
Microbial population in the biomass adhering to supporting material in a packed-bed reactor...
12
ENVIRONMENTAL BIOTECHNOLOGY Microbial population in the biomass adhering to supporting material in a packed-bed reactor degrading organic solid waste Kengo Sasaki & Shin Haruta & Yoshiyuki Ueno & Masaharu Ishii & Yasuo Igarashi Received: 11 November 2006 / Revised: 11 February 2007 / Accepted: 13 February 2007 / Published online: 3 March 2007 # Springer-Verlag 2007 Abstract An anaerobic packed-bed reactor using carbon fiber textiles (CFT) as the supporting material was continuously operated using an artificial garbage slurry. 16S rRNA gene analysis showed that many bacteria in the biomass adhering to CFT were closely related to those observed from other anaerobic environments, although a wide variety of unidentified bacteria were also found. Dot blot hybridization results clarified that 16S rRNA levels of methanogens in the adhering biomass were higher than those in the effluent. Based on microscopic observation, the adhering biomass consisted of microorganisms, organic material, and void areas. Bacteria and Archaea detected by fluorescence in situ hybridization were distributed from the surface to the inner regions of the adhering biomass. Methanosarcina sp. tended to be more abundant in the inner part of the adhering biomass than at the surface. This is the first report to elucidate the structure of the microbial community on CFT in a packed-bed reactor. Keywords Packed-bed reactor . Microbial community . Organic solid waste Introduction A huge amount of garbage is currently being generated worldwide (Williams 1994; Haruta et al. 2005). Incinera- tion and landfill, widely used for the treatment of garbage, can cause pollution in the natural environment. Anaerobic digestion is an alternative method that is able to recover energy sources from organic waste. Methanogenic packed- bed reactors, which are packed with supporting material such as carbon fiber textiles (CFT), are applicable for solid wastes (Sasaki et al. 2006b). Furthermore, packed-bed systems have many advantages over other types of bioreactors. Packed-bed systems do not need a long start- up period and have no risk of accidental washout of the biomass (Lanthier et al. 2005). A recent study by Tatara et al. (2004) showed that a packed-bed reactor with CFT was applicable under very high organic loading rates (OLR) and short hydraulic retention times (HRT), even if it was compared with other processes using supporting materials, such as anaerobic upflow fixed-film reactor or fluidized bed reactor. CFT provide the microorganisms with a place to grow and allow them to exist stably in the reactor. Therefore, the micro- organisms adhering to CFT are of great interest. In our previous study, archaeal communities were compared in a packed-bed reactor operated on an artificial garbage slurry (AGS), while the OLR was increased stepwisely and the acetate-removing methanogen, Methanosarcina sp., became dominant among methanogens in the adhering fraction at an OLR of 12.2 g/l/day (Sasaki et al. 2006b). However, the microbial population and its spatial structure in the adhering biomass in packed-bed reactors degrading organic solid waste have not yet been clarified. In the present study, we operated an anaerobic reactor packed with CFT using AGS as a model organic solid Appl Microbiol Biotechnol (2007) 75:941–952 DOI 10.1007/s00253-007-0888-x K. Sasaki (*) : S. Haruta : M. Ishii : Y. Igarashi Department of Biotechnology, Graduate School of Agricultural and Life Science, The University of Tokyo, Yayoi 1-1-1, Bunkyo-ku, Tokyo 113-8657, Japan e-mail: [email protected] Y. Ueno Environmental and Bioengineering Group, Kajima Technical Research Institute, Tobitakyu 2-19-1, Chofu-shi, Tokyo 182-0036, Japan
Transcript of Microbial population in the biomass adhering to supporting material in a packed-bed reactor...

ENVIRONMENTAL BIOTECHNOLOGY
Microbial population in the biomass adhering to supportingmaterial in a packed-bed reactor degrading organicsolid waste
Kengo Sasaki & Shin Haruta & Yoshiyuki Ueno &
Masaharu Ishii & Yasuo Igarashi
Received: 11 November 2006 /Revised: 11 February 2007 /Accepted: 13 February 2007 /Published online: 3 March 2007# Springer-Verlag 2007
Abstract An anaerobic packed-bed reactor using carbonfiber textiles (CFT) as the supporting material wascontinuously operated using an artificial garbage slurry.16S rRNA gene analysis showed that many bacteria in thebiomass adhering to CFT were closely related to thoseobserved from other anaerobic environments, although awide variety of unidentified bacteria were also found. Dotblot hybridization results clarified that 16S rRNA levels ofmethanogens in the adhering biomass were higher thanthose in the effluent. Based on microscopic observation, theadhering biomass consisted of microorganisms, organicmaterial, and void areas. Bacteria and Archaea detected byfluorescence in situ hybridization were distributed from thesurface to the inner regions of the adhering biomass.Methanosarcina sp. tended to be more abundant in theinner part of the adhering biomass than at the surface. Thisis the first report to elucidate the structure of the microbialcommunity on CFT in a packed-bed reactor.
Keywords Packed-bed reactor .Microbial community .
Organic solid waste
Introduction
A huge amount of garbage is currently being generatedworldwide (Williams 1994; Haruta et al. 2005). Incinera-tion and landfill, widely used for the treatment of garbage,can cause pollution in the natural environment. Anaerobicdigestion is an alternative method that is able to recoverenergy sources from organic waste. Methanogenic packed-bed reactors, which are packed with supporting materialsuch as carbon fiber textiles (CFT), are applicable for solidwastes (Sasaki et al. 2006b). Furthermore, packed-bedsystems have many advantages over other types ofbioreactors. Packed-bed systems do not need a long start-up period and have no risk of accidental washout of thebiomass (Lanthier et al. 2005).
A recent study by Tatara et al. (2004) showed that apacked-bed reactor with CFT was applicable under veryhigh organic loading rates (OLR) and short hydraulicretention times (HRT), even if it was compared with otherprocesses using supporting materials, such as anaerobicupflow fixed-film reactor or fluidized bed reactor. CFTprovide the microorganisms with a place to grow and allowthem to exist stably in the reactor. Therefore, the micro-organisms adhering to CFT are of great interest. In ourprevious study, archaeal communities were compared in apacked-bed reactor operated on an artificial garbage slurry(AGS), while the OLR was increased stepwisely and theacetate-removing methanogen, Methanosarcina sp., becamedominant among methanogens in the adhering fraction at anOLR of 12.2 g/l/day (Sasaki et al. 2006b). However, themicrobial population and its spatial structure in theadhering biomass in packed-bed reactors degrading organicsolid waste have not yet been clarified.
