
Gonadectomy inhibits development of experimental amoebic liver abscess in hamsters through...
10
Gonadectomy inhibits development of experimental amoebic liver abscess in hamsters through downregulation of the inflammatory immune response C. CERVANTES-REBOLLEDO, 1 N. MORENO-MENDOZA, 2 J. MORALES-MONTOR, 1 P. DE LA TORRE, 1 J. P. LACLETTE 1 & J. C. CARRERO 1 1 Department of Immunology, Instituto de Investigaciones BiomȖdicas, Universidad Nacional AutɃnoma de MȖxico, MȖxico D.F., MȖxico; and 2 Department of Cellular Biology and Physiology, Instituto de Investigaciones BiomȖdicas, Universidad Nacional AutɃnoma de MȖxico, MȖxico D.F., MȖxico SUMMARY Incidence of amoebic liver abscess (ALA) in human males is considerably higher than in females, suggesting a role for sex hormones in this parasite infection. We describe here the effect of hamster gonadectomization on the development of ALA. After monitoring the decrease of oestradiol in females and testosterone in males to undetectable levels by ELISA and Radio Immuno Assay (RIA) in serum, hamsters were intraportally infected with Entamoeba histolytica trophozo- ites and killed 7 days later. ALA was absent in 50% of male and 15% of female gonadectomized (Gdx) hamsters, in comparison with 100% infection in non-Gdx controls. This protection against ALA in Gdx hamsters was concomitant to a comparatively scarce inflammatory infiltrate and necro- sis surrounding clusters of trophozoites in the liver tissue, as well as to a lack of response of spleen cells to Con A, evalu- ated in proliferation assays. As tissue damage in ALA has been associated with a local inflammatory Th1 response, we determined the profile of response in hamsters by immuno- histochemistry on liver sections. In contrast to strong Th1 responses in non-Gdx animals, Gdx females and males exhibited Th2 and Th3 profiles of cytokines, respectively, suggesting that protection against ALA following gonadec- tomization, could be related to downregulation of liver Th1 response during amoebic infection. Keywords amoebic liver abscess, cellular immune response, goanadectomy, protection, sex hormones INTRODUCTION Evidence is accumulating to support the notion that prev- alence and intensity of parasitic infections are higher in males than in females (1–4). The interactions between the nervous, endocrine and immune systems are gaining atten- tion as determinants for the control of parasitic infections. The hypothalamic-pituitary-gonadal axis appears to be involved in these regulations through the direct action of hormones on parasite growth as well as their effect on the host immune response (1,2). Androgens in males and oes- trogens in females, have received a special consideration for their immunomodulatory properties. Androgens may decrease and oestrogens may enhance the outcome of the immune response, possibly by modulating the expression of specific genes (3–5) through the binding of sex hor- mones to functional steroid hormone receptors expressed in innate and adaptive immune cells (6–8). The triggering of steroid receptors in immune cells by hormones induces the activation of signalling pathways, including that of the nuclear factor-jb, resulting in the expression of diverse cytokines and chemokines that recruits more immune cells to the site of infection and helps to shape the type of immune response, Th1 or Th2 (8,9). Plasmodium spp. and Trypanosoma cruzi are good exam- ples about the influence of sex hormones on the suscepti- bility to parasitic infections. Different vertebrate hosts infected with Plasmodium show a pronounced sexual dimorphism (SD) (10), translated in higher parasitaemia and mortality in males than in females, suggesting the involvement of sexual hormones and immunological differ- ences between sexes as responsible for such outcomes (11,12). Moreover, male castration of mice reduces the parasitaemia and mortality caused by Plasmodium and Trypanosoma (13–16), whereas gonadectomy of females Correspondence: Julio C. Carrero, Departamento de Inmunologȷa, Instituto de Investigaciones BiomȖdicas, UNAM, A.P. 70228, 04510 MȖxico D.F., MȖxico (e-mail: [email protected]). Disclosures: None Received: 19 January 2009 Accepted for publication: 26 March 2009 Parasite Immunology, 2009, 31, 447–456 DOI: 10.1111/j.1365-3024.2009.01120.x ȑ 2009 Blackwell Publishing Ltd 447
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Gonadectomy inhibits development of experimental amoebic liver abscess in hamsters through...

Gonadectomy inhibits development of experimental amoebic liver
abscess in hamsters through downregulation of the inflammatory
immune response
C. CERVANTES-REBOLLEDO,1 N. MORENO-MENDOZA,2 J. MORALES-MONTOR,1 P. DE LA TORRE,1 J. P. LACLETTE1 &J. C. CARRERO1
1Department of Immunology, Instituto de Investigaciones Biom�dicas, Universidad Nacional Aut�noma de M�xico, M�xico D.F., M�xico;and 2Department of Cellular Biology and Physiology, Instituto de Investigaciones Biom�dicas, Universidad Nacional Aut�noma de M�xico,M�xico D.F., M�xico
SUMMARY
Incidence of amoebic liver abscess (ALA) in human malesis considerably higher than in females, suggesting a role forsex hormones in this parasite infection. We describe here theeffect of hamster gonadectomization on the development ofALA. After monitoring the decrease of oestradiol in femalesand testosterone in males to undetectable levels by ELISAand Radio Immuno Assay (RIA) in serum, hamsters wereintraportally infected with Entamoeba histolytica trophozo-ites and killed 7 days later. ALA was absent in 50% of maleand 15% of female gonadectomized (Gdx) hamsters, incomparison with 100% infection in non-Gdx controls. Thisprotection against ALA in Gdx hamsters was concomitantto a comparatively scarce inflammatory infiltrate and necro-sis surrounding clusters of trophozoites in the liver tissue, aswell as to a lack of response of spleen cells to Con A, evalu-ated in proliferation assays. As tissue damage in ALA hasbeen associated with a local inflammatory Th1 response, wedetermined the profile of response in hamsters by immuno-histochemistry on liver sections. In contrast to strong Th1responses in non-Gdx animals, Gdx females and malesexhibited Th2 and Th3 profiles of cytokines, respectively,suggesting that protection against ALA following gonadec-tomization, could be related to downregulation of liver Th1response during amoebic infection.
Keywords amoebic liver abscess, cellular immune response,goanadectomy, protection, sex hormones
INTRODUCTION
Evidence is accumulating to support the notion that prev-alence and intensity of parasitic infections are higher inmales than in females (1–4). The interactions between thenervous, endocrine and immune systems are gaining atten-tion as determinants for the control of parasitic infections.The hypothalamic-pituitary-gonadal axis appears to beinvolved in these regulations through the direct action ofhormones on parasite growth as well as their effect on thehost immune response (1,2). Androgens in males and oes-trogens in females, have received a special considerationfor their immunomodulatory properties. Androgens maydecrease and oestrogens may enhance the outcome of theimmune response, possibly by modulating the expressionof specific genes (3–5) through the binding of sex hor-mones to functional steroid hormone receptors expressedin innate and adaptive immune cells (6–8). The triggeringof steroid receptors in immune cells by hormones inducesthe activation of signalling pathways, including that of thenuclear factor-jb, resulting in the expression of diversecytokines and chemokines that recruits more immune cellsto the site of infection and helps to shape the type ofimmune response, Th1 or Th2 (8,9).
