
Reliability in endoscopic diagnosis of portal hypertensive gastropathy

EXPERIMENTAL RESEARCH
Microsurgical and endoscopic anatomy of Liliequist"smembrane and the prepontine membranes: cadaveric studyand clinical implications
Ihsan Anik & Savas Ceylan & Kenan Koc &
Mehtap Tugasaygi & Gozde Sirin & Nurperi Gazioglu &
Bulent Sam
Received: 21 December 2010 /Accepted: 15 February 2011 /Published online: 6 March 2011# Springer-Verlag 2011
AbstractBackground Liliequist’s membrane is mostly described ashaving a diencephalic leaf, mesencephalic leaf, anddiencephalic-mesencephalic leaves in the literature. Alsodifferent descriptions of the prepontine membranes werereported. In this study, we visualized the regular structuralforms of membranes without disturbing any attachmentsand defined infrachiasmatic and prepontine safety zones.We discussed the clinical significance of these structures.Materials and methods The study was carried out on 24adult human cadavers at the Morgue Specialization Depart-ment of the Forensic Medicine Institution following theinitial autopsy examination. Liliequist"s membrane and theprepontine membranes were explored after retraction of thefrontal lobes. Dissections were performed under theoperative microscope. A 0- and 30-degree, 2.7-mm angledrigid endoscope (Aesculap, Tuttlingen, Germany) wasadvanced through the prepontine cistern from the naturalholes of membranes, or small holes were opened withoutdamaging the surrounding structures.Results The basal arachnoid membrane (BAM) continuedas Liliequist"s membrane (LM) without any distinct sepa-
ration in all specimens. The LM coursed over theposterior clinoids and split into two leaves as thediencephalic leaf (DL) and mesencephalic leaf (ML) in18 specimens; the medial pontomesencephalic membrane(MPMM) coursed anterolaterally as a continuation of theML and attached to the medial surfaces of the fifth andsixth nerves, joining with the lateral pontomesencephalicmembrane (LPMM), which was also a posterolateralcontinuation of the ML in all specimens. The medialpontomedullar membrane (MPMdM) and lateral ponto-medullar membrane (LPMdM) were observed in 21specimens. The MPMdM membrane was a continuationof the MPMM, and the LPMdM was a continuation ofthe LPMM in all 21 specimens.Conclusion We observed that the LM is a borderlesscontinuation of the BAM. The MPMM and LPMM splitfrom the ML without any interruptions. The MPMdM andLPMdM were a single membrane continuing from theMPMM and LPMM. We determined infrachiasmatic andprepontine areas that can be important for inferior surgicalapproaches.
Keywords Liliequist membrane . Endoscope . Skull base .
Prepontine
Introduction
The anatomy of Liliequist"s membrane (LM) and themembranous structures in the prepontine structures is ofessential importance in minimally invasive neurosurgery.The importance of these anatomic structures has in-creased with the introduction of neuroendoscopic andendoscopic transphenoidal procedures [13, 28, 29]. An
I. Anik : S. Ceylan (*) :K. KocDepartment of Neurosurgery, Kocaeli University,School of Medicine,41380 Umuttepe, Kocaeli, Turkeye-mail: [email protected]
M. Tugasaygi :G. Sirin :N. Gazioglu :B. SamNational Forensic Institute of Ministry of Justice,Istanbul, Turkey
N. GaziogluCerrahpasa School of Medicine, Istanbul University,Istanbul, Turkey
Acta Neurochir (2011) 153:1701–1711DOI 10.1007/s00701-011-0978-5

endoscopic approach through the cisterns requires detailedknowledge of the BAM, LM and prepontine cisternanatomy [13, 23, 28].
Magendie [21] first described the arachnoid mater as aspecific layer of the meninges and arachnoidal cisterns in1842. Key and Retzius [16] described the arachnoid materand cisterns, and illustrated a distinct arachnoid membranethat divided the interpeduncular cistern into deep andsuperficial portions. The cisternal anatomy, which iscomprised of the arachnoid structures, is described in theliterature [15, 22, 24, 33].
In the literature, the LM is mostly defined as a group ofanatomically distinct arachnoid sheets [13, 17]: a dience-phalic leaf (DL), a mesencephalic leaf (ML) and a pair ofdiencephalic-mesencephalic leaves (DML). Differentdescriptions of the lateral pontomesencephalic membrane(LPMM),medial pontomesencephalic membrane (MPMM),medial pontomedullar membrane (MPMdM) and lateralpontomedullar membrane (LPMdM) in the prepontinecistern and their connections forming the cisterns havebeen described in the literature.
Several authors [27] described the basal arachnoidmembrane (BAM), which is important especially inendoscopic skull base surgery, and recently Wang et al.[32] defined the hypothalamic membrane in addition to theother membranes.
In our cadaveric study, we determined the variations ofLM, BAM and membranous structures located in theprepontine cistern. We discussed the clinical significanceof our findings for endoscopic skull-base procedures andendoscopic third ventriculostomy.
