
The Role of Indoleamine 2,3Dioxygenase in a Mouse Model of Neuroinflammation-Induced Depression
11
Journal of Alzheimer’s Disease 28 (2012) 905–915 DOI 10.3233/JAD-2011-111097 IOS Press 905 The Role of Indoleamine 2,3-Dioxygenase in a Mouse Model of Neuroinflammation- Induced Depression Nikoletta Dobos a,b , Erik F.J. de Vries c , Ido P. Kema d , Konstantinos Patas a , Marloes Prins a , Ingrid M. Nijholt a , Rudi A. Dierckx c , Jakob Korf b , Johan A. den Boer b,c , Paul G.M. Luiten a,b and Ulrich L.M. Eisel a,∗ a Department of Molecular Neurobiology, University of Groningen, Groningen, The Netherlands b Department of Psychiatry, University Medical Center Groningen, Groningen, The Netherlands c Department of Nuclear Medicine and Molecular Imaging, University Medical Center Groningen, Groningen, The Netherlands d Department of Laboratory Medicine, University Medical Center Groningen, Groningen, The Netherlands Handling Associate Editor: Tiziana Borsello Accepted 1 November 2011 Abstract. Indoleamine 2,3-dioxygenase (IDO), an enzyme which is activated by pro-inflammatory cytokines, has been sug- gested as a potential link between neuroinflammatory processes in neurodegenerative diseases (like Alzheimer’s disease) and depression. The present study aimed to determine whether neuroinflammation-induced increased IDO levels in the mammalian brain will lead to depressive-like behavior. Neuroinflammation was initiated in mice by a single intracerebroventricular injection of lipopolysaccharide (LPS). Cerebral inflammation was monitored 1, 2, 3 and 4 days after the injection with small-animal positron emission tomography (PET) using the inflammatory marker [ 11 C]-PK11195. In the presence or absence of systemically applied 1-methyl-tryptophan (1-MT), a competitive IDO-inhibitor, we assessed the development of depressive-like behavioral symptoms in parallel with IDO expression and activity. The PK11195 PET signal reached a highly significant peak 3 days after LPS injection, while these animals displayed a significant increase of depressive-like behavior in the forced swim test compared to vehicle-injected animals. These findings were paralleled by a significant increase of IDO in the brainstem, and an increased kynurenine/tryptophan ratio in the serum. Moreover, we report here for the first time, that inhibition of IDO by 1-MT in centrally induced neuroinflammation under experimental conditions can prevent the development of depressive-like behavior. Keywords: Depression, indoleamine 2,3-dioxygenase, lipopolysaccharide, neuroinflammation, positron emission tomography INTRODUCTION Neuroinflammation is defined as the activation of an immune response in the central nervous system (CNS) [1]. During neuroinflammation, microglia, the most ∗ Correspondence to: U.L.M. Eisel, Department of Molecular Neurobiology, Center for Life Sciences, Nijenborgh 7, 9747 AG Groningen, The Netherlands. Tel.: +31 50 363 2366; Fax: +31 50 363 2331; E-mail: [email protected]. important resident immune cells, become activated [2] and, as a consequence, their morphology starts to change and secretion of pro-inflammatory cytokines, such as interferon (IFN) [3], tumor necrosis factor (TNF) [4], or interleukin 6 [5], is initiated. An impor- tant hallmark of activated microglia is the expression of peripheral benzodiazepine receptors (PBRs) on the outer membrane of the mitochondria, which allows the non-invasive detection their activated state by means of labeled PBR ligands. Ligands such as the ISSN 1387-2877/12/$27.50 © 2012 – IOS Press and the authors. All rights reserved
Transcript of The Role of Indoleamine 2,3Dioxygenase in a Mouse Model of Neuroinflammation-Induced Depression

Journal of Alzheimer’s Disease 28 (2012) 905–915DOI 10.3233/JAD-2011-111097IOS Press
905
The Role of Indoleamine 2,3-Dioxygenase ina Mouse Model of Neuroinflammation-Induced Depression
Nikoletta Dobosa,b, Erik F.J. de Vriesc, Ido P. Kemad, Konstantinos Patasa, Marloes Prinsa,Ingrid M. Nijholta, Rudi A. Dierckxc, Jakob Korfb, Johan A. den Boerb,c, Paul G.M. Luitena,b
and Ulrich L.M. Eisela,∗aDepartment of Molecular Neurobiology, University of Groningen, Groningen, The NetherlandsbDepartment of Psychiatry, University Medical Center Groningen, Groningen, The NetherlandscDepartment of Nuclear Medicine and Molecular Imaging, University Medical Center Groningen, Groningen,The NetherlandsdDepartment of Laboratory Medicine, University Medical Center Groningen, Groningen,The Netherlands
Handling Associate Editor: Tiziana Borsello
Accepted 1 November 2011
Abstract. Indoleamine 2,3-dioxygenase (IDO), an enzyme which is activated by pro-inflammatory cytokines, has been sug-gested as a potential link between neuroinflammatory processes in neurodegenerative diseases (like Alzheimer’s disease) anddepression. The present study aimed to determine whether neuroinflammation-induced increased IDO levels in the mammalianbrain will lead to depressive-like behavior. Neuroinflammation was initiated in mice by a single intracerebroventricular injectionof lipopolysaccharide (LPS). Cerebral inflammation was monitored 1, 2, 3 and 4 days after the injection with small-animalpositron emission tomography (PET) using the inflammatory marker [11C]-PK11195. In the presence or absence of systemicallyapplied 1-methyl-tryptophan (1-MT), a competitive IDO-inhibitor, we assessed the development of depressive-like behavioralsymptoms in parallel with IDO expression and activity. The PK11195 PET signal reached a highly significant peak 3 days afterLPS injection, while these animals displayed a significant increase of depressive-like behavior in the forced swim test comparedto vehicle-injected animals. These findings were paralleled by a significant increase of IDO in the brainstem, and an increasedkynurenine/tryptophan ratio in the serum. Moreover, we report here for the first time, that inhibition of IDO by 1-MT in centrallyinduced neuroinflammation under experimental conditions can prevent the development of depressive-like behavior.