Plasmodium spp. and Trypanosoma cruzi are good exam-ples about the influence of sex hormones on the suscepti-bility to parasitic infections. Different vertebrate hostsinfected with Plasmodium show a pronounced sexualdimorphism (SD) (10), translated in higher parasitaemiaand mortality in males than in females, suggesting theinvolvement of sexual hormones and immunological differ-ences between sexes as responsible for such outcomes(11,12). Moreover, male castration of mice reduces theparasitaemia and mortality caused by Plasmodium andTrypanosoma (13–16), whereas gonadectomy of females
Correspondence: Julio C. Carrero, Departamento de Inmunolog�a,Instituto de Investigaciones Biom�dicas, UNAM, A.P. 70228,04510 M�xico D.F., M�xico (e-mail: [email protected]).Disclosures: NoneReceived: 19 January 2009Accepted for publication: 26 March 2009
Parasite Immunology, 2009, 31, 447–456 DOI: 10.1111/j.1365-3024.2009.01120.x
� 2009 Blackwell Publishing Ltd 447

increased the parasitaemia in mice infected with T. cruzi(16,17). Even though there is no clear evidence of SD inhumans, indirect observations suggest a lower severity andfrequency of illness by T. cruzi in women (18,19).
Amoebiasis caused by Entamoeba histolytica remains asone of the leading parasite diseases worldwide, responsiblefor about 100,000 deaths throughout the world (20). Thisparasite invades the intestinal mucosa causing amoebiccolitis and ulcers. It may also spread to other organs,mainly the liver, resulting in amoebic liver abscess (ALA)(20). Although information is scarce, epidemiological datahas pointed out that sex hormones could play a role onthe development of E. histolytica human infections, mainlyALA. Thus, is well documented in adults that dependingon the studied area and age, ALA is five to sevenfoldsmore prevalent in men than women (21,22). This differ-ence is less clear in the case of intestinal amoebiasis; how-ever, symptomatic intestinal amoebiasis in adults appears,in contrast to ALA, to be more common in women thanmen (20). These variations in the susceptibility associatedto sex suggest that sex hormones could be involved in thedeveloping of amoebic infection.
Noteworthy, in spite of such SD in human incidenceof extraintestinal amoebiasis, very few studies havefocused in determine the role of sex hormones on ani-mal models of E. histolytica infection. An early reportproposed that oestrogens confer protection against devel-opment of ALA in hamsters (23). The only reportedeffect for progesterone, deals with the positive effect ofthis hormone in the migration of trophozoites from theintestine to the liver in gerbils (24). More recently, dif-ferences within genders in the control of ALA wasreported in C57BL ⁄ 6 mice. In contrast to males, femalesrapidly cleared the parasites, recruited higher numbersof natural killer T cells (NKTCs) to the infection site,and produced higher levels of interferon-c (INF-c), sug-gesting a SD (22). We have previously reported aninhibitory action of dehydroepiandrosterone (DHEA)and stimulatory effect of cortisol on the in vitro prolifer-ation of E. histolytica trophozoites. However, sex hor-mones such as progesterone, oestradiol and testosteronedid not show effect on the in vitro trophozoite¢s prolifer-ation (25). According with the experimental evidencepresented above that supports that host sex is relevantduring amoebiasis, and due to the fact that mammalianhosts’ SD in amoebiasis it is a scarcely and superficiallystudied biological phenomenon of considerable signifi-cance for individual health, behaviour and lifestyles ofseveral mammalian species, the object of the presentstudy was to explore the possible SD in the a experi-mental model of amoebiasis in which this has not beenexplored before (hamster) and to look to important
aspects of the immune response in vivo and in vitro. Wehypothesized that differences between sexes in hamsterscould not really be detected due to the over-activationof the Th1 immune response that mask differences asso-ciated to sexual hormones, leaving open the possibilityof detect differences upon gonadectomy, as resulted inour experiments.
Our findings, shows that a sexual hormone deficiencyinduced by gonadectomy, reduced the immune responseand damage caused by E. histolytica in hamster liver, sug-gesting that immune and endocrine system interactionsplay a fundamental role in the establishment, developmentand outcome of ALA.
MATERIALS AND METHODS
Parasites
Axenic HM1 : IMSS E. histolytica trophozoites weremaintained in TYI-S-33 medium supplemented with 15%adult bovine serum (Biofluids), 3% Diamond¢s vitaminmix (JRH, Biosciences, San Jos�, CA, USA), 100 U ⁄ mLpenicillin and 100 lg ⁄ mL streptomycin sulphate at 37�Cunder anaerobic conditions.
Animals
Male and female golden Syrian hamsters (Mesocricetusauratus) of 6–8 week old and about 100 g were used. Theywere kept in plastic cages, with water and food being pro-vided ad libitum. Animals were divided into five groups;two noninfected groups: intact and gonadectomized(Gdx); and three infected groups: infected, Gdx + infectedand sham (surgery treatment without removal of gonadsfollowed by liver infection). The number of animals forgroup was as follows: intact and Gdx, 20 each (10 femalesand 10 males); infected and Gdx + infected, 80 each (40females and 40 males); sham, 28 each (14 females and 14males).
Gonadectomy
The animals were anaesthetized with an intraperitonelinjection of anesthesal (60 mg ⁄ kg) and testes or ovarieswere bilaterally removed under sterilized conditions. TheGdx and sham hamsters were maintained for 2 weeksbefore used for further experiments.
Determination of testosterone and oestradiol
Blood samples were obtained from each animal on threeoccasions. In the Gdx group, at the beginning of the
C. Cervantes-Rebolledo et al. Parasite Immunology
448 � 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456

experiment (before gonadectomy), before the infection(15 days after gonadectomy) and at killing (22 days aftergonadectomy). In the other groups, blood samples werealso collected at the same days during the running experi-ment. Samples (0Æ5 mL) were obtained from the retro-orbital sinus under anaesthesia and the serum stored at)20�C until used. Serum testosterone levels were deter-mined using a testosterone enzyme immunoassay (EIA)kit (Diagnostic System Laboratories, Inc., Webster, Texas,USA) and oestradiol levels using an enzyme-linked fluo-rescent assay (Biomerieux, Lyon, France).
