
Expression of glutamate transporters in rat optic nerve oligodendrocytes
Upload
independentCategory
view
1download
0
Plasmodium chabaudi: Immunogenicity of a Highly AntigenicGlutamate-Rich Protein
MARK F. WISER,*,1 LUIS E. GIRALDO,* HANS-PETERSCHMITT-WREDE,† AND FRANK WUNDERLICH†
*Department of Tropical Medicine, Tulane University School of Public Health and Tropical Medicine,New Orleans, Louisiana 70112, U.S.A.; and †Division of Parasitology, University of Du¨sseldorf, 4000
Dusseldorf, Germany
WISER, M. F., GIRALDO, L. E., SCHMITT-WRETE, H-P., AND WUNDERLICH, F. 1997.Plasmodiumchabaudi:Immunogenicity of a highly antigenic glutamate-rich protein.Experimental Parasitology85,43–54. The immunogenicity of a 93-kDaPlasmodium chabaudiprotein that contains glutamate-rich tandem repeats was investigated in this study. Immunoblotting with various monoclonal anti-bodies indicates that this 93-kDa protein is equivalent to a potentialP. chabaudiRESA analogue.However, the sequence of theP. chabaudiprotein does not exhibit any significant homology toPf155/RESA. Antibodies against the 93-kDa protein appear early duringP. chabaudiinfection andreach high titers. The highest antibody titers are found when the parasitemia is descending, suggestingthat this protein may play some role in immunity. Immunization of mice with the recombinant proteinalso results in high antibody titers, indicating that the protein is quite immunogenic. However, miceimmunized with recombinant protein and challenged withP. chabaudido not exhibit a delayedappearance of parasitemia, a reduced parasitemia, or a shortened duration of parasitemia. Glutamate-rich P. falciparumproteins such as Pf155/RESA, are being considered as vaccine candidates. Thestudies withP. chabaudisuggest that interpretation of serological data using glutamate-rich proteinsshould proceed with caution. The glutamate-rich repeats, although highly immunogenic, may not beimportant in host immunity against malaria. However, antibodies that appear late in theP. chabaudiinfection do appear to play a role in anti-malarial immunity.© 1997 Academic Press
INDEX DESCRIPTORS ANDABBREVIATIONS: malaria; tandem repeats; glutamate-rich protein; vaccine.EENV, Glu-Glu-Asn-Val tetrapeptide repeat; EENY, Glu-Glu-Asn-Tyr tetrapeptide; GST, glutathi-one-S-transferase; mAb, monoclonal antibody; MBP, maltose binding protein; SDS–PAGE, sodiumdodecyl sulfate–polyacrylamide gel electrophoresis.
INTRODUCTION
Malaria is a major cause of morbidity andmortality in much of the developing world.There is interest in developing vaccines againstmalaria and several candidate antigens havebeen identified. Many of thePlasmodium falci-parumproteins proposed as vaccine candidatescontain tandem repeats rich in glutamate resi-dues. Among these are Pf155/RESA (Coppeletal. 1984), Pf332 (Mattei and Scherf 1992),Pf11.1 (Scherfet al.1992), Pfs230 (Williamsonet al. 1993), MESA (Coppel 1992), GLURP(Borre et al. 1991), and D260 (Barneset al.
1995). Glu-Glu residues are quite prevalentwithin the tandem repeats of these antigens.Little work has been carried out on analogousproteins in rodent malaria models, and studiesof the efficacy of these glutamate-rich proteinsas vaccines has been limited. A putative RESAanalogue fromP. chabaudi,referred to as eitherPch105/RESA (Gabrielet al. 1986) or Pc96(Wanidworanumet al. 1987), has been identi-fied based upon its localization to the erythro-cyte membrane.Phosphoproteins fromP. berghei and P.
chabaudicontaining high levels of glutamateand aspartate residues have also been character-ized (Wiser et al. 1988). These proteins arequite acidic (isoelectric point <4) and are local-ized to the host erythrocyte membrane. Theseacidic phosphoproteins are not exposed on thesurface of infected erythrocytes, but are found
1 To whom correspondence should be addressed at theDepartment of Tropical Medicine, Tulane UniversityMedical Center, 1501 Canal Street, New Orleans, LA70112. Fax: (504) 599-6686; E-mail: [email protected].
EXPERIMENTAL PARASITOLOGY85, 43–54 (1997)ARTICLE NO. PR964126
430014-4894/97 $25.00Copyright © 1997 by Academic PressAll rights of reproduction in any form reserved.

on the cytoplasmic face of the host membraneand appear to interact with the submembranecytoskeleton (Wiseret al. 1988, 1990). TheP.chabaudiacidic phosphoprotein, referred to asPc(em)93, is the same as a protein known asPc90 (Wunderlichet al. 1988b). Cloning andsequencing of the gene for Pc90 reveals a pre-dicted amino acid sequence consisting largelyof a central block of tandem repeats rich in glu-tamate residues (Schmitt-Wredeet al. 1993).Pc(em)93 also exhibits a size and subcellularlocation similar to those of Pch105/RESA andPc96. Data presented herein indicate thatPc(em)93 and Pch105/RESA are the same pro-teins.To better characterize the potential role of
glutamate-rich proteins in malarial immunitythe immunogenicity of Pc(em)93 was charac-terized duringP. chabaudiinfections, and therecombinant protein was tested for its ability toprotect against infection. Pc(em)93 is highlyimmunogenic and anti-Pc(em)93 antibodies ap-pear early during the infection. Vaccinationwith recombinant Pc(em)93, however, does notprotect mice againstP. chabaudiinfection. Theresults of these studies imply that research onhighly immunogenic proteins as vaccine candi-dates should proceed with caution.
