
Glutamate receptor expression in rat striatum: Effect of deafferentation

Plant Molecular Biology 27: 115-128, 1995. © 1995 Kluwer Academic Publishers. Printed in Belgium. 115
Expression of ferredoxin-dependent glutamate synthase in dark-grown pine seedlings
Angel Garcia-Guti6rrez, Francisco R. Cantdn, Fernando Gallardo, Francisca Sfinchez-Jim6nez and Francisco M. Cfinovas* Laboratorio de Bioquimica y Biologia Molecular, Facultad de Ciencias, Universidad de Mdlaga, Campus de Teatinos, E-29071, Mdlaga, Spain (*author for correspondence)
Received 26 July 1994; accepted in revised form 18 October 1994
Key words: Pinus, gymnosperm, conifer, amino acid biosynthesis, glutamate synthase, cDNA cloning, light regulation
Abstract
Pine seedlings are able to accumulate chlorophylls and develop green plastids in a light-independent manner. In this work, we have characterized ferredoxin-dependent glutamate synthase (EC 1.4.7.1; Fd-GOGAT), a key enzyme in nitrogen interconversion during this process. Fd-GOGAT has been purified about 170-fold from cotyledons of maritime pine (Pinus pinaster). As occurs in angiosperms, the native enzyme is a single polypeptide with an apparent molecular mass of 163-168 kDa that is confined to the chloroplast stroma. Polyclonal antibodies generated against the purified enzyme were used to immunoscreen a 2gtl 1 expression library from Scots pine (Pinus sylvestris) seedlings and partial cDNA clones were isolated and characterized. The clone with the longest cDNA insert (pGOP44) contained the codification for the C-terminal (550 amino acids) of the pine Fd-GOGAT polypeptide. Immuno- logical cross-reactivity and comparative amino sequence analysis revealed that Fd-GOGAT is a well conserved protein in higher plants. Western blot analyses showed that protein was expressed in chloroplast-containing pine tissues and this expression pattern was not affected by exogenously supplied nitrogen. Fd-GOGAT mRNA, polypeptide and enzyme activity accumulated in substantial amounts in dark-grown pine seedlings. The presence of a functional Fd-GOGAT may be important to provide the required glutamate for the biosynthesis of nitrogen compounds during chloroplast biogenesis in the dark.
Introduction
Incorporation of inorganic nitrogen into amino acids in plants occurs through the glutamate syn- thase metabolic pathway, which implies the se- quential action of glutamine synthetase (GS; EC 6.3.1.2) and glutamate synthase (GOGAT; EC 1.4.1.14 and EC 1.4.71). Glutamine and
glutamate are the amino donors for the biosyn- thesis of major nitrogen compounds in higher plants such as proteins, nucleic acids and chlo- rophylls. GOGAT exists in higher plants as two molecular forms that differ in their respective source of reductant: NADH-GOGAT and ferre- doxin (Fd)-GOGAT. In angiosperms, Fd- GOGAT is a chloroplast-located enzyme [8] rep-

116
resenting the predominant molecular form in green tissues, where it is involved in the assimi- lation of ammonia derived from nitrate reduction and the reassimilation of ammonia released in photorespiration and protein catabolism [6, 27]. Developmental regulation of Fd-GOGAT has been studied during greening of etiolated leaves [38, 41] and its expression is affected by nitrogen supply [36]. Based on the different physico- chemical, immunological and regulatory proper- ties of both enzymes in a number of higher plants, it is assumed that they are distinct proteins [27, 42]. This assumption has been recently con- firmed by the isolation and characterization of different cDNA clones encoding Fd-dependent [ 5, 38, 49] and NADH-dependent [23 ] glutamate synthases.
Chlorophyll biosynthesis and accumulation of gene products associated with photosynthesis are regulated by light in angiosperms [16, 44]. How- ever, it is well documented that pine and other gymnosperm species are able to accumulate pho- tosynthetic pigments and develop chloroplasts even when growing in darkness [7, 29]. In several pine species, the light-independent synthesis of LHCP II (apoprotein of the light-harvesting com- plex of photosystem II) and other chloroplast polypeptides such as LSU and SSU (large and small subunits of ribulose-l,5-bisphosphate carboxylase/oxygenase) have been reported [47, 11]. Since these plants are able to develop chloroplasts in the dark, we hypothesized that the nitrogen-assimilating enzymes GS and Fd- GOGAT, key enzymes in the nitrogen economy of the developing seedling, could also accumulate in darkness maintaining the appropriate glutamine and glutamate levels for the construc- tion of the photosynthetic apparatus.
In a previous work we examined the physiology of glutamine synthetase during early development in pine and reported the molecular characteriza- tion of a full-length cDNA clone encoding the enzyme subunit [ 12, 15]. In this paper we report, for the first time, the purification and molecular characterization of Fd-GOGAT in a gymno- sperm species. Polyclonal antibodies raised against the purified protein have been used as
molecular tools for studying protein expression in pine and to isolate cDNA clones by immuno- screening. We present experimental evidence demonstrating light-independent gene expression in pine seedlings.
Materials and methods
Plant material
Conifer seeds (Pinus pinaster, P. halepensis, P. sylvestris, P. pinea, Larix decidua, A bies pinsapo ) were supplied by Instituto Nacional para la Con- servacidn de la Naturaleza, Madrid, Spain. The seeds were imbibed for three days and germinated in moistened vermiculite at 20 ° C. Experimental conditions for plant growth and the criterion for selection of seedling developmental stages have been described elsewhere [12].
Enzyme assay and protein determination
Fd-GOGAT activity was determined according to Gallardo etal. [19] with spinach Ferredoxin (Sigma, St. Louis, MI) as electron donor. Protein concentration was estimated by Bradford's pro- cedure [ 10].
