
Mechanism of Coenzyme Recognition and Binding Revealed by Crystal Structure Analysis of...
10
Mechanism of Coenzyme Recognition and Binding Revealed by Crystal Structure Analysis of Ferredoxin– NADP 1 Reductase Complexed with NADP 1 Juan A. Hermoso 1 * , Tomas Mayoral 1 , Merche Faro 2 Carlos Go ´ mez-Moreno 2 , Julia Sanz-Aparicio 1 and Milagros Medina 2 1 Grupo de Cristalografı ´a Macromolecular y Biologı ´a Estructural, Instituto Quı ´mica-Fı ´sica Rocasolano C.S.I.C., Serrano 119, 28006 Madrid, Spain 2 Departamento de Bioquı ´mica y Biologı ´a Molecular y Celular Facultad de Ciencias Universidad de Zaragoza 50009 Zaragoza, Spain The flavoenzyme ferredoxin – NADP þ reductase (FNR) catalyses the production of NADPH in photosynthesis. The three-dimensional structure of FNR presents two distinct domains, one for binding of the FAD prosthetic group and the other for NADP þ binding. In spite of extensive experiments and different crystallographic approaches, many aspects about how the NADP þ substrate binds to FNR and how the hydride ion is transferred from FAD to NADP þ remain unclear. The structure of an FNR:NADP þ complex from Anabaena has been determined by X-ray diffraction analysis of the cocrystallised units to 2.1 A ˚ resolution. Struc- tural perturbation of FNR induced by complex formation produces a narrower cavity in which the 2 0 -phospho-AMP and pyrophosphate por- tions of the NADP þ are perfectly bound. In addition, the nicotinamide mononucleotide moiety is placed in a new pocket created near the FAD cofactor with the ribose being in a tight conformation. The crystal struc- ture of this FNR:NADP þ complex obtained by cocrystallisation displays NADP þ in an unusual conformation and can be considered as an inter- mediate state in the process of coenzyme recognition and binding. Struc- tural analysis and comparison with previously reported complexes allow us to postulate a mechanism which would permit efficient hydride transfer to occur. Besides, this structure gives new insights into the postu- lated formation of the ferredoxin:FNR:NADP þ ternary complex by pre- diction of new intermolecular interactions, which could only exist after FNR:NADP þ complex formation. Finally, structural comparison with the members of the broad FNR structural family also provides an explanation for the high specificity exhibited by FNR for NADP þ /H versus NAD þ /H. q 2002 Elsevier Science Ltd. All rights reserved Keywords: ferredoxin – NADP þ reductase; coenzyme binding; NADPH; NADH; X-ray structure *Corresponding author Introduction Although the general mechanism of action for many flavoenzymes is known, the mechanism of the hydride ion transfer between the isoalloxa- zine ring and the nicotinamide ring of the coenzyme is not completely understood for most of them. This is the case for ferredoxin–NADP þ reductase (FNR) and other structurally related enzymes. Three-dimensional structures of FNR from different sources have been determined in their oxidised 1–4 and reduced states. 2 FNR con- sists of two distinct domains, one responsible for binding of the FAD prosthetic group and the other for NADP þ binding. This unique structural two-domain motif has been proposed to be a prototype for a large family of flavoproteins. 5 Structurally characterised members of this family involve phthalate dioxygenase reduc- tase 6 (PDR), the FAD-containing fragment of the NADH-dependent nitrate reductase, 7 cyto- chrome b 5 reductase, 8 NADPH-cytochrome P450 reductase, 9 flavodoxin reductase 10 and sulfite reductase. 11 0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved E-mail address of the corresponding author: [email protected] Abbreviations used: FNR, ferredoxin – NADP þ reductase; Fd, ferredoxin; NMN, nicotinamide monoucleotide portion of NADP þ /H; 2 0 P-AMP, 2 0 -phospho-AMP portion of NADP þ /H; bOG, b-octylglucoside; PEG, polyethylene glycol; PDR, phthalate dioxygenase reductase. doi:10.1016/S0022-2836(02)00388-1 available online at http://www.idealibrary.com on B w J. Mol. Biol. (2002) 319, 1133–1142
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Mechanism of Coenzyme Recognition and Binding Revealed by Crystal Structure Analysis of...

Mechanism of Coenzyme Recognition and BindingRevealed by Crystal Structure Analysis of Ferredoxin–NADP1 Reductase Complexed with NADP1
Juan A. Hermoso1*, Tomas Mayoral1, Merche Faro2
Carlos Gomez-Moreno2, Julia Sanz-Aparicio1 and Milagros Medina2
1Grupo de CristalografıaMacromolecular y BiologıaEstructural, InstitutoQuımica-Fısica RocasolanoC.S.I.C., Serrano 119, 28006Madrid, Spain
2Departamento de Bioquımica yBiologıa Molecular y CelularFacultad de CienciasUniversidad de Zaragoza50009 Zaragoza, Spain
The flavoenzyme ferredoxin–NADPþ reductase (FNR) catalyses theproduction of NADPH in photosynthesis. The three-dimensional structureof FNR presents two distinct domains, one for binding of the FADprosthetic group and the other for NADPþ binding. In spite of extensiveexperiments and different crystallographic approaches, many aspectsabout how the NADPþ substrate binds to FNR and how the hydride ionis transferred from FAD to NADPþ remain unclear. The structure of anFNR:NADPþ complex from Anabaena has been determined by X-raydiffraction analysis of the cocrystallised units to 2.1 A resolution. Struc-tural perturbation of FNR induced by complex formation produces anarrower cavity in which the 20-phospho-AMP and pyrophosphate por-tions of the NADPþ are perfectly bound. In addition, the nicotinamidemononucleotide moiety is placed in a new pocket created near the FADcofactor with the ribose being in a tight conformation. The crystal struc-ture of this FNR:NADPþ complex obtained by cocrystallisation displaysNADPþ in an unusual conformation and can be considered as an inter-mediate state in the process of coenzyme recognition and binding. Struc-tural analysis and comparison with previously reported complexes allowus to postulate a mechanism which would permit efficient hydridetransfer to occur. Besides, this structure gives new insights into the postu-lated formation of the ferredoxin:FNR:NADPþ ternary complex by pre-diction of new intermolecular interactions, which could only exist afterFNR:NADPþ complex formation. Finally, structural comparison with themembers of the broad FNR structural family also provides an explanationfor the high specificity exhibited by FNR for NADPþ/H versus NADþ/H.
