role of formin-2 in actin-microtubule coordination during ...
Upload
independentCategory
view
0download
0
www.elsevier.com/locate/cardiores
Cardiovascular Research
httD
ownloaded from
Microtubule-associated protein-4 (MAP-4) inhibits microtubule-dependent
distribution of mRNA in isolated neonatal cardiocytes
Dimitri Scholz a,c,*, Paul McDermott a,c, Maria Garnovskaya b,c, Thomas N. Gallien a,c,
Stefan Huettelmaier d, Christina DeRienzo a,c, George Cooper IV a,c
a Gazes Cardiac Research Institute, Cardiology Division, Medical University of South Carolina, United Statesb Medical and Research Services of the Ralph H. Johnson Veteran Affairs Medical Center, Nephrology Division,
Medical University of South Carolina, United Statesc Department of Veterans Affairs Medical Center, Charleston, SC 29401, United States
d Department of Anatomy and Structural Biology, Albert Einstein College of Medicine, Bronx, NY 10461, United States
Received 3 October 2005; received in revised form 27 March 2006; accepted 9 April 2006
Available online 5 June 2006
Time for primary review 23 days
by guest on June 1p://cardiovascres.oxfordjournals.org/
Abstract
Objectives: Active mRNA distribution in the form of ribonucleoprotein particles moving along microtubules has been shown in several cell
types, but not yet in cardiocytes. This study addresses two hypotheses: 1) a similar mRNA distribution mechanism operates in cardiocytes; 2)
decoration of microtubules with microtubule-associated proteins compromises this distribution.
Methods: To visualize ribonucleoproteins in cultured neonatal rat cardiocytes, they were transfected with vectors encoding zipcode binding
protein-1 and Staufen fused with GFP. The velocity of microtubular transport and elongation were calculated on time-lapse confocal pictures.
Results: ZBP-1 and Staufen labeled particles co-localized with each other and with microtubules and moved along microtubules over a
distance of 1–20 Am with a mean speed of 80 nm/s. The average speed decreased about 50% after decoration of microtubules by adenoviral
microtubule-associated protein-4 (MAP-4). The elongation speed measured using the GFP-tagged end-binding protein-1 exceeded 200 nm/s
and was not influenced by MAP-4.
Conclusions: We demonstrate for the first time ribonucleoprotein particles in cardiocytes, their microtubular-related movement, and its
inhibition (but not of the microtubular elongation), by the MAP-4 decoration of microtubules.
D 2006 European Society of Cardiology. Published by Elsevier B.V. All rights reserved.
3, 2
013Keywords: mRNA; Microtubule; Cardiocyte; Translocation
1. Introduction
Cardiocytes have a high rate of protein turnover, with
proteins of both the myofilament and extramyofilament
cytoskeleton being renewed to the extent of ¨20% per day
[1]. These processes are greatly enhanced during hypertro-
phy [2]. Thus, a consistent supply of translation-competent
mRNA for these structural proteins to the polysomes is
0008-6363/$ - see front matter D 2006 European Society of Cardiology. Publish
doi:10.1016/j.cardiores.2006.04.001
* Corresponding author. Gazes Cardiac Research Institute, Medical
University of South Carolina, 114 Doughty Street, Rm. 302 Charleston,
SC 29403, United States. Tel.: +1 843 876 5067, fax: +1 843 876 5068.
E-mail address: [email protected] (D. Scholz).
needed for normal cellular homeostasis, and it is essential
for hypertrophic growth. However, several unique aspects of
cardiocyte cytoarchitecture challenge this biosynthetic
pathway. In terms of cardiocyte volume, myofibrils are
50–60%, mitochondria are 25–35%, nuclei are 3–5%, the
T-system is 1–2%, and the sarcoplasmic reticulum is 1–3%
[3–5]. ‘‘Free’’ cytoplasm (containing glycogen, ribosomes
and the extramyofilament cytoskeleton) makes up only
¨10%. This high concentration of organelles markedly
limits the free diffusion of macromolecules within these
large cells. Therefore, mRNAs for cardiocyte structural
proteins must be actively delivered to their sites of
translation rather than simply arriving via diffusion.
71 (2006) 506 – 516
ed by Elsevier B.V. All rights reserved.

D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516 507
by guest on June 13, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
In 1993, James Wilhelm and Ron Vale [6] formulated a
hypothesis according to which the transport of mRNAs, and
not of the translated proteins, is responsible for the
localization of cytoplasmic proteins. This model, slightly
modified by Jansen [7], entails the binding of specific
shuttling factors to the so-called ‘‘zip-code’’ sequence of
nascent mRNA in the nucleus [8,9]. The term ‘‘zip code’’ as
a definition for mRNA localization signals within the
3VUTR was proposed by Singer in 1993 [10] and since
then has been broadly employed [7,10–12].
Zip code function is mediated by mRNA-binding proteins
[7,13–18]. The best described and the most promising for
the study of mRNA trafficking in cardiocytes are zipcode-
binding protein-1 (ZBP-1) [10–12,19,20] and Staufen [21–
28]. ZBP-1 was shown to participate in mRNA granule
formation and is needed for mRNA–cytoskeleton attach-
ment [19]. Further, ZBP-1 co-localizes with a h-actintranscription site [20]. Staufen, found also in skeletal muscle
[26], was co-immunoprecipitated with the microtubule-
based + end directed motor protein kinesin in Xenopus
oocytes and was proposed to represent a link between
specific mRNAs and the transportation machinery [29].
The active distribution of mRNA has been described in
several cell types, including ascidian embryos [30], mos-
quito salivary glands [31], Drosophila oocytes [9,32],
Xenopus oocytes [33], rat embryonic neurons [34,35], rat
oligodendrocytes [8], and chicken embryo fibroblasts [19].
The organization of mRNA in ribo-nucleo-protein (RNP)
granules in living cells and their movement along the
cytoskeleton was shown for the first time in dendrites of
mammalian neurons [36], then in oligodendrocytes [37] and
the dendrites of rat hippocampal neurons [38]. RNP
granules have been hypothesized to represent storage
containers for mRNA under translational arrest, which
could be poised for release to actively translated pools
[35]. However, RNP granules and their active distribution
have not been described in cardiocytes.