Protein purification
Pine cotyledons (55g) were homogenized in buffer A (60 mM potassium phosphate, 5 mM EDTA, 12.5 mM 2-mercaptoethanol, 2 mM 2-oxoglutarate pH 7.3) supplemented with 20~o (v/v) ethylene glycol, 0.05 ~o (v/v) Triton-X-100. The extract was filtered through two layers of muslin, centrifuged (150000 x g, 30 min) and the supernatant fractionated with ammonium sul- phate (20-60~o) (w/v). Proteins were resus- pended in a minimal volume of buffer A, dialysed and loaded onto a DEAE-cellulose column (4 cm × 20 cm) equilibrated in the same buffer. After washing with buffer A (75 ml/h), Fd- GOGAT activity was detected in the exclusion

volume. Protein was again concentrated by am- monium sulphate precipitation, and then resus- pended in buffer B (20 mM potasium phosphate, 5 mM EDTA, 12.5 mM 2-mercaptoethanol, 2 mM 2-oxoglutarate pH 7.3). The new extract (6 ml) was applied to a DEAE-Sephacel column (1.2 cm x 20 cm) equilibrated in buffer B. Enzyme activity was eluted by the application of a linear gradient 0-0.2 M KC1 in buffer B, at a flow rate of 50 ml/h. Active fractions were pooled, concen- trated by ammonium sulphate precipitation and applied to a phenyl-Sepharose column equili- brated in buffer A supplemented with 25 ~o (w/v) of ammonium sulphate. Column was twice washed, initially with 200 ml of the equilibration buffer, and then with an equal volume of buffer A. Fd-GOGAT activity was eluted with a linear 0-70~o (v/v) ethylene glycol gradient prepared in buffer A (35 ml/h). Active fractions were con- centrated and stored at 4 °C. Fd-GOGAT polypeptide was monitored by SDS-PAGE and enzyme activity assays throughout the purifica- tion steps.
Antibody production and western blotting
Further purification of the active fractions from the phenyl-Sepharose chromatography was car- ried out by preparative SDS-PAGE before rab- bit immunization as described for the preparation of other antisera in our laboratory [20]. In our previous article, we have also reported the immu- nization protocol and polyclonal antibody purifi- cation procedures, which were adapted for Fd- GOGAT in the present work. Western blot analysis were performed essentially as described elsewhere [12] using a 1:8000 dilution of crude antiserum.
Chloroplast isolation
Chloroplasts were isolated from light-grown pine cotyledons by differential centrifugation and fur- ther purified following the protocol of Joy and Mills [25 ].
117
Isolation of cDNA clones and sequencing
A cDNA library from P. sylvestris seedlings [26] constructed in the 2gtl l expression vector was screened with purified Fd-GOGAT polyclonal antibodies, using the method reported by Ausubel et al. [2] with minor modifications. Nitrocellulose filters were processed for immunoscreening fol- lowing the same protocol mentioned above for western blotting, except that purified IgGs (1:200 dilution) were used. Positive plaques were iso- lated and the cDNA inserts subcloned into pGEM 3Z plasmid (Promega, Madison, WI). Rescreening of the library with double-stranded DNA probes was carried out as described else- where [15]. Nucleotide sequences of cDNA clones were determined by the dideoxy termina- tion method of Sanger et al. [40] using T7 DNA polymerase (Sequenase, USB, Cleveland, OH). The sequencing of both strands was performed after multiple digestion of the inserts and con- struction of overlapping recombinant plasmids. Other basic recombinant DNA procedures were carried out following the standard protocols de- scribed by Sambrook et al. [39]. Compilation of sequencing data and comparative analysis of amino acid sequences were carried out by using the Genetics Computer Group software (Univer- sity of Wisconsin) [ 17].
Northern blot analysis
Total RNA was extracted from pine seedlings, electrophoresed, transferred to nylon membranes and hybridized with the 32p-labelled cDNA in- serts as described by Cantdn et al. [ 15].
Results
Protein purification and antibody production
The purification protocol developed for obtaining partially purified extracts of Fd-GOGAT from pine cotyledons is summarized in Table 1 and Fig. 1, panel a. This protocol used hydrophobic

118
Table 1. Purification of Fd-GOGAT from pine cotyledons.
Step Total activity, Yield, Protein, Specific activity, Purification, nkat ~o mg nkat/mg fold
Crude extract 443 100 559 0.8 1 20-60 ~o (NH4)2SO 4 379 86 301 1.3 1.6 DEAE-Cellulose 287 65 101 2.8 3.5 DEAE-Sephacel 129 29 5.2 24.8 31 Penyl-Sepharose 68 15 0.5 136 170
chromatography on phenyl-Sepharose as a basic step, that has been described as an alternative method for the rapid purification of ferredoxin- dependent enzyme [21, 37]. The sequential puri- fication steps led to a final preparation which ex- hibited a specific ferredoxin-dependent activity of 136 nkat/mg of protein with a purification degree of about 170-fold. The purification procedure was completed in five working days. After the purifi- cation procedure, the pine extract Was enriched in a major polypeptide of 168 kDa and two minor abundant polypeptides of 90 and 70 kDa, when analysed by SDS-PAGE (Fig. la, lanes 1-4). The 168 kDa band represented about 70~o of
total protein after the phenyl-Sepharose step, and its relative abundance matched the Fd-GOGAT activity profiles in the different purification steps. The molecular sizes of Fd-GOGAT polypeptide in angiosperms ranges from 140 to 180kDa [30, 49]. The identification of the 168 kDa as the Fd-GOGAT subunit was confirmed by native PAGE of the purified fraction followed by en- zyme extraction, activity measurement and pro- tein staining (results not shown); maximum en- zyme activity was coincident with the major protein band in the gel. The additional faster mi- grating bands observed in the final preparation could represent either degradation products of
Fig. 1. Electrophoretic analysis of Fd-GOGAT purification and antibody specificity. (a) Samples of active fractions from each purificatin step were subjected to 8~o SDS/PAGE separation and polypeptides stained with Coomassie blue. Lane 1, 20-60~o ammonium sulphate (40 #g); lane 2, DEAE-cellulose (40/~g); lane 3, DEAE-Sephacel (20/~g); lane 4, Phenyl-Sepharose (10 #g). (b) Phenyl-Sepharose purified fractions, (lanes 1 and 2) and crude extract preparations (lanes 3 and 4) were separated by SDS/ PAGE, transferred to nitrocellulose filters and immunoassayed with Fd-GOGAT antibodies. Amounts of protein loaded per lane were: 10 #g (lane 1), 5 #g (lane 2), 60 #g (lane 3) and 30 #g (lane 4). The positions of the molecular size markers are indicated on the left. The position of the intact Fd-GOGAT polypeptide is marked by the arrow on the right.