Infection with parasites
Hamsters were infected for induction of ALA as describedelsewhere. In brief, the animals were anaesthetized withanesthesal (60 mg ⁄ kg) and the portal vein was exposed bylaparatomy under aseptic conditions. Trophozoites(5 · 105) washed three times in PBS and resuspended in100 lL PBS were directly inoculated into the portal veinusing a tuberculin syringe, followed by the immediateapplication of a gel-foam pad at the site of inoculation inorder to avoid bleeding and loss of amoebas. Hamsterswere sacrificed after 7 days and the livers removed andweighed. Abscess were exscinded from the liver tissue andalso weighed. Liver samples containing abscesses were alsofixed in 4% paraformaldehyde in PBS for 1 h and thenstored in 30% sucrose for histological analysis.
Splenocyte proliferation
Spleens were removed and splenocyte cultures were carriedout in polystyrene 96-well flat bottom plates using 5 · 105
cells ⁄well in RPMI medium supplemented with 10% bovinefoetal serum, with or without the addition of 1 lg ⁄ well ConA. Cultures were incubated for 48 h in wet chambers at37�C in an atmosphere containing 5% CO2 and subse-quently, tritiated thymidine was added (1 lCi ⁄ well in 20 lLof medium). After an additional 24 h of incubation, cellswere filtered in special glass filters, and collected in a cellharvester. The amount of radioactivity incorporated in thecellular DNA was assessed by using scintillation liquid anda b-plate counter. Each test was carried out in triplicate.
Histological studies
Liver tissue with abscesses were embedded in paraffin andprocessed for histology by standard techniques. In brief,serial sections of 20 lm thicknesses were obtained in amicrotome, placed on slides coated with poly-L-lysine(Sigma, St Louis, MO, USA), deparaffined, and finally
stained with haematoxylin and eosin for light microscopyanalysis.
Immunohistochemistry
Antibodies anti-mouse IL2, IL1b, IL5 and TGF-b done inrabbit, anti-mouse INF-c, TNFa, IL12, IL10, IL6 andIL13 done in goat, and anti-mouse IL4 done in rat wereused as primary antibodies (Santa Cruz Biotechnology,Santa Cruz, CA, USA). Anti-rabbit IgG and anti-goatIgG conjugated with tetramethyl rhodamine iso-thiocya-nate (TRITC) and anti-rat IgG conjugated with flouresce-in isothiocyanate (ZIMED Laboratories Inc., SanFrancisco, CA, USA) were used as secondary antibodies.ALA-containing liver lobes were fixed in 4% paraformal-dehyde for 1 h, washed with PBS and stored in PBS con-taining 30% sucrose at 4�C. The next day, the sampleswere embedded in tissue-freezing medium (Leica, Nuss-loch, Germany) and frozen at )70�C (dry ice hexane).Serial sections of 20 lm thicknesses were obtained using acryomicrotome, placed on slides coated with poly-L-lysine(Sigma) and air dried. Sections were treated with 1% Tri-ton X-100 after blocking with 1% bovine serum albumin(BSA) and incubated with primary antibody overnight at4�C diluted in 1 : 1000 BSA. After rinsing in PBS, the sec-tions were incubated with the secondary antibody during1 h at room temperature diluted in 1 : 500 BSA, washedin PBS and embedded in DAKO fluorescent mountingmedium (DAKO, Carpinter�a, CA, USA). Liver sectionsprocessed without the primary antibody were used asnegative controls.
For each group, several field images of ALA were cap-tured and analysed by semi-quantitative inmunofluores-cence laser confocal microscopy. In brief, TIFF imageswere acquired with the TCS-SP1 software and importedinto Image Pro Plus (Media Cybernetics, Bethesda, MD,USA) for subsequent measuring of the immunofluores-cence intensity. In each image, representative areas wereselected and, with the exposure times kept constant, theintensity of fluorescence was quantified and expressed asthe mean pixel intensity for that region. For each animal,at least six randomly selected areas were analysed. Datawas statistically analysed and values were expressed asmean € SE.
Statistical analysis
Statistical significance between groups was determined byANOVA followed by post hoc Tukey test. The results wereexpressed as the mean € standard deviation of mean. Dif-ferences were considered significant from a P < 0Æ05.
Volume 31, Number 8, August 2009 Sex hormones deficiency and amoebic liver abscess
� 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456 449

RESULTS
Amoebic liver infection by itself induces changes in thelevel of sex hormones in hamsters
Previous to the infection experiments, levels of oestradiolin females and testosterone in males were determinedbefore and after gonadectomization. Results showed phys-iological levels of both hormones for hamsters of 4- to 6-week old, in all animals previous to gonadectomy. Fifteendays after gonadectomy and before the infection withtrophozoites, the serum concentration of oestradiol infemales and testosterone in males, decreased from14Æ2 € 0Æ7 pg ⁄ mL and 1Æ2 € 0Æ2 ng ⁄ mL, respectively, toundetectable levels, indicating that castration effectivelyresulted in a marked reduction in the novo synthesis ofsex hormones. Once confirmed the deficiency of hor-mones, hamsters of all groups excluding the intact andGdx groups, were intraportally infected with virulenttrophozoites of E. histolytica. By the time of killing(7 days post-infection), concentration of both hormonesin the Gdx animals (oestrogens in female and testosteronein males) continued undetectable (not shown). Interest-ingly, hormone determinations in the infected groupshowed that infection by itself affects the level of oestro-gens and testosterone in hamsters. Blood concentrationsof both hormones in females diminished significantlyupon infection (oestradiol from 15 € 1Æ6 pg ⁄ mL to 11Æ7 €0Æ5 pg ⁄ mL and testosterone from 0Æ07 € 0Æ01 ng ⁄ mL to0Æ03 € 0Æ01 ng ⁄ mL; P < 0Æ05 for both hormones). In con-trast, liver infection of male hamsters resulted in a bloodincrease of oestradiol (9Æ6 € 0Æ4 ng ⁄ mL before infection to12Æ7 € 0Æ5 5 ng ⁄ mL at 7 days post-infection; P < 0Æ05),whereas testosterone diminished drastically (from1Æ2 € 0Æ3 ng ⁄ mL to not detectable levels; P < 0Æ05). In theuninfected groups, the hormone levels remained constantthroughout the experiment (data not shown).