MATERIALS AND METHODS
Parasites and antibodies. P. chabaudiwas obtained fromDr. R. Walter (Bernhard-Nocht-Institut, Hamburg) andmaintained by serial passage in CD-1 outbred mice. Thisstrain was obtained originally from Dr. P. I. Trigg and wasderived from strain 54X of Landau (Landau and Killick-Kendrick, 1966). Mice were acquired from Charles RiversLaboratories and maintained at the Tulane University Medi-cal Center vivarium. Parasitemia was monitored by Giemsa-stained thin blood smears obtained from tail snips.Blood was collected from the orbital plexus of anesthe-
tized mice and incubated for 2–4 hr at 37°C. Alternatively,heavily anesthetized mice were exsanguinated by axillaryincision. The clot was removed and remaining erythrocyteswere pelleted by centrifugation. Sera were stored at −20°C.mAb-13.5 was raised against two-dimensional gel-purifiedPc(em)93 (Wiseret al. 1988). Drs. Chingchai Wanidwora-num and Hanna Shear provided mAb-6C12E7 and mAb-8F2G9. These mAb were prepared from mice immunizedwith erythrocyte ghosts isolated fromP. chabaudi-infectederythrocytes (Wanidworanumet al. 1987). Dr. WillyDeleersnijder provided mAb-43 which was prepared by im-
munizing mice with a fusion protein obtained from algt11lysogen (Deleersnijderet al. 1992).Immunoblot assay of anti-Pc(em)93 antibodies.Antibod-
ies recognizing Pc(em)93 were analyzed using affinity-purified Pc(em)93 in an immunoblot assay carried out in afashion similar to previously described enriched blotting(Wiser and Schweiger 1986). Infected erythrocytes werecollected and washed in Hanks’ balanced salt solution, re-suspended in 20 volumes of K-1 buffer, and lysed by incu-bation with 0.01% saponin at 37°C for 5 min as described(Wiser et al. 1988). Particulate matter (parasites and hostmembranes) was removed by centrifugation at 27,000g for20 min. Since approximately half of Pc(em)93 is releasedinto the erythrocyte cytosol following saponin lysis (Wun-derlichet al.1988b), the supernatant was retained and pre-cipitated with 50% saturated (NH4)2SO4. Precipitated pro-tein was collected by centrifugation at 12,000g for 10 min,resuspended in approximately four times the originalpacked cell volume of phosphate-buffered saline (20 mMphosphate, pH 7.6 containing 0.15M NaCl), and recentri-fuged. The supernatant was mixed with an equal volume ofhybridoma supernatant media containing mAb-13.5 and in-cubated for 2 hr on ice. Antigen–antibody complexes werecollected with protein-G agarose (Pharmacia) and solubi-lized in SDS gel electrophoresis sample buffer. The samplewas electrophoresed on a 7.5% polyacrylamide gel with asingle wide lanes, transferred to Immobilon (Millipore)membranes and stored at −20°C. The membrane was cutinto strips and incubated with serum samples and negative(normal mouse serum) and positive (mAb-13.5) controls.Anti-Pc(em)93 antibodies were detected with an alkalinephosphatase-conjugated anti-mouse IgG (Sigma; A3562)and developed with 5-bromo-4-chloro-3-indolyl phosphateand nitro blue tetrazolium.Preparation of recombinant constructs.The 2.5-kb insert
corresponding to Pc(em)93 was excised from clone Pc90-1(Schmitt-Wredeet al.1993) withSacI andXhoI, subclonedinto the SacI and SalI sites of pMAL-p2 (New EnglandBiolabs), and designated pMPS90. This resulted in maltosebinding protein (MBP) and Pc(em)93 fusion protein. Theinsert DNA was also subcloned from clone Pc90-1 intopGEX-5X-3 (Pharmacia) using theEcoRI andXhoI restric-tion sites and designated as pGPS90. This resulted in aglutathione-S-transferase (GST) and Pc(em)93 fusion pro-tein.Escherichia coli (JM83) transformants of pMAL,
pMPS90, pGEX, and pGPS90 were grown in 200 ml ofLuria broth containing ampicillin for 3 hr at 30°C. Isopro-pylthiogalactoside was added to a final concentration of 0.6mM and the bacteria were incubated for an additional 1.5 hr.TheA600 values were measured and the bacteria were col-lected by centrifugation. The pellet was resuspended in 10times (ml) theA600 value of saline, resulting in a concen-tration of 2 × 1010 bacteria ml−1. The samples were sub-jected to two rounds of freeze–thaw and Triton X-100 wasadded to a final concentration of 0.1%. The samples weredispensed in 1-ml aliquots and stored at −70°C until use.
WISER ET AL.44

Expression of the recombinant protein was confirmed bySDS–PAGE and immunoblotting with mAb-13.5.Vaccination of mice with recombinantE. coli. Aliquots of
the recombinantE. coliwere thawed and emulsified with anequal volume (1 ml) of Freunds’ complete adjuvant. Mice ingroups of five were injected intraparentally with 0.1 ml (109
bacteria) of the emulsion. Three weeks later mice were in-jected with 109 recombinant bacteria emulsified with Fre-unds’ incomplete adjuvant. Serum samples were collectedvia the orbital plexus 3–4 days after the boost and tested foranti-Pc(em)93 activity. Two weeks after the second inocu-lation mice were injected with 109 recombinant bacteriawithout adjuvant. Sera samples were collected again and themice were challenged with 104 P. chabaudi-infected eryth-rocytes 10–14 days after the second boost. Parasitemiaswere determined from Giemsa-stained thin blood smears.Immunofluorescence.The immunofluorescence assay
was carried out on glutaraldehyde-fixed erythrocytes per-meabilized with Triton X-100 as previously described(Wiser et al. 1993). Briefly,P. chabaudi-infected erythro-cytes were washed four times in Hanks’ balanced salt so-lution, resuspended at a 5% hematocrit, and fixed for 10 minin 0.025% glutaraldehyde. The fixed erythrocytes weretreated for 20 min with 0.1% Triton X-100 before use in theassay. The fixed and permeabilized erythrocytes were incu-bated with primary sera for 1–2 hr, washed, and then incu-bated with fluorescein-conjugated anti-mouse IgG (Sigma;F6257) for 1 hr. Samples were examined for epifluores-cence under UV illumination. Pictures were obtained byexposing 400 ASA black and white print film for 20 sec andall prints were developed under identical conditions.