119
the purified enzyme or contaminant polypeptides that were co-purified with the protein (see below). Taking into account the differences in the mo- lecular size between G O G A T polypeptide and the two major additional bands, a final attempt to further purification of the 168 kDa band was car- ried out by FPLC (fast protein liquid chromato- graphy) gel filtration, but the results were unsuc- cessful.
Molecular mass of native pine Fd -GOGAT was estimated to be about 163 kDa as judged by gel filtration chromatography of the purified preparation through a Sephacryl S-300 column calibrated with proteins of a known size (results not shown). This value was very close to that of the band observed under denaturing conditions and, therefore, it suggests that native Fd -GOGAT in gymnosperms is a monomeric protein, as oc- curs in angiosperms [27].
F d - G O G A T subunit (168 kDa) was further purified by preparative SDS-PAGE, and poly- clonal antibodies were elicited against the polypeptide in rabbits. The specificity of the antiserum was tested by Western blot analysis of purified and crude extracts. As shown in Fig. 1 (panel b, lanes 1 and 2) the antiserum strongly recognized the Fd -GOGAT polypeptide (168 kDa) as well as the 90 and 70 kDa contami- nating bands in a minor extent. In contrast, only one polypeptide was immunodetected in freshly prepared pine crude extracts which corresponded to the expected size of intact Fd-GOGAT- polypeptide (Fig. lb, lanes 3 en 4). Thus, in our opinion, 90 and 70 kDa bands could represent breakdown products generated by handling of the enzyme through the purification procedure. These results are similar to those reported by Sakaki- bara et al. [38] for the maize enzyme, which is proteolytically cleaved in two fragments of related sizes during the purification process.
The antigenic similarities between F d - G O G A T proteins in gymnosperms were studied by using the antiserum prepared against the maritime pine protein. GOGAT was extracted from several spe- cies and equal amounts of total protein were sub- j ected to S D S-PAGE, blotted onto nitr0c ellulo s e and immunoassayed with pine G O G A T antise-
rum. As shown in Fig. 2, pine G O G A T antibodies cross-reacted with polypeptides of a similar size in several coniferous species including other pine species as well as larch and pinsapo fir. These data suggest that Fd -GOGAT protein is a con- served enzyme in this group of plants. The dif- ferences observed in the intensity of the ilnmu- noreactive signals may reflect differences in the antigenicity of the proteins. Nevertheless, the same antibodies cross-reacted with a 150 kDa band in crude extracts of tomato leaves and com- parable signals were detected (A. P6rez-Garcia et al., manuscript submitted). This fact suggests that the observed differences could also be due to the relative abundance of the G O G A T polypep- tide in each preparation.
Localization of Fd-GOGA T in pine chloroplasts
In angiosperms, it is well established that Fd- GOGAT is a chloroplast-located enzyme [ 8 ] that, in coordination with chloroplastic glutamine syn- thetase (GS2), participates in the assimilation of ammonia derived from nitrate reduction and pho-
Fig. 2. Immunochemicalcross-reactivityofFd-GOGAT anti- bodies with polypeptides of several gymnosperms. Total polypeptides were prepared from cotyledons of several spe- cies, separated by SDS-PAGE (40 #g of protein per lane), electrotransferred to nitrocellulose membranes and probed with Fd-GOGAT antiserum. Lane 1, Pinus pinaster; lane 2, Pinus pinea; lane 3, Pinus halepensis; lane 4, Pinus sylvestris; lane 5, Abiespinsapo; lane 6, Larix decidua. The molecular size of the major immunoreactive band is indicated on the right.

120
torespiration [31, 42]. However, during early de- velopment of pine seedlings, GS2 has not been detected and cytosolic GS (GS 1) seems to be the predominant GS isoform [12, 15]. Based on the above data, we decided to study the subcellular distribution of Fd -GOGAT polypeptide in pine seedlings (Fig. 3). Chloroplasts were isolated from light-grown plants (cotyledon 2 cm in length) by differential centrifugation and rapid purifica- tion through Percoll gradients. Isolated organella were fractionated into stroma and thylakoid membranes. Total (T), soluble (S) and membrane (M) proteins were extracted, separated by SDS- PAGE, electrotransferred to nitrocellulose filters and stained with Coomassie blue (Fig. 3a) or probed with specific antibodies (Fig. 3b). As mo- lecular markers for polypeptide localization, we detected the large (LSU) and small (SSU) sub- units of ribulose 1,5-bisphosphate carboxylase/ oxygenase (the most abundant protein in the stroma of the plastid) and LHCP II (a major
membrane protein in the thylakoids). Chloroplast polypeptides were immunodetected on nitrocel- lulose filters with specific antibodies raised to- wards maritime pine proteins (20). As shown in Fig. 3, Fd -GOGAT was exclusively detected in the lanes containing total (T) and soluble (S) chloroplast polypeptides, and it was absent in the membrane (M) protein fraction. Therefore, we can conclude that Fd -GOGAT in pine cotyle- dons is confined to the stroma fraction of the chloroplast compartment.
Isolation of cDNA clones encoding Fd-GOGAT in pine
In order to obtain more detailed information on the structure and regulation of Fd -GOGAT in gymnosperms, we decided to use G O G A T anti- bodies to immunoscreen a cDNA library from Pinus sylvestris seedlings constructed in the 2gtl 1
Fig. 3. Localization of Fd-GOGAT polypeptide in isolated chloroplasts. (a) Intact chloroplasts were isolated from cotyledons of light-grown maritime pine seedlings. Proteins were extracted from total (T), soluble (S) and membrane (M) fractions, resolved by SDS-PAGE (40 #g of protein per lane) and stained with Coomassie blue. (b) Replicates of the same gel were processed for western analysis using specific antibodies raised against LSU, SSU, LHCP [20] and Fd-GOGAT polypeptides. The bands on the left correspond to the molecular mass standards. The positions of the immunodetected bands are indicated on the right.