q 2002 Elsevier Science Ltd. All rights reserved
Keywords: ferredoxin–NADPþ reductase; coenzyme binding; NADPH;NADH; X-ray structure*Corresponding author
Introduction
Although the general mechanism of actionfor many flavoenzymes is known, the mechanismof the hydride ion transfer between the isoalloxa-zine ring and the nicotinamide ring of thecoenzyme is not completely understood for mostof them. This is the case for ferredoxin–NADPþ
reductase (FNR) and other structurally relatedenzymes. Three-dimensional structures of FNRfrom different sources have been determined intheir oxidised1 – 4 and reduced states.2 FNR con-sists of two distinct domains, one responsiblefor binding of the FAD prosthetic group and theother for NADPþ binding. This unique structuraltwo-domain motif has been proposed to be aprototype for a large family of flavoproteins.5
Structurally characterised members of thisfamily involve phthalate dioxygenase reduc-tase6 (PDR), the FAD-containing fragment ofthe NADH-dependent nitrate reductase,7 cyto-chrome b5 reductase,8 NADPH-cytochrome P450reductase,9 flavodoxin reductase10 and sulfitereductase.11
0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved
E-mail address of the corresponding author:[email protected]
Abbreviations used: FNR, ferredoxin–NADPþ
reductase; Fd, ferredoxin; NMN, nicotinamidemonoucleotide portion of NADPþ/H; 20P-AMP,20-phospho-AMP portion of NADPþ/H; bOG,b-octylglucoside; PEG, polyethylene glycol; PDR,phthalate dioxygenase reductase.
doi:10.1016/S0022-2836(02)00388-1 available online at http://www.idealibrary.com onBw
J. Mol. Biol. (2002) 319, 1133–1142
In plants, algae and cyanobacteria FNR plays astrategic role during photosynthesis by mediatingthe reversible two-electron transfer between twomolecules of the one-electron carrier ferredoxin(Fd) and a single NADPþ/H molecule.12 It is pro-posed that this process requires the formation ofa transient ternary complex between the threepartners, FNR, NADPþ and Fd, in which oxidisedFNR is thought to form a complex with NADPþ
prior to its association with reduced Fd.13 Oncethe FNR is fully reduced, the flavin and the nico-tinamide rings must be placed in a relative orien-tation such that functional hydride transfer fromN5 of the flavin to C4 of the nicotinamide wouldbe allowed. These conditions could be onlyachieved by disruption of the stacking interactionof the FNR C-terminal Tyr side-chain with theisoalloxazine ring or by changes in the relativeorientation of the two protein domains in the
FNR. Only a few structures of flavoenzymesbound to NADPþ in a “productive” mode areavailable. In the case of glutathione reductase andNADPH peroxidase the nicotinamide and flavinring planes are parallel, making extensive contacts,and the C4 of the nicotinamide is just 3.8 A of N5from the flavin.14 – 16 In NAD(P)H–quinonereductase, however, although both rings are alsoparallel there is no superposition of the centralpart of the flavin with the nicotinamide ring.17 Thecrystal structure of a ferric reductase in complexwith NADPþ also shows a packing of theNMN (nicotinamide mononucleotide) half of thecoenzyme and the isoalloxazine ring.18 However,this enzyme differs significantly from FNR, sinceit possesses neither an NADPþ-binding domainable to discriminate between NADPþ/H andNADþ/H, nor an aromatic residue packingagainst the flavin ring in the position where the
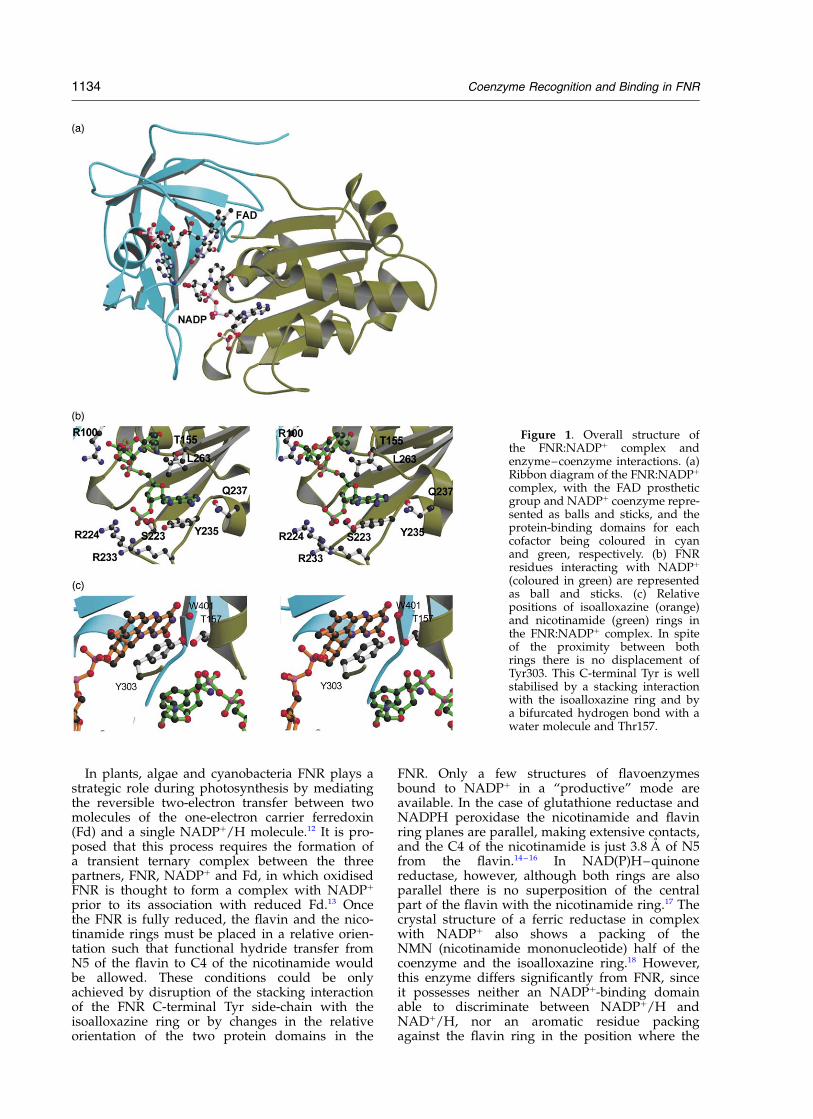
Figure 1. Overall structure ofthe FNR:NADPþ complex andenzyme–coenzyme interactions. (a)Ribbon diagram of the FNR:NADPþ
complex, with the FAD prostheticgroup and NADPþ coenzyme repre-sented as balls and sticks, and theprotein-binding domains for eachcofactor being coloured in cyanand green, respectively. (b) FNRresidues interacting with NADPþ
(coloured in green) are representedas ball and sticks. (c) Relativepositions of isoalloxazine (orange)and nicotinamide (green) rings inthe FNR:NADPþ complex. In spiteof the proximity between bothrings there is no displacement ofTyr303. This C-terminal Tyr is wellstabilised by a stacking interactionwith the isoalloxazine ring and bya bifurcated hydrogen bond with awater molecule and Thr157.