RESULTS
Several 90- to 105-kDaP. chabaudiproteinsthat are associated with the host erythrocytemembrane have been described. These includePc(em)93 (Wiseret al. 1988), Pc90 (Wun-derlich et al. 1988b), Pc96 (Wanidworanumetal. 1987), and Pch105/RESA (Gabrielet al.1986). It was previously shown that Pc(em)93and Pc90 are the same protein (Wunderlichetal. 1988b) and that Pc96 and Pch105/RESA arethe same protein (Holmquistet al. 1990). Im-munoblotting analysis indicates that mAb-6C12E7 and mAb-43, which both recognizePch105/RESA (Holmquistet al. 1990), alsorecognize Pc(em)93 (Fig. 1). mAb-8F2G9,which was also raised against Pc96 (Wanid-woranum et al. 1987), does not recognizePc(em)93 (Fig. 1). The lack of reactivity ofmAb-8F2G9 with Pc(em)93 may be due tostrain specificity, since mAb-8F2G9 only rec-ognizes some strains ofP. chabaudi(Holmquist
et al. 1990). The strong signals obtained withmAb-6C12E7 and mAb-43 (Fig. 1) suggest thatthe reactivity is probably not due to fortuitouscross-reactivity. In summary, these immunob-lots (Fig. 1) and those previously presented(Holmquist et al. 1990; Wunderlichet al.1988b) demonstrate that theP. chabaudipro-teins known as Pc(em)93, Pc90, Pc96, andPch105/RESA are all the same protein.Comparison of the clone Pc90-1 sequence
(Schmitt-Wredeet al. 1993) with the Pf155/RESA sequence (Favaloroet al. 1986) demon-strates that these two sequences are not homolo-gous (data not shown). Therefore, thisP.chabaudi protein, referred to hereafter asPc(em)93 to minimize confusion, is not a ho-mologue to Pf155/RESA, as the name Pch105/RESA implies. However, homology plots doshow some similarity between the tandem re-peats of Pf155/RESA and the tandem repeats ofPc(em)93 (Fig. 2). Pf155/RESA contains a cen-tral block (referred to as 59 repeats) and a C-terminal block (referred to as 39 repeats) of tan-dem repeats (Favaloroet al. 1986). Pc(em)93
FIG. 1. Reactivities of monoclonal antibodies raisedagainstP. chabaudierythrocyte membrane-associated pro-teins. mAb-13.5 was used to immunoprecipitate Pc(em)93from infected erythrocyte extracts, electrophoresed onsingle-lane gels, and transferred to Immobilon membranes.The membrane was cut into strips which were incubatedwith no primary antibody (lane 1), mAb-13.5 (lane 2), mAb-6C12E7 (lane 3), mAb-8F2G9 (lane 4), and mAb-43 (lane5). Bound antibody was detected with alkaline phosphatase-conjugated anti-mouse IgG and colorimetric substrates.pc(em)93 and the IgG heavy chain are indicated on theright. Molecular mass standards (in kDa) are indicated onthe left.
ANTIGENICITY OF GLU-RICH PlasmodiumPROTEINS 45

contains a large central domain consisting oftandem repeats (Wunderlichet al. 1988b). Allthree blocks of repeats are relatively rich in glu-tamic acid residues which account for theirsimilarity. In particular, the consensus sequenceof the 39 repeats of Pf155/RESA is EENV whileEENY is part of the 14-amino-acid consensussequence derived from the Pc(em)93 repeats.Some studies have implicated a role for
Pc(em)93 in anti-malarial immunity (Wun-derlich et al. 1988b; Wanidworanumet al.1987). Therefore, studies to examine the anti-genicity of Pc(em)93 during aP. chabaudiin-fection were carried out.P. chabauditypicallyexhibits a rapidly rising parasitemia that peaksbetween 20 and 50%, then declines and clears inapproximately 3 weeks. Sera collected frommice on various days after infection were ex-amined for anti-Pc(em)93 antibodies by immu-noblotting (Table I). Antibodies recognizingPc(em)93 are observed when parasites are de-tected by Geimsa staining. The antibody titersare initially low (1:200) but increase dramati-cally with increasing parasitemia. The anti-Pc(em)93 titers peak at a dilution of 1:10,000 afew days after the parasitemia had peaked. After
parasite clearance the antibody titers decrease tomoderate levels (1:1000–1:2000) and remainconstant for at least 2 months. The mice, onceclearing the initial infection, are effectively im-mune to subsequent challenge in that reinfec-tion results in either no detectable parasitemiaor a transient parasitemia of less than 1% for1–2 days. Reinfection of mice increases theanti-Pc(em)93 titers to approximately 1:100,000(Table I). The antibody levels then decrease totiters of approximately 1:10,000.To further investigate the possible role of
Pc(em)93 in anti-malarial immunity, recombi-nant Pc(em)93 was prepared and tested as avaccine. The insert from clone Pc90-1 (Schmitt-Wredeet al. 1993) was subcloned into pMALso that it was in frame with MBP. Immunoblotsof lysates from recombinantE. coli reveal sev-eral polypeptides recognized by mAb-13.5 (Fig.3), suggesting that the recombinant fusion pro-tein is not stable inE. coli and is subjected toproteolysis. The coding sequence of the insertDNA is predicted to have a molecular mass of75 kDa.Plasmodiumproteins with tandem re-peats migrate anomalously on SDS–PAGE(Kempet al. 1987). For example, Pf155/RESA
FIG. 2. Homology of Pc(em)93 vs Pf155/RESA. The amino acid sequences of Pc(em)93 (vertical axis) andPf155/RESA (horizontal axis) were subjected to homology plots using the DNAPLOT program of DNASTAR.A window of 10 amino acids with a 50% homology was used. The portions of the central repeats of Pc(em)93and the 59 and 39 repeats of Pf155/RESA are indicated. No homology outside of the repeats was detected.
WISER ET AL.46
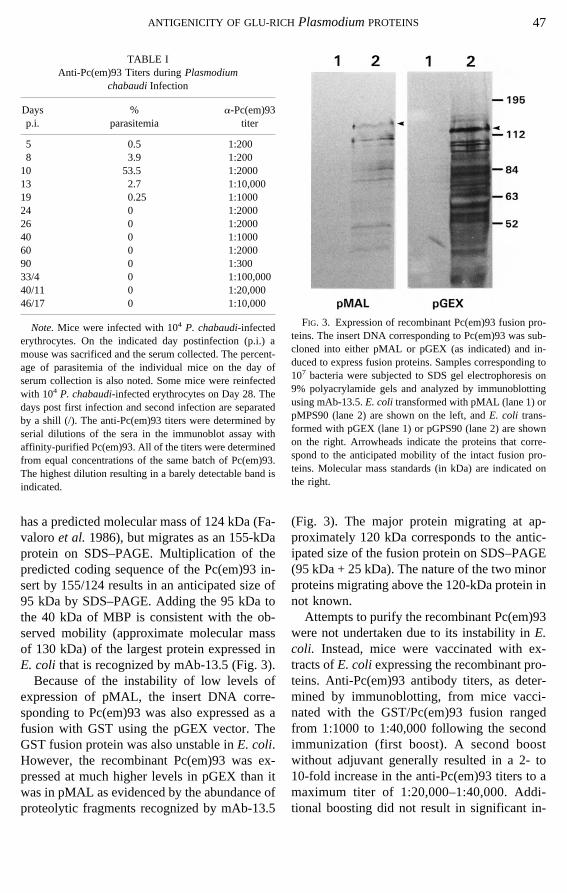
has a predicted molecular mass of 124 kDa (Fa-valoroet al.1986), but migrates as an 155-kDaprotein on SDS–PAGE. Multiplication of thepredicted coding sequence of the Pc(em)93 in-sert by 155/124 results in an anticipated size of95 kDa by SDS–PAGE. Adding the 95 kDa tothe 40 kDa of MBP is consistent with the ob-served mobility (approximate molecular massof 130 kDa) of the largest protein expressed inE. coli that is recognized by mAb-13.5 (Fig. 3).Because of the instability of low levels of
expression of pMAL, the insert DNA corre-sponding to Pc(em)93 was also expressed as afusion with GST using the pGEX vector. TheGST fusion protein was also unstable inE. coli.However, the recombinant Pc(em)93 was ex-pressed at much higher levels in pGEX than itwas in pMAL as evidenced by the abundance ofproteolytic fragments recognized by mAb-13.5
(Fig. 3). The major protein migrating at ap-proximately 120 kDa corresponds to the antic-ipated size of the fusion protein on SDS–PAGE(95 kDa + 25 kDa). The nature of the two minorproteins migrating above the 120-kDa protein innot known.Attempts to purify the recombinant Pc(em)93
were not undertaken due to its instability inE.coli. Instead, mice were vaccinated with ex-tracts ofE. coliexpressing the recombinant pro-teins. Anti-Pc(em)93 antibody titers, as deter-mined by immunoblotting, from mice vacci-nated with the GST/Pc(em)93 fusion rangedfrom 1:1000 to 1:40,000 following the secondimmunization (first boost). A second boostwithout adjuvant generally resulted in a 2- to10-fold increase in the anti-Pc(em)93 titers to amaximum titer of 1:20,000–1:40,000. Addi-tional boosting did not result in significant in-
TABLE IAnti-Pc(em)93 Titers duringPlasmodium
chabaudiInfection
Daysp.i.