As one example of active intracellular distribution,
microtubules serve as rails for the transport of vesicles,
cytoplasmic particles [39,40], membrane receptors [41], and
translation-competent mRNAs [6,42–44]. Active, microtu-
bule-based mRNA transport has been described in Dro-
sophila oocytes, yeast, neurons, oligodendrocytes, and
fibroblasts [15,16,22,37,43–45].
A major impetus to this study was our finding that in
pressure-overload cardiac hypertrophy, cardiocytes contain
an increased amount of microtubules, heavily decorated
with and stabilized by the predominant cardiac microtubule-
associated protein, MAP-4 [46], which contributes to cell
viscosity and contractility deficits [47–56]. Depolymeriza-
tion of microtubules restores both the contractility of
cardiocytes and cardiac function to normal [54,56–58].
However, the present study was based not on cardiocyte
mechanics but on the question of whether the extensive
MAP-4 decoration of a dense microtubule network that we
find in pressure-overload cardiac hypertrophy might disrupt
important cellular functions by interfering with microtubule-
based intracellular transport of vesicles and cytoplasmic
particles [6,39,40].
Thus, the aim of present study was to initially test in
neonatal cardiocytes the hypothesis that cardiocyte mRNA
transport is microtubule-dependent and that excessive MAP-
4 decoration of microtubules compromises this transport
function, thereby affecting processes important for the
protein biosynthesis that is essential for the anabolic cellular
response to hemodynamic overloads.
2. Materials and methods
2.1. Ethical statement
This investigation conforms with the Guide for the Care
and Use of Laboratory Animals published by the US
National Institutes of Health (NIH Publication No. 85-23,
revised 1996).
2.2. Animals and isolation of cells
Primary neonatal rat heart cell cultures were prepared from
ventricular myocardium of 0- to 3-day-old rats as described
[59]. Briefly, the cardiocytes were dissociated in (Ca2+–
Mg2+)-free Hank’s salt solution buffered with 30 mM
HEPES, pH 7.4, in a Celstir apparatus (Wheaton Instruments)
at 37 -C by the addition of trypsin (Cooper Biomedical Inc.),
chemotrypsin, and elastase (Sigma) at concentrations of 2.4,
2.7 and 0.94 U/ml, respectively. After each of six successive
20-min incubations, the dissociated cells were mixed with
minimum Eagle’s medium (GIBCO) containing 10% new-
born calf serum, centrifuged at 500�g, and pooled. Cells
were transfected by nucleofection (see below) and plated at a
density of about 5�105 cells in 22 mm (170 Am thick-
bottomed) gelatin-coated WillcoWells\ culture dishes for
live cell microscopy [28]. The majority of cells remained
quiescent in vitro, and only a small percentage was beating at
a low frequency spontaneously or under the laser beam;
these cells were excluded from measurements.
2.3. Plasmids and transfection of cardiocytes
The vector encoding the fusion EYFP–Staufen protein
[21–28] was constructed in the lab of Dr. M. Kiebler, Max-
Planck-Institute for Developmental Biology, Tuebingen,
Germany. The vector encoding the fusion EYFP–ZBP-1
and the ECFP–ZBP-1 proteins [10–12,19,20] were con-
structed in the lab of Dr. R. Singer, Albert Einstein College
of Medicine, NY, USA. The vector encoding the fusion
EGFP–EB-1 protein [60–62] was constructed in the lab of
Dr. Y. Mimori-Kiyosue, Exploratory Research for Advanced
Technology, Japan Science and Technology Corporation,
Kyoto, Japan. The vector encoding the fusion EYFP–a-
tubulin [63] protein was purchased from Clontech.

Fig. 1. Distribution of Staufen–EYFP fusion protein 4 h (A) and 48 h (B)
after nucleofection of neonatal rat cardiocytes. Light diffuse labeling over
the entire cytoplasm corresponds to unbound protein; the nucleus appears as
a dark oval. Granular labeling, most likely representing RNP particles,
appears in the perinuclear region shortly after nucleofection (A) and later
populates the entire cytoplasm (B). Scale bar=10 Am.
D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516508
by guest on June 13, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
A Nucleofectori kit designed for transfection of freshly
isolated primary cardiocytes from newborn rats (Amaxa
Biosystems, Giessen, Germany) was used according to the
manufacturer’s protocol. About 2�106 cells and 2–5 Ag of
plasmid per transfection (single plasmid or double) were
used for each transfection. Up to 10% of cardiocytes were
transfected.
2.4. Adenoviral infection
Adenoviruses expressing bacterial h-gal and human
MAP-4 (Adh-gal and AdMAP-4) were generated in our
laboratory as described [64]. Neonatal rat cardiocytes were
infected in serum-free medium [64] at a multiplicity of
infection (MOI) of about 10 plaque forming units per cell,
thus infecting about 90% of the cardiocytes as determined
by immunofluorescence microscopy using either our poly-
clonal MAP-4 antibody or an X-gal reaction; there was no
observable cytotoxicity.
Preliminary results have shown that MAP-4 appears
mostly diffuse until day 3 after infection, after which it
decorates the microtubules. Therefore, all experiments were
conducted after day 3.
2.5. Immunofluorescent labeling
Neonatal heart cells were fixed in freshly prepared 4%
formaldehyde for 10 min, permeabilized in 0.1% Triton
X-100 and were covered with antibody against MAP-4
and then with the anti-rabbit antiserum tagged with Alexa
Fluor\ 488 (Molecular Probes, Eugene, OR). To generate
the MAP-4 antibody, we made a bacterial expression
construct using the 1–740 NH2-terminal residues of
human MAP-4 [41]. Briefly, the recombinant protein,
which had a hexahistidine tag inserted at the COOH
terminus, was overexpressed in Escherichia coli, purified
on a nickel–chelate affinity column, and submitted to
SDS–PAGE. The purified protein band was excised,
eluted from the gel, and sent to Lampire Biological
Laboratories for preparation of a rabbit polyclonal MAP-4
antibody.
2.6. Microscopy and time lapse photography
A Zeiss LSM510META microscope with Ar-Laser (458,
477, 488 and 514 nm) 30 mW, Plan-Apochromat 63/1.4
objective and a chamber maintaining 37 -C and 5% CO2
was used. The minimal possible laser intensity was chosen
to decrease the thermal damage to cells. Excitation
wavelength and emission filters used for imaging fluores-
cence: CFP-458 nm/BP475-525, GFP-488 nm/LP505, YFP-
514 nm/LP 530. CFP and YFP were visualized simulta-
neously on Figs. 2 and 4.