1134 Coenzyme Recognition and Binding in FNR

nicotinamide is expected to bind. Complexes withNADPþ/H of FNR forms from different specieshave always resulted in a nicotinamide ring eithernot visible or away from the isoalloxazine ring.1 – 3
Only partial success has been reported with acouple of mutants in the pea enzyme, where theterminal Tyr residue was replaced either by Ser orTrp.19 In these complexes, specially in the Y308Smutant, the obtained structures of the complexeswith NADPH and NADPþ are proposed toresemble that of the productive state of thecomplex. In order to provide a structural guidelinefor elucidating the mechanism of electron transferfrom FNR to the coenzyme we have furtherstudied the interaction between the unmodifiedenzyme and NADPþ by X-ray crystallography.
Results and Discussion
Structure of the FNR:NADP1 complex
Cocrystallisation of FNR with NADPþ producedcrystals with a completely different packing fromthose of native, wild-type and any mutant so farreported. The structure was solved by molecularreplacement, and the electron density maps clearlyshowed strong density for the complete NADPþ
molecule. The crystal structure of this FNR:NADPþ
complex has been refined up to 2.1 A resolution.The overall FNR folding is equivalent to that ofthe free Anabaena FNR. Differences observed inthe 104–114 loop seem to be related to the struc-tural flexibility of this region3,20,21 that it is involvedin crystal packing contacts. No significant changesin the relative orientation of the FAD and NADPþ-binding domains were observed, and the NADPþ
molecule is perfectly complementary within acavity situated at the C-terminal edge of the paral-lel b-sheets of the NADPþ-binding domain, whichextends towards the FAD-binding domain inter-face (Figure 1(a)). The coenzyme shows a charac-teristic L-shape with the pyrophosphate grouplocated at the corner and the two branches of theL accommodating the NMN moiety and the 20-P-AMP, respectively. The adenine moiety of theNADPþ is sandwiched between the hydrophobicLeu263 side-chain and the aromatic ring of Tyr235,and is also making a hydrogen bond with theGln237 side-chain (Figure 1(b)). The 20-P group ofNADPþ is stabilised by several H-bonds with theside-chains of Ser223, Arg224 and Arg233, andthe OH group of Tyr235. Two H-bonds are alsodetected between 20-P and residues Lys105 andThr112 of a symmetry-related FNR molecule. Thepyrophosphate moiety of NADPþ is essentiallystabilised by the guanidinium group of Arg100.The nicotinamide moiety is only stabilised by twopolar interactions with Thr155 and by a van derWaals contact with Leu263. It is worth noting thevery tight conformation in which the ribose isfound, specially considering that no interactionswith the enzyme are found which could account
for this feature. This may be related to theenzymatic mechanism as will be discussed below.
Finally, Tyr303 is stacked between the isoalloxa-zine and the nicotinamide rings with the NADPC4 atom being 7.78 A far from the FAD N5 atom(Figure 1(c)). Besides, nicotinamide ring presentsits B-face to the Re-face of the FAD and thereforeis not orientated properly according to the stereo-specificity defined for FNR. Therefore, it is remark-able that although in this complex the nicotin-amide ring of the NADPþ is placed nearthe isoalloxazine ring, it does not replace theC-terminal Tyr as it has been proposed to berequired for efficient electron transfer. A similarsituation of the nicotinamide moiety has been alsoobserved in a PDR complex with NADH6 wherethe nicotinamide is sandwiched between Pro201and Phe225 in a pocket formed by the pyro-phosphate binding segment, a helical turn and thearomatic ring of Phe225.6 The authors suggest thatthe structural changes necessary to stack thenicotinamide over the flavin ring require some dis-placement of the protein backbone as well asmovement of the side-chain of the Phe225.
Structural differences between free FNR andthe FNR:NADP1 complex
In spite of the good correlation between NADPþ-complexed and free FNR forms for the backbone,rearrangements of the regions involving residues221–239 (Ca rmsd of 0.30 A) and 261–298 (Ca
rmsd of 0.52 A) are observed upon NADPþ bind-ing (Figure 2(a)). The first region contains the longloop formed by residues 221–235, and the begin-ning of a small a-helix (236–243), which movestowards the adenine ring and the 20-P of thecoenzyme. With regard to the second region, a lessextended conformation of the loop 261–265 aswell as a displacement of a-helices D (residues266–281) and E (residues 285–295) are observed.A small displacement of the adenine moiety of theFAD is also observed. These backbone displace-ments are also associated with significant changesin conformers for some residues (Figure 2(b)).Thus, a more extended conformer is observed forArg100, which allows its guanidinium group tostabilise the pyrophosphate group of the coenzymewhile, at the same time, provides the necessaryspace to accommodate the ribose from the NMNmoiety of NADPþ. Displacement of the 221–239backbone region is associated with new con-formers for Ser223, Arg224, Arg233, Tyr235 andGln237. Arginine and serine side-chains movetowards the coenzyme in order to stabilise its 20-P(Figure 2(b)). A change in the x1 torsion angle ofTyr235 is also observed allowing both, a stackinginteraction with the adenine ring, and a polar inter-action with the 20-P. In contrast with the free FNRstructure, an ordered conformation for Gln237side-chain is observed as a consequence of anH-bond with the adenine. With regard to the261–298 region, the most important change is that