Keywords: Depression, indoleamine 2,3-dioxygenase, lipopolysaccharide, neuroinflammation, positron emission tomography
INTRODUCTION
Neuroinflammation is defined as the activation of animmune response in the central nervous system (CNS)[1]. During neuroinflammation, microglia, the most
∗Correspondence to: U.L.M. Eisel, Department of MolecularNeurobiology, Center for Life Sciences, Nijenborgh 7, 9747 AGGroningen, The Netherlands. Tel.: +31 50 363 2366; Fax: +31 50363 2331; E-mail: [email protected].
important resident immune cells, become activated[2] and, as a consequence, their morphology starts tochange and secretion of pro-inflammatory cytokines,such as interferon � (IFN�) [3], tumor necrosis factor �(TNF�) [4], or interleukin 6 [5], is initiated. An impor-tant hallmark of activated microglia is the expressionof peripheral benzodiazepine receptors (PBRs) on theouter membrane of the mitochondria, which allowsthe non-invasive detection their activated state bymeans of labeled PBR ligands. Ligands such as the
ISSN 1387-2877/12/$27.50 © 2012 – IOS Press and the authors. All rights reserved

906 N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO?
isoquinoline PK11195 (1-[2-chlorophenyl]-N-methyl-N-[1-methylpropyl]-3-isoquinoline carboxamide)bind the 18-kDa subunit of the PBRs [6]. Whenlabeled with carbon-11, PK11195 can be used as asensitive marker for positron emission tomography(PET) to visualize and quantify microglial activationin vivo associated with various conditions triggered byinflammatory or neurodegenerative processes [6, 7].
Chronic inflammatory changes have been shown toplay a key role in the pathogenesis of neurodegen-erative diseases, like Alzheimer’s disease (AD) [8].The typical hallmarks of AD are extracellular depo-sitions of amyloid-�, intracellular aggregates of theprotein tau, but besides these features, activation ofmicroglia and elevated cytokine and chemokine levelshave been shown as well [8]. It has been shown thatAD is often accompanied by symptoms of depression,and patients are often being treated for depression sev-eral years before clinical diagnosis of AD occurs [9].Meta-analysis on retrospective and prospective studiesrevealed that the history of depression approximatelydoubles the risk of developing dementia. Several stud-ies support the hypothesis that depression is a likelyrisk factor for dementia in general and for AD specifi-cally [10, 11].
Inflammation is an important hallmark of sepsis aswell. It has been recently shown that severe sepsis canlead to cognitive impairment and functional disability[12].
During peripheral infection, either acute or chronic,the immune system is activated, e.g., by macrophagestimulation, and pro-inflammatory cytokines arereleased, which also act on the brain causing sicknessbehavior. Sickness behavior shares similar featureswith major depression. Both are characterized bymalaise, weakness, loss of appetite, lethargy, ordecreased interest in the surroundings. However,sickness behavior breaks off when the pathogen iseliminated. When the peripheral immune system iscontinuously activated, like in chronic inflammatorydiseases, such as rheumatoid arthritis, osteoarthritis, orinflammatory lung disease, the resulting chronic acti-vation of the immune system in the brain can lead tothe development of depression. Depression may thusrepresent a maladaptive version of cytokine-inducedsickness behavior [13]. In 1991, R.S. Smith proposedthe macrophage theory of depression [14], whichtriggered a growing body of evidence supportingthe notion that inflammation can increase the risk ofdeveloping major depression [15]. The overall ideabeing that the overexpression of pro-inflammatoryagents is associated with increased activity of the
ubiquitous intracellular enzyme indoleamine-2,3-dioxygenase (IDO), which catalyzes tryptophancatabolism through the kynurenine pathway [13, 16].The depletion of tryptophan in brain cells reduces theproduction of brain serotonin (5-HT) [13, 16]. Thedegradation of tryptophan along the kynureninepathway also generates neurotoxins, like quinolinicacid (QUIN), an NMDA receptor agonist, or 3-hydroxykynurenine (3-HK), which leads to apoptosisin neurons, which can add to local excitotoxic neuronaloverstimulation next to modulating serotonergic neu-rotransmission [17]. Taken together, cytokine-inducedIDO-mediated tryptophan depletion [18] and QUIN-mediated neurotoxicity [17, 19] are hypothesized tobe involved in the pathophysiology of mood disorders,like major depression. Nevertheless, it has been alsoshown that the metabolism of tryptophan by IDOthrough the kynurenine pathway is increased in AD[20]. Lipopolysaccharide (LPS) injection in rodentsis often used as an experimental model for systemicimmune challenges. LPS-injected mice display imme-diate sickness behavior after peripheral injection ofLPS whereas depressive-like behavior seems to emergein a shifted temporal frame, when sickness is alreadyabated [13, 21]. Accordingly, IDO activation andconcomitant depressive-like behavior has been shownto be causally interrelated in this mouse model [22].
Previous studies demonstrated that systemic expo-sure to LPS induces chronic central neuroinflammation[23] as well as cerebral IDO activation [24, 25]. Inour study, we measured IDO activity and expres-sion after intracerebroventricular LPS injection andassessed the development of depressive-like symptomsin the presence or absence of 1-methyl tryptophan(1-MT), a competitive IDO-inhibitor. Neuroinflamma-tion was monitored over time by small animal positronemission tomography (PET).
We report here for the first time that the inhibitionof IDO in centrally induced neuroinflammation is suf-ficient to prevent the development of depressive-likebehavior in mice.
MATERIALS AND METHODS
Ethics statement
All animal care and treatments were reviewed andapproved by the Ethical Committee for the use ofexperimental animals of the University of Groningenunder license number DEC5461.

N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO? 907
Animals
Three months old male C57Bl/6J mice(n = 8–10/group, groups: Placebo/PBS, 1-MT/PBS,Placebo/LPS, 1-MT/LPS) were obtained from Harlan(Horst, The Netherlands). Animals were individuallyhoused under normal laboratory conditions (air-conditioned, 21 ± 2◦C), humidity-controlled room,12/12 h light/dark cycle (light on from 08 : 00–20 : 00).Food and water were available ad libitum. In all casesthe weight of the animals was monitored on a dailybasis.
Intracerebroventricular LPS injection
Before fixation in a stereotactic apparatus mice wereanaesthetized with avertin (250 mg/kg by intraperi-toneal injection). Periprocedural analgesia was pro-vided with finadyne (2.5 mg/kg, subcutaneously).The coordinates for the bilateral intracerebroventric-ular (ICV) injections were −2.5 mm dorsal/ventral,−1.0 mm lateral, and −0.5 mm anterior/posterior frombregma [26]. The holes were drilled perpendicularly tothe previously exposed skull. 1 �l endotoxin-free phos-phate buffered saline (PBS) or 5 �g lipopolysaccharide(LPS, L-6529, serotype 055:B5, Sigma-Aldrich)dissolved in 1 �l PBS was injected into the lateral ven-tricles using a 1 �l Hamilton needle (cat. nr.170431,Omnilabo) fitted to a 25 �l Hamilton syringe. This doseof LPS was previously shown to induce significantneuroinflammation [27–30]. A syringe pump (TSEsystem, Bad Homburg, Germany) was used for injec-tion at a constant rate of 0.3 �l/ min. After injection,the needle was kept in the ventricles for an additional5 min and subsequently slowly removed from the brain.The incision was sealed with dental cement.