For Fig. 6, an Olympus IX71 microscope equipped with
a mercury lamp as a light source, and filter sets for DAPI
(Chroma, excitation S403/12, emission S457/50) and for
Alexa488 (Chroma, excitation S490/20, emission D528/38)
have been used.
Time-lapse pictures of about 100 frames gathered over
10 to 25 min without intervals between successive frames
were scanned. Fluorescence-labeled particles moving
straight in at least 3 frames in the row and over a distance
more than 1 Am were considered to be moving actively
and were measured using LSM 5 Image Browser software.
Contracting or laser-damaged cells were excluded from the
study. The movement speed was measured separately for
ZBP-1, Staufen, and EB-1 labeled particles in control and
in MAP-4 or h-gal infected cells for at least 60 particles
per group. Results were analyzed statistically using
Student’s T-test.
2.7. Radioautography
Radioautography was conducted as described [65],
applied here to neonatal cardiocytes. Briefly, cells were
cultured on cover slips in 35 mm culture dishes. 3H-uridine
(10 ACi/ml) was added to the culture medium for 1–6 h.
After that, cells were fixed with formaldehyde, dried in
alcohol and coated with undiluted Kodak NTB2 photo-
graphic emulsion at 42 -C. After 7 days of exposure at 4 -C,they were developed in Kodak D-19 developer as recom-
mended by the manufacturer, counterstained with toluidine
blue and mounted in Entellan.

Fig. 2. Co-localization of ZBP-1 (red) and Staufen (green) in the same granules. Four out of six visible ZBP-1 granules also contain Staufen. A small shift is
caused by color aberration of the lens at the critical magnification. Neonatal rat cardiocytes were double-nucleofected with plasmids encoding the CFP–ZBP-1
and EYFP–Staufen. Scale bar=10 Am.
Fig. 3. Linear movement of a single ZBP-1–EYFP granule (green, arrow) over a distance of about 20 Am. Scale bar=10 Am. A movie of this figure is available
in the Appendix. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516 509
by guest on June 13, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516510
Do
3. Results
3.1. Assembly of RNPs
In order to visualize mRNA-related proteins in rat
neonatal cardiocytes, we transfected freshly isolated cells
with plasmids encoding fluorescently tagged RNP proteins
Staufen and ZBP-1. The diffuse fluorescence became
detectable in the cytoplasm as early as 3–4 h after
transfection and persisted throughout the period of
observation (7 days). This most likely corresponds to
newly synthesized protein which is not yet bound to
mRNA. Fluorescent granules, most likely representing
RNP particles, appeared first in a perinuclear location
(Fig. 1A) and then populated after 24 h the entire
cytoplasm (Fig. 1B).
Fig. 4. Movement of two ZBP-1–EYFP granules (red, arrows) along microtubu
encoding: 1) a-Tubulin–EYFP (green) and 2) ZBP-1–ECFP (red). A–E represent
of this figure is available in the Appendix.
Double transfection of neonatal rat cardiocytes with
plasmids encoding two different and differently labeled
fusion proteins (Staufen–YFP and ZBP-1–CFP) revealed
their co-localization in many instances (Fig. 2).
3.2. Movement of RNPs
Time-lapse pictures were analyzed to measure the speed
of RNP movement. We found that in live cells the vast
majority of labeled particles were oscillating in place, and
only a small subpopulation moved straight over a distance
more than 1 Am, and only rarely up to 10–20 Am (Fig. 3
and Fig. 3 movie in the Appendix). The measured speed
of granules averaged 77.2T6.3 nm/s for ZBP-1 and 76.7T8.5 nm/s for Staufen (meanTS.E.M.). The close corre-
spondence of the transport rates for these two types of
les (green). Neonatal rat cardiocytes were nucleofected with two plasmids
sequential pictures with a time interval of 14 s. Scale bar=10 Am. A movie
by guest on June 13, 2013http://cardiovascres.oxfordjournals.org/
wnloaded from

Fig. 5. Elongation of microtubules. Neonatal rat cardiocytes were
nucleofected with the plasmid encoding the microtubule+end-binding
protein EGFP–EB-1. Scale bar=10 Am. A movie of this figure is available
in the Appendix.
D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516 511
htD
ownloaded from
proteins indicates that they may be transported by the
same cellular motor and (combined with their co-localiza-
tion) suggests indirectly that we may really be dealing
with two proteins being transported as part of the same
RNPs.
Fig. 6. Neonatal rat cardiocytes labeled with anti-MAP-4 antibodies (green)
after AdMAP-4 infection (A) and without infection (B). Nuclei are stained
with DAPI (red). Scale bar=20 Am.
tp://cardiovascres.
3.3. Co-localization of RNP and microtubules and move-
ment along the microtubules
To test our hypothesis that microtubules serve as a
railroad system for RNP movement, microscopic assess-
ment of co-localization of microtubules and RNPs was
undertaken. Cardiocytes double-transfected with the a-
tubulin–EYFP and the ZBP-1–CFP demonstrated consis-
tent co-localization: the RNP granules were localized close
to the microtubules (Fig. 4). In addition, RNP granules
moved along the microtubules from the nucleus towards the
cell periphery, as shown by Fig. 4 and Fig. 4 movie in the
Appendix.
3.4. Microtubule elongation is about 3-fold faster than RNP
transport
Elongation of microtubules in live rat neonatal cardio-
cytes was measured using the GFP-labeled EB-1, which
binds to the + end of growing microtubules (Fig. 5 and Fig.
5 movie in the Appendix). The mean speed of microtubule
elongation was calculated to be 204T6 nm/s, or about three
times as fast as the transport of the RNP particles (Fig. 7).
Thus, the RNP transport kinetics that we have measured are
not a function of the rate of microtubule polymerization-
based elongation.
Fig. 7. Average speed of labeled granules in live neonatal cardiocytes, nm/s,
meanTS.E.M.
by guest on June 13, 2013oxfordjournals.org/
D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516512
http://cardiovascres.oxfoD
ownloaded from
3.5. MAP-4 decoration slows down RNP transport but not
microtubule elongation
MAP-4 decoration of microtubules at day 3 after the
AdMAP-4 infection was documented in cardiocytes by
immunofluorescence using antibodies against MAP-4 (Fig.