Coenzyme Recognition and Binding in FNR 1135
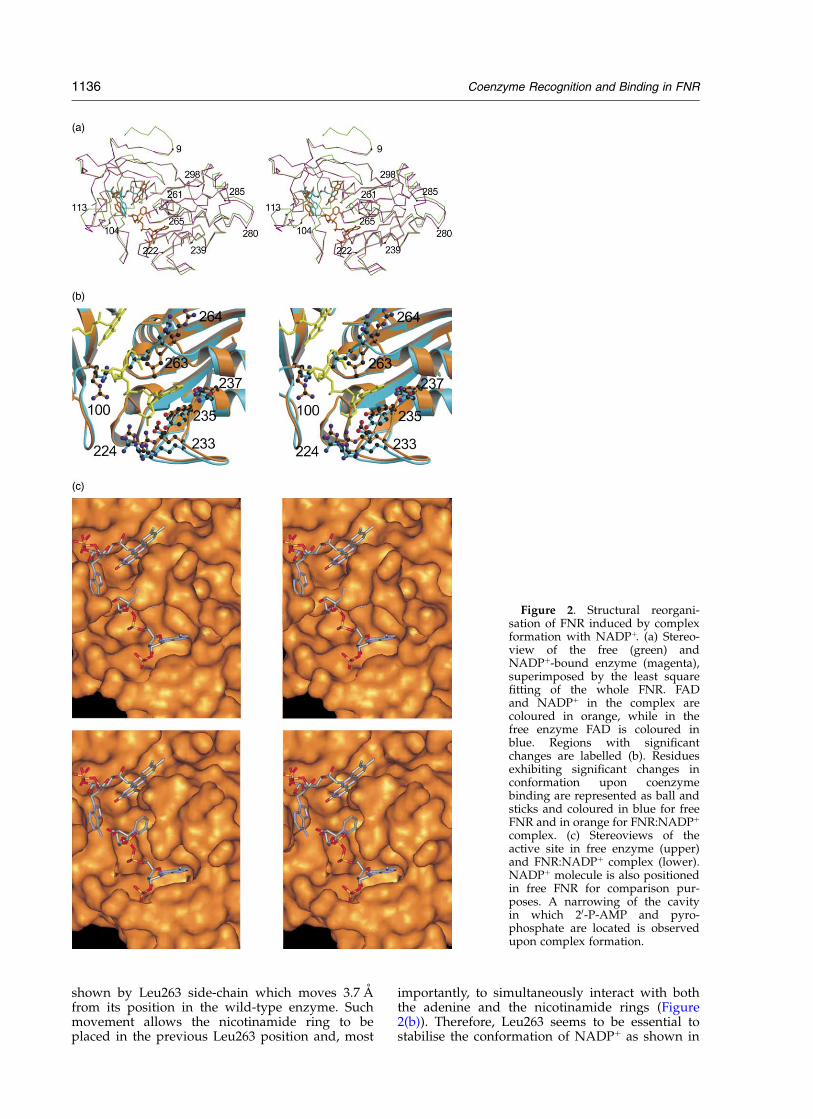
shown by Leu263 side-chain which moves 3.7 Afrom its position in the wild-type enzyme. Suchmovement allows the nicotinamide ring to beplaced in the previous Leu263 position and, most
importantly, to simultaneously interact with boththe adenine and the nicotinamide rings (Figure2(b)). Therefore, Leu263 seems to be essential tostabilise the conformation of NADPþ as shown in
Figure 2. Structural reorgani-sation of FNR induced by complexformation with NADPþ. (a) Stereo-view of the free (green) andNADPþ-bound enzyme (magenta),superimposed by the least squarefitting of the whole FNR. FADand NADPþ in the complex arecoloured in orange, while in thefree enzyme FAD is coloured inblue. Regions with significantchanges are labelled (b). Residuesexhibiting significant changes inconformation upon coenzymebinding are represented as ball andsticks and coloured in blue for freeFNR and in orange for FNR:NADPþ
complex. (c) Stereoviews of theactive site in free enzyme (upper)and FNR:NADPþ complex (lower).NADPþ molecule is also positionedin free FNR for comparison pur-poses. A narrowing of the cavityin which 20-P-AMP and pyro-phosphate are located is observedupon complex formation.
1136 Coenzyme Recognition and Binding in FNR

this complex. Thus, the combined effects describedfor backbone and side-chains movements producea narrower cavity in which the 20-P-AMP andthe pyrophosphate moieties are bound in such away that a perfect complementarity betweenthis coenzyme moiety and the protein surface isproduced (Figure 2(c)).
Comparison of different FNR:NADP1 complexes
Three-dimensional structures for FNR:NADPþ
complexes have been previously reported for theAnabaena and pea enzymes.3,19 In the case of theprevious Anabaena FNR:NADPþ complex (complexI), which was obtained by soaking native FNRcrystals in an NADPþ solution, no changes weredetected in the protein backbone with regard tothe free enzyme, and only slight rotations wereobserved for the side-chain of some residues inorder to approximate to the 20-P group.3 ComplexI only provided reliable binding information forthe 20-P-AMP coenzyme moiety, since the nicotin-amide appears pointing towards the molecular sur-face far away from the FAD isoalloxazine ring(Figure 3(a)). Thus, conformation of NADPþ in thecomplex I exhibited a more extended conformationthan that of the complex presented here (complexII). These changes have to be interpreted in lightof the different crystallisation methods used,suggesting that cocrystallisation allows the adap-tation between the FNR enzyme and the coenzymewithout any kind of steric impediment introducedby the preformed native crystal packing.
The three-dimensional structure of a complexbetween NADPþ and a pea FNR mutant in whichthe C-terminal Tyr residue has been replaced bySer and Trp, has also been reported19 (complexIII). In this complex, NADPþ presents a differentconformation from those described for complex Iand complex II (Figure 3(a)). Thus, in complex IIIthe pyrophosphate and 20-P-AMP portions ofNADPþ present a similar conformation to theanalogous moieties of the complex II and induceequivalent structural rearrangements in the proteinto stabilise them. Therefore, similar interactions ofNADPþ with residues of regions 261–265 and221–239 of the FNR are observed in pea complex(Figure 3(b)). However, in complex III removal ofthe terminal Tyr produces a different configurationof the NMN moiety and a more extended con-formation for the ribose. This conformationapproaches the nicotinamide group to the FADisoalloxazine ring and situates its C4 atom at only3 A from the N5 atom of the isoalloxazine ringallowing direct hydride transfer.
Insights into the mechanism of molecularrecognition and complex reorganisation forhydride transfer between FNR and NADP1
Although the existing structural data show threedifferent binding modes for the NADPþ–FNRinteraction, none of them represents a functionalmodel for the in vivo hydride transfer process.Thus, while in complexes I and II the relativedistance between the nicotinamide and the
Figure 3. Different NADPþ conformations in the FNR:NADPþ complexes. (a) Superposition of the two NADPþ con-formations observed for the different complexes onto the cocrystallised Anabaena FNR:NADPþ structure. Coenzymeconformation obtained by soaking3 is coloured in red (complex I), cocrystallised coenzyme is coloured in magenta(complex II) and NADPþ conformation obtained after mutation of the C-terminal Tyr in pea FNR19 is coloured ingreen (complex III). The FAD cofactor is coloured in blue. (b) Superposition of the cocrystallised Anabaena FNR:NADPþ
complex (orange) and the pea FNR:NADPþ complex (green). FAD and NADPþ cofactors are coloured in magenta forcomplex II and in cyan for complex III. Residues interacting with NADPþ in both structures are represented as balland sticks (see Figure 1(b) for labelling).