Hormone deficiency inhibits the development of ALA
Induction of hormonal deficiencies in hamsters by gonadec-tomy, resulted in a marked reduction in the number and sizeof liver abscesses evaluated 7 days post-infection (Table 1).Thus, 19 out of 40 Gdx males and 34 out of 40 Gdx femalehamsters developed any ALA, in comparison with 40 out of40 females and 40 out 40 males infected hamsters in theinfected group (Table 1), and 14 out of 14 females and 14out 14 males infected animals in the sham group (notshown). Moreover, significant statistical differences werealso observed in the weights of complete livers and abscessesbetween the Gdx animals that developed any ALA and theanimals of the infected group (Table 1). Differences betweensexes were also observed in the Gdx + infected group butnot in the infected group. Thus, whereas the number andsize of abscesses in livers was similar between sexes withinthe infected group hamsters (82Æ8% of the liver in males and80% of the liver in females with damage), the extent of dam-age in the Gdx + infected group was lower in males than infemales (32Æ8% and 45Æ3% respectively Table 1). Analysis ofhistological sections from liver abscesses of the infectedgroup animals showed extensive necrotic damage and tissueliquefaction associated with an intensive inflammatoryinfiltrate, mainly represented by neutrophils and lympho-cytes, throughout all the liver lobes. Scarce trophozoitesdistributed along the necrotic liver tissue were alsoobserved (Figure 1a, b). In contrast, focal inflammatoryinfiltrates surrounding clusters of trophozoites wereobserved in tissue sections from ALA in the Gdx +infected group hamsters. Necrosis was scarce and alsofocal, only detected beneath the cluster of trophozoitesand in the interphase between such clusters and the sur-rounding inflammatory focus (Figure 1c, d). This focalinflammation was always smaller in the ALA from Gdxmales in spite of being surrounding cluster of 20 or moretrophozoites, in comparison with Gdx females where the
Table 1 Comparison of the liver and abscess weights between not treated and gonadectomized hamsters
Group
ALA
Livers
Liver weight (g)a Abscess weight (g)aRatio ALA ⁄liver · 100
M F M F M F M F
Normal control NA NA 4Æ5 € 0Æ5 5 € 0Æ4 NA NA NA NAGonadectomized control NA NA 4 € 0Æ3 4Æ1 € 0Æ3 NA NA NA NAInfected control 40 ⁄ 40 40 ⁄ 40 14 € 2Æ3 11Æ1 € 2Æ5 11Æ6 € 2Æ7 8Æ9 € 1Æ6 82Æ8% 80%Gonadectomized and infected 19 ⁄ 40 34 ⁄ 40 7 € 1Æ2* 8Æ6 € 1Æ6* 2Æ3 € 1Æ8* 3Æ9 € 1Æ3* 32Æ8% 45Æ3%
aData are presented as mean € standard deviation. *Statistical significant differences (P < 0Æ05) are observed in the weight of the liversand abscess of the gonadectomized and infected animals respect to the infected controls (without gonadectomy). F, female; M, male; NA,not applicable; ALA, amoebic liver abscess.
C. Cervantes-Rebolledo et al. Parasite Immunology
450 � 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456
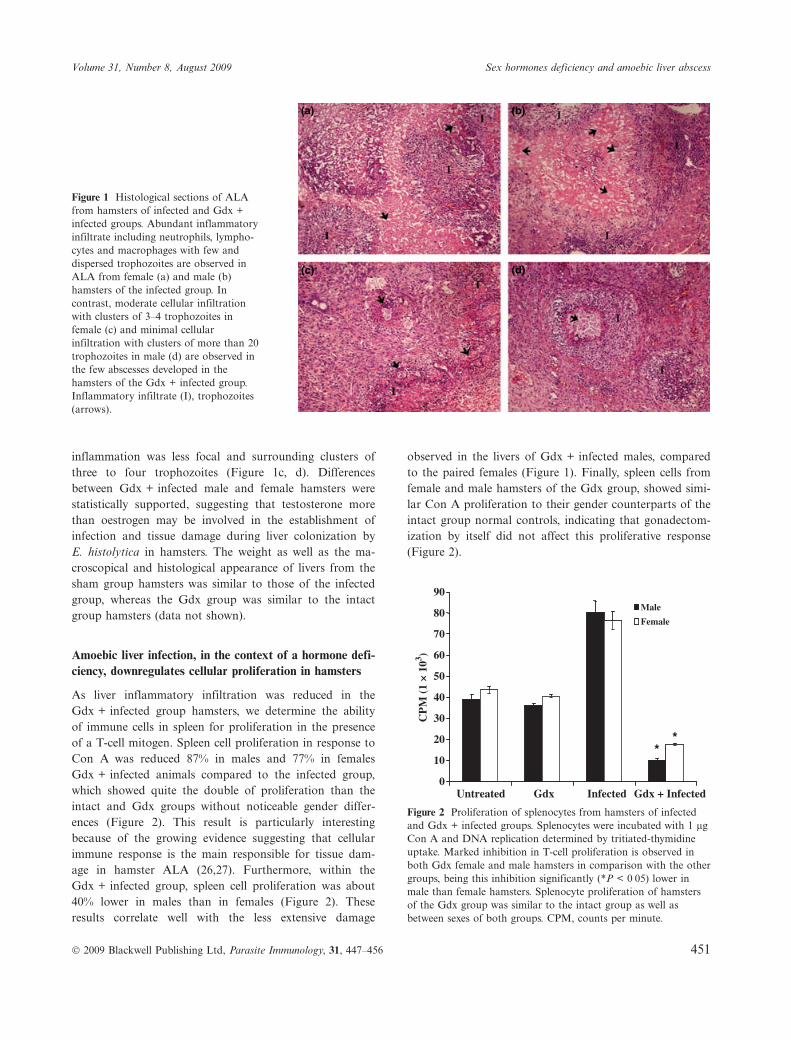
inflammation was less focal and surrounding clusters ofthree to four trophozoites (Figure 1c, d). Differencesbetween Gdx + infected male and female hamsters werestatistically supported, suggesting that testosterone morethan oestrogen may be involved in the establishment ofinfection and tissue damage during liver colonization byE. histolytica in hamsters. The weight as well as the ma-croscopical and histological appearance of livers from thesham group hamsters was similar to those of the infectedgroup, whereas the Gdx group was similar to the intactgroup hamsters (data not shown).
Amoebic liver infection, in the context of a hormone defi-ciency, downregulates cellular proliferation in hamsters
As liver inflammatory infiltration was reduced in theGdx + infected group hamsters, we determine the abilityof immune cells in spleen for proliferation in the presenceof a T-cell mitogen. Spleen cell proliferation in response toCon A was reduced 87% in males and 77% in femalesGdx + infected animals compared to the infected group,which showed quite the double of proliferation than theintact and Gdx groups without noticeable gender differ-ences (Figure 2). This result is particularly interestingbecause of the growing evidence suggesting that cellularimmune response is the main responsible for tissue dam-age in hamster ALA (26,27). Furthermore, within theGdx + infected group, spleen cell proliferation was about40% lower in males than in females (Figure 2). Theseresults correlate well with the less extensive damage
observed in the livers of Gdx + infected males, comparedto the paired females (Figure 1). Finally, spleen cells fromfemale and male hamsters of the Gdx group, showed simi-lar Con A proliferation to their gender counterparts of theintact group normal controls, indicating that gonadectom-ization by itself did not affect this proliferative response(Figure 2).
(a) (b)
(d)(c)
Figure 1 Histological sections of ALAfrom hamsters of infected and Gdx +infected groups. Abundant inflammatoryinfiltrate including neutrophils, lympho-cytes and macrophages with few anddispersed trophozoites are observed inALA from female (a) and male (b)hamsters of the infected group. Incontrast, moderate cellular infiltrationwith clusters of 3–4 trophozoites infemale (c) and minimal cellularinfiltration with clusters of more than 20trophozoites in male (d) are observed inthe few abscesses developed in thehamsters of the Gdx + infected group.Inflammatory infiltrate (I), trophozoites(arrows).
