
Salt-induced antioxidant metabolism defenses in maize ( Zea mays L.) seedlings
10
INTRODUCTION Antioxidant defense systems evolved along with aerobic metabolism to protect living organisms against the poten- tial damaging effects of reactive oxygen species (ROS). 1 ROS are produced through many metabolic reactions as a consequence of aerobic metabolism 1 and act as signalling molecules in various physiological processes. 2,3 ROS can also interact with many cellular components causing sig- nificant damage due to peroxidative chain reactions tar- geted at lipids, proteins, and nucleic acids. 1,4 Among ROS detoxifying activities, catalases (CAT) and superoxide dismutases (SOD) are front-line defenses, removing H 2 O 2 and O 2 – , respectively. They are present virtually in all tissues and throughout plant development. In maize, expression of the three CAT and nine SOD isozymes and their encoding genes is regu- lated by genetic, developmental, and environmental sig- nals. 5,6 Their regulation by abscisic acid (ABA), a stress-related growth regulator, and by osmotic stress is dependent on tissue and developmental stage. 7–10 Increased production and accumulation of ROS are also triggered by many environmental stresses. 2,11 Among these, salinity is a major problem throughout the world. 12 Soil salinity effects are complex and diverse, imposing both osmotic and ionic imbalances that in the © W. S. Maney & Son Ltd Redox Report, Vol. 9, No. 1, 2004 DOI 10.1179/135100004225003888 Research article Salt-induced antioxidant metabolism defenses in maize (Zea mays L.) seedlings Larissa Menezes-Benavente 1,2 , Sheri P. Kernodle 2 , Márcia Margis-Pinheiro 1 , John G. Scandalios 2 1 Laboratory for Plant Molecular Genetics, Department of Genetics, Federal University of Rio de Janeiro, Brazil 2 Department of Genetics, North Carolina State University, North Carolina, USA Salinity alters general metabolic processes and enzymatic activities, causing increased production of reactive oxygen species (ROS). Expression of antioxidant defense genes would, in turn, be triggered to defend the cell against oxidative damage. We report that salt disturbed antioxidant metabolism in maize seedlings, causing detrimental effects on the growth and development of maize plantlets, increased hydrogen peroxide production and altered antioxidant activities and transcripts profiles. Excessive ROS levels were accompanied by increased catalase (CAT) activity in photosynthesizing shoots, along with induction of mRNA accumulation. Increased accumulation of superoxide dismutase (SOD) transcripts was also observed although no significant changes in total SOD enzymatic activity and isozyme profiles were detected. Higher salt concentrations (above 0.25 M NaCl) were highly detrimental to the plants, causing arrested growth and severe wilting, among other effects. Histochemical detection of H 2 O 2 by 3,3-diaminobenzidine (DAB) staining indicated a collapse of the leaf veins, with hydrogen peroxide leaking to neighboring cells. In agreement to these observations, Sod1, Sod2, Sod4, Sod4A, as well as all Cat transcripts were severely inhibited in plants exposed to high salt concentrations. Keywords : Salinity, antioxidants, metabolism, defenses, maize Received 13 August 2003 Revised 10 October 2003 Accepted 3 December 2003 Correspondence to: Larissa Menezes-Benavente, Lab. Genética Molecular Vegetal, Dep. Genética, Sala A2-76, CCS, Universidade Federal do Rio de Janeiro, Ilha do Fundão, s/n, Rio de Janeiro, RJ – Brazil, CEP 21944-970 Tel: +55 21 2562 6381; Fax: +55 21 2590 0111; E-mail: [email protected] Abbreviations: CAT, catalase; SOD, superoxide dismutase; DAB, 3,3- diaminobenzidine; ABA, abscisic acid; dpi, days post imbibition; MS, Murashige and Skoog medium; PAGE, polyacrylamide gel electro- phoresis; EST, expressed sequence tag; APx, ascorbate peroxidase; GR, glutathione reductase
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Salt-induced antioxidant metabolism defenses in maize ( Zea mays L.) seedlings

INTRODUCTION
Antioxidant defense systems evolved along with aerobicmetabolism to protect living organisms against the poten-tial damaging effects of reactive oxygen species (ROS).1
ROS are produced through many metabolic reactions as aconsequence of aerobic metabolism1 and act as signallingmolecules in various physiological processes.2,3 ROS canalso interact with many cellular components causing sig-nificant damage due to peroxidative chain reactions tar-geted at lipids, proteins, and nucleic acids.1,4
Among ROS detoxifying activities, catalases (CAT)and superoxide dismutases (SOD) are front-linedefenses, removing H2O2 and O2
�–, respectively. Theyare present virtually in all tissues and throughout plantdevelopment. In maize, expression of the three CAT andnine SOD isozymes and their encoding genes is regu-lated by genetic, developmental, and environmental sig-nals.5,6 Their regulation by abscisic acid (ABA), astress-related growth regulator, and by osmotic stress isdependent on tissue and developmental stage.7–10
Increased production and accumulation of ROS arealso triggered by many environmental stresses.2,11
Among these, salinity is a major problem throughout theworld.12 Soil salinity effects are complex and diverse,imposing both osmotic and ionic imbalances that in the
© W. S. Maney & Son LtdRedox Report, Vol. 9, No. 1, 2004DOI 10.1179/135100004225003888
Research article
Salt-induced antioxidant metabolism defenses in maize (Zea mays L.) seedlings
Larissa Menezes-Benavente1,2, Sheri P. Kernodle2, Márcia Margis-Pinheiro1, John G. Scandalios2
1Laboratory for Plant Molecular Genetics, Department of Genetics, Federal University of Rio de Janeiro, Brazil2Department of Genetics, North Carolina State University, North Carolina, USA
Salinity alters general metabolic processes and enzymatic activities, causing increased productionof reactive oxygen species (ROS). Expression of antioxidant defense genes would, in turn, betriggered to defend the cell against oxidative damage. We report that salt disturbed antioxidantmetabolism in maize seedlings, causing detrimental effects on the growth and development of maizeplantlets, increased hydrogen peroxide production and altered antioxidant activities and transcriptsprofiles. Excessive ROS levels were accompanied by increased catalase (CAT) activity inphotosynthesizing shoots, along with induction of mRNA accumulation. Increased accumulation ofsuperoxide dismutase (SOD) transcripts was also observed although no significant changes in totalSOD enzymatic activity and isozyme profiles were detected. Higher salt concentrations (above 0.25M NaCl) were highly detrimental to the plants, causing arrested growth and severe wilting, amongother effects. Histochemical detection of H2O2 by 3,3-diaminobenzidine (DAB) staining indicated acollapse of the leaf veins, with hydrogen peroxide leaking to neighboring cells. In agreement tothese observations, Sod1, Sod2, Sod4, Sod4A, as well as all Cat transcripts were severely inhibitedin plants exposed to high salt concentrations.
