspin promoter activity data were expressed as the meansof two independent experiments plusmn SD Statistical significanceamong data groups was determined by one-way ANOVA andfurther two-sample t test at p lt 005 level using S-PLUS 2000
RESULTS
Ad-MnSOD increased MnSOD protein expressionin human breast and prostate cancer cells
Adenovirus has proven to be a powerful gene delivery ve-hicle for the expression of transgenes In this study we used atype 5 recombinant adenovirus vector containing the full-lengthhuman MnSOD cDNA driven by a CMV promoter (Ad-MnSOD) to infect human breast and prostate cancer cells
Similarly Ad-LacZ was used as the infection control To ver-ify the expression of MnSOD protein in the studied cells weinfected one breast cancer cell line MDA-MB-435 and oneprostate cancer cell line DU145 with Ad-MnSOD and mea-sured MnSOD protein expression by western blot analysisThe SDS-PAGE western blot results are shown in Fig 1 Asseen in Fig 1 Ad-MnSOD infection led to MnSOD expres-sion detectable as low as 10 MOI and abundantly at 100 MOIin both of the two cell types compared with controls Ad-LacZ as high as 100 MOI did not induce MnSOD protein ex-pression We also observed a band with a higher molecularweight of MnSOD in DU145 cells This is likely the MnSODprotein precursor which has an extra 24 amino acids that areresponsible for the transportation of the precursor to the mito-chondria (30) Previous immunohistochemical studies havedemonstrated the correct mitochondrial localization of thisadenovirally expressed MnSOD (14)
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells
To determine whether overexpression of MnSOD couldup-regulate maspin we performed northern blot analyses tomeasure maspin mRNA expression in a variety of untreatedAd-LacZ-infected and Ad-MnSOD-infected human breast andprostate cancer cells The northern blot results from human breastcancer cells MDA-MB-435 and MDA-MB-231 are shown inFig 2A and C The results from human prostate cancer cellsDU145 PC3 and LNCaP are shown in Fig 3A B and C re-spectively As shown in Fig 2 we observed a dose-dependentincrease of maspin mRNA expression in MDA-MB-435 andMDA-MB-231 cells infected with 50ndash250 MOI Ad-MnSODImportantly MnSOD expression also induced expression of
680 DUAN ET AL
FIG 1 Overexpression of MnSOD protein in Ad-MnSOD-infected human breast cancer cell MDA-MB-435 and humanprostate cancer cell DU145 Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Ten mi-crograms of whole cell lysate was separated by SDS-PAGE andanalyzed by immunoblotting with rabbit antiserum againsthuman MnSOD
the maspin protein in MDA-MB-231 cells as determined bywestern blot analysis shown in Fig 2B The induced maspinmRNA transcript was ~30 kb determined by its location rela-tive to the 18S and 28S RNA Infection with Ad-LacZ did notinduce maspin expression Interestingly in Ad-MnSOD-infectedhuman prostate cancer cells we observed two maspin tran-scripts corresponding to 30 kb and 12 kb respectively Care-ful inspection of the 3rsquo-untranslated region (3rsquo-UTR) of maspincDNA revealed two potential poly(A) signals located at nu-cleotides 1239 to 1244 and nucleotides 2551 to 2555 (Fig3C) relative to the transcription start site These two poly(A)signals apparently lead to the generation of the shorter 12-kband longer 30-kb maspin transcripts Between the two tran-scripts the smaller 12-kb transcript was much more induciblethan the 30-kb transcripts as seen in the northern blots Themechanisms of this change will be discussed later from theperspective of altered mRNA stability
MnSOD up-regulated maspin mRNA expression in human breast and prostate cancer cellsindependent of the methylation status of the maspin promoter
Our previous study showed that aberrant CpG methylationof maspin promoter participated in the silencing of maspin inmost of the studied breast carcinoma cell lines (6) To findout whether the effect of MnSOD on up-regulation of maspinmRNA in human breast cancer cells is through demethylatingthe maspin promoter we examined the cytosine methylationstatus of the maspin 5rsquo regulatory region from 2237 to +133in MDA-MB-435 cells before and after adenovirus infection
To increase the amplification efficiency of bisulfite-modifiedDNA we performed nested PCR reactions The two pairs ofprimers in the maspin promoter region are shown above thearrows in Fig 4A This amplified region contains 19 CpGdinucleotides that are potential sites for cytosine methylationThe bisulfite genomic sequencing results of maspin promotermethylation status in the untreated MDA-MB-435 cell Ad-LacZ-infected cell and Ad-MnSOD-infected cell are shownin Fig 4BndashD As seen in Fig 4B nearly all the 19 CpGs arehypermethylated in MDA-MB-435 cells Neither Ad-LacZ norAd-MnSOD infection changed the maspin promoter methyla-tion pattern under the same conditions that provided maximalmaspin reactivation Therefore the effect of MnSOD on maspinexpression is not due to the demethylation of maspin pro-moter in MDA-MB-435 cells
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 681
FIG 2 Ad-MnSOD infection reactivates maspin mRNA(A) and protein (B) expression in human breast cancer cellsMDA-MB-231 Ad-MnSOD infection also reactivates maspinmRNA expression in MDA-MB-435 human breast carcinomacells (C) Cells were infected with Ad-LacZ or Ad-MnSOD for24 h and recovered for 48 h Twenty micrograms of total RNAwas used for northern blot analysis of maspin mRNA expres-sion GAPDH mRNA expression was used as RNA loadingand transfer control Twenty micrograms of total cellular pro-tein was used for western blot analysis of maspin protein ex-pression
FIG 3 Ad-MnSOD infection up-regulated maspin mRNAexpression in human prostate cancer cells DU145 (A) PC3(B) and LNCaP (C) Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Twenty micrograms oftotal RNA was put to northern blot analysis of maspin mRNAexpression GAPDH mRNA expression was used as RNA load-ing and transfer control (D) Schematic representation of maspin3rsquo-UTR showing the two poly(A) signals
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells independent ofcellular p53 status
Zou et al reported that p53 up-regulated maspin expres-sion in both prostate and breast cancer cells including all threeprostate cancer cells we used here and one breast cancer cell
MCF-7 (41) Thus it is reasonable to hypothesize that MnSODup-regulated maspin in a p53-dependent manner To clarifywhether this was the case we examined the p53 status of thecells that were studied here and the MCF-7 cell which wasstudied previously (16) based on the published literature Re-sults of this analysis are summarized in Table 1 Among thethree breast cancer cell lines MCF-7 harbors wild-type p53
682 DUAN ET AL
FIG 4 Ad-MnSOD did not change the maspin promoter methylation status in human breast cancer MDA-MB-435 cells(A) DNA sequence of maspin 5rsquo regulatory region Two pairs of arrows indicate the four primers for nest PCR The 19 CpG po-tential sites for cytosine methylation are underlined Cells were infected with 100 MOI Ad-LacZ or Ad-MnSOD for 24 h and re-covered for 48 h The cytosine methylation status of maspin in untreated MDA-MB-435 cells Ad-LacZ-infected cells and Ad-MnSOD-infected cells is shown in BndashD
A
B
C
D
whereas MDA-MB-435 and MDA-MB-231 cells harbor mu-tant p53 Among the prostate cancer cell lines LNCaP har-bors wild-type p53 expression whereas DU145 has a mutantp53 expression and PC3 is a p53 null cell line Therefore theup-regulation effect of MnSOD on maspin mRNA expressionwas independent of cellular p53 status
MnSOD did not influence maspin promoter activityor mRNA transcription rate in PC3 cells
Several studies have shown that maspin promoter activityis abolished or down-regulated in human breast and prostatecancer cells compared with their normal counterpart (34 35)To elucidate whether the effect of MnSOD is due to activationof the maspin promoter we measured maspin promoter activ-ity in transient and stable MnSOD overexpression modelsThe maspin promoter luciferase reporter construct pGL3-MPconsists of the 5rsquo regulatory region from 2396 to +131 rela-tive to the maspin transcription start (Fig 5A) This regionwhich contains the binding sites of Ets p53 and AP-1 hasbeen characterized to be necessary and sufficient for the maspintransactivation (34) To test the function of this construct wefirst transfected it into an immortalized but nontumorigenichuman breast epithelial cell line MCF-10A that constitutivelyexpresses a high level of maspin As shown in Fig 5B maspinpromoter activity in MCF-10A cells was higher than the ac-tivity of pGL3-Basic construct Compared with the promoteractivity in normal MCF-10A cells the maspin promoter wasless active in human breast cancer cells MDA-MB-435 andZR-75-1 To determine whether MnSOD up-regulated themaspin promoter we cotransfected pGL3-MP and an MnSODexpression vector pcDNA3-MnSOD into MDA-MB-435 andDU145 cells As shown in Fig 5C MnSOD had no effect onmaspin promoter activity in either of the two cell lines We alsomeasured maspin promoter activity in MnSOD stable trans-fectants SOD15 Mn1 and Mn40 which were derived fromMCF-7 human breast cancer cells (33) The MnSOD enzymeactivities in the stable transfectants are 19 62 and 230 Umgof protein for SOD15 Mn1 and Mn40 respectively How-ever no significant changes were observed in maspin promoteractivity among the three MnSOD overexpressing cell lines(Fig 5D)
To explore further whether MnSOD modulates maspin mRNAexpression at the transcriptional level we measured the maspin
mRNA in vitro transcription rate The nuclear run-on assayresults in the Ad-MnSOD- and Ad-LacZ-infected PC3 cellsare shown in Fig 6 Again we detected an almost equal amountof the newly transcribed maspin mRNA signal in the two groupsThis result indicates that MnSOD has no effect on the rate oftranscriptional initiation of maspin as determined by this invitro transcription assay
MnSOD increased maspin mRNA stability in PC3 cells