Materials and methods
We studied 24 adult human cadavers in the MorgueSpecialization Department of the Forensic Medicine Insti-tution in Istanbul following the initial autopsy examinationand approval of the forensic medicine team. Age of 18 yearsor older and no history of head trauma and/or cranialsurgery were the selection criteria for the cadavers.Liliequist"s membrane and the prepontine membranes wereexplored after retraction of the frontal lobes. Dissectionswere performed under the operative microscope, and a 0-and 30-degree, 2.7-mm angled rigid endoscope (Aesculap,Tuttlingen, Germany) was advanced through the prepontinecistern from the natural holes of membranes, or small holeswere opened without damaging the surrounding structures.This allowed us to visualize the regular structural forms ofmembranes without disturbing any attachments. Surgicalprocedures were recorded with a digital video camerasystem.
Results
Basal arachnoid membrane The BAM arose from thesuperior wall of the cavernous sinus and optic canalscovering the tuberculum sellae and anterior clinoid process,and then continued as the LM without any distinctseparation in all specimens (Fig. 1). Arachnoid sleevesarose from the BAM to chiasmatic cistern and constitutedthe anterior wall of the chiasmatic cistern (Fig. 2a). Thepituitary stalk coursed to the pituitary gland, penetrating theBAM.
Liliequist’s membrane The LM coursed over the posteriorclinoids and split into two leaves as the DL and ML in 18specimens, separated the DL and ML only in 3 specimensand coursed as the ML in 3 specimens. These leaves of themembrane attached to the corpus mamillare at the posteriorinferior edges, basilar artery bifurcation anterior-inferiorly,extended to the tentorial edges anterosuperolaterally andattached to the medial uncus inferolaterally (Fig. 1).
Diencephalic leaf The DL was observed in 21 specimens,but not in 3. It had a trabecular form in 4 specimens, wasthick in 13 and opaque in 4 (Fig. 2b).
Posterior projections
The DL coursed posterosuperiorly towards the diencepha-lon, extended upwards and attached to the floor of the thirdventricle at the mammillary bodies. The posterior edge ofthe DL was attached to the mammillary bodies directly in17 specimens and attached to the pia of the diencephalonbetween the infundibulum and mammillary bodies in 4specimens.
Lateral projections
The DL coursed laterally covering the medial third nerve in14 specimens and gave rise to arachnoid trabeculations ofthe third nerve in 7 specimens. In three specimens the DLwas not observed; thus, the LM continued as the ML. Inthree specimens the DL coursed from the dorsum sellae tothe mammillary bodies as a single membrane.
It merged with the ML superolaterally and coursed in ananterior-inferior direction, forming the DML in 17 specimens.
Mesencephalic leaf The ML leaf was observed in allspecimens. The membrane was thick in 14 specimens andopaque in 10. It had various free edges in 7 specimens,trabecular attachments in 5 and was closed totally in 12(Fig. 2b).
1702 Acta Neurochir (2011) 153:1701–1711

Fig. 2 a Arachnoid sleeves arose from the BAM to chiasmatic cisternand formed the anterior wall of the chiasmatic cistern covering thepituitary stalk superiorly (arrows). The pituitary stalk coursed to thepituitary gland, penetrating the BAM. b View of the BAM, which wasseparated from the tuberculum sellae and coursed over the superiorwall of the cavernous sinus, optic canals and anterior clinoid process,
and continued as the LM without any distinction. The ML attached tothe basilar artery bifurcation. The DL is absent, and the ML and LMattached to the uncus laterally covering the third nerve. BAM: Basalarachnoid membrane, LM: Liliequist’s membrane, PS:pituitary stalk,ON: optic nerves, PS: pituitary stalk, BAM: basal arachnoidmembrane, BAB: basilar artery bifurcation
Fig. 1 a Optic chiasma was removed. The LM coursed as acontinuation of the BAM over the posterior clinoids and split intotwo leaves as the DL and ML (a large part of a single piece of the LMwas removed to visualize the ML). The LM extended laterally andcovered the proximal third nerve. The DL attached to the corpusmamillare posteriorly, extending laterally to cover the distal part of thethird nerve and attached to the tentorial edges anterosuperolaterally. Itattached to the medial uncus inferolaterally. The ML attached to thebasilar artery anterior-inferiorly and continued through the prepontinecistern. b The optic chiasma was removed. The LM coursed as acontinuation of the BAM over the posterior clinoids and split into twoleaves as the DL and ML (a large part of the DL was removed to
visualize the ML). The DL coursed over the distal part of the thirdnerve and attached to the medial uncus inferolaterally. The MLcoursed towards to the basilar artery-vertebral artery junction throughthe prepontine cistern. c Illustrative drawing of the LM andsurrounding structures. BAM: Basal arachnoid membrane, LM:Liliequist’s membrane, PS: pituitary stalk, ON: optic nerves, PS:pituitary stalk, BAM: basal arachnoid membrane, DS: dorsum sellae,III.N: third nerve, ML: mesencephalic leaf, DL: diencephalic leaf,CM: corpus mamillare, BA: basilar artery. Horizontal elliptical bluearea represents the infrachiasmatic area; vertical elliptical blue arearepresents the prepontine area
Acta Neurochir (2011) 153:1701–1711 1703

Anterior projection
In 15 specimens, the ML split from the anterior portion ofthe LM. The ML coursed anteriorly to the basilarbifurcation in 8 of 12 and coursed towards to the basilarartery-vertebral artery junction in 4 of 12 specimens. Inthree specimens the ML coursed as a single membrane andattached to the anterior part of the basilar bifurcation.