Keywords: Salinity, antioxidants, metabolism, defenses, maize
Received 13 August 2003Revised 10 October 2003Accepted 3 December 2003
Correspondence to: Larissa Menezes-Benavente, Lab. GenéticaMolecular Vegetal, Dep. Genética, Sala A2-76, CCS, UniversidadeFederal do Rio de Janeiro, Ilha do Fundão, s/n, Rio de Janeiro, RJ –Brazil, CEP 21944-970Tel: +55 21 2562 6381; Fax: +55 21 2590 0111; E-mail: [email protected]
Abbreviations: CAT, catalase; SOD, superoxide dismutase; DAB, 3,3-diaminobenzidine; ABA, abscisic acid; dpi, days post imbibition; MS,Murashige and Skoog medium; PAGE, polyacrylamide gel electro-phoresis; EST, expressed sequence tag; APx, ascorbate peroxidase;GR, glutathione reductase

long term cause arrested growth and even death.Disturbance of membrane integrity, nutrient uptake, growthregulators, and general metabolic activities, including pho-tosynthesis, are some of the physiological and biochemicalconsequences.11,13 Enhanced ROS production further aggra-vates primary effects of disruption of cell water status andionic balance.11
This study aimed to complement the array of data com-piled on the regulation of catalases and superoxide dismu-tases in maize. The information accumulated thus faroffers a unique opportunity to use this model system as atool to provide a better understanding of the correlationsbetween environmental stress and antioxidant metabolism.Antioxidant metabolism changes in response to salt treat-ment have been reported in rice,14 Nicotiana plumbaginifo-lia,15 and other species.16,17 In this study, phenotypiceffects, CAT and SOD total activity, isozyme pattern, andtranscript profiles were evaluated in response to NaCl. Theresults presented here indicate that salt induced a complexand concerted antioxidant response in maize.
MATERIALS AND METHODS
Plant cultivation and stress treatment
The maize (Zea mays) in-bred line W64A was used in thisstudy. Seeds were surface sterilized with 20% commercialbleach for 10 min and rinsed 6 times with distilled water.After 24 h imbibition in water, seeds were planted inMetromix. Plants were cultivated in a growth chamber, at 26± 0.5°C, under a 12-h photoperiod, and were watered daily.Light was provided by fluorescent lamps. At nine days postimbibition (9 dpi), seedlings were exposed to different con-centrations of NaCl (0.1 M, 0.25 M, 0.5 M, 0.75 M or 1.0 M)in water or half-strength MS salts solution (0.5x MS),18 for 24h, 48 h, or 72 h. Controls comprised of plants that remainedin soil or that were transferred to deionized water or 0.5x MS.Fresh-weight measurements were taken immediately beforeand after salt exposure. Seedlings shoot growth was alsorecorded as a second indication of physiological stress.
Protein extraction and enzymatic assays
Leaf and root protein samples were prepared by grindingfresh tissues in ice-cold 0.025 M glycylglycine bufferpH 7.4 with polyvinylpyrrolidone (PVP-40) and sand.Crude extracts were obtained after centrifugation at13,000 rpm, at 4°C, for 15 min. Total protein was deter-mined according to Lowry et al.19 Catalase activity andsuperoxide dismutase activity were assayed as describedpreviously.20,21 Activity gels were performed using equalprotein loadings to evaluate changes in isozyme pattern.For catalase activity, native 6% PAGE was prepared
using the Tris-glycine buffer system and stained asdescribed.22,23 For superoxide dismutase activity, native 9%PAGE was prepared using the Tris-glycine buffer systemand stained as described.21 For statistical analyses of thedata, the analysis of variance was performed using theGLM procedure and Fisher’s protected LSD of the SASStatistical Package (SAS Institute, Cary, NC, USA).
