
Microtubule Assembly of Isotypically Purified Tubulin and Its Mixtures
Upload
independentCategory
view
0download
0
MP
GD
DB
AfvpdtlmmbgtTsnpI
Kb
Lanidda2waNllrdtiebntTi
*EA
Neuroscience 154 (2008) 946–952
0d
ICROTUBULE STABILIZING EFFECT OF NOTCH ACTIVATION IN
RIMARY CORTICAL NEURONSiJetca
P
TtiiCsLma-aien
C
JrNmmat
Jm
R
Tcg(st
Q
Ot(mvso
. FERRARI-TONINELLI, S. A. BONINI, P. BETTINSOLI,
. UBERTI AND M. MEMO*
epartment of Biomedical Sciences and Biotechnologies, University ofrescia, viale Europa, 11, 25123 Brescia, Italy
bstract—The appropriate level of microtubule stability isundamental in neurons to assure correct polarity, migration,esicles transport and to prevent axonal degeneration. In theresent study, we have identified Notch pathway as an en-ogenous microtubule stabilizer. Stimulation of Notch recep-ors by exposure of mouse cortical neurons to the Notchigand Jagged1 resulted in increased microtubule stability, as
easured by using antibodies against post-translationallyodified � tubulin, and changes in axonal morphology and
ranching, with varicosity loss, thicker neurites and enlargedrowth cones. Similar effects were found after exposure ofhe cells to different doses of Taxol. However, contrary toaxol, Jagged1 induced downregulation of the microtubuleevering protein Spastin. We suggest that a fine-tuned ma-ipulation of Notch signaling may represent a novel ap-roach to modulate neuronal cytoskeleton plasticity. © 2008
BRO. Published by Elsevier Ltd. All rights reserved.
ey words: Jagged1, acetylated tubulin, Spastin, microtu-ules dynamics.
oss of neuronal connectivity, mainly caused by synapticlterations, is considered as one of the main causes ofeuronal degeneration in several neurological diseases
ncluding Alzheimer’s disease, tauopathies, Parkinson’sisease, multiple sclerosis and schizophrenia. Emergingata suggest that connectivity disorders can result frombnormalities in the microtubule system (Andrieux et al.,006; Cappelletti et al., 2005). One of the molecular path-ays responsible for the differentiation, morphogenesisnd functioning of the CNS is the Notch pathway. Theotch transmembrane receptor, after binding to specific
igands, such as Delta and Jagged, is cleaved and trans-ocates to the nucleus where it controls transcriptionalepressors (Kageyama and Ohtsuka, 1999). In terminallyifferentiated neurons, Notch regulates the local architec-ure of the cells probably through the contact-dependentnhibition of neurite growth (Sestan et al., 1999; Ber-zovska et al., 1999; Salama-Cohen et al., 2006). It haseen suggested that the Notch pathway role in maintainingeuronal arborization in the adult is linked to its capabilityo modulate cytoskeleton plasticity (Louvi and Artavanis-sakonas, 2006). Here we investigated on the possible
nfluence of Notch pathway on microtubules stability. In an
Corresponding author. Tel: �39-0303717516; fax: �39-0303717529.
u-mail address: [email protected] (M. Memo).bbreviation: cDNA, complementary DNA; DIV, days in vitro.
306-4522/08$32.00�0.00 © 2008 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2008.04.025
946
n vitro model of mouse cortical neurons, the effects of theagged1 ligand stimulation were compared with those ex-rted by the microtubule stabilizing drug Taxol. We foundhat exposure of the cells to Jagged1 resulted in an in-reased microtubule stabilization, suggesting Notch pathways a novel target for modulating cytoskeleton plasticity.
EXPERIMENTAL PROCEDURES
rimary cortical neurons cultures
o obtain primary cortical neurons, embryonic day 15 (E15) cor-ices were isolated from CD1 mouse brains (Charles River Breed-ng Laboratories, Calco, Italy), pooled, mechanically dissociatednto a single-cell suspension in Neurobasal medium (Invitrogenorporation, San Giuliano Milanese, Italy) containing 2% B27upplement (Invitrogen), 500 �M glutamine (Sigma-Aldrich, St.ouis, MO, USA), 100 units/ml penicillin, and 100 �g/ml strepto-ycin (Sigma-Aldrich) (N-B27 medium), and centrifuged for 5 mint 200�g. Cells were plated onto poly-L-lysine (Sigma-Aldrich)coated glass coverslips or dishes (2.5�104–2.5�106 cells/cm2)nd cultured in N-B27 medium for 7 days in vitro (DIV). All exper-
ments conformed to local and international guidelines on thethical use of animals. All efforts were made to minimize theumber of animals used and their suffering.
