Minocycline Modulates Neuroinflammation Independently of Its Antimicrobial Activity in...
16
Immunopathology and Infectious Disease Minocycline Modulates Neuroinflammation Independently of Its Antimicrobial Activity in Staphylococcus aureus-Induced Brain Abscess Tammy Kielian,* Nilufer Esen,* Shuliang Liu,* Nirmal K. Phulwani,* Mohsin M. Syed,* Napoleon Phillips,* Koren Nishina, † Ambrose L. Cheung, † Joseph D. Schwartzman, † and Jorg J. Ruhe ‡ From the Departments of Neurobiology and Developmental Sciences* and Infectious Diseases, ‡ University of Arkansas for Medical Sciences, Little Rock, Arkansas; and the Department of Microbiology and Immunology, † Dartmouth Medical School, Lebanon, New Hampshire Minocycline exerts beneficial immune modulatory ef- fects in several noninfectious neurodegenerative dis- ease models; however , its potential to influence the host immune response during central nervous system bacterial infections , such as brain abscess , has not yet been investigated. Using a minocycline-resistant strain of Staphylococcus aureus to dissect the antibiotic’s bacteriostatic versus immune modulatory effects in a mouse experimental brain abscess model , we found that minocycline significantly reduced mortality rates within the first 24 hours following bacterial expo- sure. This protection was associated with a transient decrease in the expression of several proinflamma- tory mediators , including interleukin-1 and CCL2 (MCP-1). Minocycline was also capable of protecting the brain parenchyma from necrotic damage as evi- dent by significantly smaller abscesses in minocy- cline-treated mice. In addition , minocycline exerted anti-inflammatory effects when administered as late as 3 days following S. aureus infection , which corre- lated with a significant decrease in brain abscess size. Finally , minocycline was capable of partially attenu- ating S. aureus-dependent microglial and astrocyte activation. Therefore, minocycline may afford addi- tional therapeutic benefits extending beyond its an- timicrobial activity for the treatment of central ner- vous system infectious diseases typified by a pathogenic inflammatory component through its ability to balance beneficial versus detrimental in- flammation. (Am J Pathol 2007, 171:1199 –1214; DOI: 10.2353/ajpath.2007.070231) Staphylococcus aureus is a frequently encountered or- ganism among patients with brain abscess, an entity that represents a significant medical problem despite recent advances in diagnosis and therapy. 1–3 Brain abscess has been reported to account for approximately 1 in 10,000 hospital admissions in the United States. 2 The most common sources of brain abscess are direct or indirect cranial infection arising from the paranasal si- nuses, middle ear, and teeth. Other routes include seed- ing of the brain from distant sites of systemic infection (ie, endocarditis or lung abscess) or penetrating trauma to the head. 1,2 Even with recent advances made in imaging techniques, the clinical diagnosis of brain abscess is often delayed, because patients may initially present with nonspecific clinical signs. Another factor complicating diagnosis is the tendency of brain abscesses to appear similar to necrotic tumors on computed tomography and magnetic resonance imaging scans. If not detected early, an abscess has the potential to rupture into the ventricular space, a serious complication with an 80% mortality rate. The current treatment for brain abscess includes surgical drainage to relieve pressure and long- term (6 to 8 weeks) systemic antimicrobial therapy. 4 Treatment can be complicated by the emergence of mul- Supported by the National Institutes of Health, National Institute of Neu- rological Disorders and Stroke grant 2R01 NS40730 (to T.K.), and the National Institute of Neurological Disorders and Stroke-supported Core facility at the University of Arkansas for Medical Sciences (grant P30 NS047546). Support for the Digital and Confocal Microscopy Laboratory at the University of Arkansas for Medical Sciences is provided by National Institutes of Health/IDeA Networks of Biomedical Research Excellence P20 RR6460 and National Institutes of Health/National Center for Re- search Resources S10 RR19395. Accepted for publication June 19, 2007. Address reprint requests to Tammy Kielian, Ph.D., University of Arkan- sas for Medical Sciences, Department of Neurobiology and Developmen- tal Sciences, 4301 W. Markham St., Slot 846, Little Rock, AR 72205. E-mail: [email protected]. The American Journal of Pathology, Vol. 171, No. 4, October 2007 Copyright © American Society for Investigative Pathology DOI: 10.2353/ajpath.2007.070231 1199
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Minocycline Modulates Neuroinflammation Independently of Its Antimicrobial Activity in...

Immunopathology and Infectious Disease
Minocycline Modulates NeuroinflammationIndependently of Its Antimicrobial Activity inStaphylococcus aureus-Induced Brain Abscess
Tammy Kielian,* Nilufer Esen,* Shuliang Liu,*Nirmal K. Phulwani,* Mohsin M. Syed,*Napoleon Phillips,* Koren Nishina,†
Ambrose L. Cheung,† Joseph D. Schwartzman,†
and Jorg J. Ruhe‡
From the Departments of Neurobiology and Developmental
Sciences* and Infectious Diseases,‡ University of Arkansas for
Medical Sciences, Little Rock, Arkansas; and the Department of
Microbiology and Immunology,† Dartmouth Medical School,
Lebanon, New Hampshire
Minocycline exerts beneficial immune modulatory ef-fects in several noninfectious neurodegenerative dis-ease models; however, its potential to influence thehost immune response during central nervous systembacterial infections, such as brain abscess, has not yetbeen investigated. Using a minocycline-resistant strainof Staphylococcus aureus to dissect the antibiotic’sbacteriostatic versus immune modulatory effects in amouse experimental brain abscess model, we foundthat minocycline significantly reduced mortality rateswithin the first 24 hours following bacterial expo-sure. This protection was associated with a transientdecrease in the expression of several proinflamma-tory mediators, including interleukin-1� and CCL2(MCP-1). Minocycline was also capable of protectingthe brain parenchyma from necrotic damage as evi-dent by significantly smaller abscesses in minocy-cline-treated mice. In addition, minocycline exertedanti-inflammatory effects when administered as lateas 3 days following S. aureus infection, which corre-lated with a significant decrease in brain abscess size.Finally, minocycline was capable of partially attenu-ating S. aureus-dependent microglial and astrocyteactivation. Therefore , minocycline may afford addi-tional therapeutic benefits extending beyond its an-timicrobial activity for the treatment of central ner-vous system infectious diseases typified by apathogenic inflammatory component through itsability to balance beneficial versus detrimental in-
flammation. (Am J Pathol 2007, 171:1199–1214; DOI:
10.2353/ajpath.2007.070231)
Staphylococcus aureus is a frequently encountered or-ganism among patients with brain abscess, an entity thatrepresents a significant medical problem despite recentadvances in diagnosis and therapy.1–3 Brain abscesshas been reported to account for approximately 1 in10,000 hospital admissions in the United States.2 Themost common sources of brain abscess are direct orindirect cranial infection arising from the paranasal si-nuses, middle ear, and teeth. Other routes include seed-ing of the brain from distant sites of systemic infection (ie,endocarditis or lung abscess) or penetrating trauma tothe head.1,2 Even with recent advances made in imagingtechniques, the clinical diagnosis of brain abscess isoften delayed, because patients may initially present withnonspecific clinical signs. Another factor complicatingdiagnosis is the tendency of brain abscesses to appearsimilar to necrotic tumors on computed tomography andmagnetic resonance imaging scans. If not detectedearly, an abscess has the potential to rupture into theventricular space, a serious complication with an 80%mortality rate. The current treatment for brain abscessincludes surgical drainage to relieve pressure and long-term (6 to 8 weeks) systemic antimicrobial therapy.4
Treatment can be complicated by the emergence of mul-
Supported by the National Institutes of Health, National Institute of Neu-rological Disorders and Stroke grant 2R01 NS40730 (to T.K.), and theNational Institute of Neurological Disorders and Stroke-supported Corefacility at the University of Arkansas for Medical Sciences (grant P30NS047546). Support for the Digital and Confocal Microscopy Laboratoryat the University of Arkansas for Medical Sciences is provided by NationalInstitutes of Health/IDeA Networks of Biomedical Research ExcellenceP20 RR6460 and National Institutes of Health/National Center for Re-search Resources S10 RR19395.
Accepted for publication June 19, 2007.
Address reprint requests to Tammy Kielian, Ph.D., University of Arkan-sas for Medical Sciences, Department of Neurobiology and Developmen-tal Sciences, 4301 W. Markham St., Slot 846, Little Rock, AR 72205.E-mail: [email protected].
The American Journal of Pathology, Vol. 171, No. 4, October 2007
Copyright © American Society for Investigative Pathology
DOI: 10.2353/ajpath.2007.070231
1199

tidrug-resistant strains of S. aureus and decreased cen-tral nervous system (CNS) penetration of alternativedrugs such as vancomycin.5,6 Patients surviving brainabscesses may experience any number of long-termcomplications including seizures, loss of mental acuity,and focal neurological defects that are lesion site-depen-dent.1,2 We have established a mouse experimental brainabscess model using S. aureus in which lesion sites aregreatly exaggerated compared with the relatively local-ized area of initial bacterial growth.3,7–9 This phenome-non is also observed in human disease, where lesionscan encompass a large portion of brain tissue, oftenspreading well beyond the initial focus of infection. Col-lectively, these sequelae are reminiscent of an overactiveimmune response. The continual production of bacteri-cidal compounds by both resident glia and peripheralimmune cells as well as amplification of cytokine net-works and protease production can contribute to the lossof surrounding uninfected tissue. We propose that a bal-ance exists between sufficient and overcompensatoryresponses to S. aureus in the CNS, which dictates theextent of parenchymal damage during brain abscessdevelopment. Indeed, we have found that S. aureus leadsto the immediate and sustained expression of numerousproinflammatory mediators in the brain, indicative of apotent innate immune response.7,8,10,11
Brain abscesses develop in response to a parenchy-mal infection with pyogenic bacteria, beginning as alocalized area of cerebritis and evolving into a suppura-tive lesion surrounded by a well-vascularized fibroticcapsule.1–3 Brain abscesses are typified by extensiveedema and tissue necrosis and tend to localize at white-gray matter junctions where microcirculatory flow ispoor.12,13 In addition to the sequential progression fromcerebritis to necrosis during brain abscess evolution, theactivation of resident glial cells and influx of peripheralleukocytes demonstrate temporal patterns.3,14,15 Specif-ically, microglial and astrocyte activation is evident im-mediately following the entry of bacteria into the CNSparenchyma and persists throughout abscess evolu-tion.11,16 Neutrophils are the initial leukocyte subset toinfiltrate developing abscesses and are observed asearly as 12 hours following bacterial exposure.7,11 Mac-rophages and T cells are associated with lesions as theyprogress, with infiltrates generally more pronouncedaround days 3 and 7, respectively.11,14 Beginning at 7 to10 days postinfection, a highly vascularized fibrotic wallforms around the necrotic milieu, effectively forming abarrier to contain the infection.11,17,18 It is important tonote that the kinetics of cellular activation, influx, andbordering functions represent general time frames, andthere is probably overlap between each of these pro-cesses during brain abscess evolution.
Although necessary for pathogen containment, exces-sive inflammation during the early phases of brain ab-scess development may have deleterious consequenceson disease outcome through the induction of exagger-ated edematous and/or necrotic responses.3,12,13 Similardetrimental effects have been implicated in the patho-physiology of bacterial meningitis, where the dysregu-lated production of free radicals, proinflammatory cyto-
kines, and edema lead to significant neuron damage andassociated mortality.19,20 Therefore, attenuating glial andperipheral immune cell activation without compromisingbacterial containment may result in less damage to nor-mal brain tissue and improvements in cognitive and neu-rological functions.