Coenzyme Recognition and Binding in FNR 1137
isoalloxazine rings is not adequate to allow thehydride transfer, the complex III can only beobtained upon removal of the terminal Tyr residue.However, this residue has also been proposed to beinvolved in the catalytic mechanism by modulatingpyridine nucleotide affinity.19,22 Nevertheless, noneof these structures excludes the others and can beinterpreted as different steps in molecular recog-nition and complex reorganisation to provide theadequate orientation for the hydride transfer.Thus, complex I might reflect the initial recognitionof NADPþ as an FNR coenzyme, through its20-P-AMP moiety and the 221–239 region of theprotein. The final coenzyme recognition is attainedwhen complex II is produced. Structural rearrange-ments in this second step are observed in the pro-tein chain in order to better stabilise the 20-P-AMPand the nicotinamide moieties. The regionsinvolved are the segments 221–239 and 261–298.Besides, a change in the conformation of Arg100 isessential to H-bond the NADPþ pyrophosphatebridge. These protein structural changes are alsoassociated with a change in the NADPþ moleculeconformation, which goes from the extendedconformation observed in complex I to the moretighten state observed in complex II. Finally, Tyrdisplacement would allow the nicotinamide ringto approach the isoalloxazine presenting theA-face of the NADPþ to the Re-face of the FAD, ashas been observed in complex III. (Figure 3(b)).Moreover, the movement of this Tyr must be essen-tial for catalysis, since it has been shown that itdown-modulates the strong affinity of the pro-
posed binding pocket shown in complex III for theNADPþ nicotinamide.19
Thus, the key point in this mechanism is howdoes the terminal Tyr get removed from its stack-ing position with the flavin ring. The influence ofFd in this event has been investigated by model-ling studies.4 However, the driving force for suchdisplacement still remains unknown, although thebiochemical and structural information availablepermits some hypothesis to be made. First of all,aromatic stacking interactions between activatedrings seem to be destabilising as deduced from themodel of p–p interactions proposed by Hunter &Sanders.23 Therefore, the stacking interactionbetween Tyr and isoalloxazine rings may even behindered upon FAD reduction. It is also worthnoting that Tyr303 is involved in a bifurcatedH-bond with Thr157 and a buried water moleculethat is highly conserved in all reported FNR struc-tures (Figure 1(c)). This water has been proposedas an alternative possible proton donor of Ser80to the isoalloxazine upon reduction.2 As a conse-quence, the proton transfer process might weakenthe Tyr303–water link and promote movement ofTyr303 towards Thr157. In addition, this movement(downwards in Figure 1(c)) is the only possibleway out for the Tyr303 side-chain, since FNR resi-dues prevent movement in the opposite direction.Moreover, the analysis of spectral perturbationsupon NADPþ binding to oxidised FNR shows thatthe nicotinamide ring is not able to alter theabsorption spectrum of the flavin ring of cyano-bacterial FNRs (including the Anabaena one),
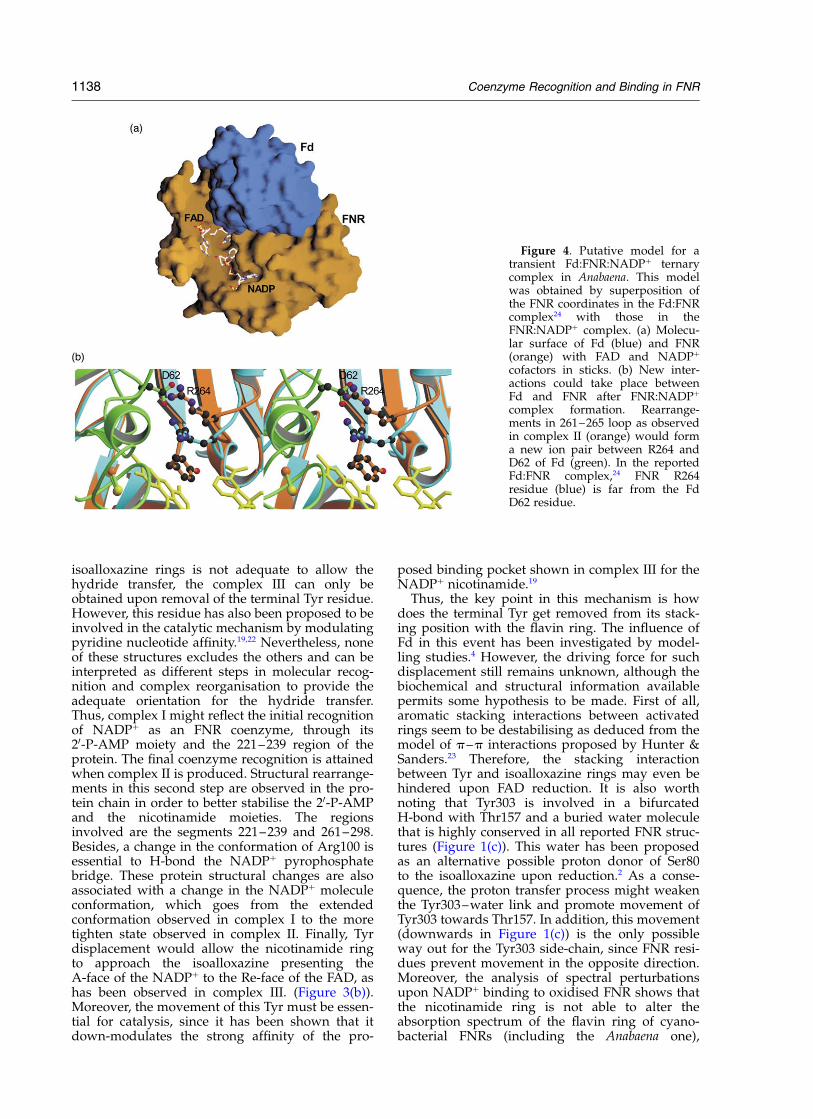
Figure 4. Putative model for atransient Fd:FNR:NADPþ ternarycomplex in Anabaena. This modelwas obtained by superposition ofthe FNR coordinates in the Fd:FNRcomplex24 with those in theFNR:NADPþ complex. (a) Molecu-lar surface of Fd (blue) and FNR(orange) with FAD and NADPþ
cofactors in sticks. (b) New inter-actions could take place betweenFd and FNR after FNR:NADPþ
complex formation. Rearrange-ments in 261–265 loop as observedin complex II (orange) would forma new ion pair between R264 andD62 of Fd (green). In the reportedFd:FNR complex,24 FNR R264residue (blue) is far from the FdD62 residue.
1138 Coenzyme Recognition and Binding in FNR

suggesting a weak interaction of the nicotinamidewith its “putative” binding site for hydride transferof these enzymes in their oxidised state.24 – 26 On thecontrary, the spectrum of FNRs from higher plantsresult clearly modified under the same conditions,pointing to an interaction between the nicotin-amide and the flavin group, which suggest somedifferences in this mechanism among higherplants and cyanobacterial FNRs.22,24 – 26 Therefore,all these pieces of structural and biochemicalinformation support the hypothesis that, at leastin the case of the Anabaena FNR, the Tyr303 dis-placement could be a direct consequence of boththe isoalloxazine reduction and the proton transferprocess.