ell treatments
agged1 (CDDYYYGFGCNKFCRPR corresponding to Jagged1esidues 188 –204) or the scrambled peptide (RCGPDCFD-YGRYKYCF) (Nickoloff et al., 2002) was added to the cultureedium at concentrations ranging from 20 �M to 200 �M, for 30in to 2 h, as indicated. Taxol (Paclitaxel, Sigma-Aldrich) wasdded to the culture media at concentrations ranging from 10 nMo 1 �M, as indicated.
�-Secretase inhibitor IV (Calbiochem, EMD Biosciences, Laolla, CA, USA) was added to the culture medium, at 10 �M, 30in before Jagged1 peptide.
NA isolation
otal RNA was isolated from untreated and Jagged-treated corti-al cells 7 DIV (about 7.5�105 cells) using the RNeasy kit (Qia-en, Milano, Italy) and digested with the RNase-Free DNAse setQiagen), according to the manufacturer protocol. RNA quality ofamples was tested by RNA electrophoresis to ensure RNA in-egrity.
-RT-PCR
ne microgram of total RNA from untreated and Jagged1-reated cortical cells was transcribed into complementary DNAcDNA) using murine leukemia virus reverse transcriptase (Pro-ega Italia, Milano, Italy) and oligodT15-18 as a primer (final
olume: 50 �l). Parallel reactions containing no reverse tran-criptase were used as negative controls to confirm the removalf all genomic DNA. Murine specific primers were designed
sing the Primer3 software (http://frodo.wi.mit.edu) (Rozen andved.
SuCGAT
itwSamoa9AsegnwoibcJpu
eJ
S
Sah
PfN
ionbg1Vaptd(
Fttm
G. Ferrari-Toninelli et al. / Neuroscience 154 (2008) 946–952 947
kaletsky, 2000). The oligonucleotide sequences of the primerssed are as follow: Spastin forward primer 5=-ATC CGA GAATG AAG CCA GA-3=, reverse primer 5=-GCC AAA GCT CTTTT TCC TG-3=, GAPDH forward primer 5=-AAC TTT GGCTT GTG GAA GG-3=, reverse primer 5=-ACA CAT TGG GGGAG GAA CA-3=.
Amplification and detection were performed with the iCYCLERQ Real Time PCR Detection System (BioRad Italia, Milano, Italy);he fluorescence signal was generated by SYBR Green I. Samplesere run in triplicate in a 25 �l reaction mix containing 12.5 �l 2�YBR Green Master Mix (BioRad Italy), 12.5 pmol of each forwardnd reverse primer and 2 �l of diluted cDNA. Each PCR experi-ent included serial dilutions of a positive control for constructionf the calibration curve, a positive and a negative DNA sample,nd water blanks. The PCR program was initiated by 10 min at5 °C before 40 cycles, each for 15 s at 95 °C and 1 min at 56 °C.