Slow-release pellet implantation
Four days prior to ICV injection, mice were ini-tially anaesthetized by 5% isoflurane in an inductionchamber. After anesthetic induction, the animals wereplaced in an open circuit system and 2% isofluranewas administered via a nose-cone. The competitiveIDO inhibitor 1-methyl-DL-tryptophan (1-MT) wasapplied via a slow release pellet implanted subcuta-neously beneath the dorsal skin surface. Pellets weredesigned to continuously release 5 mg/day of drug for21 days (Innovative Research of America, Sarasota,FL, USA) [22]. Similar drug free pellets served asplacebo.
Synthesis of [11C]-PK11195
The PET tracer [11C]-PK11195 was produced at theUniversity Medical Center Groningen according to theprocedure described by Cremer et al. [31]. The prod-uct was obtained in 32 ± 10% radiochemical yield as asterile solution in 10% ethanol in saline. The radio-chemical purity was always >95% and the specificactivity was 110 ± 69 GBq/� mol.
Micro-PET imaging
Mice were anaesthetized by 5% isoflurane (Phar-machemie, The Netherlands) mixed with medical airin an induction chamber. Anesthesia was maintainedwith 2% isoflurane in medical air at a flow of 2 ml/minvia a nose-cone. Mice were injected intravenously with7.5 ± 2.5 MBq [11C]-PK11195 via the penile vein andpositioned in a small animal PET camera (Focus 220,Siemens Medical Solutions USA, Inc.) in a transaxialposition with their heads in the field of view. A trans-mission scan with a 57Co point source was acquiredfor the correction of attenuation by tissue. Ten min-utes after tracer injection, a 30-min static emissionscan was started. After completion of the emissionscan, the animals were allowed to recover. Emissionsinograms were iteratively reconstructed (OSEM2d,4 iterations) after being normalized, corrected forattenuation and decay of radioactivity. Regions-of-interest (ROI) were manually drawn over the wholemouse brain in the reconstructed images using themicroPET® ASIProTM VM6.6.2.0 software. Traceruptake in ROI was obtained in Bq/cm3 and convertedinto the standardized uptake value (SUV), defined as:[tissue activity concentration (MBq/cm3)]/[(injecteddose (MBq)/body weight (g)].
Elevated plus maze
The elevated plus maze test was used to measureanxiety-related exploration [32]. The test consistedof a plus-shaped maze, 50 cm above the floor, withtwo opposite arms closed by high side walls and twoopposite open arms without walls (50 cm long, 5 cmwide). The mouse was placed in the center of the mazeand was allowed to explore the maze freely for 8 min.Exploratory activity, time spent in the dark arms, openarms, and center zone were recorded visually by tworaters (one of them was blind to the experiment). Theratio of time spent in the open (light) and closed (dark)arms and in the center to total time spent in the mazewas calculated for each group. A lower ratio of time

908 N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO?
spent in the open arms and total time is indicative forhigher anxiety levels.
Spontaneous alternation
Short-term spatial memory performance (workingmemory) was investigated by recording spontaneous(since it is not reinforced) alternation behavior in aY-maze paradigm [33]. The maze consisted of threetubular and transparent Plexiglas arms forming the Y.All three arms were 5 cm in diameter, 27.5 cm long,and at a 120◦ angle from each other. The experimentalroom contained visual cues for spatial orientation. Themouse (naive to the maze) was placed into the centerof the Y maze and allowed to explore the maze freelyduring an 8-min session. The series of arm entries(considered to be completed when all four paws ofthe animal entered the arm) was recorded visually bytwo raters (one of them was blind to the experiment).Alternation was defined as successive entries into thethree arms on overlapping triplet sets. Working mem-ory was assessed by the alternation percentage, whichwas calculated as the ratio of actual to possible alterna-tions (defined as the total number of arm entries minustwo). Exploratory activity was assessed by countingthe total number of arm entries.
Forced swim test
The forced swim test is a widely used paradigmto measure antidepressant activity and depressive-like behavior in animal models [34]. A cylinder(22 cm height × 11.5 cm diameter) was filled withwater (23–25◦C) up to a height of 17 cm. The animalwas placed inside the cylinder and immobility timewas recorded over a 7 min test period. Afterwards themouse was dried and returned to its cage. Betweentesting sessions the water was refreshed.
Protein analysis
Immediately after the PET scans and behavioralexperiments, mice were sacrificed by CO2 inhalation.Brains were quickly dissected, snap-frozen in liquidnitrogen and stored at −80◦C until homogenizationand Western blotting [35]. Antibodies (Santa CruzBiotechnology, Inc., Santa Cruz, CA, USA) wereused for IDO (rat monoclonal antibody), and HPRT(hypoxanthine-guanine phosphoribosyltransferase) asan internal standard to correct for variations in proteincontent (rabbit polyclonal antibody), and stainedwith horseradish peroxidase-conjugated secondary
antibodies (goat-anti-mouse, donkey-anti-rabbit, res-pectively). The proteins were detected using EnhancedChemiluminescence (Pierce ECL Western BlottingSubstrate, Rockford, IL, USA) and the MolecularImage Chemidoc XRS+ System (Biorad). The resultswere analyzed using Image Lab software (BioRad).
Serum tryptophan and kynurenine quantification
We used as an in vivo index of IDO activity the ratioof the blood levels of kynurenine/tryptophan. Never-theless, it is known, that tryptophan 2,3-dioxygenase(TDO) also catabolyzes tryptophan into kynurenine,but since 1-MT specifically inhibits IDO, but notTDO, we could use the results of the ratio of kynure-nine/tryptophan as a reflection of IDO activity [36].Tryptophan and kynurenine concentrations in serumwere determined by an automated on-line solid-phase extraction-liquid chromatographic-tandem massspectrometric (XLC-MS/MS) method with deuteratedinternal standards as described previously [37].
Statistical analysis
The data in Fig. 1F were analyzed by 2-way analy-sis of variance (ANOVA), and the data in Fig. 4. wereanalyzed by 3-way ANOVA with repeated measures.All other data were analyzed by one-way ANOVA,followed by the LSD post-hoc test to determine thedifferences between the selected groups using theprogram SPSS 16.0 for Windows. *p < 0.05 was con-sidered to be statistically significant (**p < 0.005,highly significant; ***p < 0.0005, extremely signif-icant). Data are presented as a mean value ± thestandard error of the mean (SEM).
RESULTS
Intracerebroventricular LPS injection-inducedneuroinflammation culminates 3 days afterinjection
We evaluated the temporal course of in vivo cere-bral inflammation in the mouse brain using micro-PET(Fig. 1D). PK11195 binding potential peaked 3 daysafter the ICV injection of either LPS (F(7,57) = 5.099,p < 0.0005) or, interestingly, also after injection of PBS(p ≤ 0.05) with similar kinetics. Glial activation wassignificantly higher at day 3 and 4 in LPS injected micewhen compared to PBS injected mice. Already on thefourth post-operative day, the inflammatory responsestarted to decline (Fig. 1D).