Insights into the Fd:FNR:NADP1 ternarycomplex and the electron transfer process
Fast kinetic studies suggested that the FNR cata-lysed electron transfer from Fd to NADPþ proceedsthrough the formation of a transient ternarycomplex.12,13 Thus, the binding of the proteinsappears to be ordered for efficient electron transfer,with Fd binding to a preformed FNR:NADPþ
complex.12 Recently, the crystal structure of anAnabaena Fd:FNR complex has also beenreported,27 which showed no significant structuralmodifications in FNR upon Fd binding. Themolecular interface between Fd and FNR is formedby a hydrophobic core, a net of 11 hydrogen bondsand one ion pair.27 On the basis of this structureand that of the complex here presented, we havemodelled a putative Fd:FNR:NADPþ ternary com-plex (Figure 4). This model shows that the NADPþ
binding site on FNR is not close to the Fd:FNRinterface (Figure 4(a)) and, therefore, previousNADPþ binding to FNR should not affect the inter-action between the two proteins. However, uponNADPþ binding, structural rearrangements in the261–265 loop of FNR are observed that involvechanges in the conformation and orientation ofArg264, residue that has been shown to beinvolved in productive FNR:Fd interaction forelectron transfer,28 which moves in such a waythat a putative new ion pair could be formedbetween its side-chain and that of Asp62 from Fd(Figure 4(b)). This new link would be form througha Fd loop, that has been shown to contain criticalresidues for FNR binding (like Phe65) and also tobe involved in triggering Fdox release upon electrontransfer to FNR.27,29 Therefore, in the preformedFNR:NADPþ complex such a link between Arg264FNR and Asp62 Fd could provide a larger affinityfor reduced Fd than that exhibited by the freeenzyme. Moreover, since a displacement of theloop containing Asp62 in Fd upon reoxidation hasbeen related to the different affinity of FNR forreduced and oxidised ferredoxin,27 such interactionmight also being involved in oxidised Fd releasing.Further biochemical work should be done in orderto clarify this point.
Why is FNR an NADP1 specific enzyme?
Recent studies on Anabaena FNR indicate thatthe determinants of the high specificity shown bythis enzyme for NADPþ/H versus NADþ/H is notonly provided by those residues directly inter-acting with the 20-P, despite the fact that this is theonly difference between the two coenzymes.25,30
Moreover, these studies also pointed to otherregions of the protein that, although not interactingdirectly with the coenzyme, must undergo specificstructural arrangements of the backbone chain inorder to determine coenzyme specificity. Sequenceand structure analysis of different members of theFNR family with affinity for either NADPþ/H orNADþ/H, suggested that the region formed byresidues 155–161 of Anabaena FNR might beinvolved in such discrimination. Biochemical andstructural characterisation of an Anabaena FNRmutant in which, Thr155, (a residue conserved inall NADPþ/H-dependent members of the family)had been replaced by Gly (a residue present inall NADþ/H-dependent members) has beenreported.25 It was observed that this replacementproduced an important modification of the FNR261–265 loop. The equivalent region in theNADþ/H-dependent enzymes is rich in prolineresidues and therefore quite rigid.
Superposition of the NADPþ binding domain ofthe FNR:NADPþ complex presented here with thecorresponding domain of several NADþ/H-depen-dent members of the FNR family (phthalate dioxy-genase reductase, PDB code 2PIA, cytochrome b5
reductase, code 2CND and the NADH-dependentnitrate reductase, code 1NDH) shows a closerconformation for the 261–265 loop (see Figure 2)than that obtained when comparing with the freeFNR. In particular, a proline residue of the abovementioned Pro-rich loop in the NADþ/H-depen-dent enzymes is systematically placed at the sameposition that the critical Leu263 residue of theFNR:NADPþ complex. These observations clearlysuggest different mechanisms for coenzyme recog-nition and complex reorganisation in FNR, andtherefore in NADPþ/H-dependent members of theFNR family relative to the NADþ/H-dependentmembers. Thus, in the case of the FNR, it seemsthat the free enzyme presents a large cavity toaccommodate the 20-P-AMP moiety of the coen-zyme, which upon its binding is reorganised inorder to match perfectly the charge and shape ofthe adenine portion of the substrate. However, inthe case of the NADþ/H-dependent members,such a narrow cavity is already preformed in thefree enzyme and probably does not need toundergo important structural rearrangements inorder to adapt the adenine moiety of NADþ/H asit has been described in the case of PDR:NADHcomplex.6 In summary, the coenzyme specificity inthe FNR family can be explained as a consequenceof the nature of both, the residues interactingdirectly with the 20P phosphate and the residuesshaping the pocket that accommodates the AMP.
Coenzyme Recognition and Binding in FNR 1139

Nevertheless, further mutagenesis studies on theFNR family will clarify the subtle structuralfeatures that confer coenzyme specificity.
Conclusions
In the photosynthetic electron transfer chaininvolved in converting the energy of the lightinto chemical power, under the form of NADPHreducing equivalents, FNR catalyses the flavin-mediated two-electron transfer from two ferre-doxin molecules to NADPþ. The FNR structure,formed by two domains, one for binding the FADand the other for binding NADPþ, has provideda structural prototype for a large family offlavoenzymes.
Here, we present the crystal structure of anFNR:NADPþ complex from Anabaena that hasbeen obtained by cocrystallisation. This structurediffers from those previously reported forFNR:NADPþ complexes in Anabaena and peaenzymes, the main difference being the relativeposition of the NADPþ nicotinamide ring withinthe complex. However, these different NADPþ
conformations do not exclude each other, and
together reveal new insights into the catalyticmechanism of FNR, suggesting a model forcoenzyme recognition and complex reorganisationfor an efficient hydride transfer to occur. Finally,structural comparison between the FNR familyprovides an explanation for the high specificityexhibited by FNR for NADPþ/H versus NADþ/Hthat could be extended to the rest of members ofthis family.
Materials and Methods
Expression and purification of FNR
Recombinant FNR was purified from IPTG-inducedEscherichia coli LB cultures containing the correspondingconstruct of the Anabaena pet H gene as previouslydescribed.25
Crystallisation and data collection
The FNR:NADPþ crystals were grown by cocrystalli-sation using the hanging drop method. The 5 ml dropletsconsisted of 1 ml of 27 mg protein/ml solution bufferedwith 10 mM Tris/HCl (pH 8.0), 1 ml of 10 mM NADPþ,1 ml of unbuffered bOG at 5% (w/v) and 2 ml of reservoirsolution containing 20% (w/v) PEG6000 and 0.1 Msodium acetate (pH 5.0). The droplet was equilibratedagainst 1 ml reservoir solution at 20 8C. Under theseconditions crystals grew within 1–15 days as smallneedles and only very few experiments led to prismsthat increased the maximum size to 0.8 mm £ 0.3 mm£ 0.2 mm, suitable for X-ray diffraction.
X-ray data for these crystals were collected at 100 K ona Mar Research 345 (Germany) IP area detector usinggraphite-monochromated CuKa radiation generated byan Enraf–Nonius rotating anode generator. Crystalsbelong to the I41 tetragonal space group (a ¼ b ¼151.69 A and c ¼ 35.44 A). The VM is 3.0 A3 Da21 withone FNR molecule in the asymmetric unit and 59%solvent content. The X-ray data set was processed withMOSFLM31 and scaled and reduced with SCALA fromthe CCP432 up to 2.1 A resolution (Table 1).