subsequent dissociation curve analysis verified the productspecificity. Gene expression levels were normalized to GAPDHxpression and data are presented as the fold change in targetene expression in Jagged1-treated cells normalized to the inter-al control gene (GAPDH) and relative to untreated cells. Resultsere estimated as Ct values; the Ct was calculated as the meanf the Ct for the target gene minus the mean of the Ct for the
nternal control gene. The Ct represented the mean differenceetween the Ct of untreated cells minus the Ct of Jagged1-treatedells. The N-fold differential expression in the target gene ofagged-treated cells compared with untreated cells was ex-ressed as 2���Ct. Data analysis and graphics were performedsing GraphPad Prism 4 software and were the results of a single
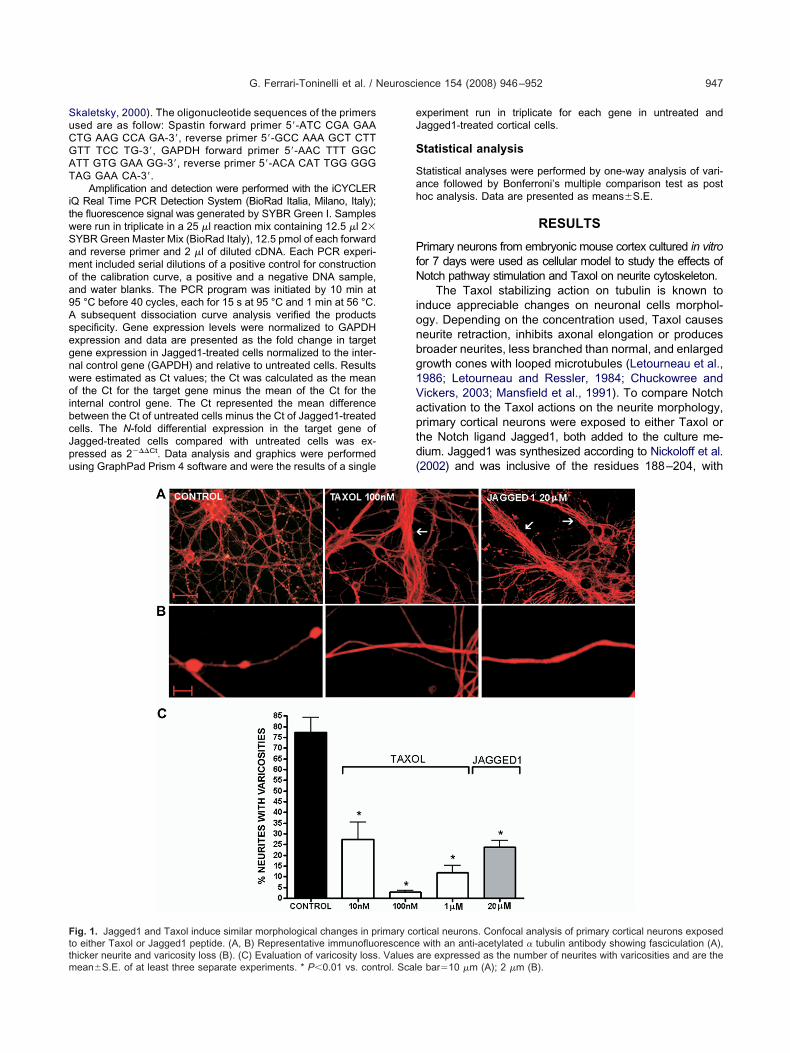
ig. 1. Jagged1 and Taxol induce similar morphological changes in po either Taxol or Jagged1 peptide. (A, B) Representative immunofluo
hicker neurite and varicosity loss (B). (C) Evaluation of varicosity loss. Valuesean�S.E. of at least three separate experiments. * P�0.01 vs. control. Scalxperiment run in triplicate for each gene in untreated andagged1-treated cortical cells.
tatistical analysis
tatistical analyses were performed by one-way analysis of vari-nce followed by Bonferroni’s multiple comparison test as postoc analysis. Data are presented as means�S.E.
RESULTS
rimary neurons from embryonic mouse cortex cultured in vitroor 7 days were used as cellular model to study the effects ofotch pathway stimulation and Taxol on neurite cytoskeleton.
The Taxol stabilizing action on tubulin is known tonduce appreciable changes on neuronal cells morphol-gy. Depending on the concentration used, Taxol causeseurite retraction, inhibits axonal elongation or producesroader neurites, less branched than normal, and enlargedrowth cones with looped microtubules (Letourneau et al.,986; Letourneau and Ressler, 1984; Chuckowree andickers, 2003; Mansfield et al., 1991). To compare Notchctivation to the Taxol actions on the neurite morphology,rimary cortical neurons were exposed to either Taxol orhe Notch ligand Jagged1, both added to the culture me-ium. Jagged1 was synthesized according to Nickoloff et al.2002) and was inclusive of the residues 188–204, with
rtical neurons. Confocal analysis of primary cortical neurons exposedwith an anti-acetylated � tubulin antibody showing fasciculation (A),
rimary corescence
are expressed as the number of neurites with varicosities and are thee bar�10 �m (A); 2 �m (B).