In the present study, we operated an anaerobic reactorpacked with CFT using AGS as a model organic solid
Appl Microbiol Biotechnol (2007) 75:941–952DOI 10.1007/s00253-007-0888-x
K. Sasaki (*) : S. Haruta :M. Ishii :Y. IgarashiDepartment of Biotechnology, Graduate School of Agriculturaland Life Science, The University of Tokyo,Yayoi 1-1-1, Bunkyo-ku,Tokyo 113-8657, Japane-mail: [email protected]
Y. UenoEnvironmental and Bioengineering Group,Kajima Technical Research Institute,Tobitakyu 2-19-1, Chofu-shi,Tokyo 182-0036, Japan

waste. The aims of this research were to gain insight intothe microbial structure of the adhering biomass on the CFTby molecular ecological analysis and to elucidate themicrobial architecture by fluorescence in situ hybridization(FISH) coupled with confocal laser scanning microscopy.
Materials and methods
Anaerobic packed-bed bioreactor
The details of the packed-bed reactor system were de-scribed in previous reports (Tatara et al. 2004; Sasaki et al.2006a). In short, a 5-l capacity jar-fermentor was used asthe bioreactor and was packed with supporting materialcomposed of CFT (125×90×2 mm). AGS containing 40 gof commercial dog food (Vita-one, Nihon Pet Food, Tokyo,Japan) and 960 ml of tap water was fed at a predeterminedHRT. The effluent from the reactor was discharged from theoverflow line. Concentrations of suspended solids (SS) andvolatile suspended solids (VSS) of the AGSwere 32.9 g/l and31.0 g/l, respectively. The total chemical oxygen demand(COD) of the AGS was 44.0 g/l. AGS in the reactor wasrecycled from the bottom to the top by an external peristalticpump to mix the contents of the reactor. The operationaltemperature was 55°C.
The packed-bed reactor was operated for 387 days at anOLR between 1.4 g/l/day and 12.2 g/l/day, followed by anadditional 48 days of operation at an OLR of 12.2 g/l/day(HRT of 3.6 days), for a total of 435 days of operation. Theaverage values of reactor conditions for the last 20 days wereas follows; pH, 6.9±0.1; gas production, 3.8±0.2 l/l/day(methane content in the biogas accounted for 62% and theremainder was carbon dioxide); acetate concentration, 18.1±4.1 mM; propionate, 4.5±2.6 mM; butyrate, 1.2±0.1 mM.The reactor performance for these days was as follows: CODremoval efficiency, 71.5±3.8%; SS digestion efficiency,86.5±2.7%; VSS digestion efficiency, 87.0±2.9%. Approx-imately 56% of the carbon in the influent was converted tomethane gas and the carbon recovery was nearly 85%.
Analysis
Gas production was monitored by the water displacementmethod. Samples were taken periodically from the reactoreffluent port. Volatile fatty acids (VFA) and COD in theeffluent and methane concentration in the biogas wereanalyzed as described previously (Tatara et al. 2004; Uenoet al. 2001). For VFA analyses, suspended solids wereremoved by centrifugation at 6,000×g for 3 min and filtrationthrough the membrane (0.45 μm pore size). SS and VSSwere determined in accordance with the methods of theJapanese Industrial Standards Committee (1986).
DNA and RNA extraction
After 402, 412, 422 and 435 days of operation, the collectedeffluents were filtered through a membrane (100 μm poresize) and divided into two fractions: solid fraction (>100 μm,designated as fraction S) and planktonic-phase fraction(<100 μm, designated as fraction P). In addition, threesheets (15×30×2 mm) of CFT were retrieved directly fromthe reactor and the adhering biomass was collected (desig-nated as fraction CF).
Fractions S, P, and CF were centrifuged to collect theprecipitate (10,000×g, 4°C, 5 min). The nucleic acids wereextracted by the benzyl chloride method (Zhu et al. 1993).Samples treated with benzyl chloride were further mixed andvortexed with 0.1 mm zirconia/silica beads for 10 s. CrudeDNA obtained by this method was purified by phenolextraction and isopropanol precipitation. Total RNAs werepurified by the low-pH, phenol–chloroform extraction proce-dure and DNA digestion with DNase (Wako, Osaka, Japan).
PCR–denaturing gradient gel electrophoresis
PCR amplification was performed using Ampli Taq Goldaccording to the manufacturer’s instructions (AppliedBiosystems, Japan) for denaturing gradient gel electropho-resis (DGGE) analysis. The PCR primers used to amplifybacterial 16S rRNA genes were B357F-GC, 5′-CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGGCCTACGGGAGGCAGCAG-3′ (Escherichia colipositions 341–357) and U517R 5′-ATTACCGCGGCTGCTGG-3′ (E. coli positions 517–534) (Muyzer et al. 1993).The sequence in italics is a GC clamp. The archaeal 16SrRNA genes were amplified by nested PCR. The primersets A21F 5′-TCCGGTTGATCCYGSCRG-3′ (E. colipositions 8–25) (Delong 1992) plus A915R 5′-GTGCTCCCCCGCCAATTCCT-3′ (E. coli positions 915–934) (Stahland Amann 1991) and A348IF 5′-GGIGCAICAGGCGCGAAA-3′ (E. coli positions 348–365) plus U806IR-GC 5′-GGACTACCIGGGTITCTAA-3′ (E. coli positions 788–806) (Hori et al. 2006) were used for the first and secondPCR, respectively. A GC clamp (CGCCCGCCGCGCCCCGCGCCCGGCCCGCCGCCCCCGCCCC) was attached toU806Ir-GC at the 5′-terminus (Myers et al. 1989). The PCRamplification and DGGE were performed as describedpreviously (Haruta et al. 2002; Sasaki et al. 2006b).