%parasitemia
a-Pc(em)93titer
5 0.5 1:2008 3.9 1:20010 53.5 1:200013 2.7 1:10,00019 0.25 1:100024 0 1:200026 0 1:200040 0 1:100060 0 1:200090 0 1:30033/4 0 1:100,00040/11 0 1:20,00046/17 0 1:10,000
Note.Mice were infected with 104 P. chabaudi-infectederythrocytes. On the indicated day postinfection (p.i.) amouse was sacrificed and the serum collected. The percent-age of parasitemia of the individual mice on the day ofserum collection is also noted. Some mice were reinfectedwith 104 P. chabaudi-infected erythrocytes on Day 28. Thedays post first infection and second infection are separatedby a shill (/). The anti-Pc(em)93 titers were determined byserial dilutions of the sera in the immunoblot assay withaffinity-purified Pc(em)93. All of the titers were determinedfrom equal concentrations of the same batch of Pc(em)93.The highest dilution resulting in a barely detectable band isindicated.
FIG. 3. Expression of recombinant Pc(em)93 fusion pro-teins. The insert DNA corresponding to Pc(em)93 was sub-cloned into either pMAL or pGEX (as indicated) and in-duced to express fusion proteins. Samples corresponding to107 bacteria were subjected to SDS gel electrophoresis on9% polyacrylamide gels and analyzed by immunoblottingusing mAb-13.5.E. coli transformed with pMAL (lane 1) orpMPS90 (lane 2) are shown on the left, andE. coli trans-formed with pGEX (lane 1) or pGPS90 (lane 2) are shownon the right. Arrowheads indicate the proteins that corre-spond to the anticipated mobility of the intact fusion pro-teins. Molecular mass standards (in kDa) are indicated onthe right.
ANTIGENICITY OF GLU-RICH PlasmodiumPROTEINS 47

creases in antibody titers. Titers of mice immu-nized three times with the MBP/Pc(em)93 fu-sions ranged from 1:4000 to 1:20,000. Noantibodies against Pc(em)93 were observed inmice vaccinated with pMAL or pGEX (data notshown). In summary, vaccination of mice withE. coli expressing GST/Pc(em)93 fusion pro-teins results in anti-Pc(em)93 titers comparableto those seen during infection withP. chabaudi.The antibodies raised against the recombinantbacteria recognize the host membrane of in-fected erythrocytes and exhibit a pattern of im-munofluorescence similar to that of mAb-13.5(Fig. 4). The differences in the immunofluores-cence patterns are primarily quantitative and
correlate with the antibody titers. No excep-tional background is seen with control mouseserum other than a weak parasite fluorescence.Mice vaccinated with the recombinantE. coli
expressing Pc(em)93 are not protected fromP.chabaudichallenge (Fig. 5), even though theyexhibit antibody titers comparable to those ob-tained during infection and the immunofluores-cence pattern is as expected. Vaccination withthe recombinant fusion proteins does not resultin a delay in the appearance of parasitemia, areduction of peak levels of parasitemia, orshortening of the duration of parasitemia. Thevaccination and challenge experiments werecarried out several different times in several dif-
FIG. 4. Analysis of sera from vaccinated mice by immunofluorescence.P. chabaudi-infected erythrocyteswere fixed in glutaraldehyde and permeabilized with Triton X-100. The fixed and permeabilized erythrocyteswere incubated with the primary antibody followed by an incubation with secondary fluorescein-conjugatedanti-mouse IgG antibody and examined for epifluorescence under UV illumination. The sera were pooled fromfive vaccinated mice and diluted 1:100. (A) Sera from mice vaccinated withE. coli expressing GST. (B) Serafrom mice vaccinated withE. coli expressing the GST-Pc(em)93 fusion protein [anti-Pc(em)93 titer 1:20,000].(C) Serum from mice vaccinated withE. coli expressing the MBP/Pc(em)93 fusion protein [anti-Pc(em)93 titer1:8000]. (D) Undiluted hybridoma supernatant media containing mAb-13.5 [anti-Pc(em)93 titer 1:100,000].