6). Non-infected cardiocytes demonstrated no detectable
labeling whereas about 90% of infected cells were positively
labeled, just as we demonstrated before [51]. In AdMAP-4-
treated cells, the mean speed of the ZBP-1 and Staufen-
labeled particles was reduced to about 50% of the control
value (Fig. 7). On the other hand, h-gal infection did not
cause significant changes.
In contrast, the mean speed of the EB-1-labeled particles
in AdMAP-4-infected cells did not change (204T6 nm/s
control vs. 227T10 nm/s AdMAP-4, n.s.).
3.6. Radioautographic dispersion of RNA
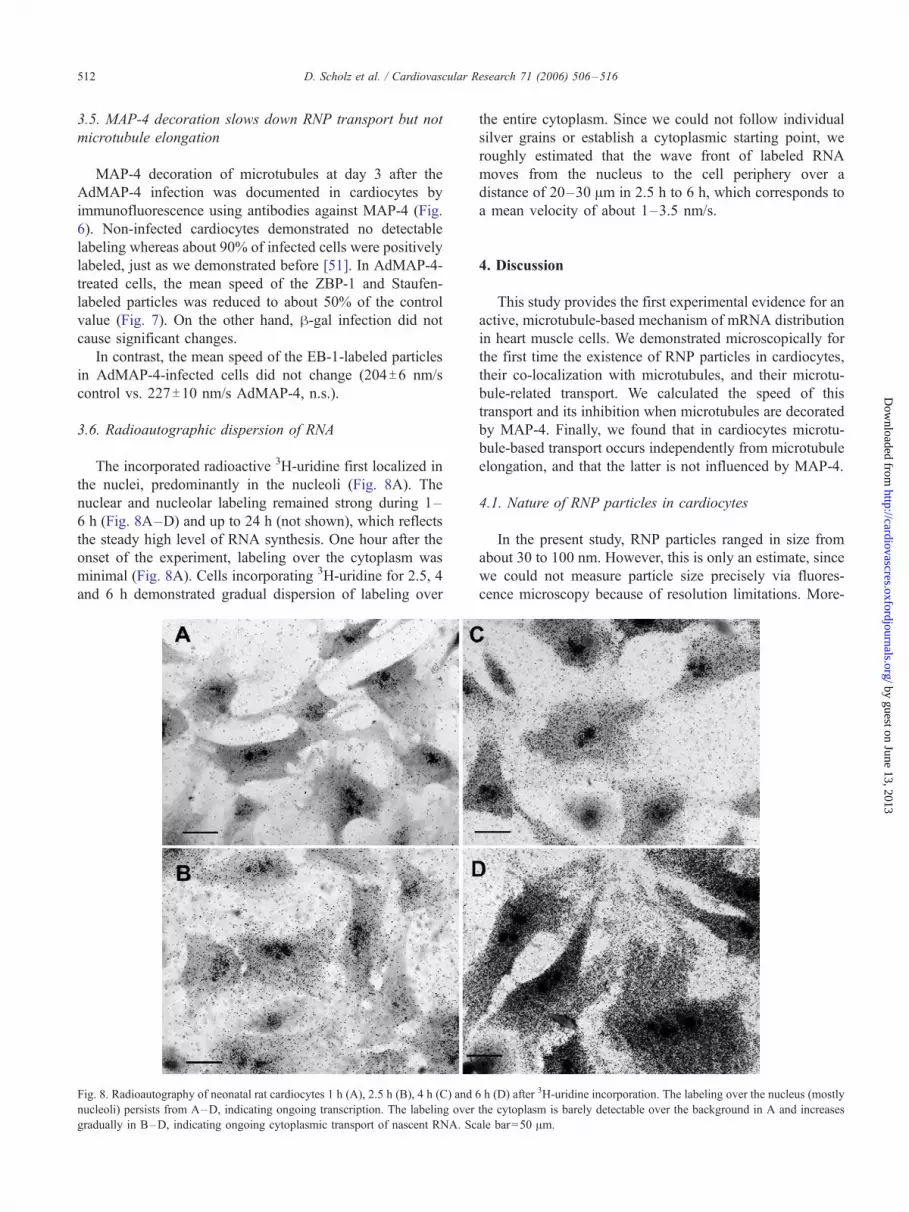
The incorporated radioactive 3H-uridine first localized in
the nuclei, predominantly in the nucleoli (Fig. 8A). The
nuclear and nucleolar labeling remained strong during 1–
6 h (Fig. 8A–D) and up to 24 h (not shown), which reflects
the steady high level of RNA synthesis. One hour after the
onset of the experiment, labeling over the cytoplasm was
minimal (Fig. 8A). Cells incorporating 3H-uridine for 2.5, 4
and 6 h demonstrated gradual dispersion of labeling over
Fig. 8. Radioautography of neonatal rat cardiocytes 1 h (A), 2.5 h (B), 4 h (C) and 6
nucleoli) persists from A–D, indicating ongoing transcription. The labeling over
gradually in B–D, indicating ongoing cytoplasmic transport of nascent RNA. Sc
the entire cytoplasm. Since we could not follow individual
silver grains or establish a cytoplasmic starting point, we
roughly estimated that the wave front of labeled RNA
moves from the nucleus to the cell periphery over a
distance of 20–30 Am in 2.5 h to 6 h, which corresponds to
a mean velocity of about 1–3.5 nm/s.
4. Discussion
This study provides the first experimental evidence for an
active, microtubule-based mechanism of mRNA distribution
in heart muscle cells. We demonstrated microscopically for
the first time the existence of RNP particles in cardiocytes,
their co-localization with microtubules, and their microtu-
bule-related transport. We calculated the speed of this
transport and its inhibition when microtubules are decorated
by MAP-4. Finally, we found that in cardiocytes microtu-
bule-based transport occurs independently from microtubule
elongation, and that the latter is not influenced by MAP-4.
4.1. Nature of RNP particles in cardiocytes
In the present study, RNP particles ranged in size from
about 30 to 100 nm. However, this is only an estimate, since
we could not measure particle size precisely via fluores-
cence microscopy because of resolution limitations. More-
h (D) after 3H-uridine incorporation. The labeling over the nucleus (mostly
the cytoplasm is barely detectable over the background in A and increases
ale bar=50 Am.
by guest on June 13, 2013rdjournals.org/

D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516 513
by guest on June 13, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
over, overexposed particles appear larger, and some studies
have reported RNP particle sizes of up to 500 nm [34] or
even 700 nm [66]. Electron microscopy studies [67]
described nuclear RNP particles of 20 to 50 nm. However,
electron microscopy data on RNP particles outside the
nucleus [22] have described them as loose and amorphous
particles associated with polyribosomes, about 200 nm in
diameter. While Oleynikov and Singer described ZBP-1
shuttling between nucleus and cytoplasm in chicken embryo
fibroblasts [20], we could not detect any nuclear ZBP-1 or
Staufen labeling.