Much attention has been focused on the transcriptional reg-ulation of maspin in normal and cancer cells Thus far an ef-fect of maspin mRNA stability has not been explored In thisstudy we examined maspin mRNA stability in PC3 cells whichhave the most abundant endogenous maspin mRNA expres-sion among the cancer cells we used here We inhibited newmaspin mRNA synthesis by treating the cells with 10 microgmlactinomycin D and measured the residual levels of the 30-kband 12-kb maspin transcripts at various time points A repre-sentative northern blot result is shown in Fig 7A Quantita-tive analysis of the maspin half-life based on two independentexperiments is shown in Fig 7B It shows that the half-life ofthe 30-kb transcript was ~20 h and the half-life of 12-kbtranscript was much longer ~30ndash36 h
To determine whether the effect of MnSOD on the up-regulation of maspin mRNA was due to the modulation ofmaspin mRNA stability we infected the PC3 cells with 100MOI Ad-LacZ and Ad-MnSOD then treated cells with actin-omycin D and assessed maspin mRNA levels at each timepoint The northern blot results are shown in Fig 8A Quanti-tative analysis of the maspin mRNA remaining as a functionof time is shown in Fig 8B for the 30-kb transcript and Fig8C for the 12-kb transcript These data suggest that MnSODincreased the stability of both transcripts compared with theAd-LacZ infection control For the 30-kb transcript the resid-ual maspin mRNA dropped to 30 within 36 h In contrastthe 12-kb transcript consistently remained at nearly 80 from12 to 36 hours Therefore the stabilizing effect of MnSODexpression on the 12-kb transcript was more significant thanthat on the 30-kb transcript Taking into account that the 12-kb transcript has a longer half-life it might explain why weobserved a significant accumulation of 12-kb transcript butnot the 30-kb transcript after MnSOD overexpression
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 683
TABLE 1 OVERVIEW OF MASPIN PROMOTER METHYLATION AND P53STATUS IN HUMAN BREAST AND PROSTATE CANCER CELL LINES
Cell types p53 status Promoter methylation status (6)
Breast cancer cellsMDA-MB-435 mt-p53 +MDA-MB-231 mt-p53 +MCF-7 wt-p53 +
Prostate cancer cellsPC3 p53(22) 2DU145 mt-p53 2LNCaP wt-p53 2
mt mutant wt wild type
DISCUSSION
Tumor cells have often been found to display lower levelsof antioxidant enzyme expression than their normal counter-parts For example tumor cells are nearly always low in MnSODand catalase activity and usually low in copperzinc superox-ide dismutase activity whereas glutathione peroxidase activ-ity is variable (3 25 33) MnSOD is a nuclear-encoded mito-chondrial matrix protein and serves a protective function by
684 DUAN ET AL
FIG 5 Maspin promoter activity in human breast andprostate cancer cells (A) Schematic representation of maspinpromoter construct with the important cis-elements labeled inthe shaded areas (B) Maspin promoter activity was down-regulated in human breast cancer cells MCF-10A and MDA-MB-435 cells were transiently transfected with 2 microg of maspinpromoter construct pGL3-MP or pGL3-Basic together with 1microg of pCMV-b-galactosidase as internal control ZR-75ndash1 cellswere transfected by electroporation Ten micrograms of pGL3-MP or pGL3-basic was transfected into 5 3 107 cells with 5 microgof pCMV-b-galactosidase as the internal control The relativeluciferase activity normalized by b-galactosidase activity wasrepresented as the fold increase compared with pGL3-Basicvector activity in each cell types (C) MnSOD did not influencemaspin promoter activity in human breast cancer MDA-MB-435 cells and human prostate cancer DU145 cells Two micro-grams of pcDNA3-MnSOD expression plasmid was cotrans-fected with 2 microg of pGL3-MP The relative maspin promoteractivity in the control cells was designated as 100 (D)Maspin promoter activity in MnSOD transfected cells was rep-resented as a percentage increase compared with control Re-sults shown here are the means and standard deviations fromtwo independent transfection experiments
FIG 6 Influence of MnSOD on the maspin mRNA invitro transcription rate in PC3 cells Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h total RNA was isolated and put into in vitro tran-scription analysis [32P]-UTP-labeled mRNA samples were hy-bridized with slot-blotted 2 microg of maspin and GAPDH cDNA32P signals were detected and quantified by PhosphorImager
FIG 7 Maspin mRNA stability in human prostate cancerPC3 cells (A) Northern blot analysis for maspin mRNA stabil-ity assay Cells were treated with 10 microgml actinomycin DTotal RNA was harvested at the times indicated and processedfor northern blot analysis The mRNA signals were quantifiedby PhosphorImager (B) The remaining maspin mRNA tran-scripts normalized to GAPDH loading control were plottedversus time
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL
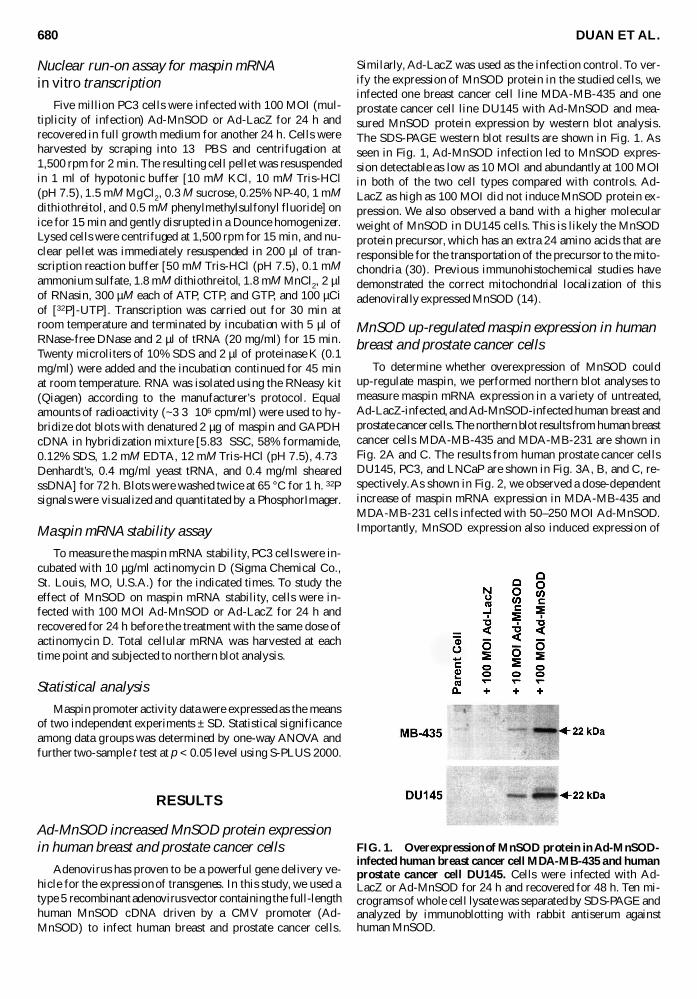
Nuclear run-on assay for maspin mRNAin vitro transcription
Five million PC3 cells were infected with 100 MOI (mul-tiplicity of infection) Ad-MnSOD or Ad-LacZ for 24 h andrecovered in full growth medium for another 24 h Cells wereharvested by scraping into 13 PBS and centrifugation at1500 rpm for 2 min The resulting cell pellet was resuspendedin 1 ml of hypotonic buffer [10 mM KCl 10 mM Tris-HCl(pH 75) 15 mM MgCl2 03 M sucrose 025 NP-40 1 mMdithiothreitol and 05 mM phenylmethylsulfonyl fluoride] onice for 15 min and gently disrupted in a Dounce homogenizerLysed cells were centrifuged at 1500 rpm for 15 min and nu-clear pellet was immediately resuspended in 200 microl of tran-scription reaction buffer [50 mM Tris-HCl (pH 75) 01 mMammonium sulfate 18 mM dithiothreitol 18 mM MnCl2 2 microlof RNasin 300 microM each of ATP CTP and GTP and 100 microCiof [32P]-UTP] Transcription was carried out for 30 min atroom temperature and terminated by incubation with 5 microl ofRNase-free DNase and 2 microl of tRNA (20 mgml) for 15 minTwenty microliters of 10 SDS and 2 microl of proteinase K (01mgml) were added and the incubation continued for 45 minat room temperature RNA was isolated using the RNeasy kit(Qiagen) according to the manufacturerrsquos protocol Equalamounts of radioactivity (~3 3 106 cpmml) were used to hy-bridize dot blots with denatured 2 microg of maspin and GAPDHcDNA in hybridization mixture [583 SSC 58 formamide012 SDS 12 mM EDTA 12 mM Tris-HCl (pH 75) 473
Denhardtrsquos 04 mgml yeast tRNA and 04 mgml shearedssDNA] for 72 h Blots were washed twice at 65 degC for 1 h 32Psignals were visualized and quantitated by a PhosphorImager
Maspin mRNA stability assay
To measure the maspin mRNA stability PC3 cells were in-cubated with 10 microgml actinomycin D (Sigma Chemical CoSt Louis MO USA) for the indicated times To study theeffect of MnSOD on maspin mRNA stability cells were in-fected with 100 MOI Ad-MnSOD or Ad-LacZ for 24 h andrecovered for 24 h before the treatment with the same dose ofactinomycin D Total cellular mRNA was harvested at eachtime point and subjected to northern blot analysis
Statistical analysis
Maspin promoter activity data were expressed as the meansof two independent experiments plusmn SD Statistical significanceamong data groups was determined by one-way ANOVA andfurther two-sample t test at p lt 005 level using S-PLUS 2000
RESULTS
Ad-MnSOD increased MnSOD protein expressionin human breast and prostate cancer cells
Adenovirus has proven to be a powerful gene delivery ve-hicle for the expression of transgenes In this study we used atype 5 recombinant adenovirus vector containing the full-lengthhuman MnSOD cDNA driven by a CMV promoter (Ad-MnSOD) to infect human breast and prostate cancer cells
Similarly Ad-LacZ was used as the infection control To ver-ify the expression of MnSOD protein in the studied cells weinfected one breast cancer cell line MDA-MB-435 and oneprostate cancer cell line DU145 with Ad-MnSOD and mea-sured MnSOD protein expression by western blot analysisThe SDS-PAGE western blot results are shown in Fig 1 Asseen in Fig 1 Ad-MnSOD infection led to MnSOD expres-sion detectable as low as 10 MOI and abundantly at 100 MOIin both of the two cell types compared with controls Ad-LacZ as high as 100 MOI did not induce MnSOD protein ex-pression We also observed a band with a higher molecularweight of MnSOD in DU145 cells This is likely the MnSODprotein precursor which has an extra 24 amino acids that areresponsible for the transportation of the precursor to the mito-chondria (30) Previous immunohistochemical studies havedemonstrated the correct mitochondrial localization of thisadenovirally expressed MnSOD (14)
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells
To determine whether overexpression of MnSOD couldup-regulate maspin we performed northern blot analyses tomeasure maspin mRNA expression in a variety of untreatedAd-LacZ-infected and Ad-MnSOD-infected human breast andprostate cancer cells The northern blot results from human breastcancer cells MDA-MB-435 and MDA-MB-231 are shown inFig 2A and C The results from human prostate cancer cellsDU145 PC3 and LNCaP are shown in Fig 3A B and C re-spectively As shown in Fig 2 we observed a dose-dependentincrease of maspin mRNA expression in MDA-MB-435 andMDA-MB-231 cells infected with 50ndash250 MOI Ad-MnSODImportantly MnSOD expression also induced expression of
680 DUAN ET AL
FIG 1 Overexpression of MnSOD protein in Ad-MnSOD-infected human breast cancer cell MDA-MB-435 and humanprostate cancer cell DU145 Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Ten mi-crograms of whole cell lysate was separated by SDS-PAGE andanalyzed by immunoblotting with rabbit antiserum againsthuman MnSOD
the maspin protein in MDA-MB-231 cells as determined bywestern blot analysis shown in Fig 2B The induced maspinmRNA transcript was ~30 kb determined by its location rela-tive to the 18S and 28S RNA Infection with Ad-LacZ did notinduce maspin expression Interestingly in Ad-MnSOD-infectedhuman prostate cancer cells we observed two maspin tran-scripts corresponding to 30 kb and 12 kb respectively Care-ful inspection of the 3rsquo-untranslated region (3rsquo-UTR) of maspincDNA revealed two potential poly(A) signals located at nu-cleotides 1239 to 1244 and nucleotides 2551 to 2555 (Fig3C) relative to the transcription start site These two poly(A)signals apparently lead to the generation of the shorter 12-kband longer 30-kb maspin transcripts Between the two tran-scripts the smaller 12-kb transcript was much more induciblethan the 30-kb transcripts as seen in the northern blots Themechanisms of this change will be discussed later from theperspective of altered mRNA stability