Library construction, sequencing, and phylogeny
PCR was performed to amplify the 16S rRNA genes withprimer sets B27F 5′-AGAGTTTGATCCTGGCTCAG-3′(E. coli positions 8–27) plus U1492R 5′-GGYTACCTTGTTACGACTT-3′ (E. coli positions 1492–1510) for Bacteria(Lane 1991) and A21F (Delong 1992) plus A915R (Stahl
942 Appl Microbiol Biotechnol (2007) 75:941–952

and Amann 1991) for Archaea. The PCR amplification wasperformed as follows: 95°C for 10 min, followed by25 cycles for Bacteria and 93°C for 60 s, 50°C for 60 s,72°C for 90 s, and a final elongation at 72°C for 5 min forArchaea. The amplified fragments were cloned into E. coliJM109 by using the pGEM-T Easy vector system (Promega,Tokyo, Japan). Randomly chosen from the bacterial andarchaeal clone libraries were 83 and 74 clones, respectively.Plasmids were extracted and purified with GenElute plasmidMiniprep Kit (Sigma, USA). Operational taxonomic units(OTUs) were defined based on mobility by PCR–DGGE.Bacterial clones were subjected to PCR with the primerset B357F-GC plus U517R. The primer set A348If plusU806IR-GC was used for archaeal clones. Sequence sim-ilarities of a few clones with the same mobility were alsochecked. Sequencing reactions were carried out with aBigDye kit (Applied Biosystems). The products wereanalyzed with an ABI 3130xl DNA sequencer (AppliedBiosystems). The internal 16S rRNA primers U515F, 5′-GTGCCAGCAGCCGCGG-3′ (E. coli positions 515–530)and B907R, 5′-CCCCGTCAATTCCTTTGAGTTT (E. colipositions 907–927) (Lane 1991) were also used in additionto primers T7 and SP6 for full-length sequencing. Wechecked all sequences using the CHIMERA_CHECK pro-gram version 2.7 in the Ribosomal Database Project II(Maidak et al. 2001). The sequences were aligned by theCLUSTAL_X program (Thompson et al. 1997). Neighbor-joining phylogeny (Saitou and Nei 1987) was constructedwith the MEGA program (Kumar et al. 2001). To evaluatethe robustness of the inferred tree, the bootstrap resamplingmethod of Felsenstein (1985) was used with 1,000 replicates.
Dot blot hybridization
Dot blot hybridization was carried out according to themethod of Raskin et al. (1994) with some modifications.
The extracted RNA was first denatured and blotted onto apositively charged nylon membrane (AmershamBiosciences,England) in triplicate by a manifold device (BioRad, Tokyo,Japan). The membrane was baked at 80°C for 30 min andthen UV-irradiated. The membrane was further prewettedwith a hybridization buffer (5×SSC, 1% blocking solution,0.1% N-lauroylsarcosine, 0.02% SDS) at 40°C for 2 h.Hybridization was performed at 40°C for 12 h by adding thedigoxigenin (DIG)-labeled oligonucleotide probe. After thehybridization, the membrane was first washed with awashing solution (1% SDS, 5×SSC) at 40°C for 2 h andthen at the desired washing temperature (Tw) for 30 min.The sequences and Tw of those domain-specific and group-specific oligonucleotide probes used are listed in Table 1.Nonspecific binding was blocked by incubation of themembrane with 1% blocking solution in buffer (0.1 Mmaleic acid, 0.15 M NaCl, pH 7.5). The DIG-labeled probewas detected as described by Sasaki et al. (2006b). Thesignal of the sample in individual hybridizations was nor-malized against the reference and expressed as the percentageof that obtained with the universal probe. Reference strainsused were E. coli (IAM1264), Methanosarcina barkeri(JCM 10043), Methanoculleus thermophilicus (DSM2373), and Methanothermobacter thermautotrophicus(JCM 10044).
Sample fixation and cryosectioning
Three sheets (15×30×2 mm) of CFT including the adheringbiomass were taken from the reactor. The adhering biomasson the CFT was fixed by incubation with 4% paraformal-dehyde in phosphate-buffered saline (PBS) and gentlywashed in PBS. After fixation, 3 freeze-and-thaw cycles(−80°C to 60°C) were done to increase the efficiency ofpenetration of oligonucleotide probes into cells (Sekiguchiet al. 1999). Samples were embedded in Tissue-Tek OCT
Table 1 Oligonucleotide probes used for dot blot hybridizations and FISH
Probe Specificity Sequence (5′–3′) Target-sitea
Twb
(°C)Formamidec
(%)Citation
UNIV1390 All organisms GACGGGCGGTGTGTACAA 1,390–1,407 43 –d Zheng et al. 1996EUB338 Subset of Bacteria GCTGCCTCCCGTAGGAGT 338–355 – 5 Stahl and Amann
1991EUB338II Subset of Bacteria GCAGCCACCCGTAGGTGT 338–355 – 5 Daims et al. 1999EUB338III Subset of Bacteria GCTGCCACCCGTAGGTGT 338–355 – 5 Daims et al. 1999ARC915 Archaea GTGCTCCCCCGCCAATTCCT 915–934 58 35 Raskin et al. 1994MS821 Methanosarcina CGCCATGCCTGACACCTAGCGAGC 821–844 60 5 Raskin et al. 1994MG1200 Methanomicrobiales CGGATAATTCGGGGCATGCTG 1,200–1,220 53 5 Raskin et al. 1994MB1174 Methanobacteriaceae TACCGTCGTCCACTCCTTCCTC 1,174–1,195 62 35 Raskin et al. 1994
aCorresponds to the numbering in the sequence of the 16S rRNA gene of E. coli.bWashing temperature used in dot blot hybridizations.c Formamide concentration in the hybridization buffer used in FISH.dNot determined.
Appl Microbiol Biotechnol (2007) 75:941–952 943

compound (Sakura, Tokyo, Japan) and frozen at −20°C.Vertical thin sections (12 μm thick) of the fixed sampleswere prepared with the cryomicrotome (HM520; Micro-edge Instruments, Tokyo, Japan). Vertical thin sectionswere fixed on gelatin-coated microscopic slides, air dried,and dehydrated in a series of increasing concentrations ofethanol (50%, 80%, and 100% [v/v]).
In situ hybridization
The following 16S rRNA-targeted oligonucleotide probeswere used: EUBMIX (a mixture of EUB338, EUB338II,and EUB338III) (for Bacteria), ARC915 (for Archaea),MB1174 (for the family Methanobacteriaceae), MG1200(for the order Methanomicrobiales), and MS821 (for thegenus Methanosarcina). All probe sequences, their speci-ficities, and hybridization conditions are given in Table 1.For in situ hybridization, the probes were labeled with eitherAlexa555 or Alexa647. For determination of probe specific-ity, the following reference organisms were used: E. coli(IAM1264), M. thermophilicus (DSM 2373), M. barkeri(JCM 10043), and M. thermautotrophicus (JCM 10044).
Hybridization was performed at 46°C for 12 h withhybridization buffer (0.9 M NaCl, 20 mM Tris–HCl[pH 7.2], 0.01% sodium dodecyl sulfate) containing 5 ngof each labeled probe (Amann 1995). The hybridizationstringency was adjusted by adding formamide to thehybridization buffer. The washing step was done at 48°Cfor 30 min with washing buffer containing the same
components as the hybridization buffer except for theprobes. The sections hybridized with the probes wereobserved by confocal laser scanning microscopy (CLSM)(FV500; Olympus, Tokyo, Japan).
Nucleotide sequence accession numbers
The nucleotide sequences reported in this paper have beendeposited in the GSDB, DDBJ, EMBL, and NCBInucleotide sequence databases under accession numbersAB274486 to AB274517.