Pre-RNP exported through the nuclear pores to the
cytoplasm [68,69] matures into RNP particle binding
proteins, recently classified [70] into 1) RNA-binding
proteins that associate with specific mRNAs such as Staufen
[42], ZBP-1 [12], and Vera [71]; 2) motor proteins that
associate with RNPs to transport them, such as dynein [45]
and kinesin [72]; 3) adaptor proteins that interact with other
elements of RNPs and are essential for their transport, such
as Barentsz [27,73], She3p [74] and Miranda [75]; 4)
cytoskeletal structures along which RNPs move, such as
microtubules [42] and actin thin filaments [76]; and 5)
specific repressors of protein synthesis that prevent trans-
lation before the target mRNA is properly localized, such as
Bruno [77]. Obviously, after passing the nuclear pore
bottleneck, tens or hundreds of particles may come together
and bind additional regulatory and transport proteins and
ribosomes [78]. They thereby grow about 10-fold in size
(which corresponds to a 1000-fold gain in volume), bind to
the microtubules, and move to their destination where the
translation may finally begin [79].
4.2. Speed of RNP movement
Twenty years ago, Lasek et al. [80] differentiated fast
(0.5–4 Am/s) axonal transport of vesicles from slow (10–
50 nm/s) transport of cytoplasmic proteins based solely on
radiolabeling, the only technique available at that time. The
measurements for mRNA showed even slower movement
than that for proteins [65].
Table 1
Literature data for the velocity of microtubule transport for RNP and other non-v
Speed (nm/s) Reference Species, tissue
0.6 [65] Rat embryonic hippocam
1.0–1.2 [87] Rat embryonic hippocam
1.17 [88] Xenopus oocyte
200 [6] Myelin basic protein (M
100–1000 [37] Mouse oligodendocytes
100 [34] Rat embryonic cortical n
200–400 [89] Yeast
70 [82] HeLa cells
500–1000 [39] Squid axon
600 [20] Fibroblasts, ZBP-1
540 [90] Human U2OS osteosarc
300–2000 [91] CHO cells
Direct measurements with fluorescent microscopy.
Since fluorescence study of live cells after microinjection
of labeled mRNA [37], labeling with mRNA-dyes [34] or
transfection with fluorescent fusion proteins [23,81] became
available, many investigators have followed intracellular
movement directly and calculated the speed; the results are
summarized in Table 1.
In the present study, we estimated an average speed of
RNPs in the cytoplasm of about 80 nm/s, which corresponds
closely to the literature data for RNP transport in neurons,
oligodendocytes and fibroblasts [20,34,37,38,82].
In cells infected with AdMAP-4, the mean speed of RNP
particles was significantly reduced. We excluded the
influence of adenoviral infection by using Adh-gal as a
negative control and suggest that the reason for the speed
reduction by MAP-4 could be the competition of the
molecular motor with the MAP for binding sites on
microtubules.
In contrast, the mean speed of microtubule elongation
was almost 3-fold higher, very close to that described
for HeLa cells [83], and it was MAP-4-independent.
Thus, microtubule elongation and microtubule-dependent
transport of RNPs are based on different molecular
mechanisms.
At any time point the majority of particles do not exhibit
net movement but instead oscillate in place. Similar
behavior of RNP-labeled granules using GFP–ZBP-1 or
GFP–Staufen has been reported in chicken embryo fibro-
blasts [20], chicken neurons [84], and rat hippocampal
neurons [23]. This transport pattern could be explained by
saltatory motion or by very short-term movement of
intracellular cargos along microtubules [85]. Further, RNPs
have been shown not to be bound to motor proteins or
microtubules most of the time [86].
To approximate the rate of movement of RNA pools over
a longer period of time, we used radiolabeling, taking into
account that the measurement could be contaminated by
rRNA movement but hoping that rRNA might be trans-
ported in the same RNP particles [22]. We roughly
estimated the mean velocity of newly synthetisized RNA
in cytoplasm to be about 1–3 nm/s or 20 to 80 times less
esicle cytoplasmic particles
Comments
pal neurons 3H-uridine incorporation
pal neurons EGFP–ZBP-1
BP) mRNA
eurons Prevented by microtubule depolymerization
slow plus-end-directed mitotic kinesin motor
oma cells Intranuclear
Vesicles, short measurements

D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516514
than that measured by our fluorescence method, which
probably means that at any given time only one out of 20 to
80 granules is moving. This result is consistent with our
observation by fluorescence microscopy that at any given
time only a small fraction of the particles show substantial
vectorial movement.
5. Conclusions
We demonstrate here for the first time RNP particles in
cardiocytes, their microtubule-related movement, and the
inhibition of this movement, but not of microtubule
elongation, by MAP-4 microtubule decoration.
http://cardiovascres.oxfordjournals.D
ownloaded from
Acknowledgments
This study was supported by Program Project Grant HL-
48788 from the National Heart, Lung, and Blood Institute
and by a Merit Award from the Research Service of the
Department of Veterans Affairs. We thank Dr. Michael
Kiebler and Dr. Bernhard Goetze from Max Planck Institute
for Developmental Biology, Tuebingen 72076, Germany
and Dr. Yuko Mimori-Kiyosue from Tsukita Cell Axis
Project, Exploratory Research for Advanced Technology,
Japan Science and Technology Corporation, Kyoto Re-
search Park, Shimogyo-ku, Kyoto 600-8813, Japan, for
providing vectors and for the fruitful critical discussion and
comments during the manuscript preparation.
by guest on June 13, 2org/
Appendix A. Supplementary data
Supplementary data associated with this article can be
found, in the online version, at doi:10.1016/j.cardiores.
2006.04.001.
013
References
[1] Simpson D, Decker M, Clark W, Decker R. Contractile activity and
cell –cell contact regulate myofibrillar organization in cultured cardiac
myocytes. J Cell Biol 1993;123:323–36.
[2] Imamura T, McDermott PJ, Kent RL, Nagatsu M, Cooper G IV,
Carabello BA. Acute changes in myosin heavy chain synthesis rate in
pressure versus volume overload. Circ Res 1994;75:418–25.