One attractive candidate to achieve this balance duringthe course of brain abscess development is minocycline.Minocycline is a second-generation tetracycline that effec-tively crosses the blood-brain barrier due to its small (495 d)and lipophilic nature.21–24 In susceptible bacterial strains,minocycline binds to the h34 ribosomal subunit, resulting ina conformational change that effectively interferes with pro-tein synthesis.25,26 Minocycline has been shown to exertneuroprotective effects distinct from its bacteriostatic activ-ity in animal models of cerebral ischemia,27–30 Parkinson’sand Huntington’s disease,31–34 Alzheimer’s disease,35,36
Down’s syndrome,37 amyotrophic lateral sclerosis,38,39 spi-nal cord injury,40–42 and experimental autoimmune en-cephalomyelitis, an animal model for multiple sclero-sis.43–45 Minocycline is thought to modulate these diseases,in part, by inhibiting the production of potentially cytotoxicmolecules by activated microglia.46 For example, minocy-cline has been shown to inhibit microglial production of NO,interleukin (IL)-1�, tumor necrosis factor (TNF)-�, and IL-6,which can impact neuron survival.47–49 In addition, minocy-cline may also protect CNS cells from apoptosis throughinhibition of caspase-3 activity.31,41,42 Minocycline has alsobeen shown to down-regulate signal transduction pathwayssuch as nuclear factor-�B and p38 mitogen-activated pro-tein kinase,32,48,50,51 both of which play pivotal roles incontrolling the expression of numerous proinflammatorygenes. Tetracyclines have also been shown to inhibit pro-duction of the proinflammatory molecules TNF-�,52,53 NO,54
and matrix metalloproteinases44,55 by leukocytes. However,it is also important to acknowledge the fact that other inves-tigators have reported that minocycline has no impact or, insome cases, exacerbates neuronal injury in several CNSdisease models,23,56–60 highlighting the need for furtherstudies in this area to identify specific conditions in whichminocycline therapy may exert beneficial effects.
Although a few recent reports have documented favor-able effects of minocycline in various viral and parasiticinfections,61–64 the consequences of this antimicrobial onmodulating inflammatory responses in the context of CNSbacterial infections, such as brain abscess, have not yetbeen examined. The relative bacteriostatic versus im-mune modulatory effects of minocycline have beendifficult to differentiate; however, in this study we havedevised a strategy to discern between these two pos-sibilities by engineering a S. aureus strain that is resistantto the bacteriostatic effects of minocycline. Our findingsindicate that an overactive host response to infection canlead to the heightened expression of proinflammatorymediators and subsequent mortality during the initialstages of brain abscess, which can be attenuated byminocycline independently of its antibacterial activity. Inaddition, minocycline was capable of reducing ongoinginflammation, as both inflammatory mediator expressionand brain abscess size were attenuated when minocy-cline treatment was initiated 3 days following S. aureus
1200 Kielian et alAJP October 2007, Vol. 171, No. 4

infection. Collectively, these results suggest that if nottightly regulated, the inflammatory response that ensuesduring brain abscess development can exert deleteriouseffects. Therefore, minocycline may represent an excel-lent candidate for brain abscess therapy through its abil-ity to attenuate chronic inflammation and reduce the ex-tent of parenchymal tissue damage that is characteristicof infection.
Materials and Methods
Bacterial Strains and Reagents
The minocycline-sensitive S. aureus strain RN6390 wasused as previously described.7–10 A minocycline-resis-tant isogenic strain of RN6390 was constructed with atetracycline resistance cassette (tetM), which also con-fers resistance to minocycline, stably inserted into the bac-terial chromosome using the integration vector pCL84,which preferentially inserts into the S. aureus lipase gene.65
Chromosomal insertion of the tetM resistance cassette wasconfirmed by sequence analysis of chromosomal polymer-ase chain reaction (PCR) products and Southern blots. Todetermine whether the insertion of the tetM cassette led toany overt alterations in the S. aureustetM mutant comparedwith the parental strain RN6390, pulse field gel electro-phoresis and sodium dodecyl sulfate-polyacrylamidegel electrophoresis of whole cell lysates and secreted pro-teins from several independent bacterial colonies wereperformed.
Minocycline-HCl was purchased from Sigma ChemicalCo. (St. Louis, MO), and a 10 mg/ml stock solution wasprepared in endotoxin-free H2O and stored in aliquots at�20°C until use. For animal injections, minocycline work-ing stocks were prepared fresh daily in sterile endotoxin-free phosphate-buffered saline (pH 7.4).
Generation of Experimental Brain Abscesses
Brain abscesses were induced in 6- to 8-week-oldC57BL/6 mice using a stereotaxic approach as previ-ously described.8,9 For all studies, an equal number ofage-matched males and females were used as previouswork has established that both genders exhibit qualita-tively similar inflammatory profiles following bacterialchallenge.7–10 The animal use protocol has been ap-proved by the University of Arkansas for Medical Sci-ences Institutional Animal Care and Use Committee andis in accord with the National Institutes of Health guide-lines for the use of rodents.
For evaluating the potential therapeutic effects of mino-cycline during brain abscess development, mice were sub-jected to either one of two treatment paradigms. In the first,the effects of minocycline when administered before S.aureus exposure were examined. For these experiments,mice received once daily intraperitoneal (i.p.) injections ofminocycline beginning 1 day before S. aureus infection andcontinuing until study termination. In the second treatmentparadigm, minocycline was evaluated for its ability to mod-ulate ongoing CNS inflammation. For these studies, mino-
cycline was administered 3 days following S. aureus infec-tion, a time point when bacterial burdens have nearlypeaked.7,11 After this initial treatment, minocycline injectionscontinued daily until the termination of the study. Minocy-cline was initially examined at three concentrations (2, 10,and 50 mg/kg/day); however, in subsequent studies onedose was selected (50 mg/kg/day), which was determinedto exert optimal effects on the ensuing host CNS antibacte-rial response. For all experiments, one group of animalsreceived minocycline treatment alone (50 mg/kg/day) toassess any effects of the drug independently of S. aureusinfection. Animals receiving 50 mg/kg/day minocyclinealone did not demonstrate any overt adverse effects to thedrug (ie, no significant alterations in body weight). Thedoses of minocycline tested are representative of concen-trations used in previously published reports of disparateCNS neurodegenerative models,46,66 and although theyrepresent concentrations well above those used in humans,this is offset by the extremely short half-life of minocycline inmice (2 to 3 hours) versus humans (15 to 18 hours).22,66,67
Simultaneous Collection of RNA, Protein, andQuantitation of Viable Bacteria from BrainAbscesses
Brain abscesses were visualized by the stab wound cre-ated during injections, sectioned within 2 to 3 mm on allsides, and homogenized in PBS supplemented with pro-tease and RNase inhibitors. Subsequently, RNA, protein,and bacterial titers were obtained as previously de-scribed.9 It is not feasible to dissect away necrotic fromviable brain tissue while collecting samples for analysis,because these microscopic borders are not easily dis-cernable by the naked eye. Although we do not specifybetween viable and necrotic tissue when collecting brainabscesses, we propose that this does not introduce arti-facts in the results obtained. For example, our resultsdemonstrate that minocycline treatment reduced brainabscess severity as typified by smaller lesions, whichtranslates into a greater amount of surviving tissue. Yet inthis case, the expression of several proinflammatory me-diators was diminished. If necrotic tissue greatly impactsthe results obtained, we would have expected a reduc-tion in cytokine production in brain abscesses of vehicle-treated animals, which was not observed.
Quantitation of Brain Abscess Size
At the indicated time points postinfection, minocycline-and vehicle-treated mice were perfused transcardiallywith PBS to eliminate leukocytes from the vasculature,whereupon brains were removed and immediately flash-frozen on dry ice. Before cryostat sectioning, brain tis-sues were embedded in optimal cutting temperature me-dium, and serial 10-�m sections were made throughoutthe entire lesioned tissue and stained with hematoxylinand eosin to demarcate abscess margins. Evaluation ofserial sections throughout the affected brain parenchymaensured that the largest abscess cross-sectional area
Minocycline Modulates Brain Abscess 1201AJP October 2007, Vol. 171, No. 4

would be identified for comparisons of lesion size. Ab-scess area (reported as square millimeters) was calcu-lated using the MetaMorph image analysis program (Uni-versal Imaging Corporation, Downingtown, PA).
Evaluation of Tissue Viability Using Nitro BlueTetrazolium (NBT) Staining
To better demarcate the border between living and non-viable tissue during the early stages of brain abscessdevelopment, NBT staining of fresh frozen tissue sectionswas performed.68 The intensity of staining is dictated bythe amount of mitochondrial diaphorase enzymes thatwere present in viable cells at the time of tissue collec-tion, which metabolize the conversion of NBT into a blueformazan product. The loss of staining is indicative of thedegree of cell damage/injury. In brief, 10-�m cryostatsections were fixed for 10 minutes in ice-cold acetoneand air-dried. Tissues were subsequently incubated for40 minutes at room temperature with 200 �l/slide NBTdevelopment solution (0.2 mol/L Tris-HCl, pH 7.1, con-taining 0.25 mg/ml NBT, 2 mg/ml NADH, both from SigmaChemical Co., and 1 mg/ml MgCl2). To terminate thereaction, slides were rinsed in double distilled water,dehydrated, and coverslipped using Permount mountingmedium.
Immunofluorescence Staining of Astrocytes andMicroglia/Macrophages
The effects of minocycline on astrocyte and microglia/macrophage activation during brain abscess develop-ment were evaluated by immunofluorescence stainingusing glial fibrillary acidic protein (GFAP) and F4/80,respectively, and confocal microscopy. Brain abscesstissues were collected at the indicated time points follow-ing minocycline treatment, and 10-�m cryostat sectionswere mounted onto SuperFrost Plus slides (Fisher Scien-tific, Houston, TX), air-dried, and stored at �80°C untiluse. To initiate staining, slides were equilibrated at roomtemperature for 15 minutes and fixed in ice-cold metha-nol. Following numerous rinses in PBS, tissues were in-cubated with PBS/10% normal donkey serum to minimizenonspecific staining. Brain abscess tissues were incu-bated with either primary anti-rat GFAP (Zymed, SouthSan Francisco, CA) or F4/80 (Serotec, Raleigh, NC) an-tibodies overnight at 4°C in a humidified chamber. Fol-lowing numerous rinses in PBS, tissues were incubatedwith either a donkey anti-rat IgG fluorescein isothiocya-nate conjugate (for GFAP) or biotinylated donkey anti-ratIgG antibody (for F4/80; both from Jackson Immunore-search, West Grove, PA) for 1 hour at room temperature.F4/80 expression was visualized by the addition of astreptavidin-Alexa Fluor 568 conjugate (MolecularProbes, Eugene, OR). On completion of the staining pro-tocol, slides were coverslipped using the Prolong anti-fade reagent (Molecular Probes) and sealed using nailpolish. Slides were imaged using a Zeiss laser scanningconfocal microscope (LSM 510; Carl Zeiss Microimaging
Inc., Thornwood, NY), and fluorescein isothiocyanate wasexcited with a 488-nm argon laser to visualize GFAPimmunoreactivity, and Alexa Fluor 568 was excited with a561-nm diode-pumped solid-state laser to demonstrateF4/80 expression, with images collected using the appro-priate emissions. The confocal pinhole was set to obtainan optical section thickness of 1.6 �m. Specific stainingof antibodies was confirmed by the absence of fluores-cence signal following incubation of brain abscess tis-sues with secondary antibodies alone (data not shown).
Preparation of Primary Microglia and AstrocyteCultures
Primary microglia and astrocytes were isolated from neo-natal C57BL/6 mice (postnatal days 2 to 4) as previouslydescribed.69 The purity of microglia and astrocyte cul-tures, as determined by immunohistochemical stainingusing antibodies against CD11b (BD PharMingen, SanJose, CA) and GFAP (Dako Corp., Carpinteria, CA), wasroutinely greater than 95 and 90%, respectively.