0
10
20
30
40
50
60
70
80
90
Untreated Gdx Infected Gdx + Infected
Male
Female
**
CP
M (
1 ×
103 )
Figure 2 Proliferation of splenocytes from hamsters of infectedand Gdx + infected groups. Splenocytes were incubated with 1 lgCon A and DNA replication determined by tritiated-thymidineuptake. Marked inhibition in T-cell proliferation is observed inboth Gdx female and male hamsters in comparison with the othergroups, being this inhibition significantly (*P < 0Æ05) lower inmale than female hamsters. Splenocyte proliferation of hamstersof the Gdx group was similar to the intact group as well asbetween sexes of both groups. CPM, counts per minute.
Volume 31, Number 8, August 2009 Sex hormones deficiency and amoebic liver abscess
� 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456 451

Hormone deficiency modulates, during infection, thecytokine expression in liver from a Th1 profile to a Th2and Treg profiles
As tissue damage in hamster ALA has been related to alocal Th1 inflammatory profile, we carried out immuno-histochemical assays in tissue sections from ALA, in orderto determine and compare the local profile of Th responsein each group tested. The following Th1 inflammatorycytokines (IL-1b, IL-2, IL-12, INF-c and TNF-a), Th2anti-inflammatory cytokines (IL-5, IL-13, IL-4 and IL-6),and regulatory cytokines (IL-10 and TGF-b), were deter-mined in hamsters of the intact, infected, Gdx andGdx + infected groups. On semi-quantitative inmunofluo-rescence laser confocal microscopy, the mean intensity ofall tested Th1 cytokines was dramatically lower or absentin ALA sections from Gdx + infected males and females,in contrast to the high level of expression of these cyto-kines in the infected hamsters of both sexes (Table 2). Thisobservation suggested that hormone deficiency in hamstersmodified the inflammatory environment, previously pro-posed as responsible for liver lesions during hepatic infec-tion by E. histolytica (26,27). Noteworthy, differencesbetween sexes for the expression of Th1 cytokines were
still clear in the Gdx + infected group animals. Thus, incontrast to the females where low but detectable expres-sion of all Th1 cytokines tested was observed (Table 2),males showed null expression of inflammatory cytokinesIL-1b, INF-c and TNF-a, in combination with a statisti-cally significant decrease in the expression of IL-2 andIL-12 when compare with the expression in females(Table 2). Moreover, the level of expression of Th1 cyto-kines in those males was similar to that determined in theintact as well as Gdx group hamsters (basal expression ofIL-2 and IL-12 and null expression of the other cytokinesin both sexes; data not shown). From these results, it isconceivable to think that the lower production of Th1inflammatory cytokines and the decreased inflammatoryinfiltrate in males (compared to females), is associatedwith the scarce or complete absence of damage in most ofthe Gdx + infected male hamsters (Table 2).
A significant decrease in the fluorescence intensity ofTh2 cytokines (with the exception of IL-4) was alsoobserved in the ALA of Gdx + infected group hamsters,compared to the ALA of animals from the infected groupthat showed a Th2 cytokines expression as high as Th1,with a higher fluorescence intensity for IL-5 and IL-13 inmales (Table 2). In addition, differences between sexes inthe Gdx + infected group were also found: lower produc-tion of IL-13, IL-5 and IL-6 in the livers of malescompared to females. In the intact and Gdx groups, Th2cytokines were undetectable in the livers with no differ-ences between sexes (data not shown).
Noteworthy, the expression of regulatory cytokines IL-10 and TGF-b was high in the ALA from Gdx + infectedmales compared to a partial expression in both sexes ofthe infected group and very low expression in theGdx + infected females. In addition, low levels of IL-10(8Æ0 € 1Æ1) were detected in males of the infected groupwhen compared with the females of the same group(32Æ6 € 6Æ1) (Table 2). Probably, the increased expressionof these regulatory cytokines in Gdx + infected males pre-vents the exacerbation of the inflammatory immuneresponse needed for ALA development, resulting in smal-ler and fewer abscesses (Table 1). No signal for Treg cyto-kines was observed in both sexes of the intact and Gdxgroups (data not shown).
DISCUSSION
We show here that induction of sex hormone deficiency bygonadectomy of hamsters resulted in the inhibition ofALA development or in the reduction of the number andsize of abscesses. This is in agreement with a number ofobservations in other parasitic infections showing thatinduction of hormonal deficiencies by gonadectomy,
Table 2 Cytokine expression in livers from female and malegonadectomized and infected hamsters
Cytokines
Groups
Infected controlaGonadectomized andinfecteda
M F M F
Th1IL-1b 46Æ5 € 4Æ1 46Æ2 € 2Æ7 0* 14Æ9 € 1Æ1*IL-2 56Æ2 € 4Æ2 54Æ6 € 1Æ1 0Æ3 € 0Æ2* 3Æ9 € 1Æ7*INF-c 46Æ6 € 4Æ1 52Æ1 € 3Æ3 0* 2Æ3 € 1Æ1*IL-12 53Æ4 € 3Æ2 52Æ5 € 3Æ8 0Æ2 € 0Æ4* 9Æ4 € 2Æ3*TNF-a 55Æ5 € 1Æ8 42Æ6 € 2Æ8 0* 5Æ6 € 1Æ7*
Th2IL-5 43Æ7 € 2Æ1 32Æ8 € 3Æ3 0Æ5 € 0Æ2* 9Æ0 € 1Æ2*IL-13 44Æ0 € 3Æ6 25Æ0 € 3Æ0 0Æ6 € 0Æ2* 7Æ8 € 1Æ5*IL-6 28Æ4 € 1Æ2 25Æ7 € 0Æ8 7Æ3 € 1Æ3* 13Æ4 € 1Æ1*IL-4 50Æ5 € 0Æ6 49Æ8 € 3Æ3 50Æ0 € 0Æ7 40Æ0 € 2Æ1*
TregIL-10 8Æ0 € 1Æ1 32Æ6 € 6Æ1 58Æ5 € 3Æ0* 5Æ0 € 2Æ0*TGF-b 30Æ0 € 3Æ0 34Æ1 € 1Æ5 66Æ6 € 3Æ1* 7Æ0 € 1Æ6*
aFluorescence intensity data are presented as mean € standarddeviation. *Statistical significant differences (P < 0Æ05) areobserved in the fluorescence intensity of Th1, Th2, and regulatorycytokines of livers from male and female gonadectomized andinfected hamsters respect to their gender counterpart infectedwithout gonadectomy. F, female; M, male.