Results
Bacterial and archaeal profiles by PCR–DGGE
The packed-bed reactor was exposed to an increasing OLRand finally operated for 48 days at an OLR of 12.2 g/l/day.PCR–DGGE profiles for the domain Bacteria or Archaea inthe effluents (fractions P and S) and the biomass adheringto the CFT (fraction CF) were taken during the last monthof operation (Fig. 1). If there were microorganisms existingin only floating or attaching conditions, microbial commu-nities would differ among fractions. From the archaeal andbacterial profiles, bands with the same mobility weredetected in all three fractions, although small differencesin the relative intensity of certain bands among the threefractions were observed. These results suggest that the
Fig. 1 Changes in 16S rRNAgene-based DGGE profilesobtained by PCR amplification;the primer sets for the domainBacteria (a) and Archaea(b) were used. Lanes 1 to 4:fraction P, 5 to 8: fraction S, 9 to12: fraction CF samples takenfrom 402, 412, 422, and435 days of the operation,respectively
944 Appl Microbiol Biotechnol (2007) 75:941–952
microbial composition was similar in each of the threefractions. The last day of operation was selected for thefollowing microbial community analyses.
Archaeal population
Four OTUs were found in the archaeal 16S rRNA geneclones from fraction CF. These OTUs were close relativesof methanogens (Table 2). OTU CFA-1, which was mostfrequently observed in the clone library, was associated withthe aceticlastic methanogen, Methanosarcina thermophila(Zinder and Robert 1979). The second most abundant OTU,CFA-2, was related to the hydrogenotrophic methanogen,Methanoculleus palmolei (Zeller et al. 1998). In addition,two OTUs, CFA-3 and CFA-4, were found to be related tothe hydrogenotrophic methanogens, M. thermautotrophicus(Zeikus andWolfe 1972) andMethanobacterium formicicum(Bryant and Boone 1987), respectively.
The distribution of each methanogen was comparedamong fractions P, S, and CF. The relative amount of 16SrRNA from each methanogen compared to that fromwhole organisms was determined by dot blot hybridizationwith the following oligonucleotide probes: probe MS821targeted for Methanosarcina sp., probe MG1200 targetedfor Methanoculleus sp., and probe MB1174 for Methano-bacterium sp. and Methanothermobacter sp. (Table 3). Thetotal amount of methanogens in fraction CF was highestamong the three fractions. The distribution of methanogensin fraction S was more similar to that in fraction CF ratherthan in fraction P. These results indicate that the methanogens
in fraction S were derived from fraction CF. On the otherhand, the specific distribution of methanogens dominated byMethanoculleus sp. was observed in fraction P.
Bacterial population
High SS, VSS, and COD removal efficiency in this reactorare characteristic of the bacterial diversity present in thereactor. Therefore, 16S rRNA gene clonal analysis wasperformed for the domain Bacteria. Twenty-eight OTUswere obtained in the bacterial library for fraction CF(Table 4). The clonal distribution within the library was asfollows: Firmicutes (49%), Chloroflexi (15%), Bacteroidetes(13%), Thermotogae (4%), and Planctomycetes (2%), whilethe remaining clonal sequences (17%) were unclassified(Figs. 2 and 3). Of the total clonal sequences, 60% (10OTUs) were related to bacterial 16S rRNA gene sequencesreported in the public databases with >97% sequencesimilarity. On the other hand, 40% of the total clones (18OTUs) were distinct from known sequences in the publicdatabases (<97% similarity).
Microbial distribution in the adhering biomass on carbonfiber textiles
Direct sampling of CFT was carried out. The thickness ofCFT covered with the adhering biomass was 2.5 to 3.0 mm(data not shown). Figure 4a represents a differentialinterference contrast image of an overview of the section.Carbon fibers are represented by dark black regions. It wasshown that the biomass adhering to CFT was rich with voidspaces where no organic material was found.
To determine the spatial distribution of Bacteria andArchaea, a bacterial-domain probe (EUBMIX) and anarchaeal-domain probe (ARC915) were applied to thesection. Figure 4b and c shows representative FISH imagesfor the surface portion of the biomass adhering to CFT andthe part well below the surface, respectively. Organicsubstrates producing autofluorescence were shown by thecolor yellow due to overlapping red and green signals. Bothbacterial and archaeal cells were detected on/in organicsubstances throughout all parts on the CFT. In addition, theobvious layered structure of Bacteria and Archaea as