MnSOD up-regulated maspin mRNA expression in human breast and prostate cancer cellsindependent of the methylation status of the maspin promoter
Our previous study showed that aberrant CpG methylationof maspin promoter participated in the silencing of maspin inmost of the studied breast carcinoma cell lines (6) To findout whether the effect of MnSOD on up-regulation of maspinmRNA in human breast cancer cells is through demethylatingthe maspin promoter we examined the cytosine methylationstatus of the maspin 5rsquo regulatory region from 2237 to +133in MDA-MB-435 cells before and after adenovirus infection
To increase the amplification efficiency of bisulfite-modifiedDNA we performed nested PCR reactions The two pairs ofprimers in the maspin promoter region are shown above thearrows in Fig 4A This amplified region contains 19 CpGdinucleotides that are potential sites for cytosine methylationThe bisulfite genomic sequencing results of maspin promotermethylation status in the untreated MDA-MB-435 cell Ad-LacZ-infected cell and Ad-MnSOD-infected cell are shownin Fig 4BndashD As seen in Fig 4B nearly all the 19 CpGs arehypermethylated in MDA-MB-435 cells Neither Ad-LacZ norAd-MnSOD infection changed the maspin promoter methyla-tion pattern under the same conditions that provided maximalmaspin reactivation Therefore the effect of MnSOD on maspinexpression is not due to the demethylation of maspin pro-moter in MDA-MB-435 cells
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 681
FIG 2 Ad-MnSOD infection reactivates maspin mRNA(A) and protein (B) expression in human breast cancer cellsMDA-MB-231 Ad-MnSOD infection also reactivates maspinmRNA expression in MDA-MB-435 human breast carcinomacells (C) Cells were infected with Ad-LacZ or Ad-MnSOD for24 h and recovered for 48 h Twenty micrograms of total RNAwas used for northern blot analysis of maspin mRNA expres-sion GAPDH mRNA expression was used as RNA loadingand transfer control Twenty micrograms of total cellular pro-tein was used for western blot analysis of maspin protein ex-pression
FIG 3 Ad-MnSOD infection up-regulated maspin mRNAexpression in human prostate cancer cells DU145 (A) PC3(B) and LNCaP (C) Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Twenty micrograms oftotal RNA was put to northern blot analysis of maspin mRNAexpression GAPDH mRNA expression was used as RNA load-ing and transfer control (D) Schematic representation of maspin3rsquo-UTR showing the two poly(A) signals
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells independent ofcellular p53 status
Zou et al reported that p53 up-regulated maspin expres-sion in both prostate and breast cancer cells including all threeprostate cancer cells we used here and one breast cancer cell
MCF-7 (41) Thus it is reasonable to hypothesize that MnSODup-regulated maspin in a p53-dependent manner To clarifywhether this was the case we examined the p53 status of thecells that were studied here and the MCF-7 cell which wasstudied previously (16) based on the published literature Re-sults of this analysis are summarized in Table 1 Among thethree breast cancer cell lines MCF-7 harbors wild-type p53
682 DUAN ET AL
FIG 4 Ad-MnSOD did not change the maspin promoter methylation status in human breast cancer MDA-MB-435 cells(A) DNA sequence of maspin 5rsquo regulatory region Two pairs of arrows indicate the four primers for nest PCR The 19 CpG po-tential sites for cytosine methylation are underlined Cells were infected with 100 MOI Ad-LacZ or Ad-MnSOD for 24 h and re-covered for 48 h The cytosine methylation status of maspin in untreated MDA-MB-435 cells Ad-LacZ-infected cells and Ad-MnSOD-infected cells is shown in BndashD
A
B
C
D
whereas MDA-MB-435 and MDA-MB-231 cells harbor mu-tant p53 Among the prostate cancer cell lines LNCaP har-bors wild-type p53 expression whereas DU145 has a mutantp53 expression and PC3 is a p53 null cell line Therefore theup-regulation effect of MnSOD on maspin mRNA expressionwas independent of cellular p53 status
MnSOD did not influence maspin promoter activityor mRNA transcription rate in PC3 cells
Several studies have shown that maspin promoter activityis abolished or down-regulated in human breast and prostatecancer cells compared with their normal counterpart (34 35)To elucidate whether the effect of MnSOD is due to activationof the maspin promoter we measured maspin promoter activ-ity in transient and stable MnSOD overexpression modelsThe maspin promoter luciferase reporter construct pGL3-MPconsists of the 5rsquo regulatory region from 2396 to +131 rela-tive to the maspin transcription start (Fig 5A) This regionwhich contains the binding sites of Ets p53 and AP-1 hasbeen characterized to be necessary and sufficient for the maspintransactivation (34) To test the function of this construct wefirst transfected it into an immortalized but nontumorigenichuman breast epithelial cell line MCF-10A that constitutivelyexpresses a high level of maspin As shown in Fig 5B maspinpromoter activity in MCF-10A cells was higher than the ac-tivity of pGL3-Basic construct Compared with the promoteractivity in normal MCF-10A cells the maspin promoter wasless active in human breast cancer cells MDA-MB-435 andZR-75-1 To determine whether MnSOD up-regulated themaspin promoter we cotransfected pGL3-MP and an MnSODexpression vector pcDNA3-MnSOD into MDA-MB-435 andDU145 cells As shown in Fig 5C MnSOD had no effect onmaspin promoter activity in either of the two cell lines We alsomeasured maspin promoter activity in MnSOD stable trans-fectants SOD15 Mn1 and Mn40 which were derived fromMCF-7 human breast cancer cells (33) The MnSOD enzymeactivities in the stable transfectants are 19 62 and 230 Umgof protein for SOD15 Mn1 and Mn40 respectively How-ever no significant changes were observed in maspin promoteractivity among the three MnSOD overexpressing cell lines(Fig 5D)
To explore further whether MnSOD modulates maspin mRNAexpression at the transcriptional level we measured the maspin
mRNA in vitro transcription rate The nuclear run-on assayresults in the Ad-MnSOD- and Ad-LacZ-infected PC3 cellsare shown in Fig 6 Again we detected an almost equal amountof the newly transcribed maspin mRNA signal in the two groupsThis result indicates that MnSOD has no effect on the rate oftranscriptional initiation of maspin as determined by this invitro transcription assay
MnSOD increased maspin mRNA stability in PC3 cells
Much attention has been focused on the transcriptional reg-ulation of maspin in normal and cancer cells Thus far an ef-fect of maspin mRNA stability has not been explored In thisstudy we examined maspin mRNA stability in PC3 cells whichhave the most abundant endogenous maspin mRNA expres-sion among the cancer cells we used here We inhibited newmaspin mRNA synthesis by treating the cells with 10 microgmlactinomycin D and measured the residual levels of the 30-kband 12-kb maspin transcripts at various time points A repre-sentative northern blot result is shown in Fig 7A Quantita-tive analysis of the maspin half-life based on two independentexperiments is shown in Fig 7B It shows that the half-life ofthe 30-kb transcript was ~20 h and the half-life of 12-kbtranscript was much longer ~30ndash36 h
To determine whether the effect of MnSOD on the up-regulation of maspin mRNA was due to the modulation ofmaspin mRNA stability we infected the PC3 cells with 100MOI Ad-LacZ and Ad-MnSOD then treated cells with actin-omycin D and assessed maspin mRNA levels at each timepoint The northern blot results are shown in Fig 8A Quanti-tative analysis of the maspin mRNA remaining as a functionof time is shown in Fig 8B for the 30-kb transcript and Fig8C for the 12-kb transcript These data suggest that MnSODincreased the stability of both transcripts compared with theAd-LacZ infection control For the 30-kb transcript the resid-ual maspin mRNA dropped to 30 within 36 h In contrastthe 12-kb transcript consistently remained at nearly 80 from12 to 36 hours Therefore the stabilizing effect of MnSODexpression on the 12-kb transcript was more significant thanthat on the 30-kb transcript Taking into account that the 12-kb transcript has a longer half-life it might explain why weobserved a significant accumulation of 12-kb transcript butnot the 30-kb transcript after MnSOD overexpression
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 683
TABLE 1 OVERVIEW OF MASPIN PROMOTER METHYLATION AND P53STATUS IN HUMAN BREAST AND PROSTATE CANCER CELL LINES
Cell types p53 status Promoter methylation status (6)
Breast cancer cellsMDA-MB-435 mt-p53 +MDA-MB-231 mt-p53 +MCF-7 wt-p53 +
Prostate cancer cellsPC3 p53(22) 2DU145 mt-p53 2LNCaP wt-p53 2
mt mutant wt wild type
DISCUSSION
Tumor cells have often been found to display lower levelsof antioxidant enzyme expression than their normal counter-parts For example tumor cells are nearly always low in MnSODand catalase activity and usually low in copperzinc superox-ide dismutase activity whereas glutathione peroxidase activ-ity is variable (3 25 33) MnSOD is a nuclear-encoded mito-chondrial matrix protein and serves a protective function by
684 DUAN ET AL
FIG 5 Maspin promoter activity in human breast andprostate cancer cells (A) Schematic representation of maspinpromoter construct with the important cis-elements labeled inthe shaded areas (B) Maspin promoter activity was down-regulated in human breast cancer cells MCF-10A and MDA-MB-435 cells were transiently transfected with 2 microg of maspinpromoter construct pGL3-MP or pGL3-Basic together with 1microg of pCMV-b-galactosidase as internal control ZR-75ndash1 cellswere transfected by electroporation Ten micrograms of pGL3-MP or pGL3-basic was transfected into 5 3 107 cells with 5 microgof pCMV-b-galactosidase as the internal control The relativeluciferase activity normalized by b-galactosidase activity wasrepresented as the fold increase compared with pGL3-Basicvector activity in each cell types (C) MnSOD did not influencemaspin promoter activity in human breast cancer MDA-MB-435 cells and human prostate cancer DU145 cells Two micro-grams of pcDNA3-MnSOD expression plasmid was cotrans-fected with 2 microg of pGL3-MP The relative maspin promoteractivity in the control cells was designated as 100 (D)Maspin promoter activity in MnSOD transfected cells was rep-resented as a percentage increase compared with control Re-sults shown here are the means and standard deviations fromtwo independent transfection experiments