expression vector [26]. About 300000 recombi- nant phages were plated out and immunoprobed with purified antibodies obtained against the pine enzyme in the same conditions used before for western blotting analysis. A single plaque show- ing strong signal was identified (2GOP4), purified through successive rounds of plating and finally picked out. Phage DNA was purified from liquid bacterial lysates, cDNA insert (150 bp) released by Eco RI digestion, subcloned into pGEM3Z plasmid and sequenced in both strands. Two lines of evidence confirmed that the isolated clone en- coded part of a pine Fd-GOGAT polypeptide: the deduced amino acid sequence was highly ho- mologous to the other Fd-GOGAT protein se- quences available in the databases; in addition, the fusion protein encoded by 2GOP4 (and in- duced in the presence of IPTG) was able to im- munoselect specific antibodies anti Fd-GOGAT (data not shown). This cDNA insert was then used, as a probe, for re-screening the library (500000 recombinants) and several positive ph- ages were isolated and their inserts subcloned. Sequencing analysis showed that all isolated clones were incomplete, encoding for the C- terminal region of pine Fd-GOGAT. The clone harbouring the longest insert of 2.1 kb (pGOP44) was completely sequenced and its derived amino acid sequence was compared to Fd-GOGAT se- quences isolated from angiosperms (Fig. 4). The comparative analysis indicated that the gymno- sperm sequence is highly homologous to both the monocot Zea mays [38] (8370 identical) ant the dicot Nicotiana tabacum [49] and Spinacia olera- cea [34] polypeptides (85-8470 of identity re- spectively), in spite of their evolutionary distance. Alignment of the deduced sequence also reveals several regions for which the amino acid identity is nearly 10070. Nevertheless, some amino acid residues are only present in the gymnosperm se- quence (Fig. 4). As in protein sequences deduced from maize and tobacco cDNA clones, the cor- responding pine polypeptide contains a conserved region which shares about 70 70 similarity with a putative FMN-binding domain in yeast flavocy- tochrome b2 [28] and three conserved cysteines potentially implicated in an iron-sulphur cluster.
121
Organ-specific protein expression
To further characterize Fd-GOGAT in pine seed- lings the distribution of the protein was deter- mined in different organs of light-grown plants by western blot analysis. Figure5a shows that GOGAT polypeptide was more abundant in cotyledons than in stems and it was barely de- tectable in roots. These results suggest an organ- specific expression of GOGAT in pine, the higher protein levels being detected in photosynthetic tissues. The possibility that exogenous supplied nitrogen could alterate the expression pattern of the protein, particularly in roots, was investigated by western blot analysis of GOGAT polypeptides in plants grown with no nitrogen source or ni- trate. Treatment with 15 mM nitrate did not af- fect the relative abundance of GOGAT polypep- tides in roots and cotyledons of pine seedlings (Fig. 5b). Therefore, Fd-GOGAT expression ap- pears to be organ-specific in pine and nitrogen nutrition source does not exert significant effect on this expression pattern.
Accumulation of Fd-GOGAT in developing pine cotyledons
The evolution of Fd-GOGAT activity and pro- tein during early development of maritime pine seedlings was also studied. The cotyledon length of the seedlings was chosen as a parameter to compare similar developmental stages in light- and dark-grown plant. Although chlorophyll con- tents were higher in illuminated plants, light- grown and dark-grown plants accumulated simi- lar amounts of photosynthetic pigments during initial stages of development [12]. Fd-GOGAT specific activity was very low in the embryos and increased markedly during seed germination in light and darkness (Fig. 6a). A net increase of about 4-fold was observed in the light whereas in darkness it was even slightly higher (6-fold). The availability of a Fd-GOGAT antiserum allowed us the analysis of the protein by western blotting in the same developmental stages in which en- zyme activity had also been determined (Fig. 6b).

122
p s m m m m 4 0 Nt 40 So 40 Zm 40
So 80 Zm D'.Io] I ~ K ~ 80
P S m m n m 120 Nt 120 SO 120 Zm 120
Nt 160 SO 160 Zm 160
00 Nt 200 So 200 Zm 200
m! m imm Ps F I A < R 7 H 240 Nt Y R D 240 So F K D 240 Zm Y K H 240
v=l lll i| oO so A s A ~Sh~:~ T A ~ m ~ ~ 280 Zm ~ N A ~ S Q ~ S ~ D ~ 280
Nt ~ w a v m N ~ 32o so s_~mEul ,,wm~ ~ i [ ] ~ 320 Zm ~]~iE [] A |,~.~I~ |~I~¢4[E [] ~ ~ F Q 320
o m Uim | vlm 0 o 600
So 400 Zm 400
Ps Lm m m 440 Nt V 440 So I 440 Zm V Q 440
p s m m m m Nt 480 So 480 Zm 480
sot msmA 520 z ~ N E I a ~ s2o
i mi o, . . . . Nt T T AAG V L Y L ~ E I PLK*555 So T M T S E A S L ~ * ..... 550 Zm S V L'AKQAb'ITmLS AK* . . .552

123
Fig. 5. Organ specific-expression of the Fd-GOGAT polypeptide in pine seedlings. (a) Soluble proteins were extracted from roots, cotyledons and stems of light-grown plants mad separated by SDS-PAGE (30 #g of protein per lane); then, western blotting was accomplished with the specific anti-GOGAT antibodies. The position of the Fd-GOGAT polypeptide is marked on the right. (b) The effect of nitrogen supply on Fd-GOGAT polypeptide abundance in roots and cotyledons was studied by western blot analysis. Pine seedlings were grown for 15 days in vermiculite with no nitrogen (-) or in the presence of nitrate (15 raM) or am- monium (15 raM). Roots of ammonium fed plants were severely damaged by the treatment, and therefore, were not processed to study. The position of Fd-GOGAT polypeptide is indicated by an arrow on the right.
Soluble proteins were extracted from pine em- bryo and cotyledons, separated by SDS-PAGE, electroblotted to nitrocellulose filters and immu- nochemically processed. Figure 6b (light and dark) shows that GOGAT polypeptide, which was undetectable in the embryo (lane 1), accu- mulated gradually during development (lanes 2-6), in a light-independent manner. A densito- metric analysis of the immunoreactive bands at the last stages (lanes 5 and 6) indicated that the relative abundance of GOGAT polypeptide was slightly higher in dark-grown than in light-grown plants (data not given), agreeing well with the results observed at enzyme activity level. The en- zyme activity evolution correlates well with the protein steady-state levels and therefore we can conclude that the increase of Fd-GOGAT activ- ity seems to be mainly supported by a parallel accumulation of the enzyme; however we cannot
exclude that enzyme modulation may also con- tribute to the observed values of enzyme activity during pine seed germination.