WISER ET AL.48
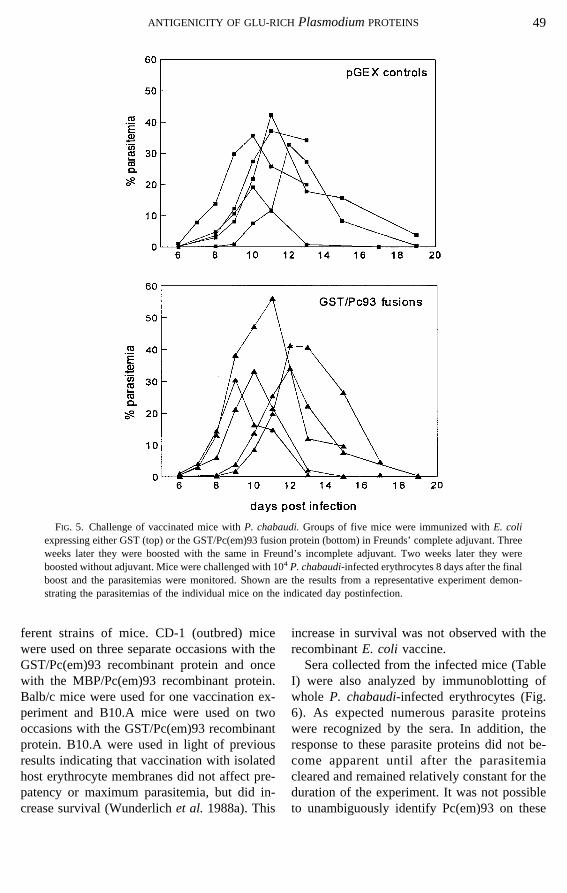
ferent strains of mice. CD-1 (outbred) micewere used on three separate occasions with theGST/Pc(em)93 recombinant protein and oncewith the MBP/Pc(em)93 recombinant protein.Balb/c mice were used for one vaccination ex-periment and B10.A mice were used on twooccasions with the GST/Pc(em)93 recombinantprotein. B10.A were used in light of previousresults indicating that vaccination with isolatedhost erythrocyte membranes did not affect pre-patency or maximum parasitemia, but did in-crease survival (Wunderlichet al. 1988a). This
increase in survival was not observed with therecombinantE. coli vaccine.Sera collected from the infected mice (Table
I) were also analyzed by immunoblotting ofwhole P. chabaudi-infected erythrocytes (Fig.6). As expected numerous parasite proteinswere recognized by the sera. In addition, theresponse to these parasite proteins did not be-come apparent until after the parasitemiacleared and remained relatively constant for theduration of the experiment. It was not possibleto unambiguously identify Pc(em)93 on these
FIG. 5. Challenge of vaccinated mice withP. chabaudi.Groups of five mice were immunized withE. coliexpressing either GST (top) or the GST/Pc(em)93 fusion protein (bottom) in Freunds’ complete adjuvant. Threeweeks later they were boosted with the same in Freund’s incomplete adjuvant. Two weeks later they wereboosted without adjuvant. Mice were challenged with 104P. chabaudi-infected erythrocytes 8 days after the finalboost and the parasitemias were monitored. Shown are the results from a representative experiment demon-strating the parasitemias of the individual mice on the indicated day postinfection.
ANTIGENICITY OF GLU-RICH PlasmodiumPROTEINS 49

blots using crude extracts as antigen. For ex-ample, the Day 13 serum exhibits a titer of 1:10,000 in the enriched immunoblot assay (TableI), but is unable to detect Pc(em)93 from crudeextracts at a 1:1000 dilution (Fig. 6, lane 4). Partof the problem is that quantitatively lessPc(em)93 is present in the crude extracts than inthe blots using affinity-purified Pc(em)93. Inaddition, apparently anotherP. chabaudipro-tein comigrates with Pc(em)93 as best illus-trated by the Day 90 serum which recognizes aprotein with the same mass as Pc(em)93 (Fig. 6,lane 8) but has very little anti-Pc(em)93 activity(Table I). This comigrating protein is not due todifferential expression of multiple alleles ofPc(em)93 since southern blot analysis revealsthat Pc(em)93 is a single-copy gene (Giraldoetal. manuscript in preparation). A possible can-didate for this comigrating protein is thePc(em)93 Lys/Asp-rich protein ofP. chabaudi(Hartzet al.1993). The approximately 250-kDaprotein on this blot is possibly merozoite sur-face protein-1.P. chabaudimerozoite surface
protein-1 has been previously demonstrated tobe an immunodominant protein as well as a pro-tective antigen (Boyleet al. 1982).
DISCUSSION
The results of the studies described hereindemonstrate that Pc(em)93 is the same as sev-eral other erythrocyte membrane-associatedproteins fromP. chabaudiand that Pc(em)93 ishighly immunogenic. Vaccination with cruderecombinantE. coli expressing Pc(em)93 in-duced antibodies against Pc(em)93 with titerscomparable to the anti-Pc(em)93 titers observedduringP. chabaudiinfections. Furthermore, thepredominant antibody response of mice immu-nized with host membranes isolated fromP.chabaudi-infected erythrocytes is also againstPc(em)93 (Wanidworanumet al. 1987; Wun-derlich et al. 1988a). The observations thatcrude membrane preparations and crude bacte-rial lysates induce high titer and specific anti-body responses indicate that Pc(em)93 is an im-munodominant protein. Purified Pc(em)93
FIG. 6. Analysis of sera by immunoblotting during the course of infection.P. chabaudi-infected erythrocyteswere isolated during late trophozoite and schizont stage, solubilized in SDS gel electrophoresis sample buffer,electrophoresed on 9% polyacrylamide gels, and transferred to Immobilon membranes which were cut intostrips. The strips were incubated with sera diluted 1:1000 which were collected on Day 8 (lane 2), Day 10 (lane3), Day 13 (lane 4), Day 24 (lane 5), Day 40 (lane 6), Day 60 (lane 7), or Day 90 (lane 8) postinfection. Somemice were reinfected on Day 28 and sera collected 4 days (lane 9) or 11 days (lane 10) after the second infection.Normal mouse serum (lane 1) and no primary antibody (lane 11) were used as negative controls and mAb-13.5was used as a positive control (lane 12). Molecular mass standards (in kDa) are indicated on the left. The smallarrow denotes presumptive merozoite surface protein-1 and the bold arrow denotes Pc(em)93.