C. Cervantes-Rebolledo et al. Parasite Immunology
452 � 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456

influence the susceptibility of various hosts. For example,gonadectomy of male mice decreases the parasitaemia ofP. chabaubi, P. berghei, Leishmania major and T. cruzi,whereas gonadectomy of females, in most cases, leads toan increase in the parasitaemia (13–17,28,29). In the caseof T. cruzi, several animal models have shown higher levelsof parasitaemia, more extensive tissue invasion, greaterweight loss, and shorter survival rates in males thanfemales (15,16). Such difference between sexes disappearedin Gdx females mice resulting in an T. cruzi increasedparasitaemia (15,16). In addition, treatment of mice withoestradiol and progesterone decreases the number of bloodtrypomastigotes to the levels observed in control animals(17). Those results demonstrated that female gonadal hor-mones, especially oestrogen, play a fundamental role inthe resistance to T. cruzi. In contrast, Gdx males showedlower parasitaemia compared to control animals, and hor-monal replacement with testosterone restored parasitaemiato control levels, confirming the influence of malehormones in susceptibility to this infection (15).
Our results on hamster ALA are in agreement withthose showing testosterone as involved in susceptibility toparasites infections. The half of Gdx + infected males didnot develop any amoebic abscesses and the ALA progres-sion in the other half was considerable reduced, suggestingthat testosterone is a host factor favouring the develop-ment of ALA. Interestingly, we also observed a decrease,less marked that Gdx male hamsters, in the incidence orsize of ALA in Gdx + infected female hamsters, in con-trast with many models showing increased parasitaemiasand symptomatology in Gdx females. Therefore, at least inthe ALA hamster model, both hormones, but testosteronein a higher grade, appear to be involved in the establish-ment and development of the liver lesions by E. histo-lytica. This experimental observation appears grossly tocorrelate well with epidemiological data showing a higherincidence (up to sevenfold) of ALA in men than in women(21,22). However, experiments of hormone replacementfollowing gonadectomy are needed before a mechanisticcorrelation is established (currently being carried out inour laboratory). In the meantime, proposals that a malehigher exposure to infection could be responsible for thisgender difference, cannot ruled out. However, is importantto mention that the incidence of ALA still remains veryhigh in males, even though the exposition of females tosources of amoebic infection has increased rapidly.
It is interesting that differences between sexes in ALAwere only registered when the animals were Gdx. As thismodel has been described as highly susceptible to liverinfection by amoeba, it is tempting to speculate that differ-ences in normal animals are not detected due to an exac-erbate inflammatory response that overcome any effect
due to the sex hormones. In this sense, growing evidenceindicates that sex hormones modulate the function of thehost immune system and thereby, influence the susceptibil-ity or resistance to parasitic infections (3–5,30). Femalesappear to develop more robust and potentially protectivehumoral and cellular immune responses to antigenic chal-lenges than their male counterparts. In contrast, andro-gens in males have frequently been related to moreaggressive and damaging immune responses to microbialstimuli (31). As both sex hormones have the capacity toaffect the cellular immune responses, the hormone defi-ciency in Gdx hamsters downregulated this response as weobserved in Figure 2. Such immunosuppressive stage couldbe responsible (or contribute) for the observed decrease inliver inflammation, and in turn, in the reduction of theliver injuries, and the inhibition of ALAs development inthe Gdx hamsters. This is in agreement with recent reportsshowing that the host cellular response appears to fosterthe establishment and development of ALA on hamsters,instead of playing a protective role (26,27).
Histological sections from the ALA of Gdx + infectedmales revealed a high number of aggregated trophozoitesforming circles with a necrotic core, and surrounded byfibrotic tissue and local inflammatory infiltrate. A similararrangement of the trophozoites was observed when ham-sters were treated with cyclosporine A, a T-cell immuno-suppressive agent that also inhibits ALA (27). Therefore,deficiency of androgens (and of oestrogens to a lowergrade) in hamsters apparently results in an immuno-suppressive state, that favours proliferation of trophozoitesin the liver and inhibits their migration or invasive activity.In agreement with our results, immunosuppressive effectsby gonadectomy was also observed in females of Calomyscallosus infected with the ‘Y’ strain of T. cruzi, in spite ofthe fact, that in this case, immunosuppression induced anincrease in the parasitaemia during the course of infection(16). This hypothesis is controversial to the notion thatandrogens play an immunosuppressive role as observed inautoimmune diseases (32,33).
In experimental ALA in hamsters, the acute inflamma-tory response is characterized by the presence of neutro-phils, macrophages and lymphocytes, that promotedestruction of the surrounding cells (26,27,34). Eventhough INF-c and TNF-a activate neutrophils and macro-phages in vitro resulting in amoebicide activity mediatedby nitric oxide (35–37), experiments in vivo have shownthat neutrophils and macrophages are not capable of kill-ing amoebas, probably due to the superoxide dismutaseand to the oxidoreductase produced by the trophozoites,which inhibit the respiratory burst (38). It is also knownthat macrophages from ALA are unresponsive to TNF-aand bacterial LPS (38). The host’s cellular immune
Volume 31, Number 8, August 2009 Sex hormones deficiency and amoebic liver abscess
� 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456 453

response against parasites is regulated by cytokines, andtherefore the Th1 ⁄ Th2 type polarization can result inprotection or susceptibility (39). Several reports havedescribed the ability of sex hormones to influence all cellu-lar types of the innate and adaptive immune systems,thereby influencing a multitude of immunological func-tions (40,41). Moreover, sex hormones influence the devel-opment, maturation and state of activation of Tlymphocytes, including the Th1 ⁄ Th2 balance (42,43).
It has been reported that in the late phase of ALA ingerbils, when animals recover from an acute immuno-suppression, a Th1 response and particularly IL-2 seemsto be the major cytokine responsible for protection,whereas Th2 cytokines (IL-4 and IL-10) seem to associatewith susceptibility (44–46). However, there are no studiesdemonstrating such apparently roles of Th1 and Th2responses in ALA, as well as on the role of regulatorycytokines on the susceptibility or resistance to invasiveamoebiasis in hamsters. Our study sampled a broad profileof the cytokines that supposedly influence the course ofamoebic liver infection in hamsters. Semi-quantitativeimmunohistochemical analysis on confocal images of ALAfrom Gdx + infected female hamsters, suggested that thedeficiency of sex hormones resulted in a polarization ofthe cellular response to the Th2 type, which could protectthe liver from the damage caused by the Th1 responsedeveloped in the animals of the infected group (Table 2).On the other hand, Gdx + infected male hamsters exhib-ited a high profile of regulatory cytokines, mainly TGF-b,suggesting an even better control of the Th1 response andtherefore, a higher level of protection against ALA(Table 1). The protective effect of the Th1 response foundin gerbils against ALA, is observed only after 20 dayspost-infection (47). The course of infection in hamsters ismore acute and aggressive; most of the animals die in1–2 weeks (47). Our results support the notion that astrong Th1 response is responsible for tissue destruction asdescribed elsewhere (26,27), being the sex hormones neces-sary, at least in part, for this Th1 response to take place(Gdx hamsters of both sexes did not develop a Th1response).