RNA extraction and Northern blot analysis
Total RNA was extracted from leaves.24 For Northern blotanalysis, 20 mg of total RNA were separated in 1.6% dena-turing agarose-formaldehyde gels and transferred to nylonmembranes.25 Blots were air dried, vacuum-baked at 70°C,25 mmHg, for 30 min and cross-linked (120 mJ, UVStratalinker, Stratagene). Duplicate blots were hybridizedwith [32P]-labeled gene-specific probes for one of the cata-lases (Cat1, Cat2, Cat3) or superoxide dismutases (Sod1,Sod2, Sod4, Sod4A), in modified Church buffer.26 Sod3 is afull-length probe that hybridizes to all four members of theSod3 gene family (Sod3.1, Sod3.2, Sod3.3, and Sod3.4).APx and GR probes were rice ESTs (Genebank accessionnumbers C28227 and AU056093, respectively), homolo-gous to the corresponding rice cDNAs, and were kindlyprovided by the Rice Genome Research Program, NIASDNA Bank, Japan. Probes were prepared by random prim-ing.27 Blots were washed once (APx and GR) or twice(CATs and SODs) under stringent conditions (0.1x SSC,0.1% SDS; 65°C). Equal RNA loading was confirmedwith an 18-S ribosomal DNA probe from pea.28
Densitometry analyses were performed using the publicdomain NIH ImageJ program from the US NationalInstitutes of Health (available at <http://rsb.info.nih.gov/ij/>).
Hydrogen peroxide detection
In situ histochemical localization of H2O2 was per-formed according to Thordal-Christensen et al.29 Thistechnique is based on H2O2-catalyzed polymerization of3,3-diaminobenzidine (DAB). Seedlings were cut in theshoot basal region and supplied with a 1 mg/ml solutionof DAB for 16 h. The DAB taken up by the leaves andpolymerized in the presence of H2O2 was visualized asan insoluble red polymer after boiling in ethanol (96%)for 10 min, and chlorophyll washing in ethanol.
RESULTS
Phenotypic effects
The fresh weight of seedlings was recorded as an indica-tion of the whole-plant effect of salt treatment. Seedlings
30 Menezes-Benavente, Kernodle, Margis-Pinheiro,Scandalios

transferred to 0.5x MS solution sustained a steady gainof fresh weight (Fig. 1). When exposed to 0.1 M NaCl,the plantlets were still able to sustain fresh weight gain.At 0.25 M NaCl, fresh weight gain was significantlyinhibited after 24 h and 48 h of exposure. At 72 h, a sig-nificant weight loss was actually observed, ~4.3–9.8%of initial fresh weight. At 0.5 M NaCl, fresh weight losswas observed at all time exposures. High standard devi-ations are likely due to the solution that remained onroots during weight measurements, besides intrinsicvariation among the seedlings. Furthermore, shootgrowth was severely compromised. Significant differ-ences were observed at all salt concentrations. In controlplantlets, growth rates varied from 25–50% of initialshoot length in both soil and 0.5x MS-cultivatedseedlings (Fig. 1). On the other hand, while plantsexposed to 0.1 M NaCl were still able to support areduced growth rate, higher concentrations almost com-pletely inhibited shoot growth. At 0.25 M NaCl, growthrates were ~2.3–4.12% of initial shoot length. Higherconcentrations (0.5 M) totally abolished shoot growth.
Formation of brownish spots and stripes in leaf blades,shoot wilting, inhibition of secondary leaf and rootgrowth were observed after 48 h. After 72 h of exposure,besides the symptoms described above, severe wiltingand formation of small salt crystals on the leaf bladewere also observed, spots and stripes formed on leafedges and tips, and leaf pigmentation was altered.
Salt stress has been associated with increased accumu-lation of hydrogen peroxide.16,30,31 To evaluate if anequivalent response occurred in maize, seedlings werestained with DAB for in situ detection of H2O2.
29 In con-trol plants, grown in soil and 0.5x MS, DAB stainingwas associated with lignification of vascular tissues.29
Staining intensity increased as seedlings developed,being stronger in older plantlets. In salt-treatedseedlings, increased H2O2 accumulation was visualizedin response to 0.1 M NaCl after 48 h and 72 h and inresponse to 0.25 M NaCl for 24 h (data not shown).Longer exposures (48 h and 72 h) to 0.25 M NaCl didnot enhance the staining intensity, which was decreasedin comparison to the intensity visualized after 24 h ofstress. Furthermore, the DAB staining restricted to thevascular tissue observed in non-stressed seedlings wasaltered upon NaCl exposure to 0.1 M for 48 h and 72 hand to 0.25 M for 24 h. In these plants, the staining pat-tern observed was suggestive of H2O2 leakage from thevascular tissue to surrounding cells (data not shown). Inseedlings exposed to 0.5 M NaCl, no staining could beobserved (data not shown).
Salt-induced antioxidant metabolism defenses in maize (Zea mays L.) seedlings 31
Fig. 1. Phenotypic effects of salt stress on maize seedlings. Fresh weightvariation and shoot growth were analyzed in response to increasingconcentrations of NaCl. Controls were plants that remained on soil or thatwere transferred to half-strength MS solution. Vertical bars indicate SD.a,bIndicate means that are significantly different from both soil andmedium controls (P < 0.05). Fig. 2. Catalase activity and isozyme profile in salt stressed maize. 9 dpi
W64A seedlings were exposed to increasing NaCl concentrations for 24 h,48 h and 72 h, and total CAT activity was determined. Leaf isozymeprofiles from seedlings exposed to 0.25 M NaCl were determined bynative 6% PAGE as described in Materials and Methods. Controls wereplants that remained on soil or that were transferred to water or half-strength MS solution. Vertical bars indicate SD. a,bIndicate means that aresignificantly different from both soil and medium controls (P < 0.05).