[3] Page E, McCallister LP. Quantitative electron microscopic description
of heart muscle cells. Application to normal, hypertrophied and
thyroxin-stimulated hearts. Am J Cardiol 1973;31:172–81.
[4] Anversa P, Loud AV, Vitali-Mazza L. Morphometry and autoradiog-
raphy of early hypertrophic changes in the ventricular myocardium of
adult rat: an electron microscopic study. Lab Invest 1976;35:475–83.
[5] Rumyantsev PP. Cardiomyocytes in the reproduction, differentiation
and gegeneration. Leningrad’ Nauka; 1982 [in Russian].
[6] Wilhelm JE, Vale RD. RNA on the move: the mRNA localization
pathway. J Cell Biol 1993;123:269–74.
[7] Jansen R. mRNA localization: message on the move. Nat Rev Mol
Cell Biol 2001;2:247–56.
[8] Hoek K, Kidd G, Carson JRS. hnRNP A2 selectively binds the
cytoplasmic transport sequence of myelin basic protein mRNA.
Biochemistry 1998;37:7021–9.
[9] Norvell A, Kelley R, Wehr KTS. Specific isoforms of squid, a
Drosophila hnRNP, perform distinct roles in Gurken localization
during oogenesis. Genes Dev 1999;13:864–76.
[10] Singer R. RNA zipcodes for cytoplasmic addresses. Curr Biol
1993;3:719–21.
[11] Oleynikov Y, Singer RH. RNA localization: different zipcodes, same
postman? Trends Cell Biol 1998;8:381–3.
[12] Ross AF, Oleynikov Y, Kislauskis EH, Taneja KL, Singer RH.
Characterization of a h-actin mRNA zipcode-binding protein. Mol
Cell Biol 1997;17:2158–65.
[13] Kislauskis E, Li Z, Singer R, Taneja K. Isoform-specific 3V-untranslated sequences sort a-cardiac and h-cytoplasmic actin
messenger RNAs to different cytoplasmic compartments. J Cell Biol
1993;123:165–72.
[14] Bashirullah A, Cooperstock R, Lipshitz H. RNA localization in
development. Annu Rev Biochem 1998;67:335–94.
[15] Bassell G, Oleynikov Y, Singer R. The travels of mRNAs through all
cells large and small. FASEB 1999;13:447–54.
[16] Farina K, Singer R. The nuclear connection in RNA transport and
localization. Trends Cell Biol 2002;12:466–72.
[17] Roegiers F. Insights into mRNA transport in neurons. Proc Natl Acad
Sci 2002;100:1465–6.
[18] Jockusch B, Huttelmaier S, Illenberger S. From the nucleus toward the
cell periphery: a guided tour for mRNAs. News Physiol Sci
2003;18:7–11.
[19] Farina K, Huttelmaier S, Musunuru K, Darnell R, Singer R. Two
ZBP1 KH domains facilitate h-actin mRNA localization, granule
formation, and cytoskeletal attachment. J Cell Biol 2003;160:77–87.
[20] Oleynikov Y, Singer R. Real-time visualization of ZBP1 association
with. Curr Biol 2003;13:199–207.
[21] Wickham L, Duchaine T, Luo M, Nabi IR, DesGroseillers L.
Mammalian Staufen is a double-stranded-RNA- and tubulin-binding
protein which localizes to the rough endoplasmic reticulum. Mol Cell
Biol 1999;19:2220–30.
[22] Kiebler MA, Hemraj I, Verkade P, Kohrmann M, Fortes P, Marion
RM, et al. The mammalian Staufen protein localizes to the
somatodendritic domain of cultured hippocampal neurons: implica-
tions for its involvement in mRNA transport. J Neurosci 1999;19:
288–97.
[23] Kohrmann M, Luo M, Kaether C, DesGroseillers L, Dotti CG, Kiebler
MA. Microtubule-dependent recruitment of Staufen–green fluores-
cent protein into large RNA-containing granules and subsequent
dendritic transport in living hippocampal neurons. Mol Biol Cell
1999;10:2945–53.
[24] Duchaine TF, Hemraj I, Furic L, Deitinghoff A, Kiebler MA,
DesGroseillers L. Staufen2 isoforms localize to the somatodendritic
domain of neurons and interact with different organelles. J Cell Sci
2002;115:3285–95.
[25] Martin S, Leclerc V, Smith-Litiere K, D.S.J.. The identification of
novel genes required for Drosophila anteroposterior axis formation in
a germline clone screen using GFP–Staufen. Development
2003;130:4201–15.
[26] Belanger G, Stocksley MA, Vandromme M, Schaeffer L, Furic L,
DesGroseillers L, et al. Localization of the RNA-binding proteins
Staufen1 and Staufen2 at the mammalian neuromuscular junction. J
Neurochem 2003;86:669–77.
[27] Macchi P, Kroening S, Palacios IM, Baldassa S, Grunewald B,
Ambrosino C, et al. Barentsz, a new component of the Staufen-containing
ribonucleoprotein particles in mammalian cells, interacts with Staufen in
an RNA-dependent manner. J Neurosci 2003;23:5778–88.
[28] Mallardo M, Deitinghoff A, Muller J, Goetze B, Macchi P, Peters C,
et al. Isolation and characterization of Staufen-containing ribonucleo-
protein particles from rat brain. Proc Natl Acad Sci U S A 2003;100:
2100–5.

D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516 515
by guest on June 13, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
[29] Yoon Y, Mowry K. Xenopus Staufen is a component of a
ribonucleoprotein complex containing Vg1 RNA and kinesin. Devel-
opment 2004;131:3035–45.
[30] Jeffery W, Tomlinson C, Brodeur R. Localization of actin messenger
RNA during early ascidian development. Dev Biol 1983;99:408–17.
[31] Mehlin H, Daneholt B, Skoglund U. Translocation of a specific
premessenger ribonucleoprotein particle through the nuclear pore
studied with electron microscope tomography. Cell 1992;69:605–13.
[32] Frigerio G, Burri M, Bopp D, Baumgartner S, Noll M. Structure of the
segmentation gene paired and the Drosophila PRD gene set as part of
a gene network. Cell 1986;47:735–46.
[33] Rebagliati M, Weeks D, Harvey R, Melton D. Identification and
cloning of localized maternal RNAs from Xenopus eggs. Cell
1985;42:769–77.