FIG 6 Influence of MnSOD on the maspin mRNA invitro transcription rate in PC3 cells Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h total RNA was isolated and put into in vitro tran-scription analysis [32P]-UTP-labeled mRNA samples were hy-bridized with slot-blotted 2 microg of maspin and GAPDH cDNA32P signals were detected and quantified by PhosphorImager
FIG 7 Maspin mRNA stability in human prostate cancerPC3 cells (A) Northern blot analysis for maspin mRNA stabil-ity assay Cells were treated with 10 microgml actinomycin DTotal RNA was harvested at the times indicated and processedfor northern blot analysis The mRNA signals were quantifiedby PhosphorImager (B) The remaining maspin mRNA tran-scripts normalized to GAPDH loading control were plottedversus time
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL
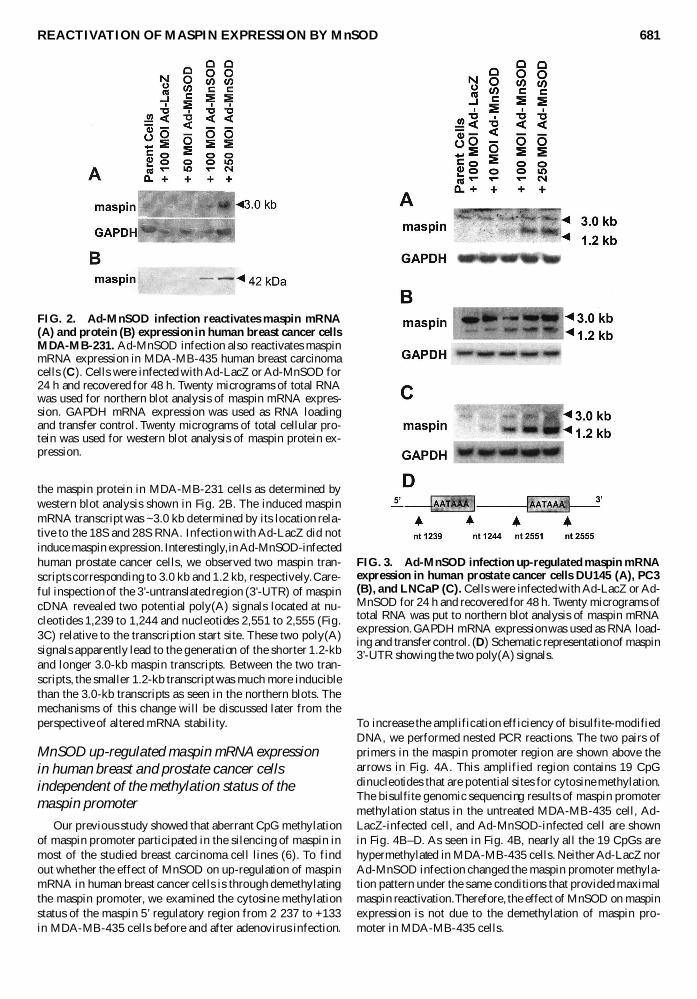
the maspin protein in MDA-MB-231 cells as determined bywestern blot analysis shown in Fig 2B The induced maspinmRNA transcript was ~30 kb determined by its location rela-tive to the 18S and 28S RNA Infection with Ad-LacZ did notinduce maspin expression Interestingly in Ad-MnSOD-infectedhuman prostate cancer cells we observed two maspin tran-scripts corresponding to 30 kb and 12 kb respectively Care-ful inspection of the 3rsquo-untranslated region (3rsquo-UTR) of maspincDNA revealed two potential poly(A) signals located at nu-cleotides 1239 to 1244 and nucleotides 2551 to 2555 (Fig3C) relative to the transcription start site These two poly(A)signals apparently lead to the generation of the shorter 12-kband longer 30-kb maspin transcripts Between the two tran-scripts the smaller 12-kb transcript was much more induciblethan the 30-kb transcripts as seen in the northern blots Themechanisms of this change will be discussed later from theperspective of altered mRNA stability
MnSOD up-regulated maspin mRNA expression in human breast and prostate cancer cellsindependent of the methylation status of the maspin promoter
Our previous study showed that aberrant CpG methylationof maspin promoter participated in the silencing of maspin inmost of the studied breast carcinoma cell lines (6) To findout whether the effect of MnSOD on up-regulation of maspinmRNA in human breast cancer cells is through demethylatingthe maspin promoter we examined the cytosine methylationstatus of the maspin 5rsquo regulatory region from 2237 to +133in MDA-MB-435 cells before and after adenovirus infection
To increase the amplification efficiency of bisulfite-modifiedDNA we performed nested PCR reactions The two pairs ofprimers in the maspin promoter region are shown above thearrows in Fig 4A This amplified region contains 19 CpGdinucleotides that are potential sites for cytosine methylationThe bisulfite genomic sequencing results of maspin promotermethylation status in the untreated MDA-MB-435 cell Ad-LacZ-infected cell and Ad-MnSOD-infected cell are shownin Fig 4BndashD As seen in Fig 4B nearly all the 19 CpGs arehypermethylated in MDA-MB-435 cells Neither Ad-LacZ norAd-MnSOD infection changed the maspin promoter methyla-tion pattern under the same conditions that provided maximalmaspin reactivation Therefore the effect of MnSOD on maspinexpression is not due to the demethylation of maspin pro-moter in MDA-MB-435 cells
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 681
FIG 2 Ad-MnSOD infection reactivates maspin mRNA(A) and protein (B) expression in human breast cancer cellsMDA-MB-231 Ad-MnSOD infection also reactivates maspinmRNA expression in MDA-MB-435 human breast carcinomacells (C) Cells were infected with Ad-LacZ or Ad-MnSOD for24 h and recovered for 48 h Twenty micrograms of total RNAwas used for northern blot analysis of maspin mRNA expres-sion GAPDH mRNA expression was used as RNA loadingand transfer control Twenty micrograms of total cellular pro-tein was used for western blot analysis of maspin protein ex-pression
FIG 3 Ad-MnSOD infection up-regulated maspin mRNAexpression in human prostate cancer cells DU145 (A) PC3(B) and LNCaP (C) Cells were infected with Ad-LacZ or Ad-MnSOD for 24 h and recovered for 48 h Twenty micrograms oftotal RNA was put to northern blot analysis of maspin mRNAexpression GAPDH mRNA expression was used as RNA load-ing and transfer control (D) Schematic representation of maspin3rsquo-UTR showing the two poly(A) signals
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells independent ofcellular p53 status
Zou et al reported that p53 up-regulated maspin expres-sion in both prostate and breast cancer cells including all threeprostate cancer cells we used here and one breast cancer cell
MCF-7 (41) Thus it is reasonable to hypothesize that MnSODup-regulated maspin in a p53-dependent manner To clarifywhether this was the case we examined the p53 status of thecells that were studied here and the MCF-7 cell which wasstudied previously (16) based on the published literature Re-sults of this analysis are summarized in Table 1 Among thethree breast cancer cell lines MCF-7 harbors wild-type p53
682 DUAN ET AL
FIG 4 Ad-MnSOD did not change the maspin promoter methylation status in human breast cancer MDA-MB-435 cells(A) DNA sequence of maspin 5rsquo regulatory region Two pairs of arrows indicate the four primers for nest PCR The 19 CpG po-tential sites for cytosine methylation are underlined Cells were infected with 100 MOI Ad-LacZ or Ad-MnSOD for 24 h and re-covered for 48 h The cytosine methylation status of maspin in untreated MDA-MB-435 cells Ad-LacZ-infected cells and Ad-MnSOD-infected cells is shown in BndashD
A
B
C
D
whereas MDA-MB-435 and MDA-MB-231 cells harbor mu-tant p53 Among the prostate cancer cell lines LNCaP har-bors wild-type p53 expression whereas DU145 has a mutantp53 expression and PC3 is a p53 null cell line Therefore theup-regulation effect of MnSOD on maspin mRNA expressionwas independent of cellular p53 status
MnSOD did not influence maspin promoter activityor mRNA transcription rate in PC3 cells
Several studies have shown that maspin promoter activityis abolished or down-regulated in human breast and prostatecancer cells compared with their normal counterpart (34 35)To elucidate whether the effect of MnSOD is due to activationof the maspin promoter we measured maspin promoter activ-ity in transient and stable MnSOD overexpression modelsThe maspin promoter luciferase reporter construct pGL3-MPconsists of the 5rsquo regulatory region from 2396 to +131 rela-tive to the maspin transcription start (Fig 5A) This regionwhich contains the binding sites of Ets p53 and AP-1 hasbeen characterized to be necessary and sufficient for the maspintransactivation (34) To test the function of this construct wefirst transfected it into an immortalized but nontumorigenichuman breast epithelial cell line MCF-10A that constitutivelyexpresses a high level of maspin As shown in Fig 5B maspinpromoter activity in MCF-10A cells was higher than the ac-tivity of pGL3-Basic construct Compared with the promoteractivity in normal MCF-10A cells the maspin promoter wasless active in human breast cancer cells MDA-MB-435 andZR-75-1 To determine whether MnSOD up-regulated themaspin promoter we cotransfected pGL3-MP and an MnSODexpression vector pcDNA3-MnSOD into MDA-MB-435 andDU145 cells As shown in Fig 5C MnSOD had no effect onmaspin promoter activity in either of the two cell lines We alsomeasured maspin promoter activity in MnSOD stable trans-fectants SOD15 Mn1 and Mn40 which were derived fromMCF-7 human breast cancer cells (33) The MnSOD enzymeactivities in the stable transfectants are 19 62 and 230 Umgof protein for SOD15 Mn1 and Mn40 respectively How-ever no significant changes were observed in maspin promoteractivity among the three MnSOD overexpressing cell lines(Fig 5D)
To explore further whether MnSOD modulates maspin mRNAexpression at the transcriptional level we measured the maspin
mRNA in vitro transcription rate The nuclear run-on assayresults in the Ad-MnSOD- and Ad-LacZ-infected PC3 cellsare shown in Fig 6 Again we detected an almost equal amountof the newly transcribed maspin mRNA signal in the two groupsThis result indicates that MnSOD has no effect on the rate oftranscriptional initiation of maspin as determined by this invitro transcription assay
MnSOD increased maspin mRNA stability in PC3 cells
Much attention has been focused on the transcriptional reg-ulation of maspin in normal and cancer cells Thus far an ef-fect of maspin mRNA stability has not been explored In thisstudy we examined maspin mRNA stability in PC3 cells whichhave the most abundant endogenous maspin mRNA expres-sion among the cancer cells we used here We inhibited newmaspin mRNA synthesis by treating the cells with 10 microgmlactinomycin D and measured the residual levels of the 30-kband 12-kb maspin transcripts at various time points A repre-sentative northern blot result is shown in Fig 7A Quantita-tive analysis of the maspin half-life based on two independentexperiments is shown in Fig 7B It shows that the half-life ofthe 30-kb transcript was ~20 h and the half-life of 12-kbtranscript was much longer ~30ndash36 h
To determine whether the effect of MnSOD on the up-regulation of maspin mRNA was due to the modulation ofmaspin mRNA stability we infected the PC3 cells with 100MOI Ad-LacZ and Ad-MnSOD then treated cells with actin-omycin D and assessed maspin mRNA levels at each timepoint The northern blot results are shown in Fig 8A Quanti-tative analysis of the maspin mRNA remaining as a functionof time is shown in Fig 8B for the 30-kb transcript and Fig8C for the 12-kb transcript These data suggest that MnSODincreased the stability of both transcripts compared with theAd-LacZ infection control For the 30-kb transcript the resid-ual maspin mRNA dropped to 30 within 36 h In contrastthe 12-kb transcript consistently remained at nearly 80 from12 to 36 hours Therefore the stabilizing effect of MnSODexpression on the 12-kb transcript was more significant thanthat on the 30-kb transcript Taking into account that the 12-kb transcript has a longer half-life it might explain why weobserved a significant accumulation of 12-kb transcript butnot the 30-kb transcript after MnSOD overexpression