Catalase activity and isozymes
Quantification of total catalase activity and evaluation ofisozyme pattern were performed. In leaves, total catalaseactivity was low. A moderate increase in CAT activity inresponse to salt was observed (Fig. 2). This effect wasmore pronounced in response to 0.25 M NaCl. The
short-term induction in total CAT activity observed at 24 hwas maintained at 48 h and 72 h (Fig. 2). In the two repe-titions performed, increased activity was observed withboth 0.1 M (24.4–31.2%) and 0.25 M (51.5–59.9%) NaCl,compared to soil and 0.5x MS controls. High standarddeviations were due to very low levels of activity; how-ever, the results were statistically significant at P < 0.05.
32 Menezes-Benavente, Kernodle, Margis-Pinheiro,Scandalios
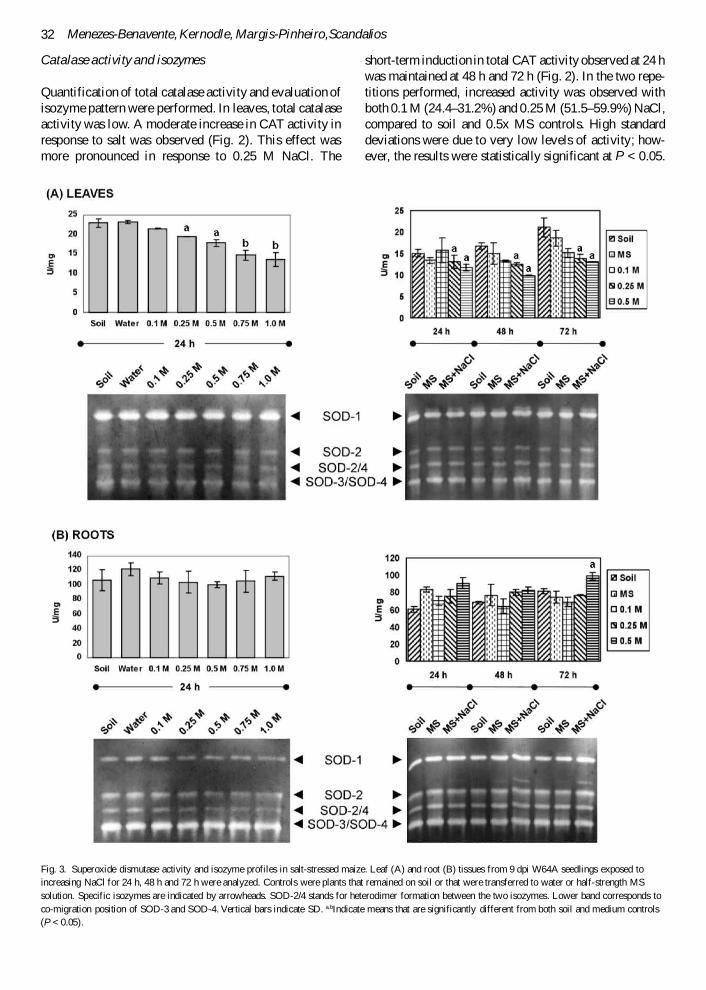
Fig. 3. Superoxide dismutase activity and isozyme profiles in salt-stressed maize. Leaf (A) and root (B) tissues from 9 dpi W64A seedlings exposed toincreasing NaCl for 24 h, 48 h and 72 h were analyzed. Controls were plants that remained on soil or that were transferred to water or half-strength MSsolution. Specific isozymes are indicated by arrowheads. SOD-2/4 stands for heterodimer formation between the two isozymes. Lower band corresponds toco-migration position of SOD-3 and SOD-4. Vertical bars indicate SD. a,bIndicate means that are significantly different from both soil and medium controls(P < 0.05).

Higher salt concentration (0.5 M) did not alter CAT activ-ity. Isozyme analysis of the 0.25 M NaCl-treated shootsrevealed CAT-2 as the main isozyme responsible forincreases in activity (Fig. 2, bottom). In roots, CAT activ-ity was extremely low and difficult to detect by the spec-trophotometric assay. Isozyme analysis was only possiblewith very sensitive staining procedures. In roots, the pre-dominant isozyme was CAT-1. However, no significantchanges were observed (data not shown).
Superoxide dismutase activity and isozymes
Total SOD activity and isozyme patterns were evaluatedin both leaves and roots. In leaves, total SOD activityshowed an inverse correlation to salt concentration, thatis, increasing salt concentrations caused diminishedactivity (Fig. 3A). Electrophoretic analysis did not showany difference in isozyme pattern and staining intensity.Longer exposures (48 h and 72 h) produced similareffects in leaves, causing diminished activity in responseto increasing salt concentrations (Fig. 3A). Furthermore,while in control plants grown in soil a clear trend ofincreasing SOD activity could be detected as the plantsaged,32 this behavior was abolished by salt treatment. Nosignificant changes in isozyme pattern were observed at0.25 M NaCl throughout the experimental period. Inroot tissues, total SOD activity was not significantlyaffected by exposing the seedlings to NaCl concentra-tions up to 0.25 M for a maximum of 72 h when com-pared to 0.5x MS control (Fig. 3B). The apparentinduction compared to soil controls may be explained asan effect of the medium itself, since this was able tocause an increase in activity. A slight induction wasobserved only at a high salt concentration (0.5 M) and
prolonged exposure (72 h). Isozyme patterns fromseedlings exposed to 0.25 M NaCl revealed one addi-tional band in 0.5x MS and salt-treated samples after 48h and 72 h, although this band was barely detectable at24 h (Fig. 3B). The proper identification of this bandrequires further investigation.