Structure determination and refinement
The FNR:NADPþ structure was solved by molecularreplacement using the program AMoRe33 on the basis ofthe 1.8 A resolution native FNR model.3 An unambigu-ous single solution for the rotation and translationfunctions was obtained. The model was subjected toalternate cycles of conjugate gradient refinement withthe program X-PLOR34 and manual model buildingwith the software package O.35 The crystallographic R ofan Rfree
36 values converged to values of 0.20 and 0.27,respectively, for reflections between 30 A and 2.1 A reso-lution. The resulting electron density was of great qualityas can be seen for the case of the NADPþ molecule(Figure 5). The final model contains 2334 non-hydrogenprotein atoms, one NADPþ, one FAD and 200 solventmolecules. The model exhibits good stereochemistry,with rmsd from ideality of 0.011 A for bond lengthsand 1.28 for bond angles (see Table 1). Pictures weregenerated with O,35 GRASP,37 MOLSCRIPT38 andRASTER.39
Table 1. Structure determination statistics
Crystal data
Space group (I41)a (A) 151.69b (A) 151.69c (A) 35.44
Data collectiona
Wavelength (A) 1.5418Resolution (A) 2.1 (2.21–2.1)Unique data 23,930Redundancy 3.8 (3.8)Completeness (%) 99.5 (99.5)I/s(I ) 4.4 (1.9)Rsym (%)b 0.10 (0.38)
RefinementResolution range (A) 29.7–2.1Rwork (Rfree)
c 0.20 (0.27)Rmsdd bonds (A) 0.011Rmsd angles (8) 1.239
Quality of Ramachandran plote
Percentage of residues inmost favoured regions
89.1
Percentage of residues inadditional allowed regions
10.9
Percentage of residues ingenerously allowed regions
0.0
Number of atoms (average B-values (A2))Protein 2334 (32.5)Water 200 (39.7)NADPþ 48 (35.8)
a Values in parentheses correspond to the highest resolutionshell.
b Rsym ¼P
lI 2 Iavl=P
I; where the summation is over sym-metry equivalent reflections.
c R calculated on 7% of data excluded from refinement.d Root-mean-square deviation.e Figures from PROCHECK.
1140 Coenzyme Recognition and Binding in FNR

Protein Data Bank accession numbers
The coordinates and structure factors for theFNR:NADPþ complex have been deposited in theProtein Data Bank and were released with the date ofpublication (accession number 1GJR).
Acknowledgments
We are grateful to Dr J. K. Hurley and ProfessorG. Tollin (University of Arizona) and to Dr Martınez-Ripoll (CMBE, Madrid) for their collaboration in discuss-ing different aspects of this work. This work was sup-ported by grant BIO97-0912C02-01 from the CICYT toC.G.-M. and by grant P006/2000 from CONSIDþDGAto M.M.
References
1. Karplus, P. A., Daniels, M. J. & Herriott, J. R. (1991).Atomic structure of ferredoxin–NADPþ reductase:prototype for a structurally novel flavoenzymefamily. Science, 251, 60–66.
2. Bruns, C. M. & Karplus, P. A. (1995). Refined crystalstructure of spinach ferredoxin reductase at 1.7 Aresolution: oxidized, reduced and 20-phospho-50-AMP bound states. J. Mol. Biol. 247, 125–145.
3. Serre, L., Vellieux, F. M. D., Medina, M., Gomez-Moreno, C., Fontecilla-Camps, J. C. & Frey, M.(1996). X-ray structure of the ferredoxin:NADPþ
reductase from the cyanobacterium Anabaena PCC7119 at 1.8 A resolution, and crystallographic studiesof NADPþ binding at 2.25 A resolution. J. Mol. Biol.263, 20–39.
4. Dorowski, A., Hofmann, A., Steegborn, C., Boicu, M.& Huber, R. (2001). Crystal structure of paprikaferredoxin–NADPþ reductase. Implications forthe electron transfer pathway. J. Biol. Chem. 276,9253–9263.
5. Correll, C. C., Ludwig, M. L., Bruns, C. M. &Karplus, P. A. (1993). Structural prototypes for anextended family of flavoprotein reductases: compari-son of phthalate dioxygenase reductase with ferre-doxin reductase and ferredoxin. Protein Sci. 2,2112–2133.
6. Correll, C. C., Batie, C. J., Ballou, D. P. & Ludwig,M. L. (1992). Phthalate dioxygenase reductase: amodular structure for electron transfer from pyridinenucleotide to [2Fe–2S]. Science, 258, 1604–1610.
7. Lu, G., Campbell, W. H., Schneider, G. & Lindqvist,Y. (1994). Crystal structure of the FAD-containingfragment of corn nitrate reductase at 2.5 A reso-lution: relationship to other flavoprotein reductases.Structure, 2, 809–821.
8. Nishida, H., Inaka, K., Yamanaka, M., Kaida, S.,Kobayashi, K. & Miki, K. (1995). Crystal structure ofNADH-cytochrome b5 reductase from pig liver at2.4 A resolution. Biochemistry, 34, 2763–2767.
9. Wang, M., Roberts, D. L., Paschke, R., Shea, T. M.,Masters, B. S. & Kim, J. J. (1997). Three-dimensionalstructure of NADPH-cytochrome P450 reductase:prototype for FMN- and FAD-containing enzymes.Proc. Natl Acad. Sci. USA, 94, 8411–8416.
10. Ingelman, M., Bianchi, V. & Eklund, H. (1997). Thethree-dimensional structure of flavodoxin reductasefrom Escherichia coli at 1.7 A resolution. J. Mol. Biol.268, 147–157.
11. Gruez, A., Pignol, D., Zeghouf, M., Coves, J., Fonte-cave, M., Ferrer, J. L. & Fontecilla-Camps, J. C.(2000). Four crystal structures of the 60 kDa flavopro-tein monomer of the sulfite reductase indicate adisordered flavodoxin-like module. J. Mol. Biol. 299,199–212.
12. Arakaki, A. K., Ceccarelli, E. A. & Carrillo, N. (1997).Plant-type ferredoxin–NADPþ reductases: a basalstructural framework and a multiplicity of functions.FASEB J. 11, 133–140.
13. Batie, C. J. & Kamin, H. (1984). Electron transferby ferredoxin:NADPþ reductase. Rapid-reactionevidence for participation of a ternary complex.J. Biol. Chem. 259, 11976–11985.
14. Karplus, P. A. & Schulz, G. E. (1989). Substrate bind-ing and catalysis by glutathione reductase as derivedfrom refined enzyme: substrate crystal structures at2 A resolution. J. Mol. Biol. 210, 163–180.
15. Pai, E., Karplus, P. A. & Schulz, G. E. (1988). Crystal-lographic analysis of the binding of NADPH,NADPH fragments, and NADPH analogues toglutathione reductase. Biochemistry, 27, 4465–4474.
16. Stehle, T., Claiborne, A. & Schulz, G. E. (1993).NADH binding site and catalysis of NADH peroxi-dase. Eur. J. Biochem. 211, 221–226.
17. Li, R., Bianchet, M. A., Talalay, P. & Amzel, L. M.(1995). The three-dimensional structure ofNAD(P)H: quinone reductase, a flavoproteininvolved in cancer chemoprotection and chemo-therapy: mechanism of the two-electron reduction.Proc. Natl Acad. Sci. USA, 92, 8846–8850.