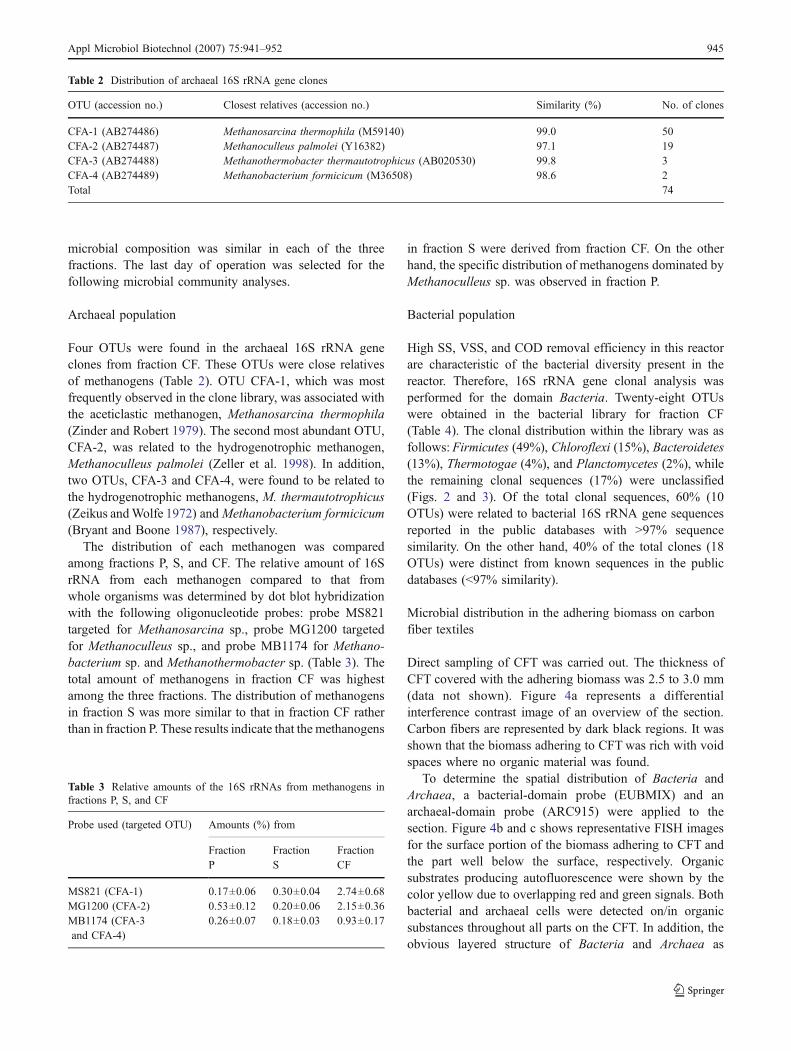
Table 2 Distribution of archaeal 16S rRNA gene clones
OTU (accession no.) Closest relatives (accession no.) Similarity (%) No. of clones
CFA-1 (AB274486) Methanosarcina thermophila (M59140) 99.0 50CFA-2 (AB274487) Methanoculleus palmolei (Y16382) 97.1 19CFA-3 (AB274488) Methanothermobacter thermautotrophicus (AB020530) 99.8 3CFA-4 (AB274489) Methanobacterium formicicum (M36508) 98.6 2Total 74
Table 3 Relative amounts of the 16S rRNAs from methanogens infractions P, S, and CF
Probe used (targeted OTU) Amounts (%) from
FractionP
FractionS
FractionCF
MS821 (CFA-1) 0.17±0.06 0.30±0.04 2.74±0.68MG1200 (CFA-2) 0.53±0.12 0.20±0.06 2.15±0.36MB1174 (CFA-3and CFA-4)
0.26±0.07 0.18±0.03 0.93±0.17
Appl Microbiol Biotechnol (2007) 75:941–952 945

observed in granules within upflow anaerobic sludgeblanket (UASB) reactors (Sekiguchi et al. 1999) was notdetected in the adhering biomass within the packed-bedreactor. However, the ratio of archaeal cells to bacterialcells at the inner part of the adhering biomass appeared tobe higher than that at the surface. In addition, the spatialdistribution of methanogens detected by clonal analysis wasevaluated by FISH with relevant probes. In the innerregions of the adhering biomass, microcolonies of Meth-anosarcina sp. were frequently observed and the ratio ofMethanosarcina sp. to bacterial cells seemed to be highcompared with that at the surface (Fig. 5a and b). On theother hand, distinguishable signal was not detected usingprobe MB1171 or MG1200, although nucleic analysesindicated that the population of Methanoculleus sp. wascomparable to that of Methanosarcina sp. (Tables 2 and 3).
Discussion
Effective decomposition and gasification of AGS in thepacked-bed reactor were ascertained by observation of highSS and COD removal and gas production for prolongedperiods. The microbial structure and architecture in theadhering biomass on the surface of the CFT within thisreactor were analyzed.
In our previous report, we analyzed the archaealcommunity in a packed-bed reactor operated at an OLRof 12.2 g/l/day (Sasaki et al. 2006b); however, operationallength (207 days in the previous report) and concentrationsof VFAs (low in the previous report) differed from those ofthe present study. The previous study also detected se-quences related to Methanosarcina sp., Methanoculleus sp.,and Methanothermobacter sp. by PCR–DGGE analysis.
Table 4 Distribution of bacterial 16S rRNA gene clones
OTU (accession no.) Closest sequences/microorganism (accession no.) Similarity (%) No. Percentage (%)
Firmicutesa
CFB-10 (AB274499) MSW cluster clone MBNA08 (AB114327) 99.5 11 13.3CFB-19 (AB274508) Anaerobaculum mobile (U50711) 99.5 1 1.2CFB-1 (AB274490) Clostridium sp. PPf35E10 (AY548785) 94.2 3 3.6CFB-14 (AB274503) Pelotomaculum thermopropionicum (AB035723) 91.3 3 3.6CFB-16 (AB274505) MSW landfill clone GZKB39 (AJ853533) 89.6 3 3.6CFB-3 (AB274492) MSW landfill clone GZKB101 (AJ853594) 88.4 3 3.6CFB-18 (AB274507) Anaerobaculum mobile (U50711) 97.4 2 2.4CFB-12 (AB274501) Clostridium sp. AC007 (AY330129) 96.8 2 2.4CFB-15 (AB274504) Syntrophic bacterium OL (AB106354) 94.1 2 2.4CFB-6 (AB274495) Clostridium stercorarium (AJ310082) 92.2 2 2.4CFB-2 (AB274491) Clostridium sp. EBR-02E-0046 (AB186360) 98.4 1 1.2CFB-17 (AB274506) Coprothermobacter proteolyticus (X69335) 97.6 1 1.2CFB-7 (AB274496) Clostridium straminisolvens (AB125279) 93.2 1 1.2CFB-11 (AB274500) Clostridium sp. AC007 (AY330129) 92.6 1 1.2CFB-4 (AB274493) MSW landfill clone GZKB101 (AJ853594) 91.4 1 1.2CFB-8 (AB274497) Syntrophomonas sapovorans (AF022249) 88.8 1 1.2CFB-5 (AB274494) Uncultured bacterium clone p-1528-b5 (AF371777) 88.0 1 1.2CFB-13 (AB274502) Desulfotomaculum thermocisternum (U33455) 87.0 1 1.2CFB-9 (AB274498) Caloranaerobacter azorenasis (AJ272422) 85.7 1 1.2BacteroidetesCFB-20 (AB274509) Sludge digestor clone 009F02_B_SD_P15 (CR933158) 97.7 10 12.0CFB-21 (AB274510) MSW landfill clone GZKB3 (AJ853498) 97.7 1 1.2UnclassifiedCFB-22 (AB274511) Uncultured bacterium clone BSA2B-19 (AB175391) 98.1 14 16.9ChloroflexiCFB-26 (AB274515) UASB granule clone STG-2 (AB109432) 100 6 7.2CFB-23 (AB274512) Anaerolinea thermophila (AB046413) 90.0 3 3.6CFB-24 (AB274513) Anaerolinea thermophila (AB046413) 90.0 1 1.2CFB-25 (AB274514) Anaerolinea thermophila (AB046413) 88.3 2 2.4ThermotogaeCFB-27 (AB274516) Sludge digester clone 060F05_B_SD_P93 (CR933305) 99.8 3 3.6PlanctomycetesCFB-28 (AB274517) Anoxic bulk soil clone PBS-71 (AJ390467) 90.3 2 2.4Total 83
a Phylum, to which OTUs in lower columns belong, is shown.