WISER ET AL.50

also induces high titers of anti-Pc(em)93 anti-bodies (Wiseret al. 1988; Wunderlichet al.1988b), indicating that the high antibody titer isnot exclusively due to the expected adjuvanteffect of the recombinantE. coli. Vaccinationwith the recombinant Pc(em)93, in spite of thehigh antibody titers, does not protect againstP.chabaudichallenge. Similarly, vaccination withPc(em)93 purified from infected erythrocytesdoes not protect against infection (Wunderlichet al. 1988b). Interestingly, vaccination withisolated erythrocyte membranes from infectedcells does protect against challenge (Wanid-woranumet al.1987; Wunderlichet al.1988a),suggesting that a less antigenic protein in thehost membrane preparations may be responsiblefor protection. Alternatively, the membrane-associated form of Pc(em)93 may be antigeni-cally different from the purified or recombinantPc(em)93, and thus results in protection.The lack of a role for the early appearing
anti-Pc(em)93 antibodies in protection againstinfection is consistent with the observation thatmice infected withP. chabaudiinitially mount astrong Th1 response which greatly reduces par-asitemia (Stevenson and Tam 1993; Taylor-Robinson and Phillips 1994; von der Weidet al.1994). Final parasite clearance and preventionof recrudescence, however, is dependent upon aswitch to a Th2 response. This suggests thatcellular mechanisms are more important thanantibody-dependent mechanisms during the ini-tial development of anti-malarial immunity. Theappearance of high titer antibodies against para-site proteins, such as merozoite surface protein-1, late in the infection suggests that the switchto a protective Th2 response involves the pro-duction of antibodies against a specific subset ofP. chabaudiproteins. The observation thatPc(em)93 is not a predominant antigen late inthe infection is further evidence that immuneresponses against Pc(em)93 are not importantfor anti-malarial immunity. However, it cannotbe excluded that parts of Pc(em)93 could elicita protective response and that this potentiallyprotective response is being diverted by the im-munodominant glutamate-rich tandem repeats.The exactP. falciparumhomologue of the
Pc(em)93-kDa protein is not known, althoughsimilar acidic proteins have been identified onhost membranes isolated fromP. falciparum-infected erythrocytes (Heidrich and Wiser, un-published). Nonetheless, manyP. falciparumantigens, such as Pf155/RESA, exhibit similari-ties to Pc(em)93 in terms of tandem repeats thatare rich in glutamic acid. Some of theseP. fal-ciparum proteins are proposed vaccine candi-dates and/or used in epidemiological studies tomake inferences about immunity. For example,high antibody titers against Pf155/RESA, espe-cially the 39 EENV repeats, are reported to cor-relate with immunity against malaria (Petersenet al.1990; Rileyet al.1991; Hoghet al.1992).In addition, high antibody titers against the glu-tamate-rich repeats of GLURP correlate withlower parasite densities (Hoghet al. 1992).Similar types of correlations could also be madefrom the results of this study withP. chabaudiin that the anti-Pc(em)93 titers are highest whenthe parasitemia is falling. Especially notable isthe high titer of antibody observed after rein-fection with virtually no parasitemia. However,vaccination experiments do not show a directrole for Pc(em)93 in anti-malarial immunity.Vaccination with Pf155/RESA has thus far beenambiguous. Initial experiments suggested thatPf155/RESA protectedAotusmonkeys fromoverwhelming parasitemia (Collinset al.1986);however, protection was not observed in latertrials (Collinset al. 1991; Pyeet al. 1991). Inaddition, several studies have found no correla-tion between antibodies against Pf155/RESAand immunity (Duboiset al. 1993; IqbalMDBUet al. 1994; Al-Yamanet al. 1995). Instead,prevalence of anti-Pf155/RESA antibodies cor-related with age which presumably reflects agreater incidence of exposure to the parasite.Similarly, the prevalence of seropositivity toGLURP increased with age, and in contrast tothe above study (Hoghet al. 1992), a positivecorrelation between seropositivity and para-sitemia was reported (Dziegielet al. 1993).Therefore, antibodies against glutamate-richproteins may only indicate a previous exposureto parasites.The fact that so many malarial antigens are
ANTIGENICITY OF GLU-RICH PlasmodiumPROTEINS 51

highly antigenic raises some questions about theusefulness of serological studies on isolatedpolypeptides or synthetic peptides in terms ofevaluating whether a particular protein may beinvolved in immunity. For example, anti-Pc(em)93 antibodies are readily detected whenaffinity-purified Pc(em)93 was used as the sub-strate in immunoblotting. However, Pc(em)93could not be positively identified on immunob-lots of crude extracts using immune sera (Fig.6). These results demonstrate that looking at anantibody response to a purified antigen maygive a misperception of the importance of thatantigen with regard to the total immune re-sponse of the host. Human serological studiesusually involve the use of recombinant or syn-thetic peptides on ELISA plates at relativelyhigh concentrations. Such serological studiesalso have the potential to exaggerate the immu-nological importance of an antigen as was foundto be the case with Pc(em)93 in this study.The fact that the malarial parasite produces
highly immunogenic proteins is an enigma. Ithas been proposed that proteins containing tan-dem repeats may serve as a smokescreens todivert the immune response toward antigens notinvolved in protection (Anders, 1986), and sucha role for Pc(em)93 cannot be excluded. Con-sistent with the smokescreen hypothesis, manyof the glutamate-rich proteins are relativelyabundant and are localized to the compartmentsoutside of the parasite but within the infectederythrocyte. Presumably, these proteins are re-leased and exposed to the host immune systemduring erythrocyte lysis and merozoite invasion.However, we propose an alternative explana-tion. Pc(em)93 was first described as a proteininteracting with cytoplasmic face of the eryth-rocyte membrane (Wiseret al. 1988, 1993).Furthermore, differential solubility data (Wiser1987) and the ability to bind to inside-outvesicles prepared from erythrocyte membranes(Wiseret al.1990) suggest that Pc(em)93 inter-acts with the submembrane cytoskeleton. BothMESA (Lustigmanet al. 1990) and Pf155/RESA (Foleyet al.1991) also interact with pro-teins of the submembrane erythrocyte cytoskel-eton. Many cytoskeletal proteins consist largely
of a-helical domains, tend to be acidic, and arelong rod-shaped, or fibrous, molecules. Analy-sis of the predicted secondary structures ofPc(em)93 suggests that the glutamate-rich tan-dem repeats may bea-helices (data not shown).Similar observations of predicteda-heliceshave been made with respect to the glutamate-rich repeats of MESA (Coppel 1992) and a cy-toskeletal protein fromTrypanosoma cruziwhich contains repeats rich in glutamate andglutamate dimers (Cotrimet al.1995). Further-more, analysis of Pc(em)93 by gel filtration andnondenaturing gel electrophoresis suggests thatit may be a long rod-shaped protein (Wiser andGrab, unpublished results). The highly immu-nogenic glutamate-rich proteins in the malarialparasite may therefore simply reflect structuralconstraints, which presumably reflect the func-tions of the proteins.
ACKNOWLEDGMENTS
We gratefully acknowledge the excellent technical assis-tance of Maryetta Brooks in carrying out these experimentsand Dr. Dennis Grab for his comments on the manuscript.We also thank Drs. Chingchai Wanidworanum and HannaShear for providing mAb-6C12E7 and mAb-8F2G9 and Dr.Willy Deleersnijder for providing mAb-43. This researchwas supported by NIH Grant AI31083.
REFERENCES
AL-YAMAN , F., GENTON, B., FALK , M., ANDERS, R. E.,LEWIS, D., HII, J., BECK, H. P.,AND ALPERS, M. P. 1995.Humoral response toPlasmodium falciparumring-infected erythrocyte surface antigen in a highly endemicarea of Papua New Guinea.American Journal of TropicalMedicine and Hygiene52, 66–71.
ANDERS, R. F. 1986. Multiple cross-reactivities amongst an-tigens ofPlasmodium falciparumimpair the developmentof protective immunity against malaria.Parasite Immu-nology8, 529–539.