In eight specimens, the ML was split from the posteriorpart of the LM and coursed anterior to the basilarbifurcation.
In one specimen there were two MLs, one split from theanterior portion and attached to the basilar artery at thelevel of the vertebral artery bifurcation, and the other splitfrom the posterior portion of the LM and attached to thebasilar artery bifurcation.
Lateral projections
The ML coursed in a posterior-lateral-inferior direction,forming the DML in 17 specimens. It attached with the DLsuperiorly. Lateral projections of the ML also included theMPPL and LPMM. The ML continued anterolaterally asthe MPMM and LPMM posterolaterally in all specimens(Fig. 3). These membranes covered the fifth and sixthnerves medially and laterally.
Diencephalic-mesencephalic leaf The DML was observedin 17 specimens (Fig. 3a). It was located anteroinferolat-erally on the distal part of the oculomotor nerve and gavearachnoid sheets to the anteromedial surface of theoculomotor nerve. The posterosuperior part of the leafwas the contination of the DL, and the anterosuperior partof the membrane was the pair of the ML. It was attached tothe LPMM inferolaterally. The inferior border of themembrane was located on the anterolateral surface of the
pons. In four specimens, arachnoid sleeves attaching to thecorpus mammillare formed the DL of the membrane.
In three specimens where the mesencephalic anddiencephalic leaves coursed separately as single mem-branes, an interpeduncular cistern was formed as a singleclosed room bordered by the DML inferolaterally and themedial carotid artery laterally and superolaterally, the DLposteriorly and ML anteriorly.
Medial and lateral pontomesencephalic membranes(Figs. 3b, 4a)
The MPMM coursed anterolaterally as a continuation of theML and attached to the medial surfaces of the fifth andsixth nerves, joining with the LPMM between these nervesin all specimens.The LPMM was a continuation of the MLin the posterolateral direction in all specimens. It extendedlaterally and attached to the lateral edges of the trigeminaland abducens nerves. The LPMM attached to the MPMMbetween the fifth and sixth nerves anteriorly in all speci-mens and continued as the LPMdM anteroinferiorly in 21specimens. The fourth nerve coursed to penetrate the LPMMbeneath the tentorium. The LPMM joined the formation of aring that covered the third nerve, attaching to the DL on thedistal posterior surface of the nerve and attaching to theparahippocampal gyrus coursing lateral to the uncus.
Medial and lateral pontomedullar membranes
The MPMdM and LPMdM were observed in 21 specimens.The MPMdM membrane was a continuation of the MPMM,and the LPMdM was a continuation of the LPMM in all 21specimens (Fig. 4b, c, d).
Lateral and medial pontomedullar membranes continuedas a single membrane in all specimens. They continued
Fig. 3 a Microscopic view of the MPML and LPML, which arecontiguous with the ML (proximal part of the third nerve was cut).The ML continued anterolaterally as the MPMM and LPMM poster-olaterally. It is attached to the basilar artery inferiorly. b The LPMMwas localized at the lateral edges of the trigeminal and abducens
nerves. The MPMM was localized at the medial part of these nerves.The tentorium was cut to clarify the course of the LPMM. The fifthnerve coursed to penetrate the LPMM, and the LPMM was dividedinto two parts at this point. *MPMM, T: tentorium
1704 Acta Neurochir (2011) 153:1701–1711

laterally covering the cranial nerves and coursed throughthe premedullary area in 17 specimens anteriorly, butattached the dura behind the clivus in 4 specimensinferiorly below the level where the vertebral arteries exit.
Occulomotor nerve
We separated the course of the third nerve into three parts(Fig. 5):
1. The entrance point (EP) represents the area between theoculomotor nerve entrance through the porus at thelevel between the anterior clinoid process and theposterior clinoids. This part was covered with the BAMmedially in all cases.
2. The proximal part represents the area between theinterpeduncular cistern and EP. If the mesencephalic anddiencephalic leaves were separated from the anteriorportion of the LM beneath the dorsum sellae, a single partof the LM covered the superior proximal part of thenerve. In case of a posterior separation level, asuperior single part of the LM and lateral aspect ofthe ML covered the proximal posterior surface of thenerve (8 specimens), and the DL covered the inferiorpart of the proximal part of the nerve. The proximalanteromedial surface of the nerve was covered withML in all cases.On the proximal anterior surface, theoculomotor nerve coursed between two leaves:medially the ML and laterally the LPMM. The
Fig. 4 a Endoscopic view ofthe MPMM and LPMM, whichsplit from the LM. The MPMMcovered the sixth nerve laterally(*) and attached to the fifthnerve medially. The LPMMcovered the fifth nerve laterally.b Membranes representing themedial pontomedullar(MPMdM) and lateral pontome-dullar (LPMdM) junction at thelevel of the basilar artery (BA).Vertebral artery (VA), bifurca-tion (arrow). c, d Endoscopicview of the membranes in theprepontine area. A: MPMM (*)coursing over the basilar artery(BA) and continuing as theMPMdM; B: LPMdM (short bigarrow) coursing over thevertebral artery (VA) laterallyand the MPMdM (thin arrow)coursing between the vertebralarteries. Note that these twomembranes cannot be separated
Fig. 5 The course of the third nerve was separated into three regions:the entrance point (EP) represents the area between the occulomotornerve entrance through the porus at the level between the anteriorclinoid process and the posterior clinoids. The proximal part (PROX)represents the area between the interpedincular cistern and EP. Thedistal part (DS) represents the region in the interpedincular cistern.The ML continued anterolaterally as the MPMM and posterolaterallyas the LPMM. The MPMM attached to the anterior and anteromedialpart of the proximal part of the third nerve. The LPMM extendedbackwards, covering the third nerve posterolaterally and laterally. Itformed a ring with arachnoid sleeves attaching the DL on the distalposterior surface of the nerve. The DL covered the distal posteriorsurface of the nerve in the interpedincular cistern. The fourth nerve (*)coursed beneath the tentorium penetrating the LPM. BAM: Basalarachnoid membrane, PS: pituitary stalk, ML: mesencephalic leaf, DL:diencephalic leaf, LPM: lateral pontomesencephalic membrane, MPM:medial pontomesencephalic membrane
Acta Neurochir (2011) 153:1701–1711 1705
proximal lateral surface of the nerve was coveredwith LPMM in all cases.