946 Appl Microbiol Biotechnol (2007) 75:941–952

However, CFA-2 identified in this study differed slightlyfrom the related organism detected previously (Sasaki et al.2006b) (96.8% similarity). In addition, Methanobacteriumsp. was not detected in the previous study. Extended reactoroperation and several differences in operating conditionsmay have allowed the succession of the methanogeniccommunity. Methanogenic archaea found in this reactor arewidely distributed in similar environments. On the otherhand, rice cluster III (RCIII)-related sequence, which hadbeen detected as a major band by previous PCR–DGGEanalysis, was not observed in this study. Members of RCIII
have not been reported to produce methane (Großkopf et al.1998) and RCIII-related organism might be washed-outafter prolonged operation.
From bacterial diversity analysis, we identified many16S rRNA gene sequences that were closely related tosequences retrieved from other anaerobic environments ormethanogenic bioreactors (not packed-bed systems). CFB-2and CFB-10 were affiliated with Clostridium sp. EBR-02E-0046 (Shiratori et al. 2006) and clone MBNA08 (Tang et al.2004), which were isolated or detected from thermophilicreactors degrading municipal solid waste (MSW). Similarly,
Fig. 2 Phylogenetic tree of theclones among the phylum Fir-micutes (low G+C gram-positivebacteria). Numbers at nodesrepresent bootstrap values(Felsenstein 1985) for the nodes(1,000 times resampling analy-sis). Arthrobacter globiformiswas used as the outgroup
Appl Microbiol Biotechnol (2007) 75:941–952 947

clone STG-2, which is related to CFB-26, was found ingranules from a thermophilic UASB operated on wastewa-ter (Yamada et al. 2005). CFB-20 and CFB-27 were closelyrelated to clones from mesophilic reactors fed with sludge(Chouari et al. 2005). In addition, CFB-22 had highsimilarity with a clone from a mesophilic reactor operatedon an artificial medium that included bovine serum albuminas the sole carbon source (Tang et al. 2005). CFB-21 wasvery similar to clone GZKB3 from the MSW landfill(Huang et al. 2005). In addition, two OTUs (CFB-18 andCFB-19) showed high similarities to Anaerobaculummobile, which was isolated from a methanogenic meso-philic lagoon treating wool-sourcing wastewater (Menesand Muxi 2002). All of these clones were related toorganisms that are ubiquitous in methanogenic conditions.Finally, one OTU (CFB-17) was closely related toCoprothermobacter proteolyticus, which was isolated froma non-methanogenic anaerobic compost digesting bio-kitchen (Kersters et al. 1994). Therefore, temperature,available substrates or operational conditions were differentfrom those in the reactor analyzed in this study. Thesebacterial groups do not appear to be specific to packed-bed
systems. CFB-2, CFB-6, CFB-17, CFB-18, and CFB-19were closely related to B-13, B-21, B-5, B-8, andB-8 detectedfrom the packed-bed reactor degrading medium, whichincluded acetic and butyric acids as major carbon sources(Sasaki et al. 2006a) with 97%, 100%, 98%, 99%, and 97%identities, respectively (Fig. 2). Thus, these bacteria wouldbe responsible for degradation of low-molecule organicmatters. One OTU (CFB-10) belonged to MSW cluster 1 towhich many clonal sequences from the MSW digester wereassigned (Tang et al. 2004).
The remaining unidentified organisms in 16S rRNAgene analysis seem to be characteristic of the bacterialcommunity in the packed-bed systems, i.e., growing onorganic substrates in the adhered state under anaerobicconditions. CFB-7 and CFB-8 belonged to Clostridiumcluster III and cluster VIII, which include cellulolytic mi-crobes and syntrophic fatty acid-oxidizing species (Collinset al. 1994), respectively. In addition, CFB-14 belonged tothe cluster composed of Pelotomaculum spp., which werereported to be propinate-oxidizing bacteria (Imachi et al.2002). CFB-23, CFB-24, CFB-25, and CFB-26 were affil-iated with theChloroflexi subphylum I (Fig. 3). In this group,
Fig. 3 Phylogenetic tree of theclones among the phyla Bacter-oidetes, Chloroflexi, Planctomy-cetes, and Thermotogae and theclones unclassified so far.Numbers at nodes are the sameas in Fig. 2. E. coli was used asthe outgroup
948 Appl Microbiol Biotechnol (2007) 75:941–952

a few strains, which were isolated from UASB reactors, werereported to utilize monomeric and dimeric carbohydrates,starch, and amino acids in the presence of yeast extract(Yamada et al. 2005; Sekiguchi et al. 2003). Members of thephyla Firmicutes, Chloroflexi, and Planctomycetes were alsofound in the sludge digester and UASB granules (Chouariet al. 2005; Sekiguchi et al. 1998). Input of complex organicsubstrates and/or CFT would cause the appearance of theirgroups. Their physiological properties, such as growth rate,adhering (or aggregating) nature, degradation rate of solidsubstances, and metabolic collaboration with methanogensare of considerable interest.
We studied the microbial architecture on CFT. Micro-scopic observation of the cryomicrotomy sections of CFTwith the adhering biomass elucidated the overall structure.CFT was covered with organic substances to whichmicrobial cells adhered. Void spaces were frequentlyobserved in the biomass adhering to CFT. Textiles and/orsolid materials may influence the development of voidareas. Open channels could connect the outside surface ofthe adhering biomass and the bulk solution with the innerregions and facilitate transport of nutrients and microbialcells. This adhering biomass structure may extend the
surface area where microbial cells efficiently obtainrelevant substrates and release their products.
Quantification of the relative amount of rRNA indicatedthat the ratio of methanogens to total organisms in thebiomass adhering to the CFT was significantly higher thanthat in the effluent. This result indicates that a certainportion of methanogens was washed-out in the effluentthrough continuous operation at a HRT of 3.6 days.However, the CFT selectively allowed the methanogens tobe stably maintained in the reactor. The aggregating natureof Methanosarcina sp. (Boone and Mah 2001) may help itto exist in the adhering biomass as previously suggested forthe reactor operated under the same conditions (Sasaki et al.2006b). Methanosarcina sp.-related microcolonies tendedto be more common in the inner regions of the adheringbiomass than on the surface. The void structure woulddiminish gradients of substrate concentrations within theadhering biomass. Therefore, the distribution of Methano-sarcina sp. cells in the adhering biomass was probablycaused by washing-out of the cells at the surface, whichwas exposed to shear stress and stream flow. Particularlyfor organisms with slow growth rates, distinct spatialarrangements are likely to be present within fractions on
Fig. 4 DIC images of adheringbiomass section on carbon fibertextiles (a) and CLSM projec-tion images of the microscopicfields enclosed by boxes after insitu hybridization withAlexa647-labeled bacterial-do-main probes (EUBMIX) (green)and Alexa555-labeled archaeal-domain probe (ARC915) (red)(b and c). The surface (b) andinner (c) regions were visual-ized. Yellow areas are due toautofluorescence
Appl Microbiol Biotechnol (2007) 75:941–952 949

the CFT. Methanosarcina sp. grows on acetate, which isproduced from complex organic matters. Such metabolismof Methanosarcina sp. would form carbon flow from thesurface to the internal part of the CFT. In contrast to thisstudy, our previous study reported that hydrogenotrophicmethanogens (which were detected as predominant specieson CFT in this study) made up small fractions among themethanogens on CFT. Extended reactor operation will
increase the amount of biomass adhering to CFT and/orchange the structure. Development of the adhering biomassmay provide space that is favorable for the growth ofhydrogenotrophic methanogens. However, FISH signals forhydrogenotrophic methanogens, which were observed inclone library analysis, could not be detected from theadhering biomass. These cells would probably distributeindividually and not aggregate as Methanosarcina sp. asreported previously (Zeikus and Wolfe 1972; Bryant andBoone 1987; Zeller et al. 1998). Thus, their signals fromsingle cells might have been concealed by signals fromadjacent organic substrate and bacterial cells. In addition,increase of cell wall thickness by syntrophically growingMethanothermobacter cells might diminish signals fromthem (Nakamura et al. 2006).