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 683
TABLE 1 OVERVIEW OF MASPIN PROMOTER METHYLATION AND P53STATUS IN HUMAN BREAST AND PROSTATE CANCER CELL LINES
Cell types p53 status Promoter methylation status (6)
Breast cancer cellsMDA-MB-435 mt-p53 +MDA-MB-231 mt-p53 +MCF-7 wt-p53 +
Prostate cancer cellsPC3 p53(22) 2DU145 mt-p53 2LNCaP wt-p53 2
mt mutant wt wild type
DISCUSSION
Tumor cells have often been found to display lower levelsof antioxidant enzyme expression than their normal counter-parts For example tumor cells are nearly always low in MnSODand catalase activity and usually low in copperzinc superox-ide dismutase activity whereas glutathione peroxidase activ-ity is variable (3 25 33) MnSOD is a nuclear-encoded mito-chondrial matrix protein and serves a protective function by
684 DUAN ET AL
FIG 5 Maspin promoter activity in human breast andprostate cancer cells (A) Schematic representation of maspinpromoter construct with the important cis-elements labeled inthe shaded areas (B) Maspin promoter activity was down-regulated in human breast cancer cells MCF-10A and MDA-MB-435 cells were transiently transfected with 2 microg of maspinpromoter construct pGL3-MP or pGL3-Basic together with 1microg of pCMV-b-galactosidase as internal control ZR-75ndash1 cellswere transfected by electroporation Ten micrograms of pGL3-MP or pGL3-basic was transfected into 5 3 107 cells with 5 microgof pCMV-b-galactosidase as the internal control The relativeluciferase activity normalized by b-galactosidase activity wasrepresented as the fold increase compared with pGL3-Basicvector activity in each cell types (C) MnSOD did not influencemaspin promoter activity in human breast cancer MDA-MB-435 cells and human prostate cancer DU145 cells Two micro-grams of pcDNA3-MnSOD expression plasmid was cotrans-fected with 2 microg of pGL3-MP The relative maspin promoteractivity in the control cells was designated as 100 (D)Maspin promoter activity in MnSOD transfected cells was rep-resented as a percentage increase compared with control Re-sults shown here are the means and standard deviations fromtwo independent transfection experiments
FIG 6 Influence of MnSOD on the maspin mRNA invitro transcription rate in PC3 cells Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h total RNA was isolated and put into in vitro tran-scription analysis [32P]-UTP-labeled mRNA samples were hy-bridized with slot-blotted 2 microg of maspin and GAPDH cDNA32P signals were detected and quantified by PhosphorImager
FIG 7 Maspin mRNA stability in human prostate cancerPC3 cells (A) Northern blot analysis for maspin mRNA stabil-ity assay Cells were treated with 10 microgml actinomycin DTotal RNA was harvested at the times indicated and processedfor northern blot analysis The mRNA signals were quantifiedby PhosphorImager (B) The remaining maspin mRNA tran-scripts normalized to GAPDH loading control were plottedversus time
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL
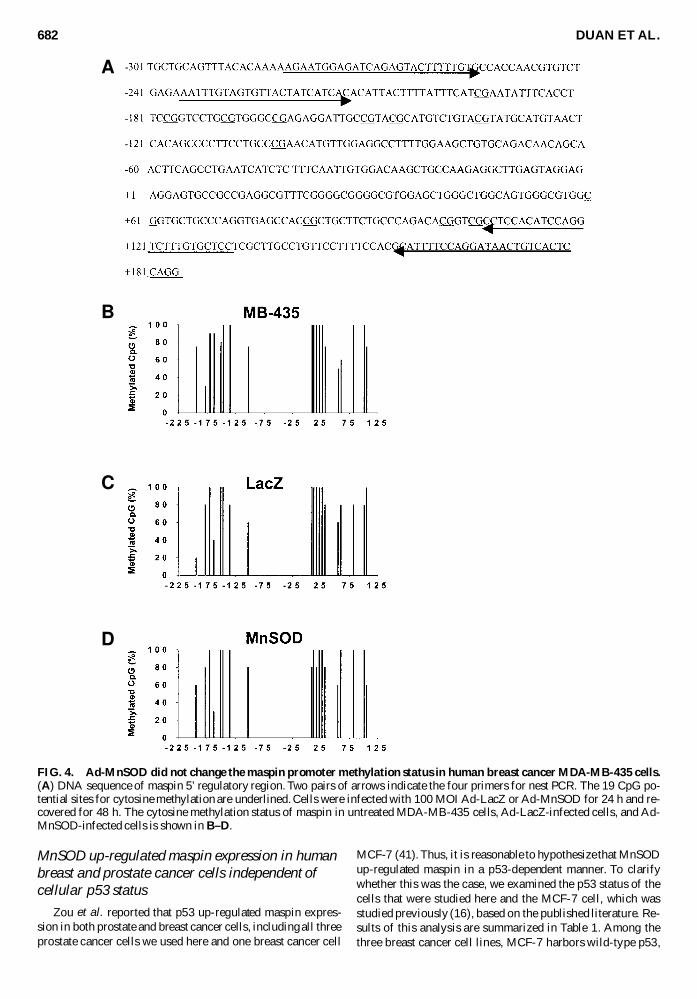
MnSOD up-regulated maspin expression in humanbreast and prostate cancer cells independent ofcellular p53 status
Zou et al reported that p53 up-regulated maspin expres-sion in both prostate and breast cancer cells including all threeprostate cancer cells we used here and one breast cancer cell
MCF-7 (41) Thus it is reasonable to hypothesize that MnSODup-regulated maspin in a p53-dependent manner To clarifywhether this was the case we examined the p53 status of thecells that were studied here and the MCF-7 cell which wasstudied previously (16) based on the published literature Re-sults of this analysis are summarized in Table 1 Among thethree breast cancer cell lines MCF-7 harbors wild-type p53
682 DUAN ET AL
FIG 4 Ad-MnSOD did not change the maspin promoter methylation status in human breast cancer MDA-MB-435 cells(A) DNA sequence of maspin 5rsquo regulatory region Two pairs of arrows indicate the four primers for nest PCR The 19 CpG po-tential sites for cytosine methylation are underlined Cells were infected with 100 MOI Ad-LacZ or Ad-MnSOD for 24 h and re-covered for 48 h The cytosine methylation status of maspin in untreated MDA-MB-435 cells Ad-LacZ-infected cells and Ad-MnSOD-infected cells is shown in BndashD
A
B
C
D
whereas MDA-MB-435 and MDA-MB-231 cells harbor mu-tant p53 Among the prostate cancer cell lines LNCaP har-bors wild-type p53 expression whereas DU145 has a mutantp53 expression and PC3 is a p53 null cell line Therefore theup-regulation effect of MnSOD on maspin mRNA expressionwas independent of cellular p53 status
MnSOD did not influence maspin promoter activityor mRNA transcription rate in PC3 cells
Several studies have shown that maspin promoter activityis abolished or down-regulated in human breast and prostatecancer cells compared with their normal counterpart (34 35)To elucidate whether the effect of MnSOD is due to activationof the maspin promoter we measured maspin promoter activ-ity in transient and stable MnSOD overexpression modelsThe maspin promoter luciferase reporter construct pGL3-MPconsists of the 5rsquo regulatory region from 2396 to +131 rela-tive to the maspin transcription start (Fig 5A) This regionwhich contains the binding sites of Ets p53 and AP-1 hasbeen characterized to be necessary and sufficient for the maspintransactivation (34) To test the function of this construct wefirst transfected it into an immortalized but nontumorigenichuman breast epithelial cell line MCF-10A that constitutivelyexpresses a high level of maspin As shown in Fig 5B maspinpromoter activity in MCF-10A cells was higher than the ac-tivity of pGL3-Basic construct Compared with the promoteractivity in normal MCF-10A cells the maspin promoter wasless active in human breast cancer cells MDA-MB-435 andZR-75-1 To determine whether MnSOD up-regulated themaspin promoter we cotransfected pGL3-MP and an MnSODexpression vector pcDNA3-MnSOD into MDA-MB-435 andDU145 cells As shown in Fig 5C MnSOD had no effect onmaspin promoter activity in either of the two cell lines We alsomeasured maspin promoter activity in MnSOD stable trans-fectants SOD15 Mn1 and Mn40 which were derived fromMCF-7 human breast cancer cells (33) The MnSOD enzymeactivities in the stable transfectants are 19 62 and 230 Umgof protein for SOD15 Mn1 and Mn40 respectively How-ever no significant changes were observed in maspin promoteractivity among the three MnSOD overexpressing cell lines(Fig 5D)
To explore further whether MnSOD modulates maspin mRNAexpression at the transcriptional level we measured the maspin
mRNA in vitro transcription rate The nuclear run-on assayresults in the Ad-MnSOD- and Ad-LacZ-infected PC3 cellsare shown in Fig 6 Again we detected an almost equal amountof the newly transcribed maspin mRNA signal in the two groupsThis result indicates that MnSOD has no effect on the rate oftranscriptional initiation of maspin as determined by this invitro transcription assay
MnSOD increased maspin mRNA stability in PC3 cells
Much attention has been focused on the transcriptional reg-ulation of maspin in normal and cancer cells Thus far an ef-fect of maspin mRNA stability has not been explored In thisstudy we examined maspin mRNA stability in PC3 cells whichhave the most abundant endogenous maspin mRNA expres-sion among the cancer cells we used here We inhibited newmaspin mRNA synthesis by treating the cells with 10 microgmlactinomycin D and measured the residual levels of the 30-kband 12-kb maspin transcripts at various time points A repre-sentative northern blot result is shown in Fig 7A Quantita-tive analysis of the maspin half-life based on two independentexperiments is shown in Fig 7B It shows that the half-life ofthe 30-kb transcript was ~20 h and the half-life of 12-kbtranscript was much longer ~30ndash36 h
To determine whether the effect of MnSOD on the up-regulation of maspin mRNA was due to the modulation ofmaspin mRNA stability we infected the PC3 cells with 100MOI Ad-LacZ and Ad-MnSOD then treated cells with actin-omycin D and assessed maspin mRNA levels at each timepoint The northern blot results are shown in Fig 8A Quanti-tative analysis of the maspin mRNA remaining as a functionof time is shown in Fig 8B for the 30-kb transcript and Fig8C for the 12-kb transcript These data suggest that MnSODincreased the stability of both transcripts compared with theAd-LacZ infection control For the 30-kb transcript the resid-ual maspin mRNA dropped to 30 within 36 h In contrastthe 12-kb transcript consistently remained at nearly 80 from12 to 36 hours Therefore the stabilizing effect of MnSODexpression on the 12-kb transcript was more significant thanthat on the 30-kb transcript Taking into account that the 12-kb transcript has a longer half-life it might explain why weobserved a significant accumulation of 12-kb transcript butnot the 30-kb transcript after MnSOD overexpression
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 683
TABLE 1 OVERVIEW OF MASPIN PROMOTER METHYLATION AND P53STATUS IN HUMAN BREAST AND PROSTATE CANCER CELL LINES
Cell types p53 status Promoter methylation status (6)
Breast cancer cellsMDA-MB-435 mt-p53 +MDA-MB-231 mt-p53 +MCF-7 wt-p53 +
Prostate cancer cellsPC3 p53(22) 2DU145 mt-p53 2LNCaP wt-p53 2
mt mutant wt wild type
DISCUSSION
Tumor cells have often been found to display lower levelsof antioxidant enzyme expression than their normal counter-parts For example tumor cells are nearly always low in MnSODand catalase activity and usually low in copperzinc superox-ide dismutase activity whereas glutathione peroxidase activ-ity is variable (3 25 33) MnSOD is a nuclear-encoded mito-chondrial matrix protein and serves a protective function by
684 DUAN ET AL