Northern blot analysis of Fd-GOGAT message
As stated above, Fd-GOGAT activity and its corresponding polypeptide accumulated in devel- oping pine seedlings in darkness. After isolating the homologous cDNA probe, northern hybrid- izations were performed and the steady-state lev- els of Fd-GOGAT transcripts compared in light and dark-grown plants (Fig. 7). Total RNA was isolated from cotyledons, size-fractionated by formaldehyde-agarose gel electrophoresis, blotted to nitrocellulose filters and hybridized with the 32p-labelled pGOP44 cDNA insert. A Fd- GOGAT transcript of about 6 kb was detected in
Fig. 4. Comparison of pine Fd-GOGAT amino acid sequence with other Fd-dependent glutamate synthase in plants. Ps, Pinus sylvestris (this work); Nt, Nicotiana tabacum [49]; So, Spinacia oleracea [34]; Zm, Zea mays [38]. Identical amino acid residues are boxed in black and conserved amino acid residues are boxed in grey. The region with homology to the FMN-binding domain is underlined. The location of three conserved cysteines putatively involved in an iron-sulphur centre is marked by arrows.
124
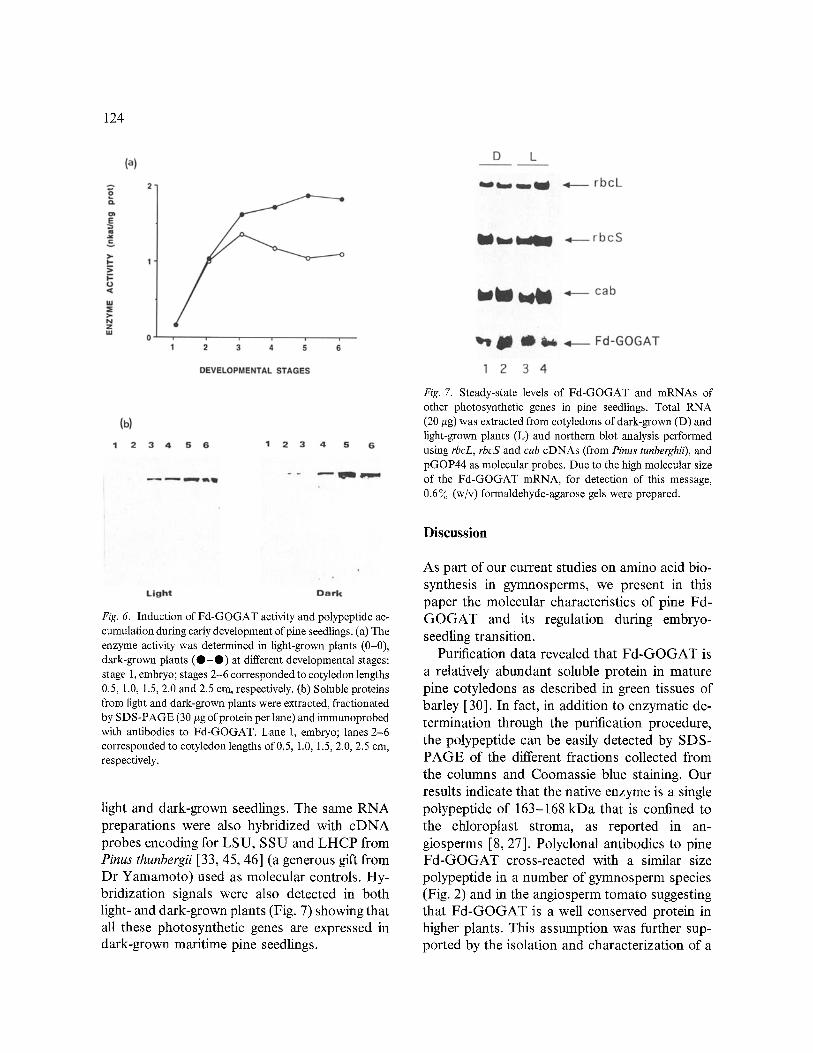
Fig. 7. Steady-state levels of Fd-GOGAT and mRNAs of other photosynthetic genes in pine seedlings. Total RNA (20 #g) was extracted from cotyledons of dark-grown (D) and light-grown plants (L) and northern blot analysis performed using rbcL, rbcS and cab cDNAs (from Pinus tunberghii), and pGOP44 as molecular probes. Due to the high molecular size of the Fd-GOGAT mRNA, for detection of this message, 0.6 7o (w/v) formaldehyde-agarose gels were prepared.
Discussion
Fig. 6. Induction of Fd-GOGAT activity and polypeptide ac- cumulation during early development of pine seedlings. (a) The enzyme activity was determined in light-grown plants (0-0), dark-grown plants ( O - Q ) at different developmental stages: stage 1, embryo; stages 2-6 corresponded to cotyledon lengths 0.5, 1.0, 1.5, 2.0 and 2.5 cm, respectively. (b) Soluble proteins from light and dark-grown plants were extracted, fractionated by SDS-PAGE (30 #g of protein per lane) and immunoprobed with antibodies to Fd-GOGAT. Lane 1, embryo; lanes 2-6 corresponded to cotyledon lengths of 0,5, 1.0, 1.5, 2.0, 2.5 cm, respectively.
light and dark-grown seedlings. The same RNA preparations were also hybridized with cDNA probes encoding for LSU, SSU and LHCP from Pinus thunbergii [33, 45, 46] (a generous gift from Dr Yamamoto) used as molecular controls. Hy- bridization signals were also detected in both light- and dark-grown plants (Fig. 7) showing that all these photosynthetic genes are expressed in dark-grown maritime pine seedlings.
As part of our current studies on amino acid bio- synthesis in gymnosperms, we present in this paper the molecular characteristics of pine Fd- GOGAT and its regulation during embryo- seedling transition.