18. Chiu, H.-J., Johnson, E., Schroder, I. & Rees, D.(2001). Crystal structures of a novel ferric reductasefrom the hyperthermophilic archaeon Archaeoglobusfulgidus and its complex with NADPþ. Structure, 9,311–319.
19. Deng, Z., Aliverti, A., Zanetti, G., Arakaki, A. K.,Ottado, J., Orellano, E. G. et al. (1999). A productiveNADPþ binding mode of ferredoxin–NADPþ
reductase revealed by protein engineering and crys-tallographic studies. Nature Struct. Biol. 6, 847–853.
20. Mayoral, T., Medina, M., Sanz-Aparicio, J., Gomez-Moreno, C. & Hermoso, J. A. (2000). Structural basisof the catalytic role of Glu301 in Anabaena PCC 7119ferredoxin–NADPþ reductase revealed by X-raycrystallography. Proteins: Struct. Funct. Genet. 38,60–69.
Figure 5. Stereoview of the electron density map forthe complete NADPþ molecule. The 2.1 A resolution2Fo–Fc map was calculated using phase informationfrom the final model and contoured at 1.5 srms.
Coenzyme Recognition and Binding in FNR 1141

21. Martinez-Julvez, M., Nogues, I., Faro, M., Hurley,J. K., Brodie, T. B., Mayoral, T. et al. (2001). Role ofa cluster of hydrophobic residues near the FADcofactor in Anabaena PCC 7119 ferredoxin–NADPþ
reductase for optimal complex formation andelectron transfer to ferredoxin. J. Biol. Chem. 276,27498–27510.
22. Piubelli, L., Aliverti, A., Arakaki, A. K., Carrillo, N.,Ceccarelli, E. A., Karplus, P. A. & Zanetti, G. (2000).Competition between C-terminal tyrosine and nico-tinamide modulates pyridine nucleotide affinity andspecificity in plant ferredoxin–NADP(þ ) reductase.J. Biol. Chem. 275, 10472–10476.
23. Hunter, C. A. & Sanders, J. K. M. (1990). The nature ofp–pi; interactions. J. Am. Chem. Soc. 112, 5525–5534.
24. Sancho, J. & Gomez-Moreno, C. (1991). Interaction offerredoxin–NADPþ reductase from Anabaena with itssubstrates. Arch. Biochem. Biophys. 288, 231–238.
25. Medina, M., Luquita, A., Tejero, J., Hermoso, J.,Mayoral, T., Sanz-Aparicio, J. et al. (2001). Probingthe determinants of coenzyme specificity in ferre-doxin–NADPþ reductase by site-directed mutagen-esis. J. Biol. Chem. 276, 11902–11912.
26. Medina, M., Martınez-Julvez, M., Hurley, J. K., Tollin,G. & Gomez-Moreno, C. (1998). Involvement ofglutamic acid 301 in the catalytic mechanism of ferre-doxin–NADPþ reductase from Anabaena PCC 7119.Biochemistry, 37, 2715–2728.
27. Morales, R., Charon, M.-H., Kachalova, G., Serre, L.,Medina, M., Gomez-Moreno, C. & Frey, M. (2000). Aredox-dependent interaction between two electron-transfer partners involved in photosynthesis. EMBORep. 1, 271–276.
28. Martınez-Julvez, M., Hermoso, J., Hurley, J. K.,Mayoral, T., Sanz-Aparicio, J., Tollin, G. et al. (1998).Role of Arg100 and Arg264 from Anabaena PCC7119 ferredoxin–NADPþ reductase for optimalNADPþ binding and electron transfer. Biochemistry,37, 17680–17691.
29. Hurley, J. K., Cheng, H., Xia, B., Markley, J. L.,Medina, M., Gomez-Moreno, C. & Tollin, G. (1993).
An aromatic amino acid is required at position 65 inAnabaena ferredoxin for rapid electron transfer toferredoxin–NADPþ reductase. J. Am. Chem. Soc. 115,11698–11701.
30. Nishida, H., Inaka, K., Yamanaka, M., Kaida, S.,Kobayashi, K. & Miki, K. (1995). Crystal structure ofNADH-cytochrome b5 reductase from pig liver at2.4 A resolution. Biochemistry, 34, 2763–2767.
31. Leslie, A. G. W. (1992). Molecular data processing.In Crystallographic Computing 5: From Chemistry toBiology (Moras, D., Podjarny, A. D. & Thierry, J. C.,eds), pp. 39–50, Oxford University Press, Oxford,UK.
32. Collaborative Computational Project Number 4(1994). The CCP4 suite: programs for protein crystal-lography. Acta Crystallog. sect. D, 50, 760–763.
33. Navaza, J. (1994). AMoRe: an automated package formolecular replacement. Acta Crystallog. sect. A, 50,157–163.
34. Brunger, A. T. (1993). X-PLOR: A System for X-rayCrystallography and NMR, Yale University Press,New Haven, CT.
35. Jones, T. A., Zou, J. Y., Cowan, S. W. & Kjeldgaard,M. (1991). Improved methods for building proteinmodels in electron density maps and the location oferrors in these models. Acta Crystallog. sect. A, 47,110–119.
36. Brunger, A. T. (1992). The free R value: a novelstatistical quantity for assessing the accuracy ofcrystal structures. Nature, 355, 472–474.
37. Nicholls, A. & Honig, B. (1991). A rapid finite dif-ference algorithm, utilizing successive over-relax-ation to solve the Poisson–Boltzmann equation.J. Comp. Chem. 12, 435–445.
38. Kraulis, P. J. (1991). MOLSCRIPT: a program to pro-duce both detailed and schematic plots of proteinstructures. J. Appl. Cryst. 24, 946–950.
39. Merritt, E. A. & Bacon, D. J. (1997). Raster3D: photo-realistic molecular graphics. Methods Enzymol. 277,505–524.
Edited by R. Huber
(Received 22 January 2002; received in revised form 3 April 2002; accepted 12 April 2002)
1142 Coenzyme Recognition and Binding in FNR




![Structure of a [2Fe–2S] ferredoxin from Rhodobacter capsulatus likely involved in Fe–S cluster biogenesis and conformational changes observed upon reduction](https://static.fdokumen.com/doc/165x107/63363fcacd4bf2402c0b6cfb/structure-of-a-2fe2s-ferredoxin-from-rhodobacter-capsulatus-likely-involved.jpg)


![A Flexible Copper(I)-Complexed [4]Rotaxane Containing Two Face-to-Face Porphyrinic Plates that Behaves as a Distensible Receptor](https://static.fdokumen.com/doc/165x107/632a7148d2eed9476207508b/a-flexible-copperi-complexed-4rotaxane-containing-two-face-to-face-porphyrinic.jpg)