To investigate the effects of minocycline on glialactivation, primary microglia and astrocytes wereseeded into 96-well plates at 2 � 105 or 1 � 105
cells/well, respectively, and incubated overnight. Thefollowing day, cells were treated with various doses ofminocycline for 1 hour before stimulation with eitherheat-inactivated S. aureus [107 colony-forming units(cfu), strain RN6390] or 10 �g/ml peptidoglycan (PGN;InvivoGen, San Diego, CA). Cell-conditioned superna-tants were collected at 24 hours following bacterialexposure, whereupon NO and cytokine/chemokineproduction was evaluated as described below. Thedoses of bacterial stimuli used were based on ourprevious studies that established optimal cytokine re-sponses without any evidence of toxicity.70,71
Nitrite Assay
Nitrite (a stable reaction product of NO) levels were de-termined by adding 50 �l of astrocyte-conditioned cul-ture medium with 50 �l of Griess reagent (0.1% naphth-ylethylenediamine dihydrochloride, 1% sulfanilamide,and 2.5% phosphoric acid; all from Sigma Chemical Co.)in a 96-well plate. The absorbance at 550 nm was mea-sured on a plate reader (Spectra Max 190; MolecularDevices, Sunnyvale, CA), and nitrite concentrations werecalculated using a standard curve with sodium nitrite(NaNO2; Sigma Chemical Co.; level of sensitivity 0.4�mol/L).
Cell Viability Assays
The effects of minocycline on microglia and astrocyteviability were evaluated using a standard 3-(4,5-dimeth-ylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT) (Sigma) assaybased on the mitochondrial conversion of MTT intoformazan crystals as previously described.72
1202 Kielian et alAJP October 2007, Vol. 171, No. 4
Protein Extraction and Western Blotting
Protein extracts were prepared from primary microglia orastrocytes as previously described.72,73 The effects ofminocycline on glial Toll-like receptor 2 (TLR2) proteinexpression were evaluated by Western blot analysis.Blots were probed using a goat anti-mouse TLR2 anti-body (R&D Systems, Minneapolis, MN) followed by adonkey anti-goat IgG–horseradish peroxidase second-ary antibody (Jackson Immunoresearch). Blots were de-veloped using the Immobilon Western substrate (Milli-pore, Billerica, MA) and visualized by exposure to X-rayfilm. Subsequently, membranes were stripped and re-probed with a rabbit anti-actin polyclonal antibody(Sigma Chemical Co.) for normalization.
Quantitative Real-Time RT-PCR (qRT-PCR)
Total RNA from brain abscesses was isolated using theTRIzol reagent and treated with DNase1 (both from In-vitrogen) before use in qRT-PCR studies. The experimen-tal procedure was performed as previously described.73
IL-1�, TNF-�, and glyceraldehyde-3-phosphate dehydro-genase primers and TAMRA TaqMan probes were de-signed69,74 and synthesized by Applied Biosystems (ABI,Foster City, CA). An ABI Assays-on-Demand TaqMan kitwas used to examine CCL2 expression.
Enzyme-Linked Immunosorbent Assay (ELISA)
Murine TNF-�, IL-1�, and CCL2 (OptiEIA; BD PharMin-gen) and CXCL2 (MIP-2, DuoSet; R&D Systems) werequantified in brain abscess homogenates using commer-cial ELISA kits. Results were normalized to the amount oftotal protein extracted from tissues to correct for differ-ences in sampling size as previously described.8,11
Statistics
Significant differences between experimental groupswere determined using the unpaired Student’s t-test atthe 95% confidence interval with Sigma Stat (SPSS Sci-ence, Chicago, IL). This analysis was determined to bemost appropriate since, although we are evaluatingchanges in proinflammatory mediator expression overtime, repeated measurements are not made on the sameanimal (mice are sacrificed to collect abscess homoge-nates at each time point), precluding analysis of varianceand post hoc analysis of the data.
In this study, we performed a minimum of three inde-pendent replicates of each study to confirm the resultsobtained. The reporting of our results as representative of“x” number of independent experiments was required,because it is difficult to achieve identical bacterial bur-dens in mice between independent brain abscess stud-ies with distinct bacterial preparations. As a result, theabsolute concentrations of the various proinflammatorymediators detected within brain abscesses of minocy-cline-treated and control mice differed between individ-ual experiments; however, the trends were consistent.
This required us to report results from a single experimentwhere at least four or five mice per time point per groupwere analyzed and statistical analysis conducted.
Results
Minocycline Inhibits S. aureus Replication inBrain Abscesses Concomitant with AttenuatedProinflammatory Mediator Expression
Initial experiments established that the S. aureus strainused in our experimental model, RN6390, was sensitiveto the antibiotic effects of minocycline (data not shown).To examine whether minocycline affected the acutephase of brain abscess development in this susceptiblestrain, the antibiotic was administered daily to animalsbeginning one day before intracerebral S. aureus infec-tion and continuing until termination of the study. Mino-cycline treatment led to a dose-dependent reduction inbacterial burdens (Figure 1) that coincided with signifi-cant decreases in the expression of several proinflamma-tory mediators that we have previously determined to bepivotal during the acute stage of brain abscess includingIL-1�, TNF-�, and CCL2 (Figure 2).8 The effects of mino-cycline on proinflammatory mediator expression wereprobably related to its ability to reduce bacterial burdensand/or modulate signaling pathways important for theinduction of cytokines/chemokines including nuclear fac-tor-�B or p38 mitogen-activated protein kinase.47,51
Minocycline Exhibits Anti-Inflammatory EffectsIndependently of Its Bacteriostatic Activity
Because we were interested in evaluating the potentialanti-inflammatory effects of minocycline in the brain ab-
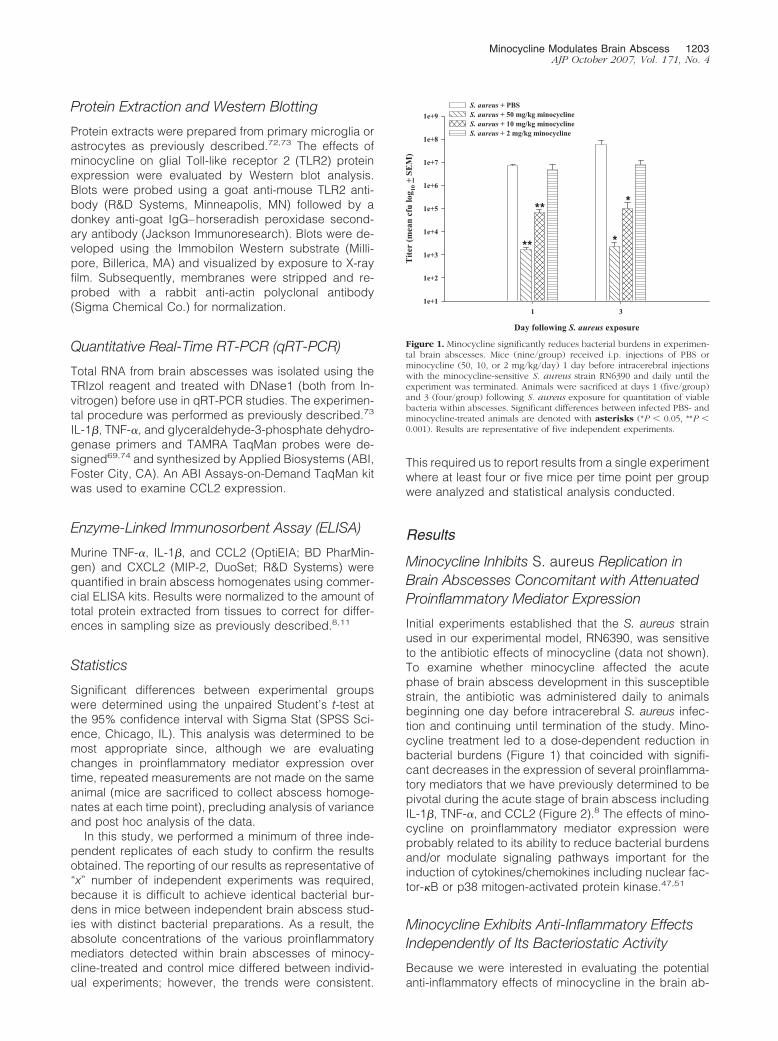
Figure 1. Minocycline significantly reduces bacterial burdens in experimen-tal brain abscesses. Mice (nine/group) received i.p. injections of PBS orminocycline (50, 10, or 2 mg/kg/day) 1 day before intracerebral injectionswith the minocycline-sensitive S. aureus strain RN6390 and daily until theexperiment was terminated. Animals were sacrificed at days 1 (five/group)and 3 (four/group) following S. aureus exposure for quantitation of viablebacteria within abscesses. Significant differences between infected PBS- andminocycline-treated animals are denoted with asterisks (*P � 0.05, **P �0.001). Results are representative of five independent experiments.
Minocycline Modulates Brain Abscess 1203AJP October 2007, Vol. 171, No. 4

scess model independently of its antibiotic properties, wegenerated an isogenic minocycline-resistant strain of S.aureus RN6390 (referred to hereafter as S. aureustetM). Itwas important to create a minocycline mutant isogenic tothe parental strain RN6390 rather than use an unrelatedS. aureus isolate that is naturally resistant to minocycline,because we have extensively characterized the host im-mune response to S. aureus RN6390 in the brain abscessmodel.7–10,16 To determine whether the insertion of theminocycline resistance cassette tetM resulted in any dra-matic changes in phenotype compared with the parentalstrain RN6390, pulse field gel electrophoresis and so-dium dodecyl sulfate-polyacrylamide gel electrophoresisof whole cell lysates and secreted proteins from severalindependent clones of both the S. aureustetM mutant andparental strains were performed. Pulse field gel electro-phoresis revealed the loss of a larger SmaI fragment inthe S. aureustetM mutant with the concomitant appear-ance of two smaller fragments (Figure 3A). This finding
can be explained by the fact that the pCL84 vector usedto insert the tetM gene into the bacterial chromosomecontains SmaI sites. Evaluation of whole cell lysates andsecreted proteins from both the parental and S. aureustetM
strains in vitro revealed that protein expression was nearlyidentical between the two strains, except for the pres-
Figure 2. Minocycline attenuates proinflammatory cytokine and chemokineexpression in experimental brain abscesses. Mice (nine/group) received i.p.injections of PBS or minocycline (50, 10, or 2 mg/kg/day) 1 day beforeintracerebral injections with the minocycline-sensitive S. aureus strainRN6390 and daily until the experiment was terminated. Abscess homoge-nates were prepared at days 1 (five/group) and 3 (four/group) following S.aureus exposure for quantitation of IL-1� (A), TNF-� (B), and CCL2 (C)expression by ELISA. Cytokine/chemokine concentrations were normalizedto the amount of total protein recovered to correct for differences in tissuesampling size. Significant differences between infected PBS- and minocy-cline-treated animals are denoted with asterisks (*P � 0.05, **P � 0.001).Results are representative of five independent experiments.
Figure 3. Characterization of the S. aureus tetM strain. A: Pulse field gelelectrophoresis (PFGE) of the parental S. aureus strain RN6390 and itsisogenic tetM mutants. Chromosomal DNAs from three individual parentaland mutant isolates were digested with SmaI and subjected to PFGE. Becausethe vector pCL84 that was used to insert the tetM gene into the lipase gene ofthe bacterial chromosome contains SmaI sites, this resulted in a loss of alarger fragment (solid arrow) and the generation of two smaller fragments(blocked arrows). B: Coomassie blue-stained gels depicting the array ofproteins in bacterial cell lysates and conditioned supernatants. Overall, theresults demonstrate that the in vitro protein expression profiles are similarbetween the parental RN6390 and S. aureus tetM strains except for the pres-ence of a faint protein band in the latter (arrow). The data presented werederived from separate colonies of parental or mutant strains that werepropagated in culture independently.