Gene expression profiles
The influence of salt stress on Cat and Sod transcriptsfrom 9 dpi W64A shoots was analyzed. Seedlings wereexposed to increasing NaCl concentrations for a shortperiod (24 h) or to 0.25 M NaCl for longer exposures (48h and 72 h). Controls comprised of plants that remainedin soil or that were transferred to deionized water.
Each catalase transcript exhibited a particular expres-sion profile (Fig. 4). In control plants, Cat3 was the mostabundant transcript detected, Cat2 was weaklyexpressed and Cat1 barely detected. Cat2 was weaklyinduced at 0.1 M and 0.25 M NaCl, but inhibited byhigher concentrations. On the other hand, Cat1 transcriptaccumulation was significantly increased, compared tocontrols, at 0.25 M and 0.5 M NaCl (9-fold and 5-fold,respectively). A moderate induction in Cat3 (2-fold) wasdetected at 0.25 M NaCl. Catalase transcript responsesto longer exposures to 0.25 M NaCl were also studied.The short-term induction on Cat1 transcript accumulationat 24 h was also observed at 48 h and 72 h (8–9-fold). Cat2transcript levels remained fairly stable, not being consid-erably affected by this particular salt concentration. Onthe other hand, Cat3 transcripts were significantlydecreased in 0.5x MS-cultivated seedlings (~0.5-fold).This inhibitory effect masked the salt induction observed
Salt-induced antioxidant metabolism defenses in maize (Zea mays L.) seedlings 33
Fig. 4. Maize catalase transcript profiles in response to salt stress.Seedlings were exposed to increasing NaCl concentrations for 24 h (left),or to 0.25 M NaCl for 24 h, 48 h and 72 h (right). Controls comprised ofRNA extracted from plants that remained on soil or that were transferredto half-strength MS medium (MS). An 18-S DNA probe from pea rRNAwas used as loading control. The results are representative of twoindependently performed experiments.
Fig. 5. Maize superoxide dismutase transcript profiles in response to saltstress. Seedlings were exposed to increasing NaCl concentrations for 24 h,or to 0.25 M NaCl for 24 h, 48 h and 72 h. Controls comprised of RNAextracted from plants that remained on soil or that were transferred to half-strength MS medium (MS). An 18-S DNA probe from pea rRNA was usedas loading control. The results are representative of two independentlyperformed experiments.

in the short-term (24 h) treatment. However, upon addi-tion of 0.25 M NaCl to the 0.5x MS solution, Cat3 tran-script accumulation was once again induced by salt incomparison to 0.5x MS solution.
Except for Sod1, the expression of SOD genes was sig-nificantly up-regulated by 0.25 M NaCl (Fig. 5). For sometranscripts (e.g. Sod3), a slightly increased accumulationwas observed at 0.1 M NaCl. The most responsive geneswere Sod3 and Sod4. Sod4 levels were almost unde-tectable in controls, but significantly increased in responseto concentrations above 0.25 M (from 7-fold in 0.25 M to2-fold in 1.0 M). A similar behavior was observed forSod3 (2–3- fold induction), although basal levels werehigher than Sod4. Furthermore, Sod3 transcripts were sus-tained at higher levels even at 1.0 M NaCl (2.4-fold).Sod4A mRNA accumulation increased 2-fold at 0.25 MNaCl but returned to control levels above this concentra-tion. Sod1 transcript accumulation was inhibited by highsalt concentrations (0.5 M and above). Sod2 was not sig-nificantly affected. SOD transcript responses to longerexposures were analogous to the ones observed in theshorter 24 h treatment. Sod3, Sod4A and Sod4 were themost affected, being significantly up-regulated, in a time-dependent manner. A moderate induction (around 2-fold)was also observed for Sod2 at all exposures. Sod1 wasweakly inhibited by the treatments.
Besides Cat and Sod, transcript profiles for two otherantioxidant enzymes – ascorbate peroxidase (APx) andglutathione reductase (GR) – were also evaluated. Theseenzymes participate in the ascorbate/glutathione cycle,in which H2O2 is converted to H2O through a series ofreactions using ascorbate and glutathione as reductants.33
The probes used in this study were rice ESTs homolo-gous to cytosolic rice cDNAs. Both probes recognizedthe maize counterparts. While APx was stronglyexpressed in control leaf tissues, GR was barely detected(Fig. 6). This may be due to either low basal expressionlevels or to poor similarity of the rice GR probe andendogenous maize mRNA. However, exposure to saltsignificantly increased APx transcript accumulation,reaching a 3-fold induction at 72 h, while GR was mod-erately induced (Fig. 6). High salt concentrations inhib-ited their expression, and transcript levels were lowerthan controls.