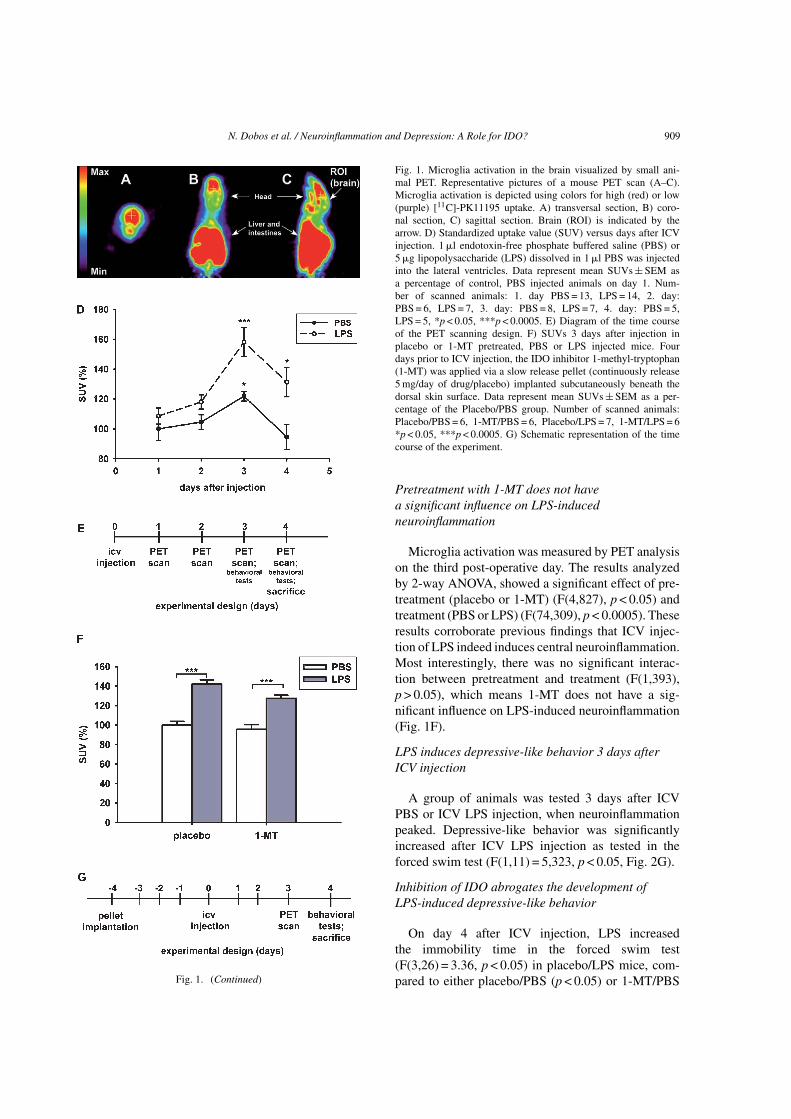
N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO? 909
Fig. 1. (Continued)
Fig. 1. Microglia activation in the brain visualized by small ani-mal PET. Representative pictures of a mouse PET scan (A–C).Microglia activation is depicted using colors for high (red) or low(purple) [11C]-PK11195 uptake. A) transversal section, B) coro-nal section, C) sagittal section. Brain (ROI) is indicated by thearrow. D) Standardized uptake value (SUV) versus days after ICVinjection. 1 �l endotoxin-free phosphate buffered saline (PBS) or5 �g lipopolysaccharide (LPS) dissolved in 1 �l PBS was injectedinto the lateral ventricles. Data represent mean SUVs ± SEM asa percentage of control, PBS injected animals on day 1. Num-ber of scanned animals: 1. day PBS = 13, LPS = 14, 2. day:PBS = 6, LPS = 7, 3. day: PBS = 8, LPS = 7, 4. day: PBS = 5,LPS = 5, *p < 0.05, ***p < 0.0005. E) Diagram of the time courseof the PET scanning design. F) SUVs 3 days after injection inplacebo or 1-MT pretreated, PBS or LPS injected mice. Fourdays prior to ICV injection, the IDO inhibitor 1-methyl-tryptophan(1-MT) was applied via a slow release pellet (continuously release5 mg/day of drug/placebo) implanted subcutaneously beneath thedorsal skin surface. Data represent mean SUVs ± SEM as a per-centage of the Placebo/PBS group. Number of scanned animals:Placebo/PBS = 6, 1-MT/PBS = 6, Placebo/LPS = 7, 1-MT/LPS = 6*p < 0.05, ***p < 0.0005. G) Schematic representation of the timecourse of the experiment.
Pretreatment with 1-MT does not havea significant influence on LPS-inducedneuroinflammation
Microglia activation was measured by PET analysison the third post-operative day. The results analyzedby 2-way ANOVA, showed a significant effect of pre-treatment (placebo or 1-MT) (F(4,827), p < 0.05) andtreatment (PBS or LPS) (F(74,309), p < 0.0005). Theseresults corroborate previous findings that ICV injec-tion of LPS indeed induces central neuroinflammation.Most interestingly, there was no significant interac-tion between pretreatment and treatment (F(1,393),p > 0.05), which means 1-MT does not have a sig-nificant influence on LPS-induced neuroinflammation(Fig. 1F).
LPS induces depressive-like behavior 3 days afterICV injection
A group of animals was tested 3 days after ICVPBS or ICV LPS injection, when neuroinflammationpeaked. Depressive-like behavior was significantlyincreased after ICV LPS injection as tested in theforced swim test (F(1,11) = 5,323, p < 0.05, Fig. 2G).