3. Distal part represents the region of the interpeduncularcistern. If mesencephalic and diencephalic leaves wereseparated from the anterior portion of the LM beneaththe dorsum sellae, the DL covered the distal part of themedial and posteriomedial surface of the nerve (13specimens). Otherwise, in the case of a posteriorseparation level, the DL covered only the distalposterior surface of the nerve in the interpeduncularcistern (8 specimens).The distal anteromedial surface ofthe nerve in the interpeduncular cistern was coveredwith DML in 17 specimens. The distal lateral surface ofthe nerve was covered with LPMM in all cases. DLcovered the distal part of the medial and posteriomedialsurface of the nerve.
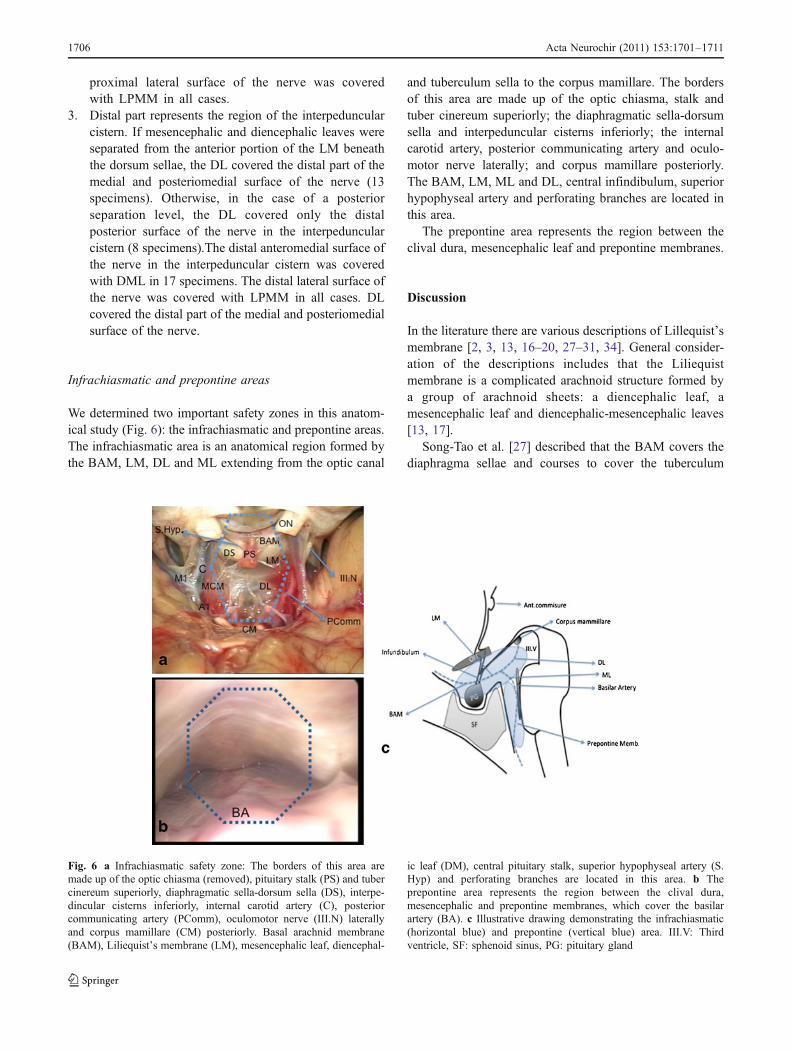
Infrachiasmatic and prepontine areas
We determined two important safety zones in this anatom-ical study (Fig. 6): the infrachiasmatic and prepontine areas.The infrachiasmatic area is an anatomical region formed bythe BAM, LM, DL and ML extending from the optic canal
and tuberculum sella to the corpus mamillare. The bordersof this area are made up of the optic chiasma, stalk andtuber cinereum superiorly; the diaphragmatic sella-dorsumsella and interpeduncular cisterns inferiorly; the internalcarotid artery, posterior communicating artery and oculo-motor nerve laterally; and corpus mamillare posteriorly.The BAM, LM, ML and DL, central infindibulum, superiorhypophyseal artery and perforating branches are located inthis area.