DISCUSSION
Disruption of redox homeostasis is one of the variousproblems inflicted by salinity. Alterations in antioxidantmetabolism and genes have been described for severalplant species in relation to salt stress and tolerance.14–17 Inthe present study, CAT and SOD total activities, isozymepatterns, and transcript profiles were analyzed in responseto different NaCl concentrations and exposures.
Salinity causes disruption of water potential, affectingplant growth, development, and reproduction.11,13 Accord-ingly, salt-stressed maize seedlings displayed severe shootwilting, fresh weight loss, and impairment of shoot growth.High salt concentrations and longer exposures can also leadto the accumulation of ions in the cells and, consequently, saltinjuries. In this study, secondary effects, such as formation ofnecrotic areas, altered pigmentation and formation of saltcrystals, were observed after 2–3 days of treatment.
A range of environmental stresses, including salinity,triggers an increase in ROS levels.2,11 Besides increasingH2O2 accumulation, as qualitatively determined by DABstaining, NaCl exposure disrupted the staining patternrestricted to vascular tissue observed in non-stressedseedlings. Instead, H2O2 accumulated in surrounding tis-sues. Cellular membranes are permeable to H2O2 and it ispossible that it diffused to adjacent cells. More likely, theunbalanced cell redox state due to higher ROS levels couldhave caused oxidative damage to membranes, resulting incell collapse and leakage of the accumulated H2O2 to thesurroundings. The inverse correlation between stainingintensity and exposure time also suggests cell damage dueto enhanced H2O2 production and a consequent oxidativestress. The lack of DAB staining in response to higher saltconcentrations is suggestive of collapse of the vascular tis-sue, avoiding DAB absorption.
The increased CAT activity was not able to detoxify thegenerated H2O2 sufficiently, which accumulated in responseto NaCl. Furthermore, salt disrupted the developmental trendof increasing SOD activity in leaves and resulted in dimin-ished SOD activity, indicating disturbances in normal plantmetabolic processes. The combination of high SOD proteinturnover in stressful conditions,1,34 high intracellular ioniccontent, and oxidative damage due to increased ROS
34 Menezes-Benavente, Kernodle, Margis-Pinheiro,Scandalios
Fig. 6. Maize ascorbate peroxidase and glutathione reductase transcriptsprofiles in response to salt stress. Seedlings were exposed to increasingNaCl concentrations for 24 h, or to 0.25 M NaCl for 24 h, 48 h and 72 h.Controls comprised of RNA extracted from plants that remained on soil orthat were transferred to half-strength MS medium (MS). An 18-S DNAprobe from pea rRNA was used as loading control. The results arerepresentative of two independently performed experiments.

levels may have caused protein damage resulting inlower SOD activity levels upon salt stress. Similar to theresults presented here, reduced foliar levels of antioxi-dant activities in response to salt have been described forSOD in rice,14 N. plumbaginifolia,15 and foxtail millet.17
However, increased SOD activity was correlated withsalt-stress tolerance in specific cultivars of rice,14
tomato,16 and foxtail millet.17
Salt stress induced major changes on maize antioxidanttranscript profiles. Cat1 showed the most significantresponse, being strongly induced by salt. This is in agree-ment with previous results on Cat1 modulation by ABA,osmotic stress, and dehydration,7,9 suggesting Cat1 as astress-responsive catalase in maize. On the other hand,Cat2 and Cat3 regulation by ABA, mannitol and dehydra-tion is dependent on developmental stage, tissue, and expo-sure.7,9 In this work, a mild and transient induction in Cat2transcript accumulation was observed at 0.25 M NaCl,while higher salt concentrations had the opposite effect.Interestingly, Cat3 expression was completely abolished in0.5x MS solution, regardless of length of exposure, mask-ing the induction effect caused by salt.
Except for Sod1, SOD transcripts were up-regulated by0.25 M NaCl. For Sod3, Sod4Aand Sod4, this effect was pos-itively correlated to exposure time. Accordingly, with theexception of Sod3.1, the Sod3 gene family was responsive toboth ABA and high osmoticum in developing and germinat-ing embryos.10 Sod4A is also induced by both ABA and highosmoticum, although responses were dependent on tissueand developmental stage.8 On the other hand, Sod4 inductionby ABA was reported to be independent of developmentalstage, representing a general protective response againstABA-mediated stress, while Sod4A would scavenge O2
�–
produced during regular metabolic processes.8 Increases inSOD transcript steady-state levels were not accompanied byincreases in enzymatic activity, similar to SOD responses tofungal toxin.34 and Mn-SOD responses to ABA and highosmoticum.10 A high protein turnover has been suggested tokeep up with a higher demand of SOD activity on stressfulconditions.1 This requirement would be a consequence of thehighly active electron transfer reactions involved in superox-ide dismutation that would cause protein inactivation.34
Similar data, increased mRNA accumulation in parallel tounaltered SOD protein levels, were reported in NaCl-stressedN. plumbaginifolia.15