NpissowmmCT
crbwrdr(baarmt1srrs(p
tbimtihpzccwisiir(wetcaaatra
ot(a
FJ(m
G. Ferrari-Toninelli et al. / Neuroscience 154 (2008) 946–952948
otch agonist activity in vitro and on organotypic hip-ocampal slices (Wang et al., 2004). The peptide specific-
ty was validated by negative control obtained with acrambled peptide without agonist activity (data nothown). Jagged1 was added to the culture media at vari-us concentrations, ranging from 20 �M to 200 �M; 20 �Mas chosen as the lowest concentration able to exertorphological effects. Taxol was also added to the cultureedium at different doses, ranging from 10 nM to 1 �M.ells were exposed to Jagged1, the scrambled peptide oraxol for 1 h.
As shown in representative pictures in Fig. 1A and B,onfocal microscopy analysis using anti-tubulin antibodyevealed similar morphological changes of the neurites inoth Jagged1 and Taxol-treated cells, when comparedith control. In particular, Taxol- and Jagged1-treated neu-
ons both showed thicker neurites; this change could beue to an increased neurite fasciculation or a microtubuleseorganization, leading single processes to become thickerA). The neurites of treated neurons appeared much lessranched than controls and varicosity loss was evident alllong the neuronal processes (B). Sometimes we wereble to detect, in both Taxol- and Jagged1-treated neu-ons, the presence of enlarged growth cones with loopedicrotubules (data not shown). Loss of varicosities along
he neurites was quantified and statistically analyzed (Fig.C). Taxol appeared more efficacious than Jagged1 inmoothing the neurites: in fact, 10 nM Taxol significantlyeduced varicose processes (27.47%�8 of varicose neu-ites versus 77.3%�7 of the control) whereas to obtainimilar results 20 �M Jagged1 peptide was needed23.9%�3.1 versus 77.3�7 of the control). Given the com-arable morphological effects, we were prompted to inves-
ig. 2. Jagged1 and Taxol treatments induce tubulin acetylation. Primagged1 peptide, then analyzed by immunofluorescence using an anti-
A) revealed a significant increase in tubulin acetylation with both the treatmenean�S.E. of at least three separate experiments. * P�0.01 vs. control. Reprigate if Jagged1 treatment, like Taxol, induced microtu-ules stabilization. Stable microtubules are commonly
dentified by the presence of tubulin post-translationalodifications, such as � tubulin acetylation or polyglu-
amylation. On the contrary, the level of tyrosinated tubulinndicates the presence of more instable microtubules (Ver-ey and Gaertig, 2007; Hammond et al., 2008). To com-are the Jagged1 effect to Taxol on microtubules stabili-ation, a fluorescence intensity analysis by confocal mi-roscopy was performed on cells treated with increasingoncentrations of Taxol or Jagged1 and immunostainedith an anti-acetylated � tubulin antibody. Representative
mages are shown in Fig. 2B. Statistical analysis in Fig. 2Ahowed that 100 nM Taxol produced a significant increase
n the acetylated � tubulin expression levels (mean pixelntensity 43.8�3.5 versus 25.3�1.9 of control). Similaresults were obtained in cells exposed to 20 �M Jagged138.6�2.9 versus 25.3�1.9). Neurons treated with Jagged1ere further tested by immunofluorescence for the pres-nce of polyglutamylated and tyrosinated � tubulin. An �
ubulin antibody was also used to evaluate possiblehanges in the total level of the protein (Fig. 3). Statisticalnalysis of each tubulin immunofluorescence level showedsignificant increased of the stabilization markers (acetylatednd polyglutamylated tubulin), evident 1 h after Jagged1
reatment, without changes in the total � tubulin levels. Down-egulation of instable, tyrosinated tubulin was also detectable,lthough it did not reach statistical significance.
Microtubule dynamics is maintained in neurons by thepposite action of stabilizing proteins, such as the micro-ubules associated proteins (MAPs) family, including tauAvila et al., 2004; Li and Feng, 2006), and proteins that acts microtubules severing, such as Katanin and Spastin.
cal neurons were treated with increasing concentrations of Taxol andd � tubulin antibody. Statistical analysis performed on confocal images
ary cortiacetylate
ts. Values are expressed as mean fluorescence intensity and are theesentative images are shown in B. Scale bar�20 �m.