Hormone deficiency could also have a direct effect onthe parasite. In this regard, amoebas have the ability tomodulate the expression of molecules related with viru-lence upon contact with host components, including hor-mones. In other parasites, specific hormone receptors havebeen shown to regulate the expression of diverse molecules(48,49). Our preliminary data indicate that a nonclassicreceptor for DHEA is present in the trophozoites ofE. histolytica (not shown). DHEA is an adrenal hormonewith multiple functions able to affect the growth of severalparasites, including amoeba (25,49).
Although the hamster model used here possibly doesnot reflect many aspects of human ALA, a set of interac-tions are established between the parasite, the immunesystem and the endocrine system, which define the out-come of the infection caused by E. histolytica. Evidence ispresented here, showing that deficiency of sex hormones inhamsters reduces progression of ALA, possibly due toreduced inflammatory and Th1 immune responses,suggesting that interactions between the immune andendocrine systems play a fundamental role in the establish-ment, development and outcome of extraintestinal amoe-bic infection in hamsters.
The evidence presented in our work illustrates theimportance of immunoendocrine interactions in an immu-nocompetent host. It strongly suggests an important rolefor sex steroids in the cytokine network. In practical mat-ters, the complexity of the immunoendocrine interactionssuggests that all physiological factors (i.e. sex, age anddevelopmental stage) should be taken into account in thedesign of vaccines and new drugs. Interventions aimed atthe hormonal network appear as a possible new thera-peutic approach to control several immune confrontations,such as amoebiasis.
ACKNOWLEDGEMENTS
This work was supported by DGAPA grants IN-227707(JCC) and IN-230207 (JPL) and CONACyT grants 61111(JCC) and 61334 (JPL). CCR has a fellowship from CO-NACyT. We thank M. Nequiz for supplying HM1 : IMSStrophozoites and his assistance during the infection ofhamsters. We also thank K. McGuigan for correcting theEnglish version of the manuscript and G. Ishiwara forhormone determinations.
REFERENCES
1 Homo-Delarche F, Fitzpatrick F, Christeff N, Nunez EA, BachJF & Dardenne M. Sex steroids, glucocorticoids, stress andautoimmunity. J Steroid Biochem Mol Biol 1991; 40: 619–637.
2 Klein SL. The effects of hormones on sex differences in infec-tion: from genes to behavior. Neurosci Biobehav Rev 2000; 24:627–638.
3 Grossman C. Possible underlying mechanisms of sexual dimor-phism in the immune response, fact and hypothesis. J SteroidBiochem 1989; 5: 325–346.
4 Bouman A, Heineman MJ & Faas MM. Sex hormones andthe immune response in humans. Hum Reprod Update 2005;11: 411–423.
5 Sapino A, Cassoni P, Ferrero E, Bongiovanni M, Righi L, For-tunati N, Crafa P, Chiarle R & Bussolati G. Estrogen receptoralpha is a novel marker expressed by follicular dendritic cellsin lymph nodes and tumor-associated lymphoid infiltrates. AmJ Pathol 2003; 163: 1313–1320.
C. Cervantes-Rebolledo et al. Parasite Immunology
454 � 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456

6 Wunderlich F, Benten WP, Lieberherr M, Guo Z, Stamm O,Wrehlke C, Sekeris CE & Mossmann H. Testosteronesignaling in T cells and macrophages. Steroids 2002; 67:535–538.
7 Benten WP, Stephan C & Wunderlich F. B cells express intra-cellular but not surface receptors for testosterone and estra-diol. Steroids 2002; 67: 647–654.
8 McKay LI & Cidlowski JA. Molecular control ofimmune ⁄ inflammatory responses: interactions between nuclearfactor-kappa B and steroid receptor-signaling pathways.Endocr Rev 1999; 20: 435–459.
9 Klein SL. Hormonal and immunological mechanisms mediat-ing sex differences in parasite infection. Parasite Immunol 2004;26: 247–264.
10 Benten WP, Ulrich P, K�hn-Velten WN, Vohr HW & Wunder-lich F. Testosterone-induced susceptibility to Plasmodium chab-audi malaria: persistence after withdrawal of testosterone.J Endocrinol 1997; 153: 275–281.
11 Wunderlich F, Maurin W, Benten WP & Schmitt-Wrede HP.Testosterone impairs efficacy of protective vaccination againstP. chabaudi malaria. Vaccine 1993; 11: 1097–1099.
12 Kamis AB & Ibrahim JB. Effects of testosterone on blood leu-kocytes in Plasmodium berghei-infected mice. Parasitol Res1989; 75: 611–613.
13 Kr�cken J, Dkhil MA, Braun JV, Schroetel RM, El-KhadragyM, Carmeliet P, Mossmann H & Wunderlich F. Testosteronesuppresses protective responses of the liver to blood-stagemalaria. Infect Immun 2005; 73: 436–443.
14 Cernetich A, Garver LS, Jedlicka AE, Klein PW, Kumar N,Scott AL & Klein SL. Involvement of gonadal steroids andgamma interferon in sex differences in response to blood-stagemalaria infection. Infect Immun 2006; 74: 3190–3203.
15 do Prado JC Jr, Levy AM, Leal MP, Bernard E & Kloetzel JK.Influence of male gonadal hormones on the parasitaemia andhumoral response of male Calomys callosus infected with the Ystrain of Trypanosoma cruzi. Parasitol Res 1999; 85: 826–829.
16 do Prado JC Jr, Leal MP, Anselmo-Franci JA, de Andradejfflniur HF & Kloetzel JK. Influence of female gonadal hor-mones on the parasitemia of female Calomys callosus infectedwith the ‘‘Y’’ strain of Trypanosoma cruzi. Parasitol Res 1998;84: 100–105.
17 Santos CD, Levy AM, Toldo MP, Azevedo AP & do Prado JCJr. Haematological and histopathological findings after ovari-ectomy in Trypanosoma cruzi infected mice. Vet Parasitol 2007;143: 222–228.
18 Mota EA, Guimaraes AC, Santana OO, Sherlock I, Hoff R &Weller TH. A nine year the prospective study of Chagas dis-ease in a defined rural population in northeast Brazil. Am JTrop Med Hyg 1990; 42: 429–440.
19 Castro C, Macedo V, Rezende JM & Prata A. Longitudinalradiologic study of esophagus, in an endemic area of Chagasdisease, in a period of 13 years. Rev Soc Bras Soc Med Trop1994; 27: 227–233.
20 Ravdin JI. Amoebiasis. In Ravdin JI (ed): Human Infection byEntamoeba Histolytica. New York, Churchill Livingstone,1988: 838.