Inhibition of IDO abrogates the development ofLPS-induced depressive-like behavior
On day 4 after ICV injection, LPS increasedthe immobility time in the forced swim test(F(3,26) = 3.36, p < 0.05) in placebo/LPS mice, com-pared to either placebo/PBS (p < 0.05) or 1-MT/PBS

910 N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO?
Fig. 2. The effect of neuroinflammation on behavior and the consequences of IDO inhibition. A) Forced swim test, a widely used paradigmto measure antidepressant activity and depressive-like behavior in animal models. The animal was placed inside the cylinder and immobilitytime was recorded over a 7 min test period. Data represent mean values ± SEM, *p < 0.05. B) Elevated plus maze test was used to measureanxiety-related exploration. The mouse was placed in the center of the maze and was allowed to explore the maze freely for 8 min. Exploratoryactivity, time spent in the dark arms, open arms and center zone were recorded visually. The ratio of time spent in the open and closed armsand in the center was calculated for each group. A lower ratio of time spent in the open arms and total time is indicative for higher anxietylevels. Data represent mean values ± SEM in percentage. C, D) Spontaneous alternation test. Short-term spatial memory performance (workingmemory) was investigated by recording spontaneous alternation behavior in a Y-maze paradigm. The mouse was placed into the centre of the Ymaze and allowed to explore the maze freely during an 8-min session. The series of arm entries was recorded visually. Alternation was definedas successive entries into the three arms on overlapping triplet sets. Working memory was assessed by the alternation percentage, which wascalculated as the ratio of actual to possible alternations (defined as the total number of arm entries minus two). Exploratory activity was assessedby counting the total number of arm entries. Data represent mean values ± SEM, *p < 0.05. E) Spontaneous alternation test 3 days after ICVPBS or ICV LPS injection. Data represent mean values ± SEM in percentage. F) Forced swim test 3 days after ICV PBS or ICV LPS injection.Data represent mean values ± SEM, *p < 0.05.

N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO? 911
Fig. 3. A) IDO protein expression in different regions of the brain. Mice were sacrificed, brains were quickly dissected, snap-frozen andWestern blotting was done. Data represent mean values ± SEM as a percentage of the Placebo/PBS group. *p < 0.05, **p < 0.005. B) IDOactivity in serum. Tryptophan and kynurenine concentrations in serum were determined by an automated on-line solid-phase extraction-liquidchromatographic-tandem mass spectrometric (XLC-MS/MS) method. IDO activity is defined as the ratio of the concentration of kynurenineand tryptophan. Data represent mean values ± SEM as a percentage of the Placebo/PBS group. ***p < 0.0005.
(p < 0.05) mice. This effect of LPS was completelyinhibited by 1-MT pretreatment (p < 0.05) (Fig. 2A).These results demonstrated that IDO inhibition prior toICV LPS injection was sufficient to prevent the devel-opment of LPS-induced depression-like behavior.
ICV administered LPS did not affectanxiety-related behavior or working memory
ICV administered LPS did not affect anxiety behav-ior in the elevated plus maze on day 4 (time spent inthe open arms (F(3,31) = 0.41, p > 0.05), in the center(F(3,31) = 0.84, p > 0.05), in the dark (F(3,31) = 0.82,p > 0.05) when compared to placebo/PBS injectedmice (Fig. 2B). 1-MT also did not affect anxietybehavior (time spent in the open arms (F(3,31) = 0.41,p > 0.05), in the center (F(3,31) = 0.84, p > 0.05), inthe dark (F(3,31) = 0.82, p > 0.05) when compared toplacebo/PBS or placebo/LPS injected animals. Work-ing memory, measured in the spontaneous alternationtask (number of alternations (F(3,33) = 0.89, p > 0.05,Fig. 2C) and exploratory behavior (number of entries(F(3,33) = 2.08, p > 0.05, Fig. 2D) were not affectedby LPS treatment nor by application of 1-MT. Over-all, these results showed that neither ICV injected LPSnor concurrent IDO inhibition could affect anxiety-likebehavior and working memory performances.
Also, a separate group of animals was tested 3days after ICV PBS or ICV LPS injection. Noabnormalities were found in the working memory(number of alternations: F(1,13) = 1.117, p > 0.05,Fig. 2E) and exploratory behavior (number of entries:
F(1,13) = 1.859, p > 0.05, data not shown) as tested inspontaneous alternation test.
LPS treatment increases IDO levels in thebrainstem 4 days after injection
A significant increase of IDO (F(3,131) = 6.388,p < 0.05) was detected in the brainstem of placebo/LPS(p < 0.05) and 1-MT/LPS (p < 0.005), compared toboth PBS groups (Fig. 3A). In the other brainregions, no significant differences among thefour groups were observed (cerebellum (F(3,49) =0.37, p > 0.05), cortex (F(3,52) = 0.24, p > 0.05),hippocampus (F(3,93) = 0.36, p > 0.05), hypothalamus(F(3,51) = 0.70, p > 0.05), olfactory bulb (F(3,109) =0.69, p > 0.05), and striatum (F(3,56) = 0.815,p > 0.05).
1-MT inhibits IDO activity in the serum after ICVLPS injection
ICV LPS injection led to increased IDO activ-ity, defined as a ratio of kynurenine and tryptophan,measured in the serum, which could be completelyinhibited by 1-MT (F(3,26) = 10.611, p < 0.0005)(Fig. 3B).
ICV injection of LPS results in a robust reductionin body weight
Cerebral inflammation was paralleled by con-siderable weight reductions in both LPS groupsafter injection, but was normalized by the time of

912 N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO?
Fig. 4. Changes of the body weight of the animals during the experiment. ICV LPS injection took place at day 0. Data represent meanvalues ± SEM.
behavioral assessment (Fig. 4). The data were ana-lyzed by 3-way ANOVA with repeated measures,showing a significant effect of time (days) (F(36,101),p < 0.0005) and also, there was a significant interactionbetween time and treatment (PBS or LPS) (F(22,041),p < 0.0005). There was no interaction effect betweenpretreatment (placebo or 1-MT) and treatment (PBS orLPS) (F(6,000), p > 0.05). Source of significance wasanalyzed by univariate measures. No significant differ-ence was observed on day −4 (F(0,454), p > 0.05), day−3 (F(0,453), p > 0.05), day 0 (F(0,777), p > 0.05), andday 4 (F(4,022), p > 0.05). There was a significant dif-ference on day 1 (F(8,661), p < 0.05), day 2 (F(10,662),p < 0.05), and day 3 (F(7,162), p < 0.05).
DISCUSSION
In the study presented here we used an animal modelfor depression in neuroinflammatory conditions, suchas AD. Our results provide some evidence that LPS-induced neuroinflammation leads to increased IDOprotein levels in the brainstem, which is accompaniedby the onset of depressive-like behavior. This behav-ioral phenomenon can be completely blocked by IDOinhibition suggesting a causal relation between thebehavioral expression and IDO activation.
Several studies have shown that peripheral inflam-mation can cause central neuroinflammatory responseswhich might be causal for depressive-like behavior.Although the question still remains whether centralinflammation alone is sufficient to lead to depressive-like behavior.
O’Connor and colleagues showed that the inhibi-tion of IDO by 1-MT in the context of systemic LPS
administration can normalize plasma and brain kynure-nine/tryptophan ratios, while only kynurenine levelswere altered [22], which is consistent with the data pre-sented here. We also found a significant increase of thekynurenine/tryptophan ratio in the serum of ICV LPS-injected animals, whereas tryptophan levels remainedunchanged.