FIG 5 Maspin promoter activity in human breast andprostate cancer cells (A) Schematic representation of maspinpromoter construct with the important cis-elements labeled inthe shaded areas (B) Maspin promoter activity was down-regulated in human breast cancer cells MCF-10A and MDA-MB-435 cells were transiently transfected with 2 microg of maspinpromoter construct pGL3-MP or pGL3-Basic together with 1microg of pCMV-b-galactosidase as internal control ZR-75ndash1 cellswere transfected by electroporation Ten micrograms of pGL3-MP or pGL3-basic was transfected into 5 3 107 cells with 5 microgof pCMV-b-galactosidase as the internal control The relativeluciferase activity normalized by b-galactosidase activity wasrepresented as the fold increase compared with pGL3-Basicvector activity in each cell types (C) MnSOD did not influencemaspin promoter activity in human breast cancer MDA-MB-435 cells and human prostate cancer DU145 cells Two micro-grams of pcDNA3-MnSOD expression plasmid was cotrans-fected with 2 microg of pGL3-MP The relative maspin promoteractivity in the control cells was designated as 100 (D)Maspin promoter activity in MnSOD transfected cells was rep-resented as a percentage increase compared with control Re-sults shown here are the means and standard deviations fromtwo independent transfection experiments
FIG 6 Influence of MnSOD on the maspin mRNA invitro transcription rate in PC3 cells Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h total RNA was isolated and put into in vitro tran-scription analysis [32P]-UTP-labeled mRNA samples were hy-bridized with slot-blotted 2 microg of maspin and GAPDH cDNA32P signals were detected and quantified by PhosphorImager
FIG 7 Maspin mRNA stability in human prostate cancerPC3 cells (A) Northern blot analysis for maspin mRNA stabil-ity assay Cells were treated with 10 microgml actinomycin DTotal RNA was harvested at the times indicated and processedfor northern blot analysis The mRNA signals were quantifiedby PhosphorImager (B) The remaining maspin mRNA tran-scripts normalized to GAPDH loading control were plottedversus time
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL

whereas MDA-MB-435 and MDA-MB-231 cells harbor mu-tant p53 Among the prostate cancer cell lines LNCaP har-bors wild-type p53 expression whereas DU145 has a mutantp53 expression and PC3 is a p53 null cell line Therefore theup-regulation effect of MnSOD on maspin mRNA expressionwas independent of cellular p53 status
MnSOD did not influence maspin promoter activityor mRNA transcription rate in PC3 cells
Several studies have shown that maspin promoter activityis abolished or down-regulated in human breast and prostatecancer cells compared with their normal counterpart (34 35)To elucidate whether the effect of MnSOD is due to activationof the maspin promoter we measured maspin promoter activ-ity in transient and stable MnSOD overexpression modelsThe maspin promoter luciferase reporter construct pGL3-MPconsists of the 5rsquo regulatory region from 2396 to +131 rela-tive to the maspin transcription start (Fig 5A) This regionwhich contains the binding sites of Ets p53 and AP-1 hasbeen characterized to be necessary and sufficient for the maspintransactivation (34) To test the function of this construct wefirst transfected it into an immortalized but nontumorigenichuman breast epithelial cell line MCF-10A that constitutivelyexpresses a high level of maspin As shown in Fig 5B maspinpromoter activity in MCF-10A cells was higher than the ac-tivity of pGL3-Basic construct Compared with the promoteractivity in normal MCF-10A cells the maspin promoter wasless active in human breast cancer cells MDA-MB-435 andZR-75-1 To determine whether MnSOD up-regulated themaspin promoter we cotransfected pGL3-MP and an MnSODexpression vector pcDNA3-MnSOD into MDA-MB-435 andDU145 cells As shown in Fig 5C MnSOD had no effect onmaspin promoter activity in either of the two cell lines We alsomeasured maspin promoter activity in MnSOD stable trans-fectants SOD15 Mn1 and Mn40 which were derived fromMCF-7 human breast cancer cells (33) The MnSOD enzymeactivities in the stable transfectants are 19 62 and 230 Umgof protein for SOD15 Mn1 and Mn40 respectively How-ever no significant changes were observed in maspin promoteractivity among the three MnSOD overexpressing cell lines(Fig 5D)
To explore further whether MnSOD modulates maspin mRNAexpression at the transcriptional level we measured the maspin
mRNA in vitro transcription rate The nuclear run-on assayresults in the Ad-MnSOD- and Ad-LacZ-infected PC3 cellsare shown in Fig 6 Again we detected an almost equal amountof the newly transcribed maspin mRNA signal in the two groupsThis result indicates that MnSOD has no effect on the rate oftranscriptional initiation of maspin as determined by this invitro transcription assay
MnSOD increased maspin mRNA stability in PC3 cells
Much attention has been focused on the transcriptional reg-ulation of maspin in normal and cancer cells Thus far an ef-fect of maspin mRNA stability has not been explored In thisstudy we examined maspin mRNA stability in PC3 cells whichhave the most abundant endogenous maspin mRNA expres-sion among the cancer cells we used here We inhibited newmaspin mRNA synthesis by treating the cells with 10 microgmlactinomycin D and measured the residual levels of the 30-kband 12-kb maspin transcripts at various time points A repre-sentative northern blot result is shown in Fig 7A Quantita-tive analysis of the maspin half-life based on two independentexperiments is shown in Fig 7B It shows that the half-life ofthe 30-kb transcript was ~20 h and the half-life of 12-kbtranscript was much longer ~30ndash36 h
To determine whether the effect of MnSOD on the up-regulation of maspin mRNA was due to the modulation ofmaspin mRNA stability we infected the PC3 cells with 100MOI Ad-LacZ and Ad-MnSOD then treated cells with actin-omycin D and assessed maspin mRNA levels at each timepoint The northern blot results are shown in Fig 8A Quanti-tative analysis of the maspin mRNA remaining as a functionof time is shown in Fig 8B for the 30-kb transcript and Fig8C for the 12-kb transcript These data suggest that MnSODincreased the stability of both transcripts compared with theAd-LacZ infection control For the 30-kb transcript the resid-ual maspin mRNA dropped to 30 within 36 h In contrastthe 12-kb transcript consistently remained at nearly 80 from12 to 36 hours Therefore the stabilizing effect of MnSODexpression on the 12-kb transcript was more significant thanthat on the 30-kb transcript Taking into account that the 12-kb transcript has a longer half-life it might explain why weobserved a significant accumulation of 12-kb transcript butnot the 30-kb transcript after MnSOD overexpression
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 683
TABLE 1 OVERVIEW OF MASPIN PROMOTER METHYLATION AND P53STATUS IN HUMAN BREAST AND PROSTATE CANCER CELL LINES
Cell types p53 status Promoter methylation status (6)
Breast cancer cellsMDA-MB-435 mt-p53 +MDA-MB-231 mt-p53 +MCF-7 wt-p53 +
Prostate cancer cellsPC3 p53(22) 2DU145 mt-p53 2LNCaP wt-p53 2
mt mutant wt wild type
DISCUSSION
Tumor cells have often been found to display lower levelsof antioxidant enzyme expression than their normal counter-parts For example tumor cells are nearly always low in MnSODand catalase activity and usually low in copperzinc superox-ide dismutase activity whereas glutathione peroxidase activ-ity is variable (3 25 33) MnSOD is a nuclear-encoded mito-chondrial matrix protein and serves a protective function by
684 DUAN ET AL
FIG 5 Maspin promoter activity in human breast andprostate cancer cells (A) Schematic representation of maspinpromoter construct with the important cis-elements labeled inthe shaded areas (B) Maspin promoter activity was down-regulated in human breast cancer cells MCF-10A and MDA-MB-435 cells were transiently transfected with 2 microg of maspinpromoter construct pGL3-MP or pGL3-Basic together with 1microg of pCMV-b-galactosidase as internal control ZR-75ndash1 cellswere transfected by electroporation Ten micrograms of pGL3-MP or pGL3-basic was transfected into 5 3 107 cells with 5 microgof pCMV-b-galactosidase as the internal control The relativeluciferase activity normalized by b-galactosidase activity wasrepresented as the fold increase compared with pGL3-Basicvector activity in each cell types (C) MnSOD did not influencemaspin promoter activity in human breast cancer MDA-MB-435 cells and human prostate cancer DU145 cells Two micro-grams of pcDNA3-MnSOD expression plasmid was cotrans-fected with 2 microg of pGL3-MP The relative maspin promoteractivity in the control cells was designated as 100 (D)Maspin promoter activity in MnSOD transfected cells was rep-resented as a percentage increase compared with control Re-sults shown here are the means and standard deviations fromtwo independent transfection experiments
FIG 6 Influence of MnSOD on the maspin mRNA invitro transcription rate in PC3 cells Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h total RNA was isolated and put into in vitro tran-scription analysis [32P]-UTP-labeled mRNA samples were hy-bridized with slot-blotted 2 microg of maspin and GAPDH cDNA32P signals were detected and quantified by PhosphorImager
FIG 7 Maspin mRNA stability in human prostate cancerPC3 cells (A) Northern blot analysis for maspin mRNA stabil-ity assay Cells were treated with 10 microgml actinomycin DTotal RNA was harvested at the times indicated and processedfor northern blot analysis The mRNA signals were quantifiedby PhosphorImager (B) The remaining maspin mRNA tran-scripts normalized to GAPDH loading control were plottedversus time
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL
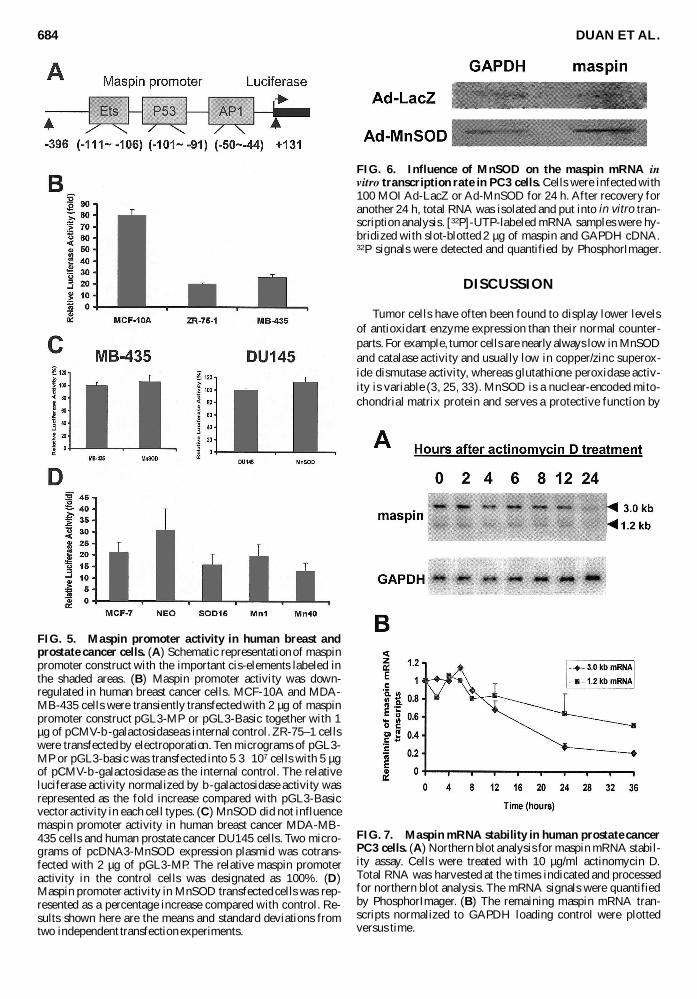
DISCUSSION
Tumor cells have often been found to display lower levelsof antioxidant enzyme expression than their normal counter-parts For example tumor cells are nearly always low in MnSODand catalase activity and usually low in copperzinc superox-ide dismutase activity whereas glutathione peroxidase activ-ity is variable (3 25 33) MnSOD is a nuclear-encoded mito-chondrial matrix protein and serves a protective function by
684 DUAN ET AL