Purification data revealed that Fd-GOGAT is a relatively abundant soluble protein in mature pine cotyledons as described in green tissues of barley [30]. In fact, in addition to enzymatic de- termination through the purification procedure, the polypeptide can be easily detected by SDS- PAGE of the different fractions collected from the columns and Coomassie blue staining. Our results indicate that the native enzyme is a single polypeptide of 163-168 kDa that is confined to the chloroplast stroma, as reported in an- giosperms [8, 27]. Polyclonal antibodies to pine Fd-GOGAT cross-reacted with a similar size polypeptide in a number of gymnosperm species (Fig. 2) and in the angiosperm tomato suggesting that Fd-GOGAT is a well conserved protein in higher plants. This assumption was further sup- ported by the isolation and characterization of a

cDNA clone encoding the C-terminal one third of the pine Fd-GOGAT polypeptide. The compari- son of the pine Fd-GOGAT amino acid sequence to the other cDNA deduced sequences revealing an unusual high degree of conservation in primary structure of this enzyme in higher plants (Fig. 4). These findings define a critical biological role for GOGAT enzyme in intermediary nitrogen meta- bolism of higher plants.
The deduced pine amino acid sequence in- cludes a putative FMN-binding domain, as re- ported for maize [38] and tobacco [49] Fd- GOGAT polypeptides. In the same region of this protein motif, three conserved cysteines poten- tially involved in an iron-sulphur cluster in alfalfa NADH-GOGAT [23] are also present. In a re- cent work, Avila et al. [5] have identified a puta- tive glutamine-amidotransferase domain in the N-terminal region of barley Fd-GOGAT, which is homologous to the corresponding domain of thepur F-type amidotransferases. Taken together, the above data suggest the existence of two func- tional parts in the same polypeptide chain, the N-terminal glutamine-amidotransferase and the C-terminal electron transfer domains that should be closely related during enzyme catalysis.
During germination, nitrogen and carbon re- serves in seeds are mobilized to the developing embryo and used in the construction of the pho- tosynthetic apparatus. Unlike angiosperms, in most gymnosperm plants this metabolic transi- tion can occur in the total absence of light [7, 29]. In maritime pine seeds, storage proteins are ex- tremely rich in arginine, about 15-25 ~o of their total amino acid content [1 ]. After nitrogen in- terconversion, most abundant amino acids in pine seedlings have been reported to be arginine and the amides glutamine and asparagine [24]. We reported earlier the accumulation of high levels of glutamine synthetase in green dark-grown mari- time pine seedlings (Pinus pinaster) [12]. The re- sults showed in this paper indicate that Fd- GOGAT activity and its corresponding polypeptide also accumulate during the initial stages of development (Fig. 6). In angiosperms, an increase in the abundance of Fd-GOGAT ac- tivity and polypeptide has also been described
125
during the greening of maize leaves [41]. More- over, the presence of the protein is apparently accounted by the expression of Fd-GOGAT mRNA in the same tissue [38]. The appearance of Fd-GOGAT activity (determined by using me- thyl viologen as reductant) has been recently re- ported to be estimulated by light in P. sylvestris seedlings [ 18]. In contrast, our findings indicate that light is not required for ferredoxin-dependent enzyme activity, polypeptide accumulation and mRNA expression during early stages of P. pin- aster development. Reducing power for Fd- GOGAT activity is supplied by light reactions in tissues with active photosynthesis. However, re- cent biochemical evidence supports the function- ality of the enzyme in other plant tissues and metabolic conditions; thus, Fd-GOGAT activity has been reported in non-photosynthetic tissues [19, 35, 43]. Roots and non-green fruits contain Fd and Fd-NADP-oxidoreductase (FNR) [22, 32, 43] and it has been demonstrated that Fd- FNR system can be activated by reductant trans- fer from NADPH [43]. Recently, Bowsher et al. [9] have described that reductant for glutamate synthase can be provided by the oxidative pen- tose phosphate pathway in tissues where no pho- tochemical activity is operating. Such redox path- way, if active in dark-grown pine seedlings, could supply the required reducing equivalents for glutamate biosynthesis.
We have shown that Fd-GOGAT protein ex- pression appears to be organ-specific in pine seedlings as occurs in tobacco [49]. Steady-state levels of Fd-GOGAT polypeptide are higher in photosynthetic than in non-photosynthetic tis- sues. We also showed that abundance of Fd- GOGAT polypeptide is not affected in a great extent by nitrogen nutrition. In close agreement with these data, Elminger et al. [ 18 ] have reported that methyl-viologen-dependent GOGAT activity is mainly found in cotyledons of Scots pine seed- lings. Other chloroplast-located proteins display a similar tissue-specific accumulation in pine seedlings [33].
In our previous studies on glutamine synthetase characterization in pine, we could not detect GS2 isoenzyme and its corresponding polypeptide in

126
Storage proteins Lipid reserves
Amino acids Carbon skeletons
NH 4 2-oxoglutarate ATP, Fd red
Chloroplast biogenesis in darkness
Fig. 8. Proposed role of the glutamine synthetase/glutamate synthase pathway in the nitrogen interconversion during early stages of pine development in the dark.
maritime [12, 13, 14] and Scot pines [15]. A full-length cDNA clone encoding GS 1 has been isolated and characterized. During the course of this work, several other additional clones were also characterized but all of them encoded for the cyto s olic polypeptide [ 13, 15 ]. Thus, the available data suggest that in a great extent glutamine bio- synthesis in pine is catalysed by GS1, at least during early stages of plant development [ 15]. As occurs in other plants, pine Fd-GOGAT is a soluble protein located in the chloroplast stroma and, consequently, glutamate synthesis is con- fined to the plastid. We still do not know if GS and GOGAT are expressed in the same cell types in pine; but, wherever glutamine is synthesized, it should be transported into the chloroplast for amino acid and chlorophyll biosynthesis in light- and dark-grown plants. A glutamine translocator has been described in spinach leaves which is also responsible for glutamate export to the cytosol [48]. If this system is working in pine it should also ~e operative in darkness.