We used microPET to monitor the ongoing cerebralinnate immune responses and dissociate cytokine-induced sickness from depressive-like behavior. Ourfindings clearly showed that cerebral inflammationwas considerably higher, as expected, in ICV LPSinjected mice and reached its peak 3 days after the ICVinjection. The 4th post-injection day (when sickness-behavior is imperceptible) was chosen to performbehavioral studies to measure depressive-like behavior.On the 4th post-injection day (when the placebo/LPStreated animals exhibited increased depressive-likebehavior in the forced swim test), cerebral inflam-mation was almost gone. This was confirmed byreduced body weight in both LPS groups during thefirst two post-injection days and subsequent restora-tion by the time of behavioral assessment (Fig. 4).The reduced food intake might affect the tryptophanlevels in the blood, but that cannot be the reason forthe higher kynurenine/tryptophan ratio, since the 1-MT/LPS group showed a dramatic decrease in the bodyweight as well, but did not show increased kynure-nine/tryptophan ratio. No differences were observedin the general activity of the mice when the behav-ioral tests were done, as seen also in the total armentries in the elevated plus maze test (data not shown)or in the spontaneous alternation test (Fig. 2). Wealso did not observe any differences in working

N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO? 913
memory, exploratory behavior and anxiety-like behav-ior between PBS and LPS-treated groups (Fig. 2),indicating that locomotor activity was normal by thetime mice were tested in the forced swim test.
Behavioral tests were also performed on day 3(Fig. 2), showing the same results, namely: workingmemory and exploratory behavior were not affected,as tested in the spontaneous, alternation test, but sig-nificantly increased immobility was observed in theforced swim test in the LPS-treated group. Statisti-cal analysis showed that depressive-like behavior afterICV LPS injection was more robust on the 4th post-operative day than on the 3rd (p values, 0.02 and0.04, respectively), making the hypothesis stronger,that depressive-like behavior lasts longer after a neu-roinflammatory response.
It is important to mention that a limitation ofthe present study, especially in relation to neurode-generative diseases, e.g., AD, is that no long-termconsequences of ICV LPS-injection and/or IDO inhi-bition were assessed. Indeed, it has been previouslyshown that chronic LPS infusion directly in the 4thventricle in rats can result in impaired cognitive per-formance [38]. We therefore hypothesize that an acuteand transient LPS challenge might not be sufficient tounequivocally confound the elaborate and pleiotropicphysiology of brain cytokines normally involved incognitive processes [39].
Taken together, our findings further support thehypothesis that depression-like behavior remains evenwhen temporally induced sickness behavior haswaned. The temporal profile showed a different pat-tern from that described in previous studies [40, 41],but it should be noted that we used larger doses ofLPS to induce the immunological challenge in ourstudy. More importantly, we mainly measured reac-tivity of resident brain cells in response to ICV LPSinjection, which might be delayed when compared tothe immediate sickness-inducing vagal afference andmonocytic infiltration to the CNS following systemicLPS exposure.
We further demonstrate for the first time that theinhibition of IDO by 1-MT pretreatment in the con-text of centrally LPS-induced neuroinflammation issufficient to prevent the development of depressive-like behavior. Since IDO in our study was inhibitedsystemically [22], we cannot ascertain whether thisfinding is associated with direct central IDO inhibitionor reduction of CNS influx of peripheral kynurenine,which has been previously shown to induce depressive-like behavior [22]. It may very well be the resultof a combined effect of peripheral and central IDO
inhibition. It has been shown that even much lowerdoses of LPS can cross the brain to blood direction[42], leading to a subsequent peripheral inflamma-tion, as it has been already also shown that lowerdose of ICV injected LPS leads to increased produc-tion of central and peripheral TNF� [43]. We alsomeasured TNF� expression in the liver and found anincrease of TNF� in both LPS treated groups (Resultsin %: Placebo/PBS: 100 ± 12, 1-MT/PBS: 99 ± 12,Placebo/LPS: 131 ± 15, 1-MT/LPS: 154 ± 15) indicat-ing peripheral inflammation in our paradigm, althoughthis effect is not because of the dose of LPS we usedin this study, but because the immune privilege of thebrain is not absolute and there is a constitutive com-munication of the brain and the periphery [44].
It was shown that IDO mRNA expression wasreduced in whole brain homogenates of 1-MT/LPStreated mice [22], which, however, was not observedin the context of Bacille Calmette-Guerin-inducedinfection [45]. In any case, those measurements wereconducted in whole brain homogenates, so we can-not exclude the possibility that brain IDO expressionor cytokine signaling is regionally altered. Indeed, ourdata show that IDO protein expression is increased inthe brainstem of LPS injected mice, independently ofpretreatment with placebo or 1-MT. Although we didnot measure cytokine expression levels in the brain,our results clearly argue in favor of a post-translationalinhibition of IDO by 1-MT. Down-regulation of IDOexpression in other brain regions was not found, buteven if it was, this does not necessarily mean it wouldbe a direct effect of 1-MT.
Our PET findings are consistent with a moderate,but not significant anti-inflammatory effect of 1-MT, asshown by the reduction of peripheral benzodiazepinereceptors expressed in the brains of 1-MT/LPS rel-ative to placebo/LPS treated mice. O’Connor et al.[22] have shown that the presence of 1-MT did notmodify the expression of pro-inflammatory cytokinesin mice upon peripheral injection of LPS. However,direct as well as accumulative immune effects of thevarious neuroactive tryptophan catabolites have to betaken into account. Indeed, Maes and coworkers [19]have shown in vitro that kynurenine and kynurenicacid (KYNA) display anti-inflammatory propertiesvia reduction of stimulated IFN� and TNF� produc-tion. In addition, Doorduin et al. [6] comment thatPBRs are also expressed in activated astrocytes, whichpreferentially produce KYNA [17, 46]. We thereforehypothesize that IDO inhibition in microglia mitigatesthe production of pro-inflammatory QUIN, which incombination with a shortage of anti-inflammatory

914 N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO?
astrocyte-derived KYNA upon LPS challenge, mightleave the brain of 1-MT/LPS mice in a neuroinflam-matory state lower than placebo/LPS mice, but greaterthan the placebo/PBS animals.
We have found significantly higher levels of IDO inthe brainstem of LPS injected mice. It has been shownthat physical stressors, such as trauma or infection,rapidly affect the brainstem, where coordinated pro-cessing of autonomic, immune, and neuroendocrineinformation takes place [47]. Interestingly, injectionof mice with LPS selectively increases immediate-early gene expression within serotonergic neurons ofthe interfascicular part of the dorsal raphe nucleus[48]. These neurons comprise a unique, anatom-ically and functionally distinct immune-responsivesubpopulation within the brainstem Raphe complexthat essentially differs from the neuronal popula-tion responding to anxiety- and stress-related stimuli[49]. This may explain the “paradox” of decreasedbehavioral activity but increased [50] or unchanged[22] serotonergic neurotransmission following acuteimmune stimulation.