FIG 5 Maspin promoter activity in human breast andprostate cancer cells (A) Schematic representation of maspinpromoter construct with the important cis-elements labeled inthe shaded areas (B) Maspin promoter activity was down-regulated in human breast cancer cells MCF-10A and MDA-MB-435 cells were transiently transfected with 2 microg of maspinpromoter construct pGL3-MP or pGL3-Basic together with 1microg of pCMV-b-galactosidase as internal control ZR-75ndash1 cellswere transfected by electroporation Ten micrograms of pGL3-MP or pGL3-basic was transfected into 5 3 107 cells with 5 microgof pCMV-b-galactosidase as the internal control The relativeluciferase activity normalized by b-galactosidase activity wasrepresented as the fold increase compared with pGL3-Basicvector activity in each cell types (C) MnSOD did not influencemaspin promoter activity in human breast cancer MDA-MB-435 cells and human prostate cancer DU145 cells Two micro-grams of pcDNA3-MnSOD expression plasmid was cotrans-fected with 2 microg of pGL3-MP The relative maspin promoteractivity in the control cells was designated as 100 (D)Maspin promoter activity in MnSOD transfected cells was rep-resented as a percentage increase compared with control Re-sults shown here are the means and standard deviations fromtwo independent transfection experiments
FIG 6 Influence of MnSOD on the maspin mRNA invitro transcription rate in PC3 cells Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h total RNA was isolated and put into in vitro tran-scription analysis [32P]-UTP-labeled mRNA samples were hy-bridized with slot-blotted 2 microg of maspin and GAPDH cDNA32P signals were detected and quantified by PhosphorImager
FIG 7 Maspin mRNA stability in human prostate cancerPC3 cells (A) Northern blot analysis for maspin mRNA stabil-ity assay Cells were treated with 10 microgml actinomycin DTotal RNA was harvested at the times indicated and processedfor northern blot analysis The mRNA signals were quantifiedby PhosphorImager (B) The remaining maspin mRNA tran-scripts normalized to GAPDH loading control were plottedversus time
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL
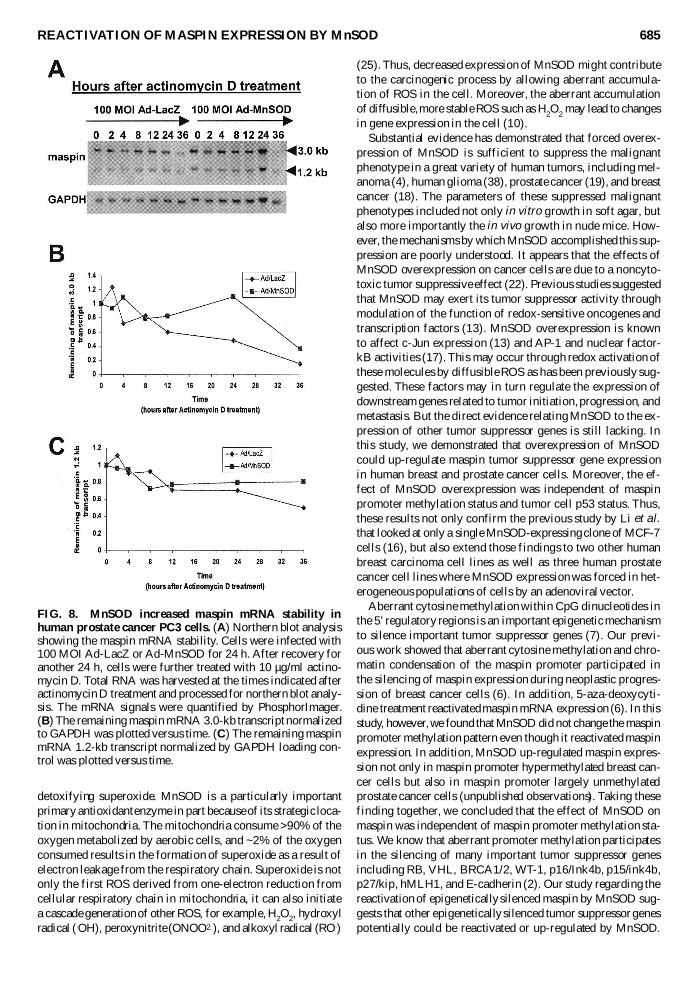
detoxifying superoxide MnSOD is a particularly importantprimary antioxidant enzyme in part because of its strategic loca-tion in mitochondria The mitochondria consume gt90 of theoxygen metabolized by aerobic cells and ~2 of the oxygenconsumed results in the formation of superoxide as a result ofelectron leakage from the respiratory chain Superoxide is notonly the first ROS derived from one-electron reduction fromcellular respiratory chain in mitochondria it can also initiatea cascade generation of other ROS for example H2O2 hydroxylradical (middotOH) peroxynitrite (ONOO2) and alkoxyl radical (ROmiddot)
(25) Thus decreased expression of MnSOD might contributeto the carcinogenic process by allowing aberrant accumula-tion of ROS in the cell Moreover the aberrant accumulationof diffusible more stable ROS such as H2O2 may lead to changesin gene expression in the cell (10)
Substantial evidence has demonstrated that forced overex-pression of MnSOD is sufficient to suppress the malignantphenotype in a great variety of human tumors including mel-anoma (4) human glioma (38) prostate cancer (19) and breastcancer (18) The parameters of these suppressed malignantphenotypes included not only in vitro growth in soft agar butalso more importantly the in vivo growth in nude mice How-ever the mechanisms by which MnSOD accomplished this sup-pression are poorly understood It appears that the effects ofMnSOD overexpression on cancer cells are due to a noncyto-toxic tumor suppressive effect (22) Previous studies suggestedthat MnSOD may exert its tumor suppressor activity throughmodulation of the function of redox-sensitive oncogenes andtranscription factors (13) MnSOD overexpression is knownto affect c-Jun expression (13) and AP-1 and nuclear factor-kB activities (17) This may occur through redox activation ofthese molecules by diffusible ROS as has been previously sug-gested These factors may in turn regulate the expression ofdownstream genes related to tumor initiation progression andmetastasis But the direct evidence relating MnSOD to the ex-pression of other tumor suppressor genes is still lacking Inthis study we demonstrated that overexpression of MnSODcould up-regulate maspin tumor suppressor gene expressionin human breast and prostate cancer cells Moreover the ef-fect of MnSOD overexpression was independent of maspinpromoter methylation status and tumor cell p53 status Thusthese results not only confirm the previous study by Li et althat looked at only a single MnSOD-expressing clone of MCF-7cells (16) but also extend those findings to two other humanbreast carcinoma cell lines as well as three human prostatecancer cell lines where MnSOD expression was forced in het-erogeneous populations of cells by an adenoviral vector
Aberrant cytosine methylation within CpG dinucleotides inthe 5rsquo regulatory regions is an important epigenetic mechanismto silence important tumor suppressor genes (7) Our previ-ous work showed that aberrant cytosine methylation and chro-matin condensation of the maspin promoter participated inthe silencing of maspin expression during neoplastic progres-sion of breast cancer cells (6) In addition 5-aza-deoxycyti-dine treatment reactivated maspin mRNA expression (6) In thisstudy however we found that MnSOD did not change the maspinpromoter methylation pattern even though it reactivated maspinexpression In addition MnSOD up-regulated maspin expres-sion not only in maspin promoter hypermethylated breast can-cer cells but also in maspin promoter largely unmethylatedprostate cancer cells (unpublished observations) Taking thesefinding together we concluded that the effect of MnSOD onmaspin was independent of maspin promoter methylation sta-tus We know that aberrant promoter methylation participatesin the silencing of many important tumor suppressor genesincluding RB VHL BRCA12 WT-1 p16Ink4b p15ink4bp27kip hMLH1 and E-cadherin (2) Our study regarding thereactivation of epigenetically silenced maspin by MnSOD sug-gests that other epigenetically silenced tumor suppressor genespotentially could be reactivated or up-regulated by MnSOD
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 685
FIG 8 MnSOD increased maspin mRNA stability inhuman prostate cancer PC3 cells (A) Northern blot analysisshowing the maspin mRNA stability Cells were infected with100 MOI Ad-LacZ or Ad-MnSOD for 24 h After recovery foranother 24 h cells were further treated with 10 microgml actino-mycin D Total RNA was harvested at the times indicated afteractinomycin D treatment and processed for northern blot analy-sis The mRNA signals were quantified by PhosphorImager(B) The remaining maspin mRNA 30-kb transcript normalizedto GAPDH was plotted versus time (C) The remaining maspinmRNA 12-kb transcript normalized by GAPDH loading con-trol was plotted versus time
Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL

Indeed recent results of microarray expression studies inMnSOD-overexpressing cells would support this hypothesis(10) These effects may explain in part the mechanisms ofMnSODrsquos extensive antitumor effect in tumor cells
Zou et al reported that p53 regulated expression of maspinin both breast and prostate cancer cells (41) They found thatp53 activated the maspin promoter by binding directly to a p53consensus site present in the maspin promoter DNA-damagingagents and cytotoxic drugs induced endogenous maspin ex-pression in cells containing the wild-type p53 Maspin expres-sion was refractory to the DNA-damaging agents in cells con-taining mutant p53 Thus it seems that wild-type p53 is a criticalupstream regulator of maspin However our study showedthat MnSOD up-regulated maspin expression not only in p53wild-type cells LNCaP but also in p53 mutant cells (MDA-MB-435 MDA-MB-231 PC3 and DU145) Taken togetherthese studies strongly suggest that even though maspin is adownstream effecter of MnSOD functional p53 is not a nec-essary mediator between them
The level of specific mRNA species depends on either theirnew transcription or posttranscriptional degradation or bothOur study in PC3 human prostate carcinoma cells showed thatthe effect of MnSOD on the up-regulation of maspin expres-sion was through increasing maspin mRNA stability but notincreasing its transcription We did not measure the effect ofMnSOD on the maspin mRNA stability and in vitro transcrip-tion in human breast cancer cells due to the undetectable en-dogenous maspin mRNA expression But we predicted thatincreasing maspin mRNA stability might also be the majormechanism for its accumulation after MnSOD treatment inhuman breast cancer cells As shown in Fig 5 maspin pro-moter activity is down-regulated in human breast cancer cellsMDA-MB-435 and ZR-75-1 Consistent with our study Zhanget al showed that the maspin promoter transactivation activ-ity was decreased in primary tumor cells and was abolishedin MDA-MB-231 cells (34) The aberrantly hypermethylatedmaspin promoter is maintained in a closed chromatin struc-ture and likely inaccessible to transcriptional factors that mayotherwise bind to their binding sites in the promoter regionEpigenetic silencing of the maspin promoter is one of the majormechanisms for the loss or down-regulation of the maspin ex-pression in human breast cancer cells MnSOD could notdemethylate the maspin promoter in MDA-MB-435 humanbreast cancer cells so it is unlikely that MnSOD can renderthe maspin promoter more accessible and lead to a strongertransactivation The possibility still exists however thatMnSOD expression could lead to other epigenetic changes atthe maspin promoter that render it more accessible to tran-scriptional transactivation as has recently been shown forp53-mediated activation of maspin expression (26) Never-theless our promoter reporter assay in MDA-MB-435 cellsshowing that MnSOD did not increase maspin promoter ac-tivity as well as the results from our nuclear run-on studiesall support the conclusion that it is unlikely that MnSOD in-creases maspin transcription
Recently the posttranscriptional regulation of mRNA sta-bility has been identified as an important control mechanismof gene expression Our study here demonstrated that MnSODincreased maspin mRNA accumulation through increasingmaspin mRNA stability and therefore identified a possiblemechanism that contributes to the regulation of other tumor