To summarize, in this work we report for the first time the molecular characterization of Fd-
GOGAT from a woody plant. We have also dem- onstrated that such a ferredoxin-dependent and chloroplast-located enzyme accumulates at high levels in cotyledons of dark-grown pine seedlings. The presence of a functional Fd-GOGAT in these conditions can be important to provide the re- quired glutamate and glutamine for the biosyn- thesis of chlorophyll and other nitrogen com- pounds during light-independent chloroplast biogenesis (Fig. 8). Current work in our labora- tory is addressed to the isolation of genomic se- quences with the aim to determine the gene struc- ture characterization and to define the regulatory elements involved in GS and GOGAT gene ex- pression in conifers.
Acknowledgements
We would like to thank Remedios Crespillo for excellent technical assistance in many experi- ments carried out in this work. We are indebted to Dr Javier M/trquez (Universidad de M/tlaga) for helping us in the fast liquid chromatography (FPLC) separations, Dr Naoki Yamamoto (For- estry and Forest Product Research Institute, Tsukuba Science city) for providing pine rbcL, rbcS and cab cDNA clones and Dr Concepcidn Avila (Centro de Investigaciones Bioldgicas) for helpful discussion. This research was supported by a Grant (PB92-0423) from the Direccidn General de Investigacidn Cientlfica y T6cnica (Spain) to F.M.C. The financial support of Junta de Andalucla is also acknowledged.
References
1. Allona I, Casado R, Aragoncillo C: Seed storage proteins from Pinus pinaster Ait.: homology of major components with 11S proteins from angiosperms. Plant Sci 87:9-18 (1992).
2. Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K: Current Protocols in Molecular Biology, vol 1. Greene Publishing Associates/ Wiley-Interscience (1989).
3. Avila C, Botella JR, Cfinovas F, Nfifiez de Castro I, Valpuesta V: Different characteristics of the two glutamate

synthases in the green leaves of Lycopersicon esculentum. Plant Physiol 85:1036-1039 (1987).
4. Avila C, C~movas F, Nfifiez de Castro I, Valpuesta V: Separation of two forms of glutamate synthase in leaves of tomato Lycopersicon esculentum. Biochem Biohpys Res Commun 122:1125-1129 (1984).
5. Avila C, Mfirquez AJ, Pajuelo P, Cannell ME, Walls- grove RM, Forde BG: Cloning and sequence analysis of a cDNA for barley ferredoxin-dependent glutamate syn- thase and molecular analysis ofphotorespiratory mutants deficient in the enzyme. Planta 189:475-483 (1993).
6. Blackwell RD, Murray AJS, Lea PJ, Kendall AC, Hall NP, Turner JC, Wallsgrove RM: The value of mutants unable to carry out photorespiration. Photosynt Res 16, 155-176 (1988).
7. Bogdanovic M: Chlorophyll formation in the dark. Clorophyll in pine seedlings. Physiol Plant 29:17-18 (1973).
8. Botella JR, Verbelen JP, Valpuesta V: Immunocytolocal- ization of ferredoxin-GOGAT in the cells of green leaves and cotyledons of Lycopersicon esculentum. Plant Physiol 87:255-257 (1988).
9. Bowsher CG, Boulton EL, Rose J, Nayagam S, Emes MJ: Reductant for glutamate synthase is generated by the oxidative penthose phosphate pathway in non photosyn- thetic root plastids. Plant J 2:893-898 (1992).
10. Bradford MM: A rapid and sensitive method for the quantitation of microgram quantities utilising the prin- ciple of protein-dye binding. Anal Biochem 72:248-254 (1976).
11. CS.novas F, McLarney B, Silverthorne J: Light-indepen- dent synthesis of LHCIIb polypeptides and assembly of the major pigmented complexes during the initial stages of Pinus palust~s seedling development. Photosynth Res 38:89-97 (1993).
12. C~.novas FM; Cant6n FR, Gallardo G, Garcia-Guti6rrez A, de Vicente A: Accumulation of glutamine synthetase during early development of maritime pine (Pinuspinaster) seedlings. Planta 185:372-378 (1991).
13. Cantdn: Caracterizaci6n molecular de la glutamina sin- tetasa de pino: asilamiento de cDNA, secuencia y expre- sidn. Ph. D. thesis, Universidad de Mfilaga (1993).
14. Cantdn FR, Garcia-Guti6rrez A, Gallardo F, de Vicente A, Cgmovas FM: Regulacidn no dependiente de luz de proteinas cloroplastidicas y glutmnina sintetasa en plfin- tulas de pino. In: Pineda M, Castitlo F (eds) Metabolismo del Nitr6geno, pp. 180-184 Sociedad Espafiola de Bio- qulmica, Cdrdoba (1992).
15. Cantdn FR, Garcia-Guti6rrez A, Gallardo F, de Vicente A, Cfinovas FM: Molecular characterization of a cDNA clone encoding glutamine synthetase from a gymnosperm: Pinus sylvestris. Plant Mol Biol 22:819-828 (1993).
16. Castelfranco PA, Beale ST: Chlorophyll biosynthesis: re- cent advances and areas of current interest. Annu Rev Plant Physiol 34:241-278 (1983).
17. Devereux J, Haeberli P, Smithies O: A comprehensive set
127
of sequence analysis programs for the VAX. Nucl Acids Res 12:387-395 (1984).
18. Elmlinger MV, Mohr H: Coaction of blue/ultraviolet-A light and light absorbed by phytochrome in controlling the appearance of ferredoxin-dependent glutamate synthase in the Scots pine (Pinus sylvestris L.) seedling. Planta 183: 374-380 (1991).
19. Gallardo F, Cantdn FR, Garcia-Guti6rrez, Cfinovas FM: Changes in photorespiratory enzymes and glutamate syn- thases in ripening tomatoes. Plant Physiol Biochem 31: 189-196 (1993).
20. Garcia-Guti6rrez A, Cantdn FR, Gallardo F, Cfinovas FM: Immunochemical analysis of chloroplast polypep- tides from maritime pine. Phytochemistry 34:337-341 (1993).
21. Gotor C, Martinez-Rivas JM, M~rquez AJ, Vega JM: Functional properties of purified ferredoxin-glutamate synthase from Chlamydomonas reinhardtii. Phytochemis- try 29:711-717 (1990).
22. Green LS, Yee BC, Buchanan B, Kamide K, Samada Y, Wada K: Ferredoxin and Ferredoxin-NADP reductase from photosynthetic and non photosynthetic tissues of tomato. Plant physiol 96:1207-1213 (1991).