1204 Kielian et alAJP October 2007, Vol. 171, No. 4
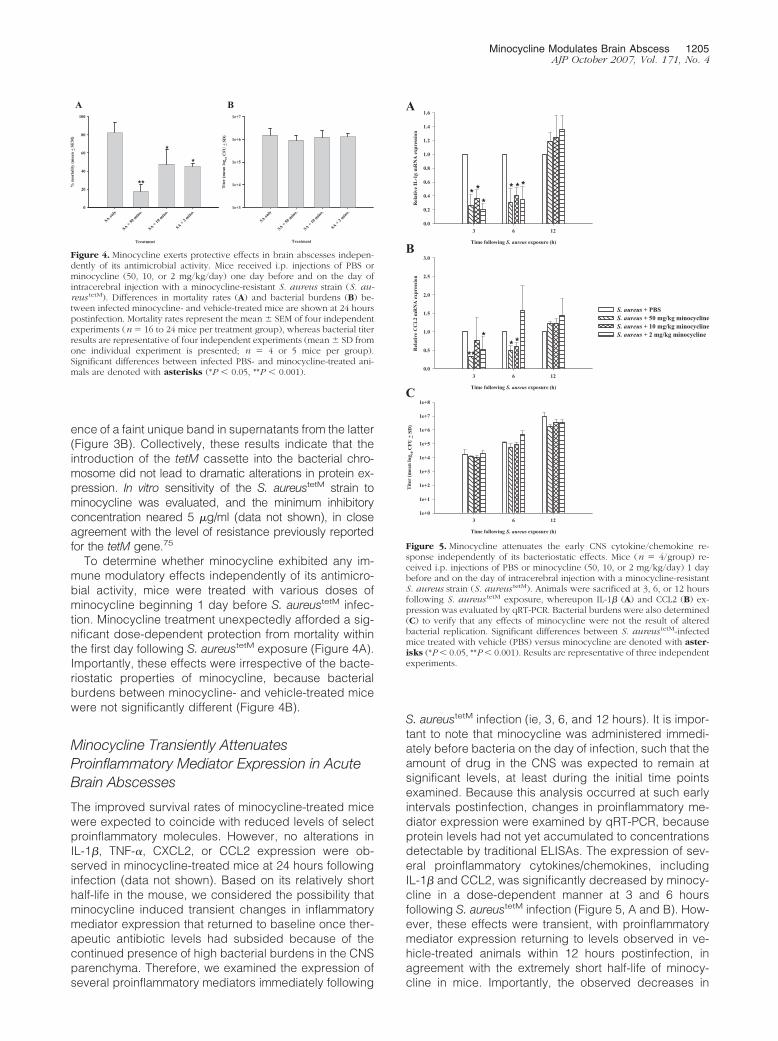
ence of a faint unique band in supernatants from the latter(Figure 3B). Collectively, these results indicate that theintroduction of the tetM cassette into the bacterial chro-mosome did not lead to dramatic alterations in protein ex-pression. In vitro sensitivity of the S. aureustetM strain tominocycline was evaluated, and the minimum inhibitoryconcentration neared 5 �g/ml (data not shown), in closeagreement with the level of resistance previously reportedfor the tetM gene.75
To determine whether minocycline exhibited any im-mune modulatory effects independently of its antimicro-bial activity, mice were treated with various doses ofminocycline beginning 1 day before S. aureustetM infec-tion. Minocycline treatment unexpectedly afforded a sig-nificant dose-dependent protection from mortality withinthe first day following S. aureustetM exposure (Figure 4A).Importantly, these effects were irrespective of the bacte-riostatic properties of minocycline, because bacterialburdens between minocycline- and vehicle-treated micewere not significantly different (Figure 4B).
Minocycline Transiently AttenuatesProinflammatory Mediator Expression in AcuteBrain Abscesses
The improved survival rates of minocycline-treated micewere expected to coincide with reduced levels of selectproinflammatory molecules. However, no alterations inIL-1�, TNF-�, CXCL2, or CCL2 expression were ob-served in minocycline-treated mice at 24 hours followinginfection (data not shown). Based on its relatively shorthalf-life in the mouse, we considered the possibility thatminocycline induced transient changes in inflammatorymediator expression that returned to baseline once ther-apeutic antibiotic levels had subsided because of thecontinued presence of high bacterial burdens in the CNSparenchyma. Therefore, we examined the expression ofseveral proinflammatory mediators immediately following
S. aureustetM infection (ie, 3, 6, and 12 hours). It is impor-tant to note that minocycline was administered immedi-ately before bacteria on the day of infection, such that theamount of drug in the CNS was expected to remain atsignificant levels, at least during the initial time pointsexamined. Because this analysis occurred at such earlyintervals postinfection, changes in proinflammatory me-diator expression were examined by qRT-PCR, becauseprotein levels had not yet accumulated to concentrationsdetectable by traditional ELISAs. The expression of sev-eral proinflammatory cytokines/chemokines, includingIL-1� and CCL2, was significantly decreased by minocy-cline in a dose-dependent manner at 3 and 6 hoursfollowing S. aureustetM infection (Figure 5, A and B). How-ever, these effects were transient, with proinflammatorymediator expression returning to levels observed in ve-hicle-treated animals within 12 hours postinfection, inagreement with the extremely short half-life of minocy-cline in mice. Importantly, the observed decreases in
Figure 4. Minocycline exerts protective effects in brain abscesses indepen-dently of its antimicrobial activity. Mice received i.p. injections of PBS orminocycline (50, 10, or 2 mg/kg/day) one day before and on the day ofintracerebral injection with a minocycline-resistant S. aureus strain (S. au-reus tetM). Differences in mortality rates (A) and bacterial burdens (B) be-tween infected minocycline- and vehicle-treated mice are shown at 24 hourspostinfection. Mortality rates represent the mean � SEM of four independentexperiments (n � 16 to 24 mice per treatment group), whereas bacterial titerresults are representative of four independent experiments (mean � SD fromone individual experiment is presented; n � 4 or 5 mice per group).Significant differences between infected PBS- and minocycline-treated ani-mals are denoted with asterisks (*P � 0.05, **P � 0.001).
Figure 5. Minocycline attenuates the early CNS cytokine/chemokine re-sponse independently of its bacteriostatic effects. Mice (n � 4/group) re-ceived i.p. injections of PBS or minocycline (50, 10, or 2 mg/kg/day) 1 daybefore and on the day of intracerebral injection with a minocycline-resistantS. aureus strain (S. aureus tetM). Animals were sacrificed at 3, 6, or 12 hoursfollowing S. aureus tetM exposure, whereupon IL-1� (A) and CCL2 (B) ex-pression was evaluated by qRT-PCR. Bacterial burdens were also determined(C) to verify that any effects of minocycline were not the result of alteredbacterial replication. Significant differences between S. aureus tetM-infectedmice treated with vehicle (PBS) versus minocycline are denoted with aster-isks (*P � 0.05, **P � 0.001). Results are representative of three independentexperiments.
Minocycline Modulates Brain Abscess 1205AJP October 2007, Vol. 171, No. 4
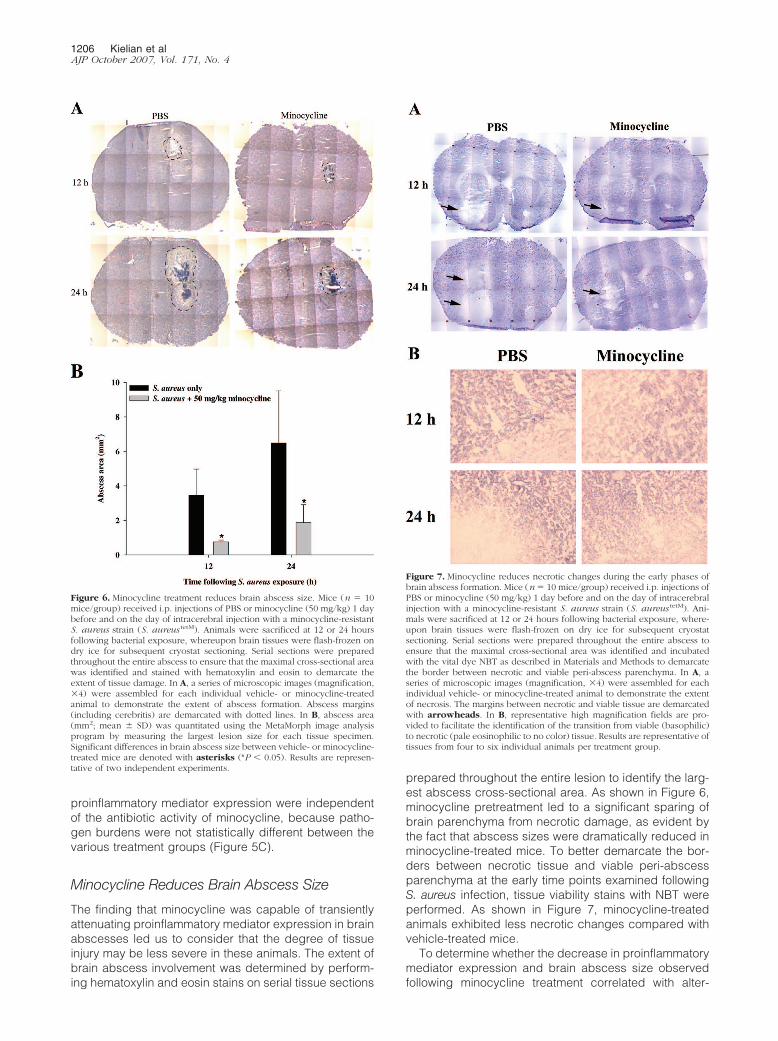
proinflammatory mediator expression were independentof the antibiotic activity of minocycline, because patho-gen burdens were not statistically different between thevarious treatment groups (Figure 5C).
Minocycline Reduces Brain Abscess Size
The finding that minocycline was capable of transientlyattenuating proinflammatory mediator expression in brainabscesses led us to consider that the degree of tissueinjury may be less severe in these animals. The extent ofbrain abscess involvement was determined by perform-ing hematoxylin and eosin stains on serial tissue sections
prepared throughout the entire lesion to identify the larg-est abscess cross-sectional area. As shown in Figure 6,minocycline pretreatment led to a significant sparing ofbrain parenchyma from necrotic damage, as evident bythe fact that abscess sizes were dramatically reduced inminocycline-treated mice. To better demarcate the bor-ders between necrotic tissue and viable peri-abscessparenchyma at the early time points examined followingS. aureus infection, tissue viability stains with NBT wereperformed. As shown in Figure 7, minocycline-treatedanimals exhibited less necrotic changes compared withvehicle-treated mice.
To determine whether the decrease in proinflammatorymediator expression and brain abscess size observedfollowing minocycline treatment correlated with alter-
Figure 6. Minocycline treatment reduces brain abscess size. Mice (n � 10mice/group) received i.p. injections of PBS or minocycline (50 mg/kg) 1 daybefore and on the day of intracerebral injection with a minocycline-resistantS. aureus strain (S. aureus tetM). Animals were sacrificed at 12 or 24 hoursfollowing bacterial exposure, whereupon brain tissues were flash-frozen ondry ice for subsequent cryostat sectioning. Serial sections were preparedthroughout the entire abscess to ensure that the maximal cross-sectional areawas identified and stained with hematoxylin and eosin to demarcate theextent of tissue damage. In A, a series of microscopic images (magnification,�4) were assembled for each individual vehicle- or minocycline-treatedanimal to demonstrate the extent of abscess formation. Abscess margins(including cerebritis) are demarcated with dotted lines. In B, abscess area(mm2; mean � SD) was quantitated using the MetaMorph image analysisprogram by measuring the largest lesion size for each tissue specimen.Significant differences in brain abscess size between vehicle- or minocycline-treated mice are denoted with asterisks (*P � 0.05). Results are represen-tative of two independent experiments.