In this study, we characterized the microbial compositionand architecture on the supporting materials, i.e., CFT in apacked-bed reactor operated on AGS. The porous structureof the adhering biomass resulted in localization of activemicroorganisms throughout the material. Furthermore, CFTprovided a stable place for slow-growing organisms toproliferate. CFT apparently allowed the development of thecharacteristic microbial structure and played a crucial rolein the efficient performance of the packed-bed reactor. It istempting to elucidate roles and networks of uniquemicrobial diversity on CFT. We expect that this informationwill aid in the development of packed-bed systems.
Acknowledgements We thank E. Kumano and T. Kuroiwa ofMicro-edge Instruments for the technical advice on cryosectioning.This work was supported by the New Energy and IndustrialTechnology Development Organization (NEDO).
References
Amann RI (1995) In situ identification of micro-organisms by whole-cell hybridization with rRNA-targeted nucleic acid probes. In:Akkerman ADL, van Elsas JD, de Bruijn FJ (eds) Molecularmicrobial ecology manual. Kluwer, The Netherlands, pp 1–15
Boone DR, Mah RA (2001) Genus I Methanosarcina. In: Boone DR,Castenholz RW (eds) Bergey’s manual of systematic bacteriol-ogy, vol. 1. Springer, New York, pp 269–276
Bryant MP, Boone DR (1987) Isolation and characterization ofMethanobacterium formicicum MF. Int J Syst Bacteriol 37:171
Chouari R, Paslier DL, Daegelen P, Ginestet P, Weissenbach J, Sghir A(2005) Novel predominant archaeal and bacterial groups revealedby molecular analysis of an anaerobic sludge digestor. EnvironMicrobiol 7:1104–1115
Collins MD, Lawson PA, Willems A, Cordoba JJ, Fernandez-Garayzabal J, Garcia P, Cai J, Hippe H, Farrow JAE (1994)The phylogeny of the genus Clostridium: proposal of five newgenera and eleven new species combinations. Int J Syst Bacteriol44:812–826
Daims H, Brühl A, Amann R, Schleifer KH, Wagner M (1999) Thedomain-specific probe EUB338 is insufficient for the detection ofall Bacteria: development and evaluation of a more comprehen-sive probe set. Syst Appl Microbiol 22:434–444
Fig. 5 FISH microscopic images of adhering biomass section oncarbon fiber textiles. Alexa647-labeled bacterial-domain probes(EUBMIX) (green) and Alexa555-labeled genus Methanosarcina probe(MS821) (red) were applied. The surface (a) and inner (b) regions werevisualized. Yellow areas are due to autofluorescence
950 Appl Microbiol Biotechnol (2007) 75:941–952

Delong EF (1992) Archaea in coastal marine environments. Proc NatlAcad Sci USA 89:5685–5689
Felsenstein J (1985) Confidence limits on phylogenies: an approachusing the bootstrap. Evolution 39:783–791
Großkopf R, Stubner S, Liesack W (1998) Novel Euryarchaeotalineages detected on rice roots and in the anoxic bulk soil offlooded rice microcosms. Appl Environ Microbiol 64:4983–4989
Haruta S, Kondo M, Nakamura K, Aiba H, Ueno S, Ishii M, Igarashi Y(2002) Microbial community changes during organic solid wastetreatment analyzed by double gradient-denaturing gradient gelelectrophoresis and fluorescence in situ hybridization. ApplMicrobiol Biotechnol 60:224–231
Haruta S, Nakamura T, Nakamura K, Hemmi H, Ishii M, Igarashi Y,Nishino T (2005) Microbial diversity in biodegradation andreutilization processes of garbage. J Biosci Bioeng 99:1–11
Hori T, Haruta S, Ueno Y, Ishii M, Igarashi Y (2006) Direct comparisonof single-strand conformation polymorphism (SSCP) and dena-turing gradient gel electrophoresis (DGGE) to characterize amicrobial community on the basis of 16S rRNA gene fragments. JMicrobiol Methods 66:165–169
Huang LN, Zhu S, Zhou H, Qu LH (2005) Molecular phylogeneticdiversity of bacteria associated with the leachate of a closedmunicipal solid waste landfill. FEMSMicrobiol Lett 242:297–303
Imachi H, Sekiguchi Y, Kamagata Y, Hanada S, Ohashi A, Harada H(2002) Pelotomaculum thermopropionicum gen. nov., sp. nov.,an anaerobic thermophilic, syntrophic propionate-oxidizing bac-terium. Int J Syst Evol Microbiol 52:1729–1735
Japanese Industrial Standards Committee (1986) Organic matter. In:Namiki H (ed) Testing methods for industrial wastewater, JIS K0102-1986. Japanese Standards Association, Tokyo, pp 32–33
Kersters I, Maestrojuan GM, Torck U, Vancanneyt M, Kersters K,Verstraete W (1994) Isolation of Coprothermobacter proteolyticusfrom an anaerobic digest and further characterization of thespecies. Syst Appl Microbiol 17:289–295
Kumar S, Tamura K, Jakobsen IB, Nei M (2001) MEGA2: molecularevolutionary genetics analysis software. Bioinformatics 17:1244–1245
Lane DJ (1991) 16S/23S rRNA sequencing. In: Stackebrandt E,Goodfellow M (eds) Nucleic acid techniques in bacterial system-atics. Wiley, New York, pp 115–175
Lanthier M, Juteau P, Lépine F, Beaudet R, Villemur R (2005) Desulfito-bacterium hafniense is present in a high proportion within thebiofilms of a high-performance pentachlorophenol-degrading meth-anogenic fixed-film reactor. Appl Environ Microbiol 71:1058–1065
Maidak B, Cole JR, Lilburn TG, Parker CT, Saxman PR, Farris RJ,Garrity GM, Olsen GJ, Schmidt TM, Tiedje JM (2001) The RDP-II (Ribosomal Database Project). Nucleic Acids Res 29:173–174