TfTSogdTpo
csttblasn
Fstit�
G. Ferrari-Toninelli et al. / Neuroscience 154 (2008) 946–952 949
hese proteins promote microtubule remodeling requiredor neurites outgrowth and branching (Baas et al., 2005;rotta et al., 2004). Interestingly, gene profile analysis ofH-SY5Y neuroblastoma cells exposed to Jagged1 dem-nstrated downregulation of Spastin mRNA levels, sug-esting that Spastin may play a role as Notch action me-iator (Ferrari-Toninelli et al., unpublished observations).o verify if a similar downregulation could occur in corticalrimary neurons, a real-time PCR analysis was performed
ig. 3. Jagged1 treatment leads to tubulin post-translational modificatained with: anti-acetylated � tubulin antibody (A), anti-polyglutamylubulin (D). Cells’ fluorescence intensity was then measured using thencrease in microtubule stabilizing post-translational modifications (aceyrosinated tubulin, typical feature of instable tubulin, and no changes inS.E., * P�0.01 vs. control.
n control and Jagged1-treated cells. The results (Fig. 4) i
onfirmed a significant decrease in Spastin mRNA levelstarting from 1 h treatment. Primary cortical neuronsreated with either Taxol or Jagged1 at various concentra-ions for 1 h were immunostained with a polyclonal anti-ody directed against Spastin, then protein expression
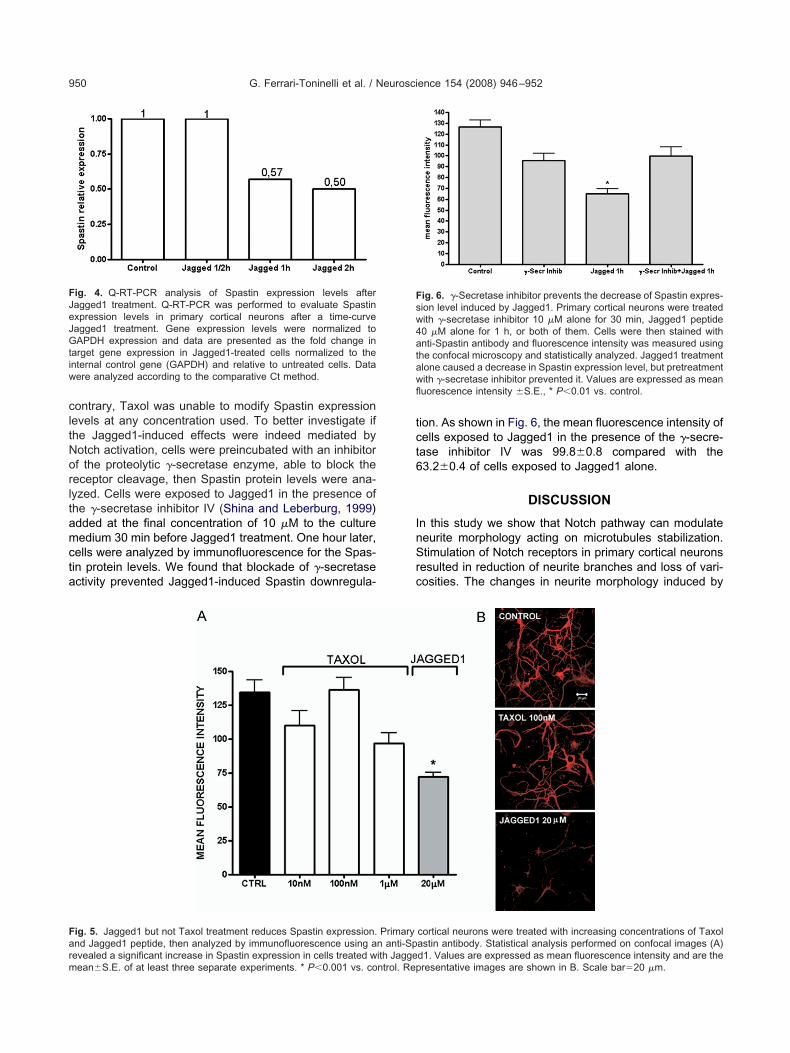
evels were quantified by densitometry and statisticallynalyzed. Representative images are shown in Fig. 5B. Ashown in Fig. 5A, 20 �M Jagged1 reduced Spastin immu-ofluorescence intensity of about 50% (mean fluorescence
ary cortical neurons were treated with Jagged1 peptide 40 �M thenlin antibody (B), anti-tyrosinated tubulin antibody (C) and anti-alphamicroscopy and statistically analyzed. Jagged1 treatment caused annd polyglutamylation), a decrease, also if no statistically significant, inha tubulin level. Values are expressed as mean fluorescence intensity
tions. Primated tubuconfocaltylation atotal alp
ntensity 72.4�3.1 versus 134.7�8.9 of the control); on the
cltNorltamcta
tct6
InSrc
Fsw4atawfl
FJeJGtiw
Farm
G. Ferrari-Toninelli et al. / Neuroscience 154 (2008) 946–952950
ontrary, Taxol was unable to modify Spastin expressionevels at any concentration used. To better investigate ifhe Jagged1-induced effects were indeed mediated byotch activation, cells were preincubated with an inhibitorf the proteolytic �-secretase enzyme, able to block theeceptor cleavage, then Spastin protein levels were ana-yzed. Cells were exposed to Jagged1 in the presence ofhe �-secretase inhibitor IV (Shina and Leberburg, 1999)dded at the final concentration of 10 �M to the cultureedium 30 min before Jagged1 treatment. One hour later,
ells were analyzed by immunofluorescence for the Spas-in protein levels. We found that blockade of �-secretasectivity prevented Jagged1-induced Spastin downregula-
ig. 4. Q-RT-PCR analysis of Spastin expression levels afteragged1 treatment. Q-RT-PCR was performed to evaluate Spastinxpression levels in primary cortical neurons after a time-curveagged1 treatment. Gene expression levels were normalized toAPDH expression and data are presented as the fold change in