The prepontine area represents the region between theclival dura, mesencephalic leaf and prepontine membranes.
Discussion
In the literature there are various descriptions of Lillequist’smembrane [2, 3, 13, 16–20, 27–31, 34]. General consider-ation of the descriptions includes that the Liliequistmembrane is a complicated arachnoid structure formed bya group of arachnoid sheets: a diencephalic leaf, amesencephalic leaf and diencephalic-mesencephalic leaves[13, 17].
Song-Tao et al. [27] described that the BAM covers thediaphragma sellae and courses to cover the tuberculum
Fig. 6 a Infrachiasmatic safety zone: The borders of this area aremade up of the optic chiasma (removed), pituitary stalk (PS) and tubercinereum superiorly, diaphragmatic sella-dorsum sella (DS), interpe-dincular cisterns inferiorly, internal carotid artery (C), posteriorcommunicating artery (PComm), oculomotor nerve (III.N) laterallyand corpus mamillare (CM) posteriorly. Basal arachnid membrane(BAM), Liliequist’s membrane (LM), mesencephalic leaf, diencephal-
ic leaf (DM), central pituitary stalk, superior hypophyseal artery (S.Hyp) and perforating branches are located in this area. b Theprepontine area represents the region between the clival dura,mesencephalic and prepontine membranes, which cover the basilarartery (BA). c Illustrative drawing demonstrating the infrachiasmatic(horizontal blue) and prepontine (vertical blue) area. III.V: Thirdventricle, SF: sphenoid sinus, PG: pituitary gland
1706 Acta Neurochir (2011) 153:1701–1711

sellae and anterior clinoid processes. The LM arose fromthe BAM, covering the dorsum sellae and posterior clinoidprocesses, and is anteriorly adjacent to the tentorial edge.
In our study we did not observe any distinct separationof the LM and BAM. In all our specimens, the BAMcontinued as the LM without any attachment or border. TheBAM continued over the anterior clinoids and tuberculumsellae, and attached to the anterior-inferior part of the opticnerves at the level of the optic canal entrance.
Wang et al. defined the DL as usually being attached to theML from underneath, and above the DL it was usually a pairof hypothalamic membranes extending superomedially [32].
In our cadaveric study, we observed that the LM coursedas a single membrane covering the dorsum sellae andposterior clinoid processes. It continued anterior- superiorlyto the pituitary stalk covering the dorsum sella and coursedalong to the optic canals and tuberculum sella in allspecimens. The LM coursed posteriorly and separated intotwo leaves as the DL and ML either close to the dorsum
sella (anterior) or at a distance from the dorsum sella(posterior).
Lü et al [17] described the MPMM as being located inthe midline and joining the BA and pons at the bottom ofthe ML of the LM. The LPMM is reported to be locatedbetween the superior cerebral artery, and the trigeminalnerve was free or attached to the lateral end of the MPMM.Froelich et al. [13] demonstrated that the LPMM was acontinuation of the LM.
In all our specimens, we observed that the ML separatedinto two parts forming the MPMM and LPMM. The MLcoursed anterolaterally as the MPMM and attached to thetrigemineal and abducens nerves. The ML coursed poster-olaterally as the LPMM. Lateral and medial pontomesen-cephalic membranes attached between the 5th and 6thnerves in the lateral part of the prepontine cistern.
Lü, Zhu et al. [20] described that the medial pontome-dullar membrane is located in the midline of the pontome-dullary sulcus, the superior extremity of the medulla
Fig. 7 Intraoperative (a),preoperative (b) and postopera-tive (c) views of a patient withtuberculum sellae meningioma.OC: Optic chiasma, PS:pituitary stalk, T: residue tumor.*BAM
Fig. 8 Intraoperative (a),preoperative (b) and postopera-tive (c) views of a patient withpre-retro infindibularcraniopharyngioma. OC: Opticchiasma, PS: pituitary stalk, T:calcified residue tumor, DS:diaphragma sellae, CM: corpusmamillare, BA: basilar artery,SC: superior cerebellar artery
Acta Neurochir (2011) 153:1701–1711 1707

oblongata and basilar membrane, which is perpendicular tothe ventral surface of the pons and approximately parallel tothe basilar artery. They also demonstrated that the lateralpontomedullar membrane is a paired membrane, which islocated in the lateral portion of the pontomedullary sulcus.
In our study we did not observe a separate basilarmembrane. The ML coursed as a membrane towards thebasilar artery, which may represent the basilar arterymembrane.
Also other differences from the literature were theformation of the medial and lateral pontomedullar mem-branes. The medial pontomedullar membrane was acontinuation of the medial pontomesencephalic mem-brane, and the lateral pontomedullar membrane was thecontinuation of the lateral pontpmesencephalic membranein 21 specimens. These membranes coursed through theprepontine cistern as a single membrane and attached tothe dura covering the superior aspects of the cranialnerves laterally.
Actually, different descriptions of the LM may berelated to the complex structural formation of the ML. Inour study, we observed that this formation of the MLprovided the continuation of the ML leaf to theneighboring membranes.