suppressor genes by MnSOD However increasing mRNAstability is not the only effect of MnSOD on other genes Me-lendez and Davies reported several years ago that MnSOD andhypoxia modulated interleukin-1a levels in HT-1080 fibro-sarcoma cells by destabilizing interleukin-1a mRNA (21)Although the mechanisms that alter mRNA stability of differ-ent genes have unique features in general sequences locatedin the 3rsquo-UTR and their interactions with specific proteins reg-ulate mRNA stability These motifs could mediate the stabi-lization or destabilization of individual mRNA depending onthe specific protein factors binding to them The binding ofsome protein factors to the 3rsquo-UTR are redox-sensitive (9 20)and the activities of these modifiers of posttranscriptional mRNAstability could be affected either directly or indirectly by dif-fusible ROS such as H2O2 the product of MnSOD Definingthe redox-sensitive cis-element in the 3rsquo-UTR and protein fac-tors binding to maspin mRNA should be an interesting area toexplore in the elucidation of redox regulation of maspin byMnSOD
Perspectives
In summary MnSOD overexpression up-regulates maspintumor suppressor gene expression in human breast and pros-tate cancer cells The effect of MnSOD on the up-regulationof maspin expression is mainly due to the posttranscriptionaleffect of increasing maspin mRNA stability Moreover the ef-fect of MnSOD on maspin expression is independent of maspinpromoter methylation status and tumor cell p53 status As theloss of p53 function and epigenetic silencing of tumor sup-pressor genes are two critical determinants of gene expres-sion and response to tumor therapy our f indings provide a ra-tionale for the use of MnSOD as a gene therapy target to treathuman breast and prostate tumors
ACKNOWLEDGMENTS
This research was supported by Public Health Servicegrants CA65662 to BWF CA73612 to FED and CA66081to LWO
ABBREVIATIONS
Ad-LacZ recombinant adenovirus containing b-galactosi-dase cDNA Ad-MnSOD recombinant adenovirus containingMnSOD cDNA GAPDH glyceraldehyde-3-phosphate dehy-drogenase H2O2 hydrogen peroxide HRP horseradish per-oxidase MnSOD manganese superoxide dismutase MOImultiplicity of infection PAGE polyacrylamide gel electro-phoresis PBS phosphate-buffered saline ROS reactive oxy-gen species SDS sodium dodecyl sulfate SSC salinendashsodiumcitrate buffer 3rsquo-UTR 3rsquo-untranslated region
REFERENCES
1 Amstad PA Liu H Ichimiya M Berezesky IK and TrumpBF Manganese superoxide dismutase expression inhibits
686 DUAN ET AL
soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL

soft agar growth in JB6 clone41 mouse epidermal cellsCarcinogenesis 18 479ndash484 1997
2 Baylin SB Herman JG Graff JR Vertino PM and Issa JPAlterations in DNA methylation a fundamental aspect ofneoplasia Adv Cancer Res 72 141ndash196 1998
3 Cerutti PA Prooxidant states and tumor promotion Sci-ence 227 375ndash381 1985
4 Church SL Grant JW Ridnour LA Oberley LW SwansonPE Meltzer PS and Trent JM Increased manganese su-peroxide dismutase expression suppresses the malignantphenotype of human melanoma cells Proc Natl Acad SciU S A 90 3113ndash3117 1993
5 Clark SJ Harrison J Paul CL and Frommer M High sen-sitivity mapping of methylated cytosines Nucleic AcidsRes 22 2990ndash2997 1994
6 Domann FE Rice JC Hendrix MJ and Futscher BW Epi-genetic silencing of maspin gene expression in humanbreast cancers Int J Cancer 85 805ndash810 2000
7 Feinberg AP DNA methylation genomic imprinting andcancer Curr Top Microbiol Immunol 249 87ndash99 2000
8 Futscher BW Oshiro MM Wozniak RJ Holtan N Hani-gan CL Duan H and Domann FE Role for DNA methyla-tion in the control of cell type specific maspin expressionNat Genet 31 175ndash179 2002
9 Goswami PC Sheren J Albee LD Parsian A Sim JE Rid-nour LA Higashikubo R Gius D Hunt CR and Spitz DRCell cycle-coupled variation in topoisomerase IIalphamRNA is regulated by the 3rsquo-untranslated region Possiblerole of redox-sensitive protein binding in mRNA accumu-lation J Biol Chem 275 38384ndash38392 2000
10 Guo G Yan-Sanders Y Lyn-Cook BD Wang T Tamae DOgi J Khaletskiy A Li Z Weydert C Longmate JAHuang T-T Spitz DR Oberley LW and Li JJ Manganesesuperoxide dismutase-mediated gene expression in radia-tion-induced adaptive responses Mol Cell Biol 23 2362ndash2378 2003
11 Izutani R Asano S Imano M Kuroda D Kato M andOhyanagi H Expression of manganese superoxide dismu-tase in esophageal and gastric cancers J Gastroenterol 33816ndash822 1998
12 Kensler TW Bush DM and Kozumbo WJ Inhibition oftumor promotion by a biomimetic superoxide dismutaseScience 221 75ndash77 1983
13 Kiningham KK and St Clair DK Overexpression of man-ganese superoxide dismutase selectively modulates the ac-tivity of Jun-associated transcription factors in fibrosar-coma cells Cancer Res 57 5265ndash5271 1997
14 Lam EW Zwacka R Engelhardt JF Davidson BL Do-mann FE Jr Yan T and Oberley LW Adenovirus-mediatedmanganese superoxide dismutase gene transfer to hamstercheek pouch carcinoma cells Cancer Res 57 5550ndash55561997
15 Lam EW Zwacka R Seftor EA Nieva DR Davidson BLEngelhardt JF Hendrix MJ and Oberley LW Effects of an-tioxidant enzyme overexpression on the invasive pheno-type of hamster cheek pouch carcinoma cells Free RadicBiol Med 27 572ndash579 1999
16 Li JJ Colburn NH and Oberley LW Maspin gene expres-sion in tumor suppression induced by overexpressing man-ganese-containing superoxide dismutase cDNA in humanbreast cancer cells Carcinogenesis 19 833ndash839 1998
17 Li JJ Oberley LW Fan M and Colburn NH Inhibition ofAP-1 and NF-kappaB by manganese-containing superox-ide dismutase in human breast cancer cells FASEB J 121713ndash1723 1998
18 Li JJ Oberley LW St Clair DK Ridnour LA and OberleyTD Phenotypic changes induced in human breast cancercells by overexpression of manganese-containing superox-ide dismutase Oncogene 10 1989ndash2000 1995
19 Li N Oberley TD Oberley LW and Zhong W Overexpres-sion of manganese superoxide dismutase in DU145 humanprostate carcinoma cells has multiple effects on cell phe-notype Prostate 35 221ndash233 1998
20 Malter JS and Hong Y A redox switch and phosphoryla-tion are involved in the posttranslational up-regulation ofthe adenosine-uridine binding factor by phorbol ester andionophore J Biol Chem 266 3167ndash3171 1991
21 Melendez JA and Davies KJ Manganese superoxide dis-mutase modulates interleukin-1alpha levels in HT-1080 fi-brosarcoma cells J Biol Chem 271 18898ndash18903 1996
22 Oberley LW Anticancer therapy by overexpression of su-peroxide dismutase Antioxid Redox Signal 3 461ndash4722001
23 Oberley LW and Buettner GR Role of superoxide dismu-tase in cancer a review Cancer Res 39 1141ndash1149 1979
24 Oberley LW and Oberley TD Role of antioxidant enzymesin the cancer phenotype In Oxygen Gene Expressionand Cellular Function edited by Clerch L and Massaro DNew York NY Marcel Dekker Inc 1997 pp 279ndash307
25 Oberley LW and Oberley TD The role of superoxide dis-mutase and gene amplification in carcinogenesis J TheorBiol 106 403ndash422 1984
26 Oshiro MM Watts GS Wozniak RJ Junk DJ Munoz-Rodriguez JL Domann FE and Futscher BW Mutant p53and aberrant cytosine methylation cooperate to silence geneexpression Oncogene 22 3624ndash3634 2003
27 Sager R Sheng S Pemberton P and Hendrix MJ Maspina tumor suppressing serpin Curr Top Microbiol Immunol213 51ndash64 1996
28 Sheng S Carey J Seftor EA Dias L Hendrix MJ andSager R Maspin acts at the cell membrane to inhibit inva-sion and motility of mammary and prostatic cancer cellsProc Natl Acad Sci U S A 93 11669ndash11674 1996
29 Slaga TJ Klein-Szanto AJ Triplett LL Yotti LP andTrosko KE Skin tumor-promoting activity of benzoyl per-oxide a widely used free radical-generating compoundScience 213 1023ndash1025 1981
30 St Clair DK and Holland JC Complementary DNA encod-ing human colon cancer manganese superoxide dismutaseand the expression of its gene in human cells Cancer Res51 939ndash943 1991
31 Sun Y Colburn NH and Oberley LW Decreased expres-sion of manganese superoxide dismutase mRNA and pro-tein after immortalization and transformation of mouseliver cells Oncol Res 5 127ndash132 1993
32 Weisiger RA and Fridovich I Mitochondrial superoxidesimutase Site of synthesis and intramitochondrial local-ization J Biol Chem 248 4793ndash4796 1973
33 Zhang HJ Yan T Oberley TD and Oberley LW Compari-son of effects of two polymorphic variants of manganesesuperoxide dismutase on human breast MCF-7 cancer cellphenotype Cancer Res 59 6276ndash6283 1999
REACTIVATION OF MASPIN EXPRESSION BY MnSOD 687
34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL

34 Zhang M Maass N Magit D and Sager R Transactivationthrough Ets and Ap1 transcription sites determines the ex-pression of the tumor-suppressing gene maspin Cell GrowthDiffer 8 179ndash186 1997
35 Zhang M Magit D and Sager R Expression of maspin inprostate cells is regulated by a positive ets element and anegative hormonal responsive element site recognized byandrogen receptor Proc Natl Acad Sci U S A 94 5673ndash5678 1997
36 Zhang M Sheng S Maass N and Sager R mMaspin themouse homolog of a human tumor suppressor gene in-hibits mammary tumor invasion and motility Mol Med 349ndash59 1997
37 Zhang M Volpert O Shi YH and Bouck N Maspin is anangiogenesis inhibitor Nat Med 6 196ndash199 2000
38 Zhong W Oberley LW Oberley TD and St Clair DK Sup-pression of the malignant phenotype of human gliomacells by overexpression of manganese superoxide dismu-tase Oncogene 14 481ndash490 1997
39 Zhong W Yan T Lim R and Oberley LW Expression ofsuperoxide dismutases catalase and glutathione peroxi-
dase in glioma cells Free Radic Biol Med 27 1334ndash13451999
40 Zou Z Anisowicz A Hendrix MJ Thor A Neveu MSheng S Rafidi K Seftor E and Sager R Maspin a serpinwith tumor-suppressing activity in human mammary ep-ithelial cells Science 263 526ndash529 1994
41 Zou Z Gao C Nagaich AK Connell T Saito S Moul JWSeth P Appella E and Srivastava S p53 regulates the ex-pression of the tumor suppressor gene maspin J Biol Chem275 6051ndash6054 2000
Address reprint requests toFrederick E Domann
Free Radical and Radiation Biology ProgramB180 Medical Laboratories
The University of IowaIowa City IA 52242
E-mail frederick-domannuiowaedu
Received for publication July 6 2003 accepted July 15 2003
688 DUAN ET AL
Copyright © 2022 FDOKUMEN




