BARNES, D. A., WOLLISH, W., NELSON, R. G., LEECH, J. H.,AND PETERSEN, C. 1995.Plasmodium falciparum:D260,an intraerythrocytic parasite protein, is a member of theglutamic acid dipeptide-repeat family of proteins.Experi-mental Parasitology81, 79–89.
BORRE, M. B., DZIEGIEL, M., HOGH, B., PETERSEN, E.,RIENECK, K., RILEY, E., MEIS, J. F., AIKAWA , M., NAKA-MURA, K., HARADA, M., WIND, A., JAKOBSEN, P. H., COW-LAND, J., JEPSEN, S., AXELSEN, N. H.,AND VUUST, J. 1991.Primary structure and localization of a conserved immu-nogenicPlasmodium falciparumglutamate rich protein(GLURP) expressed in both the preerythrocytic and
WISER ET AL.52

erythrocytic stages of the vertebrate life cycle.Molecularand Biochemical Parasitology49, 119–131.
BOYLE, D. B., NEWBOLD, C. I., SMITH, C. C., AND BROWN,K. N. 1982. Monoclonal antibodies that protect in vivoagainstPlasmodium chabaudirecognize a 250,000-Dalton parasite polypeptide.Infection and Immunity38,94–102.
COLLINS, W. E., ANDERS, R. F., PAPPAIOANOU, M., CAMP-BELL, G. H., BROWN, G. V., COPPEL, R. L., SKINNER, J. C.,ANDRYSIAK, P. M., FAVALORO, J. M., CORCORAN, L. M.,BRODERSON, J. R., MITCHELL, G. F.,AND CAMPBELL, C. C.1986. Immunization ofAotusmonkeys with recombinantprotein of an erythrocyte surface antigen ofPlasmodiumfalciparum. Nature323,259–262.
COLLINS, W. E., ANDERS, R. F., RUEBUSH, T. K., KEMP,D. J., WOODROW, G. C., CAMPBELL, G. H., BROWN, G. V.,IRVING, D. O., GOSS, N., AND FILIPSKI, V. K. 1991. Im-munization of owl monkeys with the ring-infected eryth-rocyte surface antigen ofPlasmodium falciparum. Ameri-can Journal of Tropical Medicine and Hygiene44, 34–41.
COPPEL, R. L. 1992. Repeat structures in aPlasmodium fal-ciparumprotein (MESA) that binds to human erythrocyteprotein 4.1.Molecular and Biochemical Parasitology50,335–347.
COPPEL, R. L., COWMAN, A. F., ANDERS, R. F., BIANCO,A. C., SAINT, R. B., LINGELBACH, K. R., KEMP, D. J.,ANDBROWN, G. V. 1984. Immune sera recognize an erythro-cyte Plasmodium falciparumantigen composed of re-peated amino acid sequences.Nature310,789–792.
COTRIM, P. C., PARANHOS-BACCALA, G., SANTOS, M. R.,MORTENSEN, C., CANO, M. I., JOLIVET, M., CAMARGO,M. E., MORTARA, R. A., AND DA SILVEIRA , J. F. 1995.Organization and expression of the gene encoding an im-munodominant repetitive antigen associated to the cyto-skeleton ofTrypanosoma cruzi. Molecular and Bio-chemical Parasitology71, 89–98.
DELEERSNIJDER, W., PRASOMSITTI, P., TUNGPRADUBKUL, S.,HENDRIX, D., HAMERS-CASTERMAN, C., AND HAMERS, R.1992. Structure of aPlasmodium chabaudiacidic phos-phoprotein that is associated with the host erythrocytemembrane.Molecular and Biochemical Parasitology56,59–68.
DUBOIS, B., DELORON, P., ASTAGNEAU, P., CHOUGNET, C.,AND LEPERS, J. P. 1993. Isotypic analysis ofPlasmodiumfalciparum-specific antibodies and their relation to pro-tection in Madagascar.Infection and Immunity61,4498–4500.
DZIEGIEL, M., ROWE, P., BENNETT, S., ALLEN, S. J., OLERUP,O., GOTTSCHAU, A., BORRE, M., AND RILEY, E. M. 1993.Immunoglobulin-M and immunoglobulin-G antibody re-sponses toPlasmodium falciparumglutamate-rich pro-tein—Correlation with clinical immunity in Gambianchildren.Infection and Immunity61, 103–108.
FAVALORO, J. M., COPPEL, R. L., CORCORAN, L. M., FOOTE,S. J., BROWN, G. V., ANDERS, R. F., AND KEMP, D. J.
1986. Structure of the RESA gene ofPlasmodium falci-parum. Nucleic Acids Research14, 8265–8277.
FOLEY, M., TILLEY , L., SAWYER, W. H., AND ANDERS, R. F.1991. The ring-infected erythrocyte surface antigen ofPlasmodium falciparumassociates with spectrin in theerythrocyte membrane.Molecular and BiochemicalParasitology46, 137–148.
GABRIEL, J. A., HOLMQUIST, G., PERLMANN, H., BERZINS, K.,WIGZELL, H., AND PERLMANN, P. 1986. Identification of aPlasmodium chabaudiantigen present in the membraneof ring stage infected erythrocytes.Molecular and Bio-chemical Parasitology20, 67–75.
HARTZ, D., AYANE, M., CHLUBA-DE TAPIA, J., WIRBELAUER,C., LANGHORNE, J., AND GILLARD-BLAAS, S. 1993. Clon-ing and sequencing of a cDNA fragment fromPlasmo-dium chabaudi chabaudithat contains repetitive se-quences coding for a potentially lysine-rich aspartic acid-rich protein.Parasitology Research79, 133–139.
HOGH, B., PETERSEN, E., DZIEGIEL, M., DAVID , K., HANSON,A., BORRE, M., HOLM, A., VUUST, J., AND JEPSEN, S.1992. Antibodies to a recombinant glutamate-richPlas-modium falciparumprotein—Evidence for protection ofindividuals living in a holoendemic area of Liberia.American Journal of Tropical Medicine and Hygiene46,307–313.
HOLMQUIST, G., NAGASAWA, N., BERZINS, K., SNOUNOU, G.,VIRIYAKOSOL, S., AIKAWA , M., WIGZELL, H., AND
PERLMANN, P. 1990.Plasmodium chabaudi:Polymorphicand nonpolymorphic epitopes of the antigen Pch105/RESA.Experimental Parasitology70, 436–442.
IQBAL, J., RAB, A., PERLMANN, P., AND BERZINS, K. 1994.Humoral immune responses toPlasmodium falciparumantigens in children and adults living in a hypoendemicarea of Punjab (Pakistan).American Journal of TropicalMedicine and Hygiene51, 444–453.