Figure 7. Minocycline reduces necrotic changes during the early phases ofbrain abscess formation. Mice (n � 10 mice/group) received i.p. injections ofPBS or minocycline (50 mg/kg) 1 day before and on the day of intracerebralinjection with a minocycline-resistant S. aureus strain (S. aureus tetM). Ani-mals were sacrificed at 12 or 24 hours following bacterial exposure, where-upon brain tissues were flash-frozen on dry ice for subsequent cryostatsectioning. Serial sections were prepared throughout the entire abscess toensure that the maximal cross-sectional area was identified and incubatedwith the vital dye NBT as described in Materials and Methods to demarcatethe border between necrotic and viable peri-abscess parenchyma. In A, aseries of microscopic images (magnification, �4) were assembled for eachindividual vehicle- or minocycline-treated animal to demonstrate the extentof necrosis. The margins between necrotic and viable tissue are demarcatedwith arrowheads. In B, representative high magnification fields are pro-vided to facilitate the identification of the transition from viable (basophilic)to necrotic (pale eosinophilic to no color) tissue. Results are representative oftissues from four to six individual animals per treatment group.
1206 Kielian et alAJP October 2007, Vol. 171, No. 4
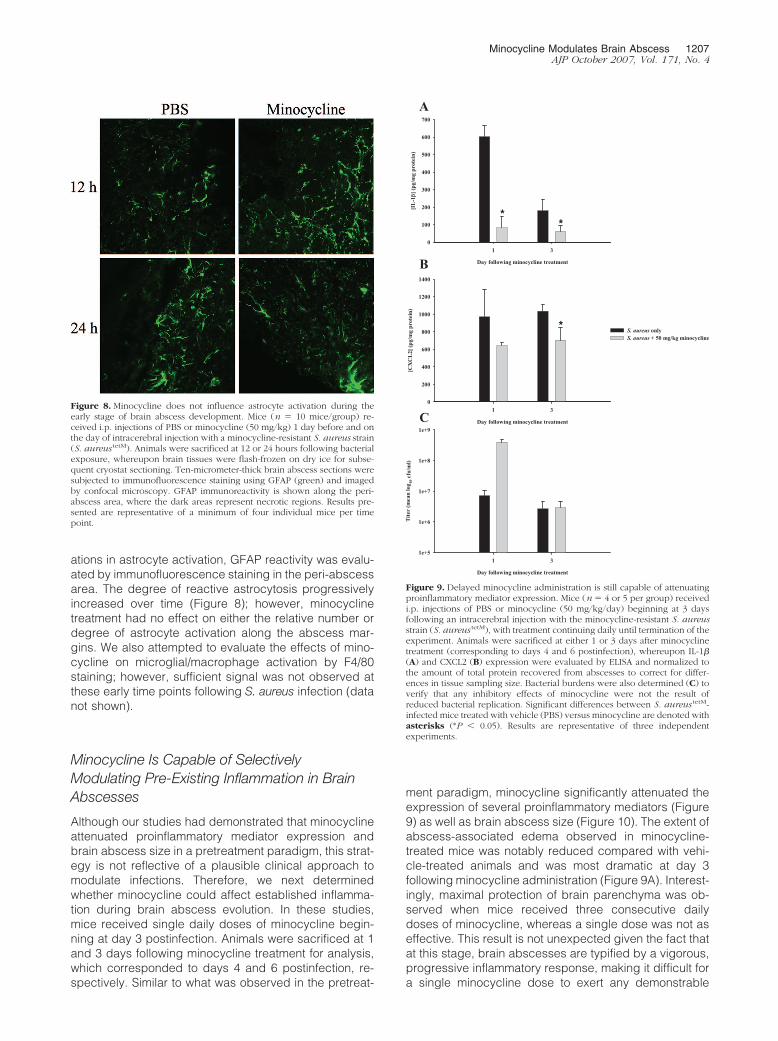
ations in astrocyte activation, GFAP reactivity was evalu-ated by immunofluorescence staining in the peri-abscessarea. The degree of reactive astrocytosis progressivelyincreased over time (Figure 8); however, minocyclinetreatment had no effect on either the relative number ordegree of astrocyte activation along the abscess mar-gins. We also attempted to evaluate the effects of mino-cycline on microglial/macrophage activation by F4/80staining; however, sufficient signal was not observed atthese early time points following S. aureus infection (datanot shown).
Minocycline Is Capable of SelectivelyModulating Pre-Existing Inflammation in BrainAbscesses
Although our studies had demonstrated that minocyclineattenuated proinflammatory mediator expression andbrain abscess size in a pretreatment paradigm, this strat-egy is not reflective of a plausible clinical approach tomodulate infections. Therefore, we next determinedwhether minocycline could affect established inflamma-tion during brain abscess evolution. In these studies,mice received single daily doses of minocycline begin-ning at day 3 postinfection. Animals were sacrificed at 1and 3 days following minocycline treatment for analysis,which corresponded to days 4 and 6 postinfection, re-spectively. Similar to what was observed in the pretreat-
ment paradigm, minocycline significantly attenuated theexpression of several proinflammatory mediators (Figure9) as well as brain abscess size (Figure 10). The extent ofabscess-associated edema observed in minocycline-treated mice was notably reduced compared with vehi-cle-treated animals and was most dramatic at day 3following minocycline administration (Figure 9A). Interest-ingly, maximal protection of brain parenchyma was ob-served when mice received three consecutive dailydoses of minocycline, whereas a single dose was not aseffective. This result is not unexpected given the fact thatat this stage, brain abscesses are typified by a vigorous,progressive inflammatory response, making it difficult fora single minocycline dose to exert any demonstrable
Figure 9. Delayed minocycline administration is still capable of attenuatingproinflammatory mediator expression. Mice (n � 4 or 5 per group) receivedi.p. injections of PBS or minocycline (50 mg/kg/day) beginning at 3 daysfollowing an intracerebral injection with the minocycline-resistant S. aureusstrain (S. aureus tetM), with treatment continuing daily until termination of theexperiment. Animals were sacrificed at either 1 or 3 days after minocyclinetreatment (corresponding to days 4 and 6 postinfection), whereupon IL-1�(A) and CXCL2 (B) expression were evaluated by ELISA and normalized tothe amount of total protein recovered from abscesses to correct for differ-ences in tissue sampling size. Bacterial burdens were also determined (C) toverify that any inhibitory effects of minocycline were not the result ofreduced bacterial replication. Significant differences between S. aureus tetM-infected mice treated with vehicle (PBS) versus minocycline are denoted withasterisks (*P � 0.05). Results are representative of three independentexperiments.
Figure 8. Minocycline does not influence astrocyte activation during theearly stage of brain abscess development. Mice (n � 10 mice/group) re-ceived i.p. injections of PBS or minocycline (50 mg/kg) 1 day before and onthe day of intracerebral injection with a minocycline-resistant S. aureus strain(S. aureus tetM). Animals were sacrificed at 12 or 24 hours following bacterialexposure, whereupon brain tissues were flash-frozen on dry ice for subse-quent cryostat sectioning. Ten-micrometer-thick brain abscess sections weresubjected to immunofluorescence staining using GFAP (green) and imagedby confocal microscopy. GFAP immunoreactivity is shown along the peri-abscess area, where the dark areas represent necrotic regions. Results pre-sented are representative of a minimum of four individual mice per timepoint.
Minocycline Modulates Brain Abscess 1207AJP October 2007, Vol. 171, No. 4

effects. Examination of astrocyte and microglial/macro-phage activation by immunofluorescence staining withGFAP and F4/80, respectively, did not reveal any signif-icant differences following minocycline treatment, al-though the relative degree of immunoreactivity of bothcell populations in the peri-abscess area progressivelyincreased over time (Figure 11), as we have previouslydemonstrated.11 Collectively, these results indicate thatminocycline could represent a potential therapeuticagent even in conditions of pre-existing inflammation toattenuate bystander destruction of surrounding normalbrain parenchyma.
Minocycline Selectively AttenuatesProinflammatory Mediator Production inS. aureus- and PGN-Activated Glia
Microglia are the resident innate immune cells in the CNSparenchyma and are uniquely poised to immediately re-spond to bacterial infections by producing numerouscytokines and chemokines.76–78 The neuroprotective ef-fects of minocycline have been attributed, in part, to its
Figure 10. Minocycline is capable of significantly reducing brain abscess sizein the face of established infection. Mice (n � 6/group) received i.p. injec-tions of PBS or minocycline (50 mg/kg/day) beginning at 3 days following anintracerebral injection with the minocycline-resistant S. aureus strain (S.aureus tetM), with treatment continuing daily until termination of the exper-iment. Animals were sacrificed at either 1 or 3 days after minocyclinetreatment (corresponding to days 4 and 6 postinfection), whereupon braintissues were flash-frozen on dry ice for subsequent cryostat sectioning. Serialsections were prepared throughout the entire abscess to ensure that themaximal cross-sectional area was identified and stained with hematoxylinand eosin to demarcate the extent of tissue damage. In A, a series ofmicroscopic images (magnification, �4) were assembled for each individualvehicle- or minocycline-treated animal to demonstrate the extent of abscessformation and edema/necrosis. Abscess margins (including cerebritis) aredemarcated with dotted lines. In B, abscess area (mm2; mean � SD) wasquantitated using the MetaMorph image analysis program by measuring thelargest lesion size for each tissue specimen. Significant differences in brainabscess size between vehicle- or minocycline-treated mice are denoted withasterisks (*P � 0.05, **P � 0.001). Results are representative of two inde-pendent experiments.
Figure 11. Minocycline does not influence astrocyte or microglial/macro-phage activation when administered subsequent to infection. Mice (n �6/group) received i.p. injections of PBS or minocycline (50 mg/kg/day)beginning at 3 days following an intracerebral injection with the minocycline-resistant S. aureus strain (S. aureus tetM), with treatment continuing daily untiltermination of the experiment. Animals were sacrificed at either 1 or 3 daysafter minocycline treatment (corresponding to days 4 and 6 postinfection),whereupon brain tissues were flash-frozen on dry ice for subsequent cryostatsectioning. Ten-micrometer-thick brain abscess sections were subjected toimmunofluorescence staining using GFAP (A; green) or F4/80 (B; red) toidentify astrocytes and microglia/macrophages, respectively, and imaged byconfocal microscopy. GFAP and F4/80 immunoreactivity is shown along theperi-abscess area, where the dark areas represent necrotic regions. Resultspresented are representative of a minimum of four independent mice pertime point.