arget gene expression in Jagged1-treated cells normalized to thenternal control gene (GAPDH) and relative to untreated cells. Dataere analyzed according to the comparative Ct method.
ig. 5. Jagged1 but not Taxol treatment reduces Spastin expression.nd Jagged1 peptide, then analyzed by immunofluorescence using a
evealed a significant increase in Spastin expression in cells treated with Jaggeean�S.E. of at least three separate experiments. * P�0.001 vs. control. Repion. As shown in Fig. 6, the mean fluorescence intensity ofells exposed to Jagged1 in the presence of the �-secre-ase inhibitor IV was 99.8�0.8 compared with the3.2�0.4 of cells exposed to Jagged1 alone.
DISCUSSION
n this study we show that Notch pathway can modulateeurite morphology acting on microtubules stabilization.timulation of Notch receptors in primary cortical neurons
esulted in reduction of neurite branches and loss of vari-osities. The changes in neurite morphology induced by
ig. 6. �-Secretase inhibitor prevents the decrease of Spastin expres-ion level induced by Jagged1. Primary cortical neurons were treatedith �-secretase inhibitor 10 �M alone for 30 min, Jagged1 peptide0 �M alone for 1 h, or both of them. Cells were then stained withnti-Spastin antibody and fluorescence intensity was measured using
he confocal microscopy and statistically analyzed. Jagged1 treatmentlone caused a decrease in Spastin expression level, but pretreatmentith �-secretase inhibitor prevented it. Values are expressed as meanuorescence intensity �S.E., * P�0.01 vs. control.
cortical neurons were treated with increasing concentrations of Taxolastin antibody. Statistical analysis performed on confocal images (A)
Primaryn anti-Sp
d1. Values are expressed as mean fluorescence intensity and are theresentative images are shown in B. Scale bar�20 �m.

Jc
cgmppmwisoNctae
iaNtml2bpt2iSfraortmlot2
Ismitaatvlscn
uNdghatNa
Af
A
A
B
B
B
C
C
E
H
K
L
L
L
L
L
M
N
N
R
G. Ferrari-Toninelli et al. / Neuroscience 154 (2008) 946–952 951
agged1 were comparable to those induced by low con-entrations of the microtubules stabilizing drug Taxol.
Post-translational modification of microtubules isonsidered as one of the main mechanisms involved inenerating functional diversity of microtubules. Theseodifications, which include detyrosination/tyrosination,olyglutamylation, polyglycylation, palmytoilation, phos-horylation, and acetylation (Verhey and Gaertig, 2007)ay occur individually or in a combinatorial fashion. In thisay, microtubule post-translational modifications play an
mportant role in regulating tubulin properties, such astability and structure (Hammond et al., 2008). Evidencesf an increased microtubule stabilization, suggesting thatotch pathway also could act through cytoskeletal modifi-ations, were provided by the upregulation of two post-ranslationally modified tubulins, the acetylated � tubulinnd the polyglutamylated tubulin, commonly used as mark-rs of stability (Hammond et al., 2008).