Abscisic acid (ABA) is a plant growth regulatorinvolved in many growth and developmental processes.ABA is also involved in plant responses to a variety ofenvironmental stresses, such as hyperosmoticum, waterdeficit, and salt.35 The results presented here show thathigh levels of Cat1, Sod3 gene family, Sod4, and Sod4Atranscripts accumulated in response to 0.25 M NaCl. Inparallel to previous data discussed above concerning theregulation of these genes by ABA, the NaCl responsesobserved may have been mediated by increased internal
levels of ABA. However, the possibility of ABA-inde-pendent pathways cannot be ruled out. Both ABA-dependent and ABA-independent pathways regulateCat1 mRNA accumulation in response to dehydrationand osmotic stress in young leaves9 and embryos.36
The increased APx and GR transcript accumulationsindicate that alterations triggered by the treatments werewidespread and not only limited to CATs and SODs. Super-oxide dismutase and catalase maize transcripts encodingmitochondrial and cytosolic isozymes were significantlyup-regulated by salt. The rice APx and GR probes used todetect the maize counterparts are homologous to cDNAsencoding cytosolic isoforms. Therefore, it seems that salttriggered a concerted response for antioxidant genes in var-ious cellular compartments. Similar results were reported inN. plumbaginifolia15 and rice.37 Increased transcript levelsof Cat2 (involved in abiotic protection), Cat3 (coding for aglyoxysomal CAT), SodA (a mitochondrial Mn-SOD), andSodCc (a cytosolic Cu,Zn-SOD), as well as for SodB (achloroplastic Fe-SOD) were reported in N. plumbaginifoliaexposed to 150 mM NaCl.15 Furthermore, rice SodA1 (amitochondrial Mn-SOD), SodB (a plastidic Fe-Sod),SodCc1 and SodCc2 (cytosolic Cu,Zn-SODs), and SodCp(a plastidic Cu,Zn-SOD) were induced by salinity; whileSodA1, SodCc2, and SodB were also induced by ABA.37
Therefore, antioxidant responses appear to be efficiently con-certed between different intracellular compartments in orderto protect cells against the potential hazards of oxidativestress successfully. Moreover, this regulated cell compart-ment response seems to operate and be conserved in differentplant species.
CONCLUSIONS
The results presented here indicate that salt stressimposes some major changes in the antioxidant metabo-lism of young maize seedlings. Salt stress has beenimplicated with enhanced ROS production due to distur-bance of normal cellular metabolic processes in manyplant species.14–17 Its effects may be exerted throughboth osmotic stress as well as ion toxicity. Besidesintrinsic cultivar and species-specific effects, other fac-tors such as salt concentration, tissue and developmentalstage, do affect antioxidant responses. Moreover, asstated before, salinity causes general metabolic distur-bances which, in turn, may interact and enhance itseffects on antioxidant responses.
ACKNOWLEDGEMENTS
We thank the Japanese Rice Genome Research Program ofthe National Institute of Agrobiological Resources (NIAR)and the Institute of the Society of Techno-Innovation in
Salt-induced antioxidant metabolism defenses in maize (Zea mays L.) seedlings 35

Agriculture, Forestry and Fisheries (STAFF) for providing usthe rice EST clones (Genebank accession numbers C28227and AU056093). The authors also wish to thank GabrieleGusmini for assistance in statistical analyses. L. Menezes-Benavente acknowledges a PhD fellowship from CNPq.This article is part of L. Menezes-Benavente’s PhD thesis.
REFERENCES
1. Scandalios JG. Oxygen stress and superoxide dismutase. PlantPhysiol 1993; 101: 7–12.
2. Dat J, Vandenabeele S, Vranová E, Van Montagu M, Inzé D, VanBreusegem F. Dual action of the active oxygen species duringplant stress responses. Cell Mol Life Sci 2000; 57: 779–795.
3. Lamb C, Dixon RA. The oxidative burst in plant diseaseresistance. Annu Rev Plant Physiol Plant Mol Biol 1997; 48:251–275.
4. Halliwell B, Gutteridge JMC. (eds) Free Radicals in Biology andMedicine, 3rd edn. Oxford: Oxford University Press, 1999.
5. Scandalios JG, Guan LM, Polidoros AN. Catalases in plants:gene structure, properties, regulation, and expression. In:Scandalios JG. (ed) Oxidative Stress and the Molecular Biologyof Antioxidant Defenses. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory Press, 1997; 343–406.
6. Scandalios JG. Molecular genetics of superoxide dismutase inplants. In: Scandalios JG. (ed) Oxidative Stress and theMolecular Biology of Antioxidant Defenses. Cold Spring Harbor,NY: Cold Spring Harbor Laboratory Press, 1997; 527–568.
7. Guan LM, Scandalios JG. Effects of the plant growth regulatorabscisic acid and high osmoticum on the developmentalexpression of the maize catalase genes. Physiol Plant 1998; 104:413–422.
8. Guan LM, Scandalios JG. Two structurally similar maizecytosolic superoxide dismutase genes, Sod4 and Sod4A, responddifferentially to abscisic acid and high osmoticum. Plant Physiol1998; 117: 217–224.
9. Guan LM, Scandalios JG. Catalase transcript accumulation inresponse to dehydration and osmotic stress in leaves of maizeviviparous mutants. Redox Report 2000; 5: 377–383.
10. Zhu D, Scandalios JG. Differential accumulation of manganese-superoxide dismutase transcripts in maize in response to abscisicacid and high osmoticum. Plant Physiol 1994; 106:173–178.
11. Zhu JK. Plant salt tolerance. Trends Plant Sci 2001; 6: 66–67.12. Dhaliwal GS, Arora R. Stresses in agroecosystems: concepts and
approaches. In: Dhaliwal GS, Arora R. (eds) EnvironmentalStress in Crop Plants. New Delhi, India: Ajay VermaCommonwealth Publishers, 1999; 1–18.