Clinical implications
Extended endoscopic transphenoidal approach for skullbase lesions
An extended endoscopic transphenoidal approach is neededfor skull base lesions extending to the suprasellar area suchas craniopharyngeomas, suprasellar cysts, pituitary ade-nomas with suprasellar extension and tuberculum sellaemeningiomas [5–8, 12]. One of the drawbacks of thisapproach is the limited visibility of the posterior part ofthe lesions. In the last decade, use of endoscopicapproaches for sellar and parasellar lesions has increased.
Fig. 9 Intraoperative (a), pre-operative (b) and postoperative(c) views of a patient withdermoid tumor extended to thesuprasellar area. Intraoperativeview of the interpeduncular cis-tern after the tumor removal. PS:Pituitary stalk, BAM: basalarachnoid membrane, AC:anterior communicating artery,A1: right anterior cerebralartery, *superior hypophysealartery with the arterial root tothe optic nerve
Fig. 10 Intraoperative (afterremoval) (a), preoperative (b)and postoperative (c) views of apatient with suprasellar pituitaryadenoma. Dashed arrows:Mesencephalic leaf. Tc:Tubercinereum, CM: corpusmamillare, BA: basilar artery,PComm: posterior communicat-ing artery, PC: posterior cerebralartery, III.N: oculomotor nerve
1708 Acta Neurochir (2011) 153:1701–1711
In our clinic, we have been performing the endoscopicendonasal transphenoidal approach since 1997 and theextended/expanded endoscopic transphenoidal approachfor the last 8 years [4, 5]. Extended and expandedendoscopic transsphenoidal approaches are used formidline skull base lesions from the lamina cribrosa toforamen magnum and have been improved by thedevelopment of a standard endoscopic technique andincreased experience.
There are still many critics of the extended endo-scopic endonasal transphenoidal approach, especiallyfor lesions with an intradural location (meningiomas,craniopharengiomas, etc.) [5–8, 12]. These critiquesinclude that these lesions can be removed much moreeasily and confidently via traditional approaches; dissec-tion and prevention of neurovascular bundles locatedbehind the tumor are performed much more easily, andcomplication rates including CSF leakage are lower.These criticisms can be justifiable for intradural anteriormidline skull base lesions including olfactory groovemeningiomas.
However, EETS provides an important alternative routefor operating on the lesions (e.g., tuberculum sellameningiomas, craniopharengioma, pituitary adenomas,etc.) located in the infra-chiasmatic region, extending tothe third ventricle, corpus mamillare and prepontine area. Inthese approaches, the optic chiasma and persistance of theprefix chiasma are important limitations of the transcranialor supraorbital keyhole approaches.
The BAM and LM are important barriers protecting theoptic chiasm and these arteries. The infrachiasmatic area isan important surgical safety zone for inferior approaches tothis area. In this region, the superior hypophyseal arteriesascend along with the infindibulum; perforating arteriesascend along with the chiasma, tuber cinereum and corpusmamillare [25, 26].
In our clinic, we preferred the extended endoscopicendonasal transsphenoidal approach especially for infra-chiasmatically located tumors including the tuberculumsella, diaphragma sella meningiomas, craniopharyngiomasand other tumors [5, 6]. In this region in pre-infundibular(Fig. 7) or retro-infundibular lesions (Fig. 8) up to thecorpus mamillare and basilar bifurcation , more secureentrances are provided by the BAM and LM. It wasobserved that in tuberculum sellae meningiomas, the BAMis displaced inferiorly; however, in pituitary adenomas,craniopharyngiomas and dermoid tumors, it is displacedsuperiorly (Fig. 9). In pituitary adenomas extending to thisregion, the mesencephalic membrane is a leading andprotecting layer (Fig. 10).
The BAM is an important structure for these lesionsbecause it can cover the posterior-superior surface of thelesions and forms a critical border that avoids damage tothe structures located posterior and superior to the lesionsduring the surgical procedures. In our study, the BAMcontinued as the LM without a distinct border, and thisflexibility of the membrane avoids its laceration duringtumor growth. Intactness and elasticity of the membranemay allow the extension of these structures rather thaninvasion.