Real time PCR analysis and immunofluorescencedentified Spastin protein as a possible mediator of Notchction on microtubules, suggesting that stimulation ofotch pathway by Jagged1 peptide inhibits the transcrip-
ion and the expression levels of this protein. Spastin geneutation has been associated with axonal degeneration
eading to hereditary spastic paraplegia (Errico et al.,002). Further studies showed that Spastin is a microtu-ule severing protein and that its microtubule destabilizingroperties are fundamental for axon outgrowth and synap-ic modulation in the long motor neurons (Wood et al.,006; Trotta et al., 2004; Sherwood et al., 2004); interest-
ngly, a recent study showed that in cultured neuronspastin is more concentrated at the sites of branches
ormation and that protein downregulation resulted in neu-ite morphology changes, with a dramatic reduction ofxonal branches (Yu et al., 2008). Experiments of siRNAr protein overexpression also allowed us to establish aelationship between posttranslational tubulin modifica-ions and microtubules severing. In fact, in a Drosophilaodel, downregulation of Spastin protein led to increased
evels of acetylated and polyglutamylated tubulin; on thether hand, Spastin overexpression reduced levels ofhese posttranslationally modified proteins (Trotta et al.,004).
CONCLUSION
n conclusion, we first established a link between Notchignaling pathway and microtubule stabilization in post-itotic neurons suggesting a novel endogenous pathway
nvolved in modulating microtubule plasticity. The fine-uned manipulation of Notch signaling is expected to guar-ntee neurons both the stability required to maintain theirxonal architecture and the structural plasticity necessaryo create new synaptic connections. The emerging rele-ance of microtubule stabilization in preventing synapticoss and preserving neuronal connectivity encourages theearch of agents endowed with cytoskeleton regulationapability. In this regard, it was recently found that anti-
eoplastic agents able to stabilize microtubules could beseful in preventing synaptic loss (Trojanowski et al., 2005;elson, 2005; Butler et al., 2007). However proposedrugs, such as Taxol and Taxol-like compounds have areat stabilizing effect on microtubules, but also a potentialigh neurotoxicity (Lee and Swain, 2006). Identification ofn endogenous, finely modulated mechanism devoid ofoxic effects is thus certainly required. In this context,otch pathway could be a target for developing therapiesgainst neurodegenerative diseases.
cknowledgments—This work was supported in part by grantrom the Italian Ministry of Education and Research, PRIN 2005.
REFERENCES
ndrieux A, Salin P, Schweitzer A, Begou M, Pachoud B, Brun P, Gory-Faure S, Kujala P, Suaud-Chagny MF, Hofle G, Job D (2006) Micro-tubule stabilizer ameliorates synaptic function and behavior in amouse model for schizophrenia. Biol Psychiatry 60(11):1224–1230.
vila J, Lucas JJ, Perez M, Hernandez F (2004) Role of tau protein in bothphysiological and pathological conditions. Physiol Rev (2):361–384.
aas PW, Karabay A, Qiang L (2005) Microtubules cut and run.Trends Cell Biol 15(10):518–524.
erezovska O, McLean P, Knowles R, Frosh M, Lu FM, Lux SE,Hyman BT (1999) Notch1 inhibits neurite outgrowth in postmitoticprimary neurons. Neuroscience 93(2):433–439.
utler D, Bendiske J, Michaelis ML, Karanian DA, Bahr BA (2007)Microtubule-stabilizing agent prevents protein accumulation-in-duced loss of synaptic markers. Eur J Pharmacol 562:20–27.
appelletti G, Surrey T, Maci R (2005) The parkinsonism producingneurotoxin MPP� affects microtubule dynamics by acting as adestabilising factor. FEBS Lett 579(21):4781–4786.
huckowree JA, Vickers JC (2003) Cytoskeletal and morphologicalalterations underlying axonal sprouting after localized transactionof cortical neuron axons in vitro. J Neurosci 23(9):3715–3725.
rrico A, Ballabio A, Rugarli EI (2002) Spastin, the protein mutated inautosomal dominant hereditary spastic paraplegia, is involved inmicrotubule dynamics. Hum Mol Genet 11(2):153–163.
ammond JW, Cai D, Verhey KJ (2008) Tubulin modifications andtheir cellular functions. Curr Opin Cell Biol 20(1):71–76.