In conclusion, our findings indicate that neuroin-flammation induces depressive-like symptoms in ananimal model, which can be abolished by inhibiting theeffects of IDO. Moreover, this study not only providesevidence of a pathogenic role of cerebral inflammationin the precipitation of depression, but also suggeststhat symptoms often concurring with depression, suchas anxiety, feelings of misery, and impaired workingmemory, are the consequences of some other processesand apparently dissociated from brainstem serotoninneurotransmission.
ACKNOWLEDGMENTS
This work was supported by grants from the Inter-national Foundation for Alzheimer Research (ISAO),grant no.06511, the Gratama Foundation and the EU-grant FP6 NeuroproMiSe LSHM-CT-2005-018637.The skillful technical assistance of Dr. J. Doorduin, W.Douwenga, J.N. Keijser, J. Krijnen, Dr. M.F. Masmanand J.W.A. Sijbesma is greatly appreciated.
Authors’ disclosures available online (http://www.j-alz.com/disclosures/view.php?id=1050).
REFERENCES
[1] Streit WJ (2006) Microglial senescence: Does the brain’simmune system have an expiration date? Trends Neurosci 29,506-510.
[2] Dobos N, Korf J, Luiten PG, Eisel UL (2010) Neuroinflam-mation in Alzheimer’s disease and major depression. BiolPsychiatry 67, 503-504.
[3] Kawanokuchi J, Mizuno T, Takeuchi H, Kato H, Wang J,Mitsuma N, Suzumura A (2006) Production of interferon-gby microglia. Multiple Sclerosis 12, 558-564.
[4] Nishioku T, Matsumoto J, Dohgu S, Sumi N, Miyao K, TakataF, Shuto H, Yamauchi A, Kataoka Y (2010) Tumor necro-sis factor-alpha mediates the blood-brain barrier dysfunctioninduced by activated microglia in mouse brain microvascularendothelial cells. J Pharmacol Sci 112, 251-254.
[5] Kakimura JI, Kitamura Y, Takata K, Umeki M, SuzukiS, Shibagaki K, Taniguchi T, Nomura Y, Gebicke-HaerterPJ, Smith MA, Perry G, Shimohama S (2002) Microglialactivation and amyloid-� clearance induced by exogenousheat-shock proteins. FASEB J 16, 601-603.
[6] Doorduin J, de Vries EF, Dierckx RA, Klein HC (2008)PET imaging of the peripheral benzodiazepine receptor:Monitoring disease progression and therapy response in neu-rodegenerative disorders. Curr Pharm Des 14, 3297-3315.
[7] Cagnin A, Kassiou M, Meikle SR, Banati RB (2007) Positronemission tomography imaging of neuroinflammation. Neu-rotherapeutics 4, 443-452.
[8] Heneka MT, O’Banion MK, Terwel D, Kummer MP (2010)Neuroinflammatory processes in Alzheimer’s disease. J Neu-ral Transm 117, 919-947.
[9] Jost BC, Grossberg GT (1996) The evolution of psychiatricsymptoms in Alzheimer’s disease: A natural history study. JAm Geriatr Soc 44, 1078-1081.
[10] Jorm AF (2000) Is depression a risk factor for dementia orcognitive decline? A review. Gerontology 46, 219-227.
[11] Jorm AF (2001) History of depression as a risk factor fordementia: An updated review. Aust N Z J Psychiatry 35, 776-781.
[12] Iwashyna TJ, Ely EW, Smith DM, Langa KM (2010) Long-term cognitive impairment and functional disability amongsurvivors of severe sepsis. JAMA 304, 1787-1794.
[13] Dantzer R, O’Connor JC, Freund GG, Johnson RW, KelleyKW (2008) From inflammation to sickness and depression:When the immune system subjugates the brain. Nat Rev Neu-rosci 9, 46-56.
[14] Smith RS (1991) The macrophage theory of depression. MedHypotheses 35, 298-306.
[15] Miller AH, Maletic V, Raison CL (2009) Inflammation andits discontents: The role of cytokines in the pathophysiologyof major depression. Biol Psychiatry 65, 732-741.
[16] Leonard BE (2007) Inflammation, depression and dementia:Are they connected? Neurochem Res 32, 1749-1756.
[17] Muller N, Schwarz MJ (2007) The immune-mediated alter-ation of serotonin and glutamate: Towards an integrated viewof depression. Mol Psychiatry 12, 988-1000.
[18] Russo S, Kema IP, Bosker F, Haavik J, Korf J (2009) Trypto-phan as an evolutionarily conserved signal to brain serotonin:Molecular evidence and psychiatric implications. World J BiolPsychiatry 10, 258-268.
[19] Maes M, Mihaylova I, Ruyter MD, Kubera M, Bosmans E(2007) The immune effects of TRYCATs (tryptophan catabo-lites along the IDO pathway): Relevance for depression- and other conditions characterized by tryptophan deple-tion induced by inflammation. Neuro Endocrinol Lett 28,826-831.
[20] Guillemin GJ, Brew BJ, Noonan CE, Takikawa O, CullenKM (2005) Indoleamine 2,3dioxygenase and quinolinic acidimmunoreactivity in Alzheimer’s disease hippocampus. Neu-ropathol Appl Neurobiol 31, 395-404.

N. Dobos et al. / Neuroinflammation and Depression: A Role for IDO? 915
[21] Moreau M, Andre C, O’Connor JC, Dumich SA, Woods JA,Kelley KW, Dantzer R, Lestage J, Castanon N (2008) Inocu-lation of Bacillus Calmette-Guerin to mice induces an acuteepisode of sickness behavior followed by chronic depressive-like behavior. Brain Behav Immun 22, 1087-1095.
[22] O’Connor JC, Lawson MA, Andre C, Moreau M,Lestage J, Castanon N, Kelley KW, Dantzer R (2009)Lipopolysaccharide-induced depressive-like behavior ismediated by indoleamine 2,3-dioxygenase activation in mice.Mol Psychiatry 14, 511-522.
[23] Qin L, Wu X, Block ML, Liu Y, Breese GR, Hong JS, KnappDJ, Crews FT (2007) Systemic LPS causes chronic neu-roinflammation and progressive neurodegeneration. Glia 55,453-462.
[24] Andre C, O’Connor JC, Kelley KW, Lestage J, Dantzer R,Castanon N (2008) Spatio-temporal differences in the pro-file of murine brain expression of proinflammatory cytokinesand indoleamine 2,3-dioxygenase in response to peripherallipopolysaccharide administration. J Neuroimmunol 200, 90-99.