[34] Knowles R, Sabry J, Martone M, Deerinck TJ, Ellisman MH, Bassell
GJ, et al. Translocation of RNA granules in living neurons. J Neurosci
1996;16:7812–20.
[35] Krichevsky A, Kosik K. Neuronal RNA granules: a link between RNA
localization and stimulation-dependent translation. Neuron
2001;32:683–96.
[36] Tiedge H, Fremeau RJ, Weinstock P, Arancio O, Brosius J. Dendritic
location of neural BC1 RNA. Proc Natl Acad Sci U S A 1991;88:
2093–7.
[37] Ainger K, Avossa D, Morgan F, Hill SJ, Barry C, Barbarese E, et al.
Transport and localization of exogenous myelin basic protein mRNA
microinjected into oligodendrocytes. J Cell Biol 1993;123:431–41.
[38] Fredj N, Grange J, Sadoul R, Richard S, Goldberg Y, V.B..
Depolarization-induced translocation of the RNA-binding protein
Sam68 to the dendrites of hippocampal neurons. J Cell Sci 2004;
117:1079–90.
[39] Prahlad V, Helfand BT, Langford GM, Vale RD, Goldman RD. Fast
transport of neurofilament protein along microtubules in squid
axoplasm. J Cell Sci 2000;113:3939–46.
[40] Vale RD. The molecular motor toolbox for intracellular transport. Cell
2003;112:467–80.
[41] Cheng G, Iijima Y, Ishibashi Y, Kuppuswamy D, Cooper G IV.
Inhibition of G protein-coupled receptor trafficking in neuroblastoma
cells by MAP-4 decoration of microtubules. Am J Physiol Heart Circ
Physiol 2002;283:H2379.
[42] Ferrandon D, Elphick L, Nusslein-Volhard C, St Johnston D.
Staufen protein associates with the 3VUTR of bicoid mRNA to form
particles that move in a microtubule-dependent manner. Cell 1994;
79:1221–32.
[43] Carson J, Worboys K, Ainger K, Barbarese E. Translocation of myelin
basic protein mRNA in oligodendrocytes requires microtubules and
kinesin. Cell Motil Cytoskeleton 1997;38:318–28.
[44] Carson J, Kwon S, Barbarese E. RNA trafficking in myelinating cells.
Curr Opin Neurobiol 1998;8:607–12.
[45] Schnorrer F, Bohmann K, Nusslein-Volhard C. The molecular
motor dynein is involved in targeting swallow and bicoid RNA to
the anterior pole of Drosophila oocytes. Nat Cell Biol 2000;2:
185–90.
[46] Takahashi M, Shiraishi H, Ishibashi Y, Blade KL, McDermott PJ,
Menick DR, et al. Phenotypic consequences of h1-tubulin expression
and MAP4 decoration of microtubules in adult cardiocytes. Am J
Physiol Heart Circ Physiol 2003;285:H2072.
[47] Tsutsui H, Ishihara K, Cooper G IV. Cytoskeletal role in the contractile
dysfunction of hypertrophied myocardium. Science 1993;260:682–7.
[48] Tsutsui H, Tagawa H, Kent RL, McCollam PL, Ishihara K, Nagatsu
M, et al. Role of microtubules in contractile dysfunction of
hypertrophied cardiocytes. Circulation 1994;90:533–55.
[49] Tagawa H, Rozich JD, Tsutsui H, Narishige T, Kuppuswamy D, Sato
H, et al. Basis for increased microtubules in pressure-hypertrophied
cardiocytes. Circulation 1996;93:1230–43.
[50] Tagawa H, Wang N, Narishige T, Ingber DE, Zile MR, Cooper G IV.
Cytoskeletal mechanics in pressure-overload cardiac hypertrophy. Circ
Res 1997;80:281–9.
[51] Sato H, Nagai T, Kuppuswamy D, Narishige T, Koide M, Menick DR,
et al. Microtubule stabilization in pressure overload cardiac hypertro-
phy. J Cell Biol 1997;139:963–73.
[52] Koide M, Nagatsu M, Zile MR, Hamawaki M, Swindle MM, Keech
G, et al. Premorbid determinants of left ventricular dysfunction in a
novel model of gradually induced pressure overload in the adult
canine. Circulation 1997;95:1601–10.
[53] Zile MR, Richardson K, Cowles MK, Buckley JM, Koide M, Cowles
BA, et al. Constitutive properties of adult mammalian cardiac muscle
cells. Circulation 1998;98:567–79.
[54] Zile MR, Koide M, Sato H, Ishiguro Y, Conrad CH, Buckley JM, et al.
Role of microtubules in the contractile dysfunction of hypertrophied
myocardium. J Am Coll Cardiol 1999;33:250–60.
[55] Koide M, Hamawaki M, Narishige T, Sato H, Nemoto S, DeFreyte G,
et al. Microtubule depolymerization normalizes in vivo myocardial
contractile function in dogs with pressure-overload left ventricular
hypertrophy. Circulation 2000;102:1045–52.
[56] Schroder EA, Tobita K, Tinney JP, Foldes JK, Keller BB. Microtubule
involvement in the adaptation to altered mechanical load in developing
chick myocardium. Circ Res 2002;91:353–9.
[57] Cicogna AC, Robinson KG, Conrad CH, Singh K, Squire R, Okoshi
MP, et al. Direct effects of colchicine on myocardial function: studies
in hypertrophied and failing spontaneously hypertensive rats. Hyper-
tension 1999;33:60–5.
[58] Roos KP, Palmer RE, Miller TW. The role of microtubules in
structural remodeling and the progression to heart failure. J Card Fail
2002;8:S300.
[59] McDermott PJ, Morgan HE. Contraction modulates the capacity for
protein synthesis during growth of neonatal heart cells in culture. Circ
Res 1989;64:542–53.
[60] Mimori-Kiyosue Y, Shiina N, Tsukita S. The dynamic behavior of the
APC-binding protein EB1 on the distal ends of microtubules. Curr
Biol 2000;10:865–8.
[61] Mimori-Kiyosue Y, Tsukita S. ‘‘Search-and-capture’’ of microtubules
through plus-end-binding proteins (+TIPs). J Biochem (Tokyo)
2003;134:321–6.
[62] Piehl M, Tulu US, Wadsworth P, Cassimeris L. Centrosome
maturation: measurement of microtubule nucleation throughout the
cell cycle by using GFP-tagged EB1. Proc Natl Acad Sci U S A
2004;101:1584–8.