ageyama R, Ohtsuka T (1999) The Notch-Hes pathway in mamma-lian neural development. Cell Res 9(3):179–188.
ee JJ, Swain SM (2006) Peripheral neuropathy induced by microtu-bule-stabilizing agents. J Clin Oncol 24(10):1633–1642.
etourneau PC, Ressler AH (1984) Inhibition of neurite initiation andgrowth by taxol. J Cell Biol 98:1355–1362.
etourneau PC, Shattuck TA, Ressler AH (1986) Branching of sensoryand sympathetic neurites in vitro is inhibited by treatment withtaxol. J Neurosci 6(7):1912–1917.
i W, Xia J, Feng Y (2006) Microtubule stability and MAP1B upregu-lation control neuritogenesis in CAD cells. Acta Pharmacol Sin27(9):1119–1126.
ouvi A, Artavanis-Tsakonas S (2006) Notch signalling in vertebrateneural development. Nat Rev Neurosci 7:93–102.
ansfield SG, Gordon Weeks PR (1991) Dynamic post-translationalmodification of tubulin in rat cerebral cortical neurons extendingneurites in culture: effects of taxol. J Neurocytol 20(8):654–666.
elson R (2005) Microtubule-stabilising drugs may be therapeutic inAD. Lancet Neurol 4(2):83–84.
ickoloff BJ, Quin JZ, Chaturvedi V, Denning MF, Bonish B, Miele L(2002) Jagged-1 mediated activation of notch signalling inducescomplete maturation of human keratinocytes through NF-�B andPPAR�. Cell Death Differ 9:842–855.
ozen S, Skaletsky H (2000) Primer3 on the WWW for general users
and for biologist programmers. Methods Mol Biol 132:365–386.
S
S
S
S
T
T
V
W
W
Y
G. Ferrari-Toninelli et al. / Neuroscience 154 (2008) 946–952952
alama-Cohen P, Arevalo MA, Grantyn R, Rodriguez-Tebar A (2006)Notch and NGF/p75NTR control dendrite morphology and thebalance of excitatory/inhibitory synaptic input to hippocampal neu-rones through Neurogenin 3. J Neurochem 97(5):1269–1278.
estan N, Artavanis-Tsakonas S, Rakic P (1999) Contact-dependentinhibition of cortical neurite growth mediated by notch signaling.Science 1999 286(5440):741–746.
herwood NT, Sun Q, Xue M, Zhang B, Zinn K (2004) Drosophilaspastin regulates synaptic microtubule networks and is required fornormal motor function. PLoS Biol 2(12):e429.
hina S, Leberburg I (1999) Cellular mechanisms of beta-amyloidproduction and secretion. Proc Natl Acad Sci U S A 96(20):11049–11053.
rojanowski JQ, Smith AB, Huryn D, Lee VM (2005) Microtubule-stabilising drugs for therapy of Alzheimer’s disease and otherneurodegenerative disorders with axonal transport impairments.
Expert Opin Pharmacother 6(5):683–686.rotta N, Orso G, Rossetto MG, Daga A, Broadie K (2004) Thehereditary spastic paraplegia gene, spastin, regulates microtubulestability to modulate synaptic structure and function. Curr Biol14(13):1135–1147.
erhey KJ, Gaertig J (2007) The tubulin code. Cell Cycle 6(17):2152–2160.
ang Y, Chan SL, Miele L, Yao PJ, Mackes J, Ingram DK, Mattson MP,Furukawa K (2004) Involvement of Notch signaling in hippocampalsynaptic plasticity. Proc Natl Acad Sci U S A 101(25):9458–9462.
ood JD, Landers JA, Bingley M, McDermott CJ, Thomas-McArthur V, Gleadall LJ, Shaw PJ, Cunliffe VT (2006) Themicrotubule-severing protein Spastin is essential for axon out-growth in the zebrafish embryo. Hum Mol Genet 15(18):2763–2771.
u W, Qiang L, Solowska JM, Karabay A, Korulu S, Baas PW (2008)The microtubule-severing proteins spastin and katanin participatedifferently in the formation of axonal branches. Mol Biol Cell
19(4):1485–1498.(Accepted 10 April 2008)(Available online 22 April 2008)
Copyright © 2022 FDOKUMEN




