[25] Lestage J, Verrier D, Palin K, Dantzer R (2002) The enzymeindoleamine 2,3-dioxygenase is induced in the mouse brainin response to peripheral administration of lipopolysaccharideand superantigen. Brain Behav Immun 16, 596-601.
[26] Keith BJ, Franklin GP (1997) The mouse brain in stereotaxiccoordinates, Academic Press.
[27] Aid S, Langenbach R, Bosetti F (2008) Neuroinflammatoryresponse to lipopolysaccharide is exacerbated in mice genet-ically deficient in cyclooxygenase-2. J Neuroinflammation 5,17.
[28] Aid S, Parikh N, Palumbo S, Bosetti F (2010) Neu-ronal overexpression of cyclooxygenase-2 does not alter theneuroinflammatory response during brain innate immune acti-vation. Neurosci Lett 478, 113-118.
[29] Choi SH, Langenbach R, Bosetti F (2008) Genetic deletionor pharmacological inhibition of cyclooxygenase-1 attenuatelipopolysaccharide-induced inflammatory response and braininjury. FASEB J 22, 1491-1501.
[30] Milatovic D, Zaja-Milatovic S, Montine KS, Shie FS, MontineTJ (2004) Neuronal oxidative damage and denritic degenera-tion following activation of CD14-dependent innate immuneresponse in vivo. J Neuroinflammation 1, 20.
[31] Cremer JE, Hume SP, Cullen BM, Myers R, Manjil LG, Tur-ton DR, Luthra SK, Bateman DM, Pike VW (1992) Thedistribution of radioactivity in brains of rats given [N-methyl-11C]-PK 11195 in vivo after induction of a cortical ischaemiclesion. Int J Rad Appl Instrum B 19, 159-166.
[32] Ostroveanu A, van der Zee EA, Eisel UL, Schmidt M, NijholtIM (2009) Exchange protein activated by cyclic AMP 2(Epac2) plays a specific and time-limited role in memoryretrieval. Hippocampus 20, 1018-1026.
[33] Dolga AM, Granic I, Nijholt IM, Nyakas C, van der Zee EA,Luiten PG, Eisel UL (2009) Pretreatment with lovastatin pre-vents N-methyl-D-aspartate-induced neurodegeneration inthe magnocellular nucleus basalis and behavioral dysfunction.J Alzheimers Dis 17, 327-336.
[34] Cryan JF, Mombereau C (2004) In search of a depressedmouse: Utility of models for studying depression-relatedbehavior in genetically modified mice. Mol Psychiatry 9,326-357.
[35] Ostroveanu A, Van der Zee EA, Dolga AM, Luiten PG, EiselUL, Nijholt IM (2007) A-kinase anchoring protein 150 in themouse brain is concentrated in areas involved in learning andmemory. Brain Res 1145, 97-107.
[36] Mellor AL, Munn DH (2004) IDO expression by dendriticcells: Tolerance and tryptophan catabolism. Nat Rev Immunol4, 762-774.
[37] de Jong WH, Smit R, Bakker SJ, de Vries EG, Kema IP (2009)Plasma tryptophan, kynurenine and 3-hydroxykynureninemeasurement using automated on-line solid-phase extractionHPLC-tandem mass spectrometry. J Chromatogr B AnalytTechnol Biomed Life Sci 877, 603-609.
[38] Hauss-Wegrzyniak B, Dobrzanski P, Stoehr JD, Wenk GL(1998) Chronic neuroinflammation in rats reproduces com-ponents of the neurobiology of Alzheimer’s disease. BrainRes 780, 294-303.
[39] McAfoose J, Baune BT (2009) Evidence for a cytokine modelof cognitive function. Neurosci Biobehav Rev 33, 355-366.
[40] Fu X, Zunich SM, O’Connor JC, Kavelaars A, Dantzer R,Kelley KW (2010) Central administration of lipopolysaccha-ride induces depressive-like behavior in vivo and activatesbrain indoleamine 2,3 dioxygenase in murine organotypichippocampal slice cultures. J Neuroinflammation 7, 43.
[41] Frenois F, Moreau M, O’Connor J, Lawson M, MiconC, Lestage J, Kelley KW, Dantzer R, Castanon N(2007) Lipopolysaccharide induces delayed FosB/DeltaFosBimmunostaining within the mouse extended amygdala, hip-pocampus and hypothalamus, that parallel the expressionof depressive-like behavior. Psychoneuroendocrinology 32,516-531.
[42] Banks WA, Robinson SM (2010) Minimal penetration oflipopolysaccharide across the murine blood-brain barrier.Brain Behav Immun 24, 102-109.
[43] Faggioni R, Fantuzzi G, Villa P, Buurman W, Van Tits LJH,Ghezzi P (1995) Independent down-regulation of central andperipheral tumor necrosis factor production as a result oflipopolysaccharide tolerance in mice. Infect Immun 63, 1473-1477.
[44] Galea I, Bechmann I, Perry VH (2007) What is immune priv-ilege (not)? Trends Immunol 28, 12-18.
[45] O’Connor JC, Lawson MA, Andre C, Briley EM, SzegediSS, Lestage J, Castanon N, Herkenham M, Dantzer R, KelleyKW (2009) Induction of IDO by bacille Calmette-Guerin isresponsible for development of murine depressive-like behav-ior. J Immunol 182, 3202-3212.
[46] Guillemin GJ, Smith DG, Kerr SJ, Smythe GA, Kapoor V,Armati PJ, Brew BJ (2000) Characterisation of kynureninepathway metabolism in human astrocytes and implications inneuropathogenesis. Redox Report 5, 108-111.
[47] Ulrich-Lai YM, Herman JP (2009) Neural regulation ofendocrine and autonomic stress responses. Nat Rev Neurosci10, 397-409.
[48] Hollis JH, Evans AK, Bruce KP, Lightman SL, LowryCA (2006) Lipopolysaccharide has indomethacin-sensitiveactions on Fos expression in topographically organized sub-populations of serotonergic neurons. Brain Behav Immun 20,569-577.
[49] Lowry CA, Hollis JH, de Vries A, Pan B, Brunet LR, Hunt JR,Paton JF, van Kampen E, Knight DM, Evans AK, Rook GA,Lightman SL (2007) Identification of an immune-responsivemesolimbocortical serotonergic system: Potential role in reg-ulation of emotional behavior. Neuroscience 146, 756-772.
[50] Linthorst AC, Flachskamm C, Muller-Preuss P, Holsboer F,Reul JM (1995) Effect of bacterial endotoxin and interleukin-1 beta on hippocampal serotonergic neurotransmission,behavioral activity, and free corticosterone levels: An in vivomicrodialysis study. J Neurosci 15, 2920-2934.




