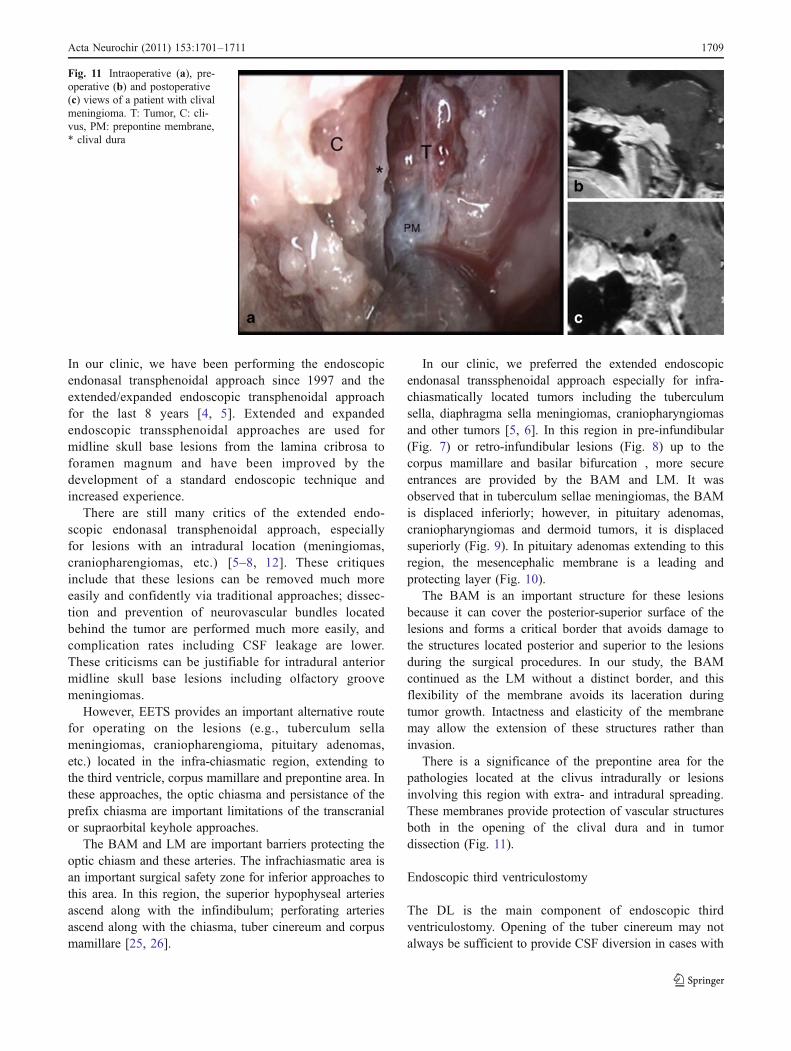
There is a significance of the prepontine area for thepathologies located at the clivus intradurally or lesionsinvolving this region with extra- and intradural spreading.These membranes provide protection of vascular structuresboth in the opening of the clival dura and in tumordissection (Fig. 11).
Endoscopic third ventriculostomy
The DL is the main component of endoscopic thirdventriculostomy. Opening of the tuber cinereum may notalways be sufficient to provide CSF diversion in cases with
Fig. 11 Intraoperative (a), pre-operative (b) and postoperative(c) views of a patient with clivalmeningioma. T: Tumor, C: cli-vus, PM: prepontine membrane,* clival dura
Acta Neurochir (2011) 153:1701–1711 1709

a dense DL [9–11, 14]. Moreover, as Froelich et al. [13]described, in some cases the DL is opened during thepuncture of the tuber cinereum. In these cases, the surgeonobserves the ML directly. Since the communicationbetween the anterior basal cisterns is variable according tothe anatomy of the LM, the success of the third ventricu-lostomy is also related to the CSF diversion through theprepontine area [9, 14]. In our study, the interpeduncularcistern was observed as a single closed space covered bythe DL, ML, DML and medial carotid membrane in threespecimens. In these cases opening of the DL may notprovide sufficient CSF diversion, and the ML becomes asignificant structure like the DL.
CSF accumulation through the prepontine cistern is alsoimportant for third ventriculostomy [1, 14]. In ourcadaveric study, it was observed that prepontine cisternsare mostly formed as a single membrane, and in fourspecimens pontomedullary membranes were attached to thedura behind the clivus, creating a wide prepontine cisternsurrounded by the membranes.
Conclusion
1. The continuity of the BAM, LM, DL and ML and therelation of the ML and prepontine membranes weredescribed. The structural formation of these membranesand their variations are important for the efficiency ofendoscopic third ventriculostomy.2. We have determined the borders of the infrachiasmaticand prepontine areas, which are described as safety zonesfor inferior approaches. The surgical importance of theseareas was emphasized by extended endoscopic transphe-noidal approaches.
Conflicts of interest None.
References
1. Anik I, Etus V, Anik Y et al. (2011) Role of Interpeduncular andPrepontine Cistern Cerebrospinal Fluid Flow Measurements inPrediction of Endoscopic Third Ventriculostomy Success inPediatric Triventricular Hydrocephalus. Pediatr Neurosurgdoi:10.1159/000323413 (in Press)
2. Brasil AV, Schneider FL (1993) Anatomy of Liliequist’s mem-brane. Neurosurgery 32:956–961
3. Buxton N, Vloeberghs M, Punt J (1998) Liliequist’s membrane inminimally invasive neurosurgery. Clin Anat 11:187–190
4. Ceylan S, Koc K, Anik I (2010) Endoscopic endonasal trans-sphenoidal approach for pituitary adenomas invading the cavernoussinus. J Neurosurg 112(1):99–107 (Erratum in: 2010, 112(1): 210)
5. Ceylan S, Koc K, Anik I (2009) Extended endoscopicapproaches for midline skull-base lesions. Neurosurg Rev 32(3):309–319
6. Ceylan S, Koc K, Anık I (2011) Extended endoscopic trans-phenoidal approach for tuberculum sellae meningiomas. ActaNeurochir (Wien) 153(1):1–9
7. de Divitiis E, Cappabianca P, Cavallo LM, Esposito F, de DivitiisO, Messina A (2007) Extended endoscopic transsphenoidalapproach for extrasellar craniopharyngiomas. Neurosurgery 61(5Suppl 2):219–227
8. de Divitiis E, Cavallo LM, Esposito F, Stella L, Messina A (2008)Extended endoscopic transsphenoidal approach for tuberculumsellae meningiomas. Neurosurgery 62(6 Suppl 3):1192–1201
9. Dinçer A, Kohan S, Ozek MM (2009) Is all "communicating"hydrocephalus really communicating? Prospective study on thevalue of 3D-constructive interference in steady state sequence at3 T. Am J Neuroradiol 30(10):1898–1906