[63] Kimble M, Kuzmiak C, McGovern KN, de Hostos EL. Microtubule
organization and the effects of GFP–tubulin expression in Dictyoste-
lium discoideum. Cell Motil Cytoskeleton 2000;47:48–62.
[64] Cheng G, Qiao F, Gallien T, Kuppuswamy D, Cooper G IV. Inhibition
of h-adrenergic receptor trafficking in adult cardiocytes by MAP4
decoration of microtubules. Am J Physiol Heart Circ Physiol 2005;
288:H1193–202.
[65] Davis L, Banker G, Steward O. Selective dendritic transport of RNA
in hippocampal neurons in culture. Nature 1987;330:477–9.
[66] Barbarese E, Koppel D, Deutscher M, Smith CL, Ainger K, Morgan F,
et al. Protein translation components are colocalized in granules in
oligodendrocytes. J Cell Sci 1995;108:2781–90.
[67] Stevens BJ, Swift H. RNA transport from nucleus to cytoplasm in
Chironomus salivary glands. J Cell Biol 1966;31:55–77.
[68] Daneholt B. Look at messenger RNP moving through the nuclear
pore. Cell 1997;88:585–8.
[69] Levesque L, Guzik B, Guan T, Coyle J, Black BE, Rekosh D, et al.
RNA export mediated by tap involves NXT1-dependent interac-
tions with the nuclear pore complex. J Biol Chem 2001;276:
44953–62.
[70] Villace P, Marion RM, Ortin J. The composition of Staufen-containing
RNA granules from human cells indicates their role in the regulated
transport and translation of messenger RNAs. Nucleic Acids Res
2004;32:2411–20.
[71] Deshler J, Highett M, Schnapp B. Localization of Xenopus Vg1
mRNA by Vera protein and the endoplasmic reticulum. Science
1997;276:1128–31.

D. Scholz et al. / Cardiovascular Research 71 (2006) 506–516516
http://cardiovascresD
ownloaded from
[72] Brendza R, Serbus L, Duffy J, Saxton W. A function for kinesin I in
the posterior transport of oskar mRNA and Staufen protein. Science
2000;289:2120–2.
[73] van Eeden F, Palacios I, Petronczki M, Weston M, St Johnston D.
Barentsz is essential for the posterior localization of oskar mRNA and
colocalizes with it to the posterior pole. J Cell Biol 2001;154:511–23.
[74] Takizawa PA, Vale RD. The myosin motor, Myo4p, binds Ash1
mRNA via the adapter protein, She3p. Proc Natl Acad Sci U S A
2000;97:5273–8.
[75] Shen C, Knoblich J, Chan Y, Jiang M, Jan L, Jan Y. Miranda as a
multidomain adapter linking apically localized Inscuteable and basally
localized Staufen and Prospero during asymmetric cell division in
Drosophila. Genes Dev 1998;12:1837–46.
[76] Takizawa PA, Sil A, Swedlow JR, Herskowitz I, Vale RD. Actin-
dependent localization of an RNA encoding a cell-fate determinant in
yeast. Nature 1997;389:90–3.
[77] Kim-Ha J, Kerr K, Macdonald P. Translational regulation of oskar
mRNA by Bruno, an ovarian RNA-binding protein, is essential. Cell
1995;81:403–12.
[78] Brendel C, Rehbein M, Kreienkamp H, Buck F, Richter D, Kindler S.
Characterization of Staufen 1 ribonucleoprotein complexes. Biochem J
2004;384:239–46.
[79] Huettelmaier S, Zenklusen D, Lederer M, Dictenberg J, Lorenz M,
Meng X, et al. Spatial regulation of h-actin translation by Src-
dependent phosphorylation of ZBP1. Nature 2005;438:512–5.
[80] Lasek R, Garner J, Brady S. Axonal transport of the cytoplasmic
matrix. J Cell Biol 1984;99:212s–21s.
[81] Chalfie M, Tu Y, Euskirchen G, Ward W, Prasher D. Green fluorescent
protein as a marker for gene expression. Science 1994;263:802–5.
[82] Abaza A, Soleilhac JM, Westendorf J, Piel M, Crevel I, Roux A, et al.
M phase phosphoprotein 1 is a human plus-end-directed kinesin-
related protein required for cytokinesis. J Biol Chem 2003;278:
27844–52.
[83] Perez F, Diamantopoulos G, Stalder R, Kreis T. CLIP-170 highlights
growing microtubule ends in vivo. Cell 1999;96:517–27.
[84] Zhang HL, Eom T, Oleynikov Y, Shenoy SM, Liebelt DA, Dictenberg
JB, et al. Neurotrophin-induced transport of a h-actin mRNP complex
increases h-actin levels and stimulates growth cone motility. Neuron
2001;31:261–75.
[85] Sheetz MP. Motor and cargo interactions. Eur J Biochem 1999;262:
19–25.
[86] Kloc M, Etkin L. RNA localization mechanisms in oocytes. J Cell Sci
2005;118:269–82.
[87] Tiruchinapalli DM, Oleynikov Y, Kelic S, Shenoy SM, Hartley A,
Stanton PK, et al. Activity-dependent trafficking and dynamic
localization of zipcode binding protein 1 and h-actin mRNA in
dendrites and spines of hippocampal neurons. J Neurosci 2003;23:
3251–61.
[88] Yisraeli J, Sokol S, Melton D. A two-step model for the localization of
maternal mRNA in Xenopus oocytes: involvement of microtubules
and microfilaments in the translocation and anchoring of Vg1 mRNA.
Development 1990;108:289–98.
[89] Bertrand E, Chartrand P, Schaefer M, Shenoy SM, Singer RH, Long
RM. Localization of ASH1 mRNA particles in living yeast. Mol Cell
1998;2:437–45.
[90] Shav-Tal Y, Darzacq X, Shenoy SM, Fusco D, Janicki SM, Spector
DL, et al. Dynamics of single mRNPs in nuclei of living cells. Science
2004;304:1797–800.
[91] Mundy D, Machleidt T, Ying Y, Anderson R, Bloom G. Dual control
of caveolar membrane traffic by microtubules and the actin cytoskel-
eton. J Cell Sci 2002;115:4327–39.
.ox
by guest on June 13, 2013fordjournals.org/
Copyright © 2022 FDOKUMEN