1208 Kielian et alAJP October 2007, Vol. 171, No. 4
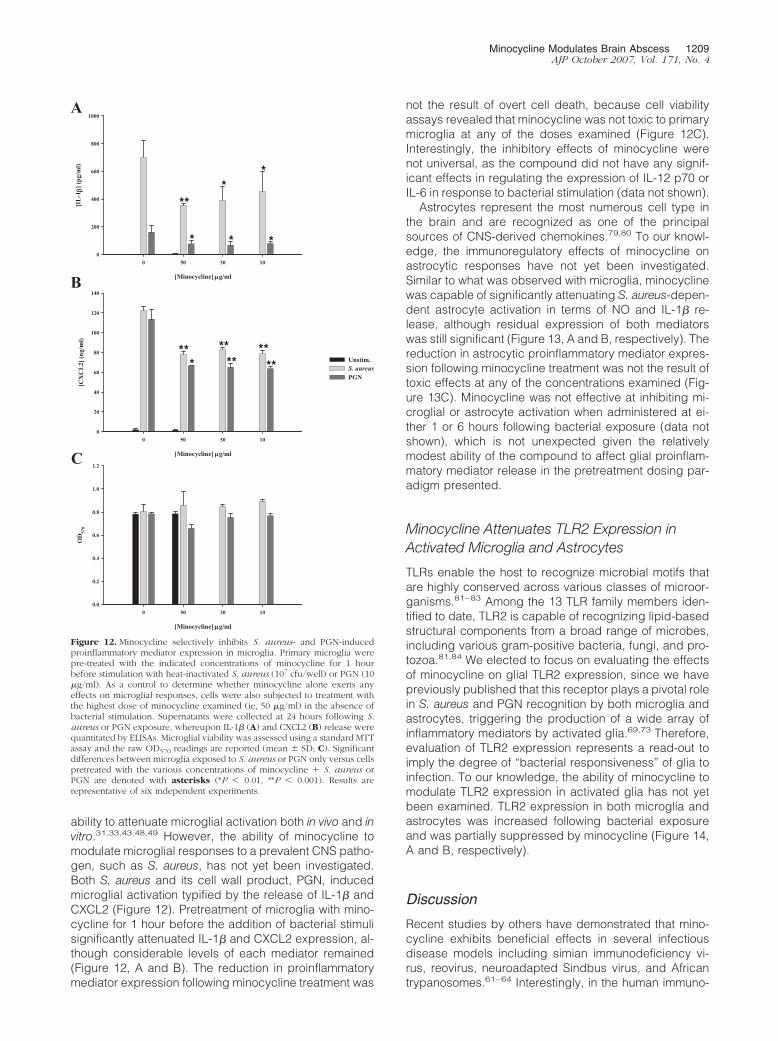
ability to attenuate microglial activation both in vivo and invitro.31,33,43,48,49 However, the ability of minocycline tomodulate microglial responses to a prevalent CNS patho-gen, such as S. aureus, has not yet been investigated.Both S. aureus and its cell wall product, PGN, inducedmicroglial activation typified by the release of IL-1� andCXCL2 (Figure 12). Pretreatment of microglia with mino-cycline for 1 hour before the addition of bacterial stimulisignificantly attenuated IL-1� and CXCL2 expression, al-though considerable levels of each mediator remained(Figure 12, A and B). The reduction in proinflammatorymediator expression following minocycline treatment was
not the result of overt cell death, because cell viabilityassays revealed that minocycline was not toxic to primarymicroglia at any of the doses examined (Figure 12C).Interestingly, the inhibitory effects of minocycline werenot universal, as the compound did not have any signif-icant effects in regulating the expression of IL-12 p70 orIL-6 in response to bacterial stimulation (data not shown).
Astrocytes represent the most numerous cell type inthe brain and are recognized as one of the principalsources of CNS-derived chemokines.79,80 To our knowl-edge, the immunoregulatory effects of minocycline onastrocytic responses have not yet been investigated.Similar to what was observed with microglia, minocyclinewas capable of significantly attenuating S. aureus-depen-dent astrocyte activation in terms of NO and IL-1� re-lease, although residual expression of both mediatorswas still significant (Figure 13, A and B, respectively). Thereduction in astrocytic proinflammatory mediator expres-sion following minocycline treatment was not the result oftoxic effects at any of the concentrations examined (Fig-ure 13C). Minocycline was not effective at inhibiting mi-croglial or astrocyte activation when administered at ei-ther 1 or 6 hours following bacterial exposure (data notshown), which is not unexpected given the relativelymodest ability of the compound to affect glial proinflam-matory mediator release in the pretreatment dosing par-adigm presented.
Minocycline Attenuates TLR2 Expression inActivated Microglia and Astrocytes
TLRs enable the host to recognize microbial motifs thatare highly conserved across various classes of microor-ganisms.81–83 Among the 13 TLR family members iden-tified to date, TLR2 is capable of recognizing lipid-basedstructural components from a broad range of microbes,including various gram-positive bacteria, fungi, and pro-tozoa.81,84 We elected to focus on evaluating the effectsof minocycline on glial TLR2 expression, since we havepreviously published that this receptor plays a pivotal rolein S. aureus and PGN recognition by both microglia andastrocytes, triggering the production of a wide array ofinflammatory mediators by activated glia.69,73 Therefore,evaluation of TLR2 expression represents a read-out toimply the degree of “bacterial responsiveness” of glia toinfection. To our knowledge, the ability of minocycline tomodulate TLR2 expression in activated glia has not yetbeen examined. TLR2 expression in both microglia andastrocytes was increased following bacterial exposureand was partially suppressed by minocycline (Figure 14,A and B, respectively).
Discussion
Recent studies by others have demonstrated that mino-cycline exhibits beneficial effects in several infectiousdisease models including simian immunodeficiency vi-rus, reovirus, neuroadapted Sindbus virus, and Africantrypanosomes.61–64 Interestingly, in the human immuno-
Figure 12. Minocycline selectively inhibits S. aureus- and PGN-inducedproinflammatory mediator expression in microglia. Primary microglia werepre-treated with the indicated concentrations of minocycline for 1 hourbefore stimulation with heat-inactivated S. aureus (107 cfu/well) or PGN (10�g/ml). As a control to determine whether minocycline alone exerts anyeffects on microglial responses, cells were also subjected to treatment withthe highest dose of minocycline examined (ie, 50 �g/ml) in the absence ofbacterial stimulation. Supernatants were collected at 24 hours following S.aureus or PGN exposure, whereupon IL-1� (A) and CXCL2 (B) release werequantitated by ELISAs. Microglial viability was assessed using a standard MTTassay and the raw OD570 readings are reported (mean � SD; C). Significantdifferences between microglia exposed to S. aureus or PGN only versus cellspretreated with the various concentrations of minocycline � S. aureus orPGN are denoted with asterisks (*P � 0.01, **P � 0.001). Results arerepresentative of six independent experiments.
Minocycline Modulates Brain Abscess 1209AJP October 2007, Vol. 171, No. 4

deficiency virus-1 and reovirus studies, minocycline wasshown to attenuate viral replication, which was thought toresult from the inhibitory effects of the drug on transcrip-tion factors known to influence human immunodeficiencyvirus-1 transcription, including p38 mitogen-activatedprotein kinase.62,63 This inhibitory effect of minocyclineon human immunodeficiency virus-1 replication has alsobeen demonstrated in vitro in macrophages62 and micro-glia.50 Our present work adds to the growing list of infec-tious neuropathologies for which minocycline has beenshown to play a beneficial role. However, to our knowl-edge, this work is the first to describe an immune mod-ulatory role for minocycline in the context of a CNS bac-
terial infection independently of the drug’s antimicrobialactivity.
In bacterial meningitis, the CNS inflammatory responsethat ensues following infection plays a role in diseasepathophysiology.19,20 Specifically, numerous cytokines,free radicals, and excitatory amino acids have all beenimplicated in negatively impacting neuron survival. Al-though it was possible that if excessive, the immediateantibacterial response to S. aureus in brain abscessmight exacerbate disease severity, this possibility re-mained unclear, since we had previously demonstratedthat several inflammatory mediators, including IL-1,TNF-�, and CXCL2, were critical during the early phases
Figure 13. Minocycline inhibits S. aureus- and PGN-induced proinflamma-tory mediator expression in astrocytes. Astrocytes were pretreated with theindicated concentrations of minocycline for 1 hour before S. aureus or PGNstimulation, whereupon NO (A) and IL-1� (B) release were quantified bynitrite assay and ELISA, respectively. Astrocyte viability was assessed using astandard MTT assay, and the raw OD570 readings are reported (mean � SD;C). As a control to determine whether minocycline alone exerts any effectson astrocyte responses, cells were also subjected to treatment with thehighest dose of minocycline examined (ie, 50 �g/ml) in the absence ofbacterial stimulation. Significant differences between astrocytes exposed to S.aureus or PGN only versus cells pretreated with the various concentrationsof minocycline � S. aureus or PGN are denoted with asterisks (*P � 0.01,**P � 0.001). Results are representative of four independent experiments.
Figure 14. Minocycline attenuates the S. aureus-dependent increase in glialTLR2 expression. Primary microglia (A) and astrocytes (B) were stimulatedwith 107 cfu of heat-inactivated S. aureus or PGN (10 �g/ml) in the presenceor absence of 50 �g/ml minocycline. Protein extracts from whole cell lysateswere prepared at 24 hours following stimulation and 40 �g of total proteinfrom each sample was evaluated for TLR2 levels by Western blotting. Mem-branes were subsequently stripped and re-probed with �-actin to verifyuniformity in gel loading. Results are representative of six independentexperiments.
1210 Kielian et alAJP October 2007, Vol. 171, No. 4

of infection.7,8 With the generation of a minocycline-resis-tant S. aureus strain, our results demonstrated that mino-cycline modulates the acute host response to S. aureus inthe CNS parenchyma as evident by the fact that minocy-cline significantly reduced abscess-associated mortalityand improved brain parenchymal survival. Importantly,these effects did not result from differences in bacterialburdens, because both minocycline- and vehicle-treatedmice displayed equivalent S. aureus levels. The ability ofminocycline to minimize tissue damage may result, inpart, from a reduction in cytokine-mediated bystanderlysis, because several cytokines/chemokines were atten-uated by minocycline in brain abscesses including IL-1�,TNF-�, CXCL2, and CCL2. The ability of minocycline toattenuate IL-1 production may be due to its ability toinhibit caspase-1 activity, as previously reported by oth-ers.31,38 These findings indicate that although inflamma-tion is a prerequisite for effective pathogen clearance,this response can become exaggerated and culminate inadverse effects to the host. This dysregulated inflamma-tory response is thought to be driven, in part, by bacterialcell wall debris and/or DNA associated with the necroticabscess milieu that serve as further triggers for patternrecognition receptors such as TLRs. In addition, manyantibiotics that are used to treat CNS gram-positive in-fections can enhance the shedding of cell wall antigens,further confounding this issue.85,86 Therefore, we pro-pose that the exaggerated release of proinflammatorymediators such as IL-1�, TNF-�, and CXCL2 may con-tribute to the nonspecific destruction of surrounding nor-mal brain parenchyma by bystander lysis. In addition,minocycline was capable of reducing the S. aureus-de-pendent increase in glial TLR2 expression, suggestingthat one way minocycline may attenuate inflammation isvia interference with bacterial recognition pathways. Col-lectively, these results indicate that minocycline mayserve to balance the ensuing inflammatory response toinfection and ensure the development of an optimal an-tibacterial immune response.
One important question that remains is defining thecomplete repertoire of neuroprotective mechanisms forminocycline in the experimental brain abscess model, inaddition to its ability to modulate proinflammatory medi-ator expression as demonstrated in this study. However,the accurate identification of affected host responses willrequire the establishment of an alternative dosing para-digm to overcome obstacles related to the short half-lifeof minocycline in the mouse (ie, 2 to 3 hours),22,66,67
compounded with the intense inflammation associatedwith a bacterial infection. For example, in brain abscess,the continued proliferation of bacteria coupled with shed-ding of cell wall determinants that are potent activators ofTLRs83 probably overwhelms the anti-inflammatory ef-fects of minocycline more rapidly compared with a non-infectious inflammatory milieu that is observed in Parkin-son’s or Huntington’s disease or following spinal cordinjury. Therefore, in brain abscess we envision that acontinuous dosing strategy for minocycline would be re-quired to achieve maximal anti-inflammatory effects. Weplan to address these issues by delivering a constantsupply of minocycline via either slow release pellets or
osmotic minipumps. By maintaining therapeutic levels ofminocycline in infected animals, we anticipate that poten-tial differences in host responses will be more pro-nounced, enabling the accurate identification of the path-ways affected by minocycline. This was considered to bebeyond the scope of this initial report demonstrating theeffectiveness of minocycline in the experimental brainabscess model. Notably, we were not able to detect anysignificant effects of minocycline on astrocyte or micro-glial/macrophage activation in the peri-abscess area,suggesting that the observed differences in inflammationwere not attributable to an overt reduction in glial activa-tion using the dosing paradigms tested in the currentstudy. These results are in agreement with the relativelymodest effects of minocycline on microglial and astrocyteactivation in vitro, suggesting that mechanisms other thancytokine/chemokine production could play a role in theability of minocycline to reduce brain abscess size. How-ever, it is important to acknowledge that differences in thedegree of astrocyte and/or microglial/macrophage acti-vation could still exist but may not be discernable bymerely examining GFAP and F4/80 expression. More de-tailed studies examining the cytokine expression profilesof these cell populations upon recovery from brain ab-scesses by fluorescence-activated cell sorting are war-ranted in the context of a more continuous dosing para-digm as discussed above.