10. Etus V, Ceylan S (2005) The role of endoscopic thirdventriculostomy in the treatment of triventricular hydrocepha-lus seen in children with achondroplasia. J Neurosurg 103(3Suppl):260–265
11. Etus V, Ceylan S (2005) Success of endoscopic third ventriculos-tomy in children less than 2 years of age. Neurosurg Rev 28(4):284–288
12. Frank G, Sciarretta V, Calbucci F et al. (2006) The endoscopictransnasal transsphenoidal approach for the treatment of cranialbase chordomas and chondrosarcomas. Neurosurgery 59(1 Suppl1):ONS50-7; discussion ONS50-7
13. Froelich SC, Abdel Aziz KM, Cohen PD, van Loveren HR,Keller JT (2008) Microsurgical and endoscopic anatomy ofLiliequist"s membrane: a complex and variable structure of thebasal cisterns.Neurosurgery 63(1 Suppl 1):ONS1-8; discussionONS8-9
14. Fushimi Y, Miki Y, Ueba T, Kanagaki M, Takahashi T,Yamamoto A, Haque TL, Konishi J, Takahashi JA, HashimotoN, Konishi J (2003) Liliequist membrane: three-dimensionalconstructive interference in steady state MR imaging. Radiol-ogy 229:360–365
15. Inoue K, Seker A, Osawa S, Alencastro LF, Matsushima T,Rhoton AL Jr (2009) Microsurgical and endoscopic anatomy ofthe supratentorial arachnoidal membranes and cisterns. Neurosur-gery 65(4):644–664
16. Key A, Retzius G (1875) Studien in der Anatomie desNervensystems und des Bindegewebes. Liliequist’s Membrane.Stockholm, vol 1
17. Lü J, Zhu XI (2003) Microsurgical anatomy of Liliequist’smembrane. Minim Invasive Neurosurg 46:149–154
18. Lü J, Zhu XL (2005) Characteristics of distribution andconfiguration of intracranial arachnoid membranes. Surg RadiolAnat 27:472–481
19. Lü J, Zhu XI (2005) Microsurgical anatomy of the interpeduncularcistern and related arachnoid membranes. J Neurosurg 103:337–341
20. Lü J, Zhu XL (2007) Cranial arachnoid membranes: some aspectsof microsurgical anatomy. Clin Anat 20(5):502–511
21. Magendie F (1842) Recherches Physiologiques et Cliniques sur leLiquide Céphalorachidienou cérébrospinal. Libraire Medicale deMequignon-marvis fils, Paris
22. Matsuno H, Rhoton AL Jr, Peace D (1988) Microsurgicalanatomy of the posterior fossa cisterns. Neurosurgery 23:58–80
23. Perneczky A (1999) Keyhole concept in neurosurgery: withendoscope-assisted microsurgery and case study. Georg Thieme,Stuttgart
24. Rhoton AL (2000) The posterior fossa cisterns. Neurosurgery 47:S287–S297
25. Rhoton AL Jr (2002) The sellar region. Neurosurgery 51(4Suppl):S335–S374
26. Rhoton AL Jr (2007) The cerebrum. Anatomy. Neurosurgery 61(1Suppl):37–118
1710 Acta Neurochir (2011) 153:1701–1711

27. Song-tao Q, Xi-an Z, Hao L, Jun F, Jun P, Yun-tao L (2010) Thearachnoid sleeve enveloping the pituitary stalk: anatomical andhistologic study. Neurosurgery 66(3):585–589
28. Sufianov AA, Sufianova GZ, Iakimov IA (2009) Microsurgicalstudy of the interpeduncular cistern and its communication withadjoining cisterns. Childs Nerv Syst 25(3):301–308
29. Vinas FC, Dujovny N, Dujovny M (1996) Microanatomical basisfor the third ventriculostomy. Minim Invasive Neurosurg 39:116–121
30. Vinas FC, Dujovny M, Fandino R, Chavez V (1996) Microsur-gical anatomy of the infratentorial trabecular membranes andsubarachnoid cisterns. Neurol Res 18:117–125
31. Vinas FC, Dujovny M, Fandino R, Chavez V (1996) Microsur-gical anatomy of the arachnoidal trabecular membranes andcisterns at the level of the tentorium. Neurol Res 18:305–312
32. Wang SS, Zheng HP, Zhang FH, Wang RM (2011) Microsurgicalanatomy of Liliequist"s membrane demonstrating three-dimensionalconfiguration. Acta Neurochir (Wien) 153(1):191–200
33. Yasargil MG, Kasdaglis K, Jain KK, Weber HP (1976) Anatom-ical observations of the subarachnoid cisterns of the brain duringsurgery. J Neurosurg 44:298–302
34. Zhang M, An PC (2000) Liliequist’s membrane is a fold of thearachnoid mater: study using sheet plastination and scanningelectron microscopy. Neurosurgery 47:902–909
Acta Neurochir (2011) 153:1701–1711 1711
Copyright © 2022 FDOKUMEN




