KEMP, D. J., COPPEL, R. L., AND ANDERS, R. F. 1987. Re-petitive proteins and genes of malaria.Annual Review ofMicrobiology41, 181–208.
LANDAU, I., AND KILLICK -KENDRICK, R. 1966. Rodent plas-modia of the Republique Centrafricaine: The sporogonyand tissue stages ofPlasmodium chabaudiandP. bergheiyoelii. Transactions of the Royal Society of TropicalMedicine and Hygiene60, 633–649.
LUSTIGMAN, S., ANDERS, R. F., BROWN, G. V., AND COPPEL,R. L. 1990. The mature parasite-infected erythrocyte sur-face antigen (MESA) associates with the erythrocytemembrane skeletal protein, band 4.1.Molecular and Bio-chemical Parasitology38, 261–270.
MATTEI, D., AND SCHERF, A. 1992. The Pf332 gene ofPlas-modium falciparumcodes for a giant protein that is trans-located from the parasite to the membrane of infectederythrocytes.Gene110,71–79.
PETERSEN, E., HOGH, B., MARBIAH, N. T., PERLMANN, H.,WILCOX, M., DOLOPAIE, E., HANSON, A. P., AND BJORK-MANN. A. 1990. A longitudinal study of antibodies to thePlasmodium falciparumantigen Pf155/RESA and immu-nity to malaria infection in adult Liberians.Transactions
ANTIGENICITY OF GLU-RICH PlasmodiumPROTEINS 53

of the Royal Society of Tropical Medicine and Hygiene84, 339–345.
PYE, D., EDWARDS, S. J., ANDERS, R. F., O’BRIEN, C. M.,FRANCHINA, P., CORCORAN, L. N., MONGER, C., PETER-SON, M. G., VANDENBERG, K. L., SMYTHE, J. A., WEST-LEY, S. R., COPPEL, R. L., WEBSTER, T. L., KEMP, D. J.,HAMPSON, A. W., AND LANGFORD, C. J. 1991. Failure ofrecombinant vaccinia viruses expressingPlasmodium fal-ciparumantigens to protectSaimirimonkeys against ma-laria. Infection and Immunity59, 2403–2411.
RILEY, E. M., ALLEN, S. J., TROYE-BLOMBERG, M., BEN-NETT, S., PERLMANN, H., ANDERSON, G., SMEDMAN, L.,PERLMANN, P., AND GREENWOOD, B. M. 1991. Associa-tion between immune recognition of the malaria vaccinecandidate antigen Pf155/RESA and resistance to clinicaldisease: A prospective study in a malaria-endemic regionof West Africa. Transactions of the Royal Society ofTropical Medicine and Hygiene85, 436–443.
SCHERF, A., CARTER, R., PETERSEN, C., ALANO, P., NELSON,R., AIKAWA , M., MATTEI, D., DASILVA , L. P.,AND LEECH,J. 1992. Gene inactivation of Pf11-1 ofPlasmodium fal-ciparum by chromosome breakage and healing—Identification of a gametocyte-specific protein with a po-tential role in gametogenesis.EMBO Journal11, 2293–2301.
SCHMITT-WREDE, H. P., QIAO, Z. D., AND WUNDERLICH, F.1993. A cDNA putatively encoding the Pc90-erythrocytemembrane antigen ofPlasmodium chabaudi. Parasitol-ogy Research79, 80–81.
STEVENSON, M. M., AND TAM, M. F. 1993. Differential in-duction of helper T-cell subsets during blood-stagePlas-modium chabaudiAS infection in resistant and suscep-tible mice.Clinical and Experimental Immunology92,77–83.
TAYLOR-ROBINSON, A. W., AND PHILLIPS, R. S. 1994. B cellsare required for the switch from Th1- to Th2-regulatedimmune responses toPlasmodium chabaudi chabaudiin-fection. Infection and Immunity62, 2490–2498.
VON DERWEID, T., KOPF, M., KOHLER, G.,AND LANGHORNE,J. 1994. The immune response toPlasmodium chabaudimalaria in interleukin-4-deficient mice.European Jour-nal of Immunology24, 2285–2293.
WANIDWORANUM, C., BARNWELL, J. W., AND SHEAR, H. L.1987. Protective antigen in the membranes of mouse
erythrocytes infected withPlasmodium chabaudi. Mo-lecular and Biochemical Parasitology25, 195–201.
WILLIAMSON, K. C., CRISCIO, M. D., AND KASLOW, D. C.1993. Cloning and expression of the gene forPlasmo-dium falciparum transmission-blocking target antigen,Pfs230.Molecular and Biochemical Parasitology58,355–358.
WISER, M. F. 1987. Phosphoproteins associated with thehost erythrocyte membrane duringPlasmodium chabaudiinfection. In ‘‘Host–Parasite Cellular and Molecular In-teractions in Protozoal Infections’’ (K. P. Chang and D.Snary, Eds.), pp. 315–319. Springer-Verlag, Berlin.
WISER, M. F., AND SCHWEIGER, H.-G. 1986. Increased sen-sitivity in antigen detection during immunoblot analysisresulting from antigen enrichment via immunoprecipita-tion. Analytical Biochemistry155,71–77.
WISER, M. F., LEIBLE, M. B., AND PLITT, B. 1988. Acidicphosphoproteins associated with the host erythrocytemembrane of erythrocytes infected withPlasmodiumberghei and P. chabaudi. Molecular and BiochemicalParasitology27, 11–22.
WISER, M. F., SARTORELLI, A. C., AND PATTON, C. L. 1990.Association ofPlasmodium bergheiproteins with thehost erythrocyte membrane: Binding to inside-outvesicles.Molecular and Biochemical Parasitology38,121–134.
WISER, M. F., FAUR, L. V. V., LANNERS, H. N., KELLY, M.,AND WILSON, R. B. 1993. Accessibility and distributionof intraerythrocytic antigens ofPlasmodium-infectederythrocytes following mild glutaraldehyde fixation anddetergent extraction.Parasitology Research79,579–588.
WUNDERLICH, F., BRENNER, H. H., AND HELWIG, M. 1988a.Plasmodium chabaudimalaria: Protective immunizationwith surface membranes of infected erythrocytes.Infec-tion and Immunity56, 3326–3328.
WUNDERLICH, F., HELWIG, M., SCHILLINGER, G., SPETH, V.,AND WISER, M. F. 1988b. Expression of the parasite pro-tein Pc90 in plasma membranes of erythrocytes infectedwith Plasmodium chabaudi. European Journal of CellBiology47, 157–164.
Received 4 June 1996; accepted with revision 24 October1996
WISER ET AL.54
Copyright © 2022 FDOKUMEN




