Menes RJ, Muxi L (2002) Anaerobaculum mobile sp. nov., a novelanaerobic, moderately thermophilic, peptide-fermenting bacteriumthat uses crotonate as an electron acceptor, and emended descriptionof the genus Anaerobaculum. Int J Syst Evol Microbiol 52:157–164
Muyzer G, de Waal ECD, Uitterlinder AG (1993) Profiling of complexmicrobial populations by denaturing gradient gel electrophoresisanalysis of polymerase chain reaction-amplified genes coding for16S rRNA. Appl Environ Microbiol 59:695–700
Myers RM, Scheffield VC, Cox DR (1989) Mutation detection byPCR, GC-clamps, and denaturing gradient gel electrophoresis. In:Erlich HA (ed) PCR-technology—principles and applications forDNA amplification. Stocken Press, New York, pp 71–88
Nakamura K, Terada T, Sekiguchi Y, Shinzato N, Meng XY, Enoki M,Kamagata Y (2006) Application of pseudomurein endoisopepti-dase to fluorescence in situ hybridization of methanogens withinthe family Methanobacteriaceae. Appl Environ Microbiol72:6907–6913
Raskin L, Stromley JM, Rittmann BE, Stahl DA (1994) Group-specific16S rRNA hybridization probes to describe natural communitiesof methanogens. Appl Environ Microbiol 60:1232–1240
Saitou N, Nei M (1987) The neighbor-joining method: a new methodfor reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Sasaki K, Haruta S, Tatara M, Yamazawa A, Ueno Y, Ishii M, Igarashi Y(2006a)Microbial community in methanogenic packed-bed reactorsuccessfully operating at short hydraulic retention time. J BiosciBioeng 101:271–273
Sasaki K, Haruta S, Ueno Y, Ishii M, Igarashi Y (2006b) Archaealpopulation on supporting material in a methanogenic packed-bedreactor. J Biosci Bioeng 102:244–246
Sekiguchi Y, Kamagata Y, Syutsubo K, Ohashi A, Harada H,Nakamura K (1998) Phylogenetic diversity of mesophilic andthermophilic granular sludges determined by 16S rRNA analysis.Microbiology 144:2655–2665
Sekiguchi Y, Kamagata Y, Nakamura K, Ohashi A, Harada H (1999)Fluorescence in situ hybridization using 16S rRNA-targetedoligonucleotides reveals localization of methanogens and select-ed uncultured bacteria in mesophilic and thermophilic sludgegranules. Appl Environ Microbiol 65:1280–1288
Sekiguchi Y, Yamada T, Hanada S, Ohashi A, Harada H, Kamagata Y(2003) Anaerolinea thermophila gen. nov., sp. nov. and Caldineaaerophila gen. nov., sp. nov., novel filamentous thermophilesthat represent a previously uncultured lineage of the domainBacteria at the subphylum level. Int J Syst Evol Microbiol 53:1843–1851
Shiratori H, Ikeno H, Ayame S, Kataoka N, Miya A, Hosono K,Beppu T, Ueda K (2006) Isolation and characterization of anew Clostridium sp. that performs effective cellulosic waste di-gestion in a thermophilic methanogenic bioreactor. Appl EnvironMicrobiol 72:3702–3709
Stahl DA, Amann R (1991) Development and application of nucleic acidprobes in bacterial systematics. In: Stackebrandt E, GoodfellowM (eds) Nucleic acid techniques in bacterial systematics. Wiley,New York, pp 205–248
Tang Y, Shigematsu T, Ikbal, Morimura S, Kida K (2004) The effectsof micro-aeration on the phylogenetic diversity of microorgan-isms in a thermophilic anaerobic municipal solid-waste digester.Water Res 38:2537–2550
Tang Y, Shigematsu T, Morimura S, Kida K (2005) Microbialcommunity analysis of mesophilic anaerobic protein degradingprocess using bovine serum albumin (BSA)-fed continuouscultivation. J Biosci Bioeng 99:150–164
Tatara M, Yamazawa A, Ueno Y, Fukui H, Goto M, Sode K (2004)High-rate thermophilic methane fermentation on short-chain fattyacids in a down-flow anaerobic packed-bed reactor. BioprocessBiosyst Eng 27:105–113
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG(1997) The clustal_X windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools.Nucleic Acids Res 25:4876–4882
Ueno Y, Haruta S, Ishii M, Igarashi Y (2001) Changes in productformation and bacterial community by dilution rate on carbohy-drate fermentation by methanogenic microflora in continuousflow stirred tank reactor. Appl Microbiol Biotechnol 57:65–73
Williams ME (1994) Integrated solid waste management. In: Kreith F(ed) Handbook of solid waste management. McGraw-Hill, NewYork
Yamada T, Sekiguchi Y, Imachi H, Kamagata Y, Ohashi A, Harada H(2005) Diversity, localization, and physiological properties offilamentous microbes belonging to Chloroflexi subphylum I inmesophilic and thermophilic methanogenic sludge granules. ApplEnviron Microbiol 71:7493–7503
Zeikus JG, Wolfe RS (1972) Methanobacterium thermoautotrophicussp. n., an anaerobic, autotrophic, extreme thermophiles. J Bacteriol109:707–713
Zeller G, Messner P, Winter J, Stackebandt E (1998) Methanoculleuspalmolei sp. nov., an irregularly coccoid methanogen from an
Appl Microbiol Biotechnol (2007) 75:941–952 951

anaerobic digester treating wastewater of a palm oil plant innorth-Sumatra, Indonesia. Int J Syst Bacteriol 48:1111–1117
Zheng D, Alm EW, Stahl DA, Raskin L (1996) Characterization ofuniversal small-subunit rRNA hybridization probes for quanti-tative molecular ecology studies. Appl Environ Microbiol 62:4504–4513
Zhu H, Qu F, Zhu LH (1993) Isolation of genomic DNAs from plants,fungi and bacteria using benzyl chrolide. Nucleic Acids Res21:5279–5280
Zinder SH, Robert AM (1979) Isolation and characterization of athermophilic strain of Methanosarcina unable to use H2–CO2
for methanogenesis. Appl Environ Microbiol 38:996–1008
952 Appl Microbiol Biotechnol (2007) 75:941–952




