Alternative pathways that may be affected by minocy-cline to reduce tissue damage during brain abscessdevelopment include the drug’s ability to attenuate matrixmetalloproteinase expression and activity, inhibit apopto-sis, and/or regulate the influx of peripheral immune cellsto minimize nonspecific tissue destruction. The ability ofminocycline to modulate leukocyte entry into evolvingbrain abscesses could be due to a number of mecha-nisms, namely, a direct action on matrix metalloprotein-ase enzymatic activity and/or matrix metalloproteinaseexpression,44,55,66 alterations in the levels of adhesionmolecules required for leukocyte extravasation acrossthe blood-brain barrier, and/or chemokine expression.On the other hand, minocycline has been reported to actas a free radical scavenger87 and to inhibit the produc-tion of neutrophil-attracting molecules from bacteria.88
Finally, in addition to these effector mechanisms, wecannot discount a potential antiapoptotic role for minocy-cline in reducing parenchymal damage through its abilityto inhibit caspase-3 expression. Although we have not yetexamined the extent of apoptosis in the brain abscessmodel, work by others using this model have demon-strated that apoptosis represents a relatively minor com-ponent of cell death.14 Cumulatively, the reversal of thesepathways that under certain circumstances can be con-sidered detrimental to cell survival could effectively res-cue brain parenchyma from excessive destruction duringbrain abscess evolution.
In addition to its ability to modulate the host responseto infection, it is also important to acknowledge that mi-nocycline may also exert direct effects on S. aureus bymodulating its virulence factor expression profile. In sus-ceptible strains, minocycline exerts its bacteriostatic ac-tivity by binding to bacterial ribosomes and preventing
Minocycline Modulates Brain Abscess 1211AJP October 2007, Vol. 171, No. 4

protein expression.25,26 The tetM gene confers resis-tance to both minocycline and tetracycline and interfereswith antibiotic binding to bacterial ribosomes. However, itis possible that minocycline selection may alter the arrayof virulence factors produced due to direct actions ongenes important for bacterial pathogenesis. Although ourin vitro analysis of the parental and S. aureustetM strainsdid not reveal any dramatic differences in protein expres-sion profiles, variations in the face of antibiotic pressure invivo are still possible. On the other hand, the ability ofminocycline to alter the host immune response to S.aureus may lead to adaptive changes in bacterial virulencefactor expression as a result of altered selective pressuresin the brain. These possibilities could be addressed byperforming gene expression profiling microarrays of bacte-rial RNA recovered from minocycline- or vehicle-treatedmice infected with the S. aureustetM strain, studies we arecurrently exploring in the laboratory.
From a clinical perspective, any candidate immuneprophylactic agent should exhibit beneficial effects whenadministered following the onset of disease. This criterionseemed to be partially satisfied in our present study asdemonstrated by the ability of minocycline to attenuateproinflammatory mediator production and brain abscesssize when treatment was initiated 3 days following S.aureus infection. Therefore, our data suggest that mino-cycline may represent an attractive candidate for thetherapeutic management of brain abscess for the follow-ing reasons. First, the bacteriostatic properties of mino-cycline effectively reduce CNS bacterial burdens in sus-ceptible strains. Second, in addition to its antimicrobialeffects, minocycline can also attenuate pathological CNSinflammation that probably contributes to the expansionof abscess size through bystander damage to surround-ing uninfected brain parenchyma. Third, minocycline ex-hibits excellent blood-brain barrier permeability becauseof its small and lipophilic nature and, based on theseproperties, would be expected to reach high therapeuticlevels within the abscess milieu, a requisite for any effec-tive antimicrobial therapy for brain abscess.
Acknowledgments
We thank Dr. Paul Drew and Dr. Joyce DeLeo for criticalreview of the manuscript, and Anessa Haney, PatrickMayes, Jennifer Johnson, and Gail Wagoner for excellenttechnical assistance.
References
1. Mathisen GE, Johnson JP: Brain abscess. Clin Infect Dis 1997,25:763–781
2. Townsend GC, Scheld WM: Infections of the central nervous system.Adv Intern Med 1998, 43:403–447
3. Kielian T: Immunopathogenesis of brain abscess. J Neuroinflamma-tion 2004, 1:16
4. Lu CH, Chang WN, Lui CC: Strategies for the management of bacte-rial brain abscess. J Clin Neurosci 2006, 13:979–985
5. Cunha BA: Methicillin-resistant Staphylococcus aureus: clinical man-ifestations and antimicrobial therapy. Clin Microbiol Infect 2005,11(Suppl 4):33–42
6. Ruhe JJ, Monson T, Bradsher RW, Menon A: Use of long-actingtetracyclines for methicillin-resistant Staphylococcus aureusinfections: case series and review of the literature. Clin Infect Dis2005, 40:1429–1434
7. Kielian T, Barry B, Hickey WF: CXC chemokine receptor-2 ligands arerequired for neutrophil-mediated host defense in experimental brainabscesses. J Immunol 2001, 166:4634–4643
8. Kielian T, Bearden ED, Baldwin AC, Esen N: IL-1 and TNF-� play apivotal role in the host immune response in a mouse model of Staph-ylococcus aureus-induced experimental brain abscess. J Neuro-pathol Exp Neurol 2004, 63:381–396
9. Kielian T, Haney A, Mayes PM, Garg S, Esen N: Toll-like receptor 2modulates the proinflammatory milieu in staphylococcus aureus-in-duced brain abscess. Infect Immun 2005, 73:7428–7435
10. Kielian T, Cheung A, Hickey WF: Diminished virulence of an �-toxinmutant of Staphylococcus aureus in experimental brain abscesses.Infect Immun 2001, 69:6902–6911
11. Baldwin AC, Kielian T: Persistent immune activation associated with amouse model of Staphylococcus aureus-induced experimental brainabscess. J Neuroimmunol 2004, 151:24–32
12. Bloch O, Papadopoulos MC, Manley GT, Verkman AS: Aquaporin-4gene deletion in mice increases focal edema associated with staph-ylococcal brain abscess. J Neurochem 2005, 95:254–262
13. Lo WD, Wolny A, Boesel C: Blood-brain barrier permeability in staph-ylococcal cerebritis and early brain abscess. J Neurosurg 1994,80:897–905
14. Stenzel W, Soltek S, Miletic H, Hermann MM, Korner H, Sedgwick JD,Schluter D, Deckert M: An essential role for tumor necrosis factor inthe formation of experimental murine Staphylococcus aureus-in-duced brain abscess and clearance. J Neuropathol Exp Neurol 2005,64:27–36
15. Kielian T, Phulwani NK, Esen N, Syed MM, Haney AC, McCastlain K,Johnson J: MyD88-dependent signals are essential for the host im-mune response in experimental brain abscess. J Immunol 2007,178:4528–4537
16. Kielian T, Hickey WF: Proinflammatory cytokine, chemokine, andcellular adhesion molecule expression during the acute phase ofexperimental brain abscess development. Am J Pathol 2000,157:647– 658
17. Flaris NA, Hickey WF: Development and characterization of an ex-perimental model of brain abscess in the rat. Am J Pathol 1992,141:1299–1307
18. Stenzel W, Soltek S, Schluter D, Deckert M: The intermediate filamentGFAP is important for the control of experimental murine Staphylo-coccus aureus-induced brain abscess and Toxoplasma encephalitis.J Neuropathol Exp Neurol 2004, 63:631–640
19. Nau R, Bruck W: Neuronal injury in bacterial meningitis: mechanismsand implications for therapy. Trends Neurosci 2002, 25:38–45
20. Koedel U, Scheld WM, Pfister HW: Pathogenesis and pathophys-iology of pneumococcal meningitis. Lancet Infect Dis 2002, 2:721–736
21. Jonas M, Cunha BA: Minocycline. Ther Drug Monit 1982, 4:137–14522. Colovic M, Caccia S: Liquid chromatographic determination of mino-
cycline in brain-to-plasma distribution studies in the rat. J ChromatogrB Analyt Technol Biomed Life Sci 2003, 791:337–343
23. Smith DL, Woodman B, Mahal A, Sathasivam K, Ghazi-Noori S, Low-den PA, Bates GP, Hockly E: Minocycline and doxycycline are notbeneficial in a model of Huntington’s disease. Ann Neurol 2003,54:186–196
24. Agwuh KN, MacGowan A: Pharmacokinetics and pharmacodynam-ics of the tetracyclines including glycylcyclines. J Antimicrob Che-mother 2006, 58:256–265
25. Speer BS, Shoemaker NB, Salyers AA: Bacterial resistance totetracycline: mechanisms, transfer, and clinical significance. Clin Mi-crobiol Rev 1992, 5:387–399
26. Roberts MC: Update on acquired tetracycline resistance genes.FEMS Microbiol Lett 2005, 245:195–203
27. Yrjanheikki J, Keinanen R, Pellikka M, Hokfelt T, Koistinaho J: Tetra-cyclines inhibit microglial activation and are neuroprotective in globalbrain ischemia. Proc Natl Acad Sci USA 1998, 95:15769–15774
28. Yrjanheikki J, Tikka T, Keinanen R, Goldsteins G, Chan PH, KoistinahoJ: A tetracycline derivative, minocycline, reduces inflammation andprotects against focal cerebral ischemia with a wide therapeuticwindow. Proc Natl Acad Sci USA 1999, 96:13496–13500
1212 Kielian et alAJP October 2007, Vol. 171, No. 4

29. Fan LW, Lin S, Pang Y, Rhodes PG, Cai Z: Minocycline attenuateshypoxia-ischemia-induced neurological dysfunction and brain injuryin the juvenile rat. Eur J Neurosci 2006, 24:341–350
30. Liu Z, Fan Y, Won SJ, Neumann M, Hu D, Zhou L, Weinstein PR, LiuJ: Chronic treatment with minocycline preserves adult new neuronsand reduces functional impairment after focal cerebral ischemia.Stroke 2007, 38:146–152
31. Chen M, Ona VO, Li M, Ferrante RJ, Fink KB, Zhu S, Bian J, Guo L,Farrell LA, Hersch SM, Hobbs W, Vonsattel JP, Cha JH, FriedlanderRM: Minocycline inhibits caspase-1 and caspase-3 expression anddelays mortality in a transgenic mouse model of Huntington disease.Nat Med 2000, 6:797–801
32. Du Y, Ma Z, Lin S, Dodel RC, Gao F, Bales KR, Triarhou LC, ChernetE, Perry KW, Nelson DL, Luecke S, Phebus LA, Bymaster FP, PaulSM: Minocycline prevents nigrostriatal dopaminergic neurodegen-eration in the MPTP model of Parkinson’s disease. Proc Natl Acad SciUSA 2001, 98:14669–14674
33. Wu DC, Jackson-Lewis V, Vila M, Tieu K, Teismann P, Vadseth C,Choi DK, Ischiropoulos H, Przedborski S: Blockade of microglialactivation is neuroprotective in the 1-methyl-4-phenyl-1,2,3,6-tetrahy-dropyridine mouse model of Parkinson disease. J Neurosci 2002,22:1763–1771