23. Gregerson RG, Miller SS, Twary SN, Gant JS, Vance CP: Molecular characterization of NADH-dependent glutamate synthase from alfalfa nodules. Plant Ceil 5: 215-226 (1993).
24. Guitton Y: M~tabolisme de l'arginine dans les premieres stades du d6veloppment de Pinus pinea L. Physiol V6g 2: 95-156 (1964).
25. Joy KW, Mills WR: Purification of chloroplasts using silica sols. Meth Enzymol 148:179-188 (1987).
26. Karpinski S, Wigsle G, Olsson O, H~lgren J-E: Charac- terization of cDNAs encoding CuZn-superoxide dismu- tases in Scots pine. Plant Mol Biol 18:545-555 (1992).
27. Lea PJ, Robinson SA, Stewart GR: The enzymology and metabolism of glutamine, glutamate, and asparagine. In: Miflin BJ, Lea PJ (eds) Biochemistry of Plants, vol 16, pp. 121-159. Academic Press, New York, (1990).
28. Lederer F, Cortial S, Becam AM, Hanmont PY, Perez L: Complete amino acid sequence of fiacocytochrome b2 from baker's yeast. Eur J Biochem 152:419-428 (1985).
29. Mariani P, De Carli ME, Rascio N, Baldan B, Casadoro G, Gennari G, Bodner M, Larcher W: Synthesis of chlo- rophyll and photosynthetic competence in eitolated and greening seedlings of Larix decidua as compared with Picea abies. J Plant Physiot 137:5-14 (1990).
30. Mfirquez AJ, Avila C, Forde BG, Wallsgrove RM: Ferredoxin-glutamate synthase from barley leaves: rapid purification and partial characterization. Plant Physiol Biocbem 26:645-651 (1988).
31. McGrath RB, Coruzzi GM: A gene network controlling glutamine and asparagine biosynthesis in plants. Plant J 1:275-280 (1991).
32. Morigasaki S, Takada K, Suzuki T, Wada K: Purifica- tion and characterization of ferredoxin-NADP + oxi-

128
doreductase-like enzyme from radish root tissues. Plant Physiol 93:896-901 (1990).
33. Mukai Y, Yamamoto N, Koshiba T: Light independent and tissue-specific accumulation of light-harvesting chlo- rophyll a/b binding protein and ribulose bisphosphate carboxylase in dark-grown pine seedlings. Plant Cell Physiol: 32:1303-1306 (1991).
34. Nalbantoglu B, Hirasawa M, Moomaw C, Nguyen H, Knaff DB, Allen R: Cloning and sequencing of the gene encoding spinach ferredoxin-dependent glutamate syn- thase. Biochim Biophys Acta 1183:557-561 (1994).
35. Oaks A, Hirel B: Nitrogen metabolism in roots. Annu Rev Plant Physiol 36:345-365 (1985).
36. R6dinbaugh MG, Campbell WH: Glutamine synthetase and ferredoxin-dependent glutamate synthase expression in the maize (Zea mays) root primary response to nitrate. Evidence for an organ-specific response. Plant Physiol 101:1249-1255 (1993).
37. Romero LC, Galv/m F, Vega JM: Purification and prop- erties of the siroheme-containing ferredoxin-nitrite reduc- tase from Chlamydomonas reinhardtii. Biochim Biophys Acta 914:55 (1987).
38. Sakakibara H, Watanabe M, Hase T, Sugiyama T: Mo- lecular cloning and characterization of complementary DNA encoding for ferredoxin-dependent glutamate syn- thase in maize leaf. J Biol Chem 266:2028-2035 (1991).
39. Sambrook J, Fritsh EF, Maniatis T: Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989).
40. Sanger F, Nicklen S, Coulson AR: DNA sequencing with chain terminating inhibitors. Proc Natl Acad Sci USA 74: 5463-5467 (1977).
41. Suzuki A, Audet C, Oaks A: Influence of light in the ferredoxin-dependent glutamate synthase in maize leaves. Plant Physiol 84:578-581 (1987).
42. Suzuki A, Gadal P: Glutamate synthase: physicochemi-
cal and functional properties of different forms in higher plants and other organisms. Physiol V6g 22:471-486 (1984).
43. Suzuki A, Oaks A, Jacquot JP, Vidal J, Gadal P: An electron transport system in maize roots for reactions of glutamate synthase and nitrite reductase. Physiological and immunochemical properties of the electron carrier and pyridine nucleotide reductase. Plant Physio178: 374- 378 (1985).
44. Tobin E, Silverthorne J: Light regulation of gene expres- sion in higher plants. Annu Rev Plant Physiol 35: 569- 593 (1985).
45. Yamamoto N, Matsuoka M, Kano-Murakami Y, Tanaka Y, Ohashi Y: Nucleotide sequence of a full-length eDNA clone of light-harvesting chlorophyll a/b binding protein gene from green dark-grown pine (Pinus thunbergii) seed- ling. Nucl Acids Res 16:11829 (1988).
46. Yamamoto N, Kano-Murakami Y, Matsuoka M, Ohashi Y, Tanaka Y: Nucleotide sequence of a full-length cDNA clone ofribulose bisphosphate carboxylase small subunit gene from green dark-grown pine (Pinus thunbergii) seed- ling. Nucl Acids Res 16:11830 (1988).
47. Yamamoto N, Mukai Y, Matsuoka M, Kano-Murakami Y, Ohashi Y, Tanaka Y, Ozeki Y, Odani K: Light- independent expression of cab and rbcS genes in dark- grown pine seedlings Plant Physiol 95:376-383 (1991).
48. Yu J, Woo KC: Glutamine transport and the role of the glutamine translocator in chloroplasts. Plant Physiol 88: 1048-1054 (1988).
49. Zehnacker C, Becker TW, Suzuki A, Carrayol E, Cabo- che M, Hirel B: Purification and properties of tobacco ferredoxin-dependent glutamate synthase, and isolation of corresponding cDNA clones. Light-inducibility and organ-specificity ofgene transcription and protein expres- sion. Planta 187:266-274 (1992).
Copyright © 2022 FDOKUMEN




