21 Hughes MA & Petri WA Jr. Amebic liver abscess. Infect DisClin North Am 2000; 14: 565–582.
22 Lotter H, Jacobs T, Gaworski I & Tannich E. Sexual dimor-phism in the control of amebic liver abscess in a mouse modelof disease. Infect Immun 2006; 74: 118–124.
23 Gonz�lez MF, Tanimoto WM, V�zquez SJA, Calder�n P &Aguirre GJ. Efecto de los estr�genos en h�msteres con amibia-sis en el h�gado. Arch Invest M�d 1972; 2: 335–341.
24 Gil JN, Ganguly KN, Majan RC, Bhusnurmath RS & DilawariBJ. Progesterone-induced liver abscess in guinea-pigs-anewmodel. Trans R Soc Trop Med Hyg 1983; 77: 53–58.
25 Carrero JC, Cervantes C, Moreno-Mendoza N, Saavedra E,Morales-Montor J & Laclette JP. Dehydroepiandrosteronedecreases while cortisol increases in vitro growth and viabilityof Entamoeba histolytica. Microbes Infect 2006; 8: 323–331.
26 Tsutsumi V, Mena-L�pez R, Anaya-Velazquez F & Mart�nez-Palomo A. Cellular bases of experimental amebic liver abscessformation. Am J Pathol 1984; 117: 81–91.
27 Olivos-Garc�a A, Carrero JC, Ramos E, Nequiz M, Tello E,Montfort I & P�rez-Tamayo R. Late experimental amebic liverabscess in hamster is inhibited by cyclosporine and N-acetyl-cysteine. Exp Mol Pathol 2007; 82: 310–315.
28 Wunderlich F, Marinovski P, Benten WP, Schmitt-Wrede HP& Mossmann H. Testosterone and other gonadal factor(s)restrict the efficacy of genes controlling resistance to Plasmo-dium chabaudi malaria. Parasite Immunol 1991; 13: 357–367.
29 Mock BA & Nacy CA. Hormonal modulation of sex differ-ences in resistance to Leishmania major systemic infections.Infect Immun 1988; 56: 3316–3319.
30 Roberts CW, Walker W & Alexander J. Sex-associated hor-mones and immunity to protozoan parasites. Clin MicrobiolRev 2001; 14: 476–488.
31 Marriott I & Huet-Hudson YM. Sexual dimorphism in innateimmune responses to infectious organisms. Immunol Res 2006;34: 177–192.
32 Malkin CJ, Pugh PJ, Jones RD, Jones TH & Channer KS. Tes-tosterone as a protective factor against atherosclerosis – immu-nomodulation and influence upon plaque development andstability. J Endocrinol 2003; 178: 373–380.
33 Cutolo M, Sulli A, Capellino S, Villaggio B, Montagna P, Seri-olo B & Straub RH. Sex hormones influence on the immunesystem: basic and clinical aspects in autoimmunity. Lupus2004; 13: 635–638.
34 Salata RA & Ravdin JI. Review of the human immune mecha-nisms directed against Entamoeba histolytica. Rev Infect Dis1986; 8: 261–272.
35 Denis M & Chadee K. Human neutrophils activated by inter-feron-gamma and tumour necrosis factor-alpha kill Entamoebahistolytica trophozoites in vitro. J Leukoc Biol 1989; 46: 270–274.
36 Lin JY, Seguin R, Keller K & Chadee K. Tumor necrosis factoralpha augments nitric oxide-dependent macrophage cytotoxicityagainst Entamoeba histolytica by enhanced expression of thenitric oxide synthase gene. Infect Immun 1994; 62: 1534–1541.
37 Ghadirian E & Denis M. In vivo activation of macrophages byIFN-gamma to kill Entamoeba histolytica trophozoites in vitro.Parasite Immunol 1992; 14: 397–404.
38 Wang W, Keller K & Chadee K. Entamoeba histolytica modu-lates the nitric oxide synthase gene and nitric oxide productionby macrophages for cytotoxicity against amoebae and tumourcells. Immunology 1994; 83: 601–610.
39 Spellberg B & Edwards JE. Type 1 ⁄ Type 2 immunity in infec-tions diseases. Clin Infect Dis 2001; 32: 76–102.
40 Lamason R, Zhao P, Rawat R, Davis A, Hall JC, Chae JJ,Agarwal R, Cohen P, Rosen A, Hoffman EP & Nagaraju K.Sexual dimorphism in immune response genes as a function ofpuberty. BMC Immunol 2006; 7: 2.
Volume 31, Number 8, August 2009 Sex hormones deficiency and amoebic liver abscess
� 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456 455

41 Roberts CW, Satoskar A & Alexander J. Sex steroids, preg-nancy-associated hormones and immunity to parasitic infec-tion. Parasitol Today 1996; 12: 382–388.
42 Radojevic K, Arsenovic-Ranin N, Kosec D, Pesic V, PilipovicI, Perisic M, Plecas-Solarovic B & Leposavic G. Neonatal cas-tration affects intrathymic kinetics of T-cell differentiation andthe spleen T-cell level. J Endocrinol 2007; 192: 669–682.
43 Seiki K & Sakabe K. Sex hormones and the thymus in relationto thymocyte proliferation and maturation. Arch Histol Cytol1997; 60: 29–38.
44 Kretschmer RR & Lopez-Osuna M. Effector mechanisms andimmunity to amebas. In Krestschmer R (ed): Amebiasis: Infec-tion and Disease by E. Histolytica. Boca Rat�n, FL, CRCPress, 1990: 105–122.
45 Campbell D, Gaucher D & Chadee K. Serum from Entamoebahistolytica-infected gerbils selectively suppresses T cell prolifer-
ation by inhibiting interleukin-2 production. J Infect Dis 1999;179: 1495–1501.
46 Campbell D & Chadee K. Interleukin (IL)-2, IL-4, and tumornecrosis factor-alpha responses during Entamoeba histolyticaliver abscess development in gerbils. J Infect Dis 1997; 175:1176–1183.
47 Tsutsumi V & Shibayama M. Experimental amebiasis: aselected review of some in vivo models. Arch Med Res 2006;37: 210–220.
48 de MendonÅa RL, Escriv� H, Bouton D, Laudet V & PierceRJ. Hormones and nuclear receptors in schistosome develop-ment. Parasitol Today 2000; 16: 233–240.
49 Escobedo G, Roberts CW, Carrero JC & Morales-Montor J.Parasite regulation by host hormones: an old mechanism ofhost exploitation? Trends Parasitol 2005; 21: 588–593.
C. Cervantes-Rebolledo et al. Parasite Immunology
456 � 2009 Blackwell Publishing Ltd, Parasite Immunology, 31, 447–456
















![[Meticilin resistant Staphylococcus aureus and liver abscess: a retrospective analysis of 117 patients]](https://static.fdokumen.com/doc/165x107/632546fd545c645c7f099e01/meticilin-resistant-staphylococcus-aureus-and-liver-abscess-a-retrospective-analysis.jpg)