34. Peng J, Xie L, Stevenson FF, Melov S, Di Monte DA, Andersen JK:Nigrostriatal dopaminergic neurodegeneration in the weaver mouseis mediated via neuroinflammation and alleviated by minocyclineadministration. J Neurosci 2006, 26:11644–11651
35. Seabrook TJ, Jiang L, Maier M, Lemere CA: Minocycline affectsmicroglia activation, A� deposition, and behavior in APP-tg mice. Glia2006, 53:776–782
36. Ryu JK, Franciosi S, Sattayaprasert P, Kim SU, McLarnon JG: Mino-cycline inhibits neuronal death and glial activation induced by �-amy-loid peptide in rat hippocampus. Glia 2004, 48:85–90
37. Hunter CL, Bachman D, Granholm AC: Minocycline prevents cholin-ergic loss in a mouse model of Down’s syndrome. Ann Neurol 2004,56:675–688
38. Zhu S, Stavrovskaya IG, Drozda M, Kim BY, Ona V, Li M, Sarang S,Liu AS, Hartley DM, Wu du C, Gullans S, Ferrante RJ, Przedborski S,Kristal BS, Friedlander RM: Minocycline inhibits cytochrome c releaseand delays progression of amyotrophic lateral sclerosis in mice.Nature 2002, 417:74–78
39. Kriz J, Nguyen MD, Julien JP: Minocycline slows disease progressionin a mouse model of amyotrophic lateral sclerosis. Neurobiol Dis2002, 10:268–278
40. Wells JE, Hurlbert RJ, Fehlings MG, Yong VW: Neuroprotection byminocycline facilitates significant recovery from spinal cord injury inmice. Brain 2003, 126:1628–1637
41. Lee SM, Yune TY, Kim SJ, Park do W, Lee YK, Kim YC, Oh YJ,Markelonis GJ, Oh TH: Minocycline reduces cell death and improvesfunctional recovery after traumatic spinal cord injury in the rat. J Neu-rotrauma 2003, 20:1017–1027
42. Stirling DP, Khodarahmi K, Liu J, McPhail LT, McBride CB, SteevesJD, Ramer MS, Tetzlaff W: Minocycline treatment reduces delayedoligodendrocyte death, attenuates axonal dieback, and improvesfunctional outcome after spinal cord injury. J Neurosci 2004,24:2182–2190
43. Popovic N, Schubart A, Goetz BD, Zhang SC, Linington C, DuncanID: Inhibition of autoimmune encephalomyelitis by a tetracycline. AnnNeurol 2002, 51:215–223
44. Brundula V, Rewcastle NB, Metz LM, Bernard CC, Yong VW: Target-ing leukocyte MMPs and transmigration: minocycline as a potentialtherapy for multiple sclerosis. Brain 2002, 125:1297–1308
45. Nessler S, Dodel R, Bittner A, Reuss S, Du Y, Hemmer B, Sommer N:Effect of minocycline in experimental autoimmune encephalomyelitis.Ann Neurol 2002, 52:689–690
46. Blum D, Chtarto A, Tenenbaum L, Brotchi J, Levivier M: Clinicalpotential of minocycline for neurodegenerative disorders. NeurobiolDis 2004, 17:359–366
47. Tikka TM, Koistinaho JE: Minocycline provides neuroprotectionagainst N-methyl-D-aspartate neurotoxicity by inhibiting microglia.J Immunol 2001, 166:7527–7533
48. Tikka T, Fiebich BL, Goldsteins G, Keinanen R, Koistinaho J: Mino-cycline, a tetracycline derivative, is neuroprotective against excito-toxicity by inhibiting activation and proliferation of microglia. J Neu-rosci 2001, 21:2580–2588
49. Familian A, Boshuizen RS, Eikelenboom P, Veerhuis R: Inhibitoryeffect of minocycline on amyloid beta fibril formation and humanmicroglial activation. Glia 2006, 53:233–240
50. Si Q, Cosenza M, Kim MO, Zhao ML, Brownlee M, Goldstein H, LeeS: A novel action of minocycline: inhibition of human immunodefi-ciency virus type 1 infection in microglia. J Neurovirol 2004, 10:284–292
51. Nikodemova M, Duncan ID, Watters JJ: Minocycline exerts inhibitoryeffects on multiple mitogen-activated protein kinases and IkappaBal-pha degradation in a stimulus-specific manner in microglia. J Neuro-chem 2006, 96:314–323
52. Kloppenburg M, Brinkman BM, de Rooij-Dijk HH, Miltenburg AM,Daha MR, Breedveld FC, Dijkmans BA, Verweij C: The tetracyclinederivative minocycline differentially affects cytokine production bymonocytes and T lymphocytes. Antimicrob Agents Chemother 1996,40:934–940
53. Shapira L, Barak V, Soskolne WA, Halabi A, Stabholz A: Effects oftetracyclines on the pathologic activity of endotoxin: in vitro and invivo studies. Adv Dent Res 1998, 12:119–122
54. D’Agostino P, Ferlazzo V, Milano S, La Rosa M, Di Bella G, Caruso R,Barbera C, Grimaudo S, Tolomeo M, Feo S, Cillari E: Anti-inflamma-tory effects of chemically modified tetracyclines by the inhibition ofnitric oxide and interleukin-12 synthesis in J774 cell line. Int Immuno-pharmacol 2001, 1:1765–1776
55. Power C, Henry S, Del Bigio MR, Larsen PH, Corbett D, Imai Y,Yong VW, Peeling J: Intracerebral hemorrhage induces macro-phage activation and matrix metalloproteinases. Ann Neurol 2003,53:731–742
56. Mievis S, Levivier M, Communi D, Vassart G, Brotchi J, Ledent C,Blum D: Lack of Minocycline Efficiency in Genetic Models of Hunting-ton’s Disease. Neuromolecular Med 2007, 9:47–54
57. Tsuji M, Wilson MA, Lange MS, Johnston MV: Minocycline worsenshypoxic-ischemic brain injury in a neonatal mouse model. Exp Neurol2004, 189:58–65
58. Fendrick SE, Miller KR, Streit WJ: Minocycline does not inhibit micro-glia proliferation or neuronal regeneration in the facial nucleus follow-ing crush injury. Neurosci Lett 2005, 385:220–223
59. Diguet E, Fernagut PO, Wei X, Du Y, Rouland R, Gross C, Bezard E,Tison F: Deleterious effects of minocycline in animal models of Par-kinson’s disease and Huntington’s disease. Eur J Neurosci 2004,19:3266–3276
60. Diguet E, Gross CE, Tison F, Bezard E: Rise and fall of minocycline inneuroprotection: need to promote publication of negative results. ExpNeurol 2004, 189:1–4
61. Masocha W, Rottenberg ME, Kristensson K: Minocycline impedesAfrican trypanosome invasion of the brain in a murine model. Antimi-crob Agents Chemother 2006, 50:1798–1804
62. Zink MC, Uhrlaub J, DeWitt J, Voelker T, Bullock B, Mankowski J,Tarwater P, Clements J, Barber S: Neuroprotective and anti-humanimmunodeficiency virus activity of minocycline. JAMA 2005,293:2003–2011
63. Richardson-Burns SM, Tyler KL: Minocycline delays disease onsetand mortality in reovirus encephalitis. Exp Neurol 2005, 192:331–339
64. Darman J, Backovic S, Dike S, Maragakis NJ, Krishnan C, RothsteinJD, Irani DN, Kerr DA: Viral-induced spinal motor neuron death isnon-cell-autonomous and involves glutamate excitotoxicity. J Neuro-sci 2004, 24:7566–7575
65. Lee CY, Buranen SL, Ye ZH: Construction of single-copy integrationvectors for Staphylococcus aureus. Gene 1991, 103:101–105
66. Yong VW, Wells J, Giuliani F, Casha S, Power C, Metz LM: Thepromise of minocycline in neurology. Lancet Neurol 2004, 3:744–751
67. Andes D, Craig WA: Animal model pharmacokinetics andpharmacodynamics: a critical review. Int J Antimicrob Agents 2002,19:261–268
68. Hickey MJ, Hurley JV, Morrison WA: Temporal and spatial relation-ship between no-reflow phenomenon and postischemic necrosis inskeletal muscle. Am J Physiol 1996, 271:H1277–H1286
69. Esen N, Tanga FY, DeLeo JA, Kielian T: Toll-like receptor 2 (TLR2)mediates astrocyte activation in response to the gram-positive bac-terium Staphylococcus aureus. J Neurochem 2004, 88:746–758
70. Esen N, Kielian T: Central role for MyD88 in the responses of microgliato pathogen-associated molecular patterns. J Immunol 2006,176:6802–6811
Minocycline Modulates Brain Abscess 1213AJP October 2007, Vol. 171, No. 4

71. Phulwani NK, Feinstein DL, Gavrilyuk V, Akar C, Kielian T: 15-Deoxy-�12,14-prostaglandin J2 (15d-PGJ2) and ciglitazone modulateStaphylococcus aureus-dependent astrocyte activation primarilythrough a PPAR-gamma-independent pathway. J Neurochem 2006,99:1389–1402
72. Kielian T, McMahon M, Bearden ED, Baldwin AC, Drew PD, Esen N:S. aureus-dependent microglial activation is selectively attenuated bythe cyclopentenone prostaglandin 15-deoxy-�12,14-prostaglandinJ2 (15d-PGJ2). J Neurochem 2004, 90:1163–1172
73. Kielian T, Esen N, Bearden ED: Toll-like receptor 2 (TLR2) is pivotal forrecognition of S. aureus peptidoglycan but not intact bacteria bymicroglia. Glia 2005, 49:567–576
74. Tanga FY, Nutile-McMenemy N, DeLeo JA: The CNS role of Toll-likereceptor 4 in innate neuroimmunity and painful neuropathy. Proc NatlAcad Sci USA 2005, 102:5856–5861
75. Trzcinski K, Cooper BS, Hryniewicz W, Dowson CG: Expression ofresistance to tetracyclines in strains of methicillin-resistant Staphylo-coccus aureus. J Antimicrob Chemother 2000, 45:763–770
76. Aloisi F: Immune function of microglia. Glia 2001, 36:165–17977. Hanisch UK: Microglia as a source and target of cytokines. Glia 2002,
40:140–15578. Kielian T: Microglia and chemokines in infectious diseases of the
nervous system: views and reviews. Front Biosci 2004, 9:732–75079. Dong Y, Benveniste EN: Immune function of astrocytes. Glia 2001,
36:180–190
80. Farina C, Aloisi F, Meinl E: Astrocytes are active players in cerebralinnate immunity. Trends Immunol 2007, 28:138–145
81. Akira S, Uematsu S, Takeuchi O: Pathogen recognition and innateimmunity. Cell 2006, 124:783–801
82. Konat GW, Kielian T, Marriott I: The role of Toll-like receptors in CNSresponse to microbial challenge. J Neurochem 2006, 99:1–12
83. Kielian T: Toll-like receptors in central nervous system glial inflamma-tion and homeostasis. J Neurosci Res 2006, 83:711–730
84. Kopp E, Medzhitov R: Recognition of microbial infection by Toll-likereceptors. Curr Opin Immunol 2003, 15:396–401
85. Weber JR, Moreillon P, Tuomanen EI: Innate sensors for Gram-posi-tive bacteria. Curr Opin Immunol 2003, 15:408–415
86. van der Flier M, Geelen SP, Kimpen JL, Hoepelman IM, Tuomanen EI:Reprogramming the host response in bacterial meningitis: how bestto improve outcome? Clin Microbiol Rev 2003, 16:415–429
87. Kraus RL, Pasieczny R, Lariosa-Willingham K, Turner MS, Jiang A,Trauger JW: Antioxidant properties of minocycline: neuroprotection inan oxidative stress assay and direct radical-scavenging activity.J Neurochem 2005, 94:819–827
88. Akamatsu H, Niwa Y, Kurokawa I, Masuda R, Nishijima S, Asada Y:Effects of subminimal inhibitory concentrations of minocycline onneutrophil chemotactic factor production in comedonal bacteria, neu-trophil phagocytosis and oxygen metabolism. Arch Dermatol Res1991, 283:524–528
1214 Kielian et alAJP October 2007, Vol. 171, No. 4









![[Meticilin resistant Staphylococcus aureus and liver abscess: a retrospective analysis of 117 patients]](https://static.fdokumen.com/doc/165x107/632546fd545c645c7f099e01/meticilin-resistant-staphylococcus-aureus-and-liver-abscess-a-retrospective-analysis.jpg)