13. Ehret DL, Plant AL. Salt tolerance in crop plants. In: DhaliwalGS, Arora R. (eds) Environmental Stress in Crop Plants. NewDelhi, India: Ajay Verma Commonwealth Publishers, 1999;69–120.
14. Dionisio-Sese ML, Tobita S. Antioxidant responses of riceseedlings to salinity stress. Plant Sci 1998; 135: 1–9.
15. Savouré A, Thorin D, Davey M et al. NaCl and CuSO4 treatmentstrigger distinct oxidative defence mechanisms in NicotianaplumbaginifoliaL. Plant Cell Environ 1999; 22: 387–396.
16. Mittova V, Tal M, Volokita M, Guy M. Salt stress induces up-regulation of an efficient chloroplast antioxidant system in thesalt-tolerant wild tomato species Lycopersicon pennellii but notin the cultivated species. Physiol Plant 2002; 115: 393–400.
17. Sreenivasulu N, Grimm B, Wobus U, Weschke W. Differentialresponse of antioxidant compounds to salinity stress in salt
tolerant and salt sensitive seedlings of foxtail millet (Setariaitalica). Physiol Plant 2000; 109: 435–442.
18. Murashige T, Skoog F. A revised medium for rapid growth andbioassays with tobacco tissue cultures. Plant Physiol 1962; 15:473–497.
19. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Proteinmeasurement with the Folin phenol reagent. J Biol Chem 1951;193: 265–275.
20. Beers RF, Sizer IW. A spectrophotometric method for measuringthe breakdown of hydrogen peroxide by catalase. J Biol Chem1952; 195: 133–140.
21. Beauchamp CO, Fridovich I. Superoxide dismutase: improvedassays and an assay applicable to acrylamide gels. Anal Biochem1971; 44: 276–287.
22. Scandalios JG. Genetic control of multiple molecular forms ofcatalase in maize. Ann NY Acad Sci 1968; 151: 274–293.
23. Scandalios JG. Genetic control of multiple molecular forms ofenzymes in maize; a review. Biochem Genet 1969; 3: 37–79.
24. Thompson WF, Everett M, Polans NO, Jorgensen RA, PalmerJD. Phytochrome control of RNA levels in developing pea andmung-bean leaves [Pisum sativum, Vigna radiata]. Planta 1983;158: 487–500.
25. Sambrook J, Fritsch EF, Maniatis T. (eds) Molecular Cloning: ALaboratory Manual, 2nd edn. Cold Spring Harbor, NY: ColdSpring Harbor Laboratory Press, 1989.
26. Church GM, Gilbert W. Genomic sequencing. Proc Natl Acad SciUSA 1984; 81: 1991–1995.
27. Feinberg AP, Vogelstein B. A technique for radiolabelling DNArestriction endonuclease fragments to high specific activity. AnalBiochem 1983; 132: 6–13.
28. Jorgensen RA, Cuellar RE, Kavanagh TA, Thompson WF.Structure and variation in ribosomal-RNA genes of pea –characterization of a cloned rDNA repeat and chromosomalrDNA variants. Plant Mol Biol 1987; 8: 3–12.
29. Thordal-Christensen H, Zhang Z, Wei Y, Collinge D. Subcellularlocalization of H2O2 in plants. H2O2 accumulation in papillae andhypersensitive response during the barley-powdery mildewinteraction. Plant J 1997; 11: 1187–1194.
30. Gómez JM, Hernández JA, Jiménez A, Del Rio LA, Sevilla F.Differential response of antioxidative enzymes of chloroplastsand mitochondria to long-term NaCl stress of pea plants. FreeRadic Res 1999; 31: S11–S18.
31. Lee DH, Kim YS, Lee CB. The inductive responses of theantioxidant enzymes by salt stress in the rice (Oryza sativa L.). JPlant Physiol 2001; 158: 737–745.
32. Kernodle SP, Scandalios JG. Structural organization, regulationand expression of the chloroplastic superoxide dismutase Sod1gene in maize. Arch Biochem Biophys 2001; 391: 137–147.
33. Noctor G, Foyer CH. Ascorbate and glutathione: keeping activeoxygen under control. Annu Rev Plant Physiol Plant Mol Biol1998; 49: 249–279.
34. Williamson JD, Scandalios JG. Differential response of maizecatalase and superoxide dismutase to the photoactivated fungaltoxin cercosporin. Plant J 1992; 2: 351–358.
35. Skriver K, Mundy J. Gene expression in response to abscisic acidand osmotic stress. Plant Cell 1990; 2: 503–512.
36. Guan LM, Zhao J, Scandalios JG. Cis-elements and trans-factorsthat regulate expression of the maize cat1 antioxidant gene inresponse to ABA and osmotic stress: H2O2 is the likelyintermediary signaling molecule for the response. Plant J 2000;22: 87–95.
37. Kaminaka H, Morita S, Tokumoto M, Masumura T, Tanaka K.Differential gene expression of rice superoxide dismutaseisoforms to oxidative and environmental stresses. Free Radic Res1999; 31: S219–S225.
36 Menezes-Benavente, Kernodle, Margis-Pinheiro,Scandalios


Copyright of Redox Report is the property of Maney Publishing and its content may not be copied or emailed to
multiple sites or posted to a listserv without the copyright holder's express written permission. However, users
may print, download, or email articles for individual use.




















